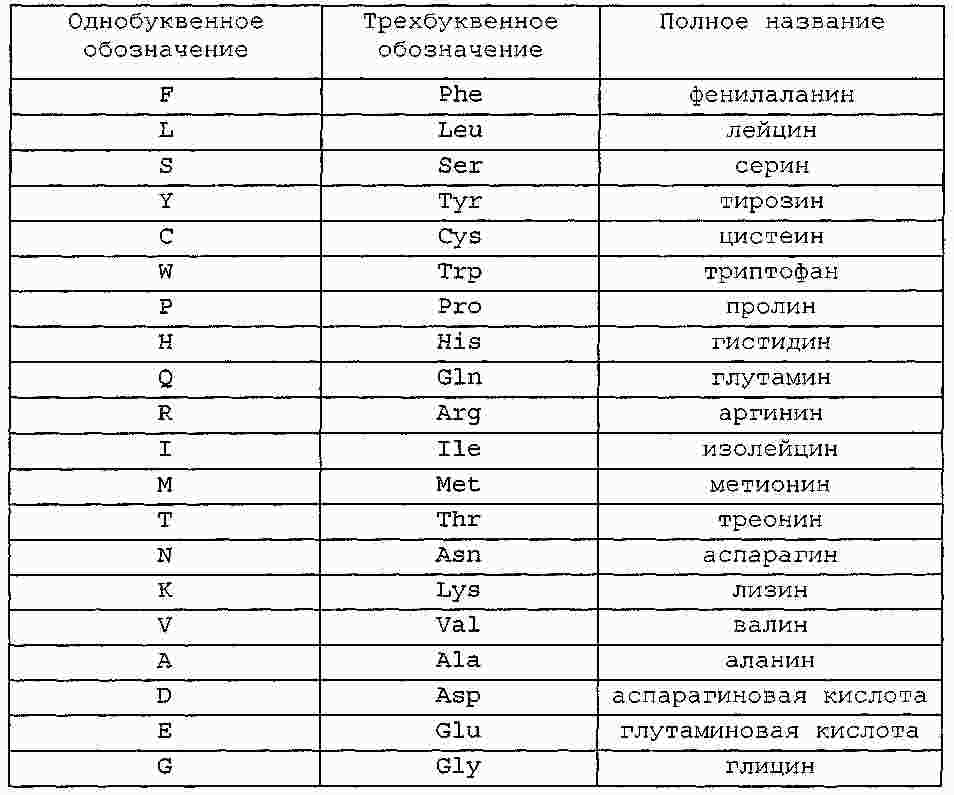
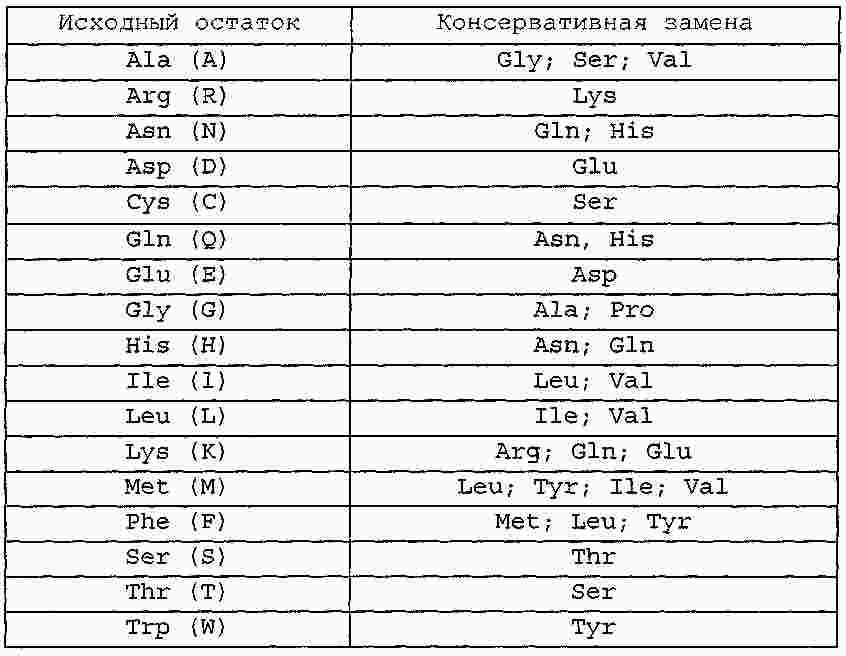
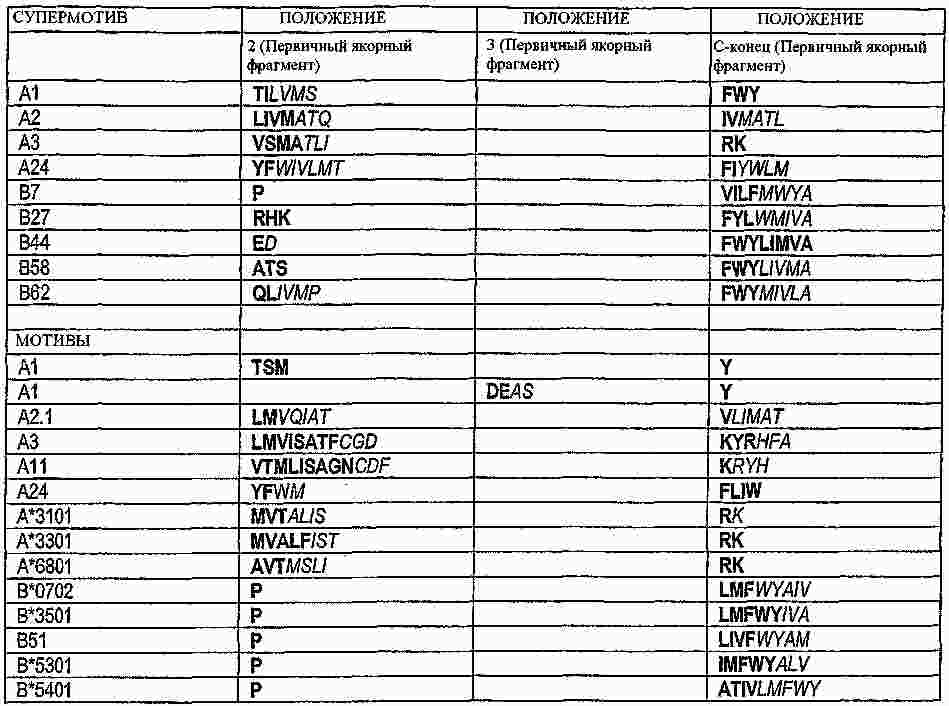
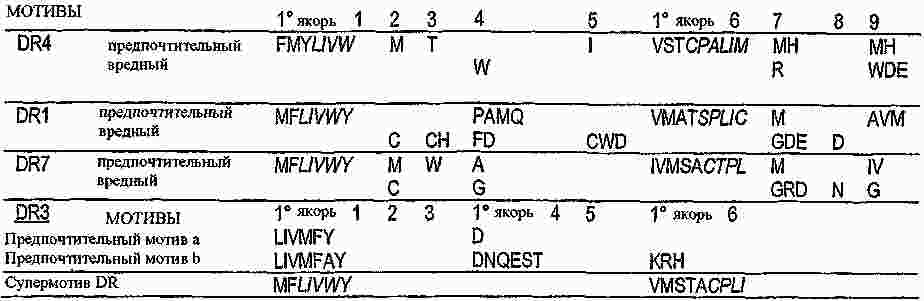
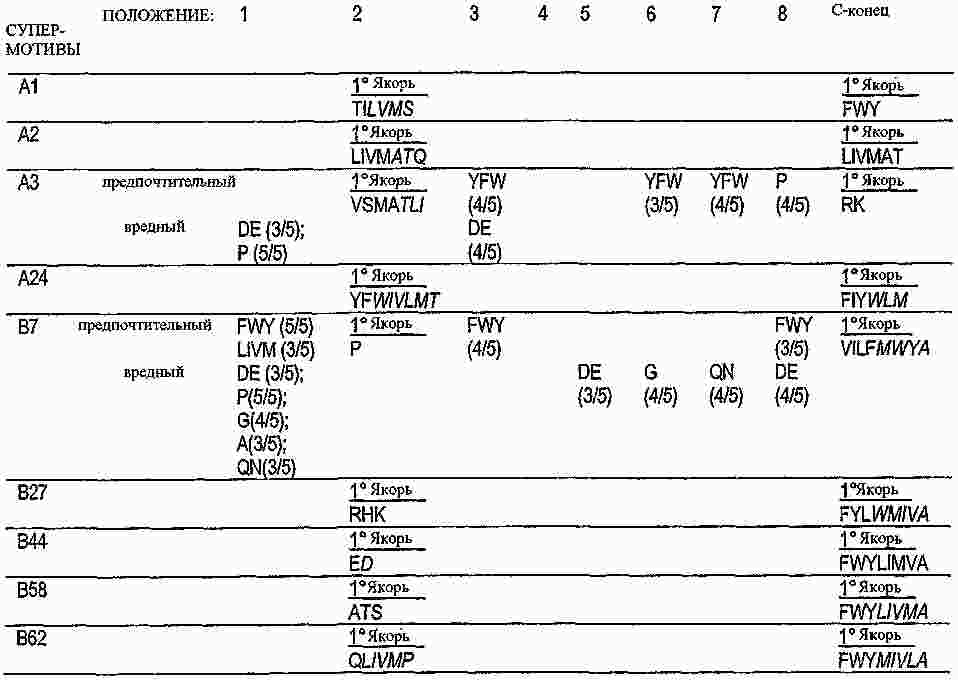
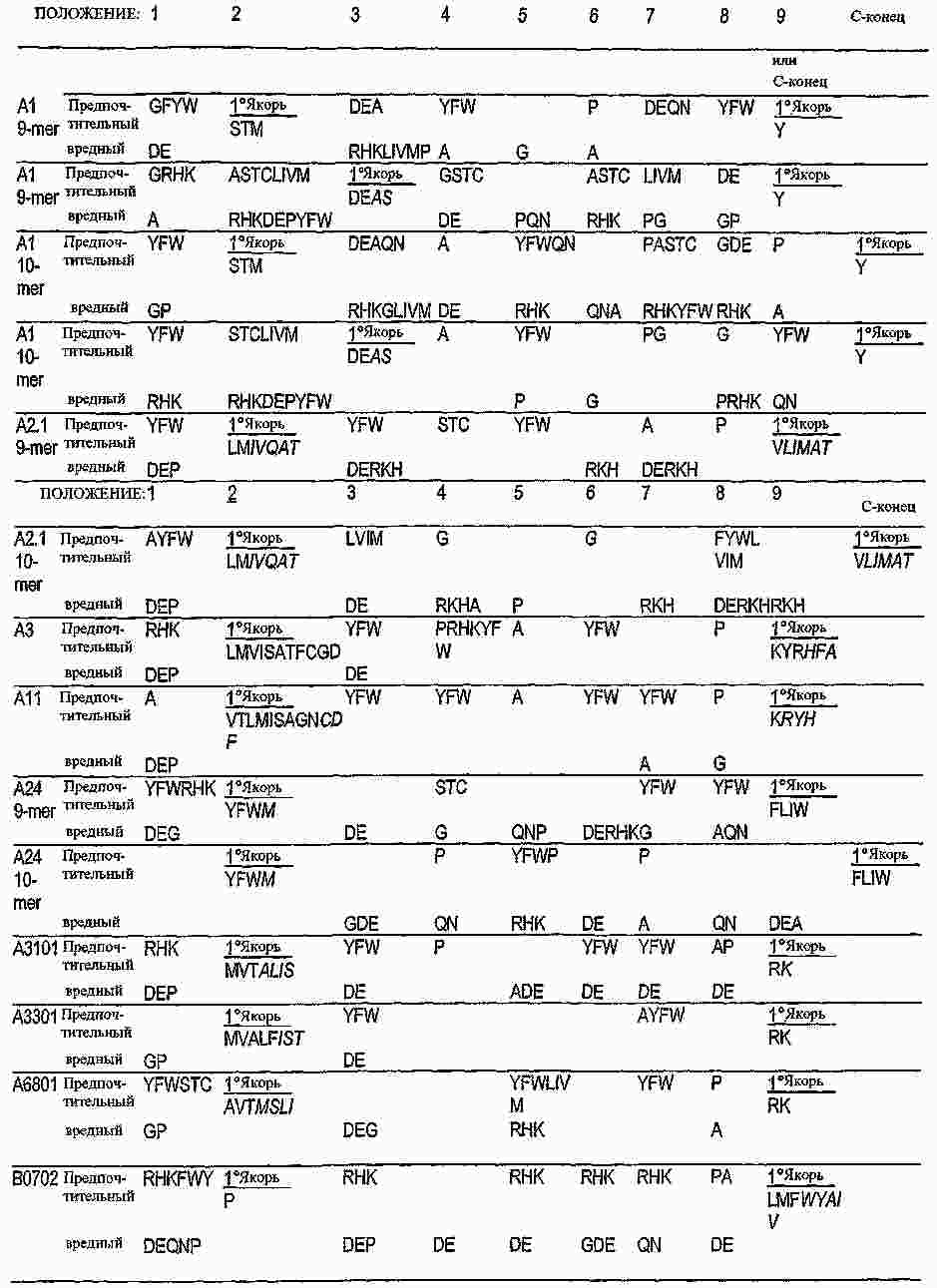
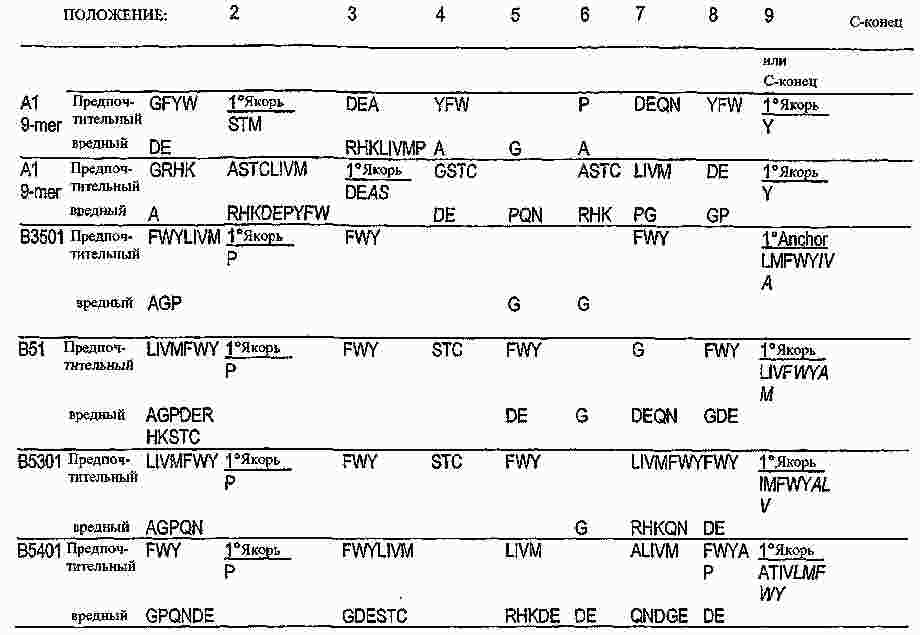
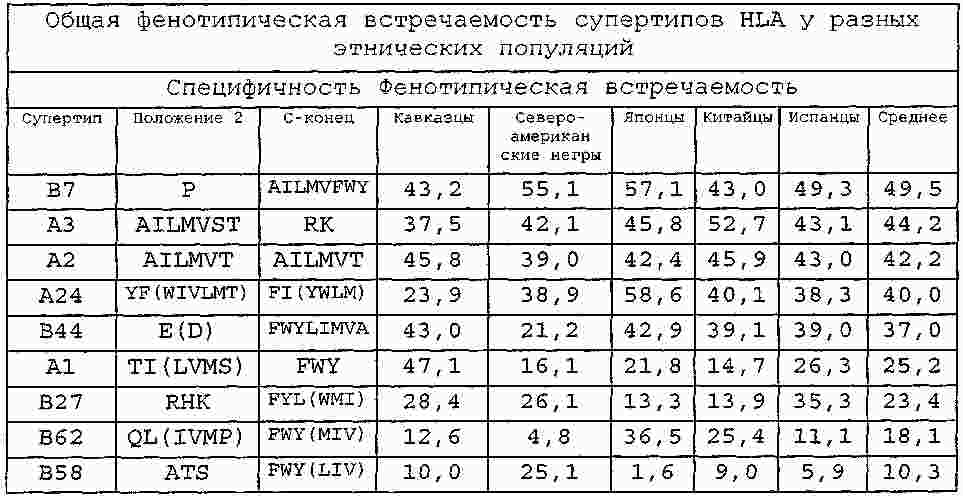
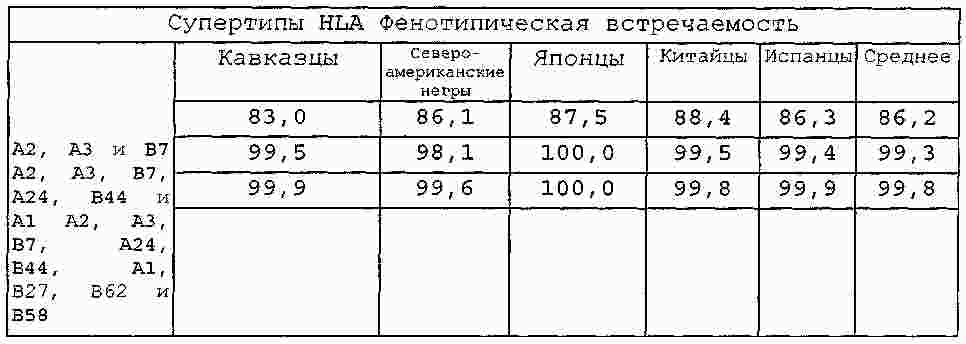
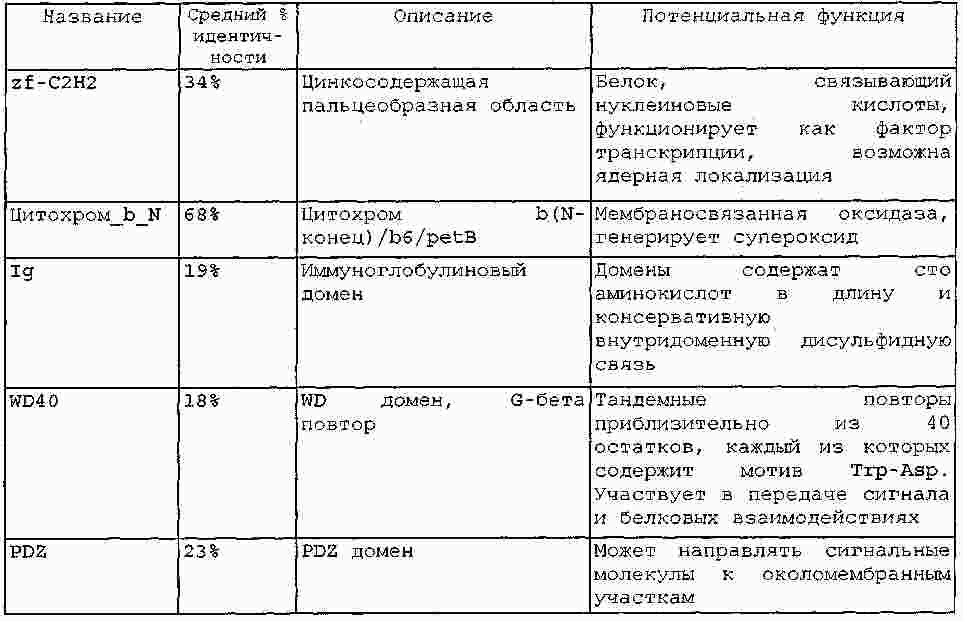
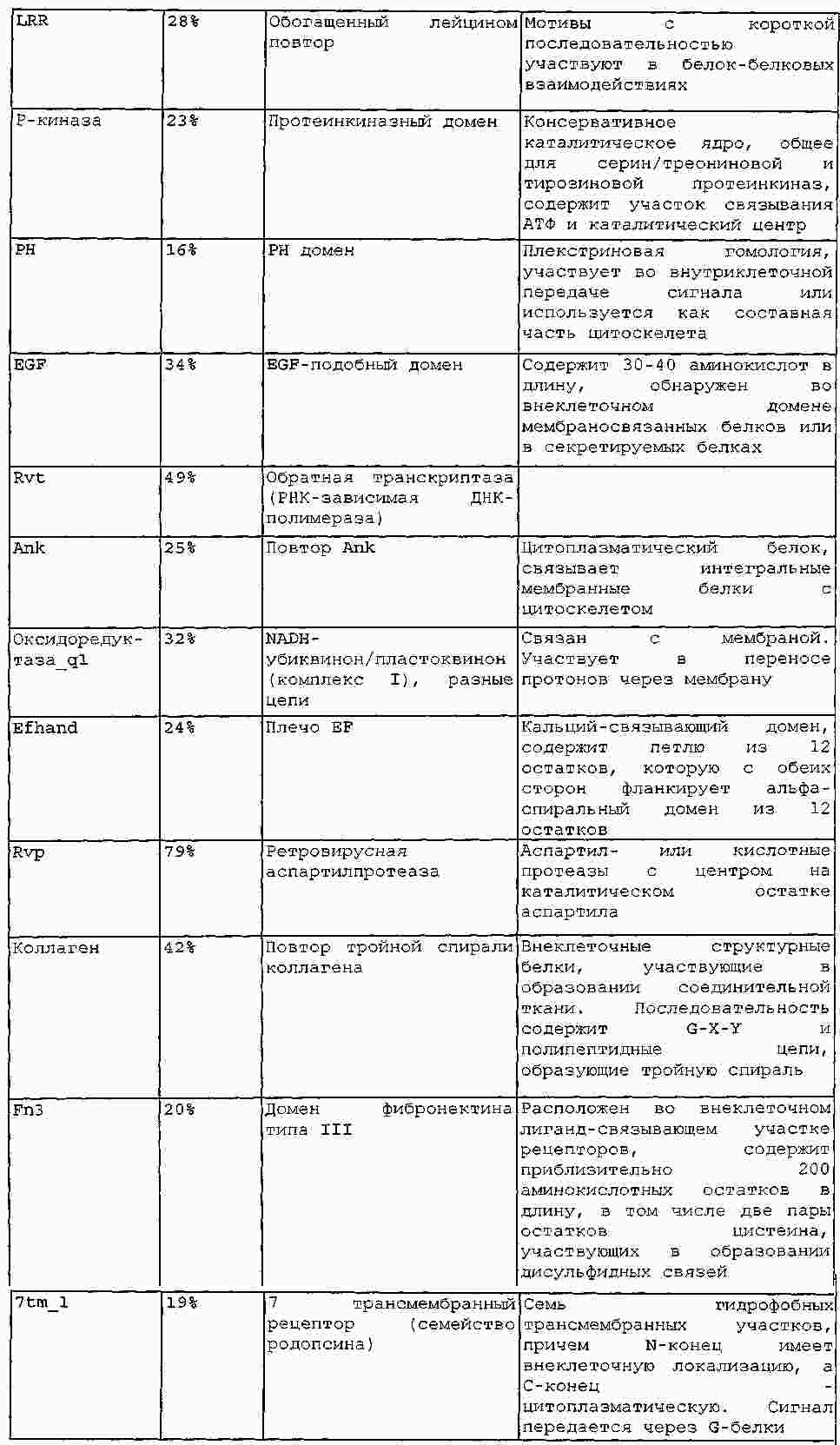
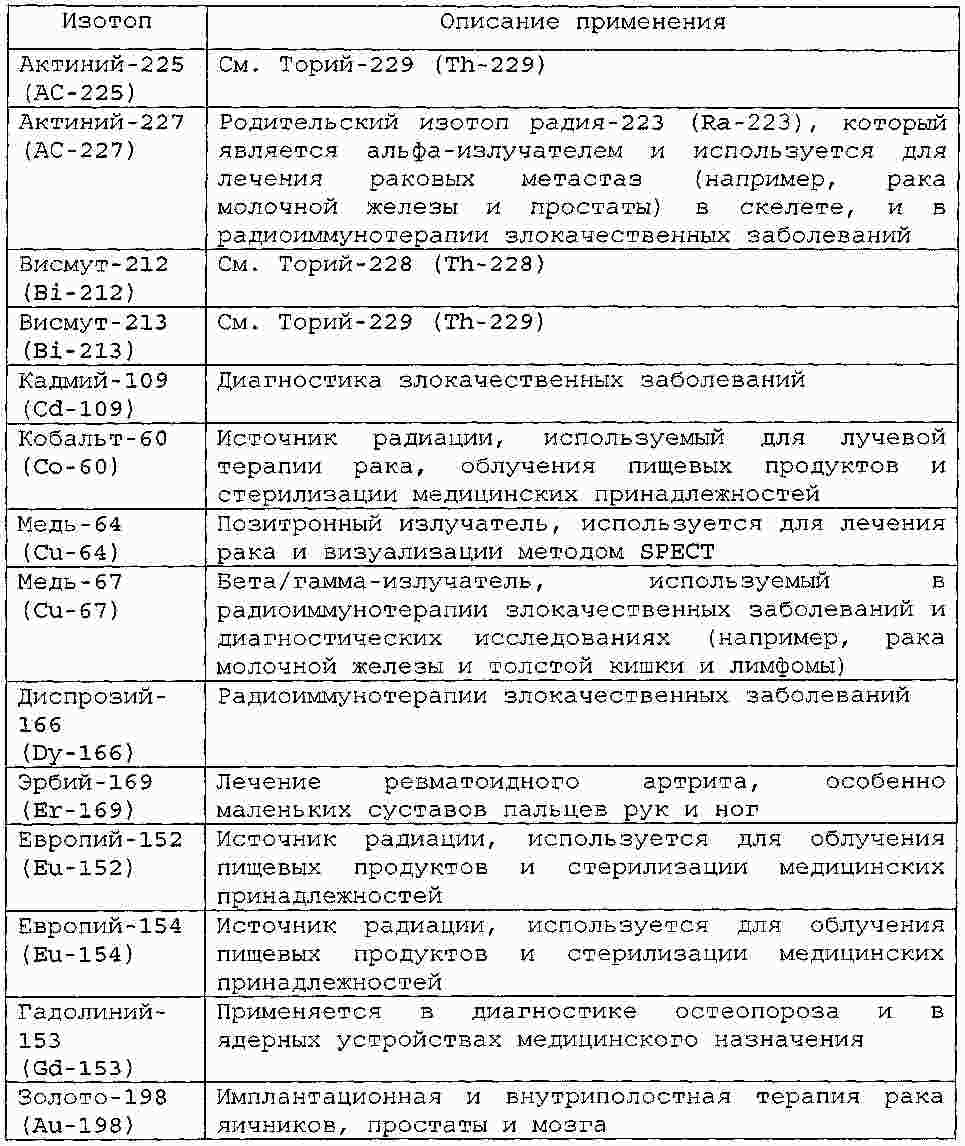
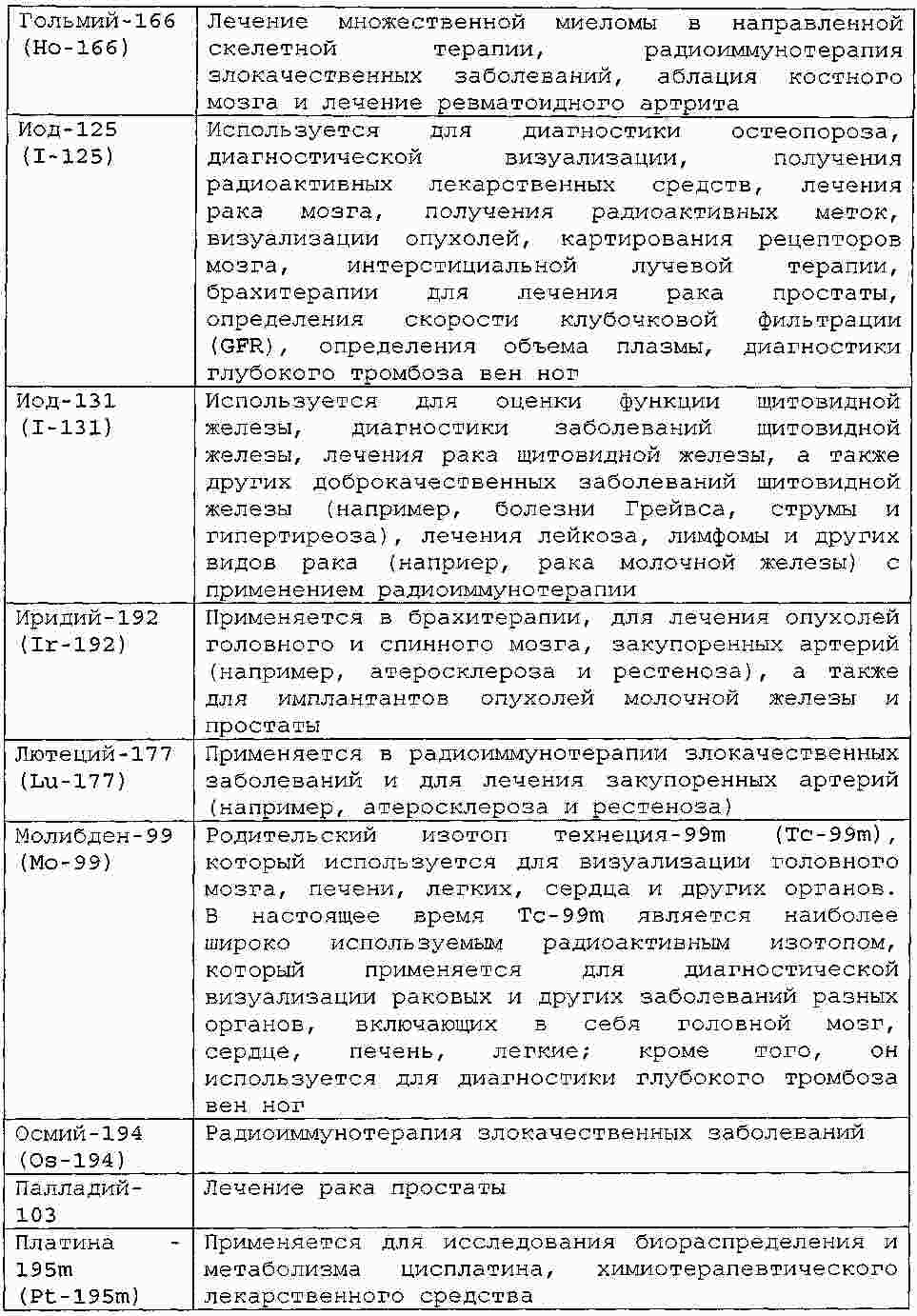
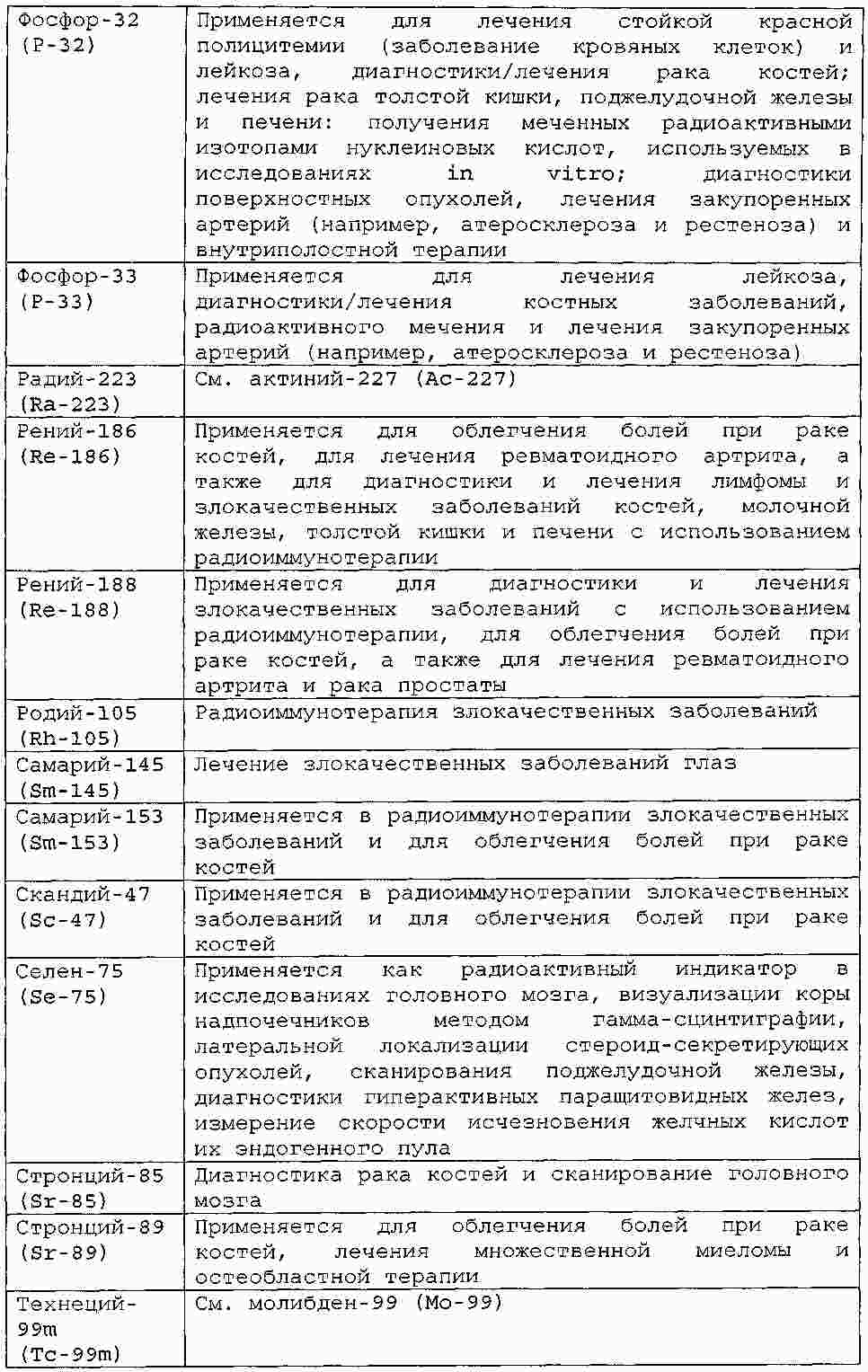
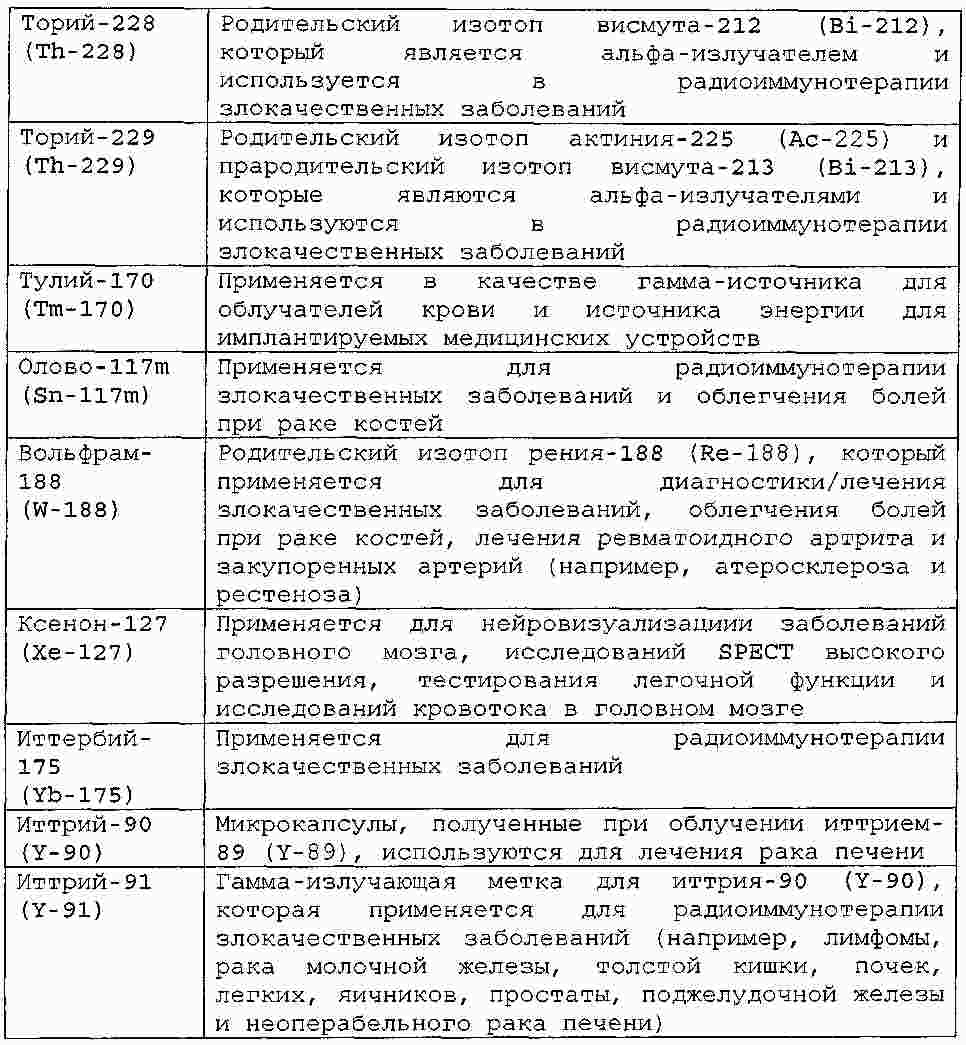
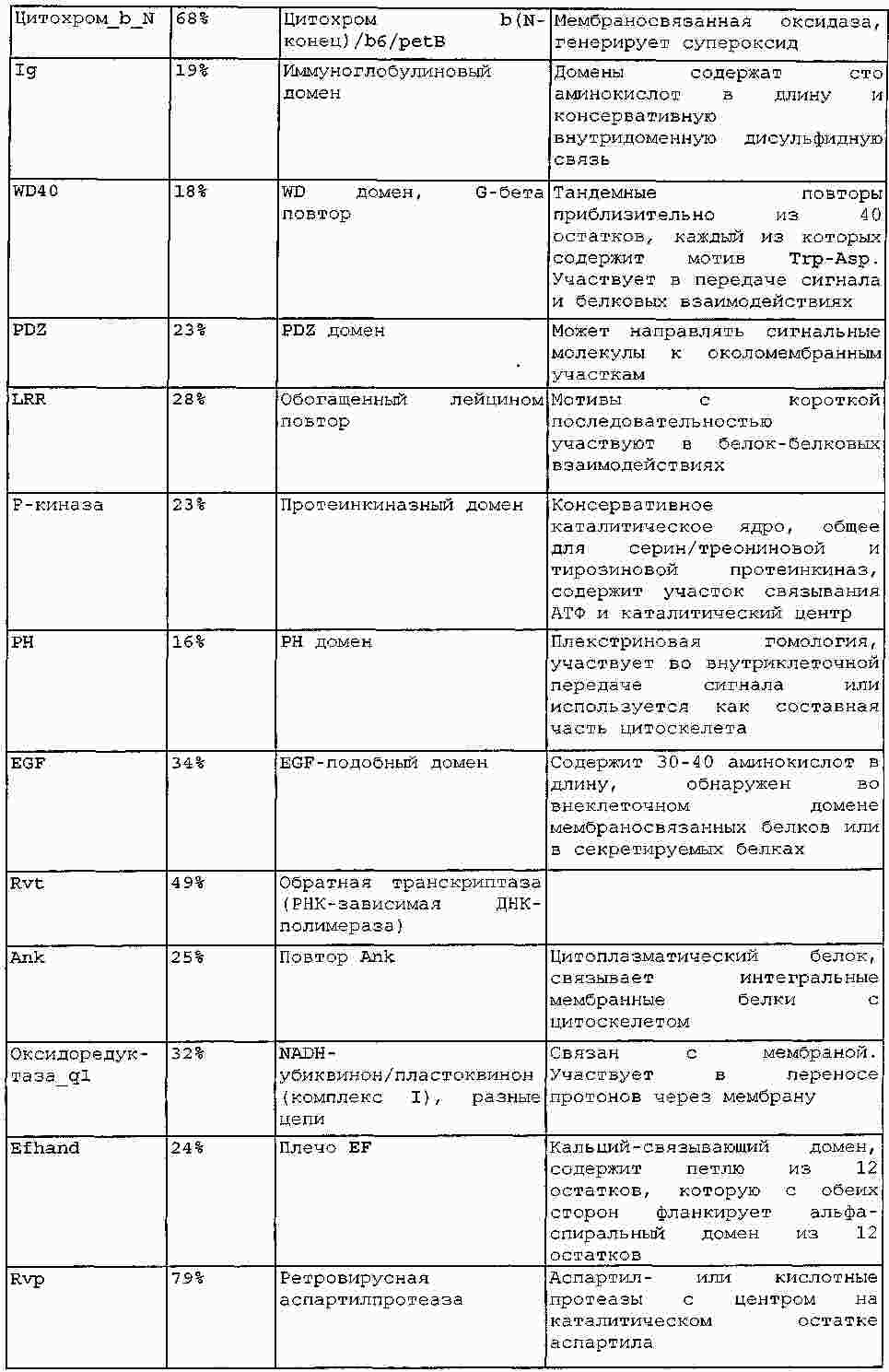
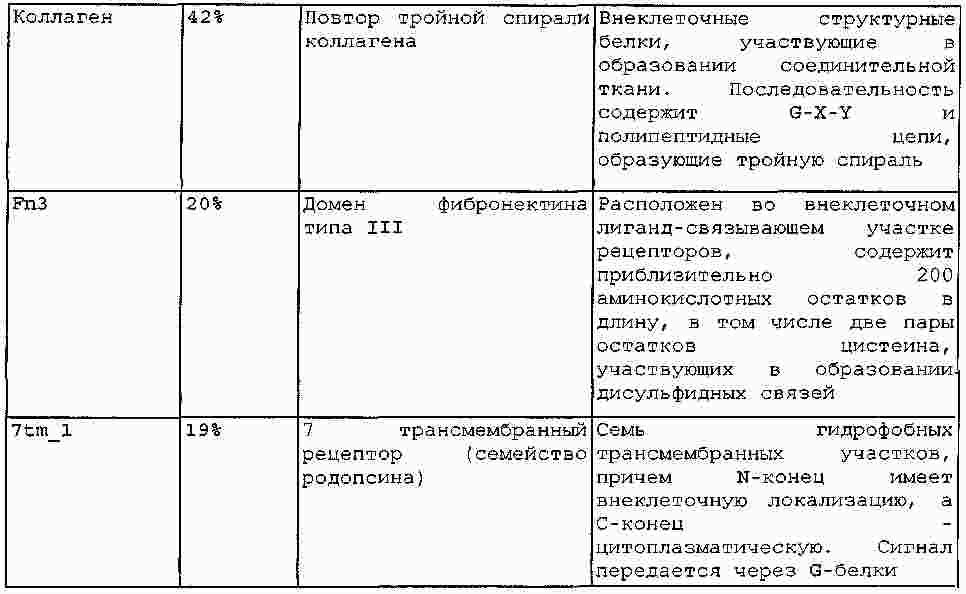
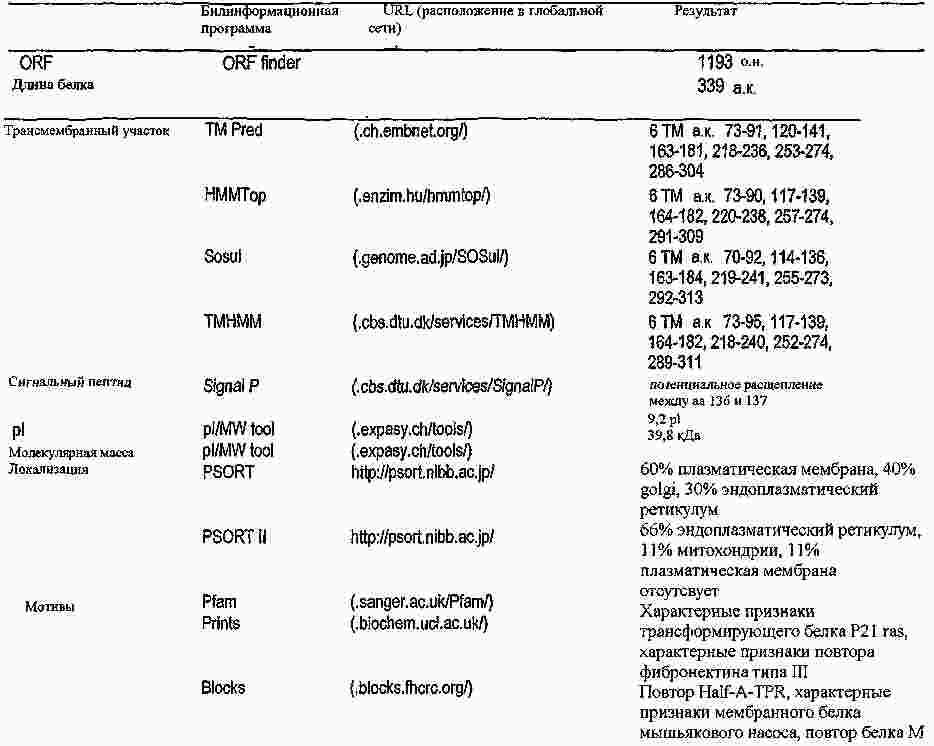
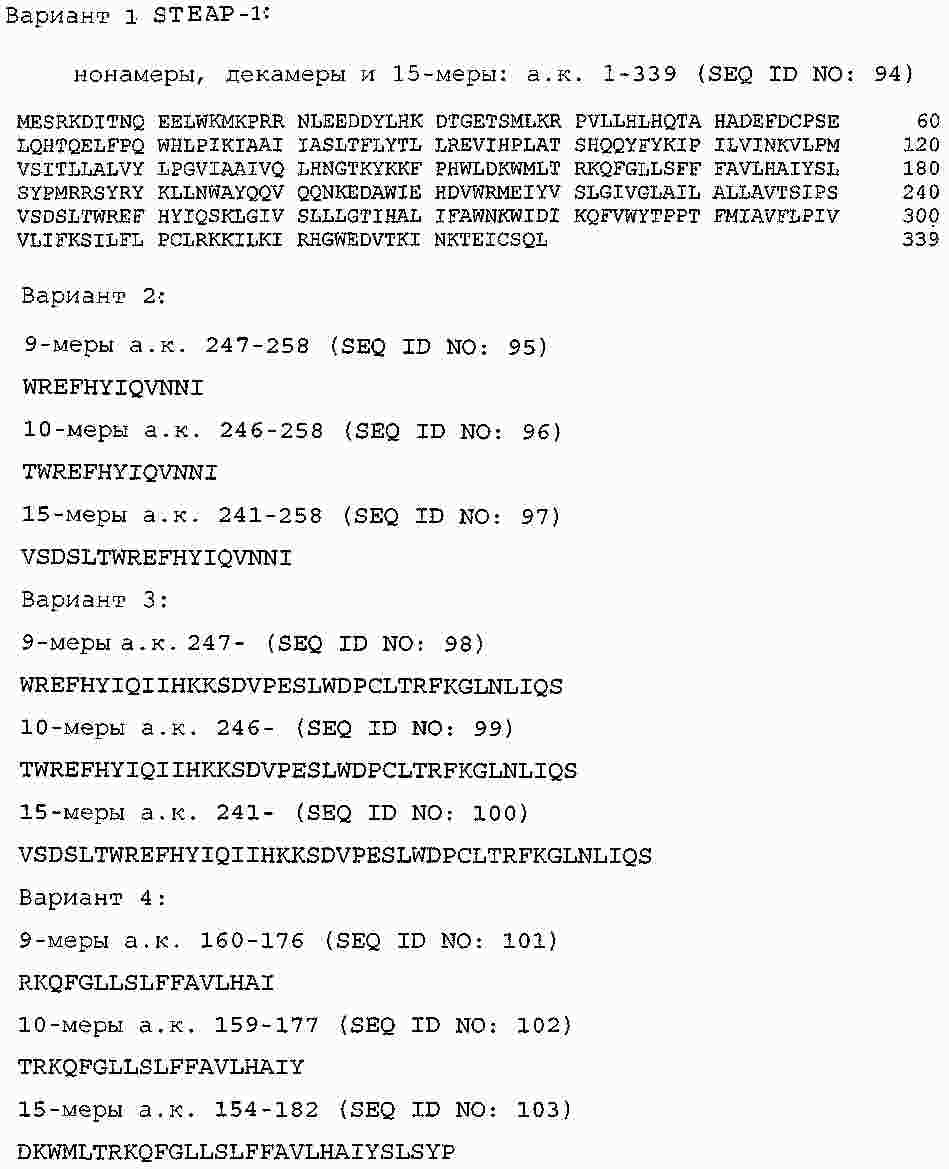
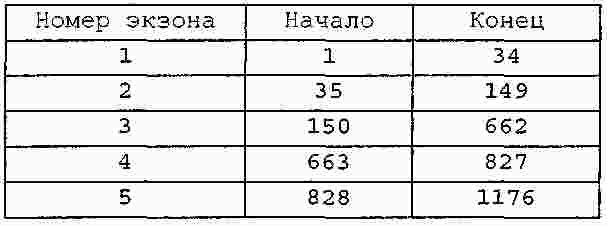
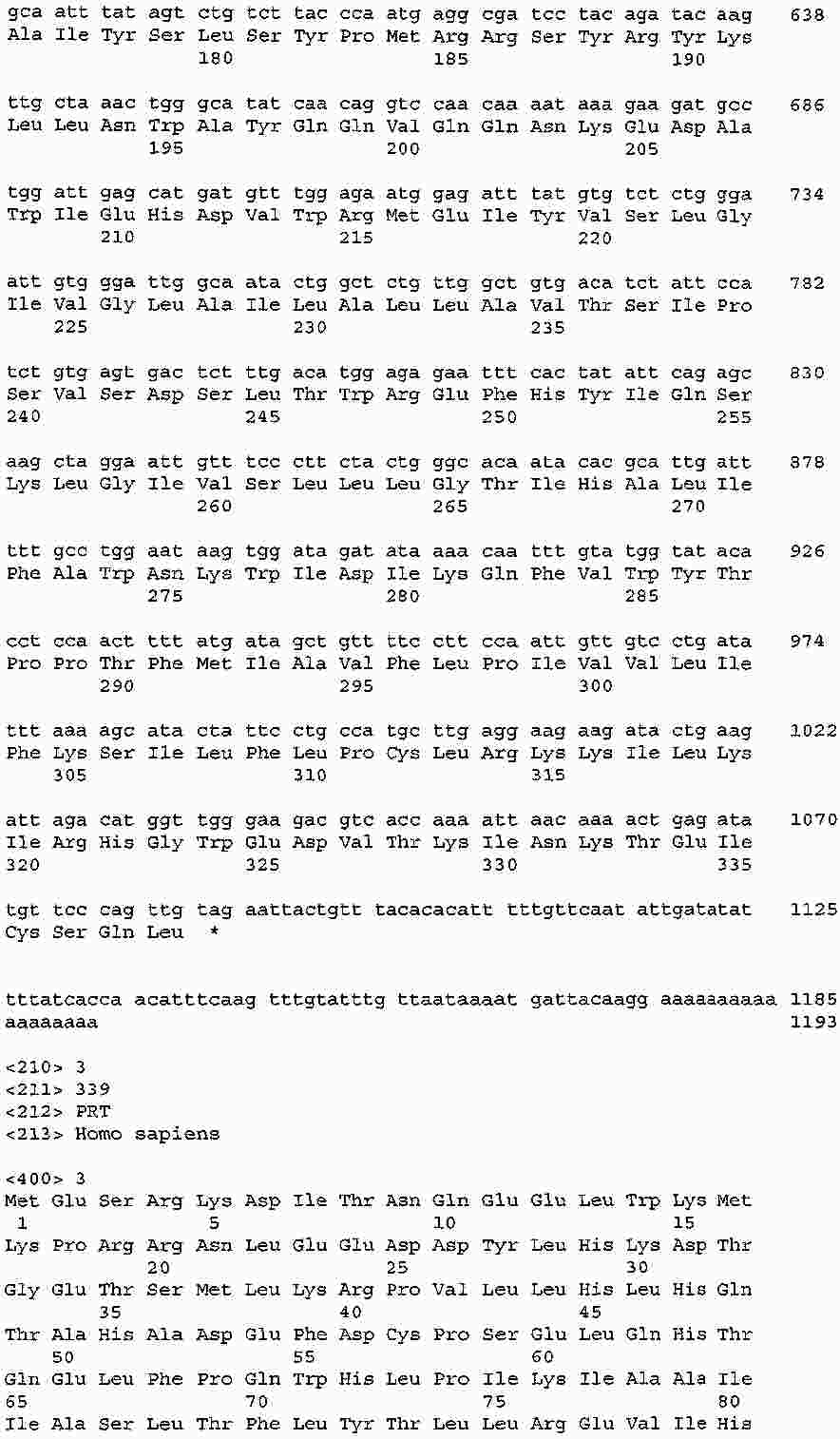
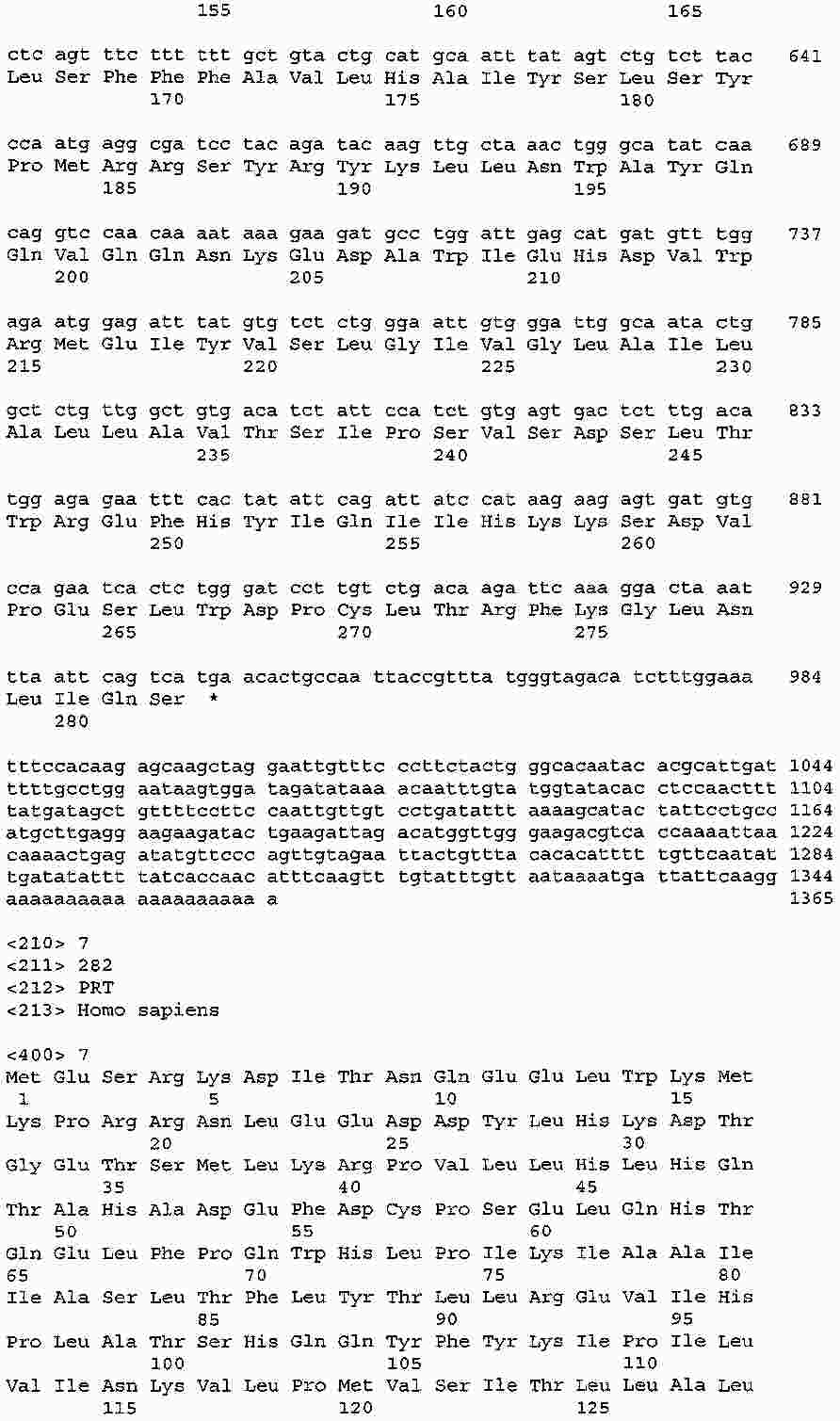
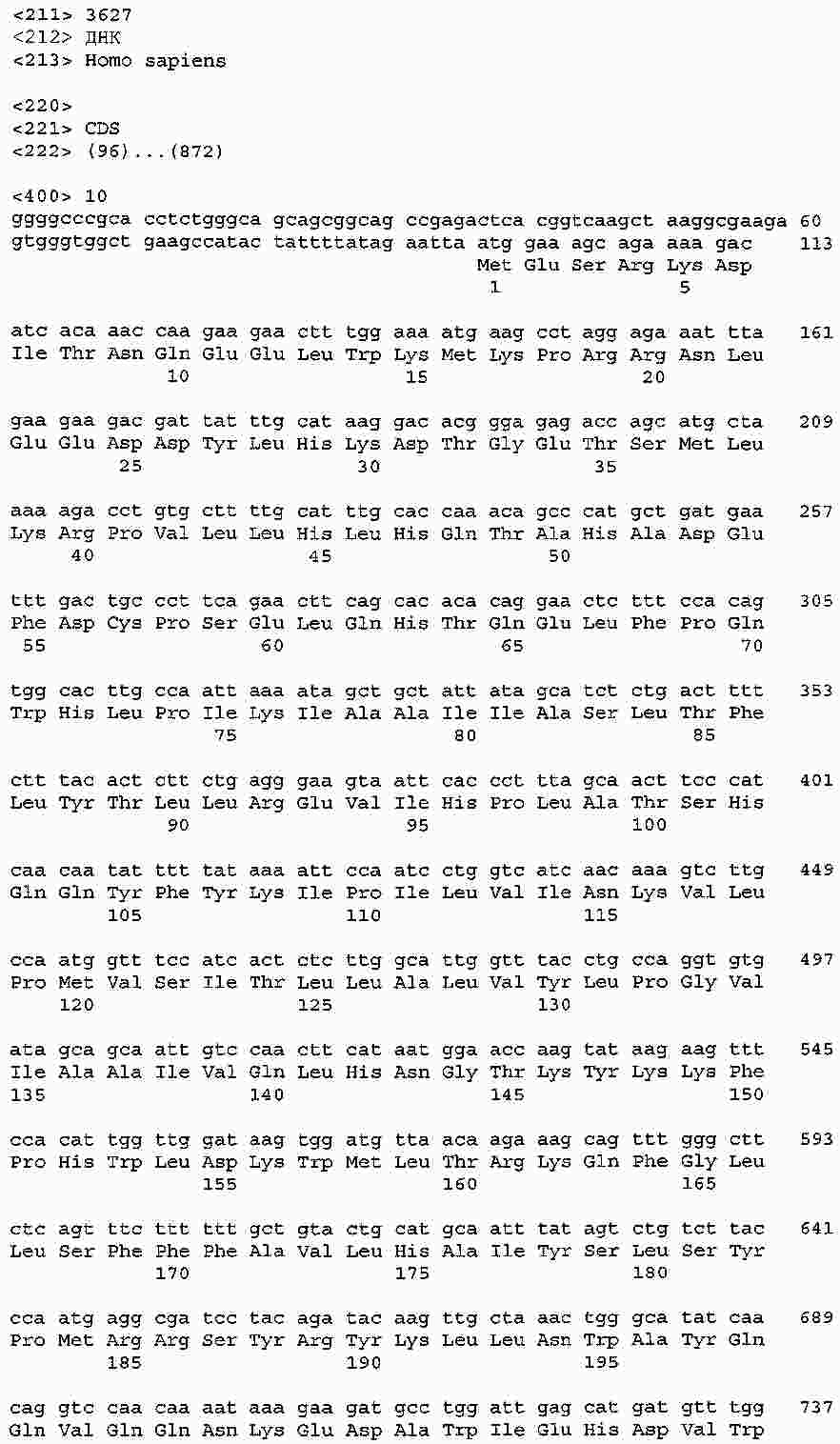
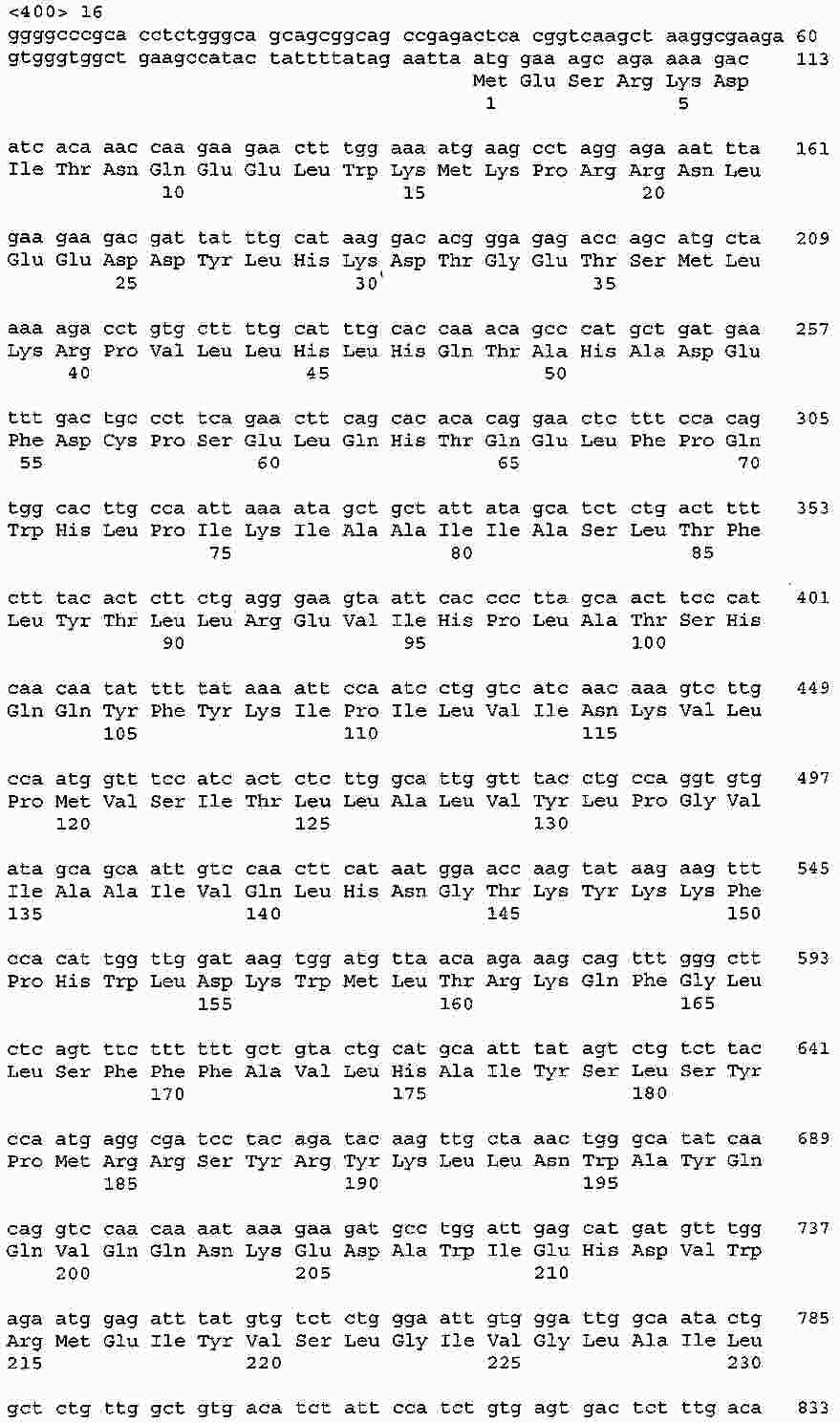
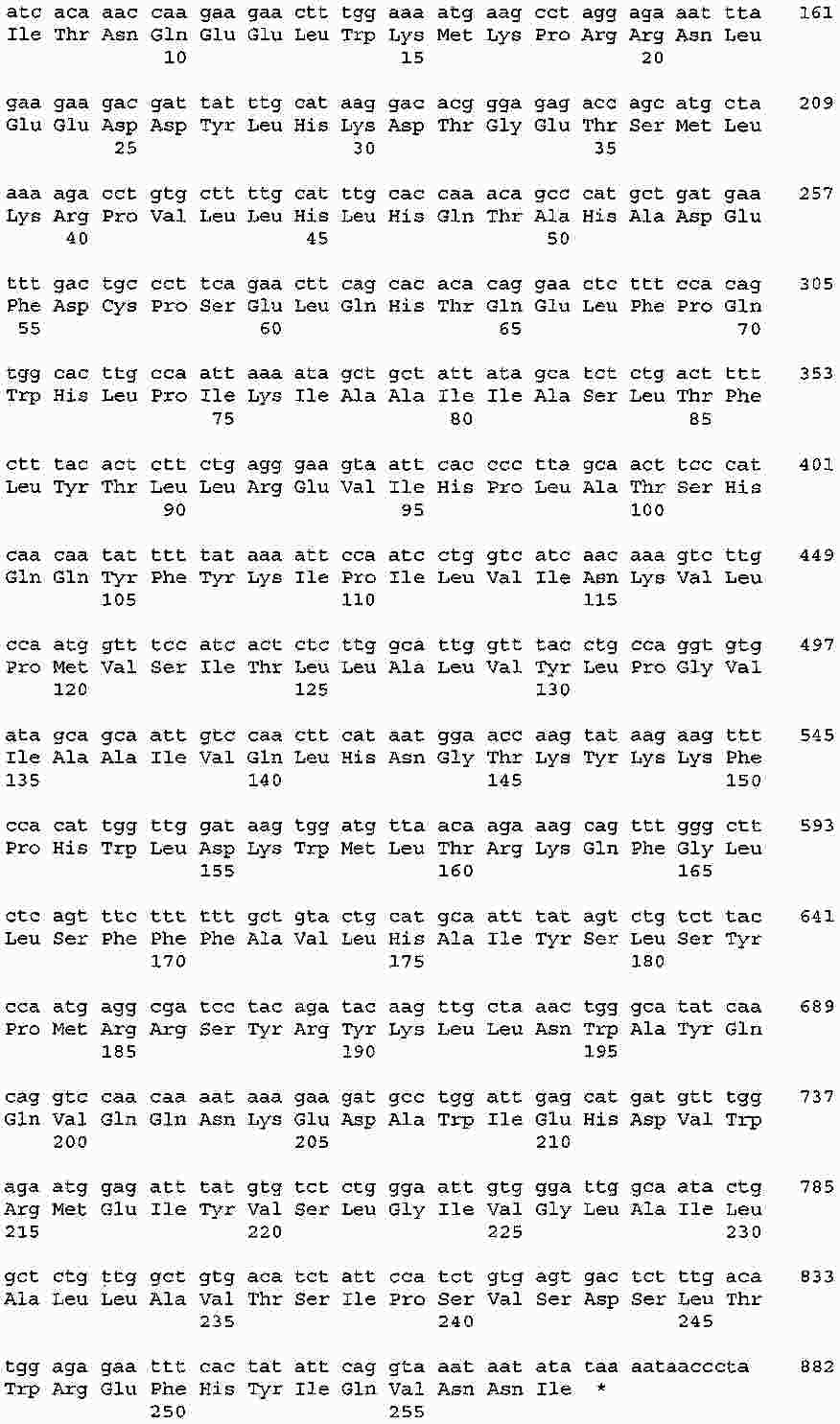
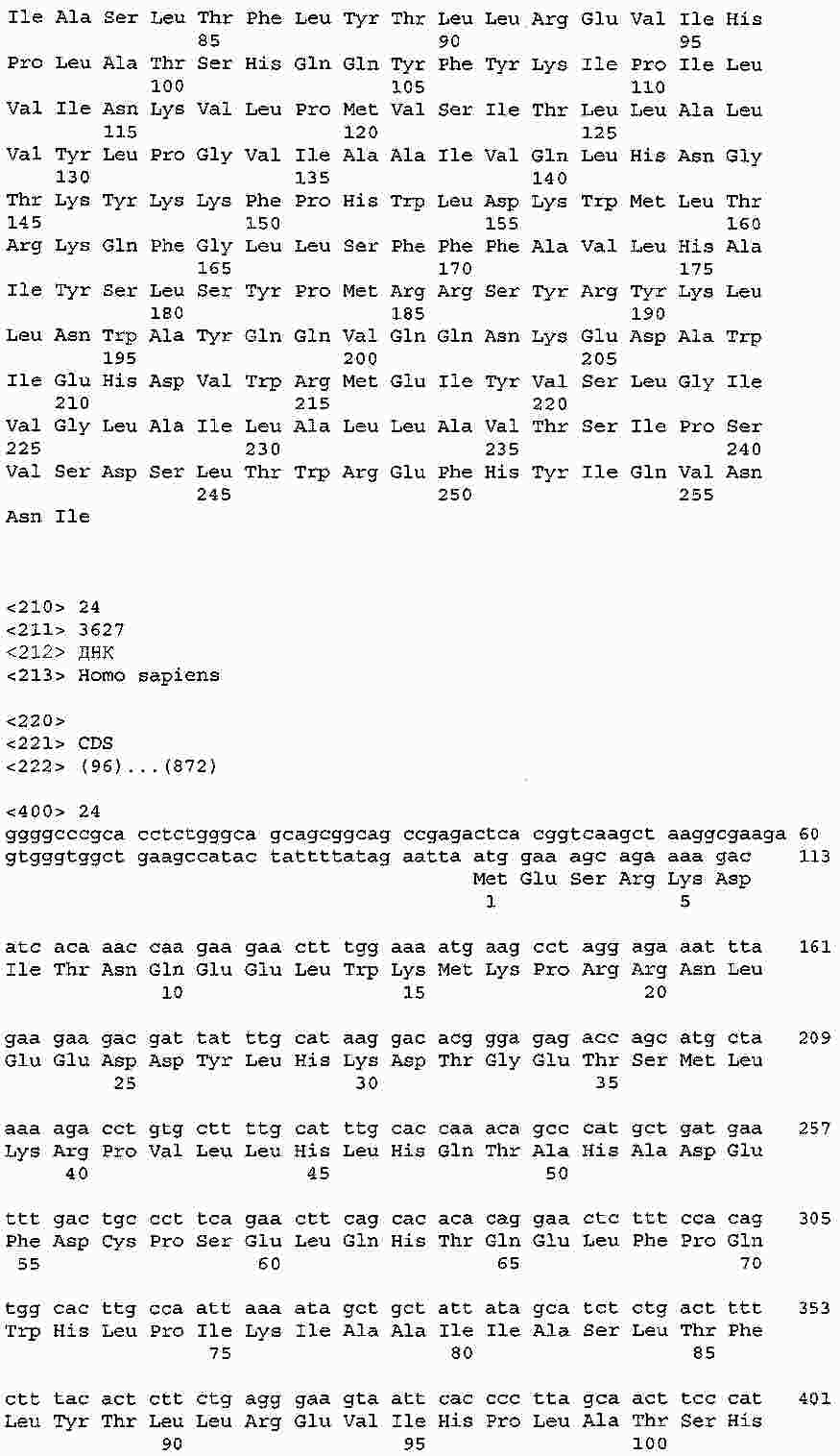
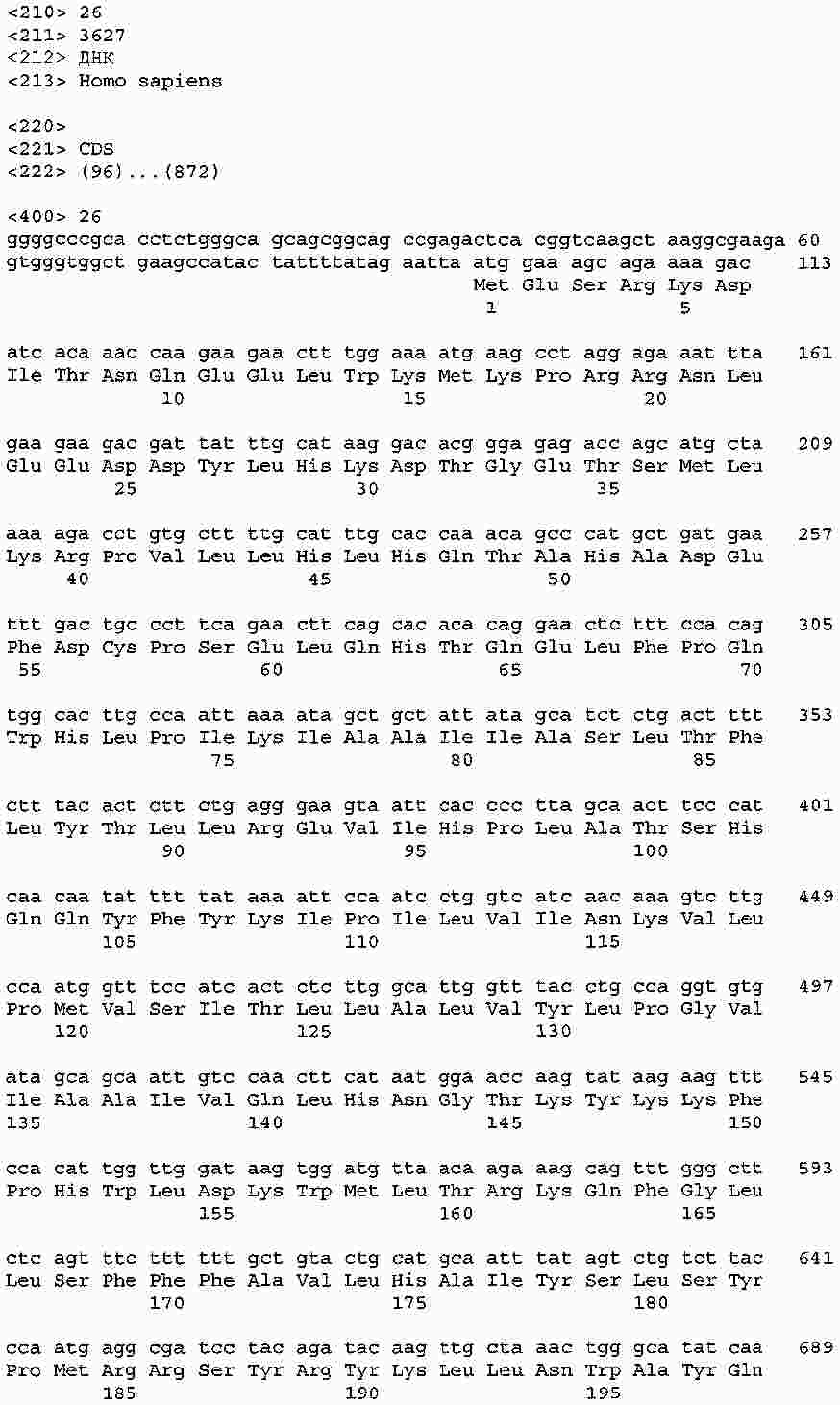
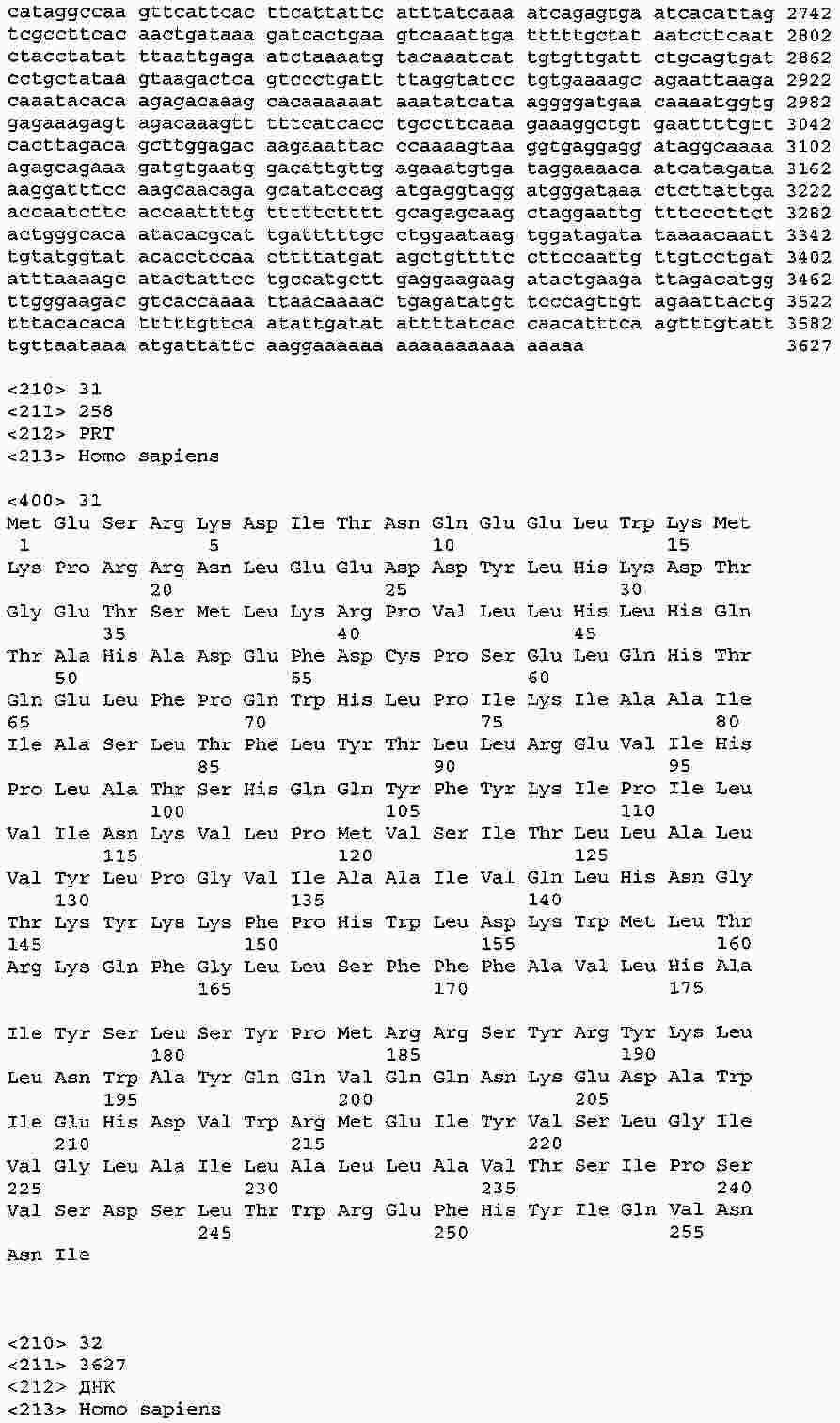
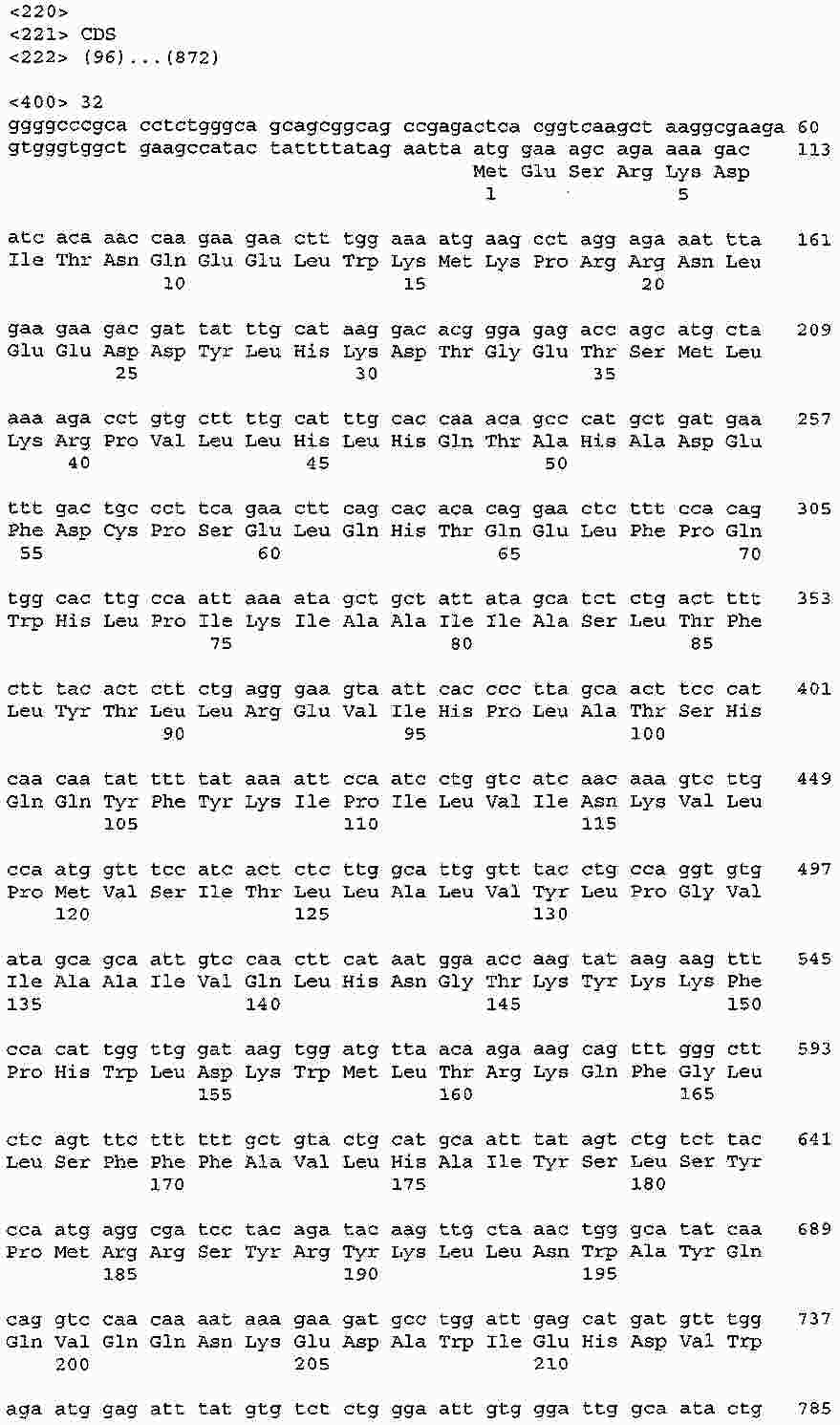
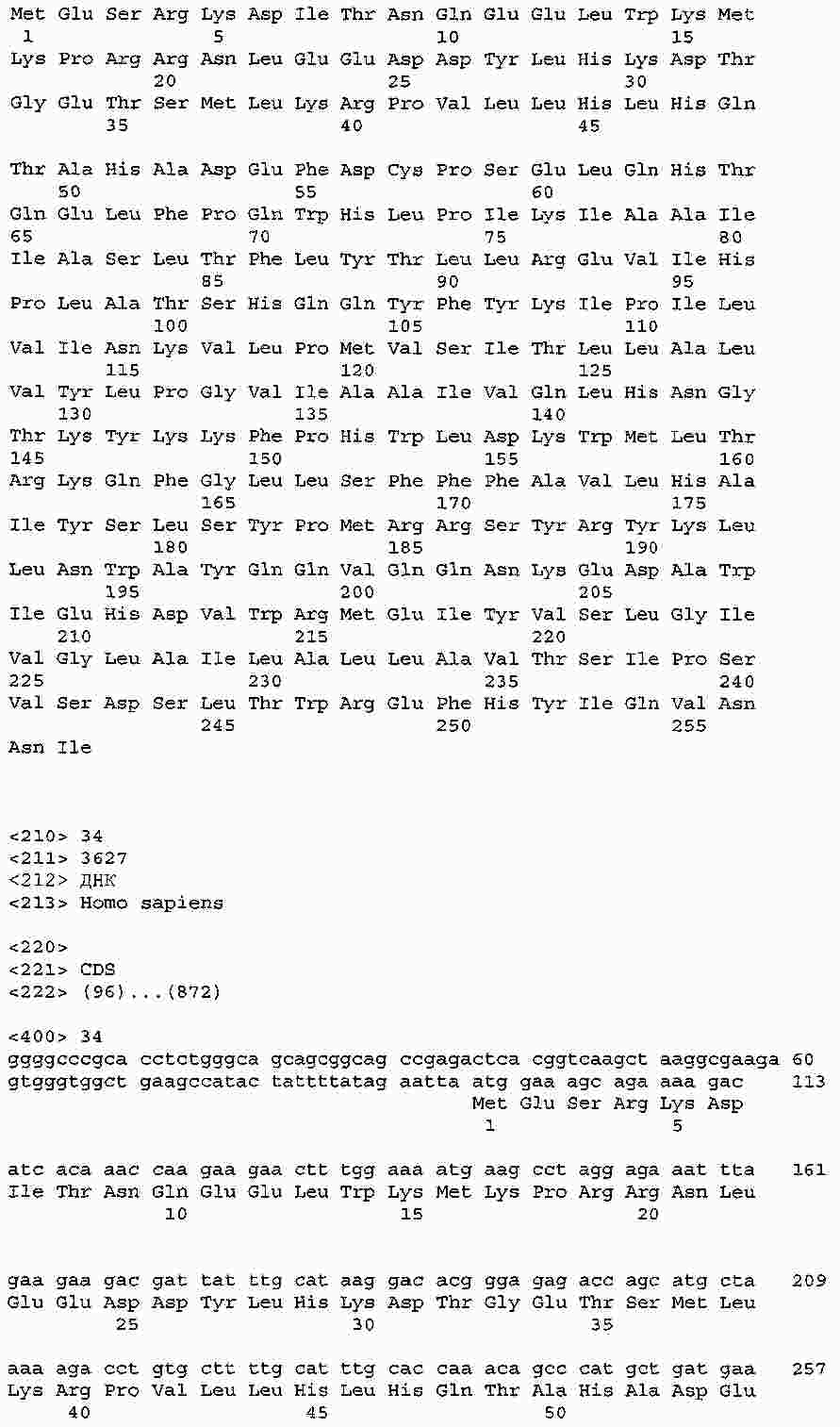
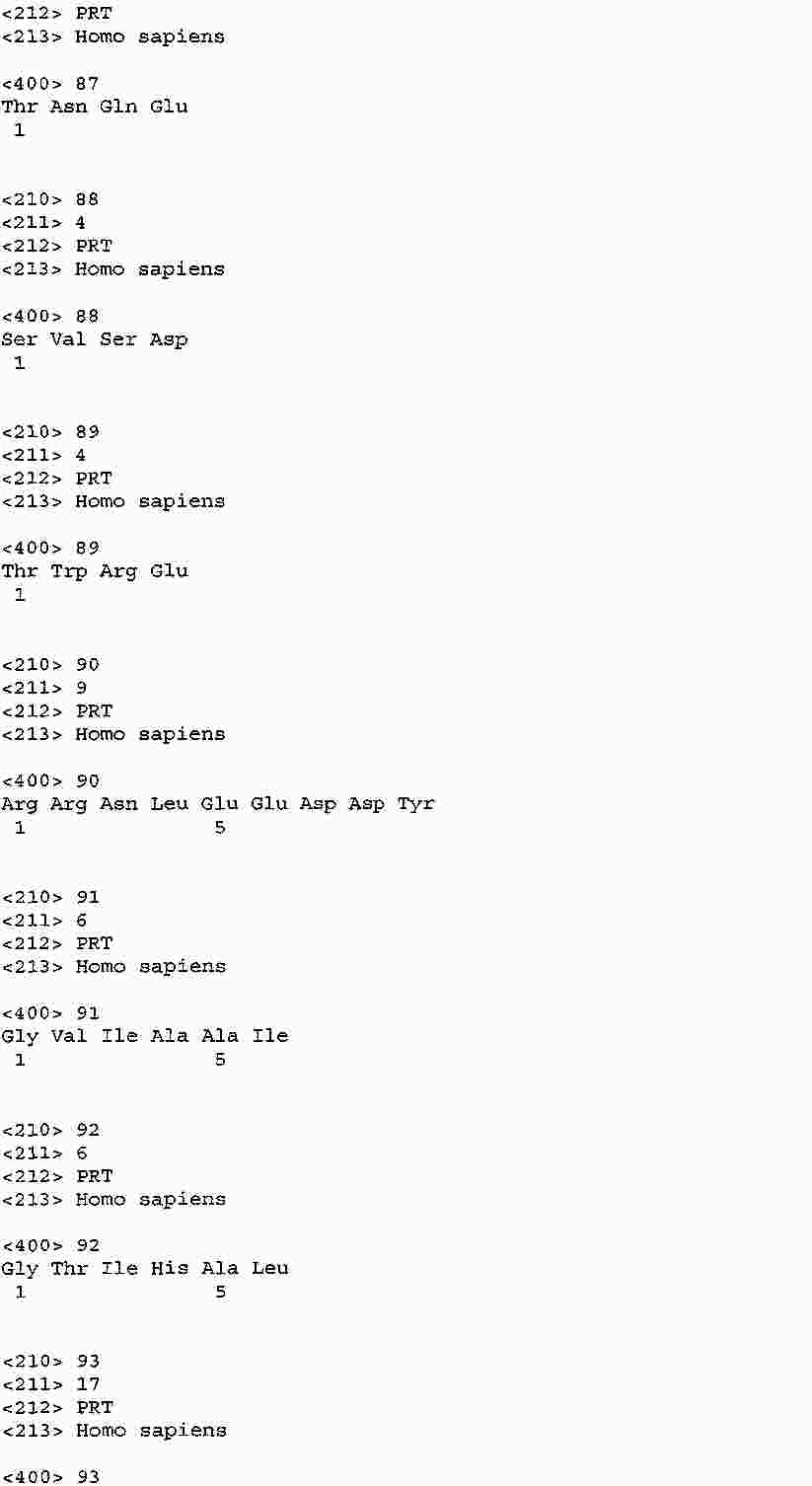
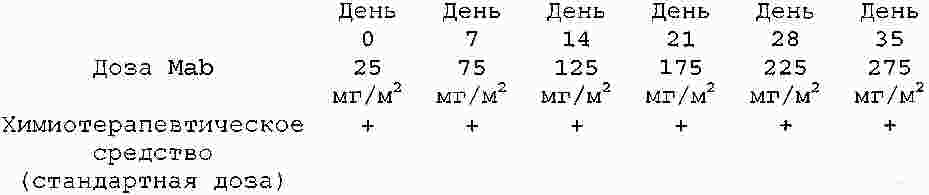|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000014870b*\id |
|
Термины запроса в документе Реферат В настоящем изобретении приведено описание антител и полученных из них молекул, которые связываются с новым белком STEAP-1 и его вариантами, где STEAP-1 обладает тканеспецифической экспрессией в нормальных взрослых тканях и аберрантно экспрессируется в раковых тканях, перечисленных в табл. I. Следовательно, STEAP-1 представляет собой диагностическую, прогностическую, профилактическую и/или терапевтическую мишень для противораковой терапии. Ген STEAP-1, или его фрагмент, или кодируемый им белок, или его варианты, или его фрагменты можно использовать для индуцирования гуморального или клеточного иммунного ответа; антитела или Т-клетки, способные взаимодействовать со STEAP-1, можно использовать для активной или пассивной иммунизации. Формула [0001] Антитело или его фрагмент, содержащие: [0002] Антитело или его фрагмент по п.1, где антитело является моноклональным антителом. [0003] Антитело или его фрагмент по п.2, где антитело представляет собой антитело Х92.1.30.1.1 (1) АТСС РТА-5802. [0004] Антитело или его фрагмент по п.2, где антитело представляет собой Х120.545.1.1 (АТСС РТА-5803). [0005] Антитело или его фрагмент по п.2, где моноклональное антитело является гуманизированным антителом. [0006] Антитело или его фрагмент по п.1, где фрагмент представляет собой фрагмент Fab, F(ab')2, Fv или Sfv. [0007] Рекомбинантный белок, содержащий антигенсвязывающий участок антитела по п.1. [0008] Антитело или его фрагмент по п.1, связанные с детектируемым маркером, токсином или терапевтическим средством. [0009] Антитело или его фрагмент по п.8, где детектируемый маркер выбран из радиоактивного изотопа; соединения, образующего хелатные комплексы с металлами; фермента; флуоресцентного соединения; биолюминесцентного соединения или хемилюминесцентного соединения. [0010] Антитело или его фрагмент по п.9, где радиоактивный изотоп выбран из 212Bi, 131I, 131In, 90Y, 186Re, 211At, 125I, 188Re, 153Sm, 213Bi, 32P и Lu. [0011] Антитело или его фрагмент по п.8, связанные с токсином. [0012] Антитело или его фрагмент по п.11, где токсин выбран из рицина, А-цепи рицина, доксорубицина, даунорубицина, маутанзиноида, таксола, бромида этидия, митомицина, этопозида, тенопозида, винкристина, винбластина, колхицина, дигидроксиантрациндиона, актиномицина, дифтерийного токсина, экзотоксина Pseudomonas (РЕ) А, РЕ40, абрина, А-цепи абрина, А-цепи модексина, альфа-сарцина, гелонина, митогеллина, ретстриктоцина, феномицина, эномицина, курицина, кротина, калихимицина, ингибитора Sapaonaria officinalis, глюкокортикоида, ауристатина, ауромицина, иттрия, висмута, комбрестатина, дуокармицинов, долостатина, сс1065 и цисплатина. [0013] Антитело или его фрагмент по п.12, связанные с ауристатином. [0014] Трансгенное животное, которое продуцирует моноклональное антитело по п.2. [0015] Гибридома, которая продуцирует моноклональное антитело по п.2. [0016] Вектор, содержащий полинуклеотид, кодирующий моноклональное антитело по п.2. [0017] Вектор по п.16, где антитело, кодируемое полинуклеотидом, представляет собой одноцепочечное моноклональное антитело, содержащее вариабельные области тяжелой и легкой цепей. [0018] Фармацевтическая композиция, которая содержит антитело или его фрагмент по п.1 в стандартной лекарственной форме для применения человеком. [0019] Способ детектирования наличия белка STEAP-1 в биологическом образце, включающий приведение в контакт указанного образца с антителом по п.1 и детектирование белка STEAP-1 в образце по его связыванию с антителом. [0020] Способ по п.19, где указанный биологический образец взят у пациента, который имеет или который предположительно имеет злокачественное заболевание ткани, приведенной в табл. I. [0021] Способ по п.20, где указанный биологический образец представляет собой сыворотку. [0022] Способ по п.20, в котором биологическим образцом является ткань предстательной железы. [0023] Способ по п.20, в котором биологическим образцом является периферическая кровь. [0024] Способ по п.23, в котором периферическая кровь содержит раковые клетки предстательной железы. [0025] Способ ингибирования роста клеток, экспрессирующих STEAP-1, у субъекта, включающий введение указанному субъекту вектора по п.17, где одноцепочечное моноклональное антитело экспрессируется в указанных клетках. [0026] Способ доставки цитотоксического или диагностического средства к клетке, которая экспрессирует белок STEAP-1, включающий получение цитотоксического или диагностического средства, конъюгированного с антителом или его фрагментом по п.1, с образованием конъюгата антитело-средство или фрагмент-средство и воздействие на клетку конъюгатом антитело-средство или фрагмент-средство. [0027] Способ по п.26, где цитотоксическое или диагностическое средство выбрано из группы, состоящей из детектируемого маркера, токсина и терапевтического средства. [0028] Способ по п.27, где диагностическое средство представляет собой детектируемый маркер, выбранный из радиоактивного изотопа; соединения, образующего хелатные комплексы с металлами; фермента; флуоресцентного соединения; биолюминесцентного соединения и хемилюминесцентного соединения. [0029] Способ по п.28, где радиоактивный изотоп выбран из 212Bi, 131I, 131In, 90Y, 186Re, 211At, 125I, 188Re, 153Sm, 213Bi, 32P и Lu. [0030] Способ по п.27, где доставляют цитотоксическое средство. [0031] Способ по п.30, где цитотоксическое средство представляет собой токсин, выбранный из рицина, А-цепи рицина, доксорубицина, даунорубицина, майтанзиноида, таксола, бромида этидия, митомицина, этопозида, тенопозида, винкристина, винбластина, колхицина, дигидроксиантрациндиона, актиномицина, дифтерийного токсина, экзотоксина Pseudomonas (РЕ) А, РЕ40, абрина, А-цепи абрина, А-цепи модексина, альфа-сарцина, гелонина, митогеллина, ретстриктоцина, феномицина, эномицина, курицина, кротина, калихимицина, ингибитора Sapaonaria officinalis, глюкокортикоида, ауристатина, ауромицина, иттрия, висмута, комбрестатина, дуокармицинов, долостатина, сс1065 и цисплатина. [0032] Способ по п.31, в котором токсином является ауристатин. [0033] Способ детектирования белка STEAP-1 в тестируемом образце, включающий приведение в контакт тестируемого образца, полученного у пациента, который имеет или который предположительно имеет злокачественную клетку, экспрессирующую STEAP-1, с антителом или фрагментом антитела по п.1 и определение количества STEAP-1 в тестируемом образце по связыванию с антителом или фрагментом антитела. [0034] Способ по п.33, дополнительно предусматривающий сравнение количества STEAP-1, связанного с антителом или фрагментом антитела, из тестируемого образца с количеством STEAP-1 из контрольного образца. [0035] Способ по п.33, в котором тестируемым образцом является образец крови, мочи, семенной жидкости, предстательной железы, толстой кишки, мочевого пузыря, поджелудочной железы, яичника, шейки матки, семенника, молочной железы, кости, лимфатического узла, легкого, печени, мозга, сыворотки или клеточного препарата. [0036] Способ по п.33, в котором тестируемым образцом является образец периферической крови. [0037] Способ по п.36, в котором периферическая кровь содержит раковые клетки предстательной железы. [0038] Способ по п.34, дополнительно включающий взятие тестируемого образца и контрольного образца у пациента, который имеет или предположительно имеет злокачественное заболевание ткани, приведенной в табл. I. [0039] Способ ингибирования роста клеток, которые экспрессируют STEAP-1, предусматривающий воздействие на клетки антителом или фрагментом антитела по п.1. [0040] Способ по п.39, в котором клетками являются раковые клетки. [0041] Способ по п.40, в котором раковые клетки представляют собой раковые клетки предстательной железы. [0042] Способ лечения рака у субъекта, предусматривающий введение субъекту фармацевтической композиции, содержащей антитело или его фрагмент по п.1. [0043] Способ по п.42, где раком является рак предстательной железы. [0044] Способ ингибирования роста раковых клеток, которые экспрессируют STEAP-1, предусматривающий воздействие на клетки антителом или фрагментом антитела по п.11. [0045] Способ по п.44, где антитело или его фрагмент связаны с токсином, выбранным из рицина, А-цепи рицина, доксорубицина, даунорубицина, майтанзиноида, таксола, бромида этидия, митомицина, этопозида, тенопозида, винкристина, винбластина, колхицина, дигидроксиантрациндиона, актиномицина, дифтерийного токсина, экзотоксина Pseudomonas (РЕ) А, РЕ40, абрина, А-цепи абрина, А-цепи модексина, альфа-сарцина, гелонина, митогеллина, ретстриктоцина, феномицина, эномицина, курицина, кротина, калихимицина, ингибитора Sapaonaria officinalis, глюкокортикоида, ауристатина, ауромицина, иттрия, висмута, комбрестатина, дуокармицинов, долостатина, сс1065 и цисплатина. [0046] Способ по п.45, в котором токсином является ауристатин. [0047] Способ по п.44, где раковые клетки представляют собой раковые клетки злокачественного заболевания ткани, приведенной в табл. I. [0048] Способ по п.47, где раковые клетки представляют собой раковые клетки предстательной железы. [0049] Способ лечения рака у субъекта, предусматривающий введение субъекту фармацевтической композиции, содержащей антитело или его фрагмент по п.11. [0050] Способ по п.49, где антитело или его фрагмент связаны с токсином, выбранным из рицина, А-цепи рицина, доксорубицина, даунорубицина, майтанзиноида, таксола, бромида этидия, митомицина, этопозида, тенопозида, винкристина, винбластина, колхицина, дигидроксиантрациндиона, актиномицина, дифтерийного токсина, экзотоксина Pseudomonas (РЕ) А, РЕ40, абрина, А-цепи абрина, А-цепи модексина, альфа-сарцина, гелонина, митогеллина, ретстриктоцина, феномицина, эномицина, курицина, кротина, калихимицина, ингибитора Sapaonaria officinalis, глюкокортикоида, ауристатина, ауромицина, иттрия, висмута, комбрестатина, дуокармицинов, долостатина, сс1065 и цисплатина. [0051] Способ по п.50, в котором токсином является ауристатин. [0052] Способ по п.49, где раком является рак предстательной железы. [0053] Антитело или его фрагмент, содержащие все определяющие комплементарность области (CDR) тяжелой цепи и легкой цепи из антитела, обозначенного Х92.1.30.1.1 (1) (АТСС РТА-5802), или антитела, обозначенного Х120.545.1.1 (АТСС РТА-5803), причем антитело или фрагмент антитела связывается с белком STEAP-1, имеющим последовательность SEQ ID NO:3. [0054] Антитело или его фрагмент, содержащие последовательность SEQ ID NO:50 вариабельной области тяжелой цепи и последовательность вариабельной области легкой цепи, выбранную из группы, состоящей из последовательности SEQ ID NO: 52, SEQ ID NO: 54 и SEQ ID NO: 56, причем антитело или фрагмент антитела связывается с белком STEAP-1, имеющим последовательность SEQ ID NO: 3. [0055] Антитело или его фрагмент, которое представляет собой гуманизированную или модифицированную форму моноклонального антитела Х92.1.30.1.1 (1) (АТСС РТА-5802), где антитело или его фрагмент специфически связывается с белком STEAP-1, имеющим последовательность SEQ ID NO:3. [0056] Антитело или его фрагмент, которое представляет собой гуманизированную или модифицированную форму моноклонального антитела Х120.545.1.1 (АТСС РТА-5803), где антитело или его фрагмент специфически связывается с белком STEAP-1, имеющим последовательность SEQ ID NO:3. [0057] Антитело или его фрагмент по пп.53-56, связанные с детектируемым маркером, токсином или терапевтическим средством. [0058] Антитело или его фрагмент по п.57, где детектируемый маркер выбран из радиоактивного изотопа; соединения, образующего хелатные комплексы с металлами; фермента; флуоресцентного соединения; биолюминесцентного соединения или хемилюминесцентного соединения. [0059] Антитело или его фрагмент по п.57, связанные с токсином. [0060] Антитело или его фрагмент по п.59, где токсин выбран из рицина, А-цепи рицина, доксорубицина, даунорубицина, маутанзиноида, таксола, бромида этидия, митомицина, этопозида, тенопозида, винкристина, винбластина, колхицина, дигидроксиантрациндиона, актиномицина, дифтерийного токсина, экзотоксина Pseudomonas (РЕ) А, РЕ40, абрина, А-цепи абрина, А-цепи модексина, альфа-сарцина, гелонина, митогеллина, ретстриктоцина, феномицина, эномицина, курицина, кротина, калихимицина, ингибитора Sapaonaria officinalis, глюкокортикоида, ауристатина, ауромицина, иттрия, висмута, комбрестатина, дуокармицинов, долостатина, сс1065 и цисплатина. [0061] Антитело или его фрагмент по п.60, связанные с ауристатином. [0062] Способ ингибирования роста раковых клеток, которые экспрессируют STEAP-1, предусматривающий воздействие на клетки антителом или фрагментом антитела по п.59. [0063] Способ по п.62, где антитело или его фрагмент связаны с токсином, выбранным из рицина, А-цепи рицина, доксорубицина, даунорубицина, майтанзиноида, таксола, бромида этидия, митомицина, этопозида, тенопозида, винкристина, винбластина, колхицина, дигидроксиантрациндиона, актиномицина, дифтерийного токсина, экзотоксина Pseudomonas (РЕ) А, РЕ40, абрина, А-цепи абрина, А-цепи модексина, альфа-сарцина, гелонина, митогеллина, ретстриктоцина, феномицина, эномицина, курицина, кротина, калихимицина, ингибитора Sapaonaria officinalis, глюкокортикоида, ауристатина, ауромицина, иттрия, висмута, комбрестатина, дуокармицинов, долостатина, сс1065 и цисплатина. [0064] Способ по п.62, в котором токсином является ауристатин. [0065] Способ по п.44, где раковые клетки представляют собой раковые клетки злокачественного заболевания ткани, приведенной в табл. I. [0066] Способ по п.65, где раковые клетки представляют собой раковые клетки предстательной железы. [0067] Способ лечения рака у субъекта, предусматривающий введение субъекту фармацевтической композиции, содержащей антитело или его фрагмент по п.59. [0068] Способ по п.67, где антитело или его фрагмент связаны с токсином, выбранным из рицина, А-цепи рицина, доксорубицина, даунорубицина, майтанзиноида, таксола, бромида этидия, митомицина, этопозида, тенопозида, винкристина, винбластина, колхицина, дигидроксиантрациндиона, актиномицина, дифтерийного токсина, экзотоксина Pseudomonas (РЕ) А, РЕ40, абрина, А-цепи абрина, А-цепи модексина, альфа-сарцина, гелонина, митогеллина, ретстриктоцина, феномицина, эномицина, курицина, кротина, калихимицина, ингибитора Sapaonaria officinalis, глюкокортикоида, ауристатина, ауромицина, иттрия, висмута, комбрестатина, дуокармицинов, долостатина, сс1065 и цисплатина. [0069] Способ по п.68, в котором токсином является ауристатин. [0070] Способ по п.67, где раком является рак предстательной железы. Полный текст патента Права на изобретения, сделанные в результате исследований, финансируемых федеральным правительством В данном случае неприменимы. Описанное в данном документе изобретение относится к антителам, а также к их связывающим фрагментам и молекулам, полученным из них методом генной инженерии, которые связываются с белками, называемыми STEAP-1. Данное изобретение также относится к диагностическим, прогностическим, профилактическим и терапевтическим способам, а также к композициям, которые можно использовать для лечения злокачественных заболеваний, сопровождающихся экспрессией STEAP-1. Рак является второй из основных причин смертности среди людей после ишемической болезни сердца. Во всем мире каждый год от рака умирают миллионы людей. Только в Соединенных Штатах, по данным Американского общества по борьбе с раком, рак является причиной смерти более полумиллиона людей ежегодно, причем каждый год диагностируется более 1,2 миллионов новых случаев. В то время как смертность от сердечных заболеваний значительно снизилась, смертность от рака в целом возрастает. Ожидается, что в начале следующего столетия рак станет главной причиной смертности. Во всем мире выделяют несколько типов рака, как наиболее опасных для жизни. В частности, карциномы легких, простаты, молочной железы, толстой кишки, поджелудочной железы, яичников и мочевого пузыря являются основными причинами смерти от злокачественных заболеваний. Указанные и практически все другие карциномы имеют общий летальный характер. За немногими исключениями, метастатическая карцинома приводит к смертельному исходу. Кроме того, опыт показывает, что, даже если пациенты выживают после первичного злокачественного заболевания, их жизнь сильно изменяется. Многие раковые пациенты испытывают сильное беспокойство, сознавая возможность рецидива или того, что лечение может быть неудачным. После лечения многие раковые пациенты испытывают сильную слабость. Кроме того, у многих раковых пациентов наблюдаются рецидивы. Во всем мире рак простаты является четвертым по распространенности среди злокачественных заболеваний у мужчин. В Северной Америке и в Северной Европе он, несомненно, является наиболее распространенным злокачественным заболеванием у мужчин и второй из основных причин смертности среди мужчин. Только в Соединенных Штатах от данного заболевания ежегодно умирает значительно более 30000 мужчин - второе место по смертности после рака легких. Несмотря на величину данных цифр, пока еще не существует эффективного лечения метастазирующего рака простаты. Основными способами лечения продолжают оставаться хирургическая простатэктомия, лучевая терапия, антигормональная терапия, хирургическая кастрация и химиотерапия. К сожалению, данные способы лечения во многих случаях являются неэффективными и зачастую связаны с нежелательными последствиями. С точки зрения диагностики отсутствие маркера опухоли простаты, который позволяет осуществлять точную диагностику локализованных опухолей на ранней стадии, остается существенным препятствием для диагностики и лечения данного заболевания. Хотя анализ сывороточного специфического антигена простаты (PSA) является очень полезным инструментом, общепризнано, что в некоторых важных аспектах он обладает недостаточной специфичностью и недостаточной общей функциональностью. Прогресс в идентификации других маркеров, специфичных для рака простаты, был достигнут в результате применения ксенотрансплантатов рака простаты, которые позволяют воспроизводить разные стадии данного заболевания у мышей. Ксенотрансплантаты LAPC (Los Angeles Prostate Cancer) представляют собой ксенотрансплантаты рака простаты, которые перенесли пересадку мышам с тяжелым комбинированным иммунодефицитом (SCID) и которые способны имитировать переход от зависимости от андрогенов к независимости от андрогенов (Klein et al., 1997, Nat. Med. 3: 402). Идентифицированные позже маркеры рака простаты включают в себя РСТА-1 (Su et al., 1996, Proc. Natl. Acad. Sci. USA, 93: 7252), простата-специфичный мембранный (PSM) антиген (Pinto et al., Clin. Cancer Res. 2 Sep., 1996 (9): 1445-51), STEAP (Hubert, et al., Proc. Natl. Acad. Sci. USA. 7 Dec., 1999; 96(25): 14523-8) и антиген стволовых клеток, дифференцирующихся в клетки простаты (PSCA) (Reiter et al., 1998, Proc. Natl. Acad. Sci. USA, 95: 1735). Хотя ранее идентифицированные маркеры, такие как PSA, PSM, РСТА и PSCA, помогают диагностировать и лечить рак простаты, еще существует потребность в идентификации других маркеров и терапевтических мишеней, специфичных для рака простаты и родственных заболеваний, которые позволили бы усовершенствовать способы диагностики и лечения. Почечно-клеточная карцинома (RCC) составляет приблизительно 3% от злокачественных заболеваний взрослых. Если диаметр аденомы достигает 2-3 см, существует возможность злокачественного перерождения. У взрослых двумя основными злокачественными опухолями почек являются почечно-клеточная аденокарцинома и переходно-клеточная карцинома почечной лоханки или мочеточника. В США зарегистрировано более 29000 случаев почечно-клеточной аденокарциномы, причем в 1998 г. от данного заболевания умерло более 11600 пациентов. Переходно-клеточная карцинома встречается реже, в Соединенных Штатах диагностируется приблизительно 500 случаев в год. В течение многих десятилетий хирургия была основным способом лечения почечно-клеточной аденокарциномы. До последнего времени метастазирующие заболевания не поддавались никакому системному лечению. Последние разработки в области системной терапии, в особенности иммунотерапии, позволили найти кардинальный подход к лечению метастазирующей почечно-клеточной карциномы у соответствующих пациентов, который обеспечивает вероятность устойчивых ответов. Тем не менее, остается потребность в эффективных способах лечения данных пациентов. Среди новых случаев рака, зарегистрированных в Соединенных Штатах, рак мочевого пузыря составляет приблизительно 5% у мужчин (пятое место среди наиболее распространенных новообразований) и 3% у женщин (восьмое место среди наиболее распространенных новообразований). Заболеваемость медленно возрастает с увеличением возраста популяции. В 1998 г. было зарегистрировано 54500 случаев, в том числе 39500 у мужчин и 15000 у женщин. Связанная с возрастом заболеваемость в Соединенных Штатах составляет 32 на 100000 у мужчин и 8 на 100000 у женщин. Исторически сложившееся соотношение заболеваемости у мужчин/женщин 3:1 может уменьшаться вследствие увеличения курящих женщин. В 1998 г. зарегистрировано 11000 смертей от рака мочевого пузыря (7800 у мужчин и 3900 у женщин). Заболеваемость раком мочевого пузыря и смертность от данного заболевания сильно увеличиваются с возрастом и представляют собой проблему, возрастающую по мере увеличения возраста популяции. Большинство злокачественных заболеваний мочевого пузыря рецидивируют в мочевом пузыре. Рак мочевого пузыря лечат сочетанием трансуретральной резекции мочевого пузыря (TUR) и внутрипузырной химиотерапии или иммунотерапии. Многоочаговый и рецидивирующий характер рака мочевого пузыря ограничивает применение TUR. Большинство проникающих в мышцы раковых опухолей нельзя вылечить с помощью одной TUR. Радикальная цистэктомия с отводом мочи является наиболее эффективным средством устранения раковой опухоли, но сопровождается нежелательным воздействием на мочевыводящие и половые функции. Соответственно, остается настоятельная потребность в способах лечения, благоприятно воздействующих на пациентов с раком мочевого пузыря. В 2000 г. в Соединенных Штатах зарегистрировано 130200 случаев колоректального рака, в том числе 93800 случаев рака толстой кишки и 36400 случаев рака прямой кишки. Колоректальный рак занимает третье место среди наиболее распространенных злокачественных заболеваний у мужчин и женщин. В период 1992-1996 гг. коэффициент заболеваемости значительно снизился (-2,1% в год). Исследования показывают, что данное снижение обусловлено увеличением скрининга населения и удалением полипов, позволяющим предотвратить их развитие до инвазивных злокачественных заболеваний. В 2000 г. зарегистрировано 56300 смертей (47700 от рака толстой кишки и 8600 от рака прямой кишки), что составляет приблизительно 11% от всех смертельных исходов злокачественных заболеваний в США. В настоящее время хирургия является наиболее распространенным способом лечения колоректального рака и при отсутствии метастазирования часто дает положительный результат. Химиотерапию, или химиотерапию в сочетании с лучевой терапией, назначают до или после хирургической операции большинству пациентов, у которых раковая опухоль глубоко проникает в стенки кишечника или метастазирует в лимфатические узлы. При раке толстой кишки и при раке прямой кишки иногда требуется постоянная колостомия (открытие брюшной полости для удаления экскрементов). Следовательно, существует потребность в эффективных способах диагностики и лечения колоректального рака. В 2000 г. зарегистрировано 164100 новых случаев рака легких и бронхов, что составляет 14% от всех раковых диагнозов в США. Коэффициент заболеваемости раком легких и бронхов значительно снизился: от 86,5 на 100000 человек в 1984 г. до 70,0 в 1996 г. В 1990-х гг. скорость увеличения заболеваемости среди женщин начала снижаться. В 1996 г. коэффициент заболеваемости у женщин составлял 42,3 на 100000 человек. В 2000 г. было зарегистрировано 156900 смертей от рака легких и бронхов, что составляет 28% от общей смертности от злокачественных заболеваний. В период 1992-1996 гг. смертность от рака легких среди мужчин значительно снизилась (-1,7% в год), тогда как смертность среди женщин еще заметно росла (0,9% в год). С 1987 г. от рака легких каждый год умирало больше женщин, чем от рака молочной железы, который в течение 40 лет был основной причиной смертей от злокачественных заболеваний среди женщин. Снижение заболеваемости раком легких и смертности от данного заболевания, скорее всего, является следствием уменьшения курения в течение предыдущих 30 лет; однако по уменьшению курения женщины отстают от мужчин. Интересно, что в то время как снижение употребления табака у взрослых замедляется, у подростков употребление табака снова увеличивается. Способы лечения рака легких и бронхов зависят от типа и стадии рака и включают в себя хирургическое лечение, лучевую терапию и химиотерапию. Для лечения многих локализованных раковых опухолей обычно выбирают хирургию. Поскольку данное заболевание обычно распространяется со времени его обнаружения, наряду с хирургическим лечением обычно требуется лучевая терапия и химиотерапия. При мелкоклеточном раке легких обычно выбирают химиотерапию, отдельно или в сочетании с лучевой терапией; при таком режиме лечения у пациентов часто наступает ремиссия, которая в некоторых случаях может быть длительной. Однако продолжает существовать потребность в эффективных способах лечения и диагностики рака легких и бронхов. В течение 2000 г. в США было зарегистрировано 182800 новых случаев инвазивного рака молочной железы. Кроме того, в 2000 г. было диагностировано приблизительно 1400 новых случаев рака молочной железы у мужчин. После увеличения приблизительно до 4% в год в 1980 г. коэффициент заболеваемости раком молочной железы у женщин снизился в 1990 г. приблизительно до 110,6 случаев на 100000 человек. В 2000 г. только в США было зарегистрировано 41200 смертей (40800 женщин, 400 мужчин) от рака молочной железы. Смертность от рака молочной железы занимает второе место среди смертности от злокачественных заболеваний у женщин. В соответствии с последними данными коэффициент смертности значительно снизился в период 1992-1996 гг., причем сильнее всего он снизился у молодых женщин как с белой кожей, так и с черной. Возможно, данное снижение является результатом более ранней диагностики и усовершенствования лечения. С учетом медицинских обстоятельств и предпочтений пациента лечение рака молочной железы может включать в себя лампэктомию (локальное удаление опухоли) и удаление лимфатических узлов под мышкой; мастэктомию (хирургическое удаление молочной железы) и удаление лимфатических узлов под мышкой; лучевую терапию; химиотерапию; или гормональную терапию. Часто используют сочетание двух или более способов. Многочисленные исследования показали, что в случае ранней стадии заболевания продолжительность жизни после лампэктомии в сочетании с лучевой терапией подобна продолжительность жизни после модифицированной радикальной мастэктомии. Значительные достижения в восстановительной технологии предоставляют некоторые возможности для воссоздания молочной железы после мастэктомии. Недавно такое воссоздание было осуществлено одновременно с мастэктомией. Локальное удаление протоковой карциномы in situ (DCIS) вместе с соответствующими количествами окружающей нормальной ткани молочной железы может предотвратить местный рецидив DCIS. Облучение молочной железы и/или обработка тамоксифеном могут уменьшить вероятность возникновения DCIS в оставшейся ткани молочной железы. Это важно, поскольку DCIS, при отсутствии лечения, может развиться в инвазивный рак молочной железы. Тем не менее, указанные способы лечения имеют серьезные побочные эффекты или последствия. Следовательно, существует потребность в эффективных способах лечения рака молочной железы. В 2000 г. в Соединенных Штатах зарегистрировано 23100 новых случаев рака яичников. Данное заболевание составляет 4% от всех злокачественных заболеваний у женщин и занимает второе место среди гинекологических злокачественных заболеваний. В период 1992-1996 гг. коэффициент заболеваемости раком яичников значительно снизился. В 2000 г. зарегистрировано 14000 смертей от рака яичников. Рак яичников вызывает больше смертей, чем любое другое злокачественное заболевание женской репродуктивной системы. Для лечения рака яичников используют хирургию, лучевую терапию и химиотерапию. Хирургия обычно включает в себя удаление одного или обоих яичников, фаллопиевых труб (удаление придатков матки) и матки (гистерэктомия). В случае некоторых опухолей на ранней стадии удаляют только пораженный яичник, особенно если пациентом является молодая женщина, желающая иметь детей. В случае запущенного заболевания, чтобы увеличить эффект химиотерапии, делают попытку удалить все внутрибрюшинные пораженные органы. Таким образом, остается потребность в эффективных способах лечения рака яичников. В 2000 г. в Соединенных Штатах зарегистрировано 28300 новых случаев рака поджелудочной железы. В течение последних 20 лет заболеваемость раком поджелудочной железы у мужчин уменьшилась. Заболеваемость среди женщин остается приблизительно постоянной, но есть тенденция к уменьшению. В 2000 г. в Соединенных Штатах зарегистрировано 28200 смертей от рака поджелудочной железы. Смертность среди мужчин за последние 20 лет немного, но значимо уменьшилась (приблизительно -0,9% в год), в то время как смертность среди женщин несколько увеличилась. Для лечения рака поджелудочной железы используют хирургию, лучевую терапию и химиотерапию. Указанные способы лечения могут увеличивать выживаемость и/или облегчать симптомы у многих пациентов, но в большинстве случаев они не приводят к излечению. Существует настоятельная потребность в дополнительных способах лечения и диагностики злокачественных заболеваний. Указанные способы лечения включают в себя применение антител, вакцин и маленьких молекул. Кроме того, существует потребность в применении данных способов в качестве инструментов исследования для диагностики, детектирования, мониторинга и развития уровня техники во всех областях лечения и исследования злокачественных заболеваний. В данном изобретении предложены антитела, а также их связывающие фрагменты и молекулы, полученные из них методом генной инженерии, которые связываются с белками STEAP-1 и полипептидными фрагментами белков STEAP-1. В данном описании термин STEAP-1 является синонимом 8P1D4. Данное изобретение включает в себя поликлональные и моноклональные антитела, антитела мышей и других млекопитающих, химерные антитела, гуманизированные и полноразмерные человеческие антитела, а также антитела, меченные детектируемым маркером или терапевтическим средством. В некоторых воплощениях существует условие, что полная нуклеотидная последовательность (фиг. 2) не кодируется и/или полная аминокислотная последовательность (фиг. 2) не синтезируется. В других воплощениях полная нуклеотидная последовательность (фиг. 2) кодируется и/или полная аминокислотная последовательность (фиг. 2) синтезируется, каждая из которых входит в состав стандартных лекарственных форм для применения человеком. В данном изобретении также предложены