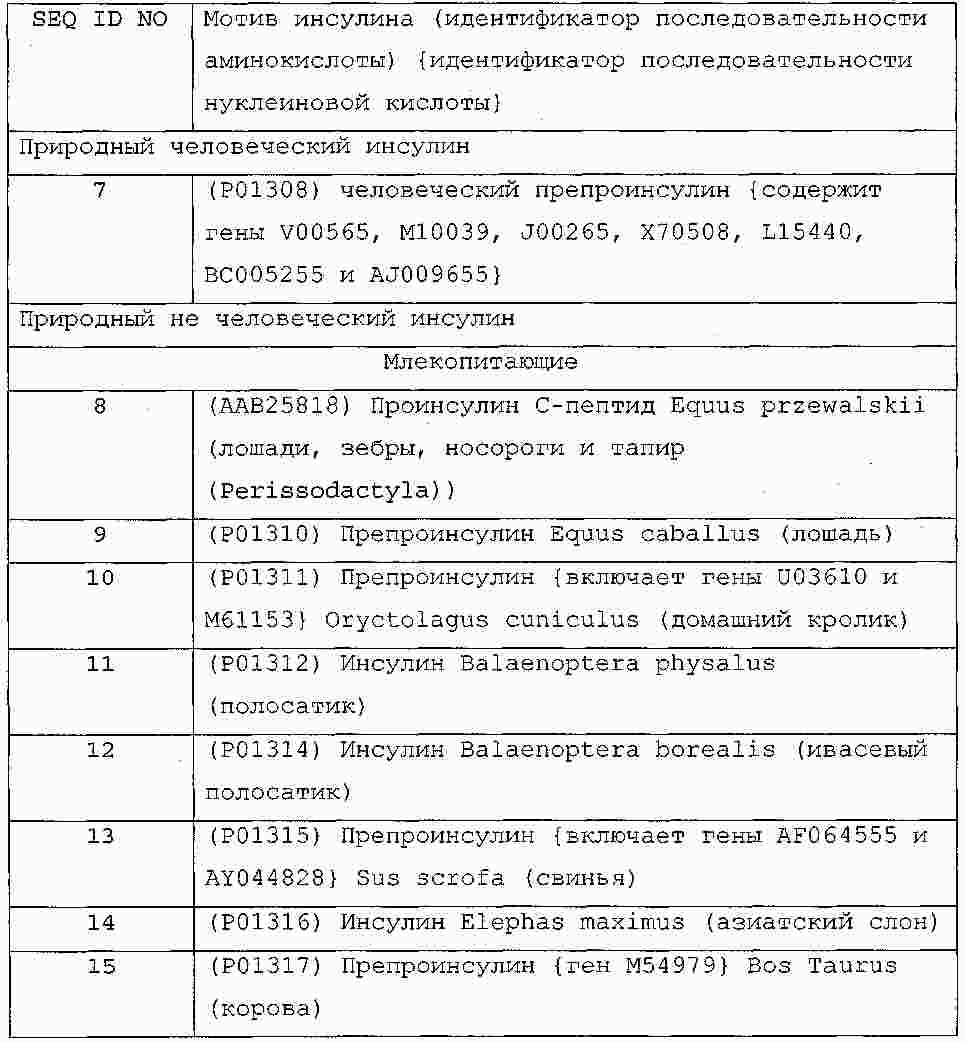
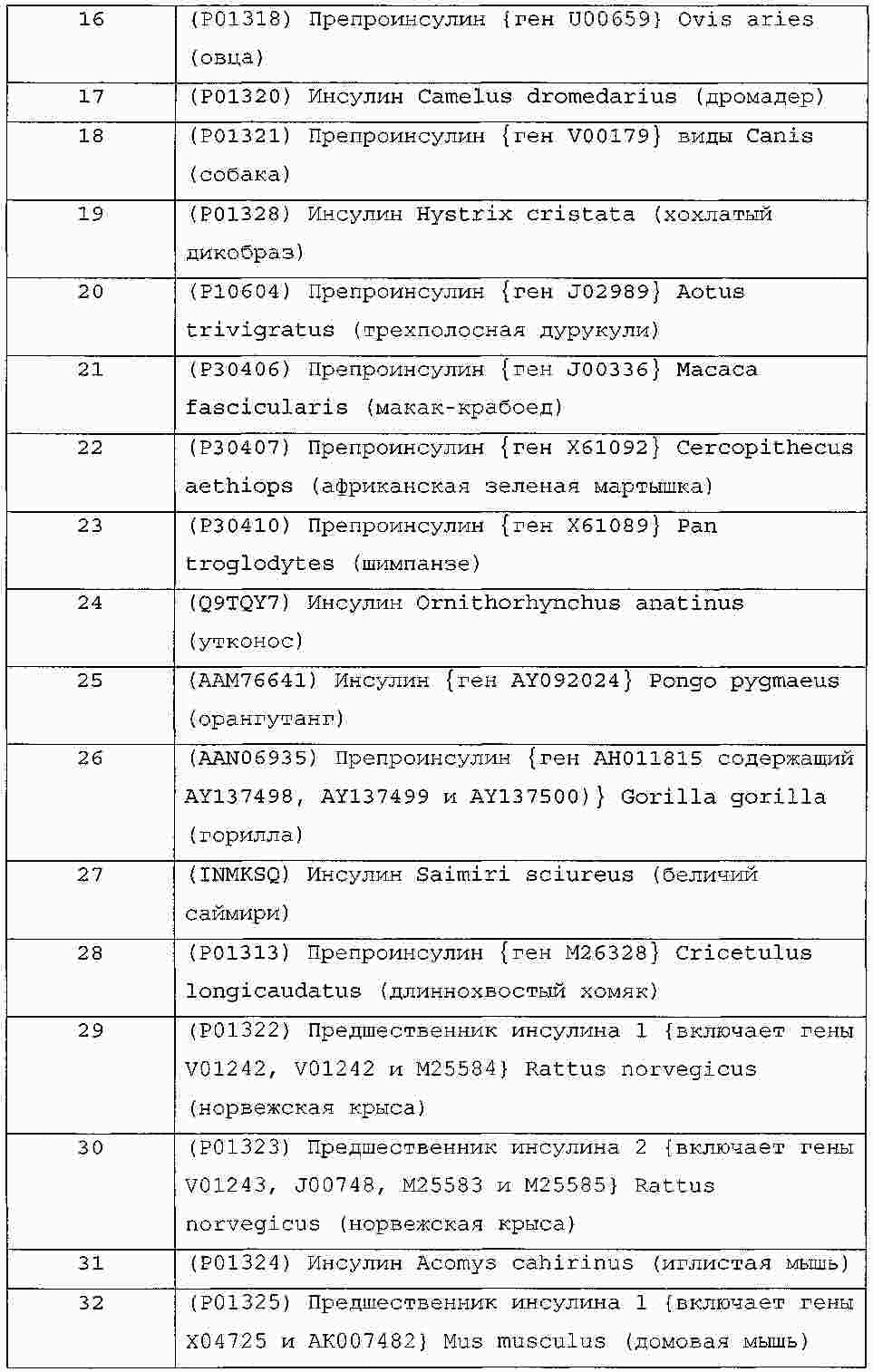
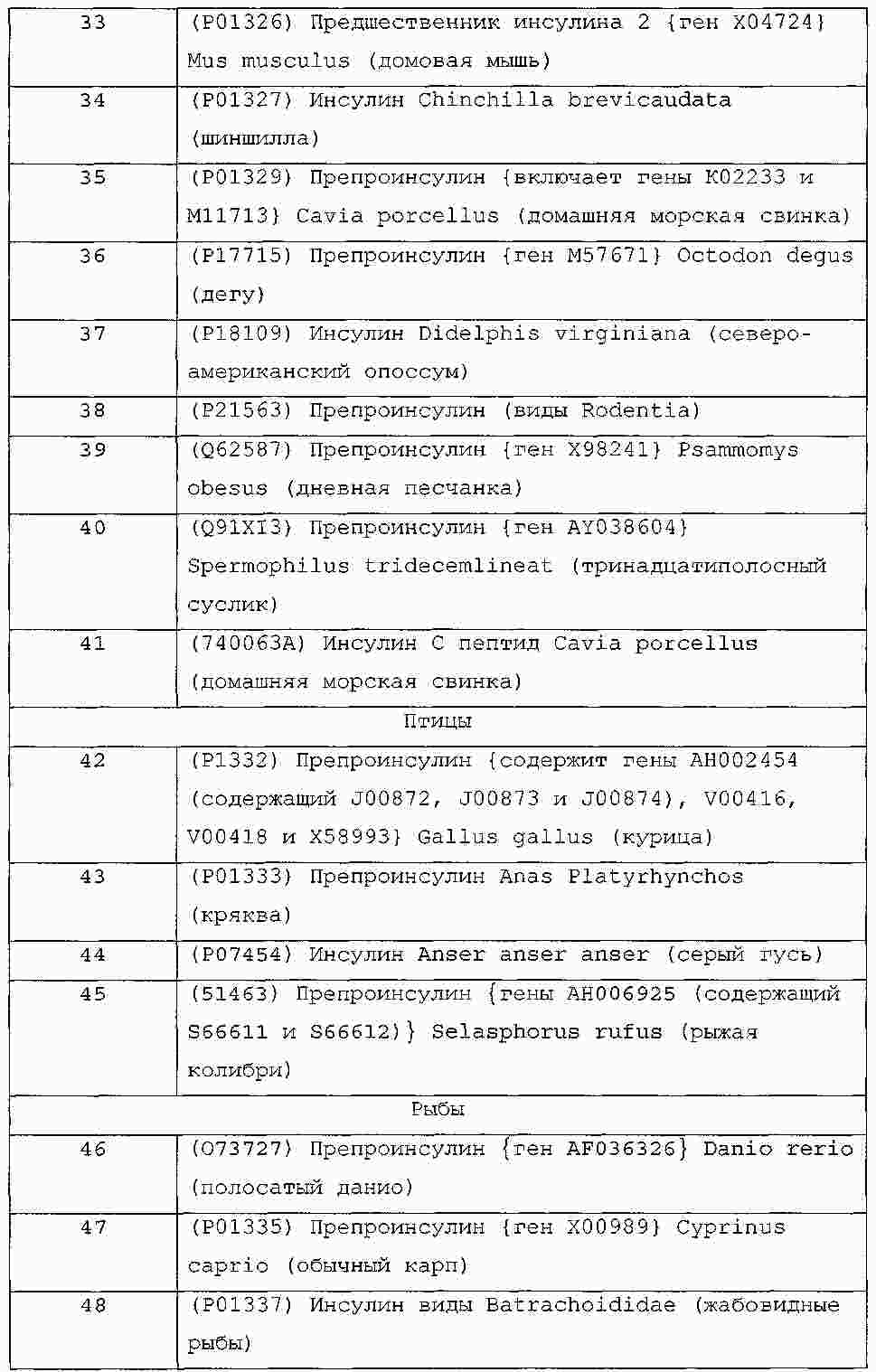
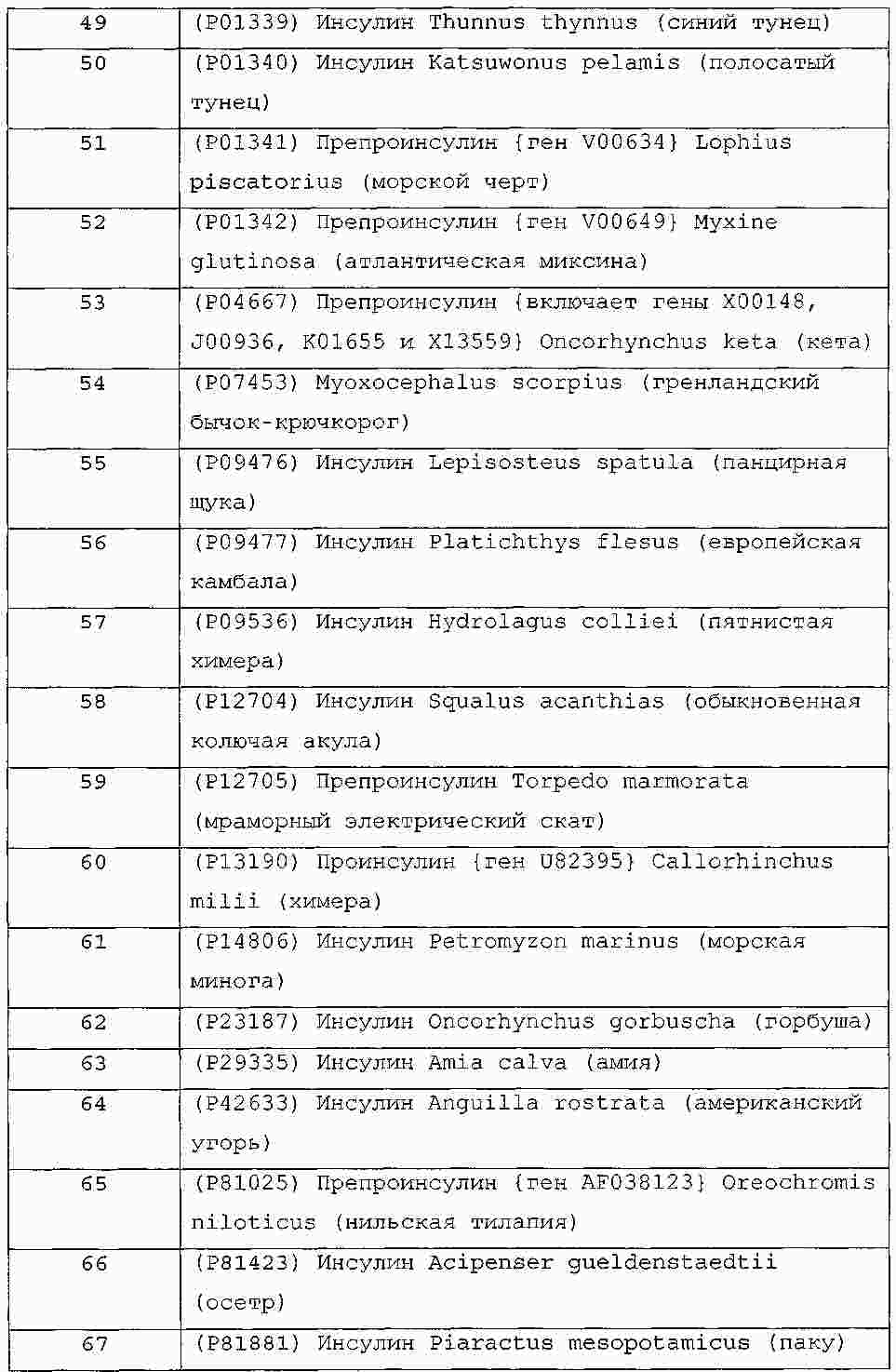
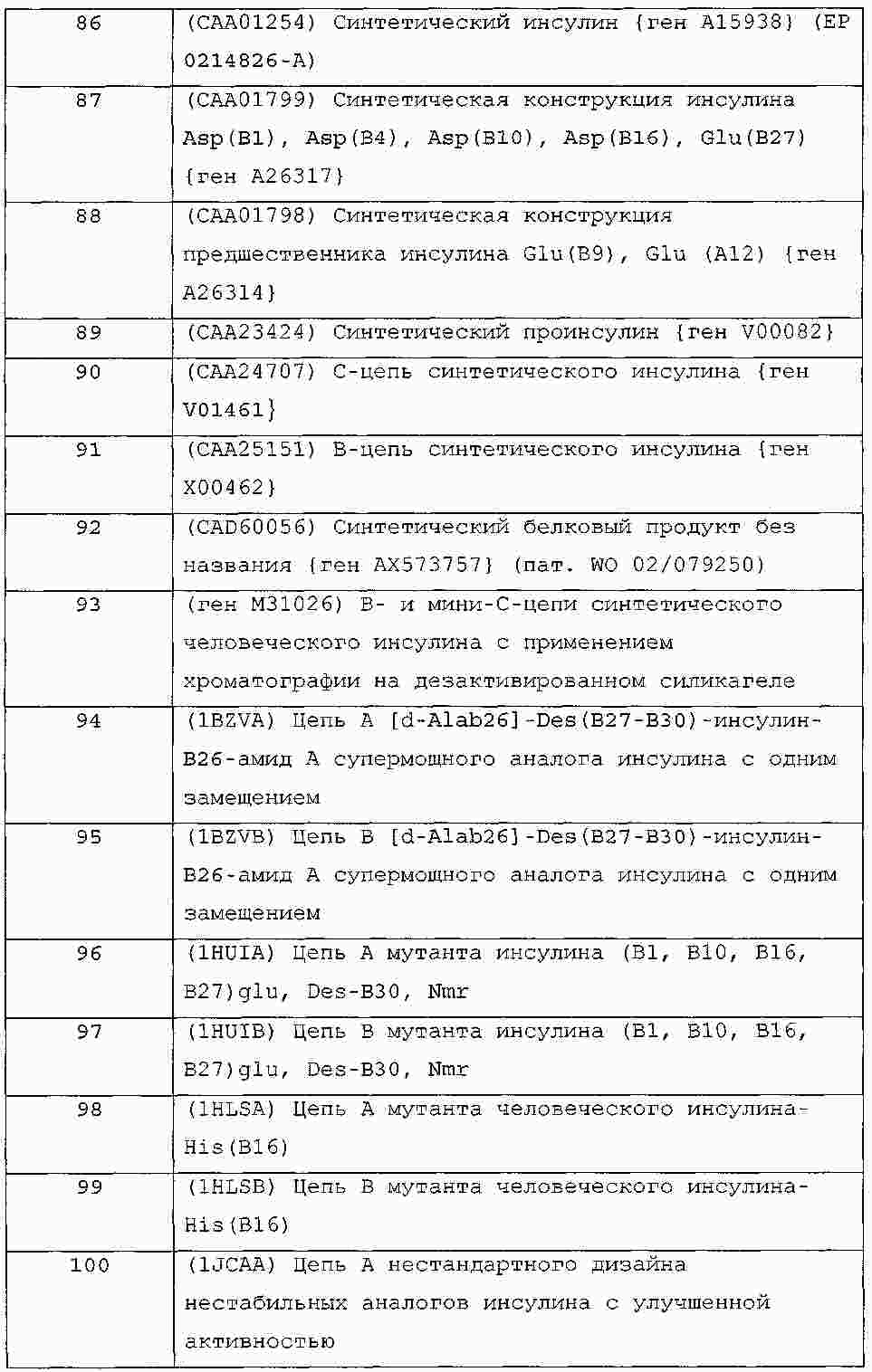
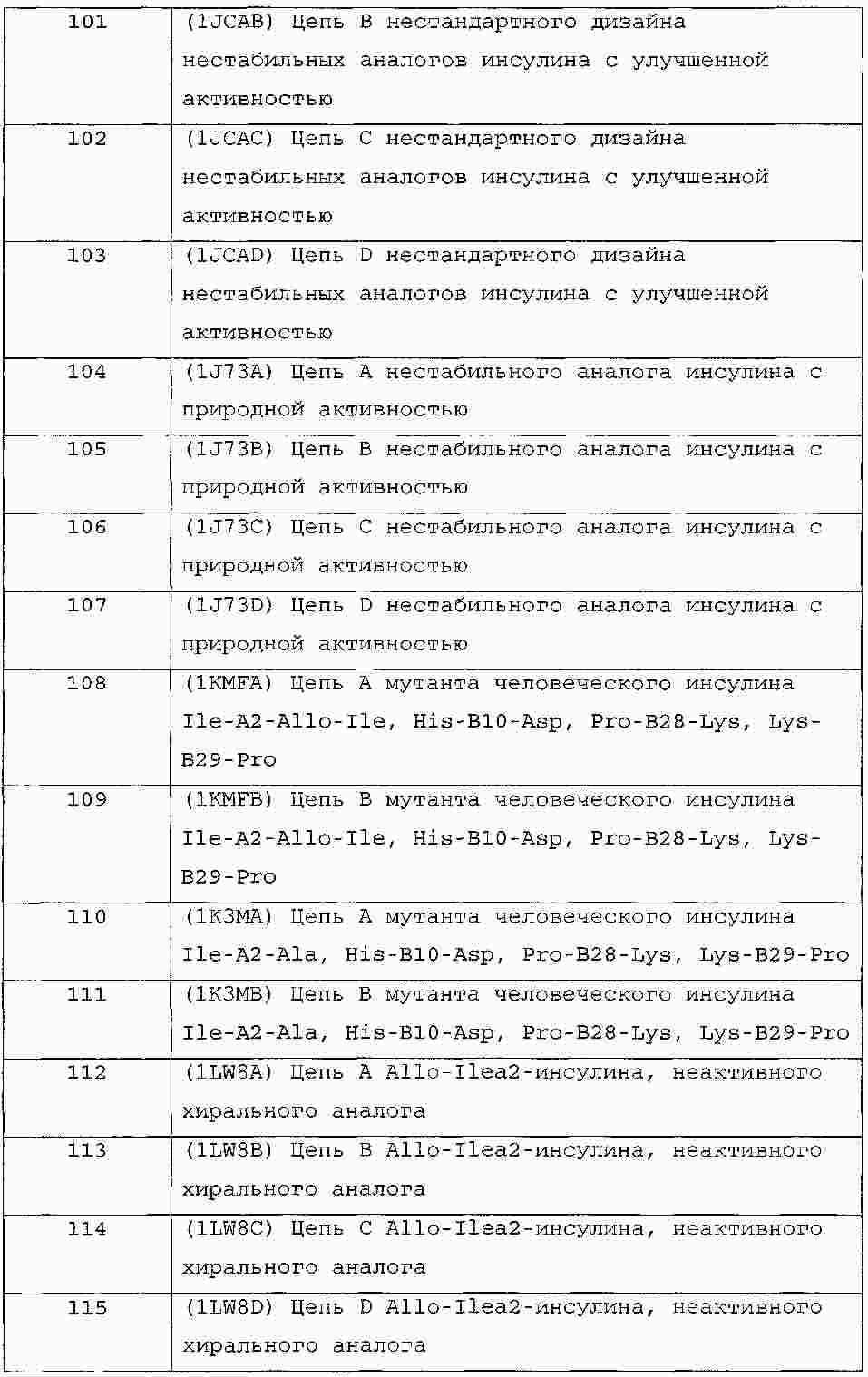
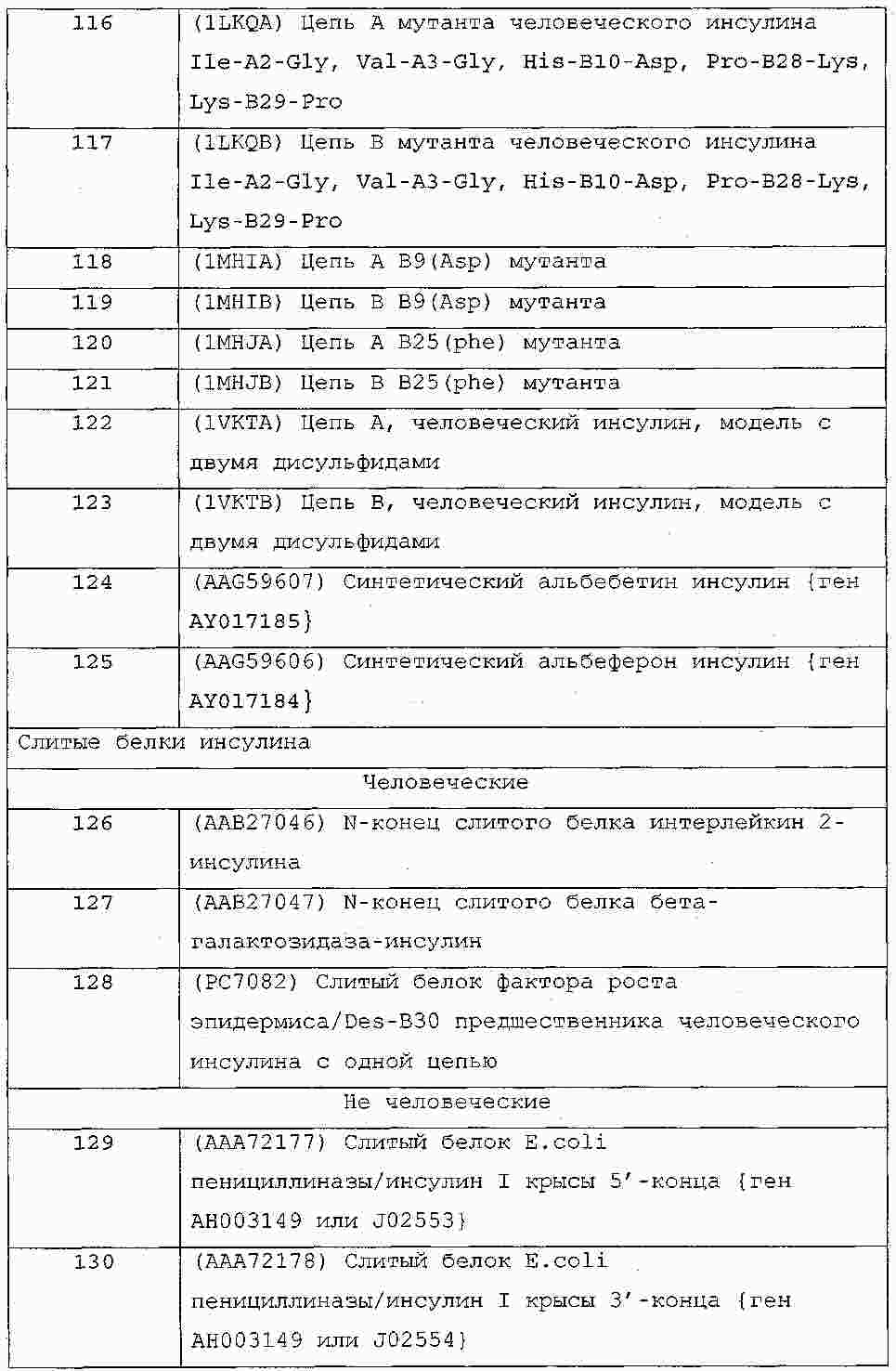
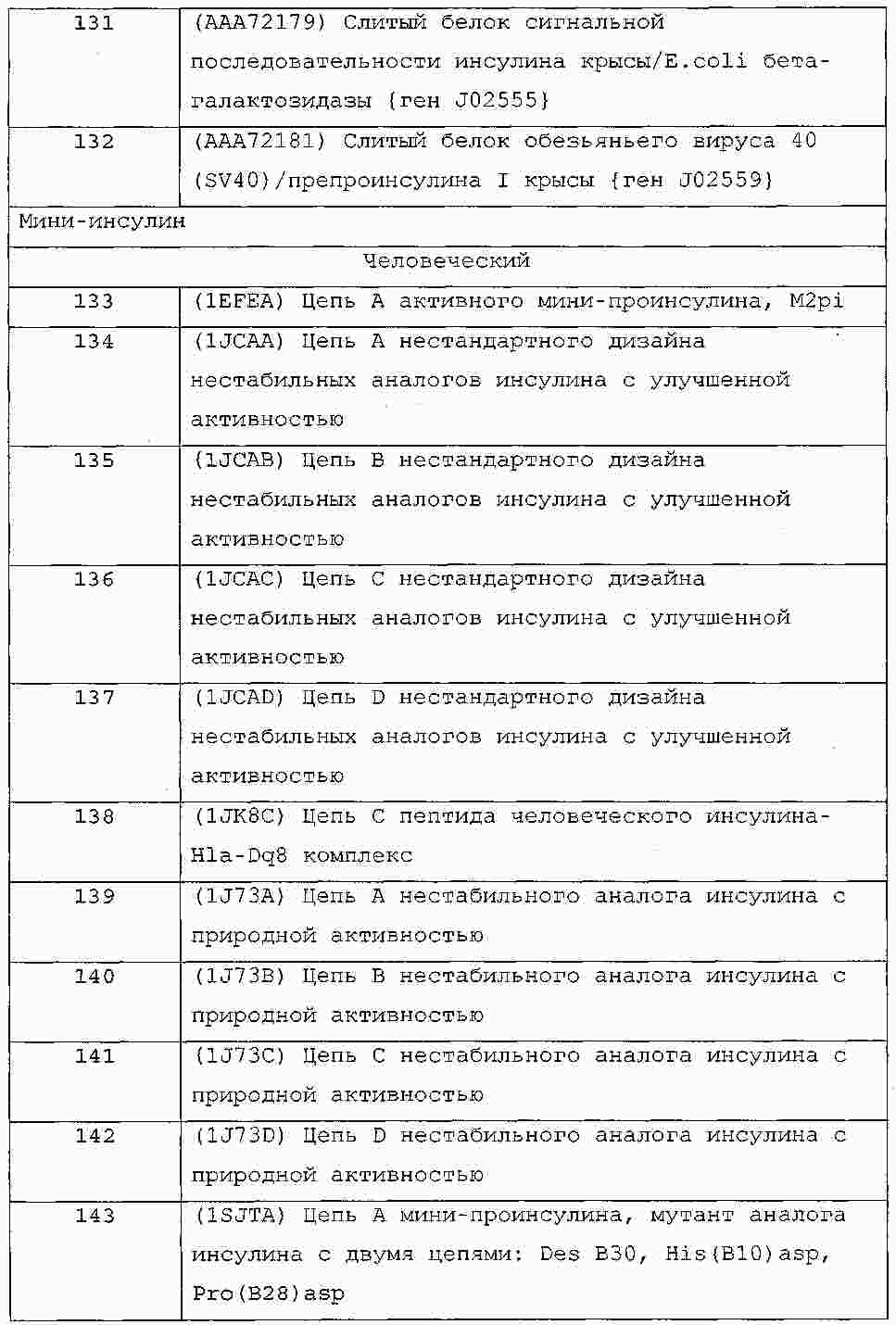
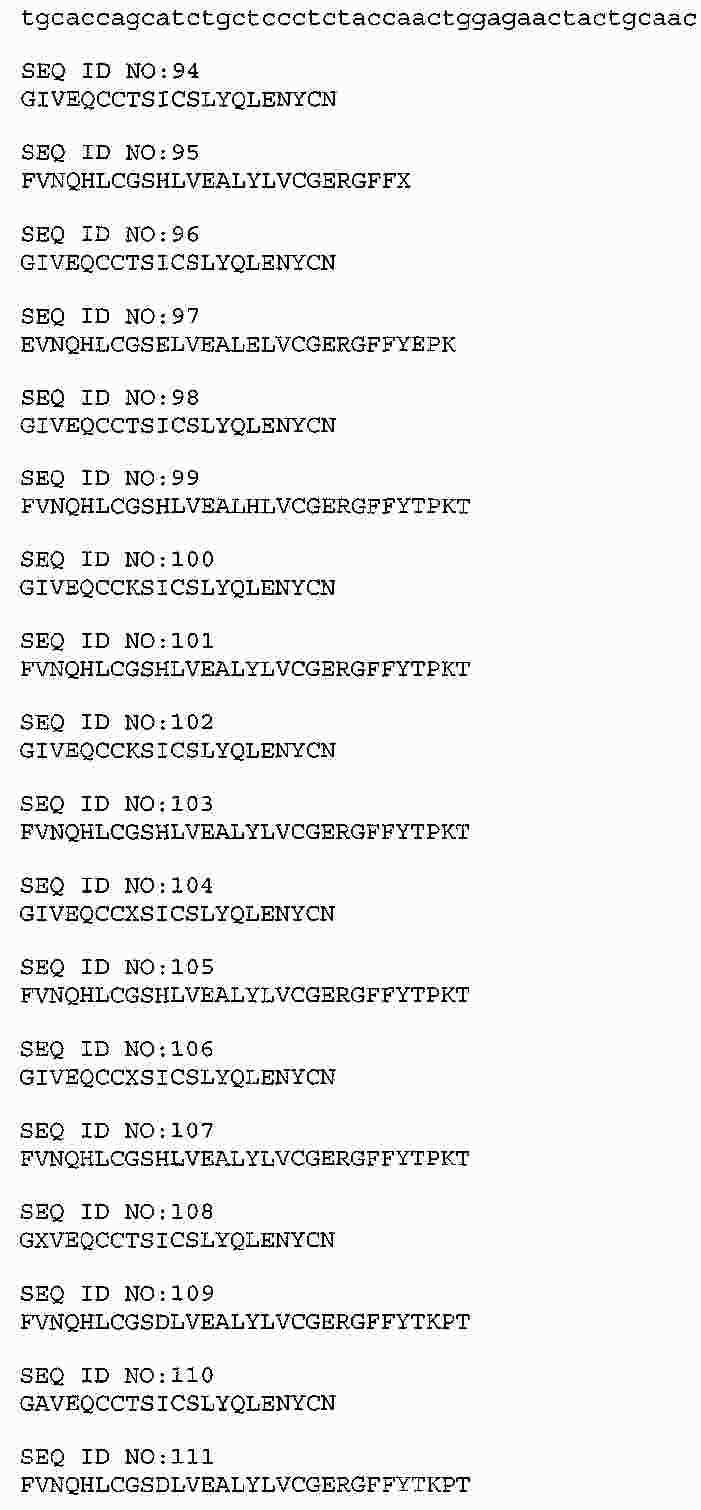
|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000014887b*\id |
|
Термины запроса в документе Реферат Описаны способы получения инсулина в растениях. В одном варианте данное изобретение относится к способу экспрессии инсулина в растениях, включающему: (а) получение конструкции химерной нуклеиновой кислоты, содержащейся в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательности нуклеиновой кислоты, способной контролировать экспрессию в клетках семян растений; и (ii) последовательности нуклеиновой кислоты, кодирующей полипептид инсулина; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения и (с) выращивание клетки растений в зрелом растении, способном давать семена, где семена экспрессируют инсулин. Формула [0001] Способ экспрессии инсулина в семенах растения, включающий: [0002] Способ по п.1, в котором указанная химерная нуклеиновая кислота дополнительно содержит последовательность, кодирующую пептид или полипептид, который способен удерживать полипептид инсулина в окруженном мембраной внутриклеточном компартменте, где указанный окруженный мембраной внутриклеточный компартмент представляет собой эндоплазматический ретикулюм (ER) или масляное тельце. [0003] Способ по п.2, в котором указанный пептид, удерживающий полипептид инсулина в ER, выбран из группы, состоящей из KDEL, HDEL, DDEL, ADEL и SDEL. [0004] Способ по п.1, где последовательность (ii) дополнительно содержит последовательность, кодирующую сигнальный пептид. [0005] Способ по п.4, в котором указанным сигнальным пептидом является сигнальный пептид, связанный с патогенезом табака (PR-S). [0006] Способ по п.5, в котором указанный сигнальный пептид имеет последовательность SEQ ID NO:161. [0007] Способ по п.2, в котором указанным полипептидом, удерживающим полипептид инсулина в окруженном мембраной внутриклеточном компартменте, является белок масляного тельца. [0008] Способ по п.7, в котором указанный белок масляного тельца выбран из группы белков масляного тельца, состоящей из олеозина, калеозина и стеролеозина. [0009] Способ по п.8, в котором указанные белки имеют последовательности, выбранные из группы, состоящей из SEQ ID NO:156, SEQ ID NO:157 и SEQ ID NO:158, SEQ ID NO:159 и SEQ ID NO:160. [0010] Способ по п.2, в котором указанная химерная нуклеиновая кислота дополнительно содержит последовательность, кодирующую стабилизирующий белок, слитую в рамке считывания с последовательностью нуклеиновой кислоты, кодирующей инсулин. [0011] Способ по п.10, в котором указанная химерная нуклеиновая кислота дополнительно содержит последовательность, кодирующую сигнальный пептид, слитую в рамке считывания с последовательностью нуклеиновой кислоты, кодирующей инсулин. [0012] Способ по п.11, в котором указанный сигнальный пептид является сигнальным пептидом, связанным с патогенезом табака (PR-S). [0013] Способ по п.12, в котором указанный сигнальный пептид имеет последовательность SEQ ID NO:161. [0014] Способ по п.10, в котором стабилизирующий белок обеспечивает объединение полипептида инсулина с масляным тельцем при сборе и измельчении семян. [0015] Способ по п.14, в котором указанный стабилизирующий белок представляет собой одноцепочечное антитело со специфичностью к масляному тельцу. [0016] Способ по п.10, в котором указанный стабилизирующий белок выбран из группы, состоящей из одноцепочечного антитела со специфичностью к масляному тельцу и В-субъединицы токсина холеры. [0017] Способ по п.1, в котором химерную последовательность нуклеиновой кислоты вводят в клетку растения в условиях ядерной геномной интеграции. [0018] Способ по п.1, в котором предпочтительным для семян промотором является промотор фазеолина. [0019] Способ по любому из пп.1-18, в котором последовательность нуклеиновой кислоты, кодирующая инсулин, выбрана из группы, включающей последовательности, кодирующие человеческий инсулин, свиной инсулин и бычий инсулин. [0020] Способ по любому из пп.1-19, в котором последовательностью нуклеиновой кислоты, кодирующей инсулин, является последовательность, кодирующая мини-инсулин. [0021] Способ по любому из пп.1-20, в котором последовательность нуклеиновой кислоты, кодирующая инсулин, оптимизирована в отношении кодонов растения. [0022] Способ получения семян растений, содержащих инсулин, включающий: [0023] Растение, способное производить семена, содержащие инсулин, который экспрессируется последовательностью химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции: [0024] Семя растения по п.23, содержащее последовательность химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции (a) первую последовательность нуклеиновой кислоты, которая содержит предпочтительный для семян промотор, функционально связанный со (b) второй последовательностью нуклеиновой кислоты, кодирующей полипептид инсулина, и последовательностью, кодирующей полипептид, который способен удерживать полипептид инсулина в окруженном мембраной внутриклеточном компартменте. [0025] Последовательность нуклеиновой кислоты, кодирующая инсулин, связанная с последовательностью нуклеиновой кислоты, содержащей предпочтительный для семян промотор, и содержащая последовательность, кодирующую полипептид, который способен удерживать полипептид инсулина в окруженном мембраной внутриклеточном компартменте, для использования в способе по любому из пп.2-21. Полный текст патента Область техники Данное изобретение относится к методам генной инженерии растений и получения инсулина. Более конкретно, данное изобретение относится к способам получения инсулина в семенах растений. Инсулин представляет собой важный пептидный гормон, требуемый для поддержания гомеостаза глюкозы в крови у млекопитающих, включая человека, и других позвоночных. У здоровых индивидуумов повышение уровня глюкозы в крови стимулирует β-клетки поджелудочной железы к выделению инсулина. Затем полипептид инсулина связывается со специфическими рецепторами в мышцах, печени и жировой ткани, что ведет к увеличению потребления глюкозы этими целевыми тканями, повышению метаболизма и снижению вырабатывания глюкозы печенью. Суммарный эффект этих реакций позволяет сохранить концентрации глюкозы в крови на постоянном уровне. У индивидуумов, страдающих сахарным диабетом, аномально низкие концентрации инсулина присутствуют сами по себе в виде хронической гипергликемии. Клинические проявления хронической гипергликемии являются многообразными и включают слепоту, почечную недостаточность и, если не лечить, в конечном счете приводят к смерти. По некоторым оценкам, сахарный диабет стоит на третьем месте среди самых частых причин смерти в промышленно развитых странах после сердечно-сосудистых заболеваний и рака (Barfoed, Н.С., 1987, Chem. Eng. Prog. 83: 49-54). Для достижения эффективного потребления и метаболизма глюкозы крови клетками пациентов, страдающих диабетом, можно лечить обычным введением инсулина. Приблизительно 0,7% населения мира страдает инсулинозависимым диабетом (сахарный диабет I типа) (Winter, J. et al., 2000, J. of Biotechnol, 84: 175-185). Кроме того, по оценкам, количество пациентов, у которых диагностирован диабет, удвоится приблизительно до 300 млн в ближайшие 25 лет (Kjeldsen, Т. et al., 2001, Biotechnol. Gen. Eng. Rev. 18: 89-121). Следовательно, возможность экономичного рентабельного производства человеческого инсулина в количествах, удовлетворяющих ожидаемую растущую мировую потребность в инсулине, крайне желательна. In vivo полипептид человеческого инсулина образуется в β-клетках поджелудочной железы в виде предшественника одинарной полипептидной цепи из 110 аминокислот, препроинсулина, который включает расположенные на N-конце 24 аминокислоты пре-последовательности, которые отщепляются сразу же после завершения биосинтеза цепи (Steiner, D.F., 2000. J. Ped. Endocrinol. Metab. 13: 229-239). Проинсулин состоит из цепей В и А, связанных связывающим пептидом (С-пептид). Во время упаковки гормона для секреции С-пептид отщепляется и удаляется конвертазами прогормона, РС2 и РС1/РС3 (Steiner, D.F., 2000. J. Ped. Endocrinol. Metab. 13: 229-239). Остается зрелый человеческий инсулин, белок из 51 аминокислоты, содержащий две полипептидные цепи, А (21 аминокислота) и В (30 аминокислот), связанные двумя межцепочечными дисульфидными связями. Кроме того, цепь А содержит одну внутрицепочечную дисульфидную связь. Человеческий инсулин получают с помощью множества различных методов. Микроорганизмы, такие как Escherichia coli (Frank et al., 1981, in Peptides: Proceedings of the 7 th American Peptide Chemistry Symposium (Rich & Gross, eds.), Pierce Chemical Co., Rockford, Ill, p. 729-739; Chan et al., 1981, Proc. Natl. Acad. Sci. USA 78: 5401-5404), Saccharomyces cerevisiae (Thim et al., 1986, Proc. Natl. Acad. Sci. USA 83: 6766-6770), обычно применяют для рекомбинантного получения инсулина. У Wang et al. (Biotechnol. Bioeng., 2001, 73: 74-79) показано, что грибы, такие как Pichia pastoris, также подходят для получения инсулина. Альтернативные производственные процессы включают продуцирование в клеточных линиях млекопитающих, отличных от человека (Yanagita, M., et al., 1992, FEBS Lett 311: 55-59), выделение из поджелудочной железы человека, пептидный синтез или полусинтетическое превращение в человеческий инсулин свиного и бычьего инсулина. Однако все эти методы дают низкий выход и требуют более высоких затрат, чем это желательно. Применение растений в качестве биореакторов для промышленного получения рекомбинантных белков хорошо известно, и было получено множество белков, включая человеческие терапевтические белки. Например, в патентах США 4956282, 5550038 и 5629175 описано получение γ-интерферона в растениях; в патентах США 5650307, 5716802 и 5763748 подробно описано получение сывороточного альбумина человека в растениях; в патентах США 5202422, 5639947 и 5959177 описано получение антител в растениях. Одним из значительных преимуществ систем получения рекомбинантного белка на основе растений является то, что при увеличении площади выращивания растений продукция белка может быть недорого масштабирована для получения значительно больших количеств белка. Наоборот, системы с применением ферментации и клеточных культур занимают много места, требуют большого количества оборудования и потребляют много энергии, что делает увеличение производственных мощностей очень дорогим. Однако, несмотря на то что применение растений в качестве биореакторов описано во множестве источников и несмотря на указанное выше ожидаемое увеличение потребности в больших количествах инсулина, в известном уровне техники представлено только ограниченное количество методов, которые наглядно показывают возможность получения инсулина в растениях (см. Arakawa et al. Nature Biotech., 1998, 16: 934-938; PCT 01/72959). У Arakawa et al. описано получение слитого белка, содержащего инсулин, в клубнях трансгенного картофеля. Однако инсулин присутствует только в количествах не более 0,05% от общего количества растворимого белка, присутствующего в трансгенных клубнях. При уровне 0,05% от общего растворимого белка большие количества биомассы должны быть обработаны для экстрагирования белка, что не выгодно с точки зрения экономического применения картофельных клубней. Более того, у Arakawa et al. не описано выделение инсулина из ткани картофельного клубня, но вместо этого предложен подход к профилактике наступления диабета I типа путем получения иммунотолерантности, который включает введение инсулина при поедании трансгенных картофельных клубней. В заявке на патент РСТ WO 01/72959 описано получение слитого белка, содержащего инсулин, в хлоропластах трансгенного табака. Однако, по существу, относя недостатки на счет уровней аккумуляции человеческих белков в тканях растений, изобретение, описанное в WO 01/72959, ограничено в том, что получение в хлоропластах дает аккумуляцию инсулина в зеленых тканях, в первую очередь в листьях табака. Из-за относительно высокого содержания воды в зеленых тканях должно быть обработано большое количество биомассы. Дальнейшее получение инсулина потребует немедленного экстрагирования из биомассы сразу же после сбора, так как листья быстро высыхают при хранении. Таким образом, с точки зрения недостатков, имеющихся в методах рекомбинантного получения инсулина в растениях известного уровня техники, в настоящее время не ясно, когда и как синтетические возможности растений могут быть использованы для достижения коммерческого получения инсулина в растениях. В данной области техники существует необходимость улучшения способов коммерческого получения инсулина в растениях. Данное изобретение относится к улучшенным способам получения инсулина в растениях. В частности, данное изобретение относится к способам получения инсулина в семенах. Следовательно, в данном изобретении представлен способ экспрессии инсулина в растениях, включающий: (а) получение конструкции химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений; и (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения и (c) выращивание клетки растений в зрелом растении, способном давать семена, где семена экспрессируют инсулин. В предпочтительном варианте данного изобретения последовательностью нуклеиновой кислоты, способной контролировать экспрессию в семенах растений, является предпочтительный для семян промотор, такой как промотор фазеолина. В предпочтительном варианте данного изобретения инсулин экспрессируется таким образом, который позволяет аккумулировать полипептид инсулина в мембране, окружающей внутриклеточный компартмент в клетках семян. Следовательно, в данном изобретении представлен способ экспрессии инсулина в растениях, который включает: (a) получение конструкции химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений; (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; и (iii) последовательность нуклеиновой кислоты, кодирующую полипептид, способный удерживать полипептид инсулина в мембране, окружающей внутриклеточный компартмент; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения и (c) выращивание клетки растений в зрелом растении, способном давать семена, где семена экспрессируют инсулин. В другом предпочтительном варианте данного изобретения мембраной, окружающей внутриклеточный компартмент, является эндоплазматическая сеть (ER) или ER-производная накопительная везикула. Следовательно, в данном изобретении представлен способ экспрессии инсулина в растениях, который включает: (a) получение конструкции химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений; и (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; и (iii) последовательность нуклеиновой кислоты, кодирующую полипептид, способный удерживать полипептид инсулина в ER или ER-производной накопительной везикуле; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения и (c) выращивание клетки растений в зрелом растении, способном давать семена, где семена экспрессируют инсулин. В другом предпочтительном варианте конструкцию химерной нуклеиновой кислоты вводят в клетку растений в условиях ядерной геномной интеграции. При таких условиях последовательность химерной нуклеиновой кислоты стабильно интегрируется в геном растения. В еще одном предпочтительном варианте последовательность нуклеиновой кислоты, кодирующая инсулин, оптимизирована для применения кодона растений и последовательность нуклеиновой кислоты, кодирующая связывающий пептид (С-пептид), укорочена. Предпочтительные последовательности нуклеиновых кислот в соответствии с данным изобретением кодируют человеческий, бычий или свиной инсулин. В соответствии с данным изобретением последовательность нуклеиновой кислоты, кодирующую последовательность проинсулина, применяют, если проинсулин модифицирован путем укорачивания длины С-пептида. В другом варианте в данном изобретении представлен способ получения семян, содержащих инсулин, из растений. Следовательно, в данном изобретении представлен способ получения семян растений, содержащих инсулин, включающий: (a) получение конструкции химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений; и (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения; (c) выращивание клетки растений в зрелом растении, способном давать семена; и (d) получение семян из указанных растений, где семена содержат инсулин. Предпочтительно по крайней мере 0,1% от общего белка семян, присутствующего в семенах, составляет инсулин. Семена могут применяться для получения популяции растений, каждое из которых содержит множество семян, экспрессирующих инсулин. В данном изобретении также представлены растения, способные давать семена, экспрессирующие инсулин. В предпочтительном варианте данного изобретения растения способны дать семена, содержащие последовательность химерной нуклеиновой кислоты, содержащую в 5'-3' направлении транскрипции: (a) первую последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений, с которыми она функционально связана; и (b) вторую последовательность нуклеиновой кислоты, кодирующую полипептид инсулина, где семена содержат инсулин. Предпочтительно по крайней мере 0,1% от общего белка семян, присутствующего в семенах, составляет инсулин. В предпочтительном варианте последовательность химерной нуклеиновой кислоты интегрируют в ядерный геном растения. В еще одном предпочтительном варианте данного изобретения применяемым растением является сафлор, лен или Arabidopsis. В еще одном аспекте данного изобретения представлены семена растений, экспрессирующие инсулин. В предпочтительном варианте данного изобретения семена растений содержат последовательность химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции: (a) первую последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений, с которыми она функционально