способы детектирования присутствия и статуса полинуклеотидов и белков STEAP-1 в разных биологических образцах, а также способы идентификации клеток, экспрессирующих STEAP-1. В одном воплощении согласно изобретению предложены способы детектирования генных продуктов STEAP-1 в ткани или гематологическом образце, в котором имеет место, или предположительно имеет место, нарушение регуляции роста, например, в образце раковой ткани. В данном изобретении также предложены разные иммуногенные или терапевтические композиции и стратегии для лечения злокачественных заболеваний, при которых происходит экспрессия STEAP-1, таких как злокачественные заболевания тканей, перечисленные в табл. I, в том числе способы лечения, обспечивающие ингибирование транскрипции, трансляции, процессинга или функционирования STEAP-1, а также противораковые вакцины. В одном аспекте согласно изобретению предложены композиции и способы с применением данных композиций для лечения рака, при котором происходит экспрессия STEAP-1, у человека, где композиция содержит носитель, подходящий для человека, и человеческую разовую дозу одного или нескольких средств, ингибирующих продукцию или функционирование STEAP-1. Предпочтительно носитель представляет собой однозначно человеческий носитель. В другом аспекте согласно изобретению средство представляет собой молекулу, обладающую иммунореактивностью по отношению к белку STEAP-1. Неограничивающие примеры таких молекул включают в себя, без ограничения, антитела (такие как одноцепочечные, моноклональные, поликлональные, гуманизированные, химерные или человеческие антитела), их функциональные эквиваленты (либо природные, либо синтетические) и их сочетания. Антитела можно конъюгировать с диагностическим или терапевтическим фрагментом. В другом аспекте средство представляет собой маленькую молекулу, как указано в данном описании. В другом воплощении средство включает в себя один или несколько пептидов, содержащих эпитоп цитотоксического Т-лимфоцита (CTL), который связывается с молекулой HLA класса I в организме человека и вызывает ответ CTL на STEAP-1, и/или один или несколько пептидов, содержащих эпитоп хелперного Т-лимфоцита (HTL), который связывается с молекулой HLA класса II в организме человека и вызывает ответ HTL. Пептиды согласно изобретению могут находиться на одной и той же или на одной или нескольких разных полипептидных молекулах. В другом аспекте согласно изобретению средство включает в себя одну или несколько молекул нуклеиновых кислот, экспрессирующих один или несколько пептидов, стимулирующих ответ CTL или HTL, как описано выше. В следующем аспекте согласно изобретению одна или несколько молекул нуклеиновых кислот могут экспрессировать фрагмент, обладающий иммунологической реактивностью в отношении STEAP-1, как описано выше. Одна или несколько молекул нуклеиновых кислот могут представлять собой молекулы, которые ингибируют продукцию STEAP-1, или они могут кодировать такие молекулы. Неограничивающие примеры таких молекул включают в себя, без ограничения, молекулы, комплементарные нуклеотидной последовательности, необходимой для продукции STEAP-1 (например, антисмысловые последовательности молекул, которые образуют тройную спираль с нуклеотидной двойной спиралью, необходимой для продукции STEAP-1), или рибозим, способный лизировать мРНК STEAP-1. Другое воплощение согласно изобретению относится к эпитопам антител, включающим в себя пептидные участки, или олигонуклеотидам, кодирующим пептидные участки, которые имеют один, два, три, четыре или пять из нижеследующих признаков: i) пептидный участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю гидрофильности, приведенному на фиг. 5, равное или превышающее 0,5, 0,6, 0,7, 0,8, 0,9 или равное 1,0; ii) пептидный участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю гидропатичности, приведенному на фиг. 6, равное или превышающее 0,5, 0,4, 0,3, 0,2, 0,1 или равное 0,0; iii) пептидный участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю процента доступных остатков, приведенному на фиг. 7, равное или превышающее 0,5, 0,6, 0,7, 0,8, 0,9 или равное 1,0; iv) пептидный участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю средней жесткости, приведенному на фиг. 8, равное или превышающее 0,5, 0,6, 0,7, 0,8, 0,9 или равное 1,0; или v) пептидный участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю бета-складки, приведенному на фиг. 9, равное или превышающее 0,5, 0,6, 0,7, 0,8, 0,9 или равное 1,0. Настоящее изобретение также относится к гену, обозначаемому STEAP-1, сверхэкспрессия которого наблюдается при злокачественных заболеваниях, перечисленных в табл. I. Анализ экспрессии гена STEAP-1 методом нозерн-блоттинга в нормальных тканях показывает ограниченный характер экспрессии во взрослых тканях. Приведены нуклеотидная (фиг. 2) и аминокислотная (фиг. 2 и 3) последовательности STEAP-1. Тканевый профиль STEAP-1 для нормальных взрослых тканей, наряду со сверхэкспрессией, наблюдающейся в тканях, перечисленных в табл. I, показывает, что, по меньшей мере, в некоторых раковых опухолях наблюдается аберрантная сверхэкспрессия STEAP-1, следовательно, STEAP-1 может служить мишенью в способах диагностики, профилактики, прогноза и/или лечения злокачественных заболеваний тканей, таких как перечисленные в табл. I. Данное изобретение предлагает полинуклеотиды, соответствующие или комплементарные полноразмерным или частичным генам, мРНК и/или кодирующим последовательностям STEAP-1, предпочтительно выделенные, в том числе полинуклеотиды, кодирующие белки, родственные STEAP-1, и фрагменты, содержащие 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более 25 последовательных аминокислот; по меньшей мере 30, 35, 40, 45, 50, 55, 60, 65, 70, 80, 85, 90, 95, 100 или более 100 последовательных аминокислот белка, родственного STEAP-1, a также сами пептиды/белки; ДНК, РНК, ДНК/РНК гибриды и родственные молекулы, полинуклеотиды или олигонуклеотиды, комплементарные или по меньшей мере на 90% гомологичные последовательностям генов или мРНК STEAP-1 или их частям, и полинуклеотиды или олигонуклеотиды, которые гибридизуются с генами и мРНК STEAP-1 или с полинуклеотидами, кодирующими STEAP-1. Также предлагаются способы выделения кДНК и генов, кодирующих STEAP-1. Кроме того, предлагаются рекомбинантные молекулы ДНК, содержащие полинуклеотиды STEAP-1, клетки, трансформированные или преобразованные такими молекулами, и системы хозяин-вектор для экспрессии генных продуктов STEAP-1. В данном описании термин STEAP-1 включает в себя все варианты, в том числе приведенные на фиг. 10, 11 и/или 12, если контекст однозначно не указывает иначе. Фиг. 1. SSH последовательность STEAP-1, содержащая 436 нуклеотидов. Фиг. 2А. кДНК и аминокислотная последовательность 1 варианта STEAP-1 (также называемого "STEAP-1 v. 1" или "1 вариант STEAP-1"). Стартовый метионин подчеркнут. В открытую рамку считывания входят нуклеотиды 66-1085, в том числе стоп-кодон. Фиг. 2В. кДНК и аминокислотная последовательность 2 варианта STEAP-1 (также называемого "STEAP-1 v. 2"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2С. кДНК и аминокислотная последовательность 3 варианта STEAP-1 (также называемого "STEAP-1 v. 3"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-944, в том числе стоп-кодон. Фиг. 2D. кДНК и аминокислотная последовательность 4 варианта STEAP-1 (также называемого "STEAP-1 v. 4"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2Е. кДНК и аминокислотная последовательность 5 варианта STEAP-1 (также называемого "STEAP-1 v. 5"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2F. кДНК и аминокислотная последовательность 6 варианта STEAP-1 (также называемого "STEAP-1 v. 6"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2G. кДНК и аминокислотная последовательность 7 варианта STEAP-1 (также называемого "STEAP-1 v. 7"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2Н. кДНК и аминокислотная последовательность 8 варианта STEAP-1 (также называемого "STEAP-1 v. 8"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2I. кДНК и аминокислотная последовательность 9 варианта STEAP-1 (также называемого "STEAP-1 v. 9"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2J. кДНК и аминокислотная последовательность 10 варианта STEAP-1 (также называемого "STEAP-1 v. 10"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2K. кДНК и аминокислотная последовательность 11 варианта STEAP-1 (также называемого "STEAP-1 v. 11"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2L. кДНК и аминокислотная последовательность 12 варианта STEAP-1 (также называемого "STEAP-1 v. 12"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2М. кДНК и аминокислотная последовательность 13 варианта STEAP-1 (также называемого "STEAP-1 v. 13"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2N. кДНК и аминокислотная последовательность 14 варианта STEAP-1 (также называемого "STEAP-1 v. 14"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2O. кДНК и аминокислотная последовательность 15 варианта STEAP-1 (также называемого "STEAP-1 v. 15"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2Р. кДНК и аминокислотная последовательность 16 варианта STEAP-1 (также называемого "STEAP-1 v. 16"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. Фиг. 2Q. кДНК и аминокислотная последовательность 17 варианта STEAP-1 (также называемого "STEAP-1 v. 17"). Кодон, кодирующий стартовый метионин, подчеркнут. В открытую рамку считывания входят нуклеотиды 96-872, в том числе стоп-кодон. В данном описании термин STEAP-1 включает в себя все варианты, в том числе приведенные на фиг. 10, 11 и/или 12, если контекст однозначно не указывает иначе. Фиг. 3A. Аминокислотная последовательность STEAP-1 v. 1, содержащая 339 аминокислот. Фиг. 3В. Аминокислотная последовательность STEAP-1 v. 2, содержащая 258 аминокислот. Фиг. 3С. Аминокислотная последовательность. STEAP-1 v. 3, содержащая 282 аминокислоты. Фиг. 3D. Аминокислотная последовательность STEAP-1 v. 4, содержащая 258 аминокислот. Фиг. 4А. Сравнительный анализ первичной структуры аминокислотной последовательности STEAP-1 v. 1 и мышиного TNFoc-индуцированного белка жировой ткани (gi/16905133). Фиг. 4В. Сравнительный анализ первичной структуры аминокислотной последовательности STEAP-1 v. 1 и крысиного белка pHyde (gi/21717655/). Фиг. 4С. Показывает гомологию STEAP-1 и мышиного эпителиального антигена простаты, содержащего шесть трансмембранных доменов (gi|20820492|). Фиг. 5A. Гидрофильность аминокислотного профиля 1 варианта STEAP-1. Фиг. 5B. Гидрофильность аминокислотного профиля 3 варианта STEAP-1. Гидрофильность определена путем компьютерного анализа последовательности по способу Норр and Woods (Норр Т.P., Woods K.R., 1981. Proc. Natl. Acad. Sci. U.S.A. 78: 3824-3828), который можно найти на web-сайте ProtScale, расположенном в глобальной сети URL ch/cgi-bin/protscale.pi), через сервер молекулярной биологии ExPasy. Фиг. 6A. Гидропатичность аминокислотного профиля 1 варианта STEAP-1. Фиг. 6B. Гидропатичность аминокислотного профиля 3 варианта STEAP-1, определенная путем компьютерного анализа последовательности по способу Kyte and Doolittle (Kyte J., Doolittle R.F., 1982. J. Mol. Biol. 157:105-132), который можно найти на web-сайте ProtScale, расположенном в глобальной сети URL expasy.ch/cgi-bin/protscale.pi) через сервер молекулярной биологии ExPasy. Фиг. 7A. Профиль процента доступных аминокислотных остатков 1 варианта STEAP-1. Фиг. 7B. Профиль процента доступных аминокислотных остатков 3 варианта STEAP-1, определенный путем компьютерного анализа последовательности по способу Janin (Janin J., 1979 Nature, 277: 491-492), который можно найти на web-сайте ProtScale, расположенном в глобальной сети URL expasy.ch/cgi-bin/protscale.pi) через сервер молекулярной биологии ExPasy. Фиг. 8A. Аминокислотный профиль средней жесткости 1 варианта STEAP-1. Фиг. 8B. Аминокислотный профиль средней жесткости 3 варианта STEAP-1, определенный путем компьютерного анализа последовательности по способу Bhaskaran and Ponnuswamy (Bhaskaran R., and Ponnuswamy P.K., 1988. Int. J. Pept. Protein Res. 32: 242-255), который можно найти на web-сайте ProtScale, расположенном в глобальной сети URL expasy.ch/cgi-bin/protscale.pi) через сервер молекулярной биологии ExPasy. Фиг. 9A. Аминокислотный профиль бета-складки 1 варианта STEAP-1. Фиг. 9B. Аминокислотный профиль бета-складки 3 варианта STEAP-1, определенный путем компьютерного анализа последовательности по способу Deleage and Roux (Deleage, G., Roux B. 1987 Protein Engineering, 1: 289-294), который можно найти на web-сайте ProtScale, расположенном в глобальной сети URL expasy.ch/cgi-bin/protscale.pi) через сервер молекулярной биологии ExPasy. Фиг. 10. Схематический сравнительный анализ вариантов SNP STEAP-1. Варианты STEAP-1 v. 4-v. 17 представляют собой варианты с единичными нуклеотидными различиями по сравнению со STEAP-1 v. 2. Хотя указанные варианты SNP показаны отдельно, они могут встречаться в любых сочетаниях и в любых транскрипционных вариантах, которые содержат пары оснований, например STEAP-1 v. 1 и v. 3. Нумерация соответствует нумерации STEAP-1 v. 2. Черные прямоугольники обозначают последовательности, совпадающие с последовательностями STEAP-1 v. 2. SNP указаны над прямоугольниками. Фиг. 11. Состав экзонов транскрипционных вариантов STEAP-1. На данной фигуре показана структура транскрипционных вариантов без хвоста поли А. Варианты STEAP-1 v. 1, v. 2 и v. 3 представляют собой транскрипционные варианты, содержащие одинаковые экзоны 2 и 3. Первый экзон STEAP-1 v. 1 на 30 оснований короче по 5'-концу, чем первые экзоны двух других транскрипционных вариантов. Четвертый экзон STEAP-1 v. 2 аналогичен объединенным экзону 4, интрону 4 и экзону 5 STEAP-1 v. 1. По сравнению со STEAP-1 v. 1 вариант STEAP-1 v. 3 имеет другой экзон, полученный в результате сплайсинга интрона 4 STEAP-1 v. 1. Интроны и экзоны имеют неодинаковую длину. Фиг. 12. Схематический сравнительный анализ первичных структур белковых вариантов STEAP-1. Белковые варианты соответствуют нуклеотидным вариантам. Нуклеотидные варианты STEAP-1 v. 5 - v. 17, приведенные на фиг. 10, кодируют белок, аналогичный STEAP-1 v. 2. Белки, транслируемые из транскрипционных вариантов STEAP-1 v. 1 и v. 3, показанных на фиг. 11, в положении 169 могут содержать аминокислоту F (Phe) или L (Leu). Единичные аминокислотные различия указаны над рамками. Черными прямоугольниками отмечены последовательности, аналогичные последовательности STEAP-1 v. 1. Прямоугольники, заштрихованные разными способами, обозначают разные последовательности. Номера под прямоугольниками соответствуют STEAP-1 v. 1. Фиг. 13A-13C. Прогнозирование вторичной структуры и трансмембранных доменов для белковых вариантов STEAP-1. Вторичную структуру белковых вариантов 1 STEAP-1 (SEQ ID NO: 46), 2 (SEQ ID NO: 47) и 3 (SEQ ID NO: 48) (фиг. 13A-13C соответственно) прогнозируют по способу HNN - Hierarchical Neural Network (Guermeur, 1997, http://pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_nn.html), который можно найти через сервер молекулярной биологии ExPasy, расположенному в глобальной сети, на сайте (.expasy.ch/tools/). Данный способ позволяет прогнозировать присутствие и расположение альфа-спиралей, развернутых цепей и статистических клубков на основе первичной последовательности белка. Также приводится процент белковых молекул, имеющих данную вторичную структуру. Фиг. 13D-13H. Схематически показана вероятность существования трансмембранных участков и ориентация вариантов 1-3 STEAP-1 соответственно, полученные с помощью алгоритма TMpred, Hofmann and Stoffel, в котором используется TMBASE (K. Hofmann, W. Stoffel. TMBASE - А database of membrane spanning protein segments Biol. Chem. Hoppe-Seyler 374: 166, 1993). Фиг. 13E-13I. Схематически показана вероятность существования трансмембранных участков, а также внеклеточная и внутриклеточная ориентация вариантов 1-3 STEAP-1 соответственно, полученные с помощью алгоритма ТМНММ, Sonnhammer, von Heijne, and Krogh (Erik L.L. Sonnhammer, Gunnar von Heijne, and Anders Krogh: A hidden Markov model for predicting transmembrane helices in protein sequences. In Proc. of Sixth Int. Conf. on Intelligent Systems for Molecular Biology, p. 175-182, Ed. J. Glasgow, T. Littlejohn, F. Major, R. Lathrop, D. Sankoff, and C. Sensen Menlo Park, CA: AAAI Press, 1998). Алгоритмы TMpred и ТМНММ можно найти через сервер молекулярной биологии ExPasy в глобальной сети на сайте (.expasy.ch/tools/). Фиг. 14A. Экспрессия STEAP-1 в образце пациента с раком желудка. РНК экстрагируют из нормального желудка (N) и из образцов от 10 разных пациентов с раком желудка (Т). Наличие последовательности STEAP-1 определяют методом нозерн-блоттинга, используя 10 мкг общей РНК на полосу. Результаты показывают, что в тканях опухоли желудка происходит интенсивная экспрессия STEAP-1 размером приблизительно 1,6 т.о. На нижней панели представлено окрашивание блоттинга бромидом этидия, демонстрирующее качество образцов РНК. Фиг. 14B. Экспрессия STEAP-1 в тканях пациента с раком прямой кишки. РНК экстрагируют из нормальной прямой кишки (N), из опухолевых тканей пациентов с раком прямой кишки (Т) и из метастаз рака прямой кишки (М). Наличие последовательности STEAP-1 определяют методом нозерн-блоттинга, используя 10 мкг общей РНК на полосу. Результаты показывают, что в тканях пациентов с раком прямой кишки происходит интенсивная экспрессия STEAP-1. На нижней панели представлено окрашивание блоттинга бромидом этидия, демонстрирующее качество образцов РНК. Фиг. 14C. Экспрессия STEAP-1 в эндотелиальных клетках пупочной вены человека (HUVEC). Одноцепочечную кДНК получают из клеток HUVEC, ксенотрансплантатов рака простаты LAPC-4AD и LAPC-9AD, а также из тканей мозга человека. Нормализацию проводят методом ПЦР, используя праймеры для актина и GAPDH. Полуколичественную ПЦР с праймерами для STEAP-1, проводят, используя 27 и 30 циклов амплификации (А). В качестве контроля проводят ПЦР с использованием праймеров для актина (В). Результаты показывают, что в клетках HUVEC происходит интенсивная экспрессия STEAP-1, как и в тканях ксенотрансплантатов рака простаты. Экспрессия STEAP-1 в клетках HUVEC показывает, что STEAP-1 можно использовать в качестве маркера эндотелиальных клеток при реваскуляризации опухолей. Фиг. 14D и 14E. Экспрессия STEAP-1 в нормальных и раковых тканях. Одноцепочечную кДНК получают из нормальных тканей (мочевого пузыря, мозга, сердца, почек, печени, легких, простаты, селезенки, скелетной мускулатуры, яичек, поджелудочной железы, толстой кишки и желудка) и из пулов раковых тканей пациентов (из пула рака простаты, пула рака мочевого пузыря, пула рака почек, пула рака прямой кишки, пула рака легких, пула рака яичников, пула рака молочной железы, пула раковых метастаз, пула рака поджелудочной железы, пула ксенотрансплантатов рака простаты и пула метастаз рака простаты в лимфоузлах). Нормализацию проводят методом ПЦР с использованием праймеров для актина. Полуколичественную ПЦР с праймерами для STEAP-1 проводят, используя 26 и 30 циклов амплификации. Фиг. 14D. Изображение агарозного геля с продуктами ОТ-ПЦР. Фиг. 14E. Результаты количественного определения продуктов ПЦР с помощью программного обеспечения Alphalmager. Результаты показывают, что среди всех анализируемых нормальных тканей интенсивная экспрессия наблюдается в простате. Повышающая регуляция экспрессии STEAP-1 обнаружена в пуле рака простаты, пуле рака мочевого пузыря, пуле рака почек, пуле рака прямой кишки, пуле рака легких, пуле рака яичников, пуле рака молочной железы и пуле рака поджелудочной железы. Интенсивная экспрессия STEAP-1 обнаружена в пуле раковых метастаз, пуле ксенотрансплантатов рака простаты и в пуле метастаз рака простаты в лимфоузлах. Фиг. 14F. Экспрессия STEAP-1 в лимфомных образцах пациентов. Нормализацию проводят методом ПЦР с использованием праймеров для актина. Полуколичественную ПЦР с праймерами для STEAP-1 проводят, используя 26 и 30 циклов амплификации. Образцы анализируют на агарозном геле и с помощью программного обеспечения Alphalmager определяют количество продуктов ПЦР. Экспрессию считают сильной или средней, если сигнал детектируется после 26 или 30 циклов амплификации соответственно, и считают, что экспрессия отсутствует, если сигнал не детектирутся даже после 30 циклов амплификации. Результаты показывают, что экспрессия STEAP-1 наблюдается в 8 из 11 анализируемых опухолевых образцов (72,7%). Фиг. 15. Специфическое окрашивание STEAP-1 на поверхности клеток с помощью Mab M2/92.30. Левые секторы: анализ методом FACS рекомбинантных клеток 3Т3 и RAT1, стабильно экспрессирующих либо STEAP-1 (темные линии), либо контрольный ген устойчивости к неомицину (светлые линии), где окрашивание проводят с помощью Mab M2/92.30 против STEAP (10 мкг/мл), а связанные с клеточной поверхностью Mab детектируют, используя в качестве вторичного реагента конъюгат козлиных антител против мышиных IgG с РЕ. Затем окрашенные клетки анализируют методом FACS. Сдвиг флуоресценции в клетках RAT1-STEAP-1 и 3T3-STEAP-1 по сравнению с контрольными клетками показывает, что Mab M2/92.30 специфически связываются со STEAP-1 на клеточной поверхности. Правый сектор: флуоресцентная микроскопия клеток 3T3-STEAP-1, окрашенных Mab M2/92.30, демонстрирует интенсивную флуоресценцию клеточной поверхности. Фиг. 16. STEAP-1 М2/92.30 Mab распознает STEAP-1 на поверхности клеток ксенотрансплантатов рака простаты. Клетки рака простаты LAPC9 окрашивают, используя 10 мкг/мл либо Mab М2/92.30, либо контрольные Mab против KLH. Связанные с поверхностью Mab детектируют, используя в качестве вторичных Ab конъюгат козлиных антител против мышиных IgG с РЕ. Затем окрашенные клетки анализируют методом FACS. Полученные результаты показывают, что Mab M2/120.545 против STEAP-1 специфически связывается с эндогенным STEAP-1, экспрессирующимся на поверхности клеток ксенотрансплантатов рака простаты. Фиг. 17. STEAP-1 М2/92.30 Mab распознает мышиный STEAP-1. Клетки 293Т транзиторно трансфицируют либо pCDNA3.1, кодирующим кДНК мышиного STEAP-1, либо пустым вектором. Через 2 дня клетки собирают и окрашивают Mab M2/92.30 против STEAP-1 (10 мкг/мл), после чего связанные с клеточной поверхностью Mab детектируют, используя в качестве вторичного реагента конъюгат козлиных антител против мышиных IgG с РЕ. Затем клетки анализируют методом FACS. Сдвиг флуоресценции на клетках 293Т, трансфицированных мышиным STEAP-1, по сравнению с клетками, трансфицированными пустым вектором, показывает, что Mab М2/92.30 специфически связываются с мышиным белком STEAP-1. Фиг. 18. STEAP-1/120.545 Mab распознает STEAP-1 на клеточной поверхности. Сектор А и сектор В. Клетки 3Т3-neo (А, закрашенная гистограмма) и 3Т3-STEAP-1 (А, незакрашенная гистограмма) и клетки RAT1-neo (В, закрашенная гистограмма) и RAT1-STEAP (В, незакрашенная гистограмма) окрашивают Mab М2/120.545 (10 мкг/мл) и связанные с поверхностью Mab детектируют, используя в качестве вторичных Ab конъюгат козлиных антител против мышиных IgG с РЕ. Затем клетки анализируют методом FACS. Сдвиг флуоресценции на клетках 3Т3-STEAP-1 и RAT1-STEAP-1 по сравнению с соответствующими neo-контролями показывает, что Mab M2/120.545 специфически связываются с STEAP-1 на клеточной поверхности. Сектор С. Клетки LNCaP окрашивают либо Mab М2/120.545, либо контрольными Mab против KLH и анализируют методом FACS, как описано выше. Сектор D. Флуоресцентная микроскопия клеток LNCaP, окрашенных М2/120.545, демонстрирует интенсивную флуоресценцию клеточной поверхности. Полученные результаты показывают, что Mab М2/120.545 специфически связывается с эндогенным STEAP-1 на поверхности клеток LNCaP. Фиг. 19A. кДНК (SEQ ID NO: 49) и аминокислотная последовательность (SEQ ID NO: 50) М2/Х92.30 V H , клон #2. Фиг. 19B кДНК (SEQ ID NO: 51) и аминокислотная последовательность (SEQ ID NO: 52) М2/Х92.30 V L , клон #2. Фиг. 19C. кДНК (SEQ ID NO: 53) и аминокислотная последовательность (SEQ ID NO: 54) М2/Х92.30 V L , клон #6. Фиг. 19D. кДНК (SEQ ID NO: 55) и аминокислотная последовательность (SEQ ID NO: 56) М2/Х120.545 V L , клон #8. Фиг. 20A. Аминокислотная последовательность (SEQ ID NO: 57) М2/Х92.30 V H , клон #2. Фиг. 20B. Аминокислотная последовательность (SEQ ID NO: 58) М2/Х92.30 V L , клон #2. Фиг. 20C. кДНК (SEQ ID NO: 59) и аминокислотная последовательность (SEQ ID NO: 60) М2/Х92.30 V L , клон #6. Фиг. 20D. Сравнительный анализ амнокислотной последовательности М2/Х92.30 V L , клон #2 (SEQ ID NO: 61), и М2/М92.30 V L , клон #6 (SEQ ID NO: 62). Фиг. 20E. Аминокислотная последовательность (SEQ ID NO: 63) М2/Х120.545 V L , клон #8. Последовательность сигнального пептида подчеркнута. Фиг. 21. STEAP-1 М2/92.30 Mab распознает STEAP-1 на поверхности клеток ксенотрансплантатов рака простаты и мочевого пузыря человека. Клетки мочевого пузыря UGB1 (левый сектор) и клетки рака простаты LAPC9 (правый сектор) окрашивают, используя 10 мкг/мл либо Mab M2/92.30, либо контрольных Mab против KLH. Связанные с поверхностью Mab детектируют, используя в качестве вторичных Ab конъюгат козлиных антител против мышиных IgG с РЕ. Затем окрашенные клетки анализируют методом FACS. Полученные результаты показывают, что Mab M2/92.30 против STEAP-1 специфически связывается с эндогенным STEAP-1, экспрессирующимся на поверхности клеток ксенотрансплантатов рака мочевого пузыря и простаты. Фиг. 22. Интернализация STEAP-1 под действием STEAP-1/92.30 Mab. Клетки 3T3-STEAP-1 окрашивают при 4 °С с помощью М2/92.30 Mab (10 мкг/мл), промывают и инкубируют с конъюгатом козлиные антитела против мышиных IgG-РЕ в качестве вторичных Ab при 4 °С. Половину клеток инкубируют при 37 °С в течение 30 мин, а вторую половину оставляют при 4 °С. Затем клетки из каждой партии анализируют методом флуоресцентной микроскопии. На поверхности клеток, оставленных инкубироваться при 4 °С, наблюдается яркая "кольцеобразная" флуоресценция. На поверхности клеток, инкубировавшихся при 37 °С, "кольцеобразная" флуоресценция отсутствует, а появление точечной и агрегированной флуоресценции указывает на кэпирование и интернализацию. Фиг. 23. Интернализация STEAP-1 под действием STEAP-1 М2/120.545 Mab. Клетки PC3-STEAP-1 окрашивают при 4 °С с помощью М2/120.545 Mab (10 мкг/мл), промывают и инкубируют с конъюгатом козлиные антитела против мышиных IgG-РЕ в качестве вторичных Ab. Половину клеток инкубируют при 37 °С в течение 30 мин, а вторую половину оставляют при 4 °С. Затем клетки из каждой партии анализируют методом флуоресцентной микроскопии. На поверхности клеток, оставленных инкубироваться при 4 °С, наблюдается яркая "кольцеобразная" флуоресценция. На поверхности клеток, инкубировавшихся при 37 °С, "кольцеобразная" флуоресценция отсутствует, а появление точечной и агрегированной флуоресценции указывает на кэпирование и интернализацию. Фиг. 24. Интернализация STEAP-1. Для того чтобы продемонстрировать, что мышиный STEAP-1 M2/120.545 проникает в клетки-мишени благодаря экспрессии STEAP-1 на поверхности клеток LNCaP, используют конъюгаты антител против мышиных IgG с сапорином (Advanced Targeting Systems, San Diego, CA). Анализ проводят следующим образом. Клетки LNCaP помещают в количестве 5000 клеток/90 мкл/лунку в 96-луночный планшет и инкубируют в течение ночи. Получают растворы конъюгатов вторичных антител с иммунотоксином (антитела против мышиных IgG-сапорин и антитела против козлиных IgG-сапорин) или антител против мышиных IgG в среде для культивирования клеток с конечной концентрацией 100 нг/мл. Добавляют первичное антитело в концентрации, варьирующей от 1 до 1000 нг/мл. Планшеты инкубируют в течение 72 ч, после чего методом МТТ определяют жизнеспособность. Результаты показывают, что клетки LNCaP погибают в присутствии М2/120.545 и конъюгата антитело против мышиных IgG-сапорин. При использовании только вторичных антител (против мышиных IgG) или конъюгатов неспецифичных вторичных антител (антител против козлиных IgG с сапорином) эффект не детектируется. При использовании только первичного антитела (М2/120.545) вплоть до концентрации 1 мкг/мл токсичность не наблюдается. Фиг. 25. Иммунопреципитация STEAP-1 под действием Mab против STEAP-1 M2/92.30 и М2/120.545. Клетки 3T3-STEAP-1 и 3Т3-neo лизируют в буфере RIPA (25 мМ Tris-Cl pH 7,4; 150 мМ NaCl, 0,5 мМ EDTA, 1% тритон Х-100, 0,5% дезоксихолевая кислота, 0,1% SDS и смесь ингибиторов протеаз). Клеточные лизаты предварительно осветляют с помощью гранул сефарозы, содержащих белок G, и затем инкубируют с 5 мкг Mab либо М2/92.30, либо М2/120.545 в течение 2 ч при комнатной температуре. Добавляют гранулы, содержащие белок G, и смесь инкубируют еще 1 ч. Иммунные комплексы промывают и солюбилизируют в буфере для SDS-PAGE. Затем солюбилизированные образцы анализируют методами SDS-PAGE и вестерн-блоттинга, используя кроличьи pAb против STEAP. В качестве положительного контроля используют лизаты целых клеток 293Т, трансфицированных STEAP-1. Иммунореактивная полоса, соответствующая ~37 кДа, наблюдается только в образцах, полученных из клеток 3T3-STEAP-1, и указывает на специфичную иммунопреципитацию STEAP-1 под действием как Mab М2/92.30, так и Mab М2/120.545. Фиг. 26. Влияние STEAP-1 Mab на рост ксенотрансплантатов рака простаты человека LAPC9 у мышей. STEAP-1 M2/92.30 и М2/120.545 тестируют в двух разных дозах, 100 и 500 мкг. В качестве контролей используют PBS и Mab против KLH. В исследовании участвуют 6 групп по 10 мышей в каждой. Mab вводят в.б. два раза в неделю, всего 12 раз, начиная со дня инъекции опухолевых клеток. Размер опухоли измеряют штангенциркулем два раза в неделю. Объем опухоли рассчитывают по формуле W 2 ×L/2, где L - самый длинный размер; W - перпендикулярный ему размер. Концентрацию PSA в сыворотке измеряют на 40 день после начала обработки с помощью коммерческого набора ELISA. Чтобы учесть различия в росте опухоли и уровне PSA между группами, используют тест Крускала-Валлиса и U-тест Манна-Уитни. Все тесты являются двусторонними с α=0,05. Полученные результаты показывают, что STEAP-1 M2/92.30 и М2/120.545 значительно замедляют рост ксенотрансплантата опухоли простаты человека в зависимости от используемой дозы. Фиг. 27. Влияние STEAP-1 Mab на рост ксенотрансплантатов рака простаты человека LAPC9 у мышей. STEAP-1 M2/92.30 и М2/120.545 тестируют в двух разных дозах, 100 и 500 мкг. В качестве контролей используют PBS и Mab против KLH. В исследовании участвуют 6 групп по 10 мышей в каждой. Mab вводят в.б. два раза в неделю, всего 12 раз, начиная со дня инъекции опухолевых клеток. Размер опухоли измеряют штангенциркулем два раза в неделю. Измеряют самый длинный размер L и перпендикулярный ему размер W, после чего объем опухоли определяют по формуле W 2 ×L/2. Концентрацию PSA в сыворотке измеряют на 40 день после начала обработки с помощью коммерческого набора ELISA. Чтобы учесть различия в росте опухоли и уровне PSA между группами, используют тест Крускала-Валлиса и U-тест Манна-Уитни. Все тесты являются двусторонними с α=0,05. Полученные результаты показывают, что STEAP-1 М2/92.30 и М2/120.545 значительно замедляют рост ксенотрансплантата опухоли простаты человека в дозозависимой манере. Фиг. 28. STEAP-1-индуцированное фосфорилирование ERK-1 и ERK-2. Левые секторы: клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 в векторе pSR α. Клетки выращивают в течение ночи в 0,5% FBS, затем стимулируют 10% FBS в течение 5 мин в присутствии или в отсутствие 10 мкг/мл ингибитора MEK PD98058. Клеточные лизаты анализируют методами 12,5% SDS-PAGE и вестерн-блоттинга с использованием антител против фосфо-ERK (Cell Signaling) и антител против ERK (Zymed). Правые секторы: клетки NIH-3T3 трансфицируют только геном устойчивости к неомицину или STEAP-1 в векторе pSR α. Клетки обрабатывают, как описано выше, но в отсутствие ингибитора MEK. Кроме того, клетки NIH-3T3-neo обрабатывают 10 мг/мл салицилата Na. Экспрессия STEAP-1 индуцирует фосфорилирование ERK-1 и ERK-2 в сыворотке, которое ингибируется вышестоящим ингибитором киназы MEK PD98058. Фиг. 29. STEAP-1 опосредует межклеточное взаимодействие. Клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1, или котрольным геном в векторе pSR α. Реципиентные клетки метят конъюгатом декстран-техасовый красный в концентрации 1 мг/мл, а донорные клетки метят кальцеином AM в концентрации 2,5 мкг/мл. Донорные (зеленые) и реципиентные (красные) клетки совместно культивируют при 37 °С в течение 18-24 ч, после чего с помощью микроскопии анализируют расположение флуоресцентных красителей. Левый сектор: в качестве доноров и реципиентов используют клетки РС3-neo. Центральный сектор: в качестве доноров и реципиентов используют клетки PC3-STEAP-1. Правый сектор: в качестве доноров и реципиентов используют РС3-контрольные клетки. STEAP-1 индуцирует перенос кальцеина в клетки, содержащие конъюгат декстран-техасовый красный, данный результат указывает на то, что STEAP-1 облегчает межклеточные взаимодействия. Фиг. 30. Для межклеточного взаимодействия требуется экспрессия STEAP-1 на донорных и реципиентных клетках. Клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 в векторе pSR α. Реципиентные клетки метят конъюгатом декстран-техасовый красный в концентрации 1 мг/мл, а донорные клетки метят кальцеином AM в концентрации 2,5 мкг/мл. Донорные (зеленые) и реципиентные (красные) клетки совместно культивируют при 37 °С в течение 18-24 ч, после чего с помощью микроскопии анализируют расположение флуоресцентных красителей. Верхние секторы: оптическая микроскопия; нижние секторы: УФ-флуоресценция. Левые секторы: в качестве доноров и реципиентов используют клетки РС3-neo. Центральные секторы: в качестве донорных клеток используют РС3-neo, а в качестве реципиентных клеток используют PC3-STEAP-1. Правые панели: в качестве доноров и реципиентов используют клетки PC3-STEAP-1. Межклеточные взаимодействия детектируют только в том случае, если STEAP-1 экспрессируется и на донорных, и на реципиентных клетках. Фиг. 31. Влияние Mab STEAP-1/120.545 на щелевой контакт. Клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 в векторе pSR α. Реципиентные клетки метят конъюгатом декстран-техасовый красный в концентрации 1 мг/мл, а донорные клетки метят кальцеином AM в концентрации 2,5 мкг/мл. Донорные (зеленые) и реципиентные (красные) клетки совместно культивируют при 37 °С в течение 18-24 ч, после чего с помощью микроскопии анализируют расположение флуоресцентных красителей. Во всех экспериментах в качестве доноров и реципиентов используют одинаковые клетки. Клетки инкубируют с указанными количествами Mab STEAP-1/120.545 в течение 10 мин перед внесением в планшеты, затем Mab оставляют в культуре на 24 ч до анализа. STEAP-1/120.545 уменьшает щелевой контакт, опосредованный STEAP-1, в зависимости от используемой дозы. Фиг. 32. Ингибирование фосфорилирования ERK-1 и ERK-2 под действием STEAP-1 Mab и РНКи. Клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 и Mab в векторе pSR α. Для удаления РНКи клетки РС3-STEAP-1 стабильно трансфицируют вектором pPUR-U6-27-STEAP-1, содержащим киРНК (короткие интерферирующие РНК) STEAP-1. Клетки держат в условиях недостаточного количества питательных веществ в 0,1% FBS в течение 18 ч при 37 °С, затем помещают на лед, где держат в течение 10 мин в присутствии или в отсутствие 10 мкг/мл Mab M2/92.30, после чего в течение 3 мин нагревают до КТ и стимулируют 10% FBS в течение 5 мин. Клетки лизируют в буфере RIPA, лизаты целых клеток анализируют методом 12,5% SDS-PAGE, а белки детектируют методом вестерн-блоттинга. Фосфо-ERK детектируют с помощью кроличьей антисыворотки (Cell Signaling), a ERK детектируют с помощью кроличьих антител против ERK (Zymed). STEAP-1 детектируют с помощью овечьих антител против STEAP-1, а актин детектируют с помощью Mab против актина (Santa Cruz). Фосфорилирование ERK-1 и ERK-2, индуцированное 10% сывороткой, ингибируется под действием Mab M2/92.30 и киРНК STEAP-1. Фиг. 33. Влияние РНКи STEAP-1 на межклеточное взаимодействие. Клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 в векторе pSR α. Для удаления РНКи клетки PC3-STEAP-1 стабильно трансфицируют вектором pPUR-U6-27-STEAP-1, содержащим киРНК STEAP-1, или пустым вектором. Реципиентные клетки метят конъюгатом декстран-техасовый красный в концентрации 1 мг/мл, а донорные клетки метят кальцеином AM в концентрации 2,5 мкг/мл. Донорные (зеленые) и реципиентные (красные) клетки совместно культивируют при 37 °С в течение 18-24 ч, после чего с помощью микроскопии анализируют расположение флуоресцентных красителей. Во всех экспериментах в качестве доноров и реципиентов используют одинаковые клетки. Специфичная РНКи STEAP-1, стабильно экспрессирующаяся в клетках PC3-STEAP-1, уменьшает межклеточное взаимодействие, индуцированное STEAP-1. Оглавление разделов. I. Определения. II. Полинуклеотиды STEAP-1. II.А. Применение полинуклеотидов STEAP-1. II.A.1. Выявление генетических нарушений. II.А.2. Антисмысловые воплощения. II.A.3. Праймеры и пары праймеров. II.А.4. Выделение молекул нуклеиновых кислот, кодирующих STEAP-1. II.А.5. Молекулы рекомбинантных нуклеиновых кислот и системы векторов-хозяев. III. Белки, родственные STEAP-1. III.А. Воплощения белков, несущих мотив. III.В. Экспрессия белков, родственных STEAP-1. III.С. Модификации белков, родственных STEAP-1. III.D. Применение белков, родственных STEAP-1. IV. Антитела против STEAP-1. V. Клеточные иммунные ответы на STEAP-1. VI. Трансгенные животные, несущие STEAP-1. VII. Способы детектирования STEAP-1. VIII. Способы определения статуса генов, родственных STEAP-1, и их продуктов. IX. Идентификация молекул, взаимодействующих со STEAP-1. X. Терапевтические способы и композиции. Х.А. Противораковые вакцины. Х.В. STEAP-1 как мишень для терапии антителами. Х.С. STEAP-1 как мишень для клеточных иммунных ответов. Х.С.1. Вакцины на основе мини-генов. Х.С.2. Сочетания пептидов CTL с хелперными пептидами. Х.С.3. Сочетания пептидов CTL с факторами, сенсибилизирующими Т-клетки. Х.С.4. Вакцинные композиции, содержащие DC, сенсибилизированные пептидами CTL и/или HTL. X.D. Адоптивная иммунотерапия. Х.Е. Введение вакцин с целью лечения или профилактики. XI. Диагностические и прогностические воплощения STEAP-1. XII. Ингибирование белковой функции STEAP-1. XII.А. Ингибирование STEAP-1 внутриклеточными антителами. XII.В. Ингибирование STEAP-1 рекомбинантными белками. XII.С. Ингибирование транскрипции или трансляции STEAP-1. XII.D. Общие положения стратегий лечения. XIII. Идентификация, характеристика и применение модуляторов STEAP-1. XIV. РНКи и терапевтическое применение маленьких интерферирующих РНК (siRNA). XV. Наборы/готовые изделия. I. Определения. Если не указано иначе, подразумевается, что все используемые в данном описании научные термины, номенклатура или терминология имеют значения, общепринятые в области, к которой относится данное изобретение. В некоторых случаях для ясности и/или в качестве справочного материала в данном описании приводятся определения терминов, имеющих общепринятые значения, но не следует считать, что включение таких определений в данное описание означает, что они существенно отличаются от общепринятых в данной области. Многие методы и процедуры, описанные в данном документе, являются хорошо известными и широко применяются специалистами в данной области в рамках традиционной методологии, такой, например, как широко используемые методы молекулярного клонирования, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2 nd edition (1989), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. Если не указано иначе, процедуры, включающие в себя применение коммерческих наборов и реагентов, обычно проводят в соответствии с инструкциями и/или параметрами производителя. Термины "распространенный рак простаты", "местно распространенный рак простаты", "распространенное заболевание" и "местно распространенное заболевание" относятся к заболеваниям простаты, распространившимся из капсулы простаты, и включают в себя заболевания стадии С по системе американской урологической ассоциации (AUA), заболевания стадий С1-С2 по системе Уитмора-Джеветта (Whitmore-Jewett) и заболевания стадий Т3-Т4 и N+ по системе TNM (опухоль, узел, метастазы). Как правило, не рекомендуется применять хирургическое лечение для пациентов с местно распространенным заболеванием, такие пациенты имеют гораздо менее благоприятный исход, чем пациенты с клинически локализованным (ограниченным органом) раком простаты. Местно распространенное заболевание идентифицируют при обнаружении путем пальпации уплотнения за боковой границей простаты, или асимметрии, или уплотнения выше основания простаты. Местно распространенный рак простаты в настоящее время диагностируют патологически после радикальной простатэктомии, если опухоль внедряется или проникает в простатическую капсулу, простирается за границы оперируемой области или проникает в семенные пузырьки. Термин "изменение природного характера гликозилирования" в данном описании означает делецию одного или нескольких углеводных фрагментов, присутствующих в нативной последовательности STEAP-1 (либо в результате удаления нижестоящего участка гликозилирования, либо в результате делеции гликозилирования химическими или ферментативными способами), и/или добавление одного или нескольких участков гликозилирования, не присутствующих в нативной последовательности STEAP-1. Кроме того, данная фраза включает в себя количественные изменения в гликозилировании нативных белков, включающие в себя изменения природы и соотношения разных присутствующих углеводных фрагментов. Термин "аналог" относится к молекуле, которая имеет структуру, подобную структуре другой молекулы, или признаки, подобные или аналогичные признакам другой молекулы (например, белка, родственного STEAP-1). Например, аналог белка STEAP-1 может специфически связываться с антителом или Т-клеткой, которые специфически связываются со STEAP-1. Если однозначно не указано иначе, термин "антитело" используется в самом широком смысле. Следовательно, "антитело" может быть природным или искусственным, таким как моноклональные антитела, полученные с помощью традиционной гибридомной технологии. Антитела против STEAP-1 включают в себя моноклональные и поликлональные антитела, а также фрагменты, содержащие антигенсвязывающий домен и/или один или несколько гипервариабельных участков данных антител. В данном описании термин "антитело" относится к антителам любого вида или к их фрагментам, которые специфически связываются со STEAP-1 и/или обладают желательной биологической активностью, и, в частности, включает в себя моноклональные антитела (в том числе полноразмерные моноклональные антитела), поликлональные антитела, поливалентные антитела (например, биспецифические антитела) и фрагменты антител, которые специфически связываются со STEAP-1 и/или обладают желательной биологической активностью. В способах и композициях, описанных в данном документе, можно использовать любое специфическое антитело. Так, в одном воплощении термин "антитело" включает в себя молекулу, содержащую по меньшей мере один вариабельный участок легкой цепи молекулы иммуноглобулина и по меньшей мере один вариабельный участок тяжелой цепи, которые вместе образуют участок специфического связывания антигена-мишени. В одном воплощении антитело представляет собой антитело IgG. Например, антитело может представлять собой IgG 1 , IgG 2 , IgG 3 или IgG 4 . Антитела, используемые в способах и композициях согласно изобретению, могут быть получены в клеточной культуре, в фагах или в разных животных, включающих в себя, без ограничения, коров, кроликов, коз, мышей, крыс, хомяков, морских свинок, овец, собак, кошек, мартышек, шимпанзе, человекообразных обезьян. Таким образом, в одном воплощении антитело согласно изобретению представляет собой антитело млекопитающего. Для выделения исходного антитела или получения вариантов с измененными характеристиками специфичности или авидности можно использовать фаговые технологии. Такие технологии являются стандартными и хорошо известны в данной области. Например, рекомбинантное антитело можно получить путем трансфекции клетки-хозяина вектором, содержащим последовательность ДНК, кодирующую антитело. Для трансфекции последовательности ДНК, экспрессирующей по меньшей мере один V L - и один V H -участок в клетке-хозяине, можно использовать один или несколько векторов. Примеры рекомбинантных способов получения и продукции антител описаны в Delves, ANTIBODY PRODUCTION: ESSENTIAL TECHNIQUES (Wiley, 1997); Shephard, et al., MONOCLONAL ANTIBODIES (Oxford University Press, 2000); Goding, MONOCLONAL ANTIBODIES: PRINCIPLES AND PRACTICE (Academic Press, 1993); CURRENT PROTOCOLS IN IMMUNOLOGY (John Wiley & Sons, most recent edition). Антитело согласно изобретению можно модифицировать рекомбинантными способами с увеличением эффективности антитела при опосредовании целевой функции. Таким образом, в объем согласно изобретению входят модификации антител путем замен с использованием рекомбинантных методов. Как правило, замены являются консервативными. Например, по меньшей мере одна аминокислота в константном участке антитела может быть заменена на другой остаток. См., например, патент США № 5624821, патент США № 6194551, заявка WO 9958572 и Angal, et al., Mol. Immunol. 30: 105-08 (1993). Модификации аминокислотного состава включают в себя делеции, добавления, замены аминокислот. В некоторых случаях такие изменения делают, чтобы уменьшить нежелательную активность, например комплемент-зависимую цитотоксичность. Зачастую антитела метят путем присоединения либо ковалентного, либо нековалентного, вещества, генерирующего детектируемый сигнал. Известен широкий ряд меток и методов присоединения, которые широко описаны в научной и патентной литературе. Данные антитела можно анализировать на связывание с нормальным или дефектным STEAP-1. См., например, ANTIBODY ENGINEERING: A PRACTICAL APPROACH (Oxford University Press, 1996). Подходящие антитела с желательной биологической активностью можно идентифицировать с помощью in vitro анализов, включающих в себя, без ограничения, пролиферацию, миграцию, адгезию, выращивание в мягком агаре, ангиогенез, межклеточные взаимодействия, апоптоз, транспорт, передачу сигнала, а также с помощью in vivo анализов, таких как ингибирование опухолевого роста. Антитела, предлагающиеся в данном описании, также можно использовать для диагностических целей. Улавливающие или не-нейтрализующие антитела можно анализировать на способность связываться со специфическим антигеном, при этом не ингибируя связывание с рецептором или биологическую активность антигена. Нейтрализующие антитела можно использовать в конкурентном анализе связывания. Их также можно использовать для количественного определения STEAP-1 или его рецептора. "Фрагмент антитела" определяют по меньшей мере как часть вариабельного участка молекулы иммуноглобулина, которая связывается с мишенью антитела, т.е. как антигенсвязывающий участок. В одном воплощении данный термин, в частности, охватывает отдельные антитела против STEAP-1 и их клоны (в том числе агонистические, антагонистические и нейтрализующие антитела), а также композиции антител против STEAP-1 с полиэпитопной специфичностью. В способах и композициях согласно изобретению можно использовать моноклональные или поликлональные антитела. Антитело может находиться в виде антигенсвязывающего фрагмента антитела, включающего в себя фрагмент Fab, фрагмент F(ab') 2 , одноцепочечный вариабельный участок и т.п. Фрагменты интактных молекул можно получить с помощью способов, хорошо известных в данной области и включающих в себя ферментативное расщепление и рекомбинантные способы. В данном описании для получения антитела, специфичного к STEAP-1, можно использовать любую форму "антигена". Так, антиген, вызывающий образование антител, может представлять собой один эпитоп, несколько эпитопов или целый белок и может использоваться один или в сочетании с одним или несколькими известными в данной области средствами, повышающими иммуногенность. Антиген, вызывающий образование антител, может представлять собой выделенный полноразмерный белок, белок клеточной поверхности (например, при иммунизации клетками, трансфицированными по меньшей мере частью антигена) или растворимый белок (например, при иммунизации только фрагментом внеклеточного домена белка). Антиген может продуцироваться в генетически модифицированных клетках. ДНК, кодирующая антиген, может быть геномной или негеномной (например, кДНК) и может кодировать по меньшей мере часть внеклеточного домена. В данном описании термин "фрагмент" относится к минимальному числу аминокислот или нуклеиновых кислот, по обстоятельствам, образующих иммуногенный эпитоп рассматриваемого антигена. Для трансформации клеток можно использовать любые подходящие генетические векторы, включающие в себя, без ограничения, аденовирусные векторы, плазмиды и невирусные векторы, такие как катионные липиды. В одном воплощении антитело, используемое в способах и композициях согласно изобретению, специфически связывается по меньшей мере с частью внеклеточного домена рассматриваемого STEAP-1. Описываемые в данном документе антитела или их антигенсвязывающие фрагменты можно конъюгировать с "биоактивным средством". В данном описании термин "биоактивное средство" относится к любому синтетическому или природному соединению, которое связывается с антигеном и/или усиливает или опосредует желательный биологический эффект, связанный с увеличением уровня цитотоксичных веществ. В одном воплощении связывающие фрагменты, используемые в настоящем изобретении, представляют собой биологически активные фрагменты. В данном описании термин "биологически активный" относится к антителу или фрагменту антитела, способному связываться с целевым антигенным эпитопом и непосредственно или косвенно оказывать биологический эффект. Непосредственные эффекты включают в себя, без ограничения, модуляцию, стимуляцию и/или ингибирование сигнала роста; модуляцию, стимуляцию и/или ингибирование антиапоптотического сигнала; модуляцию, стимуляцию и/или ингибирование апоптотического или некротического сигнала; модуляцию, стимуляцию и/или ингибирование каскада ADCC и модуляцию, стимуляцию и/или ингибирование каскада CDC. В способах и композициях согласно изобретению также используются "биспецифические" антитела. В данном описании термин "биспецифическое антитело" относится к антителу, как правило, моноклональному, способному специфично связываться по меньшей мере с двумя антигенными эпитопами. В одном воплощении эпитопы относятся к одному антигену. В другом воплощении эпитопы относятся к двум разным антигенам. Способы получения биспецифических антител известны в данной области. Например, биспецифические антитела можно получить рекомбинантными способами путем совместной экспрессии пар тяжелых и легких цепей двух иммуноглобулинов. См., например, Milstein et al., Nature, 305: 537-39 (1983). Альтернативно, биспецифические антитела можно получить путем химического присоединения. См., например, Brennan, et al., Science, 229: 81 (1985). Биспецифические антитела включают в себя фрагменты биспецифических антител. См., например, Hollinger, et al., Proc. Natl. Acad. Sci. USA. 90: 6444-48 (1993), Gruber, et al., J. Immunol. 152: 5368 (1994). Описанные в данном документе моноклональные антитела, в частности, включают в себя "химерные" антитела, в которых фрагмент тяжелой и/или легкой цепи идентичен или гомологичен соответствующим последовательностям антител, полученных из конкретных видов или принадлежащих к конкретному классу или подклассу антител, тогда как другая цепь (цепи) идентична или гомологична соответствующим последовательностям антител, полученных из других видов или принадлежащих к другому классу или подклассу антител, а также фрагменты таких антител, пока они специфически связываются с целевым антигеном и/или обладают желательной биологической активностью (патент США № 4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984)). Термин "кодон-оптимизированные последовательности" относится к нуклеотидным последовательностям, оптимизированным для конкретных видов путем замены любых кодонов с периодичностью использования менее чем приблизительно 20%. Нуклеотидные последовательности, оптимизированные для экспрессии в определенных видах хозяев путем устранения мнимых последовательностей полиаденилирования, устранения сигналов сплайсинга экзонов/интронов, устранения транспозон-подобных повторов и/или оптимизации содержания GC в добавление к оптимизации кодонов, в данном описании называют "последовательностями с повышенной экспрессией". "Комбинаторная библиотека" представляет собой набор разных химических соединений, полученных путем химического или биологического синтеза в результате объединения ряда химических "структурных элементов", таких как реагенты. Например, линейную комбинаторную химическую библиотеку, такую как библиотеку полипептидов (например, мутеина), получают путем соединения ряда химических структурных элементов, называемых аминокислотами, с получением молекулы заданной длины (т.е. полипептидного соединения, содержащего определенное число аминокислот). Многие химические соединения синтезируют путем такого комбинаторного смешивания химических структурных элементов (Gallop et al., J. Med. Chem. 37(9): 1233-1251 (1994)). Способы получения и скрининга комбинаторных библиотек хорошо известны специалистам в данной области. Такие комбинаторные химические библиотеки включают в себя, без ограничения, библиотеки пептидов (см., например, патент США № 5010175, Furka, Pept. Prot. Res. 37: 487-493 (1991), Houghton et al., Nature, 354: 84-88 (1991)), пептоиды (публикация РСТ № WO 91/19735), кодированные пептиды (публикация РСТ № WO 93/20242), статистические биоолигомеры (публикация РСТ № WO 92/00091), бензодиазепины (патент США № 5288514), диверсомеры, такие как гидантоины, бензодиазепины и дипептиды (Hobbs et al., Proc. Nat. Acad. Sci. USA, 90: 6909-6913 (1993)), винилогические полипептиды (Hagihara et al., J. Amer. Chem. Soc. 114: 6568 (1992)), непептидные пептидомиметики с бета-D-глюкозным скелетом (Hirschmann et al., J. Amer. Chem. Soc. 114: 9217-9218 (1992)), библиотеки маленьких соединений, полученные путем аналогового органического синтеза (Chen et al., J. Amer. Chem. Soc. 116: 2661 (1994)), олигокарбаматы (Cho, et al., Science, 261: 1303 (1993)) и/или пептидилфосфонаты (Campbell et al., J. Org. Chem. 59: 658 (1994)). См., в общем, Gordon et al., J. Med. Chem. 37: 1385 (1994), библиотеки нуклеиновых кислот (см., например, Stratagene, Corp.), библиотеки пептидов и нуклеиновых кислот (см., например, патент США № 5539083), библиотеки антител (см., например, Vaughn et al., Nature Biotechnology, 14 (3), (1996) и PCT/US96/10287), библиотеки углеводов (см., например, Liang et al., Science, 274: 1520-1522 (1996) и патент США № 5593853) и библиотеки маленьких органических молекул, таких как бензодиазепины (Baum, C &EN, Jan 18, p. 33 (1993)); изопреноиды (патент США № 5569588); тиазолидиноны и метатиазаноны (патент США № 5549974); пирролидины (патенты США № 5525735 и 5519134); морфолиновые соединения (патент США № 5506337); бензодиазепины (патент США № 5288514) и т.п. Устройства для получения комбинаторных библиотек имеются в продаже (см., например, 357 NIPS, 390 NIPS, Advanced Chem Tech, Louisville KY; Symphony, Rainin, Woburn, MA; 433A, Applied Biosystems, Foster City, CA; 9050, Plus, Millipore, Bedford, NIA). Для решений фазовой химии также разработан ряд роботизированных систем. Данные системы включают в себя автоматизированные рабочие станции, такие как автоматизированные аппараты синтеза, разработанные Takeda Chemical Industries, LTD. (Osaka, Japan), а также многие системы, в которых используются роботизированные руки (Zymate Н., Zymark Corporation, Hopkinton, Mass.; Orca, Hewlett-Packard, Palo Alto, Calif.), имитирующие синтетические операции, проводимые химиками. Любое из вышеуказанных устройств можно использовать в настоящем изобретении. Специалисты в данной области могут осуществлять модификации данных устройств (если таковые имеют место) в пределах их работоспособности в соответствии с настоящим изобретением. Кроме того, ряд комбинаторных библиотек является коммерчески доступным (см., например, ComGenex, Princeton, NJ; Asinex, Moscow, RU; Tripos, Inc., St. Louis, MO; ChemStar, Ltd., Moscow, RU; 3D Pharmaceuticals, Exton, PA; Martek Biosciences, Columbia, MD; etc.). В данном описании термин "консервативная замена" относится к заменам аминокислот, известным специалистам в данной области, которые, как правило, могут быть осуществлены без изменения биологической активности полученной молекулы. Специалистам в данной области известно, что единичные аминокислотные замены в неэссенциальных участках, как правило, не приводят к существенному изменению биологической активности (см., например, Watson, et al., MOLECULAR BIOLOGY OF THE GENE, The Benjamin/Cummings Pub. Co., p. 224 (4 th Edition, 1987)). Предпочтительные примеры таких замен описаны в табл. III(A), III(B). Например, такие замены включают в себя замещение любого из изолейцина (I), валина (V) и лейцина (L) на любую другую из указанных гидрофобных аминокислот: аспарагиновой кислоты (D) на глутаминовую кислоту (Е), и наоборот; глутамина (Q) на аспарагин (N), и наоборот; и серина (S) на треонин (Т), и наоборот. Другие замены также могут считаться консервативными, если это позволяет их окружение и роль в трехмерной структуре белка. Например, глицин (G) и аланин (А) часто являются взаимозаменяемыми, как и аланин (А) и валин (V). Метионин (М), который является относительно гидрофобным, как правило, можно заменить на лейцин и изолейцин, а иногда на валин. Лизин (K) и аргинин (R) часто являются взаимозаменяемыми по положениям, для которых имеет значение заряд аминокислотного остатка, но не pK. Консервативными можно считать и другие замены, в частности, в окружающей области (см., например, табл. III(A), приведенную в данном описании; p. 13-15 "Biochemistry", 2 nd ed. Lubert Stryer ed. (Stanford University); Henikoff et al., PNAS, 1992. Vol. 89, 10915-10919; Lei et al., J. Bio. Chem. 19 May, 1995; 270(20): 11882-6). Другие допустимые замены можно идентифицировать эмпирически или по аналогии с известными консервативными заменами. Термин "цитотоксическое средство" относится к веществу, которое ингибирует или предотвращает экспрессионную активность клеток, функционирование клеток и/или вызывает деструкцию клеток. Подразумевается, что данный термин включает в себя радиоактивные изотопы, химиотерапевтические средства и токсины, такие как токсины с маленькими молекулами или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, в том числе их фрагменты и/или варианты. Примеры цитотоксических средств включают в себя, без ограничения, ауристатины, ауристатин е, ауромицины, маутанзиноиды, иттрий, висмут, рицин, А-цепь рицина, комбрестатин, дуокармицины, долостатины, доксорубицин, даунорубицин, таксол, цисплатин, сс1065, бромид этидия, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, дигидроксиантрациндион, актиномицин, дифтерийный токсин, экзотоксин Pseudomonas (РЕ) А, РЕ40, абрин, А-цепь абрина, А-цепь модексина, альфа-сарцин, гелонин, митогеллин, ретстриктоцин, феномицин, эномицин, курицин, кротин, каличемицин, ингибитор Sapaonaria officinalis, глюкокортикоиды и другие химиотерапевтические средства, а также радиоактивные изотопы, такие как 211 At, 131 I, 125 I, 90 Y, 186 Re, 188 Re, 153 Sm, 212 или 213 Bi, 32 p и радиоактивные изотопы Lu, в том числе 177 Lu. К антителам также можно присоединить фермент, активирующий пролекарственную форму противоракого средства, который способен превращать пролекарство в активную форму. В данном описании термин "диантитела" относится к маленьким фрагментам антител с двумя антигенсвязывающими участками, данные фрагменты содержат вариабельный домен тяжелой цепи (V H ), соединенный с вариабельным доменом легкой цепи (V L ) в одной полипептидной цепи (V H -V L ). Путем применения линкера, который является слишком коротким, чтобы позволить спариваться двум доменам на одной цепи, домены вынуждают спариваться с комплементарными доменами другой цепи с образованием двух антигенсвязывающих участков. Более подробно диантитела описаны, например, в ЕР 404097; WO 93/11161 и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-48 (1993). Термин "генный продукт" в данном описании обозначает пептид/белок или мРНК. Например, термин "генный продукт согласно изобретению" в данном описании иногда относится к "раковой аминокислотной последовательности", "раковому белку", "белку злокачественного заболевания", приведенного в табл. I, "раковой мРНК", "мРНК злокачественного заболевания, приведенного в табл. I" и др. В одном воплощении раковый белок кодируется нуклеиновой кислотой, приведенной на фиг. 2. Раковый белок может представлять собой фрагмент или, альтернативно, полноразмерный белок, кодируемый нуклеиновыми кислотами фиг. 2. В одном воплощении раковую аминокислотную последовательность используют для определения идентичности или подобия последовательностей. В другом воплощении последовательности представляют собой природные аллельные варианты белка, кодируемого нуклеиновой кислотой, приведенной на фиг. 2. В следующем воплощении последовательности представляют собой варианты последовательностей, как описано в данном документе ниже. В способах и композициях согласно изобретению можно использовать "гетероконъюгатные" антитела. В данном описании термин "гетероконъюгатное антитело" относится к двум ковалентно связанным антителам. Такие антитела можно получить с помощью известных способов белкового синтеза, в том числе путем использования сшивающих агентов. См., например, патент США № 4676980. Способы проведения "высокопроизводительного скрининга" с целью определения присутствия, отсутствия количества или других характеристик конкретных нуклеиновых кислот или белковых продуктов хорошо известны специалистам в данной области. Подобным образом, хорошо извстны способы проведения анализов связывания и анализов репортерных генов. Так, например, патент США № 5559410 раскрывает способы проведения высокопроизводительного скрининга белков; патент США № 5585639 раскрывает способы проведения высокопроизводительного скрининга связывания нуклеиновых кислот (т.е. в массивах); а патенты США № 5576220 и 5541061 раскрывают способы проведения высокопроизводительного скрининга на связывание лиганд/антитело. Кроме того, существуют коммерческие системы высокопроизводительного скрининга (см., например, Amersham Biosciences, Piscataway, NJ; Zymark Corp., Hopkinton, MA; Air Technical Industries, Mentor, OH; Beckman Instruments, Inc. Fullerton, CA; Precision Systems, Inc., Natick, MA; etc.). Как правило, в данных системах автоматизированы все процедуры, в том числе отмеривание пипетками всех образцов и реагентов, распределение жидкостей, инкубации в течение заданного времени и конечное считывание микропланшета в детекторе, используемом в данном анализе. Данные настраиваемые системы обеспечивают высокую пропускную способность, быстрый запуск и высокую степень приспособляемости и удовлетворения требований потребителя. Производители таких систем предоставляют подробные инструкции для разных систем с высокой пропускной способностью. Так, например, Zymark Corp. предлагает технические бюллетени, описывающие системы скрининга с детекцией модуляции транскрипции генов, связывания лиганда и т.п. Термин "гомолог" относится к молекуле, которая гомологична другой молекуле, т.е. такие молекулы имеют последовательности химических остатков, совпадающие или подобные по соответствующим положениям. В одном воплощении антитело, предлагаемое в данном описании, представляет собой "человеческое антитело". В данном описании термин "человеческое антитело" относится к антителу, в котором практически все последовательности легких и тяжелых цепей, включая гипервариабельные участки (CDRs), кодируются человеческими генами. В одном воплощении для получения человеческих моноклональных антител используют триомы, человеческие В-клетки (см., например, Kozbor, et al., Immunol. Today, 4: 72 (1983)), трансформацию EBV (см., например, Cole et al. MONOCLONAL ANTIBODIES AND CANCER THERAPY, 77-96 (1985)) или фаговые дисплеи (см., например, Marks et al., J. Mol. Biol. 222: 581 (1991)). В отдельном воплощении человеческое антитело получают в трансгенной мыши. Можно использовать любые известные в данной области способы получения таких частично или полностью человеческих антител. В соответствии с особо предпочтительным воплощением последовательности полностью человеческого антитела получают в трансгенной мыши, несущей гены тяжелой и легкой цепей человеческого антитела. Примеры способов получения трансгенных мышей и их потомства, продуцирующих человеческие антитела, можно найти в заявке № WO 02/43478 и патенте США № 6657103 (Abgenix). В-клетки трансгенных мышей, продуцирующие целевое антитело, затем можно гибридизовать с получением гибридомных клеточных линий, непрерывно продуцирующих антитело (см., например, патенты США № 5569825; 5625126; 5633425; 5661016 и 5545806; Jakobovits, Adv. Drug Del. Rev. 31: 33-42 (1998); Green, et al., J. Exp. Med. 188: 483-95 (1998)). "Человеческий лейкоцитарный антиген", или "HLA" представляет собой человеческий белок главного комплекса гистосовместимости (МНС) класса I или класса II (см., например, Stites, et al., IMMUNOLOGY, 8 th ED., Lange Publishing, Los Altos, CA (1994)). В данном описании термин "гуманизированное антитело" относится к формам антител, которые содержат последовательности не человеческих (например, мышиных) и человеческих антител. Такие антитела представляют собой химерные антитела, которые содержат минимальную последовательность не человеческого иммуноглобулина. В большинстве случаев гуманизированное антитело содержит, по существу, полностью по меньшей мере один, как правило, два вариабельных доменов, в которых все или почти все гипервариабельные петли относятся к не человеческому иммуноглобулину, а все или почти все участки FR представляют собой последовательности человеческого иммуноглобулина. Гуманизированное антитело также необязательно содержит по меньшей мере часть константного участка (Fc) иммуноглобулина, обычно человеческого иммуноглобулина. См., например, Cabilly патент США № 4816567; Queen et al. (1989), Proc. Nat'1 Acad. Sci. USA 86: 10029-10033 и ANTIBODY ENGINEERING: A PRACTICAL APPROACH (Oxford University Press, 1996). Подразумевается, что термины "гибридизовать", "гибридизация", "гибридизуется" и т.