связана; и (b) вторую последовательность нуклеиновой кислоты, кодирующую полипептид инсулина. Предпочтительно по крайней мере 0,1% от общего белка семян, присутствующего в семенах, составляет инсулин. Семена являются источником, из которого желаемый полипептид инсулина, который синтезируется в клетках семян, может быть экстрагирован, и инсулин может применяться для лечения пациентов, страдающих диабетом. Другие характеристики и преимущества данного изобретения становятся понятны из представленного ниже подробного описания. Должно быть понятно, что подробное описание и конкретные примеры, показывающие предпочтительные варианты, даны только для иллюстрации, так как множество изменений и модификаций в пределах объема данного изобретения становятся очевидны специалистам в данной области техники при прочтении подробного описания. Далее описаны чертежи, относящиеся к данному изобретению. На фиг. 1 изображена нуклеотидная последовательность (SEQ ID NO:1) и выведенная последовательность аминокислот (SEQ ID NO:2) слитого белка инсулина (PRS-D9scFv-Klip27-MI-KDEL) pSBS4404. Расчетная последовательность аминокислот представлена однобуквенным кодом. Выведенная последовательность аминокислот PRS сигнального пептида показана курсивом, выведенная последовательность аминокислот D9scFv показана жирным шрифтом, выведенная последовательность аминокислот Klip27 подчеркнута, выведенная последовательность аминокислот мини-инсулина показана жирным курсивом и последовательность KDEL показана жирным подчеркнутым шрифтом. На фиг. 2 изображена нуклеотидная последовательность (SEQ ID NO:3) и выведенная последовательность аминокислот (SEQ ID NO:4) слитого белка инсулина (OLEO-Klip8-Klip7-MI) pSBS4405. Расчетная последовательность аминокислот представлена однобуквенным кодом. Выведенная последовательность аминокислот Arabidopsis thaliana 18 кДа олеозина показана курсивом, выведенная последовательность аминокислот Klip8 показана жирным шрифтом, выведенная последовательность аминокислот Klip27 подчеркнута и выведенная последовательность аминокислот мини-инсулина показана жирным курсивом. На фиг. 3 изображена нуклеотидная последовательность (SEQ ID NO:5) и выведенная последовательность аминокислот (SEQ ID NO:6) слитого белка инсулина (PRS-MI-тетраосновный линкер-D9scFv-KDEL) 4414. Расчетная последовательность аминокислот показана однобуквенным кодом. Выведенная последовательность аминокислот PRS сигнального пептида показана курсивом, выведенная последовательность аминокислот мини-инсулина (В30 тетраосновная) показана жирным шрифтом, выведенная последовательность аминокислот тетраосновного линкера подчеркнута, выведенная последовательность аминокислот D9scFv показана жирным курсивом и последовательность KDEL показана жирным подчеркнутым шрифтом. На фиг. 4 (A-D) показана рекомбинантная экспрессия слитых белков инсулина в трансформированных линиях Arabidopsis thaliana (4404-2, -17, -20 и 4405-4) на основе окрашенного кумасси SDS-PAGE и вестерн-блот анализа. Стрелки указывают на положение миграции 38,5 и 34,2 кДа слитых полипептидов, PRS-D9scFv-Klip27-MIw/KDEL и OLEO-Klip8-Klip27-MI соответственно в восстанавливающих условиях. На фиг. 4А (окрашенный кумасси гель) и 4В (соответствующий вестерн-блот с антиинсулином Е2Е3) изображен общий белок семян дикого типа (wt) и трансгенных семян, экспрессирующих 4404 и 4405 конструкции. На фиг. 4В (окрашенный кумасси гель) и 4D (соответствующий вестерн-блот с антиинсулином Е2Е3) показан белок масляного тельца, полученный из семян дикого типа и трансгенных семян, экспрессирующих 4404 и 4405 конструкции. На фиг. 4 (E-F) показана рекомбинантная экспрессия слитых белков инсулина в трансформированных линиях Arabidopsis thaliana (4419-9 и 4414-20) на основе окрашенного кумасси SDS-PAGE и вестерн-блот анализа. Молекулярные массы маркеров (М) равны 10, 15, 25, 37, 50, 75, 100, 150 кДа. Контроль включает hIN (стандарт рекомбинантного человеческого инсулина) и hProIN (стандарт рекомбинантного человеческого проинсулина), выделенные в невосстанавливающих условиях. На фиг. 5 изображены определенные уровни экспрессии в доступных линиях семян Т3 (4404-2, -17, -20, 4405-4, -13, -19) и линиях семян Т2 (4414-9 и -20). Уровни трансгена и % молярной экспрессии MI определяют на основе денситометрии. На фиг. 6 изображен анализ окрашенного кумасси SDS-PAGE (15%) препаратов масляных телец до элюирования (-ОВ), препаратов ОВ после элюирования муравьиной кислотой (-ОВ') и концентрированного элюированного материала (-Е). Стрелки указывают положение мигрирующего слитого полипептида. Контрольная группа дикого типа практически не содержит какие-либо основные белки после элюирования, в то время как концентрированный 4404 материал содержит слитый белок, некоторое количество укороченных продуктов (возможно, гидролизованный слитый белок) и, возможно, некоторое количество альбуминов, элюированных совместно. На фиг. 7 изображены хроматограммы, показывающие характерное время удержания стандарта человеческого инсулина по сравнению с элюированным расщепленным трипсином 4404 слитым белком на колонке С18. Стандарт hIN представляет собой стандарт рекомбинантного человеческого инсулина (0,5 мкг). На фиг. 8 изображен масс-спектральный анализ стандарта человеческого инсулина (А) по сравнению с расщепленными трипсином и очищенными ВЭЖХ 4404 (В) фракциями, собранными через 17,0-17,5 мин. На фиг. 9 изображен анализ окрашенного кумасси SDS-PAGE (15%) общего экстрагированного белка семян и белка, полученного из масляного тельца (ОВ) из колоний, экспрессирующих 4405 по сравнению с диким типом (не рекомбинантным) семян. Стрелка указывает положение мигрирующего слитого белка. На фиг. 10 изображена хроматограмма характерного времени удержания стандарта человеческого инсулина по сравнению с расщепленным трипсином препаратом 4405 ОВ, полученная обращенно-фазовой ВЭЖХ на колонке С18. Стандарт hIN представляет собой стандарт рекомбинантного человеческого инсулина (0,5 мкг). На фиг. 11 изображен масс-спектральный анализ стандарта человеческого инсулина (А) по сравнению с расщепленными трипсином очищенными ВЭЖХ 4405 (В) фракциями, собранными через 17,0-17,5 мин. На фиг. 12 изображена хроматограмма расщепленного трипсином препарата 4405 масляного тельца (пунктирная линия) по сравнению со стандартом человеческого инсулина (сплошная линия). Фракции элюированного расщепленного инсулина собирают между 7-35 мС/см и концентрируют лиофилизацией для биологического исследования инсулина. На фиг. 13 изображены изменения уровней глюкозы в сыворотке у самцов мышей В6 после инъекций отрицательного контроля (пустые круги = плацебо с физиологическим раствором, сплошные круги = расщепленные трипсином масляные тельца дикого типа), положительного контроля (пустые квадраты = Humulin R, пустые треугольники = Roche hIN) по сравнению с инсулином, полученным из растений из 4405 масляных телец (сплошные ромбы = SDS hIN DesB 30 ). На фиг. 14 изображено исследование с окрашенным кумасси гелем белков масляных телец от двух характерных колоний (4409-6 и 4409-8) по сравнению с миграцией слитого белка Oleosin-hPIN (показано черными стрелками) для нетрансформированного (wt) Arabidopsis. Уровни экспрессии определяют денситометрией с получением средних значений около 0,10% от общего белка семян. Этот уровень оценивают свыше и за пределами комиграции эндогенного белка с той же молекулярной массой из нетрансформированных семян (wt), который составляет приблизительно 0,04% от общего белка семян. Как указано выше, данное изобретение относится к улучшенному способу получения инсулина в трансгенных растениях. Авторы данного изобретения неожиданно обнаружили, что уровни аккумуляции инсулина, превышающие 0,1% от общего белка клетки, могут быть получены в растениях путем рекомбинантного получения инсулина в семенах растений. Такие уровни экспрессии, которые по крайней мере в десять раз больше тех, которые были получены ранее, позволяют коммерческое производство инсулина в растениях. Получение в семенах обеспечивает гибкость при хранении и транспортировке инсулина в виде сырья, так как инсулин сохраняет свою активность при экстрагировании из хранившихся семян. Более того, количество биомассы, которую необходимо подвергнуть экстрагированию, ограничено из-за относительно низкого содержания воды в семенах растений. Следовательно, в данном изобретении представлен способ экспрессии инсулина в растениях, включающий: (a) получение конструкции химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений; и (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения и (c) выращивание клетки растений в зрелом растении, способном давать семена, где семена экспрессируют инсулин. В соответствии с данным изобретением было неожиданно обнаружено, что инсулин аккумулируется в семенах растений в количествах, которые ранее не достигались, если инсулин экспрессируется в семенах таким образом, который позволяет изоляцию полипептида инсулина внутри клеток семян в мембране, окружающей внутриклеточный компартмент. Следовательно, в данном изобретении представлен способ экспрессии инсулина в растениях, который включает: (a) получение конструкции химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений; и (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; и (iii) последовательность нуклеиновой кислоты, кодирующую полипептид, способный удерживать полипептид инсулина в мембране, окружающей внутриклеточный компартмент; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения и (c) выращивание клетки растений в зрелом растении, способном давать семена, где семена экспрессируют инсулин. Если не указано иначе, все технические и научные термины, применяемые в данном описании, имеют значения, общепринятые среди специалистов в данной области техники, к которой принадлежит данное изобретение. Где это можно, все патенты, заявки, опубликованные заявки и другие публикации, включая последовательности нуклеиновых кислот и полипептидов из GenBank, SwissPro и других баз данных, упомянутых в данном описании, представлены здесь в качестве ссылок во всей полноте. Термин "последовательность нуклеиновой кислоты" в данном описании относится к последовательности нуклеозидных или нуклеотидных мономеров, состоящей из оснований, сахаров и межсахарных (скелетных) связей. Термин также включает модифицированные или замещенные последовательности, включающие не существующие в природе мономеры или их части. Последовательности нуклеиновых кислот в соответствии с данным изобретением могут быть последовательностями дезоксирибонуклеиновой кислоты (ДНК) или последовательностями рибонуклеиновой кислоты (РНК) и могут включать существующие в природе основания, включая аденин, гуанин, цитозин, тимидин и урацил. Последовательности также могут содержать модифицированные основания. Примеры таких модифицированных оснований включают аза- и деаза- аденин, гуанин, цитозин, тимидин и урацил; и ксантин и гипоксантин. Термин "последовательность нуклеиновой кислоты, кодирующая инсулин" и "последовательность нуклеиновой кислоты, кодирующая полипептид инсулина", которые могут быть взаимозаменяемыми, относятся к любой и всем последовательностям нуклеиновых кислот, кодирующим полипептид инсулина, включая полипептиды инсулина, перечисленные в таблице (SEQ ID NO:7-145), а также любой полипептид инсулина млекопитающего, и к любым последовательностям нуклеиновых кислот, которые кодируют проинсулин и препроинсулин. В данном описании "проинсулин" относится к полипептиду инсулина, который включает связывающий пептид или "С-пептид", связывающий А и В цепи полипептида инсулина. В природном человеческом инсулине С-пептид представляет собой цепь из 31 остатка аминокислоты, связывающую остаток В30 с остатком А1. Термин "препроинсулин" относится к молекуле проинсулина, дополнительно содержащей N-концевую сигнальную последовательность, которая направляет возникновение трансляции в ER рибосомах. Последовательности нуклеиновых кислот, кодирующие полипептид инсулина, также включают любую и все последовательности нуклеиновых кислот, которые (i) кодируют полипептиды, которые практически идентичны последовательности полипептида инсулина, представленной выше; или (ii) гибридизируются с любой последовательностью нуклеиновой кислоты, указанной выше, в условиях, по крайней мере, умеренно жесткой гибридизации или которые будут гибридизироваться, по крайней мере, в умеренно жестких условиях, но для применения синонимичных кодонов. Под термином "практически идентичный" понимают то, что две полипептидные последовательности предпочтительно идентичны по крайней мере на 75%, более предпочтительно по крайней мере на 85% и наиболее предпочтительно идентичны по крайней мере на 95%, например идентичны на 96, 97, 98 или 99%. Для определения процента идентичности между двумя полипептидными последовательностями последовательности аминокислот двух таких последовательностей совмещают предпочтительно с применением алгоритма Clustal W (Thompson, J.D., Higgins D.G., Gibson T.J., 1994, Nucleic Acids Res. 22(22): 4673-4680) вместе с матрицей количественной оценки BLOSUM 62 (Henikoff S., and Henikoff J.G., 1992, Proc. Natl. Acad. Sci. USA 89: 10915-10919) и штрафом раскрытия интервала 10 и штрафом протяженности интервала 0,1 таким образом, что совпадение высшего порядка достигается между двумя последовательностями, где по крайней мере 50% от общей длины одной из последовательностей включено в выравнивание. Другие способы, которые могут применяться для выравнивания последовательностей, включают способ выравнивания Нидльмана и Вунша (J. Mol. Biol., 1970, 48: 443), исправленный Смитом и Ватерманом (Adv. Appl. Math., 1981, 2: 482) таким образом, что совпадение высшего порядка получают между двумя последовательностями и между двумя последовательностями определяют количество идентичных аминокислот. Другие методы расчета процента совпадения между двумя аминокислотными последовательностями известны в данной области техники и включают, например, описанные Carillo и Lipton (SIAM J. Applied Math., 1998, 48: 1073) и описанные в Computational Molecular Biology, Lesk, e.d. Oxford University Press, New York, 1988, Biocomputing: Informatics and Genomics Projects). В основном для таких расчетов применяют программное обеспечение. Программное обеспечение, которое может применяться для этой цели, включает, но не ограничено ими, GCG (Devereux et al., Nucleic Acids Res., 1984, 12: 387), BLASTP, BLASTN и FASTA (Altschul et al., J. Molec. Biol., 1990: 215: 403). Под "по крайней мере, умеренно жесткими условиями гибридизации" понимают условия, которые способствуют селективной гибридизации между двумя комплементарными молекулами нуклеиновой кислоты в растворе. Гибридизация может происходить со всей или с частью последовательности молекулы нуклеиновой кислоты. Гибридизируемая часть обычно содержит по крайней мере 15 (например, 20, 25, 30, 40 или 50) нуклеотидов. Специалист в данной области техники поймет, что стабильность дуплекса или гибрида нуклеиновой кислоты определяется T m , которая в буферах, содержащих натрий, является функцией концентрации иона натрия и температуры (T m =81,5 °С-16,6(Log 10 [Na + ])+0,41(%(G+C)-600/1) или подобное уравнение). Следовательно, параметры условий промывания, которые определяют стабильность гибрида, включают концентрацию иона натрия и температуру. Для идентификации молекул, которые сходны, но не идентичны, известной молекуле нуклеиновой кислоты, приемлемо 1% несовпадение, что дает снижение около 1 °С в T m , например, если считается, что молекулы нуклеиновой кислоты идентичны на > 95%, температура конечной промывки будет снижена на около 5 °С. Основываясь на этих соображениях специалист в данной области техники легко подберет подходящие условия гибридизации. В предпочтительных вариантах выбирают жесткие условия гибридизации. Например, следующие условия могут применяться для получения жесткой гибридизации: гибридизация в 5 × хлориде натрия/цитрате натрия (SSC)/5 × раствора Денхардта/0,1% SDS при T m -5 °C на основании представленного выше уравнения, с последующей промывкой 0,2 × SSC/0,1% SDS при температуре 60 °С. Умеренно жесткие условия гибридизации включают промывку 3 × SSC при температуре 42 °С. Однако понятно, что эквивалентные ограничения могут быть достигнуты с применением альтернативных буферов, солей и температур. Дополнительные инструкции по проведению гибридизации можно найти в Current Protocols in Molecular Biology, John Wiley & Sons, N.Y., 1989, 6.3.1-6.3.6 и в Sambrook et al., Molecular Cloning, a Laboratory manual, Cold Spring Harbor Laboratory Press, 1989, vol. 3. В данном описании термины "инсулин" и "полипептид инсулина", которые могут быть взаимозаменяемыми, относятся к любому и всем полипептидам инсулина, включая полипептиды инсулина, перечисленные в таблице (SEQ ID NO:7-145), а также к молекуле полипептида, содержащей последовательность остатков аминокислот, которые (i) практически идентичны последовательностям аминокислот, составляющих любой из представленных здесь полипептидов инсулина, или (ii) кодированной последовательности нуклеиновой кислоты, способной гибридизироваться, по крайней мере, в умеренно жестких условиях с любой последовательностью нуклеиновой кислоты, кодирующей представленный здесь инсулин, или способной гибридизироваться, по крайней мере, в умеренно жестких условиях с любой последовательностью нуклеиновой кислоты, кодирующей представленный здесь инсулин, но для применения синонимичных кодонов. Термины "инсулин" и "полипептид инсулина" включают полипептиды проинсулина и полипептиды мини-инсулина. Полипептид инсулина предпочтительно имеет человеческое, бычье или свиное происхождение. Термин "полипептид, способный сохранять полипептид инсулина в мембране, окружающей внутриклеточный компартмент" в данном описании относится к любому полипептиду, который, будучи связанным с полипептидом инсулина, способен изолировать полипептид инсулина во внутриклеточной структуре, окруженной мембраной, и располагаться во внутриклеточном пространстве клетки растения, как определено плазматической мембраной клетки растения. Термин "полипептид, способный удерживать полипептид инсулина в ER или ER-производной накопительной везикуле" относится к любому полипептиду, который, будучи связанным с полипептидом инсулина, способен изолировать полипептид инсулина либо в эндоплазматической сети, либо в накопительном компартменте, который является производным из эндоплазматической сети, таком как, например, масляное тельце, в клетке растения. Термин "масляное тельце" или "масляные тельца" относится к любой органелле, накапливающей масло или жир, в клетке семени растения (описаны, например, в Huang (1992), Ann. Rev. Plant Mol. Biol., 43: 177-200). Термин "химерная", применяемый в данном описании в контексте с последовательностью нуклеиновой кислоты, относится по крайней мере к двум связанным последовательностям нуклеиновых кислот, которые не связаны в природе. Химерные последовательности нуклеиновых кислот включают связанные последовательности нуклеиновых кислот различного природного происхождения. Например, последовательность нуклеиновой кислоты, содержащаяся в промоторе растения, связанная с последовательностью нуклеиновой кислоты, кодирующей человеческий инсулин, считается химерной. Химерные последовательности нуклеиновых кислот также могут содержать последовательности нуклеиновой кислоты одного и того же природного происхождения при условии, что они не связаны в природе. Например, последовательность нуклеиновой кислоты, содержащаяся в промоторе, полученном из определенного типа клеток, может быть связана с последовательностью нуклеиновой кислоты, кодирующей полипептид, полученный из того же типа клеток, но который в природе не связан с последовательностью нуклеиновой кислоты, содержащейся в промоторе. Химерные последовательности нуклеиновых кислот также включают последовательности нуклеиновых кислот, содержащие любые природные последовательности нуклеиновых кислот, связанные с любыми не существующими в природе последовательностями нуклеиновых кислот. Последовательности нуклеиновых кислот, кодирующие инсулин, которые могут применяться в соответствии со способами и композициями, представленными в данном описании, могут включать любые последовательности нуклеиновых кислот, кодирующие полипептид инсулина, включая любой проинсулин и препроинсулин. Примеры последовательностей нуклеиновых кислот, кодирующих инсулин, хорошо известны в данной области техники и обычно легко доступны из множества различных источников млекопитающих, включая человека (Bell, G.I. et al., 1980, Nature 284: 26-32), свинью (Chance, R.E. et al., 1968, Science 161: 165-167), быка (D'Agostino, J. et al., 1987, Mol. Endocrinol. 