п., используемые в применении к полинуклеотидам, относятся к традиционным условиям гибридизации, предпочтительно таким, как гибридизация в 50% формамиде/6 × SSC/0,1% SDS/100 мкг/мл ss-ДHK, где температура гибридизации выше 37 °С, а температура промывания в 0,1 × SSC/0,1% SDS выше 55 °С. Фразы "выделенный" или "биологически чистый" относятся к веществу, которое по существу или практически не содержит компонентов, которые обычно окружают вещество в природном состоянии. Таким образом, выделенные пептиды в соответствии с данным изобретением предпочтительно не содержат веществ, обычно присутствующих в окружении пептидов in situ. Например, полинуклеотид считают "выделенным", если он практически не содержит примеси полинуклеотидов, соответствующих или комплементарных генам, отличным от генов STEAP-1, или кодирующих полипептиды, отличные от генного продукта STEAP-1 или его фрагментов. Опытный специалист может легко использовать способы выделения нуклеиновых кислот для получения выделенного полинуклеотида STEAP-1. Белок считают "выделенным", если, например, с помощью физических, механических или химических методов белок STEAP-1 отделяют от компонентов клеток, обычно связанных с белком. Опытный специалист может легко использовать стандартные методы очистки для получения выделенного белка STEAP-1. Альтернативно, выделенный белок можно получить химическими методами. Подходящие "метки" включают в себя радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные фрагменты, хемилюминесцентные фрагменты, магнитные частицы и т.п. Патенты, описывающие применение таких меток, включают в себя патенты США № 3817837; 3850752; 3939350; 3996345; 4277437; 4275149 и 4366241. Кроме того, антитела, предлагаемые в настоящем описании, можно использовать в качестве антигенсвязывающего компонента флуоресцентных антител. См., например, Zeytun et al., Nat. Biotechnol. 21: 1473-79 (2003). Термин "млекопитающее" относится к любому организму класса млекопитающих и включает в себя мышей, крыс, кроликов, собак, кошек, коров, лошадей и людей. В одном воплощении согласно изобретению млекопитающее представляет собой мышь. В другом воплощении согласно изобретению млекопитающее представляет собой человека. Термины "метастазирующий рак простаты" и "метастазирующее заболевание" относятся к раку простаты, распространившемуся в региональные лимфатические узлы или в отдаленные участки организма, и включает в себя заболевание стадии D по системе AUA и заболевание стадии T ×N ×M+ по системе TNM. Как и в случае с местно распространенным раком простаты, для пациентов с метастазирующим заболеванием хирургическая операция не рекомендуется, а предпочтительным способом лечения является гормональная (антиандрогенная) терапия. У пациентов с метастазирующим раком простаты с течением времени, через 12-18 месяцев после начала лечения, развивается андроген-устойчивое состояние. Приблизительно половина таких андроген-устойчивых пациентов умирает в течение 6 месяцев после развития данного состояния. Чаще всего рак простаты метастазирует в кости. Костные метастазы рака простаты чаще являются остеобластными, чем остеолитическими (т.е. приводят к образованию костей с сетчатой структурой). Костные метастазы чаще всего можно обнаружить в позвоночнике, затем в бедренной кости, тазовых костях, грудной клетке, черепе и плечевой кости. Другие распространенные участки метастазирования включают в себя лимфатические узлы, легкие, печень и мозг. Метастазирующий рак простаты обычно диагностируют путем открытия или лапароскопического иссечения тазовых лимфатичских узлов, радионуклидного анализа целой крови, радиографического исследования скелета и/или биопсии патологического изменения кости. Термины "модулятор", или "тестируемое соединение", или "лекарственное средство-кандидат", или используемые в данном описании грамматические эквиваленты относятся к любой молекуле, например белку, олигопептиду, маленькой органической молекуле, полисахариду, полинуклеотиду и т.п., тестируемым на способность непосредственно или косвенно изменять раковый фенотип, или экспрессию раковых последовательностей, например последовательностей нуклеиновых кислот или белков, или эффекты раковых последовательностей (например, сигнальные эффекты, экспрессию генов, взаимодействие белков и др.). В одном аспекте модулятор нейтрализует эффект ракового белка согласно изобретению. "Нейтрализация" означает ингибирование или блокировку активности белка, наряду с последующим эффектом на клетку. В другом аспекте модулятор нейтрализует эффект гена и соответствующего ему белка согласно изобретению путем нормализации уровней указанного белка. В предпочтительных воплощениях модуляторы изменяют профили экспрессии или профиль экспрессии нуклеиновых кислот или белков, предлагаемых в данном описании, или пути, лежащие ниже эффектора. В одном воплощении модулятор подавляет раковый фенотип, например, до нормальных характеристик ткани. В другом воплощении модулятор индуцирует раковый фенотип. Обычно несколько аналитических смесей анализируют в параллелях с разными концентрациями средства с получением разных ответов на разные концентрации. Как правило, одна из данных концентраций служит отрицательным контролем, т.е. нулевой концентрацией или концентрацией, вызывающей эффект ниже уровня детектирования. Модуляторы, лекарственные средства-кандидаты или тестируемые соединения охватывают разные классы химических соединений, однако обычно они представляют собой органические молекулы, предпочтительно маленькие органические соединения, имеющие молекулярную массу более 100 и менее чем приблизительно 2500 Да. Предпочтительные маленькие молекулы имеют молекулярную массу меньше 2000, меньше 1500 или меньше 1000 или 500 Да. Средства-кондидаты содержат функциональные группы, необходимые для структурного взаимодействия с белками, в особенности способные образовывать водородные связи, и обычно включают в себя, по меньшей мере, аминовую, карбонильную, гидроксильную или карбоксильную группу, предпочтительно, они содержат две функциональные химические группы. Средства-кандидаты часто содержат циклический атом углерода или гетероциклические структуры и/или ароматические или полиароматические структуры, замещенные одной или несколькими из вышеуказанных функциональных групп. Модуляторы также включают в себя такие биомолекулы, как пептиды, сахариды, жирные кислоты, стероиды, пурины, пиримидины, их производные, структурные аналоги или сочетания. Особенно предпочтительными молекулами являются пептиды. Одним классом модуляторов являются пептиды, например, содержащие приблизительно от 5 до 35 аминокислот, предпочтительно приблизительно от 5 до 20 аминокислот, особенно предпочтительно приблизительно от 7 до 15 аминокислот. Предпочтительно раковый модуляторный белок является растворимым, содержит участок, отличный от трансмембранного, и/или содержит N-концевой Cys, повышающий растворимость. В одном воплощении С-конец фрагмента держат в виде свободной кислоты, а N-конец - в виде свободного амина, чтобы обеспечить конденсацию, например, с цистеином. В одном воплощении раковый белок согласно изобретению конъюгируют с иммуногенным агентом, как описано в данном документе. В одном воплощении раковый белок конъюгируют с БСА. Пептиды согласно изобретению, например, имеющие предпочтительную длину, можно соединить друг с другом или с другой аминокислотой с получением более длинного пептида/белка. Модуляторные пептиды могут представлять собой продукты расщепления природных белков, как указано выше, рандомизированные пептиды или пептиды, рандомизированные "с отклонением". В предпочтительном воплощении пептидные/белковые модуляторы представляют собой антитела и их фрагменты, определенные в данном описании. Модуляторами злокачественного заболевания также могут быть нуклеиновые кислоты. Нуклеиновые модулирующие средства могут представлять собой природные нуклеиновые кислоты, рандомизированные нуклеиновые кислоты или нуклеиновые кислоты, рандомизированные "с отклонением". Например, в подходе, аналогичном описанному выше для белков, можно использовать продукты расщепления прокариотических или эукариотических геномов. В данном описании термин "моноклональное антитело" относится к антителу, полученному из популяции, по существу, гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных природных мутаций, которые могут присутствовать в минорных количествах. Моноклональные антитела являются высокоспецифичными по отношению к одному антигенному эпитопу. Наоборот, препараты традиционных (поликлональных) антител, как правило, включают в себя множество антител, направленных против разных эпитопов (или специфичных по отношению к разным эпитопам). В одном воплощении поликлональные антитела включают в себя совокупность моноклональных антител, обладающих специфичностью, аффинностью или авидностью по отношению к разным эпитопам в одном антигене, который содержит несколько антигенных эпитопов. Определение "моноклональный" указывает на характер антитела, полученного из практически гомогенной популяции антител, и не подразумевает получение антитела каким-либо конкретным способом. Например, моноклональные антитела, используемые в соответствии с настоящим изобретением, могут быть получены гибридомным методом, впервые описанным Kohler et al., Nature, 256: 495 (1975), или методами рекомбинантных ДНК (см., например, патент США № 4816567). "Моноклональные антитела" также могут быть выделены из фаговых библиотек антител с помощью методов, описанных, например, в Clackson et al., Nature, 352: 624-628 (1991) и Marks et al., J. Mol. Biol. 222: 581-597 (1991). K d связывания указанных моноклональных антител составляет по меньшей мере приблизительно 1 мкМ, чаще по меньшей мере приблизительно 300 нМ, как правило, по меньшей мере приблизительно 30 нМ, предпочтительно по меньшей мере приблизительно 10 нМ, более предпочтительно по меньшей мере приблизительно 3 нМ или менее и обычно определяется методом ELISA. Термин "мотив", например биологический мотив белка, родственного STEAP-1, относится к любой образованной аминокислотами части первичной последовательности белка, которая связана с конкретной функцией (например, белок-белковым взаимодействием, взаимодействием белок-ДНК и др.), или модификацией (например, фосфорилированная, гликозилированная или амидированная часть), или локализацией (например, секреторная последовательность, последовательность ядерной локализации и др.), или к последовательности, связанной с иммунными функциями, либо гуморальными, либо клеточными. Мотив может быть либо смежным с некоторыми положениями, коррелирующими с определенной функцией или определенным свойством, либо совпадать с такими положениями. В контексте мотивов HLA данный термин относится к совокупности остатков в пептиде определенной длины, как правило, содержащем приблизительно от 8 до 13 аминокислот в случае мотива HLA класса I и приблизительно от 6 до 25 аминокислот в случае мотива HLA класса II, которая распознается конкретной молекулой HLA. Пептидные мотивы для связывания HLA обычно различаются в белках, кодируемых разными аллелями человеческого HLA, и различаются по совокупности первичных и вторичных анкорных остатков. Термин "фармацевтический наполнитель" включает в себя такие вещества, как вспомогательные средства, носитель, средства, регулирующие рН, и забуферивающие средства, средства, регулирующие тоничность, увлажняющие средства, консерванты и т.п. Термин "фармацевтически приемлемый" относится к нетоксичной и/или инертной композиции, физиологически совместимой с людьми или другими млекопитающими. Термин "полинуклеотид" относится к полимерной форме нуклеотидов длиной по меньшей мере в 10 оснований или пар оснований, либо рибонуклеотидов, либо дезоксинуклеотидов, либо модифицированных нуклеотидов любого типа, подразумевается, что данный термин включает в себя одно- или двухцепочечные формы ДНК и/или РНК. В данной области указанный термин часто используется как взаимозаменяемый с термином "олигонуклеотид". Полинуклеотид может содержать нуклеотидную последовательность, раскрытую в данном описании, где тимидин (Т), как показано, например, на фиг. 2, может быть заменен на урацил (U); данное определение связано с различиями в химических структурах ДНК и РНК, в особенности относится к тому, что одним из четырех доминирующих оснований в РНК является урацил (U), вместо которого в ДНК присутствует тимидин (Т). Термин "полипептид" относится к полимеру, содержащему по меньшей мере приблизительно 4, 5, 6, 7 или 8 аминокислот. В данном описании для обозначения аминокислот используют стандартные трехбуквенные или однобуквенные символы. В данной области указанный термин часто используется как взаимозаменяемый с терминами "пептид" или "белок". "Первичный анкорный остаток" HLA представляет собой аминокислоту в конкретном положении пептидной последовательности, которая, как полагают, участвует в образовании точки контакта между иммуногенным пептидом и молекулой HLA. От одного до трех, как правило два, первичных анкорных остатковв пептиде определенной длины обычно образуют "мотив" иммуногенного пептида. Полагают, что данные остатки находятся в тесном контакте с пептидсвязывающей бороздкой молекулы HLA, причем их боковые цепи погружены в специфические карманы связывающей бороздки. В одном воплощении, например, первичные анкорные остатки молекулы HLA класса I расположены в положении 2 (с аминоконца), а с карбоксильной стороны - в положениях 8, 9, 10, 11 или 12 пептидного эпитопа в соответствии с данным изобретением. Альтернативно, в другом воплощении первичные анкорные отатки пептида, связывающие молекулу HLA класса II, отдалены в пространстве друг от друга в большей степени, чем от концов пептида, где длина пептида обычно составляет по меньшей мере 9 аминокислот. Первичные анкорные положения для каждого мотива и супермотива приведены в табл. IV(A). Например, аналоговые пептиды можно создать путем изменения присутствия или отсутствия конкретных остатков в первичных и/или вторичных анкорных положениях, приведенных в табл. IV. Такие аналоги используют для модуляции связывающего сродства и/или установления популяционного распределения пептида, содержащего конкретный мотив или супермотив HLA. "Радиоактивные изотопы" включают в себя, без ограничения, нижеследующие неограничивающие примеры применения, которые приведены в табл. IV(I). Под термином "рандомизированный" или его грамматическими эквивалентами в данном описании понимают нуклеиновые кислоты и белки, состоящие, по существу, из случайных нуклеотидов и аминокислот соответственно. Данные рандомизированные пептиды (или нуклеиновые кислоты, описанные в данном документе) в каждом положении могут содержать любой нуклеотид или любую аминокислоту. Можно разработать способ синтеза рандомизированных белков или нуклеиновых кислот, содержащих все или почти все возможные сочетания по всей длине последовательности, с получением библиотеки рандомизированных биологически активных белковых средств-кандидатов. В одном воплощении библиотека является "полностью рандомизированной" и не имеет последовательностей отнесения или постоянных элементов в каком-либо положении. В другом воплощении библиотека представляет собой библиотеку, "рандомизированную с отклонением". То есть некоторые положения в последовательности либо оставляют постоянными, либо выбирают из ограниченного числа вариантов. Например, нуклеотидные или аминокислотные остатки рандомизируют в пределах определенного класса, например гидрофобных аминокислот, гидрофильных остатков, стерически отклоняющихся (либо маленьких, либо больших) остатков, для создания доменов, связывающих нуклеиновые кислоты, для введения остатков цистеина, для образования поперечных связей, для введения остатков пролина в домены SH-3, остатков серина, треонина, тирозина или гистидина в участки фосфорилирования и др., или в пределах класса пуринов и др. "Рекомбинантная" молекула ДНК или РНК представляет собой молекулу ДНК или РНК, полученную в результате молекулярных манипуляций in vitro. В данном описании термин "одноцепочечный Fv" или "scFv", или "одноцепочечное" антитело относится к фрагментам антитела, содержащим домены антитела V H и V L , где указанные домены находятся на одной полипептидной цепи. Обычно полипептид Fv содержит также полипептидный линкер между доменами V H и V L , который позволяет scFv образовывать структуру, необходимую для связывания антигена. Обзор по scFv см. в Pluckthun, THE PHARMACOLOGY OF MONOCLONAL ANTIBODIES, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, p. 269-315 (1994). Неограничивающие примеры "маленьких молекул" включают в себя соединения, которые связывают STEAP-1 или взаимодействуют с ним, лиганды, в том числе гормоны, нейропептиды, хемокины, отдушки, фосфолипиды и их функциональные эквиваленты, которые связывают и предпочтительно ингибируют белковую функцию STEAP-1. Такие маленькие молекулы, без ограничения, предпочтительно имеют молекулярную массу менее чем приблизительно 10 кДа, более предпочтительно менее чем приблизительно 9, приблизительно 8, приблизительно 7, приблизительно 6, приблизительно 5 или приблизительно 4 кДа. В некоторых воплощениях маленькие молекулы физически связаны с белком STEAP-1 или связывают данный белок; не обнаружены в природных метаболических путях и/или лучше растворяются в водных, чем в неводных растворах. В данном описании термин "специфический" относится к селективному связыванию антитела с целевым антигенным эпитопом. Специфичность связывания антител можно анализировать путем сравнения связывания с целевым антигеном со связыванием с другим антигеном или смесью антигенов в заданной комбинации условий. Если антитело связывается с целевым антигеном по меньшей мере в 2, 5, 7 и предпочтительно в 10 раз лучше, чем с другим антигеном или смесью антигенов, то оно считается специфическим. В одном воплощении специфическое антитело представляет собой антитело, которое связывает только антиген STEAP-1, но не связывает другие антигены. В другом воплощении специфическое антитело представляет собой антитело, которое связывает человеческий антиген STEAP-1, но не связывает антиген STEAP-1, отличный от человеческого и обладающий 70, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99% или более аминокислотной гомологией с антигеном STEAP-1. В другом воплощении специфическое антитело представляет собой антитело, которое связывает человеческий антиген STEAP-1 и связывает мышиный антиген STEAP-1, но в большей степени оно связывает человеческий антиген. В другом воплощении специфическое антитело представляет собой антитело, которое связывает человеческий антиген STEAP-1 и антиген STEAP-1 приматов, но в большей степени оно связывает человеческий антиген. В следующем воплощении специфическое антитело связывает человеческий антиген STEAP-1 и любой отличный от человеческого антиген STEAP-1 или любое их сочетание, но в большей степени оно связывает человеческий антиген. "Жесткость" гибридизации легко определяется рядовым специалистом в данной области и, как правило, эмпирически рассчитывается в зависимости от длины зонда, температуры промывания и концентрации соли. Как правило, для соответствующего отжига более длинных зондов требуется более высокая температура, тогда как более короткие зонды требуют более низких температур. Как правило, гибридизация зависит от способности денатурированных последовательностей нуклеиновых кислот к повторному отжигу, если комплементарные цепи присутствуют в среде при температуре ниже точки плавления. Чем выше степень желаемой гомологии между зондом и гибридизующейся с ним последовательностью, тем выше относительная температура, которую можно использовать. Следовательно, более высокая относительная температура может делать условия реакции более жесткими, а более низкая температура - менее жесткими. Дополнительные подробности и разъяснения, касающиеся жесткости реакций гибридизации, можно найти в Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995). "Условия жесткости" или "условия высокой жесткости", как описано в данном документе, включают в себя, без ограничения, следующие: (1) применение низкой ионной силы и высокой температуры для промывания, например, 0,015 М хлорид натрия/0,0015 М цитрат натрия/0,1% додецилсульфат натрия при 50 °С; (2) применение в процессе гибридизации денатурирующего средства, такого как формамид, например 50% (об./об.) формамид и 0,1% бычий сывороточный альбумин/0,1% фиколл/0,1% поливинилпирролидон/50 мМ натрий-фосфатный буфер, рН 6,5, содержащий 750 мМ хлорид натрия, 75 мМ цитрат натрия, при 42 °С или (3) применение 50% формамида, 5 × SSC (0,75 М NaCl, 0,075 М цитрат натрия), 50 мМ фосфата натрия (рН 6,8), 0,1% пирофосфата натрия, 5 × раствора Денхардта, ДНК спермы лосося, обработанной ультразвуком (50 мкг/мл), 0,1% SDS и 10% сульфата декстрана при 42 °С с промыванием при 42 °С в 0,2 × SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55 °С и затем с промыванием в условиях высокой жесткости, включающих в себя применение 0,1 × SSC, содержащей EDTA, при 55 °С. "Условия средней жесткости" описаны, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989 и включают в себя применение раствора для промывания и условий гибридизации (например, температуры, ионной силы и % SDS), менее жестких, чем описанные выше. Примером условий средней жесткости является инкубация в течение ночи при 65 °С в растворе, содержащем 1% бычий сывороточный альбумин, 0,5 М фосфат натрия, рН 7,5, 1,25 мМ EDTA и 7% SDS 5 × SSC (150 мМ NaCl, 15 мМ трицитрат натрия), с последующим промыванием фильтров в 2 × SSC/1% SDS при 50 °С и 0,2 × SSC/0,1% SDS при 50 °С. Специалисту в данной области известно, как выбрать температуру, ионную силу и др. в зависимости от таких факторов, как длина зонда и т.п. "Супермотив" HLA представляет собой пептидсвязывающий специфичный участок, общий для молекул HLA, кодируемых двумя или более аллелями HLA. Общая частота фенотипической встречаемости HLA в разных этнических популяциях приведена в табл. IV(F). Разные супертипы включают в себя, без ограничения, следующие разновидности: Рассчитанное популяционное распределение разных сочетаний супертипов HLA приведено в табл. IV(G). В данном описании термины "лечить" или "терапевтический", а также грамматически родственные им термины относятся к какому-либо улучшению любого последствия заболевания, такому как повышение выживаемости, уменьшение процента смертности и/или уменьшение побочных эффектов, свойственных альтернативным терапевтическим средствам; специалистам в данной области известно, что полное излечение заболевания является предпочтительным, но не обязательным требованием для акта лечения. "Трансгенное животное" (например, мышь или крыса) представляет собой животное, несущее клетки, содержащие трансген, который вводят животному или его предку в пренатальном периоде, например на эмбриональной стадии. "Трансген" представляет собой ДНК, интегрированную в геном клетки, из которой развивается трансгенное животное. В данном описании HLA или "вакцина" клеточного иммунного ответа представляет собой композицию, которая содержит или кодирует один или несколько пептидов согласно изобретению. Существует несколько воплощений таких вакцин, например смесь одного или нескольких отдельных пептидов; один или несколько пептидов согласно изобретению, содержащих полиэпитопный пептид; или нуклеиновые кислоты, которые кодируют такие отдельные пептиды или полипептиды, например мини-ген, который кодирует полиэпитопный пептид. Термин "один или несколько пептидов" может включать в себя любое целое число от 1 до 150 или больше, например по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145 или 150, пептидов согласно изобретению. Пептиды или полипептиды, необязательно, можно модифицировать, например, путем липидизации, добавления меток или других последовательностей. Пептиды HLA класса I согласно изобретению можно смешать или связать с пептидами HLA класса II, чтобы обеспечить активацию как цитотоксических, так и хелперных Т-лимфоцитов. HLA вакцины также могут содержать сенсибилизированные пептидом клетки, экспонирующие антиген, например дендритные клетки. Термин "вариант" относится к молекуле, которая отличается от описываемого или нормального типа, такой как белок, который содержит один или несколько других аминокислотных остатков в соответствующем положении (соответствующих положениях) конкретно описываемого белка (например, белка STEAP-1, приведенного на фиг. 2 или 3). Аналог является примером вариантного белка. Другими примерами вариантов являются изоформы, образующиеся в результате сплайсинга и полиморфизма отдельных нуклеотидов (SNP). "STEAP-1-родственные белки" согласно изобретению включают в себя белки, конкретно описанные в данном документе, а также их аллельные варианты, варианты, содержащие консервативные замены, аналоги и гомологи, которые могут быть выделены/получены и охарактеризованы без проведения излишних экспериментов с помощью способов, описанных в данном документе или хорошо известных в данной области. В объем изобретения также входят гибридные белки, в которых объединены части разных белков или их фрагментов, а также гибридные белки, состоящие из белка STEAP-1 и гетерологичного полипептида. Такие белки STEAP-1 в совокупности называют белками, родственными STEAP-1, белками согласно изобретению или STEAP-1. Термин "белок, родственный STEAP-1", относится к полипептидному фрагменту или белковой последовательности STEAP-1, содержащей 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более аминокислот или по меньшей мере 30, 35, 40, 45, 50, 55, 60, 65, 70, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145, 150, 155, 160, 165, 170, 175, 180, 185, 190, 195, 200, 225, 250, 275, 300, 325, 330, 335, 339 или более аминокислот. II) Полинуклеотиды STEAP-1. В одном аспекте данное изобретение предлагает полинуклеотиды, соответствующие или комплементарные полноразмерным гену, мРНК и/или кодирующей последовательности STEAP-1 или их части, предпочтительно в выделенном виде, в том числе полинуклеотиды, кодирующие белок, родственный STEAP-1, и его фрагменты, ДНК, РНК, гибрид ДНК/РНК и родственные молекулы, полинуклеотиды или олигонуклеотиды, комплементарные гену STEAP-1 или последовательности мРНК или ее части, и полинуклеотиды или олигонуклеотиды, гибридизующиеся с геном STEAP-1, мРНК или с полинуклеотидом, кодирующим STEAP-1 (вместе их называют "полинуклеотиды STEAP-1"). Во всех случаях упоминаемый в данном разделе Т может быть заменен на U на фиг. 2. Воплощения полинуклеотида STEAP-1 включают в себя полинуклеотид STEAP-1, имеющий последовательность, изображенную на фиг. 2, причем в нуклеотидной последовательности STEAP-1, изображенной на фиг. 2, Т может обозначать U; по меньшей мере 10 смежных нуклеотидов полинуклеотида, имеющего последовательность, изображенную на фиг. 2; или по меньшей мере 10 смежных нуклеотидов полинуклеотида, имеющего последовательность, изображенную на фиг. 2, где Т может обозначать U. Например, воплощения нуклеотидов STEAP-1 включают в себя, без ограничения: (I) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2, где Т также может обозначать U; (II) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2А, от нуклеотидного остатка номер 66 до нуклеотидного остатка номер 1085, включая стоп-кодон, где Т также может обозначать U; (III) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2В, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (IV) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2С, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 944, включая стоп-кодон, где Т также может обозначать U; (V) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2D, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (VI) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2Е, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (VII) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2F, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (VIII) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2G, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (IX) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2Н, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (X) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2I, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XI) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2J, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XII) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2K, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XIII) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2L, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XIV) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2М, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XV) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2N, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XVI) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2O, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XVII) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2Р, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XVIII) полинуклеотид, по существу содержащий или содержащий последовательность, изображенную на фиг. 2Q, от нуклеотидного остатка номер 96 до нуклеотидного остатка номер 872, включая стоп-кодон, где Т также может обозначать U; (XIX) полинуклеотид, кодирующий белок, родственный STEAP-1, который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% гомологичен полноразмерной аминокислотной последовательности, изображенной на фиг. 2A-2Q; (XX) полинуклеотид, кодирующий белок, родственный STEAP-1, который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен полноразмерной аминокислотной последовательности, изображенной на фиг. 2A-2Q; (XXI) полинуклеотид, кодирующий по меньшей мере один из пептидов, приведенных в табл. V-XVIII и XXII-LI, как описано в патентной заявке США 10/236878, поданной 6 сентября 2002 г., конкретное содержание которой полностью включено в данное описание в качестве ссылки; (XXII) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3А с любым целочисленным приращением вплоть до 339, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю гидрофильности, приведенному на фиг. 5; (XXIII) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3А с любым целочисленным приращением вплоть до 339, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением меньше 0,5 по профилю гидрофобности, приведенному на фиг. 6; (XXIV) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3А с любым целочисленным приращением вплоть до 339, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю процента доступных остатков, приведенному на фиг. 7; (XXV) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3А с любым целочисленным приращением вплоть до 399, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю средней жесткости, приведенному на фиг. 8; (XXVI) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3А с любым целочисленным приращением вплоть до 339, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю бета-складки, приведенному на фиг. 9; (XXVII) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3В и 3D с любым целочисленным приращением вплоть до 258, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю гидрофильности, приведенному на фиг. 5; (XXVIII) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3В и 3D с любым целочисленным приращением вплоть до 258, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением меньше 0,5 по профилю гидрофобности, приведенному на фиг. 6; (XXIX) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3В и 3D с любым целочисленным приращением вплоть до 258, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю процента доступных остатков, приведенному на фиг. 7; (XXX) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3В и 3D с любым целочисленным приращением вплоть до 258, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю средней жесткости, приведенному на фиг. 8; (XXXI) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3В и 3D с любым целочисленным приращением вплоть до 258, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю бета-складки, приведенному на фиг. 9; (XXXII) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3С с любым целочисленным приращением вплоть до 282, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю гидрофильности, приведенному на фиг. 5; (XXXIII) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3С с любым целочисленным приращением вплоть до 282, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением меньше 0,5 по профилю гидрофобности, приведенному на фиг. 6; (XXXIV) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3С с любым целочисленным приращением вплоть до 282, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю процента доступных остатков, приведенному на фиг. 7; (XXXV) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3С с любым целочисленным приращением вплоть до 282, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю средней жесткости, приведенному на фиг. 8; (XXXVI) полинуклеотид, кодирующий пептидный участок, состоящий по меньшей мере из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот пептида фиг. 3С с любым целочисленным приращением вплоть до 282, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю бета-складки, приведенному на фиг. 9; (XXXVII) полинуклеотид, полностью комплементарный полинуклеотиду по любому из пп.(I)-(XXXVI); (XXXVIII) полинуклеотид, полностью комплементарный полинуклеотиду по любому из пп.(I)-(XXXVII); (XXXIX) пептид, который кодируется полинуклеотидом по любому из пп.(I)-(XXXVIII); и (XL) композиция, содержащая полинуклеотид по любому из пп.(I)-(XXXVIII) или пептид по п.(XXXIX) вместе с фармацевтическим наполнителем и/или в составе человеческой стандартной лекарственной формы; (XLI) способ применения полинуклеотида по любому из пп.(I)-(XXXVIII), или пептида по п.(XXXIX), или композиции по п.(XL) для модуляции экспрессии STEAP-1 в клетке; (XLII) способ применения полинуклеотида по любому из пп.(I)-(XXXVIII), или пептида по п.(XXXIX), или композиции по п.(XL) для диагностики, профилактики, прогнозирования или лечения субъекта, несущего клетку, экспрессирующую STEAP-1; (XLIII) способ применения полинуклеотида по любому из пп.(I)-(XXXVIII), или пептида по п.(XXXIX), или композиции по п.(XL) для диагностики, профилактики, прогнозирования или лечения субъекта, несущего клетку, экспрессирующую STEAP-1, где указанная клетка относится к раковой ткани, приведенной в табл. I; (XLIV) способ применения полинуклеотида по любому из пп.(I)-(XXXVIII), или пептида по п.(XXXIX), или композиции по п.(XL) для диагностики, профилактики, прогнозирования или лечения рака; (XLV) способ применения полинуклеотида по любому из пп.(I)-(XXXVIII), или пептида по п.(XXXIX), или композиции по п.(XL) для диагностики, профилактики, прогнозирования или лечения рака ткани, приведенной в табл. I; и (XLVI) способ применения полинуклеотида по любому из пп.(I)-(XXXVIII), или пептида по п.(XXXIX), или композиции по п.(XL) для идентификации или характеристики модулятора экспрессии STEAP-1 в клетке. В данном описании подразумевается, что интервал конкретно включает в себя все находящиеся внутри него целочисленные значения. Типичные воплощения согласно изобретению, раскрытые в данном описании, включают в себя полинуклеотиды STEAP-1, которые кодируют конкретные фрагменты последовательностей мРНК STEAP-1 (а также полинуклеотиды, комплементарные таким последовательностям), такие как полинуклеотиды, кодирующие, например, нижеследующие белки и/или их фрагменты: 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145, 150, 155, 160, 165, 170, 175, 180, 185, 190, 195, 200, 225, 250, 275, 300, 325, 330, 335, 339 или больше смежных аминокислот STEAP-1, вариант 1; другие варианты имеют следующую максимальную длину: вариант 2 - 258 аминокислот; вариант 3 - 282 аминокислот и вариант 4 - 258 аминокислот. Например, типичные воплощения согласно изобретению, раскрытые в данном описании, включают в себя полинуклеотиды и кодируемые ими пептиды, полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 1 до аминокислоты 10 белка STEAP-1, изображенного на фиг. 2 или 3; полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 10 до аминокислоты 20 белка STEAP-1, изображенного на фиг. 2 или 3; полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 20 до аминокислоты 30 белка STEAP-1, изображенного на фиг. 2 или 3, полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 30 до аминокислоты 40 белка STEAP-1, изображенного на фиг. 2 или 3; полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 40 до аминокислоты 50 белка STEAP-1, изображенного на фиг. 2 или 3; полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 50 до аминокислоты 60 белка STEAP-1, изображенного на фиг. 2 или 3; полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 60 до аминокислоты 70 белка STEAP-1, изображенного на фиг. 2 или 3; полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 70 до аминокислоты 80 белка STEAP-1, изображенного на фиг. 2 или 3; полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 80 до аминокислоты 90 белка STEAP-1, изображенного на фиг. 2 или 3; полинуклеотиды, кодирующие фрагменты приблизительно от аминокислоты 90 до аминокислоты 100 белка STEAP-1, изображенного на фиг. 2 или 3, с приращением приблизительно 10 аминокислот, заканчивая аминокислотой карбоксильного конца, изображенной на фиг. 2 или 3. Соответственно полинуклеотиды, кодирующие фрагменты аминокислотной последовательности (приблизительно из 10 аминокислот), включающие в себя аминокислоты белка STEAP-1 от 100 до С-концевой аминокислоты, являются воплощениями согласно изобретению, где подразумевается, что каждое конкретное аминокислотное положение включает в себя указанное положение плюс или минус пять аминокислотных остатков. Полинуклеотиды, кодирующие относительно длинные фрагменты белка STEAP-1, также входят в объем согласно изобретению. Например, полинуклеотиды, кодирующие фрагмент приблизительно от аминокислоты 1 (или 20, или 30, или 40 и т.д.) до аминокислоты 20 (или 30, или 40, или 50 и т.д.) белка или "варианта" STEAP-1, изображенного на фиг. 2 или 3, можно получить с помощью ряда методов, хорошо известных в данной области. Данные полинуклеотидные фрагменты могут содержать любой фрагмент последовательности STEAP-1, изображенной на фиг. 2. Другие иллюстративные воплощения согласно изобретению, раскрытые в данном описании, включают в себя фрагменты полинуклеотида STEAP-1, кодирующие один или несколько биологических мотивов, входящих в состав последовательности белка или варианта STEAP-1, включающей в себя одну или несколько из мотив-несущих субпоследовательностей белка или варианта STEAP-1, приведенных в табл. V-XVIII и XXII-LI. В другом воплощении типичные полинуклеотидные фрагменты согласно изобретению кодируют один или несколько участков белка или варианта STEAP-1, гомологичных известной молекуле. В другом воплощении согласно изобретению типичные полинуклеотидные фрагменты могут кодировать один или несколько участков N-гликозилирования белка или варианта STEAP-1, участков фосфорилирования под действием цАМФ- и цГМФ-зависимой протеинкиназы, участков фосфорилирования под действием казеинкиназы II или участков N-миристоилирования и амидирования. Чтобы определить исходное положение любого пептида, приведенного в табл. V-XVIII и XXII-LI (собирательные таблицы пептидов HLA), относительно родительского белка, например варианта 1, варианта 2 и др., следует учитывать три фактора: конкретный вариант, длину пептида, приведенную в таблице пептидов HLA, и поисковые пептиды, приведенные в табл. LII. Как правило, поисковые пептиды используют для получения пептидов HLA конкретного варианта. Положение каждого поискового пептида относительно соответствующей родительской молекулы приведено в табл. LII. Соответственно, если поисковый пептид начинается в положении "X", нужно добавить значение "X минус 1" к каждому положению в табл. V-XVIII и XXII-LI, чтобы определить действительное положение пептидов HLA в родительской молекуле. Например, если конкретный поисковый пептид начинается в положении 150 родительской молекулы, нужно добавить 150-1, т.е. 149, к каждому аминокислотному положению пептида HLA, чтобы рассчитать положение аминокислоты в родительской молекуле. II.А. Применение полинуклеотидов STEAP-1. II.A.1. Выявление генетических нарушений. Полинуклеотиды, описанные в предыдущих разделах, имеют ряд конкретных применений. Хромосомное расположение человеческого гена STEAP-1 описано в примере, озаглавленном "Хромосомное картирование STEAP-1". Например, поскольку ген STEAP-1 находится в определенной хромосоме, полинуклеотиды, кодирующие разные участки белков STEAP-1, используют для характеристики цитогенетических нарушений в данном хромосомном регионе, например нарушений, которые, как установлено, связаны с различными раковыми заболеваниями. Было обнаружено, что ряд хромосомных нарушений в некоторых генах, включающих в себя реаранжировки, представляют собой цитогенетические нарушения, часто наблюдающиеся при разных злокачественных заболеваниях (см., например, Krajinovic et al., Mutat. Res. 382(3-4): 81-83 (1998); Johansson et al., Blood, 86(10): 3905-3914 (1995) и Finger et al., P.N.A.S. 85(23): 9158-9162 (1988)). Так, полинуклеотиды, кодирующие конкретные участки белков STEAP-1, можно использовать для более точного, чем это было возможно ранее, выявления цитогенетических нарушений в хромосомном участке, кодирующем STEAP-1, которые могут вносить вклад в формирование злокачественного фенотипа. В данном контексте указанные полинуклеотиды удовлетворяют существующую в данной области потребность в увеличении чувствительности скрининга хромосом, которая позволила бы идентифицировать более трудноуловимые и менее распространенные хромосомные нарушения (см., например, Evans et al., Am. J. Obstet. Gynecol. 171(4): 1055-1057 (1994)). Далее, поскольку было обнаружено, что STEAP-1 интенсивно экспрессируется при раке простаты и других злокачественных заболеваниях, полинуклеотиды STEAP-1 можно использовать для определения статуса генных продуктов STEAP-1 в нормальной ткани по сравнению с раковой. Как правило, полинуклеотиды, кодирующие конкретные участки белков STEAP-1, используют для оценки изменений (таких как делеции, вставки, точечные мутации или изменения, приводящие к утрате антигена, и др.) в конкретных участках гена STEAP-1, таких как участки, содержащие один или несколько мотивов. Примеры анализов включают в себя анализы ОТ-ПЦР и анализы полиморфизма одноцепочечной конформации (SSCP) (см., например, Marrogi et al., J. Cutan. Pathol. 26(8): 369-378 (1999)), в которых используются полинуклеотиды, кодирующие конкретные участки белка, для идентификации данных участков в белке. II.А.2. Антисмысловые воплощения. Другие конкретные воплощения нуклеиновых кислот согласно изобретению, раскрытые в данном описании, представляют собой геномные ДНК, кДНК, рибозимы и антисмысловые молекулы, а также молекулы нуклеиновых кислот с альтернативным скелетом или содержащие альтернативные основания либо полученные из природных источников, либо синтезированные и включают в себя молекулы, способные ингибировать экспрессию РНК или белка STEAP-1. Например, антисмысловые молекулы могут представлять собой РНК или другие молекулы, в том числе пептидные нуклеиновые кислоты (PNAs) или молекулы, отличные от нуклеиновых кислот, такие как фосфотиоатные производные, которые специфически связываются с ДНК или РНК по принципу комплементарности оснований. Специалист в данной области может легко получить указанные классы молекул нуклеиновых кислот с использованием полинуклеотидов STEAP-1 и полинуклеотидных последовательностей, описанных в данном документе. Антисмысловая технология включает в себя введение экзогенных олигонуклеотидов, которые связываются с целевым полинуклеотидом, расположенным в клетках. Термин "антисмысловой" относится к олигонуклеотидам, комплементарным их внутриклеточным мишеням, например STEAP-1. См., например, Jack Cohen, Oligodeoxynucleotides, Antisense Inhibitors of Gene Expression, CRC Press, 1989 и Synthesis, 1: 1-5 (1988). Антисмысловые олигонуклеотиды STEAP-1 согласно изобретению включают в себя такие производные, как S-олигонуклеотиды (фосфоротиоатные производные или S-олиги, см., Jack Cohen, приведено выше), которые ингибируют рост раковых клеток с повышенной эффективностью. S-олигонуклеотиды (нуклеозидфосфоротиоаты) являются изоэлектронными аналогами олигонуклеотидов (О-олигонуклеотид), в которых не мостиковый атом кислорода фосфатной группы заменен на атом серы. S-олигонуклеотиды согласно изобретению можно получить путем обработки соответствующих О-олигонуклеотидов 3Н-1,2-бензодитиол-3-он-1,1-диоксидом, который представляет собой реагент, переносящий серу. См., например, Iyer, R.P. et al., J. Org. Chem. 55: 4693-4698 (1990) и Iyer, R.P. et al., J. Am. Chem. Soc. 112: 1253-1254 (1990). Другие антисмысловые олигонуклеотиды STEAP-1 согласно изобретению включают в себя известные в данной области антисмысловые морфолиноолигонуклеотиды (см., например, Partridge et al., 1996, Antisense & Nucleic Acid Drug Development, 6: 169-175). Антисмысловые олигонуклеотиды STEAP-1 согласно изобретению, как правило, представляют собой РНК или ДНК, комплементарные первым 100 5'-кодонам, или последним 100 3'-кодонам геномной последовательности STEAP-1, или соответствующей мРНК, или стабильно гибридизующиеся с ними. Абсолютная комплементарность не требуется, хотя высокая степень комплементарности является предпочтительной. Применение олигонуклеотида, комплементарного к данному участку, обеспечивает селективную гибридизацию с мРНК STEAP-1, но не с мРНК, кодирующей другие регуляторные субъединицы протеинкиназы. В одном воплощении антисмысловые олигонуклеотиды STEAP-1 согласно изобретению представляют собой фрагменты антисмысловой молекулы ДНК, содержащие от 15 до 30 мономеров, которые имеют последовательности, гибридизующиеся с мРНК STEAP-1. Необязательно, антисмысловой олигонуклеотид STEAP-1 содержит 30 мономеров и является комплементарным участку, включающему в себя первые 10 5'-кодонов или последние 10 3'-кодонов STEAP-1. Альтернативно, антисмысловые молекулы модифицируют, чтобы использовать рибозимы для ингибирования экспрессии STEAP-1, см., например, L.A. Couture & D.Т. Stinchcomb; Trends Genet. 12: 510-515 (1996). II.A.3. Праймеры и пары праймеров. Другие конкретные воплощения нуклеотидов согласно изобретению включают в себя праймеры и пары праймеров, обеспечивающие специфическую амплификацию полинуклеотидов согласно изобретению или любых конкретных фрагментов указанных полинуклеотидов, а также зонды, которые селективно или специфично гибридизуются с молекулами нуклеиновых кислот согласно изобретению или с любыми их фрагментами. Зонды можно пометить детектируемым маркером, таким как, например, радиоактивный изотоп, флуоресцентное соединение, биолюминесцентное соединение, хемилюминесцентное соединение, соединение, способное образовывать комплексы с металлами, или фермент. Такие зонды и праймеры используют для детектирования присутствия полинуклеотида STEAP-1 в образце и в качестве средств, позволяющих детектировать клетки, экспрессирующие белок STEAP-1. Примеры таких зондов включают в себя полинуклеотиды, содержащие полноразмерную последовательность кДНК человеческого STEAP-1, изображенную на фиг. 2, или ее часть. Примеры пар праймеров, способных обеспечить специфическую амплификацию мРНК STEAP-1, также описаны в примерах. Для опытного специалиста очевидно, что на основе последовательностей, описанных в данном документе, можно получить большое количество разных праймеров и зондов, которые можно эффективно использовать для амплификации и/или детектирования мРНК STEAP-1. Полинуклеотиды STEAP-1 согласно изобретению используют для разных целей, включающих в себя, без ограничения, применение в качестве зондов и праймеров для амплификации и/или детектирования гена (генов) STEAP-1, мРНК или его (их) фрагментов; применение в качестве реагентов для диагноза и/или прогнозирования рака простаты и других злокачественных заболеваний; применение в качестве кодирующих последовательностей, способных управлять экспрессией полипептидов STEAP-1; применение в качестве средств, модулирующих или ингибирующих экспрессию гена (генов) STEAP-1 и/или трансляцию транскрипта (транскриптов) STEAP-1; применение в качестве терапевтических средств. Настоящее изобретение включает в себя применение любого описанного в данном документе зонда для идентификации и выделения нуклеотидной последовательности STEAP-1 или нуклеотидной последовательности, родственной STEAP-1, из природного источника, такого как человек или другие млекопитающие, а также самой выделенной нуклеотидной последовательности, которая содержит полноразмерную последовательность, присутствующую в используемом зонде, или ее большую часть. II.A.4. Выделение молекул нуклеиновых кислот, кодирующих STEAP-1. Последовательности кДНК STEAP-1, описанные в данном документе, можно использовать для выделения других полинуклеотидов, кодирующих генные продукты STEAP-1, а также для выделения полинуклеотидов, кодирующих гомологи генных продуктов STEAP-1, изоформ, образующихся в результате альтернативного сплайсинга, аллельных вариантов и мутантных форм генного продукта STEAP-1, а также полинуклеотидов, кодирующих аналоги белков, родственных STEAP-1. Для выделения полноразмерных кДНК, кодирующих ген STEAP-1, можно использовать хорошо известные методы молекулярного клонирования (см., например, Sambrook, J. et al., Molecular Cloning: A Laboratory Manual, 2 nd edition, Cold Spring Harbor Press, New York, 1989; Current Protocols in Molecular Biology. Ausubel et al., Eds., Wiley and Sons, 1995). Например, можно использовать методы клонирования на основе лямбда-фага с помощью коммерчески доступных систем клонирования (например, Lambda ZAP Express, Stratagene). Фаговые клоны, содержащие кДНК генов STEAP-1, можно идентифицировать путем зондирования меченой кДНК STEAP-1 или ее фрагментом. Например, в одном воплощении кДНК STEAP-1 (например, показанная на фиг. 2), или ее фрагмент, можно синтезировать и использовать в качестве зонда для идентификации перекрывающихся и полноразмерных кДНК, соответствующих гену STEAP-1. Сам ген STEAP-1 можно выделить путем скрининга библиотек геномных ДНК, библиотек бактериальных искусственных хромосом (ВАС), библиотек дрожжевых искусственных хромосом (YAC) и т.п. с использованием зондов или праймеров на основе ДНК STEAP-1. II.А.5. Рекомбинантные молекулы нуклеиновых кислот и системы векторов-хозяев. Данное изобретение также предлагает рекомбинантные молекулы ДНК или РНК, содержащие полинуклеотид STEAP-1, его фрагмент, аналог или гомолог, включающие в себя, без ограничения, фаги, плазмиды, фагмиды, космиды, YAC, ВАС, разные вирусные и невирусные векторы, хорошо известные в данной области, а также клетки, трансформированные или трансфицированные такими рекомбинантными молекулами ДНК или РНК. Способы получения таких молекул хорошо известны в данной области (см., например, Sambrook et al., 1989, приведено выше). Данное изобретение также предлагает систему вектора-хозяина, включающую в себя рекомбинантную молекулу ДНК, содержащую полинуклеотид STEAP-1, его фрагмент, аналог или гомолог в подходящей прокариотической или эукариотической клетке-хозяине. Примеры подходящих эукариотических клеток-хозяев включают в себя дрожжевую клетку, растительную клетку или животную клетку, такую как клетка млекопитающего или клетка насекомого (например, клетку, которую можно инфицировать бакуловирусом, такую как клетка Sf9 или клетка HighFive). Примеры подходящих клеток млекопитающих включают в себя различные клеточные линии рака простаты, такие как DU145 и TsuPr1, другие клеточные линии рака простаты, которые можно трансфицировать или трансдуцировать, первичные клетки (PrEC), a также ряд клеток млекопитающих, обычно используемых для экспрессии рекомбинантных белков (таких как клетки COS, CHO, 293, 293Т). Более конкретно, полинуклеотид, включающий в себя кодирующую последовательность STEAP-1 или ее фрагмент, аналог или гомолог, можно использовать для получения белков STEAP-1 или их фрагментов с использованием любого числа традиционно используемых и широко известных в данной области систем векторов-хозяев. Широкий ряд систем векторов-хозяев, подходящих для экспрессии белков STEAP-1 или их фрагментов, является доступным (см., например, Sambrook et al., 1989, приведено выше; Current Protocols in Molecular Biology, 1995, приведено выше). Векторы, предпочтительные для экспрессии в клетках млекопитающих, включают в себя, без ограничения, ркДНК 3.1 myc-His-tag (Invitrogen) и ретровирусный вектор pSR αtkneo (Muller et al., 1991, MCB, 11: 1785). С помощью указанных векторов экспрессии STEAP-1 можно экспрессировать в некоторых клеточных линиях рака простаты и клеточных линиях, не связанных с простатой, включающих в себя, например, 293, 293Т, RAT1, NIH 3Т3 и TsuPr1. Системы векторов-хозяев согласно изобретению можно использовать для получения белка STEAP-1 или его фрагмента. Такие системы векторов-хозяев можно также использовать для изучения функциональных свойств STEAP-1 и мутантов или аналогов STEAP-1. Рекомбинантный человеческий белок STEAP-1 или его аналог, гомолог или фрагмент также можно получить с использованием клеток млекопитающих, трансфицированных конструкцией, кодирующей нуклеотид, родственный STEAP-1. Например, клетки 293Т можно трансфицировать экспрессионной плазмидой, кодирующей STEAP-1 или его фрагмент, аналог или гомолог, с последующими экспрессией белка, родственного STEAP-1, в клетках 293Т и выделением рекомбинантного белка STEAP-1 с помощью стандартных методов очистки (таких как аффинная очистка с использованием антител против STEAP-1). В другом воплощении последовательность, кодирующую STEAP-1, субклонируют в ретровирусном векторе pSR αMSVtkneo и используют для инфицирования различных клеточных линий млекопитающих, таких как NIH 3Т3, TsuPrl, 293 и RAT1, с последующей экспрессией STEAP-1 в клеточных линиях. Можно использовать и другие системы экспрессии, хорошо известные в данной области. Для получения секретируемой формы рекомбинантного белка STEAP-1 можно использовать экспрессионные конструкции, включающие в себя последовательность, кодирующую лидерный пептид, и объединенную с ней в рамке считывания последовательность, кодирующую STEAP-1. Как указано в данном описании, избыточность генетического кода приводит к возникновению вариаций в генных последовательностях STEAP-1. В частности, как известно в данной области, конкретные виды-хозяева часто имеют характерные кодоновые предпочтения, и, следовательно, раскрываемую последовательность можно адаптировать для определенного хозяина. Например, в предпочтительных аналоговых кодоновых последовательностях редкие кодоны (т.е. кодоны, периодичность встречаемости которых в известных последовательностях конкретного хозяина составляет менее чем приблизительно 20%) обычно заменяют на кодоны с более высокой встречаемостью. Кодоновые предпочтения для конкретных видов рассчитывают, например, с помощью таблиц использования кодонов, которые можно найти в Интернете, например, на сайте URL dna.affrc.go.jp/-nakamura/codon.html. Известны другие модификации последовательности с целью увеличения экспрессии белка в клетке-хозяине. Данные модификации включают в себя удаление последовательностей, кодирующих мнимые сигналы полиаденилирования, сигналы сплайсинга экзонов/интронов, транспозон-подобные повторы и/или другие подобные хорошо охарактеризованные последовательности, которые оказывают неблагоприятное влияние на экспрессию генов. Содержание GC в последовательности доводят до среднего уровня, характерного для конкретной клетки-хозяина, который рассчитывают по известным генам, экспрессирующимся в данном хозяине. По возможности последовательность модифицируют так, чтобы избежать прогнозированного образования шпилек во вторичной структуре мРНК. Другие полезные модификации включают в себя добавление консенсусной последовательности инициации трансляции в начало открытой рамки считывания, как описано Kozak, Mol. Cell Biol., 9: 5073-5080 (1989). Опытные специалисты понимают, что общее правило, согласно которому эукариотические рибосомы инициируют трансляцию исключительно по 5'-проксимальному кодону AUG, нарушается только в редких случаях (см., например, Kozak. PNAS. 92(7): 2662-2666 (1995) и Kozak. NAR. 15(20): 8125-8148 (1987)). III. Белки, родственные STEAP-1. В другом аспекте настоящее изобретение предлагает белки, родственные STEAP-1. Конкретные воплощения белков STEAP-1 включают в себя полипептид, содержащий полноразмерную аминокислотную последовательность человеческого STEAP-1, изображенную на фиг. 2 или 3, предпочтительно на фиг. 2А, или ее фрагмент. Альтернативно, воплощения белков STEAP-1 включают в себя вариантные, гомологичные или аналоговые полипептиды, которые содержат изменения в аминокислотной последовательности STEAP-1, изображенной на фиг. 2 или 3. Воплощения полипептида STEAP-1 включают в себя полипептид STEAP-1, имеющий последовательность, изображенную на фиг. 2, пептидную последовательность STEAP-1, изображенную на фиг. 2, где Т может обозначать U; по меньшей мере 10 смежных нуклеотидов полипептида, имеющего последовательность, изображенную на фиг. 2; или по меньшей мере 10 смежных пептидов полипептида, имеющего последовательность, изображенную на фиг. 2, где Т может обозначать U. Например, воплощения пептидов STEAP-1 включают в себя, без ограничения: (I) белок, по существу содержащий или содержащий аминокислотную последовательность, изображенную на фиг. 2A-2Q или 3A-3D; (II) белок, родственный STEAP-1, который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% гомологичен полноразмерной аминокислотной последовательности, изображенной на фиг. 2A-2Q или 3A-3D; (III) белок, родственный STEAP-1, который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен полноразмерной аминокислотной последовательности, изображенной на фиг. 2A-2Q или 3A-3D; (IV) белок, содержащий по меньшей мере один пептид из перечисленных в табл. V-LI, как описано в патентной заявке США 10/236878, поданной 6 сентября 2002 г., конкретное содержание которой полностью включено в данное описание в качестве ссылки, необязательно при условии, что данный белок не является полноразмерным, изображенным на фиг. 2; (V) белок, содержащий по меньшей мере один пептид из перечисленных в общей сложности в табл. V-XVIII, данные пептиды также приведены в общей сложности в табл. XXII-LI, необязательно при условии, что данный белок не является полноразмерным, изображенным на фиг. 2; (VI) белок, содержащий по меньшей мере два пептида, выбранных из пептидов, приведенных в табл. V-LI, необязательно при условии, что данный белок не является полноразмерным, изображенным на фиг. 2; (VII) белок, содержащий по меньшей мере два пептида, выбранных из пептидов, приведенных в общей сложности в табл. V-LI, при условии, что данный белок не является смежной последовательностью по отношению к аминокислотной последовательности фиг. 2; (VIII) белок, содержащий по меньшей мере один пептид, выбранный из пептидов, приведенных в табл. V-XVIII; и по меньшей мере один пептид, выбранный из пептидов, приведенных в табл. XXII-LI, при условии, что данный белок не является смежной последовательностью по отношению к аминокислотной последовательности фиг. 2; (IX) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3А, с любым целочисленным приращением вплоть до 339 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю гидрофильности, приведенному на фиг. 5; (X) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3А, с любым целочисленным приращением вплоть до 339 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением меньше 0,5 по профилю гидрофобности, приведенному на фиг. 6; (XI) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3А, с любым целочисленным приращением вплоть до 339 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю процента доступных остатков, приведенному на фиг. 7; (XII) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3А, с любым целочисленным приращением вплоть до 339 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю средней жесткости, приведенному на фиг. 8; (XIII) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 аминокислот белка, изображенного на фиг. 3А, с любым целочисленным приращением вплоть до 339 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю бета-складки, приведенному на фиг. 9; (XIV) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3В или 3D, с любым целочисленным приращением вплоть до 258 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю гидрофильности, приведенному на фиг. 5; (XV) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3В или 3D, с любым целочисленным приращением вплоть до 258 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением меньше 0,5 по профилю гидрофобности, приведенному на фиг. 6; (XVI) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3В или 3D, с любым целочисленным приращением вплоть до 258 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю процента доступных остатков, приведенному на фиг. 7; (XVII) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3В или 3D, с любым целочисленным приращением вплоть до 258 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю средней жесткости, приведенному на фиг. 8; (XVIII) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 аминокислот белка, изображенного на фиг. 3В или 3D, с любым целочисленным приращением вплоть до 258 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю бета-складки, приведенному на фиг. 9; (XIX) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3С, с любым целочисленным приращением вплоть до 282 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю гидрофильности, приведенному на фиг. 5; (XX) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3С, с любым целочисленным приращением вплоть до 282 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением меньше 0,5 по профилю гидрофобности, приведенному на фиг. 6; (XXI) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3С, с любым целочисленным приращением вплоть до 282 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю процента доступных остатков, приведенному на фиг. 7; (XXII) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислот белка, изображенного на фиг. 3С, с любым целочисленным приращением вплоть до 282 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю средней жесткости, приведенному на фиг. 8; (XXIII) полипептид, содержащий по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 аминокислот белка, изображенного на фиг. 3С, с любым целочисленным приращением вплоть до 282 соответственно, который содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 аминокислотных положений со значением больше 0,5 по профилю бета-складки, приведенному на фиг. 9; (XXIV) пептид, который встречается в общей сложности по меньшей мере два раза в табл. V-XVIII и XXII-LI; (XXV) пептид, который встречается в общей сложности по меньшей мере три раза в табл. V-XVIII и XXII-LI; (XXVI) пептид, который встречается в общей сложности по меньшей мере четыре раза в табл. V-XXVIII и XXII-LI; (XXVII) пептид, который встречается в общей сложности по меньшей мере пять раз в табл. V-XVIII и XXII-LI; (XXVIII) пептид, который встречается по меньшей мере один раз в табл. V-XVIII и по меньшей мере один раз в табл. XXII-LI; (XXIX) пептид, который встречается по меньшей мере один раз в табл. V-XVIII и по меньшей