1: 327-331), овцу (Peterson, J.D. et al., 1972, Biol. Chem. 247: 4866-4671) и подобные, а также из растительных источников (Oliveira, A.E.A. et al., 1999, Protein Pept. Lett. 6: 15-21). Последовательности, кодирующие инсулин, которые могут применяться, включают кодирующие полипептидные цепи, представленные как SEQ ID NO:7-145. Соответствующие последовательности нуклеиновых кислот, кодирующие полипептидные цепи инсулина, могут быть легко идентифицированы с помощью номеров идентификатора Swiss Protein, представленных в таблице. Применяя эти последовательности нуклеиновых кислот, дополнительные новые последовательности нуклеиновых кислот, кодирующие инсулин, могут быть легко идентифицированы с применением методик, известных специалистам в данной области техники. Например, библиотеки, такие как экспрессионные библиотеки, кДНК и геномные библиотеки, могут быть исследованы и базы данных, содержащие информацию о последовательностях из проектов, устанавливающих последовательность, могут быть проверены на предмет схожих последовательностей. Могут применяться альтернативные методы выделения дополнительных последовательностей нуклеиновых кислот, кодирующих полипептид инсулина, и новые последовательности могут быть обнаружены и использованы в соответствии с данным изобретением. В предпочтительных вариантах последовательности нуклеиновых кислот, кодирующие инсулин, включают человеческий, свиной и бычий инсулин. Множество аналогов инсулина известно в данной области техники (см., например, патенты США 5461031, 5474987, 5164366 и 5008241) и могут применяться в соответствии с данным изобретением. Аналоги, которые могут применяться, включают молекулы человеческого инсулина, в которых остаток аминокислоты 28 цепи В (В28) изменен из природного пролинового остатка на аспартат, лизин или изолейцин. В другом варианте лизиновый остаток В29 модифицирован в пролин. Далее, аспарагин А21 может быть заменен на аланин, глутамин, глутамат, глицин, гистидин, изолейцин, лейцин, метионин, серин, треонин, триптофан, тирозин или валин. Также аспарагин В3 может быть модифицирован в лизин. Другие примеры аналогов инсулина, которые могут применяться в соответствии с данным изобретением, включают человеческий инсулин, не содержащий остатка В30, также часто называемый "desB30" или "В(1-29)"; инсулин, не содержащий последние 3 аминокислотных остатка "В(1-27)"; инсулин, не содержащий фенилаланиновый остаток В1; и аналоги, в которых А-цепь или В-цепь имеет N-концевое или С-концевое удлинение, например В-цепь может быть продлена на N-конце путем добавления двух аргининовых остатков. В предпочтительных вариантах последовательность нуклеиновой кислоты, кодирующая инсулин, представляет собой проинсулин. В других предпочтительных вариантах применяют последовательность нуклеиновой кислоты, кодирующую инсулин, в которой С-пептид модифицирован относительно его природной формы. Остатки аминокислот в С-пептиде могут быть замещены и С-пептид может быть удлинен или укорочен. В этом случае термин "мини-инсулин" относится к полипептиду инсулина, который был модифицирован таким образом, что С-пептид был укорочен относительно его природной формы. В предпочтительных вариантах применяют мини-инсулин. Предпочтительно С-пептид молекулы мини-инсулина короче чем 20 остатков аминокислот, более предпочтительно короче чем 15 остатков аминокислот и наиболее предпочтительно короче чем 9 остатков аминокислот, например 7, 5 или 3 остатка. Предпочтительно, как и в случае с природными молекулами инсулина, С-пептид мини-инсулина включает сайт расщепления на его С- и N-концах. Такие сайты расщепления могут представлять собой любые удобные сайты расщепления, известные в данной области техники, например метионин, расщепляемый бромидом цианогена, единичный основный остаток или пара основных остатков, расщепляемых трипсином или протеазами, подобными трипсину, или карбоксипептидазой. Например, С-пептид может содержать лизин на С-конце, например, Ala-Ala-Lys (SEQ ID NO:146) или двухосновный сайт обработки непосредственно перед остатком GlyA1, например, Asn-Lys-Arg (SEQ ID NO:147) или Arg-Arg-Lys-Gln-Lys-Arg (SEQ ID NO:148) или тетраосновный сайт обработки непосредственно перед остатком GlyA1, например, Arg-Arg-Lys-Arg (SEQ ID NO:149). Следовательно, молекулы мини-инсулина, которые могут применяться в соответствии с данным изобретением, включают где X 1 является аминокислотой; Х 2 является аминокислотой; Х 3 является Lys или Arg; Y 1 является пептидной связью или 1-17 остатками аминокислот; В(1-29/30) является В-цепью человеческого инсулина, содержащей остатки аминокислот 1-29 или 1-30; и А(1-21) является А-цепью человеческого инсулина, содержащей остатки аминокислот 1-21. В предпочтительном варианте X 1 является остатком основной аминокислоты (Lys или Arg) и Y 1 является либо пептидной связью, либо 1-17 остатками аминокислот, где С-концевой остаток является остатком основной аминокислоты (Lys или Arg). Далее, молекулы мини-инсулина, которые могут применяться в соответствии с данным изобретением, включают те, которые представлены формулой где X 1 является пептидом из 1-8 остатков аминокислот, включающим по крайней мере один остаток ароматической аминокислоты; Х 2 является одним из Pro, Asp, Lys или Ile в положении 28 В-цепи; Х 3 является одним из Pro, Lys, Ala, Arg или Pro-Thr в положении 29 В-цепи; Y является Lys или Arg; В(1-27) является В-цепью человеческого инсулина, содержащей остатки аминокислот 1-27; и А(1-21) является А-цепью человеческого инсулина, содержащей остатки аминокислот 1-21. Другие примеры молекул нуклеиновых кислот, кодирующих полипептиды мини-инсулина, которые могут применяться в соответствии с данным изобретением, включают описанные в Markussen et al., Walter de Gruyter & Co. 1987, in: Peptides, p. 189-194; Thim et al., 1989, in: Genetics and molecular biology of industrial microorganisms, American Society for Microbiology, p. 322-328; и те, которые представлены в патентах США 4916212, 5324641 и 6521738. Изменения последовательности нуклеиновой кислоты, кодирующей инсулин, для получения аналогов инсулина могут быть произведены с применением множества методик модификации нуклеиновых кислот, известных специалистам в данной области техники, включая, например, сайт-направленный мутагенез, целевой мутагенез, произвольный мутагенез, добавление органических растворителей, перестановку генов или сочетание этих методов, и другие методики, известные специалистам в данной области техники (Shraishi et al., 1988, Arch. Biochem. Biophys., 358: 104-115; Galkin et al., 1997, Protein Eng. 10: 687-690; Carugo et al., 1997, Proteins 28: 10-28; Hurley et al., 1996, Biochem., 35: 5670-5678; Holmberg et al., 1999, Protein Eng. 12: 851-856). В соответствии с данным изобретением неожиданно было обнаружено, что инсулин аккумулируется в семенах растений в количествах, которые до сих пор не были получены, если инсулин экспрессируется в семенах, предпочтительно таким образом, что полипептид инсулина внутри клетки семени изолирован в мембране, окружающей внутриклеточный компартмент. В предпочтительных вариантах данного изобретения полипептид инсулина изолирован в ER или ER-производной накопительной везикуле. Для достижения такого накопления инсулина в ER или ER-производной накопительной везикуле в соответствии с данным изобретением полипептид, кодирующий инсулин, связан с полипептидом, который удерживает полипептид инсулина в ER или ER-производной накопительной везикуле, вместо транспортировки из ER, например, в апопласт. Полипептиды, которые могут применяться в соответствии с данным изобретением для удержания полипептида инсулина в ER, включают любые полипептиды, способные изолировать инсулин в ER. Такие полипептиды могут быть синтезированы или получены из любых биологических источников. В предпочтительном варианте данного изобретения полипептиды, которые способны удерживать инсулин, включают полипептиды, содержащие С-концевой мотив удержания в ER. Примеры таких С-концевых мотивов удержания в ER включают KDEL, HDEL, DDEL, ADEL и SDEL последовательности (SEQ ID NO:150-154 соответственно). Другие примеры включают HDEF (SEQ ID NO:155) (Lehmann et al., 2001, Plant Physiol. 127(2): 436-49) или два аргининовых остатка, близких к N-концу, расположенные в положениях 2 и 3, 3 и 4 или 4 и 5 (Abstract from Plant Biology 2001 Program, ASPB, July 2001, Providence, Rhode Island, USA). Последовательности нуклеиновых кислот, кодирующие С-концевой мотив ER удержания, предпочтительно связаны с последовательностью нуклеиновой кислоты, кодирующей полипептид инсулина таким образом, что полипептид, способный удерживать инсулин в ER, связан с С-концом полипептида инсулина. Для достижения накопления полипептида инсулина в ER-производной накопительной везикуле полипептид инсулина связан с полипептидом, который способен удерживать полипептид инсулина в ER-производной накопительной везикуле. Полипептид, способный удерживать полипептид инсулина в ER-производной накопительной везикуле, который может применяться в соответствии с данным изобретением, включает любой полипептид, способный изолировать полипептид инсулина в ER-производной накопительной везикуле. Полипептиды, способные удерживать инсулин в ER-производной накопительной везикуле, могут быть синтезированы или получены из любого биологического источника. В предпочтительном варианте ER-производная накопительная везикула представляет собой масляное тельце, и полипептид инсулина связан с белком масляного тельца или его значительной частью, способной удерживать полипептид инсулина в ER-производной накопительной везикуле. Белки масляного тельца, которые могут применяться в соответствии с данным изобретением, включают любые белки, которые естественным образом связаны с масляным тельцем. Особенно предпочтительные белки масляного тельца включают олеозины, например олеозин Arabidopsis (van Rooijen et al., (1991), Plant Mol Biol. 18: 1177-1179), кукурузный олеозин (Bowman-Vance et al., 1987, J. Biol. Chem. 262: 11275-11279; Qu et al., 1990, J. Biol. Chem. 265: 2238-2243), олеозин моркови (Hatzopoulos et al., (1990), Plant Cell 2: 457) или олеозин Brassica (Lee et al., 1991, Plant Physiol. 96: 1395-1397), калеозины (см., например, Genbank, номер доступа AF067857) и стеролеозины (Lin et al., 2002, Plant Physiol. 128(4): 1200-11). В предпочтительном варианте белком масляного тельца является растительный олеозин, который имеет сходство последовательности с другими растительными олеозинами, такими как олеозин, выделенный из Arabidopsis thaliana (SEQ ID NO:156) или Brassica napus (SEQ ID NO:157). В другом варианте белком масляного тельца является калеозин или связывающий кальций белок из растительного, грибного или другого источника, который отличается гомологией последовательности с растительными калеозинами, такими как калеозин, выделенный из Arabidopsis thaliana (SEQ ID NO:158 и SEQ ID NO:159). В другом варианте белком масляного тельца является стеролеозин (SEQ ID NO:160), дегидрогеназа, связывающая стерин (Lin L.-J. et al., (2002), Plant Physiol. 128: 1200-1211). Полипептид, кодирующий инсулин, может быть связан с белком масляного тельца через N-конец, так же как и через С-конец, и через фрагменты белка масляного тельца, такие как, например, центральный домен олеозина. Новые белки масляного тельца могут быть обнаружены, например, при получении масляных телец (методики получения масляных телец см., например, в патенте США 6650554) и идентификации белков в препаратах масляных телец с помощью, например, SDS электрофореза в геле. Против таких белков могут быть получены поликлональные антитела, которые применяют для скрининга библиотек кДНК для идентификации последовательностей нуклеиновых кислот, кодирующих белки масляных телец. Новые белки масляных телец также могут быть получены с применением известных последовательностей нуклеиновых кислот, кодирующих белки масляных телец, с применением, например, указанных здесь последовательностей белков масляных телец, кодирующих белки масляных телец, для исследования, например, кДНК или геномных библиотек на присутствие белков масляных телец. Полипептиды, способные удерживать инсулин в ER или ER-производной накопительной органелле, обычно не отщепляются, и инсулин может накапливаться в виде слитого белка, что, например, обычно происходит, если KDEL сигнал удержания применяется для удержания полипептида в ER или если белок масляного тельца применяется для удержания полипептида в ER-производной накопительной органелле. Химерная последовательность нуклеиновой кислоты дополнительно может содержать нуклеотидную последовательность, которая нацеливает последовательность нуклеиновой кислоты на эндомембранную систему ("сигнальный пептид"). В вариантах данного изобретения, в которых полипептид инсулина удерживается в ER с помощью последовательности, способной удерживать полипептид в ER, такой как KDEL, HDEL или SDEL полипептид, особенно желательно включать последовательность нуклеиновой кислоты, кодирующую сигнальный пептид. Примеры сигнальных пептидов, которые могут применяться в соответствии с данным изобретением, включают сигнальную последовательность белка, связанного с патогенезом табака (PR-S) (SEQ ID NO:161) (Sijmons et al., 1990, Bio/technology, 8: 217-221), сигнальную последовательность лектина (Boehn et al., 2000, Transgenic Res, 9(6): 477-86), сигнальную последовательность насыщенного гидроксипролином гликопротеина из Phaseolus vulgaris (Yan et al., 1997, Plant Physiol. 115(3): 915-24 и Corbin et al., 1987, Mol. Cell Biol. 7(12): 4337-44), сигнальную последовательность пататина картофеля (Iturriaga, G. et al., 1989, Plant Cell 1: 381-390 и Beyan et al., 1986, Nuc. Acids Res. 41: 4625-4638) и сигнальную последовательность альфа-амилазы ячменя (Rasmissen and Johansson, 1992, Plant Mol. Biol. 18(2): 423-7). Такие целевые сигналы могут быть in vivo отщеплены от последовательности инсулина, что обычно происходит когда применяется целевой сигнал апопласта, такой как сигнальная последовательность S-белка, связанного с патогенезом табака (PR-S) (Sijmons et al., 1990, Bio/technology, 8: 217-221). Другие сигнальные пептиды могут быть определены с помощью сервера SignalP World Wide Web (http://www.cbs.dtu.dk/services/SignalP), в котором можно определить присутствие и расположение сайтов отщепления сигнального пептида в последовательностях аминокислот различных организмов. В общем, существует незначительное сохранение первичной последовательности аминокислоты, хотя общие физико-химические свойства сохраняются до некоторого предела. Общая структура сигнальных пептидов имеет три области, короткую аминоконцевую "n-область", содержащую положительно заряженные остатки, центральную гидрофобную "h-область", изменяющуюся в размере от 7 до 15 аминокислот, и карбоксиконцевую "с-область", содержащую полярные аминокислоты и сайт расщепления, который распознается мембранно-связанными сигнальными ферментами пептидазы (Nakai K., 2000, Advances in Protein Chem. 54: 277-344). Целевой сигнал, который также может применяться в соответствии с данным изобретением, включает природную сигнальную последовательность инсулина (длина человеческой последовательности составляет 24 аминокислоты). В предпочтительных вариантах расположенную на N-конце целевую последовательность апопласта, такую как указанная выше PR-S последовательность табака, применяют в сочетании с расположенной на С-конце удерживающей в ER последовательностью, такой как последовательность KDEL. В другом предпочтительном варианте последовательность нуклеиновой кислоты, кодирующая лидерную последовательность α-фактора дрожжей, связана с N-концом последовательности нуклеиновой кислоты, кодирующей инсулин. Лидерные последовательности дрожжей или последовательности, полученные из лидерных последовательностей дрожжей, которые могут применяться в соответствии с данным изобретением, включают последовательности, перечисленные в SEQ ID NO:162-171 (Kjeldsen et al., 2001, Biotechnilogy and Genetic Engineering Reviews 18: 89-121). Такие лидерные последовательности могут также содержать пептид спейсера, расположенный на С-конце нуклеиновой кислоты, кодирующей лидерную последовательность, и на N-конце последовательности, кодирующей инсулин. В соответствии с данным изобретением такие последовательности спейсеров обычно имеют от 2 до 20 аминокислот в длину. Таким образом, например, могут применяться последовательности спейсеров SEQ ID NO:172 и SEQ ID NO:173 (Kjeldsen et al., 2001, Biotechnilogy and Genetic Engineering Reviews 18: 89-121). В вариантах данного изобретения, в которых применяется лидерная последовательность дрожжей, последовательность нуклеиновой кислоты, кодирующая полипептид инсулина, предпочтительно представляет собой полипептид мини-инсулина. В соответствии с данным изобретением в особенно предпочтительном варианте применяют последовательность нуклеиновой кислоты, кодирующую антитело с одной цепью, связанной с последовательностью нуклеиновой кислоты, кодирующей лидерный пептид секреции дрожжей, как далее описано в примере 1. Химерная последовательность нуклеиновой кислоты также может содержать полипептиды, дающие стабилизирующие удлинения белка на N- и/или С-концах. Такие удлинения могут применяться для стабилизации и/или помогают свертыванию цепи полипептида инсулина и дополнительно могут применяться для облегчения очистки инсулина. Полипептидные удлинения, которые могут применяться для этой цели, включают, например, последовательность нуклеиновой кислоты, кодирующую антитело с одной цепью, нуклеиновую кислоту, кодирующую молекулу Affibody ® (Affibody AB), последовательность нуклеиновой кислоты, кодирующую нетоксичную В субъединицу токсина холеры (СТВ) (Arakawa, Т. et al., 1998, Nat. Biotechnol. 16: 938) или сочетания таких полипептидов. В особенно предпочтительном варианте полипептид инсулина удерживается в мембране, окружающей компартмент, такой как ER, с помощью, например, последовательности KDEL, как описано выше, объединенной со стабилизирующим полипептидом, который позволяет объединение полипептида инсулина с масляным тельцем при нарушении целостности клетки растения, что может произойти, если полипептид инсулина выделяют из клетки растения. Примером такого стабилизирующего полипептида является антитело с одной цепью со специфичностью к масляному тельцу. Последовательности нуклеиновых кислот, кодирующие антитела с одной цепью со специфичностью к масляному тельцу, могут быть получены из колонии клеток гибридомы, экспрессирующих моноклональные антитела, действующие против белка масляного тельца. В одном варианте антитело с одной цепью специфически связывается с олеозином, как описано в Alting-Mees et al. (2000), IBC's International Conference on Antibody Engineering, Poster #1. Этот вариант данного изобретения более подробно описан в примере 1 ниже. В другом варианте сайт расщепления может быть расположен выше N-конца и ниже С-конца инсулина, что позволяет полипептиду инсулина отщепляться от партнера слияния, тем самым получая выделенный инсулин. Примеры таких сайтов расщепления могут быть найдены в WO 98/49326 (способ расщепления слитых белков) и родственных заявках, и у LaVille et al. (1994), Enzymatic and chemical cleavage of fusion proteins In Current Protocols in Molecular Biology, p. 16.4.5-16.4.17, John Wiley and Sons, Inc., New York NY. В предпочтительном варианте сайтом расщепления является тетраосновный линкер (например, Arg-Arg-Lys-Arg - SEQ ID NO:149), который расщепляется трипсином. В другом предпочтительном варианте сайтом расщепления является Klip8 (SEQ ID NO:174), который расщепляется аспартат протеазой, включая химозин. В данном изобретении также представлены способы отделения гетерологичных белков от компонентов клетки-хозяина разделением фракции масляного тельца и последующим выделением гетерологичного белка путем специфического расщепления гетерологичного белка - слиянием белка масляного тельца. Необязательно сайт расщепления может быть расположен выше N-конца и ниже С-конца гетерологичного полипептида, что позволяет слитому полипептиду расщепляться и отделяться путем разделения фаз на составляющие их пептиды. Последовательность нуклеиновой кислоты, кодирующая инсулин, может быть изменена для дальнейшего улучшения уровней экспрессии, например, оптимизацией последовательности нуклеиновой кислоты в соответствии с предпочтительным применением кодона для определенного типа клетки растения, которую выбирают для экспрессии полипептида инсулина, или изменением мотивов, которые известны как дестабилизирующие мРНК (см., например, заявку на патент РСТ 97/02352). Сравнение применения кодона последовательности нуклеиновой кислоты, кодирующей полипептид инсулина, и применения кодона клетки растения позволяют идентификацию кодонов, которые могут быть изменены. Построение синтетических генов путем изменения применения кодона описано, например, в заявке на патент РСТ 93/07278. В предпочтительном варианте применяемая последовательность нуклеиновой кислоты, кодирующая инсулин, представлена SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:195. В соответствии с данным изобретением последовательность нуклеиновой кислоты, кодирующая инсулин, связана с промотором, способным контролировать экспрессию полипептида инсулина в клетке семени растения. Следовательно, в данном изобретении также представлена последовательность нуклеиновой кислоты, кодирующая инсулин, связанная с промотором, способным контролировать экспрессию в клетке семени растения. Применяемые здесь промоторы обычно являются известными в данной области техники промоторами и включают любой промотор растительного происхождения, способный контролировать экспрессию полипептидов в растениях. В общем, промоторы, полученные из двудольных растений, применяют, если двудольное растение выбирают в соответствии с данным изобретением, а промотор однодольного растения используют, если выбирают однодольное растение. Основные промоторы, которые могут применяться, включают, например, промотор вируса мозаики цветной капусты 35S (CaMV) (Rothstein et al., 1987, Gene 53: 153-161), промотор актина риса (McElroy et al., 1990, Plant Cell 2: 163-171; патент США 6429357), промотор убиквитина, такой как промотор убиквитина кукурузы (патенты США 5879903; 5273894) и промотор убиквитина петрушки (Kawalleck, P. et al., 1993, Plant Mol. Biol. 21: 673-684). В предпочтительных вариантах применяемым промотором является промотор, который дает предпочтительную экспрессию полипептида инсулина в ткани семени. "Предпочтительными для семян промоторами" в этом смысле являются промоторы, которые контролируют экспрессию рекомбинантного белка (а именно, инсулина) таким образом, что предпочтительно по крайней мере 80% от общего количества рекомбинантного белка, присутствующего во взрослом растении, присутствовало в семени. Более предпочтительно по крайней мере 90% от общего количества рекомбинантного белка, присутствующего во взрослом растении, должно присутствовать в семени. Наиболее предпочтительно по крайней мере 95% от общего количества рекомбинантного белка, присутствующего во взрослом растении, должно присутствовать в семени. Предпочтительные для семян промоторы, которые могут применяться в этом смысле, включают, например, промотор фазеолина бобов (Sengupta-Gopalan et al., 1985, Proc. Natl. Acad. Sci. USA 82: 3320-3324); промотор олеозина Arabidopsis 18 кДа (патент США 5792922) или промотор олеозина льна (WO 01/16340); промотор аккумулирующего белка (линина) семени льна, подобного легумину (WO 01/16340); промотор аккумулирующего белка 2S льна (WO 01/16340); предпочтительный промотор эндосперма, такой как промотор Amy32b (Rogers and Milliman, J. Biol. Chem., 1984, 259: 12234-12240), промотор Amyb-4 (Kursheed and Rogers, J. Biol. Chem., 1988, 263: 18953-18960), или промотор Алеураина (Whittier et al., 1987, Nucleic Acids Res., 15: 2515-2535), или промотор арселина бобов (Jaeger G.D., et al., 2002, Nat. Biotechnol. Dec; 20: 1265-8). Постоянно находят новые промоторы, применяемые в различных растениях. Множество примеров промоторов растений можно найти у Ohamuro et al. (Biochem. of Plants., 1989, 15: 1-82). В данном изобретении могут применяться определенные генетические элементы, способные улучшать экспрессию полипептида инсулина. Эти элементы включают нетранслированные лидерные последовательности определенных вирусов, такие как лидерная последовательность AMV (Jobling and Gehrke, 1987, Nature, 325: 622-625) и интрон, связанный с промотором убиквитина кукурузы (патент США 5504200). Обычно химерную последовательность нуклеиновой кислоты получают таким образом, чтобы генетические элементы, способные усиливать экспрессию, были расположены в положении 5' последовательности нуклеиновой кислоты, кодирующей полипептид инсулина. В соответствии с данным изобретением химерные последовательности нуклеиновых кислот, содержащие промотор, способный контролировать экспрессию в семенах растений, связанный с последовательностью нуклеиновой кислоты, кодирующей полипептид инсулина, могут быть интегрированы в рекомбинантный вектор экспрессии, который обеспечивает хорошую экспрессию в клетке семени. Следовательно, данное изобретение включает рекомбинантный вектор экспрессии, содержащий в направлении 5'-3' транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетки семени растения; и (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; где вектор экспрессии подходит для экспрессии в клетке растения. Термин "подходящий для экспрессии в клетке растения" означает, что рекомбинантный вектор экспрессии содержит химерную последовательность нуклеиновой кислоты в соответствии с данным изобретением, связанную с генетическими элементами, требуемыми для достижения экспрессии в клетке семени. Генетические элементы, которые могут быть включены в вектор экспрессии в этом смысле, включают транскрипционную область терминации, одну или более последовательностей нуклеиновой кислоты, кодирующую маркерные гены, одно или более начало репликации и подобные. В предпочтительных вариантах вектор экспрессии также содержит генетические элементы, требуемые для интеграции вектора или его части в ядерный геном клетки растения, например, левую и правую крайние последовательности Т-ДНК, которые способствуют интеграции в ядерный геном растения в вариантах в соответствии с данным изобретением, в которых клетки растений трансформируют с помощью Agrobacterium. Как указано выше, рекомбинантный вектор экспрессии обычно содержит транскрипционный терминатор, который, кроме того что он служит в качестве сигнала для окончания транскрипции, также может служить в качестве защитного элемента, способного продлевать период полураспада мРНК (Guarneros et al., 1982, Proc. Natl. Acad. Sci. USA, 79: 238-242). Терминатор транскрипции обычно содержит от около 200 нуклеотидов до около 1000 нуклеотидов, и вектор экспрессии получают таким образом, что терминатор транскрипции расположен в положении 3' последовательности нуклеиновой кислоты, кодирующей инсулин. Концевые последовательности, которые могут применяться в соответствии с данным изобретением, включают, например, конечную область нопалина (Bevan et al., 1983, Nucl. Acids. Res., 11: 369-385), терминатор фазеолина (van der Geest et al., 1994, Plant J. 6: 413-423), терминатор арселина (Jaeger G.D., et al., 2002, Nat. Biotechnol. Dec; 20: 1265-8), терминатор для генов октопинсинтазы Agrobacterium tumefaciens или других подобных функциональных элементов. Транскрипционные терминаторы могут быть получены, как описано у An (An, 1987, Methods in Enzym. 153: 292). В соответствии с данным изобретением вектор экспрессии также может содержать маркерный ген. Маркерные гены, которые могут применяться в соответствии с данным изобретением, включают все гены, которые позволяют отличать трансформированные клетки от нетрансформированных клеток, включая все селектируемые и скринабельные маркерные гены. Маркерный ген может быть маркером устойчивости, таким как маркер устойчивости к антибиотикам, например канамицину (патент США 6174724), ампициллину, G418, блеомицину, гигромицину, что позволяет селекцию характерных особенностей химическими методами, или маркеры, толерантные к химическому агенту, такому как сахар манноза с обычной фитотоксичностью (Negrotto et al., 2000, Plant Cell Rep. 19: 798-803). Другие подходящие маркеры, которые могут применяться в соответствии с данным изобретением, включают маркеры, способные передавать устойчивость к гербицидам, таким как глифосат (патенты США 4940935; 5188642), фосфинотрицин (патент США 5879903) или сульфонилмочевины (патент США 5633437). Маркеры устойчивости, будучи тесно связаны с последовательностью нуклеиновой кислоты, кодирующей полипептид инсулина, могут применяться для сохранения давления отбора на популяцию клеток растения или растения, которые не потеряли последовательность нуклеиновой кислоты, кодирующую полипептид инсулина. Скринабельные маркеры, которые могут применяться для идентификации трансформантов путем визуальной проверки, включают β-глюкуронидазу (GUS) (патенты США 5268463 и 5599670) и зеленый флуоресцентный белок (GFP) (Niedz et al., 1995, Plant Cell Rep., 14: 403). Рекомбинантные векторы, подходящие для введения последовательностей нуклеиновых кислот в растения, включают векторы на основе Agrobacterium и Rhizobium, такие как Ti и Ri плазмиды, включая, например, pBIN19 (Bevan, Nuci. Acid. Res., 1984, 22: 8711-8721), pGKB5 (Bouchez et al., 1993, С R Acad. Sci. Paris, Life Sciences, 316: 1188-1193), ряд бинарных векторов pCGN (McBride and Summerfelt, 1990, Plant Mol. Biol., 14: 269-276) и другие бинарные векторы (например, патент США 4940838). Рекомбинантные векторы экспрессии, последовательности нуклеиновых кислот и химерные последовательности нуклеиновых кислот в соответствии с данным изобретением могут быть получены в соответствии с методиками, хорошо известными специалистам в области молекулярной биологии. Такое получение обычно включает виды бактерий Escherichia coli в качестве промежуточного клонирующего хозяина. Получение векторов Е.coli, а также векторов трансформации растений может быть осуществлено с применением известных методик, таких как рестрикционное расщепление, лигирование, гелевый электрофорез, секвенирование ДНК, полимеразная цепная реакция (PCR) и другие методики. Эти методики позволяют связывать последовательности нуклеиновых кислот и полипептиды, относящиеся к данному изобретению. Большой выбор векторов клонирования доступен для осуществления необходимых стадий, требуемых для получения рекомбинантного вектора экспрессии. Среди векторов с системой репликации, функционирующей в Е.coli, имеются векторы, такие как pBR322, ряд векторов pUC, ряд векторов M13mp, pBluescript и т.д. Обычно такие векторы клонирования содержат маркер, позволяющий отбор трансформированных клеток. Последовательности нуклеиновых кислот могут быть введены в эти векторы и векторы могут быть введены в Е.coli, выращенную в подходящей среде. Рекомбинантные векторы экспрессии могут быть легко выделены из клеток при сборе и лизисе клеток. Общие инструкции по получению рекомбинантных векторов представлены, например, в Sambrook et al., Molecular Cloning, a Laboratory Manual, Cold Spring Harbor Laboratory Press, 1989, vol. 3. В соответствии с данным изобретением химерную последовательность нуклеиновой кислоты вводят в клетку растения и клетки выращивают во взрослых растениях, способных давать семена, где семена экспрессируют полипептид инсулина. В соответствии с данным изобретением может быть выбран любой вид растения или клеток растения. Конкретные клетки, применяемые в соответствии с данным изобретением, включают клетки, полученные из Arabidopsis thaliana, бразильского ореха (Betholettia excelsa); клещевины обыкновенной (Riccinus communis), кокоса (Cocus nucifera); кориандра (Coriandrum sativum); хлопка (род Gossypium); арахиса (Arachis Hypogaea); жожоба (Simmondsia chinensis); льняного семени/льна (Linum usitatissimum); кукурузы (Zea mays); горчицы (род Brassica и Sinapis alba); гвинейской масличной пальмы (Elaeis guineeis); оливы (Olea eurpaea); рапса (вид Brassica); риса (Oryza sativa); сафлора (Carthamus tinctorius); сои (Glycine max); тыквы (Cucurbita maxima); ячменя (Hordeum vulgare); пшеницы (Traeticum aestivum) и подсолнечника (Helianthus annuus). В соответствии с данным изобретением в предпочтительном варианте применяют растения или клетки растений из семян масличных растений. Масличные растения, семена которых применяются в соответствии с данным изобретением, включают арахис (Arachis Hypogaea)/горчицу (род Brassica и Sinapis alba); рапс (вид Brassica); нут (Cicer arientinum), сою (Glycine max); хлопок (Gossypium hirsutum); подсолнечник (Helianthus annuus); (Lentil Lens culinaris); льняное семя/лен (Linum usitatissimum); белый клевер (Trifolium repens); оливу (Olea eurpaea); гвинейскую масличную пальму (Elaeis guineeis); сафлор (Carthamus tinctorius) и бобы нарбона (Vicia narbonensis). В соответствии с данным изобретением в особенно предпочтительном варианте применяют сафлор, Arabidopsis или лен. Методики введения растительных рекомбинантных векторов экспрессии в клетку растения, также обозначенные далее как "трансформация", хорошо известны специалистам в данной области техники и обычно зависят от выбранной клетки растения. Общие методики введения рекомбинантных векторов экспрессии включают электропорацию; методики, опосредованные химическими соединениями, например поглощение нуклеиновой кислоты, опосредованное CaCl 2 ; бомбардировку частицами (биолистику); применение природных инфекционных последовательностей нуклеиновых кислот, например последовательности нуклеиновых кислот, полученные из вирусов, или последовательности, полученные из Agrobacterium или Rhizobium, поглощение нуклеиновой кислоты, опосредованное полиэтиленгликолем (ПЭГ), микроинъекции и применение нитевидных кристаллов карбида кремния. В предпочтительных вариантах выбирают такую методику трансформации, которая позволяет интеграцию химерной последовательности нуклеиновой кислоты в геном клетки растения и предпочтительно ядерный геном клетки растения. В соответствии с данным изобретением это считается особенно желательным, так как применение такой методики позволяет перенос химерной последовательности нуклеиновой кислоты в растения потомства при половой репродукции. Методы трансформации, которые могут применяться в соответствии с данным изобретением, включают методы, опосредованные биолистиками и Agrobacterium. Методики трансформации для двудольных видов растений хорошо известны. Обычно применяют трансформацию, опосредованную Agrobacterium, благодаря ее высокой эффективности, а также общую восприимчивость большинством, если не всеми, двудольными видами растений. Трансформация с Agrobacterium обычно включает перенос бинарного вектора, такого как любой из упомянутых выше бинарных векторов, включающего химерную последовательность нуклеиновой кислоты в соответствии с данным изобретением, из Е.coli в подходящий штамм Agrobacterium (например, ЕНА101 и LBA4404) с помощью, например, скрещивания трех родителей со штаммом Е.coli, содержащим рекомбинантный бинарный вектор, и штаммом Е.coli, содержащим вспомогательную плазмиду, мобилизующую бинарный вектор по отношению к целевому штамму Agrobacterium, или трансформации ДНК штамма Agrobacterium (Hofgen et al., Nucl. Acids Res., 1988, 16: 9877). Другие методики, которые могут применяться для трансформации клеток двудольных растений, включают биолистики (Sanford, 1988, Trends in Biotechn. 