мере два раза в табл. XXII-LI; (XXX) пептид, который встречается по меньшей мере два раза в табл. V-XVIII и по меньшей мере один раз в табл. XXII-LI; (XXXI) пептид, который встречается по меньшей мере два раза в табл. V-XVIII и по меньшей мере два раза в табл. XXII-LI; (XXXII) пептид, который имеет один, два, три, четыре или пять из нижеследующих характерных признаков, или олигонуклеотид, кодирующий такой пептид: i) участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю гидрофильности, приведенному на фиг. 5, равное или превышающее 0,5, 0,6, 0,7, 0,8, 0,9 или равное 1,0; ii) участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю гидропатичности, приведенному на фиг. 6, равное или превышающее 0,5, 0,4, 0,3, 0,2, 0,1 или равное 0,0; iii) участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю процента доступных остатков, приведенному на фиг. 7, равное или превышающее 0,5, 0,6, 0,7, 0,8, 0,9 или равное 1,0; iv) участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю средней жесткости, приведенному на фиг. 8, равное или превышающее 0,5, 0,6, 0,7, 0,8, 0,9 или равное 1,0; или v) участок, состоящий по меньшей мере из 5 аминокислот конкретного пептида, приведенного на фиг. 3, с любым целочисленным приращением вплоть до полного размера белка, изображенного на фиг. 3, который содержит аминокислотное положение, имеющее значение по профилю бета-складки, приведенному на фиг. 9, равное или превышающее 0,5, 0,6, 0,7, 0,8, 0,9 или равное 1,0$ (XXXIII) композиция, содержащая пептид по любому из пп.I-XXXII или антитело или его связывающий фрагмент, вместе с фармацевтическим наполнителем и/или в составе человеческой стандартной лекарственной формы; (XXXIV) способ применения пептида по любому из пп.I-XXXII или антитела или его связывающего фрагмента либо композиции по п.XXXIII для модуляции экспрессии STEAP-1 в клетке; (XXXV) способ применения пептида по любому из пп.I-XXXII или антитела или его связывающего фрагмента либо композиции по п.XXXIII для диагностики, профилактики, прогнозирования или лечения субъекта, несущего клетку, экспрессирующую STEAP-1; (XXXVI) способ применения пептида по любому из пп.I-XXXII или антитела или его связывающего фрагмента либо композиции по п.XXXIII для диагностики, профилактики, прогнозирования или лечения субъекта, несущего клетку, экспрессирующую STEAP-1, где указанная клетка относится к раковой ткани, приведенной в табл. I; (XXXVII) способ применения пептида по любому из пп.I-XXXII или антитела или его связывающего фрагмента либо композиции по п.XXXIII для диагностики, профилактики, прогнозирования или лечения рака; (XXXVIII) способ применения пептида по любому из пп.I-XXXII или антитела или его связывающего фрагмента либо композиции по п.XXXIII для диагностики, профилактики, прогнозирования или лечения рака ткани, приведенной в табл. I; и (XXXIX) способ применения пептида по любому из пп.I-XXXII или антитела или его связывающего фрагмента либо композиции по п.XXXIII для идентификации или характеристики модулятора экспрессии STEAP-1 в клетке. В данном описании подразумевается, что интервал конкретно включает в себя все находящиеся внутри него целочисленные значения. Типичные воплощения согласно изобретению, раскрытые в данном описании, включают в себя полинуклеотиды STEAP-1, которые кодируют конкретные положения последовательностей мРНК STEAP-1 (а также комплементарные таким последовательностям), такие как полинуклеотиды, кодирующие, например, следующие белки и/или их фрагменты: 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145, 150, 155, 160, 165, 170, 175, 180, 185, 190, 195, 200, 225, 250, 275, 300, 325, 330, 335, 339 или более смежных аминокислот STEAP-1, вариант 1; максимальная длина других вариантов составляет: вариант 2 - 258 аминокислот; вариант 3 - 282 аминокислоты, вариант 4 - 258 аминокислот. Как правило, природные аллельные варианты человеческого STEAP-1 обладают высокой степенью структурной идентичности и гомологии (например, гомология составляет 90% или более). Обычно аллельные варианты белка STEAP-1 содержат консервативные аминокислотные замены в последовательностях STEAP-1, описанных в данном документе, или содержат аминокислотную замену в соответствующем положении гомолога STEAP-1. Один класс аллельных вариантов STEAP-1 включает в себя белки, которые являются в высокой степени гомологичными, по меньшей мере, маленькому участку конкретной аминокислотной последовательности STEAP-1, но также содержат существенное отличие от структуры последовательности, такое как неконсервативная замена, усечение, вставка или сдвиг рамки считывания. В области генетики считается, что термины подобие, идентичность и гомология, относящиеся к сравнению белковых последовательностей, имеют разные значения. Более того, большое значение для описания взаимоотношений между членами конкретного семейства белков в одном организме и членами такого же семейства в других организмах могут иметь ортология и паралогия. Сокращенные наименования аминокислот приведены в табл. II. Консервативные аминокислотные замены в белке зачастую могут быть осуществлены без изменения конформации или функции белка. Белки согласно изобретению могут содержать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 консервативных замен. Воплощения согласно изобретению, раскрытые в данном описании, включают в себя широкий ряд принятых в данной области вариантов или аналогов белков STEAP-1, таких как полипептиды, содержащие вставки, делеции и замены аминокислот. Варианты STEAP-1 можно получить с помощью известных в данной области методов, таких как сайт-направленный мутагенез, сканирование по аланину и мутагенез методом ПЦР. Сайт-направленный мутагенез (Carter et al., Nucl. Acids Res., 13: 4331 (1986); Zoller et al., Nucl. Acids Res., 10: 6487 (1987)), кассетный мутагенез (Wells et al., Gene, 34: 315 (1985)), рестрикционный селекционный мутагенез (Wells et al., Philos. Trans. R. Soc. London SerA, 317: 415 (1986)) или другие известные методы можно проводить на клонированной ДНК с получением варианта ДНК STEAP-1. Анализ с аминокислотным сканированием также можно использовать для идентификации одной или нескольких аминокислот по всей длине непрерывной последовательности, участвующей в конкретной биологической активности, такой как белок-белковое взаимодействие. К предпочтительным аминокислотам для сканирования относятся сравнительно маленькие нейтральные аминокислоты. Такие аминокислоты включают в себя аланин, глицин, серин и цистеин. Как правило, аланин является предпочтительной сканирующей кислотой в данной группе, поскольку он не содержит боковых цепей за бета-атомом углерода и с меньшей вероятностью изменяет конформацию основной цепи варианта. Аланин также часто предпочитают из-за того, что он является наиболее распространенной аминокислотой. Кроме того, он зачастую присутствует как в заглубленных, так и в экспонированных положениях (Creighton, The Proteins (W.H. Freeman & Co., N.Y.); Chothia, J. Mol. Biol., 150: 1 (1976)). Если аланиновая замена не дает адекватных количеств варианта, можно использовать изостерическую аминокислоту. Как указано в данном описании, отличительным признаком вариантов, аналогов или гомологов STEAP-1 является то, что они содержат по меньшей мере один эпитоп, способный "перекрестно взаимодействовать" с белком STEAP-1, имеющим аминокислотную последовательность, приведенную на фиг. 3. Использованный термин "перекрестное взаимодействие" означает, что антитело или Т-клетка, которая специфически связывается с вариантом STEAP-1, также может специфически связываться с белком STEAP-1, имеющим аминокислотную последовательность, приведенную на фиг. 3. Полипептид перестает быть вариантом белка, изображенного на фиг. 3, если он больше не содержит эпитопа, который может распознаваться антителом или Т-клеткой, специфически связывающимися с исходным белком STEAP-1. Специалистам в данной области известно, что антитела, распознающие белки, связываются с эпитопами разного размера, причем минимальным эпитопом, как правило, считается эпитоп, который содержит совокупность приблизительно из четырех или пяти аминокислот, смежных или нет. См., например, Nair et al., J. Immunol. 2000, 165(12): 6949-6955; Hebbes et al., Mol. Immunol. (1989), 26(9): 865-73; Schwartz et al., J. Immunol. (1985), 135(4): 2598-608. Другие классы вариантов белков, родственных STEAP-1, имеют 70, 75, 80, 85 или 90% или более процентов подобия с аминокислотной последовательностью, приведенной на фиг. 3, или с ее фрагментом. Другие конкретные классы вариантов или аналогов STEAP-1 включают в себя один или несколько биологических мотивов STEAP-1, описанных в данном документе или известных в настоящее время в данной области. Таким образом, настоящее изобретение охватывает аналоги фрагментов STEAP-1 (нуклеотидных или аминокислотных), которые имеют измененные функциональные (например, иммуногенные) свойства по сравнению с исходным фрагментом. Следует понимать, что мотивы, описанные в данном документе или известные из предыдущего уровня техники, относятся к нуклеотидным или аминокислотным последовательностям, приведенным на фиг. 2 или 3. Как указано в данном документе, воплощения заявляемого изобретения включают в себя полипептиды, содержащие последовательность меньше полноразмерной аминокислотной последовательности белка STEAP-1, изображенного на фиг. 2 или 3. Например, типичные воплощения согласно изобретению включают в себя пептиды/белки, содержащие 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или больше смежных аминокислот белка STEAP-1, изображенного на фиг. 2 или 3. Кроме того, типичные воплощения согласно изобретению, раскрытые в данном описании, включают в себя полипептиды, содержащие последовательность приблизительно от аминокислоты 1 до аминокислоты 10 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 10 до аминокислоты 20 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 20 до аминокислоты 30 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 30 до аминокислоты 40 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 40 до аминокислоты 50 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 50 до аминокислоты 60 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 60 до аминокислоты 70 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 70 до аминокислоты 80 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 80 до аминокислоты 90 белка STEAP-1, изображенного на фиг. 2 или 3; полипептиды, содержащие последовательность приблизительно от аминокислоты 90 до аминокислоты 100 белка STEAP-1, изображенного на фиг. 2 или 3, и т.д. на протяжении всей аминокислотной последовательности STEAP-1. Более того, полипептиды, содержащие последовательность приблизительно от аминокислоты 1 (или 20, или 30, или 40 и т.д.) до аминокислоты 20 (или 130, или 140, или 150 и т.д.) белка STEAP-1, изображенного на фиг. 2 или 3, являются воплощениями согласно изобретению. Следует понимать, что начальные и конечные положения, указанные в данном разделе, относятся к конкретному положению, а также к положению в интервале плюс или минус 5 остатков. Белки, родственные STEAP-1, получают с помощью стандартных методов пептидного синтеза или с помощью методов химического расщепления, хорошо известных в данной области. Альтернативно, для получения нуклеиновых кислот, кодирующих белок, родственный STEAP-1, можно использовать рекомбинантные методы. В одном воплощении молекулы нуклеиновых кислот являются средством для получения определенных фрагментов белка STEAP-1 (или его вариантов, гомологов или аналогов). III.А. Мотив-несущие воплощения белков. Другие иллюстративные воплощения согласно изобретению, раскрытые в данном описании, включают в себя полипептиды STEAP-1, содержащие аминокислотные остатки одного или нескольких биологических мотивов, входящих в состав полипептидной последовательности STEAP-1, приведенной на фиг. 2 или 3. Многие мотивы известны в данной области и присутствие таких мотивов в белке можно определить с помощью ряда широко доступных сайтов Интернета (см., например, адреса URL: pfam.wustl.edu/; searchlauncher.bcm.tmc.edu/seq-search/struc-predict.html; psort.ims.u-tokyo.ас.jp/; cbs.dtu.dk/; ebi.ac.uk/interpro/scan.html; expasy.ch/tools/scnpsitl.html; Epimatrix ™ and Epimer ™, Brown University, brown.edu/Research/TB-HIV_Lab/epimatrix/epimatrix.html и BIMAS, bimas.dcrt.nih.gov/). Мотив-несущие субпоследовательности всех белковых вариантов STEAP-1 приведены и идентифицированы в табл. V-XVIII и XXII-LI. В табл. IV(H) приведено несколько часто ввстречающихся мотивов, обнаруженных в результате поиска pfam (см. адрес URL pfam.wustl.edu/). В колонках таблицы IV(H) приведены (1) сокращенные наименования мотивов; (2) процент идентичности для разных членов семейства мотивов; (3) наименование или описание мотива и (4) наиболее общая функция; информация о расположении указывается, если мотив связан с определенным положением. Полипептиды, содержащие один или несколько описанных выше мотивов STEAP-1, можно использовать для объяснения характерных признаков злокачественного фенотипа, поскольку существуют данные, что описанные выше мотивы STEAP-1 связаны с нарушением регуляции роста и что при некоторых злокачественных заболеваниях происходит сверхэкспрессия STEAP-1 (см., например, табл. I). Известно, что такие ферменты, как, например, казеинкиназа II, цАМФ- и camp-зависимая протеинкиназа и протеинкиназа С, связаны с развитием злокачественного фенотипа (см., например, Chen et al., Lab Invest., 78(2): 165-174 (1998); Gaiddon et al., Endocrinology, 136(10): 4331-4338 (1995); Hall et al., Nucleic Acids Research, 24(6): 1119-1126 (1996); Peterziel et al., Oncogene, 18(46): 6322-6329 (1999) и O'Brian, Oncol. Rep. 5(2): 305-309 (1998)). Кроме того, с раковыми заболеваниями и их развитием также связаны модификации белков, такие как гликозилирование и миристоилирование (см., например, Dennis et al., Biochem. Biophys. Acta 1473(1):21-34 (1999); Raju et al., Exp. Cell Res. 235(1): 145-154 (1997)). Амидирование представляет собой другую модификацию белка, тоже связанную с раковыми заболеваниями и их развитием (см., например, Treston et al., J. Natl. Cancer Inst. Monogr. (13): 169-175 (1992)). В другом воплощении белки согласно изобретению содержат один или несколько иммунореактивных эпитопов, идентифицированных в соответствии с принятыми в данной области методами, таких как пептиды, приведенные в табл. V-XVIII и XXII-LI. Эпитопы CTL можно определить с помощью специальных алгоритмов, используемых для идентификации пептидов в белке STEAP-1, способных связываться с определенными аллелями HLA с оптимальной эффективностью (например, табл. IV; Epimatrix ™ and Epmer ™, Brown University, URL brown.edu/Research/TB-HIV_Lab/epimatrix/epimatrix.html и BIMAS, URL bimas.dcrt.nih.gov/). Кроме того, способы идентификации пептидов, которые обладают достаточным сродством к молекулам HLA и по имеющимся данным могут являться иммуногенными эпитопами, хорошо известны в данной области и не требуют проведения излишних экспериментов. Способы идентификации пептидов, которые являются иммуногенными эпитопами, также хорошо известны в данной области и могут проводиться без излишнего экспериментирования либо in vitro, либо in vivo. В данной области также известны принципы создания аналогов таких эпитопов с целью модулирования иммуногенности. Например, в качестве исходного можно использовать эпитоп, который несет мотив CTL или HTL (см., например, мотивы/супермотивы HLA класса I и HLA класса II в табл. IV). Аналог эпитопа получают путем замены аминокислоты в одном определенном положении на другую аминокислоту, специфичную для данного положения. Например, на основе остатков, приведенных в табл. IV, неблагоприятный остаток можно заменить на любой другой остаток, такой как предпочтительный остаток; менее предпочтительный остаток можно заменить на предпочтительный остаток или заменить изначально присутствующий предпочтительный остаток на другой предпочтительный остаток. Замены можно осуществлять по первичным анкорным положениям или по другим положениям пептида; см., например, табл. IV. Ряд ссылок отражает состояние уровня техники, связанного с идентификацией и получением эпитопов белка, представляющего интерес, а также его аналогов. См., например, WO 97/33602, Chesnut et al.; Sette, Immunogenetics, 1999 50(3-4): 201-212; Sette et al., J. Immunol. 2001, 166(2): 1389-1397; Sidney et al., Hum. Immunol. 1997, 58(1): 12-20; Kondo et al., Immunogenetics, 1997, 45(4): 249-258; Sidney et al., J. Immunol. 1996, 157(8): 3480-90 и Falk et al., Nature, 351: 290-6 (1991); Hunt et al., Science, 255: 1261-3 (1992); Parker et al., J. Immunol. 149: 3580-7 (1992); Parker et al., J. Immunol. 152: 163-75 (1994)); Kast et al., 1994 152(8): 3904-12; Borras-Cuesta et al., Hum. Immunol. 2000 61(3): 266-278; Alexander et al., J. Immunol. 2000, 164(3): 1625-1633; Alexander et al., PMID: 7895164, UI: 95202582; O'Sullivan et al., J. Immunol. 1991, 147(8): 2663-2669; Alexander et al., Immunity, 1994, 1(9): 751-761 и Alexander et al., Immunol. Res. 1998, 18(2): 79-92. Родственные воплощения согласно изобретению включают в себя полипептиды, содержащие сочетания разных мотивов, приведенных в табл. IV(A)-IV(D) и IV(H), и/или один или несколько предсказанных эпитопов CTL, приведенных в табл. V-XVIII и XXII-LI, и/или один или несколько предсказанных эпитопов HTL, приведенных в табл. XLVIII-LI, и/или один или несколько известных в данной области мотивов, связывающих Т-клетки. Предпочтительные воплощения не содержат вставок, делеций или замен внутри мотивов или внутри последовательностей, встроенных в полипептиды. Кроме того, желательными могут быть воплощения, которые включают в себя ряд любых N-концевых и/или С-концевых аминокислотных остатков на любой стороне данных мотивов (например, которые включают в себя более крупный фрагмент полипептидной структуры, содержащий мотив). Как правило, число N-концевых и/или С-концевых аминокислотных остатков на любой стороне мотива находится в интервале приблизительно от 1 до 100 аминокислотных остатков, предпочтительно приблизительно от 5 до 50 аминокислотных остатков. Белки, родственные STEAP-1, могут воплощаться в разных формах, предпочтительно в форме выделенного белка. Очищенная молекула белка STEAP-1 практически не содержит других белков или молекул, ухудшающих связывание STEAP-1 с антителом, Т-клеткой или другим лигандом. Природа и степень выделения и очистки зависят от предполагаемого применения. Воплощения белков, родственных STEAP-1, включают в себя очищенные STEAP-1-родственные белки и функциональные растворимые STEAP-1-родственные белки. В одном воплощении функциональный растворимый белок STEAP-1 или его фрагмент сохраняет способность связывать антитело, Т-клетку или другой лиганд. Данное изобретение также предлагает белки STEAP-1, содержащие биологически активные фрагменты аминокислотной последовательности STEAP-1, изображенной на фиг. 2 или 3. Такие белки обладают свойствами исходного белка STEAP-1, такими, как способность вызывать образование антител, специфически связывающихся с эпитопом, ассоциированным с исходным белком STEAP-1; способность связываться с такими антителами; способность вызывать активацию HTL или CTL и/или способность к распознаванию HTL или CTL, которые специфически связываются с исходным белком. STEAP-1-родственные полипептиды, которые содержат особенно важные структуры, можно прогнозировать и/или идентифицировать с помощью разных аналитических методов, хорошо известных в данной области, включающих в себя, например, методы Чоу-Фасмана, Гарньер-Робсона, Кит-Дулитла, Эйсенберга, Карплус-Шультца или Джеймсон-Вульфа, или на основе иммуногенности. Фрагменты, содержащие такие структуры, особенно пригодны для получения субъединица-специфичных антител против STEAP-1 или Т-клеток либо для идентификации клеточных факторов, связывающих STEAP-1. Например, для получения профилей гидрофильности и идентификации иммуногенных пептидных фрагментов можно использовать метод, описанный в Норр, Т.P. and Woods, K.R., 1981, Proc. Natl. Acad. Sci. U.S.A. 78: 3824-3828. Для получения профилей гидропатичности и идентификации иммуногенных пептидных фрагментов можно использовать метод, описанный в Kyte, J. and Doolittle, R.F., 1982, J. Mol. Biol. 157: 105-132. Для получения профилей процента (%) доступных остатков и идентификации иммуногенных пептидных фрагментов можно использовать метод, описанный в Janin J., 1979, Nature, 277: 491-492. Для получения профилей средней жесткости и идентификации иммуногенных пептидных фрагментов можно использовать метод, описанный в Bhaskaran R., Ponnuswamy P.K., 1988, Int. J. Pept. Protein Res. 32: 242-255. Для получения профилей бета-складки и идентификации иммуногенных пептидных фрагментов можно использовать метод, описанный в Deleage, G., Roux В., 1987, Protein Engineering, 1: 289-294. Эпитопы CTL можно определить с помощью специальных алгоритмов, используемых для идентификации пептидов в белке STEAP-1, способных связываться с определенными аллелями HLA с оптимальной эффективностью (например, с помощью алгоритмов, которые можно найти на сайте SYFPEITHI в глобальной сети Интернет URL syfpeithi.bmi-heidelberg.com/; перечисленных в табл. IV(А)-IV(Е); Epimatrix ™ and Epimer ™, Brown University, URL (brown. edu/Research/TB-HIV_Lab/epimatrix/epimatrix. html) и BIMAS, URL bimas.dcrt.nih.gov/). В качестве иллюстрации были предсказаны пептидные эпитопы из STEAP-1 применительно к человеческим молекулам МНС класса I, например HLA-A1, А2, A3, A11, A24, B7 и В35 (см., например, табл. V-XVIII, XXII-LI). А именно, полную аминокислотную последовательность белка STEAP-1 и соответствующие фрагменты других вариантов, т.е. в случае HLA класса I прогнозы 9 фланкирующих участков с каждой стороны точечной мутации или сочленения экзона и в случае HLA класса II прогнозы 14 фланкирующих участков с каждой стороны точечной мутации или сочленения экзона, соответствующие данному варианту, вводят в алгоритм поиска пептидного мотива HLA, который можно найти на указанном выше web-сайте Раздела Биоинформатики и молекулярного анализа (BIMAS); а также на сайте SYFPEITHI, URL syfpeithi.bmi-heidelberg.com/. Алгоритм поиска пептидного мотива HLA разработан Dr. Ken Parker на основе связывания специфических пептидных последовательностей в бороздках молекул HLA класса I, в частности HLA-A2 (см., например, Falk et al., Nature, 351: 290-6 (1991); Hunt et al., Science 255: 1261-3 (1992); Parker et al., J. Immunol. 149: 3580-7 (1992); Parker et al., J. Immunol. 152: 163-75 (1994)). С помощью данного алгоритма можно определить местоположение и ранжировать пептиды, состоящие из 8, 9 и 10 мономеров, в полной последовательности белка с целью прогнозирования связывания с HLA-A2, а также с рядом других молекул HLA класса I. Многие пептиды, связывающие HLA класса I, состоят из 8, 9, 10 или 11 мономеров. Например, в случае HLA-A2 класса I эпитопы предпочтительно содержат лейцин (L) или метионин (М) в положении 2 и валин (V) или лейцин (L) на С-конце (см., например, Parker et al., J. Immunol. 149: 3580-7 (1992)). Отдельные результаты прогнозирования STEAP-1-связывающих пептидов приведены в табл. V-XVIII и XXII-LI данного описания. В табл. V-XVIII и XXII-XLVIII приведены некоторые кандидаты семейства, каждый член которого содержит 9 и 10 мономеров, а также их расположение, аминокислотная последовательность каждого пептида и теоретический показатель связывания. В табл. XLVIII-LI приведены некоторые кандидаты семейства, каждый член которого содержит 15 мономеров, а также их расположение, аминокислотная последовательность каждого пептида и теоретический показатель связывания. Показатель связывания соответствует теоретическому полупериоду диссоциации комплексов, содержащих пептид, при 37 °С и рН 6,5. Предположительно пептиды с наивысшим показателем связывания наиболее прочно связываются с HLA класса I на поверхности клетки в течение наибольшего периода времени и, следовательно, представляют собой наилучшие иммуногенные мишени для распознавания Т-клетками. Реальное связывание пептидов с аллелью HLA можно определить путем стабилизации экспрессии HLA в антигенпроцессирующей дефектной клеточной линии Т2 (см., например, Xue et al., Prostate, 30: 73-8 (1997) и Peshwa et al., Prostate, 36: 129-38 (1998)). Иммуногенность конкретных пептидов можно определить in vitro путем стимуляции CD8+ цитотоксических Т-лимфоцитов (CTL) в присутствии антигенпрезентирующих клеток, таких как дендритные клетки. Следует понимать, что каждый эпитоп, предсказанный с помощью сайта BIMAS, сайтов Epimer ™ and Epimatrix ™ или определенный с помощью мотивов HLA класса I или класса II, доступных в данной области или которые стали частью данной области, таких как перечисленные в табл. IV (или определенные с помощью сайта глобальной сети URL syfpeithi.bmi-heidelberg.com/ или BIMAS, bimas.dcrt.nih.gov/), "применяется" к белку STEAP-1 в соответствии с данным изобретением. Специалисту в соответствующей области должно быть понятно, что в данном контексте термин "применяется" означает, что белок STEAP-1 анализируют, например, визуально или с помощью компьютерных программ поиска. В объем согласно изобретению входит каждая субпоследовательность белка STEAP-1, состоящая из 8, 9, 10 или 11 аминокислотных остатков, которая несет мотив HLA класса I, или субпоследовательность, состоящая из 9 или более аминокислотных остатков, которые несут мотив HLA класса II. III.В. Экспрессия белков, родственных STEAP-1. В воплощении, описанном в нижеследующих примерах, STEAP-1 можно с помощью традиционных способов экспрессировать в клетках (таких как клетки 293Т), трансфицированных коммерчески доступным вектором экспрессии, таким как CMV-зависимый вектор экспрессии, кодирующий STEAP-1 с С-концевым 6 × His и MYC-маркером (ркДНК3.1/mycHis, Invitrogenor tag-5, GenHunter Corporation, Nashville TN). Вектор tag-5 обеспечивает секреторный сигнал IgGK, который можно использовать для облегчения продукции секретируемого белка STEAP-1 в трансфицированных клетках. Секретированный в культуральную среду His-меченый STEAP-1 можно очистить, например, стандартными методами с использованием никелевой колонки. III.C. Модификации белков, родственных STEAP-1. В объем согласно изобретению входят модификации белков, родственных STEAP-1, таких как ковалентные модификации. Один тип ковалентной модификации включает в себя взаимодействие меченых аминокислотных остатков полипептида STEAP-1 с органическим дериватизирующим средством, которое способно взаимодействовать с отдельными боковыми цепями N- или С-концевых остатков белка STEAP-1. Другой тип ковалентной модификации полипептида STEAP-1, входящий в объем согласно изобретению, включает в себя изменение природного характера гликозилирования белка согласно изобретению. Следующий тип ковалентной модификации STEAP-1 включает в себя присоединение полипептида STEAP-1 к одному из ряда небелковых полимеров, таких как полиэтиленгликоль (PEG), полипропиленгликоль или полиоксиалкилены, с помощью способов, описанных в патентах США № 4640835; 4496689; 4301144; 4670417; 4791192 или 4179337. STEAP-1-родственные белки согласно изобретению также можно модифицировать с получением химерной молекулы, в которой STEAP-1 гибридизован с другой, гетерологичной полипептидной или аминокислотной последовательностью. Такую химерную молекулу можно получить химическими или рекомбинантными способами. Химерная молекула может содержать белок согласно изобретению, гибридизованный с другим опухолевым специфическим антигеном или его фрагментом. Альтернативно, белок в соответствии с данным изобретением может включать в себя гибрид фрагментов последовательности STEAP-1 (аминокислотной или нуклеотидной) так, что полученная молекула, несмотря на ее длину, не является непосредственно гомологичной аминокислотным или нуклеотидным последовательностям, изображенным на фиг. 2 или 3. Такая химерная молекула может содержать несколько одинаковых субпоследовательностей STEAP-1. Химерная молекула может включать в себя гибрид STEAP-1-родственного белка, содержащего в качестве маркера полигистидиновый эпитоп, который может селективно связывать иммобилизованный никель, с цитокинами или факторами роста. Эпитопный маркер обычно помещают на амино- или карбоксильные концы белка STEAP-1. В альтернативном воплощении химерная молекула может включать в себя гибрид белка, родственного STEAP-1, с иммуноглобулином или отдельным фрагментом иммуноглобулина. В случае бивалентной формы химерной молекулы (также называемой "иммуноадгезин") такой гибрид можно получить с использованием Fc-участка молекулы IgG. Гибриды Ig предпочтительно содержат растворимую форму полипептида STEAP-1 (трансмембранный домен удален или инактивирован) вместо по меньшей мере одного вариабельного участка молекулы Ig. В предпочтительном воплощении иммуноглобулиновый гибрид содержит шарнирные участки СН2 и СН3 или шарнирные участки CH1, CH2 и СН3 молекулы IgG 1 . Получение иммуноглобулиновых гибридов описано, например, в патенте США № 5428130, поданном 27 июня 1995 г. III.D. Применение STEAP-1-родственных белков. Белки согласно изобретению имеют несколько разных применений. Поскольку при раке простаты и при других злокачественных заболеваниях происходит интенсивная экспрессия STEAP-1, белки, родственные STEAP-1, используют для определения статуса генных продуктов STEAP-1 в нормальных тканях по сравнению с раковыми и выявления злокачественного фенотипа на основе полученных результатов. Обычно полипептиды из конкретных участков белка STEAP-1 используют для анализа изменений (таких как делеции, инсерции, точечные мутации и др.) в данных участках (таких как участки, содержащие один или несколько мотивов). В типичных анализах используют антитела или Т-клетки, специфичные по отношению к STEAP-1-родственным белкам, которые содержат аминокислотные остатки одного или нескольких биологических мотивов, входящих в состав полипептидной последовательности STEAP-1, чтобы сравнить характерные особенности данного участка в нормальной и раковой тканях или чтобы вызвать иммунный ответ на данный эпитоп. Альтернативно, STEAP-1-родственные белки, которые содержат аминокислотные остатки одного или нескольких биологических мотивов белка STEAP-1, используют для скрининга факторов, взаимодействующих с данным участком STEAP-1. Фрагменты/субпоследовательности белка STEAP-1 в особенности подходят для получения и характеристики домен-специфичных антител (например, антител, распознающих внеклеточный или внутриклеточный эпитоп белка STEAP-1), для идентификации агентов или клеточных факторов, связывающихся со STEAP-1 или его отдельным структурным доменом, а также для применения в разных терапевтических и диагностических контекстах, включающих в себя, без ограничения, диагностические анализы, противораковые вакцины и способы получения таких вакцин. Белки, кодируемые генами STEAP-1 или их аналогами, гомологами или фрагментами, имеют разные применения, включающие в себя, без ограничения, получение антител, а также способы идентификации лигандов и других агентов и клеточных компонентов, которые связываются с генным продуктом STEAP-1. Антитела, полученные против белка STEAP-1 или его фрагмента, можно использовать в диагностических и прогностических анализах, а также в методах визуализации, использующихся при лечении злокачественных заболеваний человека, характеризующихся экспрессией белка STEAP-1, таких как перечисленные в табл. I. Такие антитела, экспрессирующиеся внутри клетки, можно использовать в способах лечения пациентов с такими раковыми заболеваниями. Родственные STEAP-1 нуклеиновые кислоты или белки также используются для получения HTL- или CTL-ответов. Для детектирования белков STEAP-1 можно использовать разные биологические анализы, включающие в себя, без ограничения, разные типы радиоиммунологических анализов: твердофазный иммуноферментный анализ (ELISA), флуоресцентный иммуноферментный анализ (ELIFA), иммунохимические анализы и т.