6: 299-302); электропорацию (Fromm et al., 1985, Proc. Natl. Acad. Sci. USA, 82: 5824-5828); опосредованное ПЭГ поглощение ДНК (Potrykus et al., 1985, Mol. Gen. Genetics, 199: 169-177); микроинъекции (Reich et al., Bio/Techn., 1986, 4: 1001-1004) и нитевидные кристаллы карбида кремния (Kaeppler et al., 1990, Plant Cell Rep., 9: 415-418) или трансформацию in planta с применением, например, методики погружения цветка (Clough and Bent, 1998, Plant J., 16: 735-743). Однодольные виды растений могут быть трансформированы с применением различных методик, включая бомбардировку частицами (Christou et al., 1991, Biotechn. 9: 957-962; Weeks et al., Plant Physiol., 1993, 102: 1077-1084; Gordon-Kamm et al., Plant Cell, 1990, 2: 5603-618); опосредованное ПЭГ поглощение ДНК (европейские патенты 0292435; 0392225) или трансформацию, опосредованную Agrobacterium (Goto-Fumiyuki et al., 1999, Nature-Biotech. 17: 282-286). Конкретная методика трансформации растения может варьироваться в зависимости от вида растения и типа клетки растения (например, полученные из рассады типы клеток, такие как гипокотилы и котиледоны или ткань эмбриона), которые выбирают в качестве клетки-мишени для трансформации. Как указано выше, в особенно предпочтительном варианте применяют сафлор, Arabidopsis или лен. Методика получения трансформатов сафлора описана у Baker and Dyer (Plant Cell Rep., 1996, 16: 106-110). Другие протоколы трансформации определенных видов растений могут быть найдены в Biotechnology in Agriculture and Forestry 46: Transgenic Crops I (Y.P.S. Bajaj ed.), Springer-Verlag, New York (1999) и Biotechnology in Agriculture and Forestry 47: Transgenic Crops II (Y.P.S. Bajaj ed.), Springer-Verlag, New York (2001). Для трансформации клетки растений выращивают и при появлении дифференцирующей ткани, такой как ростки и корни, взрослые растения регенерируют. Обычно регенерируют множество растений. Методики регенерации растений зависят от вида растений и типа клеток и известны специалисту в данной области техники. Дальнейшие указания по культивированию тканей растений можно найти, например, в Plant Cell and Tissue Culture, 1994, Vasil and Thorpe Eds., Kluwer Academic Publishers и Plant Cell Culture Protocols (Methods in Molecular Biology 111), 1999, Hall Eds., Humana Press. В одном аспекте данное изобретение относится к способу выделения семян растений, содержащих инсулин. Следовательно, данное изобретение относится к способу получения семян растений, содержащих инсулин, который включает: (a) получение конструкции химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений; и (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения; (c) выращивание клетки растений в зрелом растении, способном давать семена; и (d) получение семян из указанных растений, где семена содержат инсулин. В предпочтительных вариантах множество трансформированных растений получают, выращивают и исследуют на присутствие желаемой химерной последовательности нуклеиновой кислоты, присутствие которой в предполагаемых трансформатах может быть протестировано, например, выращиванием в селективной среде, в которой применяют маркеры, устойчивые к гербицидам, путем непосредственного нанесения гербицида на растение или саузерн-блоттингом. Если определяется присутствие химерной последовательности нуклеиновой кислоты, трансформированные растения могут быть отобраны для выращивания потомства и, в конечном счете, взрослых растений, содержащих множество семян, содержащих желаемую последовательность химерной нуклеиновой кислоты. Таким семена могут применяться для выделения инсулина или они могут быть посажены для получения двух или более последующих поколений. Обычно желательно высаживать множество трансгенных семян для получения трансгенных растений, каждое из которых содержит семена, содержащие химерную последовательность нуклеиновой кислоты, кодирующую инсулин. Более того, обычно желательно удостовериться в гомозиготности растений для того, чтобы удостовериться в непрерывном наследовании рекомбинантного полипептида. Методы отбора гомозиготных растений хорошо известны специалистам в данной области техники. Методы получения гомозиготных растений, которые могут применяться в соответствии с данным изобретением, включают получение и трансформацию гаплоидных клеток или тканей с последующей регенерацией гаплоидных растений и последующим превращением в диплоидные растения, например, обработкой колхином или другими агентами, повреждающими микротрубочки. Растения можно выращивать в соответствии с обычной сельскохозяйственной практикой. В другом аспекте данное изобретение относится к растениям, способным давать семена, экспрессирующие инсулин. В предпочтительном варианте данного изобретения растения, способные давать семена, содержат последовательность химерной нуклеиновой кислоты, содержащую в 5'-3' направлении транскрипции: (a) первую последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений, с которыми она функционально связана; и (b) вторую последовательность нуклеиновой кислоты, кодирующую полипептид инсулина. В предпочтительном варианте химерная последовательность нуклеиновой кислоты стабильно интегрирована в ядерный геном растения. В еще одном варианте данное изобретение относится к семенам растений, экспрессирующим инсулин. В предпочтительном варианте данного изобретения семена растений содержат химерную последовательность нуклеиновой кислоты, содержащую в 5'-3' направлении транскрипции: (a) первую последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений, с которыми она функционально связана; и (b) вторую последовательность нуклеиновой кислоты, кодирующую полипептид инсулина. В соответствии с данным изобретением получают семена, в которых предпочтительно по крайней мере 0,1% от общего растворимого белка, присутствующего в семени, составляет инсулин. В другом предпочтительном варианте данного изобретения получают семена, в которых по крайней мере 0,2, 0,3, 0,5 или 1,0% от общего растворимого белка, присутствующего в семени, составляет инсулин. Полипептид инсулина может присутствовать в различных типах клеток, включая, например, гипокотилы и ось эмбриона, включая корни эмбриона и листья эмбриона, где однодольные виды растений, включая злаки и кукурузу, применяют в ткани эндосперма. После получения семян растения белок инсулина может быть выделен из семян с помощью любой методики очистки белка, известной в данной области техники. Следовательно, данное изобретение относится к способу выделения инсулина из семян растений, где способ включает: (a) получение конструкции химерной нуклеиновой кислоты, содержащей в 5'-3' направлении транскрипции в виде функционально связанных компонентов: (i) последовательность нуклеиновой кислоты, способную контролировать экспрессию в клетках семян растений; и (ii) последовательность нуклеиновой кислоты, кодирующую полипептид инсулина; (b) введение конструкции химерной нуклеиновой кислоты в клетку растения; (c) выращивание клетки растений в зрелом растении, способном давать семена, где семена экспрессируют инсулин; (d) получение семян, экспрессирующих инсулин; и (e) выделение инсулина из семян. Семена растений могут быть измельчены с применением любого общепринятого способа, дающего значительное разрушение мембраны клетки и стенок клетки. Могут применяться условия сухого и влажного измельчения (патент США 3971856; Lawhon et al., 1977, J. Am. Oil Chem. Soc., 63: 533-534). Подходящее оборудование для измельчения включает коллоидные мельницы, дисковые мельницы, IKA мельницы, гомогенизаторы промышленного масштаба и подобные. Выбор оборудования для измельчения зависит от типа семян и общих требований. Твердые примеси, такие как оболочка семян, волокнистый материал, нерастворимые углеводы, белки и другие не растворимые в воде примеси, могут быть удалены из фракции семян с помощью, например, методик на основе отбора по размеру, таких как процессы на основе фильтрации или гравитации, такие как центрифугирование. В предпочтительных вариантах избегают применения органических растворителей, обычно применяемых при извлечении масла, таких как гексан, так как такие растворители могут повредить полипептид инсулина. Практически чистый инсулин может быть выделен из семян с помощью множества дополнительных методик очистки, таких как методики на основе центрифугирования; методики на основе отбора по размеру, включая, например, ультрафильтрацию через мембрану и ультрафильтрацию в поперечном потоке; и методики хроматографии, включая, например, ионообменную хроматографию, хроматографию с отбором по размеру, аффинную хроматографию, высокоэффективную жидкостную хроматографию (ВЭЖХ), жидкостную экспресс-хроматографию белком (ЖХББ), хроматографию с гидрофобным взаимодействием, и подобные. Обычно для получения практически чистого инсулина применяют совокупность таких методик. В особенно предпочтительном варианте данного изобретения полипептид инсулина выделяют из примесей семян взаимодействием полипептида инсулина с масляными тельцами. Этот метод считается особенно преимущественным, так как он позволяет удалять примеси семян, включая белок семян особенно эффективным и недорогим способом. Как указано выше, взаимодействие полипептида инсулина с масляными тельцами может быть достигнуто связыванием полипептида инсулина с белком масляного тельца или связыванием полипептида инсулина с полипептидом, обладающим сродством с масляным тельцем, таким как антитело с одной цепью, обладающее сродством к масляному тельцу. В первом варианте полипептид инсулина будет изолирован в клетках масляных телец и, следовательно, очищаться совместно с масляными тельцами. Во втором варианте будучи экспрессированным в мембране, окружающей внутриклеточный компартмент, такой как ER, полипептид инсулина будет связываться с масляным тельцем при разрушении клеток семени во время процесса измельчения. Способ выделения масляных телец описан в патенте США 5650554. Фармацевтические композиции инсулина могут быть получены из очищенного инсулина и такие композиции могут применяться для лечения диабета. Обычно очищенный инсулин смешивают с фармацевтически приемлемым носителем или разбавителем в количествах, достаточных для оказания терапевтически полезного действия при отсутствии нежелательных побочных эффектов на пациента. Для получения композиции инсулина массовую часть инсулина растворяют, суспендируют, диспергируют или смешивают другим способом в выбранном носителе или разбавителе до эффективной концентрации, позволяющей улучшить лечимое состояние. Фармацевтические композиции инсулина предпочтительно имеют вид единичных дозированных форм. Терапевтически эффективные дозы для парентерального введения человеческого инсулина известны в данной области техники. При применении аналогов инсулина или других способов введения применяемые терапевтически эффективные дозы могут быть эмпирически определены специалистом в данной области техники с применением известных протоколов тестирования, или экстраполяцией данных тестирования in vivo или in vitro. Однако понятно, что концентрации и дозы могут варьироваться в соответствии с тяжестью облегчаемого состояния. Также понятно, что для любого конкретного пациента могут применяться определенные режимы дозирования во времени, определяемые лечащим или наблюдающим лечение специалистом. Фармацевтические растворы или суспензии могут включать, например, стерильные разбавители, такие как, например, вода, лактоза, сахароза, дикальцийфосфат или карбоксиметилцеллюлоза. Применяемые носители включают воду, физиологический раствор, водную декстрозу, глицерин, гликоли, этанол и подобные, которые могут образовывать раствор или суспензию. При желании, фармацевтические композиции также могут содержать нетоксичные вспомогательные вещества, такие как смачивающие агенты; эмульгирующие агенты; солюбилизаторы; противомикробные агенты, такие как бензиловый спирт и метилпарабены; антиокислители, такие как аскорбиновая кислота и бисульфит натрия; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота (EDTA); рН буферные агенты, такие как ацетатные, цитратные или фосфатные буферы; и их сочетания. Конечная композиция препаративной формы инсулина в основном зависит от способа введения инсулина. Инсулин, полученный в соответствии с данным изобретением, может вводиться любым желательным способом; однако парентеральный, пероральный, легочный, буккальный и назальный способы введения считаются наиболее часто применяемыми способами введения. Парентеральные препаративные формы могут быть в ампулах, одноразовых шприцах или флаконах, содержащих одну или несколько доз, сделанных из стекла, пластика или других подходящих материалов. Представленные ниже примеры приведены только для иллюстрации и не ограничивают данное изобретение. Пример 1. Получение белка инсулина, экспрессированного в виде слитого белка мини-инсулина (MI) с расщепляемым трипсином пропептидом. Получение pSBS4404: PRS-D9scFv-Klip27-MI-KDEL слитого белка. Один из изучаемых слитых белков начинается с последовательности, связанной с патогеном табака (PRS) (Sijmons et al., 1990, Bio/technology, 8: 217-221), которая служит в качестве сигнального пептида для целевой экспрессии в ER котрансляционным методом. Сразу же после нее идет последовательность, кодирующая Fv антитело с одной цепью (scFv) со специфическим к видам сродством против 18 кДа олеозина из Arabidopsis thaliana, обозначенная D9scFv, далее идет расщепляемый трипсином пропептид (Klip27), полученный из ТА57 пропептида дрожжей (Kjeldsen et al., 2001, Biotechnology and Genetic Engineering Reviews 18: 89-121). За ней следует мини-инсулин (MI), описанный у Kjeldsen et al. (2001), с добавлением сигнала ER-удержания KDEL на С-конце полипептида. Основная цепь этой плазмиды, pSBS4055, основана на бинарном векторе растения, pPZP200, описанном у Hajdukiewicz et al. (Plant Molecular Biology, 1994, 25: 989-994). Вместо описанного сайта множественного клонирования, pat ген, придающий растению-хозяину устойчивость к фосфинотрицину (Wohlleben et al., 1988, Gene 70: 25-37), управляемый промотором/терминатором убиквитина из Petroselinum crispum (Kawalleck et al., 1993, Plant Mol. Bio., 21: 673-684), был введен между левой и правой пограничными последовательностями. В дополнение к этой кассете промотор/терминатор β-фазеолина из Phaseolus vulgaris (Slightom et al., 1983, Proc. Natl. Acad. Sci. USA 80: 1897-1901), управляющий PRS, был субклонирован. Стандартную PCR (Horton et al., 1989, Gene 77: 61-68) применяли для слияния синтетической PRS-кодирующей последовательности с присоединенными сайтами рестрикции SphI/HindIII эндонуклеазы с 3'-конца промотора фазеолина с получением pSBS4011. SphI-D9scFv-XhoI, SwaI, HindIII введенную последовательность генерировали PCR амплификацией D9scFv кДНК клона (Sean Hemmingsen lab, неопубликовано) с праймерами 1325 (GCATGCTGACATTGTGATGACACAGTC) - SEQ ID NO:175 и 1326 (AAGCTTGCATTTAAATACTCGAGACTGTGAGAGTGGTGCCTTG) - SEQ ID NO:176. Последующее лигирование этого фрагмента в сайтах SphI/HindIII pSBS4011 дает плазмиду pSBS4055. Последовательность Klip27-MI синтезируют из четырех частично перекрывающихся олигонуклеотидов, в которые введен кодон Arabidopsis thaliana, применяемый для повышения эффективности трансляции в системах экспрессии, основанных на растениях. Олигонуклеотиды 1324 (GAAGAAGGAGAGCCTAAGTTTGTTAATCAACATCTTTGTGGATCTCATCTTGTTGAGGCTCTCTACCTTG) - SEQ ID NO:177 и 1323 (CCTTAGGAGTGTAGAAAAATCCTCTTTCTCCACACACAAGGTAGAGAGCCTCAACA) SEQ ID NO:178 гибридизируют в их комплементарном наложении 20 нуклеотидов и удлиняют с получением 5'-конца Klip27-MI слияния, то же самое проводят с олигонуклеотидами 1322 (CTAAGGCTGCTAAGGGAATTG() - SEQ ID NO:179 и 1321 (AAGCTTCAGTTGCAATAGTTCTCCAATTGGTAAAGTGAGCAAATAGAAGTGCAACATTGTTCAACAATTCCCTTAGCAGCCTT) - SEQ ID NO:180 с получением 3'-конца. Две половины лигируют с последующим рестрикционным расщеплением с Bsu36I с получением полной Klip27-MI кодирующей последовательности. PCR такого слияния гена с применением праймеров 1364 (CTCGAGTCAACCAATTGATGACACTGAATC) - SEQ ID NO:181 и 1334 (AAGCTTCAAAGTTCATCCTTGTTGCAATAGTTCTCCAATTG) - SEQ ID NO:182 присоединяют 5' сайт расщепления XhoI рестрикционной эндонуклеазы и 3' KDEL ДНК последовательность плюс HindIII сайт расщепления для последующего лигирования в XhoI/HindIII-разрыв pSBS4055. Результатом является плазмида pSBS4404: последовательность ДНК, кодирующая слитый белок PRS-D9scFv-Klip27-MI-KDEL, помещенный в бинарный вектор под контролем экспрессии промотором/терминатором фазеолина. Промотор фазеолина контролирует специфически временную и специфическую к тканям экспрессию трансгена во время развития семян. Полная последовательность нуклеиновой кислоты (SEQ ID NO:1) и последовательность аминокислоты (SEQ ID NO:2) слитого белка инсулина 4404 (PRS-D9scFv-Klip27-MI-KDEL) показана на фиг. 1. Получение pSBS4405: OLEO-Klip8-Klip27-MI слитого белка. Второй изучаемый слитый белок начинается с 18 кДа олеозина из Arabidopsis thaliana, далее продолжается расщепляемым химозином пропептидом (Klip8) - SEQ ID NO:175. Сразу же за ним идет последовательность, кодирующая расщепляемый трипсином пропептид (Klip27), полученная из ТА57 пропептида дрожжей, как описано выше (Kjeldsen et al., 2001, Biotechnology and Genetic Engineering Reviews 18: 89-121). Она слита с мини-инсулином (MI), описанным выше (Kjeldsen et al. 2001). Экспрессия этого слитого белка нацелена на новые масляные тельца, образующиеся в процессе развития эмбриона. Основная цепь этой плазмиды, pSBS4055, основана на бинарном векторе растения, pPZP200, описанном у Hajdukiewicz et al. (Plant Molecular Biology, 1994, 25: 989-994). Вместо описанного сайта множественного клонирования, pat ген, придающий растению-хозяину устойчивость к фосфинотрицину (Wohlleben et al., 1988, Gene 70: 25-37), управляемый промотором/терминатором убиквитина из Petroselinum crispum (Kawalleck et al., 1993, Plant Mol. Bio., 21: 673-684), был введен между левой и правой пограничными последовательностями. В дополнение к этой кассете промотор/терминатор β-фазеолина из Phaseolus vulgaris (Slightom et al., 1983, Proc. Natl. Acad. Sci. USA 80: 1897-1901), управляющий слиянием геномной последовательности-Klip8 Arabidopsis 18 кДа олеозина, был субклонирован. Стандартную PCR (Horton et al., 1989, Gene 77: 61-68) применяли для слияния последовательности ген олеозина-Klip8 с сайтами XhoI/HindIII рестрикционной эндонуклеазы с 3'-конца промотора фазеолина с получением pSBS4010. Последовательность Klip27-MI синтезируют из четырех частично перекрывающихся олигонуклеотидов, в которые введен кодон Arabidopsis thaliana, применяемый для повышения эффективности трансляции в системах экспрессии, основанных на растениях. Олигонуклеотиды 1324 (GAAGAAGGAGAGCCTAAGTTTGTTAATCAACATCTTTGTGGATCTCATCTTGTTGAGGCTCTCTACCTTG) - SEQ ID NO:177 и 1323 (CCTTAGGAGTGTAGAAAAATCCTCTTTCTCCACACACAAGGTAGAGAGCCTCAACA) SEQ ID NO:178 гибридизируют в их комплементарном наложении 20 нуклеотидов и продлевают с получением 5'-конца Klip27-MI слияния, то же самое проводят с олигонуклеотидами 1322 (CTAAGGCTGCTAAGGGAATTG) - SEQ ID NO:179 и 1321 (AAGCTTCAGTTGCAATAGTTCTCCAATTGGTAAAGTGAGCAAATAGAAGTGCAACATTGTTCAACAATTCCCTTAGCAGCCTT) - SEQ ID NO:180 с получением 3'-конца. Две половины лигируют с последующим рестрикционным расщеплением с Bsu36I с получением полной Klip27-MI кодирующей последовательности. PCR такого слияния гена с применением праймеров 1364 (CTCGAGTCAACCAATTGATGACACTGAATC) - SEQ ID NO:181 и 1329 (AAGCTTCAGTTGCAATAGTTC) - SEQ ID NO:183 присоединяют 5' сайт расщепления XhoI рестрикционной эндонуклеазы и 3' HindIII сайт расщепления соответственно для последующего лигирования в XhoI/HindIII-разрыв pSBS4010. Результатом является плазмид pSBS4405: последовательность ДНК, кодирующая слитый белок OLEO-Klip8-Klip27-MI, помещенный в бинарный вектор под контролем экспрессии промотором/терминатором фазеолина. Промотор фазеолина контролирует специфически временную и специфическую к тканям экспрессию трансгена во время развития семян. Полная последовательность нуклеиновой кислоты (SEQ ID NO:3) и последовательность аминокислоты (SEQ ID NO:4) слитого белка инсулина 4405 (OLEO-Klip8-Klip27-MI) показана на фиг. 2. Получение pSBS4414: PRS-MI-тетраосновный линкер-D9scFv-KDEL слитого белка. Другой из изучаемых слитых белков начинается с последовательности, связанной с патогеном табака (PRS) (Sijmons et al., 1990, Bio/technology, 8: 217-221), которая служит в качестве сигнального пептида для целевой экспрессии в ER котрансляционным методом. Сразу же после нее идет последовательность, кодирующая мини-инсулин (MI), описанный у Kjeldsen et al. (2001), за исключением того, что область мини-С пептида (AAK-SEQ ID NO:146) заменяют промежуточным В 30 треонин-тетраосновным сайтом (B 30 -T-RRKR) (SEQ ID NO:149), последовательностью между В( 1-29 )- и A( 1-21 )-цепями человеческого инсулина. Затем сразу же следует последовательность, кодирующая второй тетраосновный линкер, а затем Fv антитело с одной цепью (scFv) с видоспецифичным сродством против 18 кДа олеозина из Arabidopsis thaliana, обозначенная D9scFv. На С-конце полипептида добавлен сигнал ER-удержания KDEL. Основная цепь этой плазмиды, pSBS4055, основана на бинарном векторе растения, pPZP200, описанном у Hajdukiewicz et al. (Plant Molecular Biology, 1994, 25: 989-994). Вместо описанного сайта множественного клонирования, pat ген, придающий растению-хозяину устойчивость к фосфинотрицину (Wohlleben et al., 1988, Gene 70: 25-37), управляемый промотором/терминатором убиквитина из Petroselinum crispum (Kawalleck et al., 1993, Plant Mol. Bio., 21: 673-684), был введен между левой и правой пограничными последовательностями. В дополнение к этой кассете промотор/терминатор β-фазеолина из Phaseolus vulgaris (Slightom et al., 1983, Proc. Natl. Acad. Sci. USA 80: 1897-1901), управляющий PRS, был субклонирован. Стандартную PCR (Horton et al., 1989, Gene 77: 61-68) применяли для слияния синтетической PRS-кодирующей последовательности с присоединенными сайтами SphI/HindIII рестрикционной эндонуклеазы с 3'-конца промотора фазеолина с получением pSBS4011. Последовательность Klip27-MI синтезируют из четырех частично перекрывающихся олигонуклеотидов, в которые введен кодон Arabidopsis thaliana, применяемый для повышения эффективности трансляции в системах экспрессии, основанных на растениях. Олигонуклеотиды 1324 (GAAGAAGGAGAGCCTAAGTTTGTTAATCAACATCTTTGTGGATCTCATCTTGTTGAGGCTCTCTACCTTG) - SEQ ID NO:177 и 1323 (CCTTAGGAGTGTAGAAAAATCCTCTTTCTCCACACACAAGGTAGAGAGCCTCAACA) - SEQ ID NO:178 гибридизируют в их комплементарном наложении 20 нуклеотидов и продлевают с получением 5'-конца Klip27-MI слияния, то же самое проводят с олигонуклеотидами 1322 (CTAAGGCTGCTAAGGGAATTG) - SEQ ID NO:179 и 1321 (AAGCTTCAGTTGCAATAGTTCTCCAATTGGTAAAGTGAGCAAATAGAAGTGCAACATTGTTCAACAATTCCCTTAGCAGCCTT) - SEQ ID NO:180 с получением 3'-конца. Две половины лигируют с последующим рестрикционным расщеплением с Bsu36I с получением полной Klip27-MI кодирующей последовательности. PCR такого слияния гена с применением праймеров 1363 (GCATGCCCAACCAATTGATGACACTG) - SEQ ID NO:184 и 1334 (AAGCTTCAAAGTTCATCCTTGTTGCAATAGTTCTCCAATTG) - SEQ ID NO:182 присоединяют 5' сайт расщепления SphI рестрикционной эндонуклеазы и 3' KDEL ДНК последовательность плюс HindIII сайт расщепления для последующего лигирования в SphI/HindIII-разрыв pSBS4011. Результатом является плазмида pSBS4402: последовательность ДНК, кодирующая слитый белок PRS-Klip27-MI-KDEL, помещенный в бинарный вектор под контролем экспрессии промотором/терминатором фазеолина. Промотор фазеолина контролирует специфически временную и специфическую к тканям экспрессию трансгена во время развития семян. Вектор экспрессии растения, pSBS4402, служит в качестве образца введения сайта между В- и А-цепями инсулина и между MI и D9scFv. Промежуточный тетраосновный сайт (B 30 -T-RRKR) помещают между настоящими B( 1-29 )- и A( 1-21 )-цепями человеческого инсулина с помощью PCR с применением праймеров 1515 (GCATGCATGCCTTTGTTAATCAACATCTTTGTGG) SEQ ID NO:185 и 1518 (ACATTGTTCAACAATTCCTCTCTTTCTTCTAGTCTTAGGAGTGTAGAAAAATCC) SEQ ID NO:186 с применением pSBS4402 в качестве образца. Полученный 124 п.н. фрагмент применяют в сочетании с праймером 1517 (GCATAAGCTTCAAAGCTCATCCTTTGAGC) SEQ ID NO:187 с применением pSBS3400 в качестве образца. Необходимо отметить, что pSBS3400 представляет собой плазмиду, которая содержит D9scFv-KDEL фрагмент с сайтом рестрикции HindIII. Реакция PCR дает 955 п.н. продукт, который вводит тетраосновный (RRKR)-D9scFv-KDEL-HindIII в 124 п.н. SphI-MI фрагмент. Фрагмент 955 п.н. затем лигируют и субклонируют в pGEM-T (Promega) с получением pSBS3403. Весь SphI-MI (с B 30 -T-RRKR модифицированным С пропептидом)-RRKR-D9scFv-KDEL-HindIII фрагмент вводят в предварительно разрезанный (SphI/HindIII) pSBS4402 с получением pSBS4414. Полная последовательность нуклеиновой кислоты (SEQ ID NO:5) и последовательность аминокислоты (SEQ ID NO:6) слитого белка инсулина 4414 (PRS-MI-тетраосновный линкер-D9scFv-KDEL) показаны на фиг. 3. После подтверждения целостности кДНК кодирования для слитого белка анализом последовательности плазмиды pSBS4404, pSBS4405 и pSBS4414 трансформируют в штамм Е.coli DH5 α для получения высокого уровня экспрессии. Выделенную плазмиду ДНК (100 нг) смешивают на льду со 100 мкл компетентных клеток DH5 α в течение 20 мин. Затем клетки подвергают тепловому удару при температуре 42 °С в течение 45 с и возвращают на лед на 2 мин. Затем добавляют 1 мл среды SOC и клетки инкубируют при температуре 37 °С на энвиро-шейкере при 225 об/мин в течение 1 ч перед помещением трансформированных клеток на планшеты с LB-спектиномицином (10 г/л триптона, 5 г/л экстракта дрожжей, 5 г/л NaCl, 15 г/л агара) и инкубированием в течение ночи при температуре 37 °С. Одну колонию используют для инокулирования 5 мл LB-спектиномицинового бульона. Эти культуры выращивают в течение ночи при температуре 37 °С. Рекомбинантную плазмиду выделяют из 1 мл среды, выстаиваемой в течение ночи, с применением набора QIAprep ®Spin Miniprep (Qiagen). Выделенную плазмиду затем применяют для трансформации компетентного штамма Agrobacterium EH101 (Hood et al., 1986; J. Bacteriol. 144: 732-743) электропорацией (25 мкФ, 2,5 кВ, 200 Ом). Рекомбинантные Agrobacteruim помещают на планшеты с АВ-спектиномицином/канамицином (20 × АВ соли, 2М глюкозы, 0,25 мг/мл FeSO 4 ∙7H 2 O, 1M MgSO 4 , 1M CaCl 2 ) и одну колонию применяют для инокулирования 5 мл АВ-спектиномицинового/канамицинового бульона. Эти культуры выращивают в течение ночи при температуре 28 °С. Рекомбинантные Agrobacteruim затем применяют для трансформации растений Arabidopsis thaliana методом погружения цветков (Clough et al., 1998, Plant J., 16: 735-743). Вид Arabidopsis thaliana (C24) применяют для всех экспериментов. Семена высевают на поверхности смеси почв (две трети земли Redi и одна треть перлита с pH 6,7) или смеси почв для Arabidopsis от Lehle Seeds (перлит, вермикулит, торф, терра-грин с pH 5,5) в 4-дюймовые горшки. Саженцы выращивают до стадии розеток из 6-8 листьев диаметром приблизительно 2,5 см. Горшки помещают внутрь купола при температуре 4 °С на четыре дня для холодной обработки и затем переносят в комнату для выращивания с температурой 24 °С с постоянным светом около 150 мкЕ и относительной влажностью 60-70%. Растения орошают с интервалом 2-3 дня и еженедельно подкармливают удобрением 1% Peters 20-19-18. Каждый горшок содержит от пяти до шести растений. Когда растения достигают около 2 см в высоту, первые стрелки срезают для стимулирования роста вторых и третьих стрелок. Через 4-5 дней после среза первичных стрелок растения готовы к заражению Agrobacterium. Горшки с растениями Arabidopsis опрокидывают для заражения растений Arabidopsis в 500 мл ресуспензии культуры Agrobacterium, выращенной в течение ночи, содержащей целевой вектор трансформации растения, в течение 20 с. Важно, чтобы культура Agrobacterium содержала 5% сахарозы и 0,05% поверхностно-активного вещества Silwet L-77 (Lehle Seeds). Далее горшки накрывают прозрачными пластиковыми крышками на 24 ч для поддержания более высокой влажности. Растения выращивают до взрослого состояния и собирают смесь семян, трансформированных и нетрансформированных. Для выбора трансгенных линий предполагаемые трансформированные семена стерилизуют быстрым промыванием 70% этанолом, затем 20% коммерческим отбеливателем в течение 15 мин и затем ополаскивают по крайней мере четыре раза ddH 2 O. Около 1000 стерилизованных семян смешивают с 0,6% расплавленным агаром и равномерно распределяют на MS планшете с половинной силой (Murashige and Skoog, 1962, Physiologia Plantarum 15: 473-497), содержащем 0,3% сахарозы и 80 мкМ гербицида фосфинотрицина (РРТ) DL. Затем планшеты помещают в комнату для выращивания с режимом освещения 8 ч темноты и 16 ч света при температуре 24 °С. Через 7-10 дней предполагаемые трансгенные саженцы остаются зелеными и растут, в то время как нетрансформированные саженцы бледнеют. После появления корней предполагаемые трансгенные саженцы индивидуально переносят в горшки (отдельные растения орошают с интервалом 3 дня и подкармливают удобрениями 1% Peters 20-19-18 с интервалом 7 дней) и выращивают до взрослого состояния. Горшки накрывают прозрачными пластиковыми крышками на 3 дня для защиты чувствительных саженцев. Через 7 дней саженцы накрывают системой сбора семян от Lehle Seeds для предотвращения потери семян из-за рассеивания. Семена трансгенных растений собирают отдельно и готовят для анализа. Пример 2. Уровни экспрессии инсулина у Arabidopsis thaliana. Во втором примере определяют уровни экспрессии слитого белка D9scFv-Klip27-MI KDEL (4404), OLEO-Klip8-Klip27-MI (4405) и PRS-MI-RRKR-D9scFv-KDEL (4414) в трансгенных зрелых семенах Arabidopsis thaliana. Показано, что трансгенный продукт присутствует в клеточных экстрактах зрелых семян. Приблизительно 40 трансгенных семян Arabidopsis thaliana измельчают в ступе пестиком в 50 мкл 50 мМ Трис-HCl, рН 8,0. Затем к суспензии добавляют восстанавливающий SDS-PAGE образцовый буфер (6 × SDS образцовый буфер, 0,35М Трис-HCl, рН 6,8, 30% глицерин, 10% SDS, 0,012% голубого бромфенола, 5% β-меркаптоэтанола) и перемешивают короткими завихрениями. Затем образец быстро центрифугируют и выдерживают при температуре 99 °С в течение 10 мин. После охлаждения на льду в течение 2 мин образец быстро центрифугируют. Образцы помещают (10 мкл - эквивалент приблизительно 7 семенам) в условия восстановления. Для подготовленных образцов масляных телец трансгенные и дикие семена (20 мг) измельчают в 250 мкл буфера экстрагирования масляных телец (0,4М сахарозы, 0,5М NaCl, 50 мМ Трис-HCl, рН 8,0). Образцы микроцентрифугируют при 10000 g в течение 10 мин. Растворимые водные фракции удаляют с помощью 26 G 5/8 1 мл шприца и жирный слой повторно суспендируют в 100 мкл фосфатного буфера с добавлением соли (20 мМ Na 2 HPO 4 , рН 8,0, 0,5М NaCl). Повторно суспендированный жирный слой переносят в чистую пробирку микроцентрифуги и снова центрифугируют при 10000 g в течение 10 мин. Процедуру повторяют еще 3 раза с последним повторным суспендированием жирного слоя в 100 мкл фосфатном буфере без соли (20 мМ Na 2 HPO 4 , рН 8,0). Два дополнительных промывания в фосфатном буфере без соли проводят поочередно с центрифугированием, как описано выше. Конечную жирную лепешку повторно суспендируют в 10 мкл фосфатного буфера (20 мМ Na 2 HPO 4 , рН 8,0). Берут 5 мкл аликвоту и белок масляного тельца солюбилизируют кипячением в 1/10 (об./об.) 50 мМ Трис-HCl, рН 8,0 с 2% SDS. Образец охлаждают на льду в течение 2 мин и центрифугируют при 10000 g в течение 5 мин. Содержание белка в подосадочной жидкости определяют ИСФ анализом белка (Pierce, Rockford, IL). Для анализа окрашенных кумасси гелей и вестерн-блоттинга 20 мкг общего белка отделяют на 15% SDS-PAGE гелях в условиях восстановления с применением SDS-PAGE образцового буфера. Затем образцы загружают в дискретные 15% SDS-PAGE гели и разделяют при 150 В в течение приблизительно 1,5 ч. Затем гели либо окрашивают кумасси, либо наносят на PVDF мембрану (Immobilon-P, Millipore Corporation, Bedford, MA) для вестерн-блоттинга. Нанесенные образцы исследуют с. помощью моноклональных антител, направленных против инсулина (Clone Е2-Е3; Roth et al., 1992) от Abcam (Cambridge, UK). Полосы инсулина определяют с применением вторичного Sheep X IgG мыши F(ab') 2 АР-конъюгата (Chemicon International, Temecula, CA) и проявляют с помощью NBT-BCIP в GARAP буфере (Трис-HCl, рН 9,5, 100 мМ NaCl, 5 мМ MgCl 2 ). Иммунореакционноспособная полоса соответствует полипептидной полосе, мигрируя при расчетном молекулярном весе слитого белка, как показано на фиг. 4A-4F. На фиг. 4A-4F показана рекомбинантная экспрессия слитых белков инсулина в трансформированных линиях Arabidopsis thaliana (4404-2, -17, -20, 4405-4, 4414-19 и 4414-20) на основе окрашенного кумасси SDS-PAGE и вестерн-блоттинга. Стрелки указывают на положение мигрирующих 38,5 кДа, 34,2 кДа и 34,2 кДа слитых белков, PRS-D9scFv-Klip27-MIw/KDEL (4404), OLEO-Klip8-Klip27-MI (4405) и PRS-MI-RRKR-D9scFv-KDEL (4414) соответственно в условиях восстановления. Необходимо отметить, что слитый белок 4414 имеет ожидаемый молекулярный вес 34,2 кДа, но имеет более высокий реальный молекулярный вес на SDS-PAGE геле. На фиг. 4А (окрашенный кумасси гель) и 4В (соответствующий вестерн-блоттинг, исследованный с антиинсулином Е2Е3) показан общий белок семян из диких (wt) и трансгенных линий семян, экспрессирующих 4404 и 4405 конструкции. На фиг. 4С (окрашенный кумасси гель) и 4D (соответствующий вестерн-блоттинг, исследованный с антиинсулином Е2Е3) показан белок масляных телец, полученный из диких и трансгенных семян, экспрессирующих те же самые 4404 и 4405 конструкции. На фиг. 4D (окрашенный кумасси гель) и 4Е (соответствующий вестерн-блоттинг, исследованный с антиинсулином Е2Е3) показан