п. Антитела можно метить и использовать в качестве визуализирующих реагентов, позволяющих детектировать STEAP-1-экспрессирующие клетки (например, в радиосцинтиграфических методах визуализации). Белки STEAP-1 в особенности подходят для получения противораковых вакцин, как описано ниже в данном документе. IV. Антитела STEAP-1. В другом аспекте данное изобретение предлагает антитела, которые связывают STEAP-1-родственные белки. Предпочтительные антитела в физиологических условиях специфично связываются с белком, родственным STEAP-1, и не связываются (или слабо связываются) с пептидами или белками, отличными от белков, родственных STEAP-1. В данном контексте примеры физиологических условий включают в себя 1) забуференный фосфатом физиологический раствор; 2) забуференный Tris физиологический раствор, содержащий 25 мМ Tris и 150 мМ NaCl; или обычный физиологический раствор (0,9% NaCl); 4) животную сыворотку, например человеческую сыворотку; или 5) сочетание любых условий 1)-4); указанные реакции предпочтительно проводят при рН 7,5, альтернативно, в интервале рН от 7,0 до 8,0 или, альтернативно, в интервале рН от 6,5 до 8,5 и при температуре от 4 до 37 °С. Например, антитела, которые связывают STEAP-1, могут связывать STEAP-1-родственные белки, например, их гомологи или аналоги. Антитела STEAP-1 согласно изобретению в особенности подходят для применения в диагностике и прогнозировании злокачественных заболеваний (см., например, табл. I), а также в визуализирующих методах. Подобным образом, такие антитела можно использовать для лечения, диагностики и/или прогнозирования рака простаты и других злокачественных заболеваний, которые сопровождаются экспрессией или сверхэкспрессией STEAP-1. Более того, антитела, экспрессирующиеся внутри клеток (например, одноцепочечные антитела), можно использовать для лечения злокачественных заболеваний, которые сопровождаются экспрессией STEAP-1, таких как запущенный или метастазирующий рак простаты или другие запущенные или метастазирующие злокачественные заболевания. Данное изобретение также предлагает различные иммунологические методы для детектирования и количественного определения STEAP-1 и мутантных STEAP-1-родственных белков. Такие анализы могут включать в себя применение одного или нескольких антител STEAP-1, способных распознавать и связывать белок, родственный STEAP-1, в зависимости от обстоятельств. Данные анализы проводят в формате разных иммунологических анализов, хорошо известных в данной области, включающих в себя, без ограничения, разные типы радиоиммунологических анализов: твердофазный иммуноферментный анализ (ELISA), флуоресцентный иммуноферментный анализ (ELIFA) и т.п. Иммунологические анализы согласно изобретению, в которых не используются антитела, включают в себя анализ иммуногенности Т-клеток (ингибиторный или стимуляторный), а также анализ связывания главного комплекса гистосовместимости (МНС). Данное изобретение также предлагает способы иммунологической визуализации, позволяющие диагностировать рак простаты и другие злокачественные заболевания, сопровождающиеся экспрессией STEAP-1, данные способы включают в себя, без ограничения, радиосцинтиграфические методы визуализации с использованием меченых антител STEAP-1. Такие анализы можно использовать в клинике для диагностирования, мониторинга и прогнозирования злокачественных заболеваний, сопровождающихся экспрессией STEAP-1, таких как рак простаты. Антитела STEAP-1 также используют в способах очистки белков, родственных STEAP-1, а также в способах выделения гомологов STEAP-1 и родственных молекул. Например, способ очистки белка, родственного STEAP-1, включает в себя инкубацию антитела STEAP-1, иммобилизованного на твердом носителе, с лизатом или другим раствором, содержащим белок, родственный STEAP-1, в условиях, которые позволяют антителу STEAP-1 связываться с белком, родственным STEAP-1; промывание твердого носителя для удаления примесей и элюирование белка, родственного STEAP-1, с иммобилизованного антитела. Другие применения антител STEAP-1 в соответствии с данным изобретением включают в себя получение антиидиотипических антител, которые имитируют белок STEAP-1. В данной области известно много способов получения антител. Например, антитела можно получить путем иммунизации подходящего млекопитающего-хозяина STEAP-1-родственным белком, пептидом или фрагментом в выделенной или иммуноконъюгированной форме (Antibodies: A Laboratory Manual, CSH Press, Eds., Harlow, and Lane (1988); Harlow, Antibodies, Cold Spring Harbor Press, NY (1989)). Также можно использовать гибридные белки STEAP-1, такие как гибридный белок STEAP-1-GST. В отдельном воплощении получают гибридный белок GST, содержащий полноразмерную аминокислотную последовательность, приведенную на фиг. 2 или 3, или ее большую часть, и используют его в качестве иммуногена для получения соответствующих антител. В другом воплощении синтезируют белок, родственный STEAP-1, и используют его в качестве иммуногена. Кроме того, используют известные в данной области методы иммунизации оголенной ДНК (в присутствии или в отсутствие белка, родственного STEAP-1, или клеток, экспрессирующих STEAP-1), чтобы получить иммунный ответ на кодируемый антиген (обзор можно найти в Donnelly et al., 1997, Ann. Rev. Immunol. 15: 617-648). Аминокислотную последовательность белка STEAP-1, изображенную на фиг. 2 или 3, можно проанализировать, чтобы выбрать конкретные участки белка STEAP-1 для получения антител. Например, для идентификации гидрофильных участков в структуре STEAP-1 проводят анализы гидрофобности и гидрофильности аминокислотной последовательности STEAP-1. Участки белка STEAP-1, обладающие иммуногенной структурой, а также другие участки и домены можно легко идентифицировать с помощью других методов, известных в данной области, таких как анализы Чоу-Фасмана, Гарньер-Робсона, Кит-Дулитла, Эйсенберга, Карплус-Шультца или Джеймсон-Вульфа. Профили гидрофильности можно получить с помощью метода, описанного в Норр, Т.P. and Woods, K.R., 1981, Proc. Natl. Acad. Sci. U.S.A. 78: 3824-3828. Профили гидропатичности можно получить с помощью метода, описанного Kyte, J. and Doolittle, R.F., 1982, J. Mol. Biol. 157: 105-132. Профили процента (%) доступных остатков можно получить с помощью метода, описанного Janin J., 1979, Nature, 277: 491-492. Профили средней жесткости можно получить с помощью метода, описанного Bhaskaran R., Ponnuswamy Р.K., 1988, Int. J. Pept. Protein Res. 32: 242-255. Профили бета-складки можно получить с помощью метода, описанного Deleage, G., Roux В., 1987, Protein Engineering, 1: 289-294. Таким образом, все участки, идентифицированные с помощью любых указанных программ или методов, входят в объем согласно изобретению. Предпочтительные способы получения антител STEAP-1 дополнительно иллюстрируются с помощью примеров, описанных в данном документе. Способы получения белка или полипептида для применения в качестве иммуногена хорошо известны в данной области. В данной области также хорошо известны способы получения иммуногенных конъюгатов белка с носителем, таким как БСА, KLH или другой белок-носитель. В некоторых случаях проводят непосредственную конъюгацию с использованием, например, карбодиимидных реагентов; в других случаях эффективными являются конденсирующие реагенты, например, поставляемые Pierce Chemical Co., Rockford, IL. Введение иммуногена STEAP-1 часто осуществляют путем введения в течение подходящего периода времени и с использованием подходящего адъюванта, как известно специалистам в данной области. Степень образования антител в процессе схемы иммунизации оценивают путем определения титра антител. Моноклональные антитела STEAP-1 можно получить с помощью известных в данной области способов. Например, широко известно, что иммортализованные клеточные линии, которые секретируют целевое моноклональное антитело, получают с помощью стандартного гибридомного метода Келера и Милштейна (Kohler and Milstein) или с помощью модификаций, приводящих к иммортализации антитело-продуцирующих В-клеток. Иммортализованные клеточные линии, которые секретируют целевые антитела, подвергают скринингу с помощью иммунологического анализа, в котором антиген представляет собой белок, родственный STEAP-1. После идентификации подходящей иммортализованной клеточной культуры клеткам дают расти и продуцировать антитела либо в культурах in vitro, либо в асцитной жидкости. Антитела или фрагменты согласно изобретению также можно получить рекомбинантными способами. Участки, которые специфически связываются с целевыми участками белка STEAP-1, можно получить в составе антител с привитыми химерными или гипервариабельными (CDR) участками, относящимися к другим видам. Также можно получить гуманизированные или человеческие антитела, которые являются предпочтительными при применении в терапии. Способы гуманизации мышиных и других, отличных от человеческих антител путем замещения одного или нескольких CDR из нечеловеческих антител на соответствующие последовательности человеческих антител хорошо известны (см., например, Jones et al., 1986, Nature, 321: 522-525; Riechmann et al., 1988, Nature, 332: 323-327; Verhoeyen et al., 1988, Science, 239: 1534-1536, а также Carter et al., 1993, Proc. Natl. Acad. Sci. USA, 89: 4285 и Sims et al., 1993, J. Immunol. 151: 2296). Способы получения полностью человеческих антител включают в себя методы фаговых дисплеев и трансгенные методы (обзор см. в Vaughan et al., 1998, Nature Biotechnology, 16: 535-539). Полностью человеческие моноклональные антитела STEAP-1 можно получить с помощью методов клонирования с использованием больших комбинаторных библиотек генов человеческих Ig (т.е. фаговых дисплеев) (Griffiths and Hoogenboom, Building an in vitro immune system: human antibodies from phage display libraries. In: Protein Engineering of Antibody Molecules for Prophylactic and Therapeutic Applications in Man, Clark, M. (Ed.), Nottingham Academic, p. 45-64 (1993); Burton and Barbas, Human Antibodies from combinatorial libraries. Id., p. 65-82). Полностью человеческие моноклональные антитела STEAP-1 также можно получить с помощью трансгенных мышей, содержащих локус гена человеческого иммуноглобулина, как описано в патентной заявке РСТ WO 98/24893, Kucherlapati and Jakobovits et al., опубликованной 3 декабря 1997 г. (см. также Jakobovits, 1998, Exp. Opin. Invest. Drugs, 7(4): 607-614; патенты США 6162963, выданный 19 декабря 2000 г.; 6150584, выданный 12 ноября 2000 г.; и 6114598, выданный 5 сентября 2000 г.). Данный метод позволяет эффективно получать аутентичные человеческие антитела с высоким сродством, избегая манипуляций in vitro, необходимых в методе фаговых дисплеев. Реакционноспособность антител STEAP-1 по отношению к белку, родственному STEAP-1, можно определить с помощью ряда хорошо известных способов, включающих в себя вестерн-блоттинг, иммунопреципитацию, анализы ELISA и FACS, с использованием в зависимости от обстоятельств STEAP-1-родственные белки, STEAP-1-экспрессирующие клетки или их экстракты. Антитело STEAP-1 или его фрагмент можно пометить детектируемым маркером или конъюгировать с другой молекулой. Подходящие детектируемые маркеры включают в себя, без ограничения, радиоактивные изотопы, флуоресцентные соединения, хемилюминесцентные соединения, соединения, образующие комплексы с металлами или ферменты. Далее, биспецифические антитела, специфичные по отношению к двум или более эпитопам STEAP-1, получают с помощью методов, широко известных в данной области. Гомодимерные антитела можно получить с помощью известных в данной области методов поперечной сшивки (например, Wolff et al., Cancer Res. 53: 2560-2565). В одном воплощении данное изобретение предлагает моноклональные антитела, идентифицированные как мышиная гибридома Х92.1.30.1.1 (1) и мышиная гибридома Х120.545.1.1, помещенные на хранение в Американскую коллекцию типовых культур, 10801 University Blvd. Manassas, VA 20110-2209, 6 февраля 2004 г., с присвоением номеров доступа в АТСС РТА-5802 и РТА-5803 соответственно. V. Клеточные иммунные ответы STEAP-1. Механизм, посредством которого Т-клетки распознают антигены, известен. Эффективные вакцинные композиции согласно изобретению, содержащие пептидные эпитопы, индуцируют терапевтические или профилактические иммунные ответы в очень широких слоях населения во всем мире. Ниже приводится краткий обзор связанной с иммунологией технологии, позволяющий лучше понять значение и эффективность композиций согласно изобретению, индуцирующих клеточные иммунные ответы. Комплекс молекулы HLA и пептидного антигена действует как лиганд, распознаваемый HLA-ограниченными Т-клетками (Buus, S. et al., Cell, 47: 1071, 1986; Babbitt, В.P. et al., Nature, 317: 359, 1985; Townsend, A. and Bodmer, H., Annu. Rev. Immunol. 7: 601, 1989; Germain, R.N., Annu. Rev. Immunol. 11: 403, 1993). В результате анализа аналогов антигена с единичной аминокислотной заменой и секвенирования эндогенно связанных, естественно процессированных пептидов были идентифицированы критические остатки, относящиеся к мотивам, необходимым для специфического связывания с молекулами антигена HLA, указанные критические остатки приведены в табл. IV (см. также, например, Southwood, et al., J. Immunol. 160: 3363, 1998; Rammensee, et al., Immunogenetics, 41: 178, 1995; Rammensee et al., SYFPEITHI, доступный в Интернете на сайте URL (134.2.96.221/scripts.hiaserver.dll/home.htm); Sette, A. and Sidney, J. Curr. Opin. Immunol. 10: 478, 1998; Engelhard, V.H., Curr. Opin. Immunol. 6: 13, 1994; Sette, A. and Grey, H.M., Curr. Opin. Immunol. 4: 79, 1992; Sinigaglia, F. and Hammer, J. Curr. Biol. 6: 52, 1994; Ruppert et al., Cell. 74: 929-937, 1993; Kondo et al., J. Immunol. 155: 4307-4312, 1995; Sidney et al., J. Immunol. 157: 3480-3490, 1996; Sidney et al., Human Immunol. 45: 79-93, 1996; Sette, A. and Sidney, J. Immunogenetics, 1999, Nov.; 50 (3-4): 201-12, Review). Кроме того, с помощью рентгеноструктурного анализа комплексов HLA-пептид были обнаружены карманы в пептидсвязывающей щели/бороздке молекул HLA, которые принимают в аллель-специфичном режиме остатки пептидных лигандов; данные остатки, в свою очередь, определяют HLA-связывающую способность пептидов, в которых они находятся (см., например, Madden, D.R. Annu. Rev. Immunol. 13: 587, 1995; Smith, et al., Immunity, 4: 203, 1996; Fremont et al., Immunity, 8: 305, 1998; Stern et al., Structure, 2: 245, 1994; Jones, E.Y. Curr. Opin. Immunol. 9: 75, 1997; Brown, J.H. et al., Nature, 364: 33, 1993; Guo, H.С. et al., Proc. Natl. Acad. Sci. USA, 90: 8053, 1993; Guo, H.С. et al., Nature, 360: 364, 1992; Silver, M.L. et al., Nature, 360: 367, 1992; Matsumura, M. et al., Science, 257: 927, 1992; Madden et al., Cell, 70: 1035, 1992; Fremont, D.H. et al., Science, 257: 919, 1992; Saper, M.A., Bjorkman, P.J. and Wiley, D.C., J. Mol. Biol. 219: 277, 1991). Соответственно определение аллель-специфичных HLA-связывающих мотивов класса I и класса II или супермотивов класса I и класса II позволяет идентифицировать участки белка, отвечающие за связывание с конкретным антигеном (антигенами) HLA. Таким образом, в результате идентификации мотива HLA были идентифицированы вакцины-кандидаты на основе эпитопов; такие кандидаты можно также оценивать с помощью анализов связывания HLA-пептид, чтобы определить сродство связывания и/или период существования комплекса эпитопа с соответствующей молекулой HLA. Чтобы выбрать среди вакцин-кандидатов эпитопы с предпочтительными характеристиками, касающимися популяционного распределения и/или иммуногенности, нужно провести другие отборочные анализы. Для определения иммуногенности клеток можно использовать разные стратегии, в том числе: 1) анализ первичных Т-клеточных культур от нормальных субъектов (см., например, Wentworth, P.A. et al., Mol. Immunol. 32: 603, 1995; Celis, E. et al., Proc. Natl. Acad. Sci. USA, 91: 2105, 1994; Tsai, V. et al., J. Immunol. 158: 1796, 1997; Kawashima, I. et al., Human Immunol. 59: 1, 1998). Данная процедура включает в себя стимуляцию лимфоцитов периферической крови (PBL) от нормальных субъектов тестируемым пептидом в присутствии антигенпрезентирующих клеток in vitro в течение нескольких недель. Специфичные к пептиду Т-клетки активируются в течение указанного времени и их детектируют, например, с помощью анализа высвобождения лимфокинов или 51 Cr из клеток-мишеней, сенсибилизированных пептидом; 2) иммунизация трансгенных HLA-мышей (см., например, Wentworth, P.A. et al., J. Immunol. 26: 97, 1996; Wentworth, P.A. et al., Int. Immunol. 8: 651, 1996; Alexander, J. et al., J. Immunol. 159: 4753, 1997). Например, в таких способах пептиды вводят подкожно трансгенным HLA-мышам в неполном адъюванте Фрейнда. Через несколько недель после иммунизации спленоциты выделяют и культивируют in vitro в присутствии тестируемого пептида в течение приблизительно одной недели. Пептидспецифичные Т-клетки детектируют, например, с помощью анализа высвобождения 51 Cr, в котором участвуют клетки-мишени, сенсибилизированные пептидом, и клетки-мишени, экспрессирующие эндогенный антиген; 3) демонстрация повторных Т-клеточных ответов у иммунных субъектов, которые были эффективно вакцинированы и/или являются хронически больными пациентами (см., например, Rehermann, В. et al., J. Exp. Med. 181: 1047, 1995; Doolan, D.L. et al., Immunity 7: 97, 1997; Bertoni, R. et al., J. Clin. Invest. 100: 503, 1997; Threlkeld, S.C. et al., J. Immunol. 159: 1648, 1997; Diepolder, H.M. et al., J. Virol. 71: 6011, 1997). Соответственно повторные ответы детектируют путем культивирования PBL, полученных от субъектов, испытывающих воздействие антигена в результате заболевания, и, следовательно, генерирующих "естественный" иммунный ответ, или от пациентов, которые были вакцинированы против данного антигена. Полученные от субъектов PBL культивируют in vitro в течение 1-2 недель в присутствии тестируемого пептида и антигенпрезентирующих клеток (АРС), чтобы обеспечить активацию Т-клеток "памяти" по сравнению с "наивными" Т-клетками. В конце периода культивирования активность Т-клеток детектируют с помощью анализов, включающих в себя высвобождение 51 Cr из сенсибилизированных пептидом мишеней, пролиферацию Т-клеток и высвобождение лимфокинов. VI. STEAP-1-трансгенные животные. Нуклеиновые кислоты, кодирующие белок, родственный STEAP-1, также можно использовать для получения либо трансгенных животных, либо животных с "нокаутным" геном, которые, в свою очередь, можно использовать для разработки и скрининга терапевтических средств. В соответствии с общепринятыми методами, кДНК, кодирующую STEAP-1, можно использовать для клонирования геномной ДНК, кодирующей STEAP-1. Затем клонированные геномные последовательности можно использовать для получения трансгенных животных, содержащих клетки, которые экспрессируют ДНК, кодирующую STEAP-1. Способы получения трансгенных животных, в частности, таких как мыши или крысы, широко используются в данной области и описаны, например, в патентах США № 4736866, выданном 12 апреля 1988 г., и 4870009, выданном 26 сентября 1989 г. Как правило, для внедрения трансгена STEAP-1 вместе с тканеспецифичным энхансером используют особые клетки. Трансгенных животных, которые содержат копию трансгена, кодирующего STEAP-1, можно использовать для детектирования повышенной экспрессии ДНК, кодирующей STEAP-1. Таких животных можно использовать для тестирования реагентов, предположительно эффективных, например, против патологических состояний, связанных с повышенной экспрессией указанной ДНК. В соответствии с данным аспектом согласно изобретению уменьшение числа случаев патологического состояния у животных, получающих реагент, по сравнению с необработанными животными, несущими трансген, указывает на возможное терапевтическое воздействие на данное патологическое состояние. Альтернативно, нечеловеческие гомологи STEAP-1 можно использовать для конструирования животного с "нокаутным" геном STEAP-1, у которого ген, кодирующий STEAP-1, становится дефективным или изменяется в результате гомологичной рекомбинации между эндогенным геном, кодирующим STEAP-1, и измененной геномной ДНК, кодирующей STEAP-1, введенной в эмбриональную клетку животного. Например, кДНК, кодирующую STEAP-1, можно использовать для клонирования ДНК, кодирующей STEAP-1, в соответствии с широко используемыми в данной области методами. Фрагмент геномной ДНК, кодирующей STEAP-1, можно удалить или заменить на другой ген, такой как ген, кодирующий селектируемый маркер, используемый для отслеживания интеграции. Обычно в состав вектора включают несколько тысяч оснований неизмененной фланкирующей ДНК (как с 5'-, так и с 3'-конца) (описание гомологичных рекомбинантных векторов можно найти, например, в Thomas and Capecchi, Cell, 51: 503 (1987)). Вектор вводят в линию эмбриональных стволовых клеток (например, методом электропорации), после чего отбирают клетки, в которых произошла гомологичная рекомбинация введенной ДНК и эндогенной ДНК (см., например, Li et al., Cell, 69: 915 (1992)). Затем отобранные клетки вводят в бластоцисту животного (например, мыши или крысы) с получением агрегационных химер (см., например, Bradley, in Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E.J. Robertson, ed. (IRL, Oxford, 1987), p. 113-152). Полученный химерный эмбрион имплантируют подходящей псевдобеременной самке животного для вынашивания, после чего эмбрион считается эмбрионом животного с "нокаутным" геном. Потомство, в гаметах которого содержится ДНК с гомологичной рекомбинацией, можно идентифицировать стандартными методами и использовать для выведения животных, у которых все клетки содержат ДНК с гомологичной рекомбинацией. Животные с "нокаутным" геном могут характеризоваться, например, способностью сопротивляться некоторым патологическим состояниям или развитием патологических состояний, являющихся следствием отсутствия полипептида STEAP-1. VII. Способы детектирования STEAP-1. Другой аспект согласно изобретению относится к способам детектирования полинуклеотидов STEAP-1 и белков, родственных STEAP-1, а также к способам идентификации клеток, экспрессирующих STEAP-1. Профиль экспрессии STEAP-1 позволяет использовать его в качестве маркера для диагностики метастазирующего заболевания. Соответственно, на основе статуса генных продуктов STEAP-1 можно прогнозировать ряд факторов, в том числе возможность развития поздней стадии заболевания, скорость развития и/или агрессивность опухоли. Как описано в данном документе, статус генных продуктов STEAP-1 в образцах пациента можно анализировать с помощью ряда методов, хорошо известных в данной области и включающих в себя иммуногистохимический анализ, ряд методов нозерн-блоттинга, в том числе с использованием гибридизации in situ, анализ ОТ-ПЦР (например, на микроиссеченных образцах с лазерным улавливанием), вестерн-блоттинг и анализ тканей с использованием чипов. Более конкретно, данное изобретение предлагает анализы для детектирования полинуклеотидов STEAP-1 в биологическом образце, таком как сыворотка, кость, простата и другие ткани, моча, семенная жидкость, препараты клеток и т.п. Детектируемые полинуклеотиды STEAP-1 включают в себя, например, ген STEAP-1 или его фрагмент, мРНК STEAP-1, альтернативные сплайсинговые варианты мРНК STEAP-1 и рекомбинантные молекулы ДНК или РНК, которые содержат полинуклеотид STEAP-1. При осуществлении данного аспекта согласно изобретению можно использовать ряд способов амплификации и/или детектирования присутствия полинуклеотидов STEAP-1, хорошо известных в данной области. В одном воплощении способ детектирования мРНК STEAP-1 в биологическом образце включает в себя получение кДНК из образца путем обратной транскрипции с использованием по меньшей мере одного праймера; амплификацию полученной кДНК с использованием полинуклеотидов STEAP-1 в качестве смыслового и антисмыслового праймеров и детектирование присутствия амплифицированной кДНК STEAP-1. Необязательно можно определить последовательность амплифицированной кДНК STEAP-1. В другом воплощении способ детектирования гена STEAP-1 в биологическом образце включает в себя выделение геномной ДНК из образца; амплификацию выделенной геномной ДНК с использованием полинуклеотидов STEAP-1 в качестве смысловых и антисмысловых праймеров и детектирование присутствия амплифицированного гена STEAP-1. Из нуклеотидной последовательности STEAP-1 можно получить любое число подходящих сочетаний смысловых и антисмысловых зондов, подходящих для применения в вышеуказанном анализе (см., например, фиг. 2). Данное изобретение также предлагает анализы для детектирования присутствия белка STEAP-1 в ткани или другом биологическом образце, таком как сыворотка, семенная жидкость, кость, простата, моча, препараты клеток и т.п. Способы детектирования белка, родственного STEAP-1, также хорошо известны и включают в себя, например, иммунопреципитацию, иммуногистохимический анализ, вестерн-блоттинг, анализ молекулярного связывания, ELISA, ELIFA и т.п. Например, способ детектирования присутствия белка, родственного STEAP-1, в биологическом образце включает в себя приведение образца в контакт с антителом STEAP-1, его реакционноспособным по отношению к STEAP-1 фрагментом или рекомбинантным белком, содержащим антигенсвязывающий участок антитела STEAP-1; и затем детектирование связывания белка, родственного STEAP-1, в образце. Способы идентификации клеток, экспрессирующих STEAP-1, также входят в объем согласно изобретению. В одном воплощении способ идентификации клетки, экспрессирующей ген STEAP-1, включает в себя детектирование присутствия мРНК STEAP-1 в клетке. Способы детектирования конкретных мРНК в клетках хорошо известны и включают в себя, например, гибридизационные анализы с использованием в качестве зондов комплементарных ДНК (такие как гибридизация in situ с использованием меченых рибозондов STEAP-1, нозерн-блоттинг и родственные методы) и различные анализы с использованием амплификации нуклеиновых кислот (такие как ОТ-ПЦР с использованием комплементарных праймеров, специфичных для STEAP-1, а также другие способы детектирования на основе амплификации, такие как, например, метод многоцепьевых ДНК, SISBA, ТМА и т.п.). Альтернативно, способ идентификации клеток, экспрессирующих ген STEAP-1, включает в себя детектирование присутствия STEAP-1-родственного белка в клетке или его секреции клеткой. В данной области известно много способов детектирования белков, которые можно использовать для детектирования STEAP-1-родственных белков и клеток, экспрессирующих STEAP-1-родственные белки. Анализ экспрессии STEAP-1 также используется для идентификации и характеристики средств, модулирующих экспрессию гена STEAP-1. Например, при раке простаты и злокачественных заболеваниях тканей, перечисленных в табл. I, происходит значительное увеличение экспрессии STEAP-1. Идентификация молекул или биологических средств, ингибирующих экспрессию или сверхэкспрессию STEAP-1, в раковых клетках имеет терапевтическое значение. Например, такое средство можно идентифицировать с помощью скрининга, в котором проводят количественное определение экспрессии STEAP-1 с использованием ОТ-ПЦР, гибридизации нуклеиновых кислот или связывания антител. VIII. Способы мониторинга статуса STEAP-1-родственных генов и их продуктов. Онкогенез представляет собой многостадийный процесс, характеризующийся нарастающим нарушением клеточного роста и развитием клеток из нормального физиологического состояния до предракового и затем ракового состояния (см., например, Alers et al., Lab Invest. 77(5): (1997) и Isaacs et al., Cancer Surv. 23: 19-32 (1995)). В данном контексте анализ биологического образца на признаки нарушения клеточного роста (такие как аберрантная экспрессия STEAP-1 в раковых тканях) позволяет осуществить раннюю диагностику такого аберрантного физиологического состояния до того, как патологическое состояние, такое как рак, разовьется до стадии, при которой терапевтические средства имеют ограниченный эффект и/или которой соответствует плохой прогноз. В таких анализах статус STEAP-1 в исследуемом биологическом образце можно сравнить, например, со статусом STEAP-1 в соответствующей нормальной ткани (например, в образце, который не имеет патологических изменений и получен от того же субъекта или, альтернативно, от другого субъекта). Изменение статуса STEAP-1 в биологическом образце (по сравнению с нормальной тканью) свидетельствует о нарушении регуляции клеточного роста. Помимо применения в качестве нормального образца биологического образца, не имеющего патологических изменений, для сравнения статуса STEAP-1 в образце также можно использовать предварительно определенное нормативное значение, такое как предварительно определенный нормальный уровень экспрессии мРНК (см., например, Grever et al., J. Comp. Neurol. 1996, Dec 9; 376(2) и патент США № 5837501). Термин "статус" в данном контексте используют в соответствии с принятым в данной области значением и относят к состоянию или положению гена и его продуктов. Как правило, чтобы охарактеризовать состояние или положение гена и его продуктов, опытные специалисты используют ряд параметров. Данные параметры включают в себя, без ограничения, местоположение продуктов экспрессируемого гена (в том числе местоположение клеток, экспрессирующих STEAP-1), а также уровень и биологическую активность экспрессируемых генных продуктов (таких как мРНК, полинуклеотиды и полипептиды STEAP-1). Как правило, изменение статуса STEAP-1 включает в себя изменение местоположения STEAP-1 и/или STEAP-1-экспрессирующих клеток и/или увеличение экспрессии мРНК и/или белка STEAP-1. Статус STEAP-1 в образце можно определить с помощью ряда способов, хорошо известных в данной области и включающих в себя, без ограничения, иммуногистохимический анализ, гибридизацию in situ, анализ ОТ-ПЦР на микроиссеченных образцах с лазерным улавливанием, вестерн-блоттинг и анализ тканей с использованием чипов. Типичные способы определения статуса гена и генных продуктов STEAP-1 можно найти, например, в Ausubel et al. eds., 1995, Current Protocols In Molecular Biology, Units 2 (Northern Blotting), 4 (Southern Blotting), 15 (Immunoblotting) and 18 (PCR Analysis). Так, статус STEAP-1 в биологическом образце можно определить с помощью разных методов, используемых специалистами в данной области, которые включают в себя, без ограничения, саузерн-анализ генома (для определения, например, изменений в гене STEAP-1), нозерн-анализ и/или ПЦР-анализ мРНК STEAP-1 (для определения, например, изменений в полинуклеотидных последовательностях или уровнях экспрессии мРНК STEAP-1) и вестерн-блоттинг и/или иммуногистохимический анализ (для определения, например, изменений в полипептидных последовательностях, в местоположении полипептида в образце, в уровнях экспрессии белков STEAP-1 и/или комплексов белков STEAP-1 с полипептидсвязывающими партнерами). Детектируемые полинуклеотиды STEAP-1 включают в себя, например, ген STEAP-1 или его фрагмент, мРНК STEAP-1, варианты, образованные в результате альтернативного сплайсинга, мРНК STEAP-1 и рекомбинантные молекулы ДНК или РНК, содержащие полинуклеотид STEAP-1. Профиль экспрессии STEAP-1 позволяет использовать его в качестве диагностического маркера для локального и/или метастазирующего заболевания и предоставляет информацию о росте или онкогенном потенциале биологического образца. В частности, на основе статуса генных продуктов STEAP-1 можно прогнозировать ряд факторов, в том числе возможность развития конкретных стадий заболевания, скорость развития и/или агрессивность опухоли. Данное изобретение предлагает способы и анализы для определения статуса STEAP-1 и диагностики злокачественных заболеваний, таких как злокачественные заболевания тканей, перечисленных в табл. I. Например, поскольку мРНК STEAP-1 интенсивно экспрессируется в раковых тканях простаты, а также в других раковых тканях по сравнению с нормальной тканью простаты, анализы, позволяющие определить уровни транскриптов мРНК или белков STEAP-1 в биологическом образце, можно использовать для диагностики заболевания, связанного с нарушением регуляции STEAP-1, и получения информации, позволяющей выбрать подходящие способы лечения. Статус экспрессии STEAP-1 предоставляет информацию о присутствии, стадии и местоположении диспластических, предраковых и раковых клеток и позволяет прогнозировать возможность развития разных стадий заболевания и/или оценивать агрессивность опухоли. Более того, профиль экспрессии позволяет использовать STEAP-1 для визуализации метастазирующего заболевания. Следовательно, один аспект согласно изобретению направлен на разные методы молекулярного прогнозирования и молекулярной диагностики, позволяющие определить статус STEAP-1 в биологических образцах, таких как образцы, полученные от субъектов, которые имеют, или предположительно имеют патологию, характеризующуюся нарушением регуляции клеточного роста, такую как рак. Как описано выше, статус STEAP-1 в биологическом образце можно определить с помощью ряда хорошо известных в данной области методов. Например, статус STEAP-1 в биологическом образце, взятом из конкретной области организма, можно определить путем анализа образца на присутствие или отсутствие клеток, экспрессирующих STEAP-1 (например, клеток, экспрессирующих мРНК или белки STEAP-1). Результаты данного анализа могут свидетельствовать о нарушении регуляции клеточного роста, например, если STEAP-1-экспрессирующие клетки обнаружены в образце, который в нормальном состоянии не содержит таких клеток (таком как лимфатический узел), поскольку такие изменения статуса STEAP-1 в биологическом образце часто связаны с нарушением регуляции клеточного роста. А именно, показателем нарушения регуляции клеточного роста является метастазирование раковых клеток из органа, несущего первичную опухоль (такого как простата), в другую область организма (например, в лимфатический узел). В данном контексте нарушение регуляции клеточного роста является важной информацией, поскольку оно позволяет диагностировать скрытые метастазы в лимфатических узлах у значительной части пациентов с раком простаты, а такие метастазы связаны с известными прогнозами развития заболевания (см., например, Murphy et al., Prostate, 42(4): (2000); Su et al., Semin. Surg. Oncol. 