белок масляных телец, полученный из диких и трансгенных семян, экспрессирующих ту же самую 4414 конструкцию. Маркеры молекулярного веса (М) равны 10, 15, 20, 25, 37, 50, 75, 100, 150, 250 кДа. Контрольные группы включают hIN (стандарт рекомбинантного человеческого инсулина) и hProIN (стандарт рекомбинантного человеческого проинсулина), выделенные в невосстанавливающих условиях. Различия в уровнях экспрессии являются результатом клональных вариаций между трансформантами. Приблизительные уровни белка трансгенов в экспрессии MI показаны на фиг. 5. Уровни экспрессии определяют с применением ленты 18 кДа олеозина в качестве внутреннего стандарта (эквивалентно 1,5% общего белка семян) денситометрией ленты трансгена. Средний уровень экспрессии для PRS-D9scFv-Klip27-MIw/KDEL (4404), OLEO-Klip8-Klip27-MI (4405) и PRS-MI-RRKR-D9scFv-KDEL (4414) конструкций составляет 0,21% от общего белка семени, 0,12% от общего белка семени и 0,79% от общего белка семени соответственно. Пример 3. Расщепление pSBS4404 и очистка ВЭЖХ. Элюирование из масляного тельца. В третьем примере 1 г трансгенных семян гомогенизируют в 12 мл буфера экстрагирования (0,4М сахарозы, 0,5М NaCl, 50 мМ Трис-HCl, рН 8,0) и центрифугируют при 10000 g в течение 10 мин, жирные слои удаляют и помещают в 1 мл 20 мМ Na 2 HPO 4 , 0,5M NaCl и повторно центрифугируют, как описано выше. Процедуру повторяют дважды перед промыванием и центрифугированием жирного слоя дважды в 750 мкл 20 мМ Na 2 HPO 4 . Слитый белок 4404 элюируют из масляного тельца в подосадочную жидкость промыванием конечного жирного слоя 5 раз в 750 мкл 20 мМ муравьиной кислоты, рН 4,1, при центрифугировании при 10000 g между каждой промывкой. Собранные элюированные фракции (подосадочные жидкости) объединяют и нейтрализуют 2н. NaOH до рН 8,0. Весь раствор охлаждают до температуры -80 °С до замерзания и лиофилизируют в течение ночи для концентрации слитого белка. Лиофилизированный образец повторно суспендируют в 500 мкл 50 мМ Трис-HCl, рН 8,0. Повторно суспендированный слитый белок 4404 затем опресняют на колонке NAP-5 (Amersham Pharmacia Biotech Ab, Uppsala, Sweden) и повторно заменяют буфером (50 мМ Трис-HCl, рН 8,0). Опресненную фракцию затем снова замораживают и лиофилизируют в течение ночи для концентрации. Конечный концентрированный образец повторно суспендируют в конечном объеме 105 мкл дважды дистиллированной Н 2 О. Результаты элюирования представлены на фиг. 6. На фиг. 6 изображен анализ окрашенного кумасси SDS-PAGE (15%) препаратов масляных телец до элюирования (-ОВ), препаратов ОВ после элюирования муравьиной кислотой (-ОВ') и концентрированного элюированного материала (-Е). Стрелки указывают на положение миграции слитого полипептида. Контрольная группа дикого типа практически не содержит какие-либо основные белки после элюирования, в то время как концентрированный материал 4404 содержит слитый белок, некоторое количество укороченных продуктов (возможно, гидролизованный слитый белок) и, возможно, некоторые коэлюированные альбумины. Расщепление и ВЭЖХ анализ 4404, экспрессируемого семенами Arabidopsis. Концентрированный образец повторно суспендируют в 105 мкл дважды дистиллированной воды и содержание белка оценивают с помощью теста для белка ВСА согласно инструкции производителя (Pierce, Rockford, IL, USA). Затем образцы расщепляют трипсином (1:300 трипсин:общий белок, в 50 мМ Трис-HCl, рН 8,0, на льду в течение 90 мин). Реакции останавливают добавлением 10-кратного молярного избытка TLCK (N-п-тозил-L-лизин хлорметилкетона). Затем все реакционные смеси фильтруют через 0,2 мкм фильтры (Aerodisc ®, 13 мм фильтр для шприца с 0,2 мкм мембраной Supof ®, Pall Corporation, Ann Arbor, MI, USA) и анализируют ВЭЖХ с обратимой фазой (ОФ) с применением С18 колонки (Zorbax 300SB-С18, Agilent Technologies, Waldbronn, Germany). Образцы загружают в колонку и элюируют со скоростью 1,0 мл/мин с применением 19-минутного линейного градиента 5-50% (об./об.) ацетонитрила в 0,1% (об./об.) TFA. Хроматограмма этого анализа представлена на фиг. 7. Следы показывают расщепленный трипсином продукт из слитого белка 4404 со свойствами на колонке, практически идентичными стандарту человеческого инсулина (время удержания 17,011 и 17,179 мин соответственно). ВЭЖХ фракцию собирают на 17,0-17,5 мин и анализируют масс-спектрометрией PSD MALDI/TOF с применением масс-спектрометра Voyager-DE STR (Applied Biosystems). MC анализ, проведенный с помощью услуги Биоаналитической спектроскопии, предлагаемой NRC-Plant Biotechnology Institute, Saskatoon, Saskatchewan, Canada. Повторное растворение расщепленного продукта 4404, очищенного ВЭЖХ, как описано выше, показано на фиг. 8В по сравнению со стандартом человеческого инсулина, показанного на фиг. 8А. Полученная масса расщепленного слитого белка 4404 с трипсином составляет 6191,51 Да. Расхождение между стандартом человеческого инсулина (фиг. 8А) и расщепленным продуктом 4404 (фиг. 8В) соответствует Des-B 30 инсулину с KDEL сигналом, удерживаемым на цепи А расщепленного продукта (des-В 30 инсулин-KDEL). Пример 4. Расщепление pSBS4405 и очистка ВЭЖХ. Элюирование из масляного тельца. Слитый белок (OLEO-Klip8-Klip27-MI) может быть частично очищен получением препаратов масляных телец, как описано выше. Приблизительно 1 г трансгенных семян гомогенизируют в 12 мл в буфере экстрагирования (0,4М сахарозы, 0,5М NaCl, 50 мМ Трис-HCl, рН 8,0) и центрифугируют при 10000 g в течение 10 мин, жирные слои удаляют и помещают в 1 мл 50 мМ Трис-HCl, рН 8,0, 0,5М NaCl и повторно центрифугируют, как описано выше. Процедуру повторяют дважды, перед промыванием и центрифугированием жирного слоя дважды в 750 мкл 50 мМ Трис-HCl, рН 8,0. Препарирование масляных телец в результате дает удаление большинства фоновых белков. Типовой профиль белков препарата масляных телец из трансгенных семян Arabidopsis, экспрессирующих 4405 конструкцию, показан на фиг. 9. Расщепление и ВЭЖХ анализ 4405, экспрессируемого семенами Arabidopsis. Общее содержание белка из повторно суспендированных масляных телец оценивают путем солюбилизации фракции препаратов (5 мкл), разбавленной в 10 раз в 2% SDS, 50 мМ Трис-HCl, рН 8,0, кипяченой в течение 5 мин и центрифугированной в течение 3 мин при 10000 g. Затем содержание белка оценивают с помощью теста для белка ВСА согласно инструкции производителя (Pierce, Rockford, IL, USA). Затем образцы расщепляют трипсином (1:300 трипсин: общий белок, в 50 мМ Трис-HCl, рН 8,0, на льду в течение 90 мин) для выделения Klip27-MI фрагмента из слитого белка. Реакции останавливают добавлением 10-кратного молярного избытка TLCK (N-п-тозил-L-лизин хлорметилкетона). Образцы центрифугируют при 10000 g в течение 10 мин и подосадочные жидкости всех реакционных смесей фильтруют через 0,2 мкм фильтры (Aerodisc ®, 13 мм фильтр для шприца с 0,2 мкм мембраной Supof ®, Pall Corporation, Ann Arbor, MI, USA). На фиг. 9 изображен анализ окрашенного кумасси SDS-PAGE (15%) общего экстрагируемого белка семян и белка, полученного из масляных телец (ОВ) из колоний клеток, экспрессирующих 4405, по сравнению с диким типом (нерекомбинантным) семян. Стрелки указывают на положение миграции слитого полипептида. Подосадочные жидкости затем анализируют ВЭЖХ с обратимой фазой (ОФ) с применением С18 колонки (Zorbax 300 SB-C18, Agilent Technologies, Waldbronn, Germany). Образцы загружают в колонку и элюируют со скоростью 1,0 мл/мин с применением 19-минутного линейного градиента 5-50% (об./об.) ацетонитрила в 0,1% (об./об.) TFA. Хроматограмма этого анализа представлена на фиг. 10. Следы показывают расщепленный трипсином продукт из слитого белка 4405 со свойствами на колонке, практически идентичными стандарту человеческого инсулина (время удержания 17,220 и 17,179 мин соответственно). ВЭЖХ фракцию собирают на 17,0-17,5 мин и анализируют масс-спектрометрией PSD MALDI/TOF с применением масс-спектрометра Voyager-DE STR (Applied Biosystems). МС-анализ, проведенный с помощью услуги биоаналитической спектроскопии, предлагаемой NRC-Plant Biotechnology Institute, Saskatoon, Saskatchewan, Canada. Как показано на фиг. 11, полученная масса расщепленного слитого белка 4405 с трипсином составляет 5706,30 Да. Расхождение между стандартом человеческого инсулина (фиг. 8А) и расщепленным продуктом 4405 (фиг. 11) соответствует Des-В 30 инсулину (Des-В 30 инсулин). Des-В 30 инсулин представляет собой продукт, ожидаемый при коррекции созревания трипсина слияния 4405. Пример 5. Очистка расщепленного трипсином MI с применением AKTA Explorer (FPLC). Очистка отщепленного MI от 4405 также обеспечивает частичную очистку от реакций расщепления более высокого уровня анионным обменом (Mono Q FF 1 мл, Amersham Pharmacia) на AKTA Explorer (Amersham Pharmacia). Реакции расщепления проводят с масляными тельцами 4405, как описано выше, полученными из 30 г трансгенных семян. Подосадочные жидкости из реакций расщепления либо фильтруют через 0,2 мкм фильтры, либо концентрируют лиофилизацией на Savant Speed Vac. Фильтрованные реакционные смеси образцов могут быть нанесены на колонку непосредственно, но концентрированные образцы требуют удаления солей для эффективного связывания колонки. Концентрированные образцы могут быть опреснены пропусканием расщепленного материала через колонку PD-10 (Amersham Pharmacia), диализом или разбавлением до концентрации соли, эквивалентной или менее чем 5 мС/см. Опресненные образцы уравновешивают 20 мМ Трис-HCl, рН 6,5. Образцы отделяют с применением пошагового градиента с NaCl 0-40% NaCl со скоростью потока 1 мл/мин. Определение проводят при 214 нм (определение при 280 нм относительно слабое из-за низкого содержания ароматических аминокислот в инсулине). Растворитель А состоит из 20 мМ Трис-HCl, рН 6,5, а растворитель В состоит из 20 мМ Трис-HCl, рН 6,5, 1,0М NaCl. Фракции (1 мл), элюированные при той же проводимости, что и стандарт инсулина Roche, между 7-35 мС/см, собирают (см. фиг. 12). На фиг. 12 представлена хроматограмма расщепленных трипсином препаратов масляных телец 4405 (пунктирная линия) по сравнению со стандартом человеческого инсулина (сплошная линия). Присутствие инсулина подтверждается в собранных фракциях с помощью ВЭЖХ, ELISA или вестерн-блоттинга (данные не показаны). Собранные образцы концентрируют лиофилизацией и применяют для биотестирования инсулина, описанного в примере 6. Пример 6. Тестирование переносимости инсулина: биотестирование на самцах мышей C57BI/6 (В6). Это биотестирование проводят для определения in vivo действия рекомбинантного, полученного в растениях (DesB 30 IN) из расщепленного трипсином 4405, по сравнению с человеческим инсулином. Уровни глюкозы в плазме мышей В6 определяют до и после внутрибрюшинной инъекции стандартов инсулина, отрицательного контроля и SBS инсулина. Пятнадцать мышей C57BI/6 (В6) возрастом приблизительно 2 месяца приобретают у Jackson Laboratories (Bar Harbor, ME). Уровни глюкозы в плазме определяют автоматическим глюкометром (OneTouch Ultra, Lifescan, Johnson and Johnson). Положительный контроль включает HumulinR ® (Eli Lilly) и дрожжевой рекомбинантный стандарт человеческого инсулина от Roche. В качестве плацебо применяют физиологический раствор. Отрицательный контроль включает расщепленные трипсином масляные тельца, очищенные из дикого типа (нерекомбинантных) семян Arabidopsis, которые обрабатывают идентично рекомбинантным препаратам расщепленных трипсином масляных телец 4405. Мышей В6 помещают в клетки и кормят ad libitum при 12-часовом цикле день-ночь. Для проведения тестов на переносимость инсулина мышам вводят внутрибрюшинные инъекции (ВБ) инсулина (1 Ед/кг массы тела) и уровни глюкозы определяют через 0, 15, 30 и 60 мин с применением автоматического глюкометра. Все тесты на переносимость инсулина проводят с одинаковым интервалом каждый день (9:00). Тесты на переносимость инсулина проводят по крайней мере в течение 2 дней с промежутками времени перед введением очередного теста. Результаты тестов переносимости инсулина представлены на фиг. 13. SBS DesB 30 инсулин, полученный из семян 4405 (закрашенные ромбы), действует практически идентично (статистически не отличается, р <0,05) с HumulinR ® (пустые квадраты) и инсулином от Roche (пустые треугольники) после введения всего курса исследования. Все тестированные инсулины значительно снижают уровни глюкозы в плазме (р <0,05) по сравнению с плацебо физиологическим раствором (пустые круги) и расщепленных трипсином масляных телец из семян дикого типа Arabidopsis (закрашенные круги) (отрицательный контроль). Пример 7. Получение pSBS4401: PRS-Klip27-MI слитого белка. Один из изучаемых слитых белков начинается с последовательности, связанной с патогеном табака (PRS) (Sijmons et al., 1990, Bio/technology, 8: 217-221), которая служит в качестве сигнального пептида для целевой экспрессии в ER котрансляционным методом. Сразу же после нее идет расщепляемый трипсином пропептид (Klip27), полученный из ТА57 пропептида дрожжей (Kjeldsen et al., 2001, Biotechnology and Genetic Engineering Reviews 18: 89-121). Затем идет мини-инсулин (MI), описанный у Kjeldsen et al. (2001). Основная цепь этой плазмиды, pSBS4055, основана на бинарном векторе растения, pPZP200, описанном у Hajdukiewicz et al. (Plant Molecular Biology, 1994, 25: 989-994). Вместо описанного сайта множественного клонирования, pat ген, придающий растению-хозяину устойчивость к фосфинотрицину (Wohlleben et al., 1988, Gene 70: 25-37), управляемый промотором/терминатором убиквитина из Petroselinum crispum (Kawalleck et al., 1993, Plant Mol. Bio., 21: 673-684), был введен между левой и правой пограничными последовательностями. В дополнение к этой кассете промотор/терминатор β-фазеолина из Phaseolus vulgaris (Slightom et al., 1983, Proc. Natl. Acad. Sci. USA 80: 1897-1901), управляющий PRS, был субклонирован. Стандартную PCR (Horton et al., 1989, Gene 77: 61-68) применяли для слияния синтетической PRS-кодирующей последовательности с присоединенными сайтами SphI/HindIII рестрикционной эндонуклеазы с 3'-конца промотора фазеолина с получением pSBS4011. Последовательность Klip27-MI синтезируют из четырех частично перекрывающихся олигонуклеотидов, в которые введен кодон Arabidopsis thaliana, применяемый для повышения эффективности трансляции в системах экспрессии, основанных на растениях. Олигонуклеотиды 1324 (GAAGAAGGAGAGCCTAAGTTTGTTAATCAACATCTTTGTGGATCTCATCTTGTTGAGGCTCTCTACCTTG) - SEQ ID NO:177 и 1323 (CCTTAGGAGTGTAGAAAAATCCTCTTTCTCCACACACAAGGTAGAGAGCCTCAACA) - SEQ ID NO:178 гибридизируют в их комплементарном наложении 20 нуклеотидов и продлевают с получением 5'-конца Klip27-MI слияния, то же самое проводят с олигонуклеотидами 1322 (CTAAGGCTGCTAAGGGAATTG) - SEQ ID NO:179 и 1321 (AAGCTTCAGTTGCAATAGTTCTCCAATTGGTAAAGTGAGCAAATAGAAGTGCAACATTGTTCAACAATTCCCTTAGCAGCCTT) - SEQ ID NO:180 с получением 3'-конца. Две половины лигируют с последующим рестрикционным расщеплением с Bsu36I с получением полной Klip27-MI кодирующей последовательности. PCR такого слияния гена с применением праймеров 1363 (GCATGCCCAACCAATTGATGACACTG) - SEQ ID NO:184 и 1329 (AAGCTTCAGTTGCAATAGTTC) - SEQ ID NO:183 присоединяют 5' сайт расщепления SphI и 3' HindIII рестрикционной эндонуклеазы для последующего лигирования в SphI/HindIII-разрыв pSBS4011 (как описано выше). Результатом является плазмида pSBS4401: последовательность ДНК (SEQ ID NO:188), кодирующая слитый белок PRS-Klip27-MI (SEQ ID NO:189), помещенный в бинарный вектор под контролем экспрессии промотором/терминатором фазеолина. Промотор фазеолина контролирует специфически временную и специфическую к тканям экспрессию трансгена во время развития семян. После подтверждения целостности кДНК кодирования для слитого белка анализом последовательности плазмиду pSBS4401 трансформируют в штамм Е.coli DH5 α для получения высокого уровня экспрессии. Выделенную плазмиду ДНК (100 нг) смешивают на льду со 100 мкл компетентных клеток DH5 α в течение 20 мин. Затем клетки подвергают тепловому удару при температуре 42 °С в течение 45 с и возвращают в лед на 2 мин. Затем добавляют 1 мл среды SOC и клетки инкубируют при температуре 37 °С на энвиро-шейкере при 225 об/мин в течение 1 ч перед помещением трансформированных клеток на планшеты с LB-спектиномицином (10 г/л триптона, 5 г/л экстракта дрожжей, 5 г/л NaCl, 15 г/л агара) и инкубированием в течение ночи при температуре 37 °С. Одну колонию используют для инокулирования 5 мл LB-спектиномицинового бульона. Эти культуры выращивают в течение ночи при температуре 37 °C. Рекомбинантную плазмиду выделяют из 1 мл среды, выстаиваемой в течение ночи, согласно получению мини-препаратов Qiagen. Выделенную плазмиду затем применяют для трансформации компетентного штамма Agrobacterium ЕН101 (Hood et al., 1986; J. Bacteriol. 144: 732-743) электропорацией (25 мкФ, 2,5 кВ, 200 Ом). Рекомбинантные Agrobacteruim помещают на планшеты с АВ-спектиномицином/канамицином (20 × АВ соли, 2М глюкозы, 0,25 мг/мл FeSO 4 ∙7H 2 O, 1M MgSO 4 , 1M CaCl 2 ) и одну колонию применяют для инокулирования 5 мл АВ-спектиномицинового/канамицинового бульона. Эти культуры выращивают в течение ночи при температуре 28 °С. Рекомбинантные Agrobacteruim затем применяют для трансформации растений Arabidopsis thaliana методом погружения цветков (Clough et al., 1998, Plant J., 16: 735-743), как описано в примере 1. Уровни экспрессии слитого белка Klip27-MI (4401) определяют в трансгенных созревших семенах Arabidopsis thaliana с применением методики примера 2 выше. Трансгенный продукт не был найден в клеточных экстрактах зрелых семян. Пример 8. Получение pSBS4409: OLEO-человеческий проинсулин (OLEO-hPIN) слитого белка. Этот слитый белок начинается с 18 кДа олеозина из Arabidopsis thaliana, далее продолжается геном, кодирующим человеческий проинсулин (hPIN). Экспрессия этого слитого белка нацелена на новые масляные тельца, образующиеся в процессе развития эмбриона. Основная цепь этой плазмиды, pSBS4008, основана на бинарном векторе растения, pPZP200, описанном у Hajdukiewicz et al. (Plant Molecular Biology, 1994, 25: 989-994). Вместо описанного сайта множественного клонирования, pat ген, придающий растению-хозяину устойчивость к фосфинотрицину (Wohlleben et al., 1988, Gene 70: 25-37), управляемый промотором/терминатором убиквитина из Petroselinum crispum (Kawalleck et al., 1993, Plant Mol. Bio., 21: 673-684), был введен между левой и правой пограничными последовательностями. В дополнение к этой кассете промотор/терминатор β-фазеолина из Phaseolus vulgaris (Slightom et al., 1983, Proc. Natl. Acad. Sci. USA 80: 1897-1901), управляющий геномной последовательностью Arabidopsis 18 кДа олеозина, был субклонирован. Стандартную PCR (Horton et al., 1989, Gene 77: 61-68) применяли для слияния последовательности гена олеозина (минус стоп-кодон) с присоединенными сайтами NcoI и HindIII рестрикционной эндонуклеазы с 3'-конца промотора фазеолина с получением pSBS4008. Ген-HindIII NcoI-человеческого препроинсулина синтезируют в виде одинарного 335 п.н. куска с помощью аптагена с применением предпочтительного использования кодона растения. Последующее лигирование в NcoI/HindIII-разрыв pSBS4008 дает плазмиду pSBS4400: последовательность ДНК, кодирующая слитый белок олеозин-человеческий препроинсулин, помещенный в бинарный вектор под контролем экспрессии промотором/терминатором фазеолина. Плазмида pSBS4400 служит в качестве образца для получения человеческого проинсулина (hPIN) стандартной PCR с применением pfu ДНК полимеразы с праймерами, направленными против 5'-конца (1457 олиго TTCGTGAACCAACACTTG - SEQ ID NO:190) и 3'-конца (1458 олиго AAGCTT TCAGTTACAGTAGT - SEQ ID NO:191), включая сайт HindIII существующей области проинсулина вектора. Второй фрагмент увеличивают с применением pfu ДНК полимеразы с праймером, направленным против доступного сайта SphI (олиго 1455 GCATGC ATGTGTTGAGC - SEQ ID NO:192) к 3'-концу гена олеозина Arabidopsis (олиго 1456 GGTAGTGTGCTGGCCA - SEQ ID NO:193) в векторе pSBS4400. После PCR продукты разделяют на агарозном геле и полосы, соответствующие 267 п.н. (hPIN-HindIII) и 360 п.н. (SphI-OLEO (3'-конец)) фрагменты очищают на геле с применением набора для экстрагирования геля (Qiagen). Два фрагмента сливают по второму кругу увеличения PCR с применением Taq ДНК полимеразы с праймерами 1455 (SEQ ID NO:192) и 1458 (SEQ ID NO:193) в сочетании с 0,01 мкМ перекрывающего мостикового PCR праймера (олиго 1459 GGTGGCCAGCACACTACCTTCGTGAACCAACACTTGTG - SEQ ID NO:194) для двух циклов с температурой отжига 58 °С с последующим 31 циклом при температуре 52 °С для увеличения 627 п.н. SphI-OLEO (3'-конец)-hPIN-HindIII фрагмента. 627 п.н. SphI-OLEO (3'-конец)-hPIN-HindIII фрагмент затем лигируют в Т/А выступ pGEMT Easy Vector System ™ (Promega) и применяют для трансформации бактерий DH5 α с получением pSBS3409 (pGEMT-SphI-OLEO (3'-конец)-hPIN-HindIII). Фрагмент SphI/HindIII из pSBS3409 заменяют фрагментом SphI/HindIII из pSBS4400. Стандартное ограничение гидролизатов в обоих pSBS3409 и pSBS4400 проводят с применением SphI/HindIII (New England Biolabs). Фрагменты разделяют на 1,5% агарозном геле и очищают с применением набора для экстрагирования геля (Qiagen). 617 п.н. SphI/HindIII фрагмент освобождают из pSBS3409 и затем лигируют в акцепторный сайт SphI/HindIII в предварительно разорванной основной цепи вектора pSBS4400 (удален внутренний фрагмент SphI/HindIII) с применением Т4 ДНК лигазы (NEB) в течение ночи при температуре 15 °С. Результатом является плазмида pSBS4409: последовательность ДНК (SEQ ID NO:195, кодирующая слитый белок OLEOSIN-hPIN (SEQ ID NO:196), помещенный в бинарный вектор под контролем экспрессии промотором/терминатором фазеолина. Промотор фазеолина контролирует специфически временную и специфическую к тканям экспрессию трансгена во время развития семян. После подтверждения целостности кДНК кодирования для слитого белка анализом последовательности плазмиду pSBS4409 трансформируют в штамм E.coli DH5 α для получения высокого уровня экспрессии. Выделенную плазмиду ДНК (100 нг) смешивают на льду со 100 мкл компетентных клеток DH5 α в течение 20 мин. Затем клетки подвергают тепловому удару при температуре 42 °С в течение 45 с и возвращают на лед на 2 мин. Затем добавляют 1 мл среды SOC и клетки инкубируют при температуре 37 °С на энвиро-шейкере при 225 об/мин в течение 1 ч перед помещением трансформированных клеток на планшеты с LB-спектиномицином (10 г/л триптона, 5 г/л экстракта дрожжей, 5 г/л NaCl, 15 г/л агара) и инкубированием в течение ночи при температуре 37 °С. Одну колонию используют для инокулирования 5 мл LB-спектиномицинового бульона. Эти культуры выращивают в течение ночи при температуре 37 °С. Рекомбинантную плазмиду выделяют из 1 мл среды, выстаиваемой в течение ночи, согласно получению мини-препаратов Qiagen. Выделенную плазмиду затем применяют для трансформации компетентного штамма Agrobacterium ЕН101 (Hood et al., 1986; J. Bacteriol. 144: 732-743) электропорацией (25 мкФ, 2,5 кВ, 200 Ом). Рекомбинантные Agrobacteruim помещают на планшеты с АВ-спектиномицином/канамицином (20 × АВ соли, 2М глюкозы, 0,25 мг/мл FeSO 4 ∙7H 2 O, 1M MgSO 4 , 1M CaCl 2 ) и одну колонию применяют для инокулирования 5 мл АВ-спектиномицинового/канамицинового бульона. Эти культуры выращивают в течение ночи при температуре 28 °С. Рекомбинантные Agrobacteruim затем применяют для трансформации растений Arabidopsis thaliana методом погружения цветков (Clough et al., 1998, Plant J., 16: 735-743), как описано в примере 1. Уровни экспрессии слитого белка OLEO-hPIN (4409) определяют в трансгенных созревших семенах Arabidopsis thaliana с применением методики примера 2 выше. Белки масляных телец, меченные окрашенным кумасси гелем, из двух характерных колоний (4409-6 и 4409-8) сравнивают с миграцией слитого белка олеозин-hPIN (как показано черной стрелкой) у нетрансформированных (wt) Arabidopsis (фиг. 14). Уровень экспрессии определяют денситометрией с получением в среднем 0,10% от общего белка семян. Этот уровень рассчитывают выше и ниже комиграции эндогенного белка с такой же молекулярной массой в нетрансформированных семенах (wt), который составляет приблизительно 0,04% от общего белка семян. Пример 9. Трансформация сафлора. Представленный протокол трансформации схож с описанным у Orlilcowska T.K., et al. ((1995), Plant Cell, Tissue and Organ Culture 40: 85-91), но с модификациями и улучшениями как для трансформации S-317, так и для применения фосфинотрицина в качестве выбранного маркера. Дезинфицируют семена вида сафлора S-317 California, которые не повреждены, поцарапаны или больны, в 0,1% HCl 2 в течение 12 мин, затем 4-5 раз ополаскивают стерильной дистиллированной водой. Проращивают стерильные семена в темноте в среде MS (Murashige Т. & Skoog F. (1962), Physiol. Plant 15: 473-497) с 1% сахарозой и 0,25% Gerlite. Инициируют культуры Agrobacterium из замороженного сырья в глицерине в 5 мл АВ минимальной жидкой среды с выбранным антибиотиком, и выращивают в течение 48 ч при температуре 28 °С. Выращивают аликвоту этой культуры, выращенной в течение ночи в 5 мл бульона Luria с выбором для трансформации. Дважды промывают 6-8 мл бактериальных клеток средой АВ и доводят до конечной плотности клеток 0,4-0,5 (OD 600 ). Удаляют двухдневные семядоли из проращенных семян, погружают в полученные клетки Agrobacterium и помещают в среду MS с 3% сахарозой, 4 мкМ N6-бензиладенина (ВА) и 0,8 мкМ нафталинуксусной кислоты (NAA). Инкубируют планшеты при температуре 21 °С в темноте. Через 3 дня переносят в ту же среду с 300 мг/л тиментина. Через еще 4 дня все культуры переносят на свет. Через 3 дня помещают эксплантаты в среду выбора с добавлением 0,5 мг/л фосфинотрицина. Для длительного вытягивания почек эксплантаты еженедельно переносят в среду MS без фитогормонов, но с увеличенным в два раза количеством KNO 3 . Срезают ростки, которые вытянулись более чем на 10 мм от исходного уровня, и отдельно выращивают для селекции. Для образования корней помещают зеленые ростки, представляющие предполагаемую трансгенную ткань, в среду MS с 2% сахарозой, 10 мкМ индолмасляной кислоты и 0,5 мкМ NAA. Переносят укоренившиеся ростки в хорошо увлажненную смесь почв и выращивают в условиях высокой влажности и 12-часового света. Пример 10. Протокол трансформации льна. Указанная методика трансформации близка к методике, описанной у Dong J. и McHugen A. (Plant Cell Reports (1991). 10: 555-560), Dong J. и McHugen A. (Plant Sciences (1993). 88: 61-71) и Mlynarova et al. (Plant Cell Reports (1994). 13: 282-285). Дезинфицируют семена льна, которые не повреждены, поцарапаны или больны, в 70% этаноле в течение 5-7 мин, затем в течение 25 мин в 50% отбеливающем растворе с Tween 20 (3-4 капли на 100 мл) при непрерывном перемешивании. Семена 5-7 раз ополаскивают стерильной дистиллированной водой. Проращивают стерильные семена на свету в среде MS (Murashige Т. & Skoog F. (1962). Physiol. Plant 15: 473-497) с 2% сахарозой и 0,3% Gerlite в сосудах Magenta. Для трансформации выращивают культуры Agrobacterium в течение ночи в бульоне АВ с добавлением подходящего антибиотика для селекции. Дважды промывают 6-8 мл выдержанных в течение ночи клеток и повторно суспендируют в 5 мл АВ бульона; добавляют 2 мл этого сырья к 98 мл индукционной среды (основная среда MS с 3% сахарозой, 5 мкМ 6-бензиламинопурина (ВА) и 0,25 мкМ альфа-нафталинуксусной кислоты (NAA)) и доводят до конечной OD 600 1,0. Отсекают гипокотильные эксплантаты и инокулируют в полученном растворе клеток Agrobacterium в течение около 4 ч (осторожно перемешивают планшеты 1-2 раза во время этого периода). После периода заражения удаляют эксплантаты из жидкой инокуляционной среды и наносят на стерильную фильтровальную бумагу. Помещают 15-20 эксплантатов в 0,7% отвержденную агаром индукционную среду в планшет для культивирования тканей. Герметично закрывают планшеты пластиковыми крышками и совместно культивируют эксплантаты в течение 48 ч при освещении (23-24 °С). Через 2 дня переносят зеленые меристематические эксплантаты в ту же среду, содержащую 300 мг/л Тиментина (среда для предварительной селекции), и закрывают пластиковыми крышками. Через 3 дня переносят культуры в указанную выше среду, содержащую 10 мг/л DL РРТ (селекция 1). Закрывают планшеты Parafilm ® и инкубируют при температуре 24 °С при освещении. Переносят культуры каждые две недели и хранят в этой среде в течение одного месяца. Для удлинения ростков переносят культуры каждые две недели в среду для селекции II (основная среда MS, содержащая 2% сахарозы, 500 мг/л буфера MES, 300 мг/л Тиментина и 10 мг/л DL РРТ) в сосудах Magenta. Предполагаемые трансформированные ростки, которые выживают после селекции, являются темно-зелеными и образуют значительное количество корней через 7-10 дней при отдельном высаживании в среду для селекции II. Переносят ростки с корнями в стерилизованную тепличную почву в небольших горшках и накрывают растения прозрачными пластиковыми крышками для акклиматизации. Для созревания переносят активно растущие растения в одногаллоновые горшки с хорошо увлажненной смесью почв в тепличных условиях. Хотя данное изобретение описано с помощью четко определенных предпочтительных примеров, должно быть понятно, что данное изобретение не ограничено описанными примерами. Наоборот, данное изобретение включает различные модификации и эквивалентные структуры, входящие в сущность и объем представленной формулы изобретения. Все публикации, патенты и заявки на патенты включены сюда в качестве ссылок полностью, в том же объеме, в котором каждая отдельная публикация, патент или заявка на патент определенно и индивидуально включена сюда в качестве ссылки. SEQ ID NO:1 и 2 являются нуклеотидной последовательностью и выведенной последовательностью аминокислот соответственно PRS-D9scFv-Klip27-MI-KDEL слитого белка в плазмиде pSBS4404. SEQ ID NO:3 и 4 являются нуклеотидной последовательностью и выведенной последовательностью аминокислот соответственно OLEO-Klip8-Klip27-MI слитого белка в плазмиде pSBS4405. SEQ ID NO:5 и 6 являются нуклеотидной последовательностью и выведенной последовательностью аминокислот соответственно PRS-MI-тетраосновный линкер-D9scFv-KDEL слитого белка в плазмиде pSBS4414. SEQ ID NO:7-145 являются известными последовательностями инсулина, которые описаны в таблице. SEQ ID NO:146-148 являются последовательностями аминокислот фрагментов С-пептида инсулина. SEQ ID NO:149 является последовательностью аминокислот тетраосновного процессингового пептида. SEQ ID NO:150-155 являются последовательностями аминокислот полипептидов, способных удерживать полипептид инсулина в ER. SEQ ID NO:156-160 являются последовательностями аминокислот полипептидов, способных удерживать полипептид инсулина в ER-производной накопительной органелле. SEQ ID NO:161 является последовательностью аминокислот PRS сигнальной последовательности. SEQ ID NO:162-171 являются последовательностями аминокислот лидерных последовательностей дрожжей и производных от них последовательностей. SEQ ID NO:172-173 являются последовательностями аминокислот промежуточных пептидов. SEQ ID NO:174 является последовательностью аминокислот последовательности Klip8. SEQ ID NO:175 является последовательностью аминокислот прямого праймера 1325, который является комплементарным к 5'-области D9scFv кДНК клона и существует для добавления сайта SphI к 5'-области для способствования последующему лигированию. SEQ ID NO:176 является последовательностью аминокислот обратного праймера 1326, который является комплементарным к 3'-области D9scFv кДНК клона и существует для добавления сайта XhoI к 3'-области для способствования последующему лигированию. SEQ ID NO:177 является последовательностью аминокислот прямого праймера 1324, который является комплементарным к области 20 нуклеотида обратного праймера 1323 и существует для образования 5'-конца слитого Klip27-MI. SEQ ID NO:178 является последовательностью аминокислот обратного праймера 1323, который является комплементарным к области 20 нуклеотида прямого праймера 1324 и существует для образования 5'-конца слитого Klip27-MI. SEQ ID NO:179 является последовательностью аминокислот прямого праймера 1322, который является комплементарным к области 19 нуклеотида обратного праймера 1321 и существует для образования 3'-конца слитого Klip27-MI. SEQ ID NO:180 является последовательностью аминокислот обратного праймера 1321, который является комплементарным к области 19 нуклеотида прямого праймера 1322 и существует для образования 3'-конца слитого Klip27-MI. SEQ ID NO:181 является последовательностью аминокислот прямого праймера 1364, который является комплементарным к 5'-области Klip27-MI последовательности и существует для добавления сайта XhoI к 5'-области для способствования последующему лигированию. SEQ ID NO:182 является последовательностью аминокислот прямого праймера 1334, который является комплементарным к 3'-области Klip27-MI последовательности и существует для добавления сайта HindI к 3'-области для способствования последующему лигированию. SEQ ID NO:183 является последовательностью аминокислот прямого праймера 1329, который является комплементарным к 3'-области Klip27-MI последовательности и существует для добавления сайта HindIII к 3'-области для способствования последующему лигированию. SEQ ID NO:184 является последовательностью аминокислот прямого праймера 1363, который является комплементарным к 5'-области Klip27-MI последовательности и существует для добавления сайта SphI к 5'-области для способствования последующему лигированию. SEQ ID NO:185 является последовательностью аминокислот прямого праймера 1515, который является комплементарным к 5'-области цепи В последовательности инсулина и существует для введения промежуточного тетраосновного сайта между исходными А- и В-цепями человеческого инсулина в сочетании с обратным праймером 1518. SEQ ID NO:186 является последовательностью аминокислот обратного праймера 1518, который является комплементарным к 3'-области цепи В инсулина и 5'-области цепи А с промежуточной последовательностью тетраосновного мини-с-пептида и существует для введения промежуточного тетраосновного сайта между исходными А- и В-цепями человеческого инсулина. SEQ ID NO:187 является последовательностью аминокислот обратного праймера 1517, который является комплементарным к 3'-области D9scFv-KDEL последовательности и существует для увеличения всего MI-тетраосновный линкер-D9scFv-KDEL с получением вставки pSBS4414. SEQ ID NO:188 и 189 являются нуклеотидной последовательностью и выведенной последовательностью аминокислот соответственно PRS-Klip27-MI слитого белка в плазмиде pSBS4 401. SEQ ID NO:190 является последовательностью аминокислот прямого праймера 1457, который является комплементарным к 5'-области В цепи последовательности инсулина и существует для получения фрагмента человеческого проинсулина (hPIN) в сочетании с обратным праймером 1591. SEQ ID NO:191 является последовательностью аминокислот обратного праймера 1458, который является комплементарным к 3'-области человеческого проинсулина (hPIN) и существует для получения человеческого про(hPIN) и добавления 3' HindIII сайта клонирования. SEQ ID NO:192 является последовательностью аминокислот прямого праймера 1455, который является комплементарным к 5'-области сайта SphI pSBS4404 и существует для увеличения гена олеозина Arabidopsis в сочетании с обратным праймером 1456. SEQ ID NO:193 является последовательностью аминокислот обратного праймера 1456, который является комплементарным к 3'-области сайта гена олеозина Arabidopsis и существует для увеличения гена олеозина Arabidopsis в сочетании с прямым праймером 1455. SEQ ID NO:194 является последовательностью аминокислот перекрывающего мостикового PCR праймера, который является комплементарным к 3'-области сайта гена олеозина Arabidopsis и 5'-конца гена человеческого проинсулина и существует для получения вставки pSBS4409 в сочетании с прямым праймером 1455 и обратным праймером 1456. SEQ ID NO:195 и 196 являются нуклеотидной последовательностью и выведенной последовательностью аминокислот соответственно OLEO-hPIN слитого белка в плазмиде pSBS4409. |