18(1): 17-28 (2000) и Freeman et al., J. Urol. 1995 Aug. 154 (2 Pt. 1): 474-8). В одном аспекте данное изобретение предлагает способы мониторинга генных продуктов STEAP-1, экспрессируемых клетками, полученными от субъекта, предположительно имеющего заболевание, связанное с нарушением регуляции клеточного роста (такое как гиперплазия или рак), с последующим сравнением статуса, определенного с помощью данных способов, со статусом генных продуктов STEAP-1 в соответствующем нормальном образце. Присутствие аберрантных генных продуктов STEAP-1 в тестируемом образце по сравнению с нормальным образцом свидетельствует о нарушении клеточного роста в клетках субъекта. В другом аспекте изобретение предлагает анализы, использующиеся для диагностирования рака у субъекта, включающие в себя детектирование значительного повышения экспрессии мРНК или белка STEAP-1 в тестируемой клетке или в образце ткани по сравнению с уровнями экспрессии в соответствующей нормальной клетке или ткани. Присутствие мРНК STEAP-1, например, можно определить в тканях, включающих в себя, без ограничения, перечисленные в табл. I. Присутствие значительной экспрессии STEAP-1 в любой из данных тканей может указывать на возникновение, присутствие и/или тяжесть рака, поскольку соответствующие нормальные ткани не экспрессируют мРНК STEAP-1 или экспрессируют на низком уровне. В родственном воплощении статус STEAP-1 определяют чаще на уровне белка, чем на уровне нуклеиновой кислоты. Например, такой способ включает в себя определение уровня белка STEAP-1, экспрессируемого клетками в тестируемом образце ткани, и сравнение полученного уровня с уровнем STEAP-1, экспрессируемого в соответствующем нормальном образце. В одном воплощении присутствие белка STEAP-1 определяют, например, иммуногистохимическими методами. Антитела или связывающие партнеры STEAP-1, позволяющие детектировать экспрессию белка STEAP-1, применяют в ряде аналитических форматов, широко использующихся в данной области для указанной цели. В следующем воплощении предлагается способ определения статуса нуклеотидных и аминокислотных последовательностей STEAP-1 в биологическом образце с целью идентификации изменений в структуре данных молекул. Указанные изменения могут включать в себя вставки, делеции, замены и т.п. Такие анализы имеют большое значение, поскольку изменения в нуклеотидных и аминокислотных последовательностях наблюдаются в большом числе белков, связанных с фенотипом, сопутствующим нарушению клеточного роста (см., например, Marrogi et al., 1999, J. Cutan. Pathol. 26(8): 369-378). Например, мутация в последовательности STEAP-1 может указывать на присутствие фактора, стимулирующего развитие опухоли. Следовательно, такие анализы имеют диагностическое и прогностическое значение, если мутация в STEAP-1 указывает на возможную утрату функции или повышение опухолевого роста. В данной области известен широкий ряд анализов, позволяющих детектировать изменения в нуклеотидных и аминокислотных последовательностях. Например, размер и структуру нуклеотидных или аминокислотных последовательностей генных продуктов STEAP-1 можно определить с помощью нозерн-, саузерн-, вестерн-блоттингов, методов ПЦР и секвенирования ДНК, описанных в данном документе. Кроме того, в данной области хорошо известны другие методы, позволяющие определять изменения в нуклеотидных и аминокислотных последовательностях, такие как анализ полиморфизма одноцепочечной конформации (см., например, патенты США № 5382510, выданный 7 сентября 1999 г., и 5952170, выданный 17 января 1995 г.). Кроме того, можно определить статус метилирования гена STEAP-1 в биологическом образце. Аберрантные деметилирование и/или гиперметилирование островков CpG в 5'-регуляторных участках гена часто встречаются в иммортализованных и трансформированных клетках и могут приводить к изменению экспрессии разных генов. Например, гиперметилирование промотора глутатион-S-трансферазы pi-класса (белка, экспрессирующегося в ткани нормальной простаты, но не экспрессирующегося в > 90% тканей карциномы простаты) приводит к постоянно "молчащей" транскрипции данного гена и является наиболее часто детектируемым геномным изменением при карциноме простаты (De Marzo et al., Am. J. Pathol. 155(6): 1985-1992 (1999)). Данное изменение также присутствует по меньшей мере в 70% случаев простатической внутриэпителиальной неоплазии (PIN) с высокой степенью злокачественности (Brooks et al., Cancer Epidemiol. Biomarkers Prev., 1998, 7: 531-536). В другом примере экспрессия опухолеспецифичного гена LAGE-I (который не экспрессируется в нормальной простате, но экспрессируется в 25-50% случаев рака простаты) индуцируется дезоксиазацитидином в лимфобластоидных клетках, это позволяет предположить, что опухолевая экспрессия является следствием деметилирования (Lethe et al., Int. J. Cancer, 76(6): 903-908 (1998)). В данной области широко известен ряд анализов для определения статуса метилирования. Например, статус метилирования островков CpG можно определить методом саузерн-гибридизации с использованием рестриктаз, чувствительных к метилированию, которые не могут расщеплять последовательности, содержащие метилированные участки CpG. Кроме того, с помощью MSP (ПЦР, специфичная для метилирования) можно быстро определить статус метилирования всех участков CpG, присутствующих в островке CpG конкретного гена. Данный метод включает в себя модификацию ДНК бисульфитом натрия (который превращает все неметилированные остатки цитозина в остатки урацила) с последующей модификацией с использованием праймеров, специфичных для метилированной, но не для неметилированной ДНК. Методы, включающие в себя воздействие метилированием, можно найти, например, в Current Protocols In Molecular Biology, Unit 12, Frederick M. Ausubel et al., eds., 1995. Амплификация гена является другим способом определения статуса STEAP-1. Амплификацию гена измеряют непосредственно в образце, например, с помощью традиционного саузерн-блоттинга или нозерн-блоттинга путем количественного определения транскрипции мРНК (Thomas, 1980, Proc. Natl. Acad. Sci. USA, 77: 5201-5205), дот-блоттинга (анализ ДНК) или гибридизации in situ с использованием соответственно меченого зонда на основе описанных здесь последовательностей. Альтернативно можно использовать антитела, которые распознают конкретные двойные спирали, в том числе двойные спирали ДНК, двойные спирали РНК и двойные спирали гибридов ДНК-РНК или двойные спирали комплексов ДНК-белок. Данный анализ проводят с использованием меченых антител и при условии, что двойная спираль связана с поверхностью так, чтобы при образовании двойной спирали на поверхности можно было детектировать антитело, связанное с двойной спиралью. Образцы ткани или периферической крови можно анализировать на присутствие раковых клеток путем детектирования экспрессии STEAP-1 с помощью традиционных методов, таких как нозерн-блоттинг, дот-блоттинг или анализ ОТ-ПЦР. Присутствие амплифицируемой методом ОТ-ПЦР мРНК STEAP-1 свидетельствует о наличии злокачественного заболевания. Анализы ОТ-ПЦР хорошо известны в данной области. В настоящее время изучается возможность применения детекционных анализов опухолевых клеток в периферической крови с использованием ОТ-ПЦР для диагностики и контролирования ряда солидных опухолей человека. В области рака предстательной железы они включают в себя анализы ОТ-ПЦР для детектирования клеток, экспрессирующих PSA и PSM (Verkaik et al., 1997, Urol. Res. 25: 373-384; Ghossein et al., 1995, J. Clin. Oncol. 13:1195-2000; Heston et al., 1995, Clin. Chem. 41: 1687-1688). Следующий аспект согласно изобретению заключается в определении предрасположенности субъекта к развитию злокачественного заболевания. В одном воплощении способ прогнозирования предрасположенности к злокачественному заболеванию включает в себя детектирование мРНК STEAP-1 или белка STEAP-1 в образце ткакни, их присутствие указывает на предрасположенность к злокачественному заболеванию, а степень экспрессии мРНК STEAP-1 коррелирует со степенью предрасположенности. В конкретном воплощении определяют присутствие STEAP-1 в простате или в другой ткани, причем присутствие STEAP-1 в образце свидетельствует о наличии предрасположенности к раку простаты (либо о возникновении или существовании опухоли простаты). Подобным образом можно определить правильность нуклеотидных и аминокислотных последовательностей STEAP-1 в биологическом образце с целью идентификации изменений в структуре данных молекул, таких как вставки, делеции, замены и т.п. Присутствие одного или нескольких изменений в генных продуктах STEAP-1 образца свидетельствует о предрасположенности к злокачественному заболеванию (либо о возникновении или существовании опухоли простаты). Данное изобретение также включает в себя способы оценки агрессивности опухоли. В одном воплощении способ оценки агрессивности опухоли включает в себя определение уровня мРНК STEAP-1 или белка STEAP-1, экспрессируемых опухолевыми клетками, сравнение полученного уровня с уровнем мРНК STEAP-1 или белка STEAP-1, экспрессируемых в соответствующей нормальной ткани, взятой у того же субъекта, или в нормальной ткани эталонного образца, где степень экспрессии мРНК STEAP-1 или белка STEAP-1 в опухолевом образце по сравнению с нормальным образцом соответствует степени агрессивности. В конкретном воплощении агрессивность опухоли оценивают путем определения степени экспрессии STEAP-1 в опухолевых клетках, причем более высокие уровни экспрессии соответствуют более агрессивным опухолям. Другое воплощение относится к оценке правильности нуклеотидных и аминокислотных последовательностей STEAP-1 в биологическом образце с целью идентификации изменений в структуре данных молекул, таких как вставки, делеции, замены и т.п. Присутствие одного или нескольких изменений свидетельствует о наличии более агрессивной опухоли. Другое воплощение согласно изобретению направлено на способы наблюдения за развитием злокачественного заболевания у субъекта с течением времени. В одном воплощении способы наблюдения за развитием злокачественного заболевания у субъекта с течением времени включают в себя определение уровня мРНК STEAP-1 или белка STEAP-1, экспрессируемых опухолевыми клетками, сравнение полученного уровня с уровнем мРНК STEAP-1 или белка STEAP-1, экспрессируемых в образце равнозначной ткани, взятой у того же субъекта в другое время, где изменение степени экспрессии мРНК STEAP-1 или белка STEAP-1 в опухолевом образце с течением времени предоставляет информацию о развитии злокачественного заболевания. В отдельном воплощении развитие злокачественного заболевания оценивают путем определения степени экспрессии STEAP-1 в опухолевых клетках в разное время, причем более высокие уровни экспрессии свидетельствуют о развитии злокачественного заболевания. Кроме того, можно оцененить правильность нуклеотидных и аминокислотных последовательностей STEAP-1 в биологическом образце с целью идентификации изменений в структуре данных молекул, таких как вставки, делеции, замены и т.п., где присутствие одного или нескольких изменений свидетельствует о развитии злокачественного заболевания. Описанные выше диагностические способы можно объединить с любым из широкого ряда прогностических и диагностических методов, известных в данной области. Например, другое воплощение согласно изобретению направлено на способы наблюдения за соответствием между экспрессией гена STEAP-1 и генных продуктов STEAP-1 (или изменений в гене STEAP-1 и генных продуктах STEAP-1) и фактором, связанным со злокачественным заболеванием, как на средства диагностики и прогнозирования статуса образца ткани. Можно использовать широкий ряд факторов, связанных со злокачественным заболеванием, таких как экспрессия генов, ассоциированных со злокачественным заболеванием (например, экспрессия PSA, PSCA и PSM при раке простаты и др.), а также обширные цитологические наблюдения (см., например, Bocking et al., 1984, Anal. Quant. Cytol. 6(2): 74-88; Epstein, 1995, Hum. Pathol. 26(2): 223-9; Thorson et al., 1998, Mod. Pathol. 11(6): 543-51; Baisden et al., 1999, Am. J. Surg. Pathol. 23(8): 918-24). Способы наблюдения за соответствием между экспрессией гена STEAP-1 и генных продуктов STEAP-1 (или изменений в гене STEAP-1 и генных продуктах STEAP-1) и фактором, связанным со злокачественным заболеванием, являются полезными, например, поскольку присутствие ряда факторов, сопутствующих заболеванию, предоставляет важную информацию для диагностики и прогнозирования статуса образца ткани. В одном воплощении способы наблюдения за соответствием между экспрессией гена STEAP-1 и генных продуктов STEAP-1 (или изменений в гене STEAP-1 и генных продуктах STEAP-1) и фактором, связанным со злокачественным заболеванием, включают в себя детектирование сверхэкспрессии мРНК или белка STEAP-1 в образце ткани, детектирование сверхэкспрессии мРНК или белка PSA в образце ткани (или экспрессии PSCA или PSM) и наблюдение за соответствием сверхэкспрессии мРНК или белка STEAP-1 либо мРНК или белка PSA (или экспрессии PSCA или PSM). В отдельном воплощении определяют экспрессию мРНК STEAP-1 и PSA в ткани простаты, где соответствие сверхэкспрессии мРНК STEAP-1 и PSA в образце указывает на существование рака простаты, на предрасположенность к раку простаты и на появление или на статус опухоли простаты. Способы детектирования и количественного определения экспрессии мРНК или белка STEAP-1 мРНК описаны в данном документе, а стандартные методы детектирования и количественного определения нуклеиновых кислот и белков хорошо известны в данной области. Стандартные методы детектирования и количественного определения мРНК STEAP-1 включают в себя гибридизацию in situ с использованием меченых рибозондов STEAP-1, нозерн-блоттинг и родственные методы с использованием в качестве зондов полинуклеотидов STEAP-1, анализ ОТ-ПЦР с использованием праймеров, специфичных для STEAP-1, а также другие методы детектирования на основе амплификации, такие как, например, метод многоцепьевых ДНК, SISBA, ТМА и т.п. В отдельном воплощении для детектирования и количественного определения экспрессии мРНК STEAP-1 используют полуколичественный метод ОТ-ПЦР. В данном методе можно использовать любое число праймеров, способных обеспечивать амплификацию STEAP-1, которые включают в себя, без ограничения, разные наборы праймеров, конкретно описанные в данном документе. В другом воплощении в иммуногистохимическом анализе образца ткани для биопсии можно использовать поликлональные или моноклональные антитела, специфически взаимодействующие с белком STEAP-1 дикого типа. IX. Идентификация молекул, взаимодействующих со STEAP-1. С помощью белковых и нуклеотидных последовательностей STEAP-1, раскрытых в данном документе, специалист в данной области может идентифицировать белки, маленькие молекулы и другие реагенты, взаимодействующие со STEAP-1, а также пути, активируемые STEAP-1, с помощью любого из используемых в данной области методов. Например, можно использовать одну из так называемых систем улавливания взаимодействия (также называемых "двухгибридный анализ"). В таких системах молекулы подвергают взаимодействию с фактором транскрипции, который управляет экспрессией репортерного гена, с преобразованием фактора транскрипции, после чего анализируют экспрессию репортерного гена. Другие системы позволяют идентифицировать белок-белковые взаимодействия in vivo посредством преобразования эукариотного активатора транскрипции (см., например, патенты США № 5955280, выданный 21 сентября 1999 г., 5925523, выданный 20 июля 1999 г., 5846722, выданный 8 декабря 1998 г., и 6004746, выданный 21 декабря 1999 г.). Алгоритмы прогнозирования белковой функции на основе структуры генома также доступны в данной области (см., например, Marcotte, et al., Nature, 402: 4, November, 1999, 83-86). Альтернативно, молекулы, взаимодействующие с белковыми последовательностями STEAP-1, можно идентифицировать путем скрининга пептдных библиотек. В таких способах пептиды, которые связываются со STEAP-1, идентифицируют путем скрининга библиотек, кодирующих статистическое или упорядоченное расположение аминокислот. Пептиды, кодируемые библиотеками, экспрессируют в виде гибридов с белками оболочки бактериофагов, после чего бактериофаговые частицы подвергают скринингу против белка (белков) STEAP-1. Соответственно, пептиды, имеющие широкий ряд применений, таких как применения в качестве терапевтических, прогностических или диагностических средств, идентифицируют таким образом без какой-либо предварительной информации о структуре предполагаемого лиганда или рецепторной молекулы. Типичные пептидные библиотеки и способы скрининга, которые можно использовать для идентификации молекул, взаимодействующих с белковыми последовательностями STEAP-1, раскрыты, например, в патентах США № 5723286, выданном 3 марта 1998 г., и 5733731, выданном 31 марта 1998 г. Альтернативно, для идентификации белок-белковых взаимодействий, опосредованных STEAP-1, можно использовать клеточные линии, экспрессирующие STEAP-1. Такие взаимодействия можно исследовать с помощью методов иммунопреципитации (см., например, Hamilton B.J., et al. Biochem. Biophys. Res. Commun. 1999, 261: 646-51). Белок STEAP-1 из клеточных линий, экспрессирующих STEAP-1, может участвовать в реакциях иммунопреципитации с антителами против STEAP-1. Альтернативно, при применении клеточной линии, созданной для экспрессии гибридов STEAP-1 и His-маркера (векторы указаны выше), можно использовать антитела против His-маркера. Ассоциацию белков в преципитированном иммунокомплексе можно анализировать с помощью таких методов, как вестерн-блоттинг, мечение белков 35 S-метионином, микросеквенирование белков, окрашивание серебром и двухмерный гель-электрофорез. Маленькие молекулы и лиганды, взаимодействующие со STEAP-1, можно идентифицировать с помощью родственных воплощений таких скрининговых анализов. Например, можно идентифицировать маленькие молекулы, которые препятствуют выполнению белковой функции, в том числе молекулы, которые препятствуют способности STEAP-1 опосредовать фосфорилирование и дефосфорилирование, взаимодействовать с молекулами ДНК или РНК, что может служить показателем регуляции клеточного цикла, участвовать в передаче сигнала, опосредованной вторичными мессенджерами, или в онкогенезе. Подобным образом, маленькие молекулы, модулирующие STEAP-1-родственный ионный канал, белковый насос или функции клеточной коммуникации, идентифицируют и используют для лечения пациентов с злокачественным заболеванием, сопровождающимся экспрессией STEAP-1 (см., например, Hille, В., Ionic Channels of Excitable Membranes, 2 nd Ed., Sinauer Assoc., Sunderland, MA, 1992). Кроме того, лиганды, регулирующие функцию STEAP-1, можно идентифицировать по их способности связывать STEAP-1 и активировать репортерную конструкцию. Типичные способы описаны, например, в патенте США № 5928868, выданном 27 июля 1999 г., и включают в себя способы получения гибридных лигандов, в которых по меньшей мере один лиганд является маленькой молекулой. В иллюстративном воплощении клетки, созданные для экспрессии гибридного белка STEAP-1 и ДНК-связывающего белка, используют для совместной экспрессии гибридного белка гибридный лиганд/маленькая молекула и белка, активирующего транскрипцию библиотеки кДНК. Данные клетки также содержат репортерный ген, экспрессия которого ограничена степенью приближения первого и второго гибридных белков друг к другу и происходит только в том случае, если гибридный лиганд связывается с участками-мишенями на обоих гибридных белках. Клетки, экспрессирующие репортерный ген, отбирают и идентифицируют неизвестную маленькую молекулу или неизвестный лиганд. Данный способ позволяет идентифицировать модуляторы, которые активируют или ингибируют STEAP-1. Воплощение согласно изобретению включает в себя способ скрининга с целью выявления молекулы, взаимодействующей с аминокислотной последовательностью STEAP-1, изображенной на фиг. 2 или 3, включающий в себя следующие стадии: приведение в контакт популяции тестируемых молекул с аминокислотной последовательностью STEAP-1; взаимодействие популяции молекул с аминокислотной последовательностью STEAP-1 в условиях, способствующих взаимодействию; определение присутствия молекулы, взаимодействующей с аминокислотной последовательностью STEAP-1, и затем разделение молекул, которые не взаимодействуют с аминокислотной последовательностью STEAP-1, от молекул, взаимодействующих с данной последовательностью. В отдельном воплощении данный способ дополнительно включает в себя очистку, характеризацию и идентификацию молекулы, которая взаимодействует с аминокислотной последовательностью STEAP-1. Идентифицированную молекулу можно использовать для модуляции функции, выполняемой STEAP-1. В предпочтительном воплощении аминокислотную последовательность STEAP-1 приводят в контакт с библиотекой пептидов. X. Терапевтические способы и композиции. Идентификация STEAP-1 как белка, который в норме экспрессируется в ограниченном диапазоне тканей, но который также экспрессируется в раковых тканях, таких как перечисленные в табл. I, открывает ряд терапевтических подходов к лечению указанных злокачественных заболеваний. Следует отметить, что направленную на определенную мишень противоопухолевую терапию можно использовать, даже если белок-мишень экспрессируется в нормальных тканях, в том числе и в нормальных тканях жизненно важных органов. Жизненно важным органом считается орган, необходимый для поддержания жизни, такой как сердце или прямая кишка. Органом, не имеющим жизненно важных функций, считается орган, после удаления которого субъект продолжает жить. Примерами органов, не имеющих жизненно важных функций, являются яичники, молочная железа и простата. Например, Herceptin ® представляет собой фармацевтическое средство, утвержденное Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) США, которое содержит антитело, иммунореактивное по отношению к белку, известному под разными наименованиями, а именно HER2, HER2/neu и erb-b-2. Он поставляется Genentech и является коммерчески успешным противоопухолевым средством. Доход от продажи Herceptin ® в 2002 г. составил почти 400 миллионов долларов. Herceptin ® представляет собой лекарственное средство для лечения HER2-положительного метастазирующего рака простаты. Однако экспрессия HER2 не ограничивается такими опухолями. Данный белок экспрессируется в ряде нормальных тканей. В частности, HER2/neu обнаружен в нормальных тканях почек и сердца, которые присутствуют у всех людей, принимающих Herceptin. Присутствие HER2/neu в нормальных почках также подтверждают Latif, Z., et al., B.J.U. International (2002), 89: 5-9. Как показано в данной статье (в которой исследуется, действительно ли почечно-клеточная карцинома является предпочтительным показанием для применения антител против HER2, таких как Herceptin), в доброкачественных тканях почек продуцируются и белок, и мРНК. Примечательно, что в доброкачественной ткани почек обнаружена интенсивная сверхэкспрессия белка HER2/neu. Несмотря на то что HER2/neu экспрессируется в таких жизненно важных тканях, как ткани сердца и почек, Herceptin является очень полезным, утвержденным FDA и коммерчески успешным лекарственным средством. Влияние Herceptin на ткани сердца, т.е. "кардиотоксичность", является просто побочным эффектом лечения. Если пациенты принимают только Herceptin, существенная кардиотоксичность наблюдается лишь у очень небольшой части пациентов. Для минимизации кардиотоксичности ужесточают требования, касающиеся лечения с использованием HER2/neu. Перед лечением оценивают такие факторы, как предрасположенность к заболеваниям сердца. Следует особо отметить, что, несмотря на то что в тканях почек наблюдается нормальная экспрессия, возможно, даже более высокая, чем в тканях сердца, никакие побочные эффекты со стороны Herceptin в почках не обнаружены. Более того, существует очень маленькая вероятность какого-либо побочного эффекта в других нормальных тканях, экспрессирующих HER2. Вообще, все обнаруженные побочные эффекты относятся только к тканям сердца. Такая ткань, как ткань почек, где экспрессия HER2/neu является особенно значительной, не является платформой для возникновения какого-либо побочного эффекта. Далее, благоприятные терапевтические эффекты наблюдаются при использовании противоопухолевой терапии, направленной на рецептор эпидермального фактора роста (EGFR), Erbitux (ImClone). EGFR также экспрессируется в ряде нормальных тканей. При применении терапевтических средств, направленных против EGFR, существуют очень ограниченные побочные эффекты в нормальных тканях. Общим побочным эффектом лечения, направленного против EGFR, является сильная кожная сыпь, наблюдающаяся у 100% пациентов, подвергающихся лечению. Таким образом, экспрессия целевого белка в нормальной ткани, даже в жизненно важной нормальной ткани, не является причиной для отказа от применения средства, направленного против данного белка, при лечении некоторых опухолей, в которых также происходит сверхэкспрессия указанного белка. Например, экспрессия в жизненно важных органах сама по себе не является вредной. Кроме того, органы, не являющиеся жизненно необходимыми, такие как простата и яичники, можно удалить, не создавая угрозы для жизни. Наконец, экспрессия в некоторых жизненно важных нормальных органах не вносит вклад в эффект препарата по причине иммунологической привилегии данных органов. Иммунопривилегированными органами являются органы, которые защищены от некоторых переносимых кровью веществ гематоорганным барьером и, следовательно, не доступны для иммунотерапии. Примерами иммунопривилегированных органов являются мозг и яички. Соответственно, терапевтические способы, направленные на ингибирование активности белка STEAP-1, можно использовать для лечения пациентов, страдающих от рака, сопровождающегося экспрессией STEAP-1. Данные терапевтические способы обычно делят на три класса. Первый класс включает в себя модулирование функции STEAP-1, связанной с опухолевым клеточным ростом, которое приводит к подавлению или замедлению опухолевого клеточного роста или вызывает его прекращение. Второй класс включает в себя разные способы ингибирования связывания или ассоциации белка STEAP-1 с его партнером по связыванию или с другими белками. Третий класс включает в себя ряд способов ингибирования транскрипции гена STEAP-1 или трансляции мРНК STEAP-1. Х.А. Противораковые вакцины. Данное изобретение предлагает противораковые вакцины, содержащие белок, родственный STEAP-1, или нуклеиновую кислоту, родственную STEAP-1. С точки зрения экспрессии STEAP-1, с помощью противораковых вакцин можно предотвращать и/или лечить злокачественные заболевания, сопровождающиеся экспрессией STEAP-1, при этом эффект, оказываемый данными вакцинами на другие органы, является минимальным или отсутствует. Применение в качестве противораковой терапии вакцины, содержащей опухолевый антиген, который вызывает клеточный или гуморальный иммунные ответы, хорошо известно в данной области и используется для лечения рака простаты с применением в качестве иммуногенов человеческого PSMA и РАР грызунов (Hodge et al., 1995, Int. J. Cancer, 63: 231-237; Fong et al., 1997, J. Immunol. 159: 3113-3117). Такие способы можно легко осуществить с использованием белка, родственного STEAP-1, молекулы нуклеиновой кислоты, кодирующей STEAP-1, и рекомбинантных векторов, обеспечивающих экспрессию и презентирование иммуногена STEAP-1 (который обычно содержит ряд Т-клеточных эпитопов или антитело). В данной области известен широкий ряд вакцинных систем для доставки иммунореактивных эпитопов (см., например, Heryln et al., Ann Med. 1999 Feb. 31(1): 66-78; Maruyama et al., Cancer Immunol. Immunother, 2000 Jun. 49(3): 123-32). Коротко говоря, такие способы индуцирования иммунного ответа (например, клеточного и/или гуморального) у млекопитающего включают в себя стадии воздействия на иммунную систему млекопитающего иммунореактивного эпитопа (например, эпитопа, присутствующего в белке STEAP-1, изображенном на фиг. 3, или в его аналоге или гомологе) с индуцированием у млекопитающего иммунного ответа, специфичного для данного эпитопа (например, с образованием антител, которые специфично распознают данный эпитоп). В предпочтительном способе иммуноген STEAP-1 содержит биологический мотив, см., например, табл. V-XVIII и XXII-LI, или пептид, размер которого варьирует в соответствии со STEAP-1, изображенным на фиг. 5-9. Полноразмерный белок STEAP-1, его иммуногенные участки или эпитопы можно объединять и доставлять разными способами. Такие вакцинные композиции могут включать в себя, например, липопептиды (например, Vitiello, A. et al., J. Clin. Invest. 95: 341, 1995), пептидные композиции, заключенные в полусферах из поли(DL-лактид-со-гликолида) (см., например, Eldridge, et al., Molec. Immunol. 28: 287-294, 1991; Alonso et al., Vaccine, 12: 299-306, 1994; Jones et al., Vaccine, 13: 675-681, 1995), пептидные композиции в составе иммуностимулирующих комплексов (ISCOMS) (см., например, Takahashi et al., Nature 344: 873-875, 1990; Hu et al., Clin. Exp. Immunol. 113: 235-243, 1998), совокупные антиген-пептидные системы (MAP) (см., например, Tam, J.P., Proc. Natl. Acad. Sci. U.S.A. 85: 5409-5413, 1988; Tam, J.P., J. Immunol. Methods? 196: 17-32, 1996), композиции, содержащие поливалентные пептиды; пептиды для применения в баллистических системах доставки, обычные кристаллические пептиды, вирусные векторы для доставки (Perkus, M.E. et al. In: Concepts in vaccine development, Kaufmann, S.H.E., ed., p. 379, 1996; Chakrabarti, S. et al. Nature, 320: 535, 1986; Hu, S.L. et al. Nature, 320: 537, 1986; Kieny, M.-P. et al. AIDS Bio/Technology, 4: 790, 1986; Top, F.H. et al. J. Infect. Dis. 124: 148, 1971; Chanda, P.K. et al. Virology, 175: 535, 1990), частицы вирусного или синтетического происхождения (например, Kofler, N. et al., J. Immunol. Methods, 192: 25, 1996; Eldridge, J.H. et al., Sem. Hematol. 30: 16, 1993; Falo, L.D., Jr. et al., Nature Med. 7: 649, 1995), адъюванты (Warren, H.S., Vogel, F.R., and Chedid, L.A. Annu. Rev. Immunol. 4: 369, 1986; Gupta, R.K. et al., Vaccine, 11: 293, 1993), липосомы (Reddy, R. et al., J. Immunol. 148: 1585, 1992; Rock, K.L., Immunol. Today, 17: 131, 1996) либо оголенную или адсорбированную на частицах кДНК (Ulmer, J.В. et al., Science, 259: 1745, 1993; Robinson, H.L., Hunt, L.A., and Webster, R.G., Vaccine, 11: 957, 1993; Shiver, J.W. et al., In: Concepts in vaccine development, Kaufmann, S.H.E., ed., p. 423, 1996; Cease, K.B., and Berzofsky, J.A., Annu. Rev. Immunol. 12: 923, 1994 и Eldridge, J.H. et al., Sem. Hematol. 30: 16, 1993). Также можно использовать токсин-направленные методы доставки, также известные как рецептор-опосредованные направленные методы, такие как поставляемые Avant Immunotherapeutics, Inc. (Needham, Massachusetts). В случае пациентов со STEAP-1-ассоциированным раком вакцинные композиции согласно изобретению также можно использовать в сочетании с другими противораковыми способами лечения, такими как хирургия, химиотерапия, медикаментозная терапия, лучевая терапия и др., включая применение в сочетании с иммунными вспомогательными средствами, такими как IL-2, IL-12, GM-CSF и т.п. Клеточные вакцины. Эпитопы CTL можно анализировать с помощью определенных алгоритмов с целью идентификации в белке STEAP-1 пептидов, которые связывают соответствующие аллели HLA (см., например, табл. IV; Epimer ™ и Epimatrix ™, Brown University (URL brown.edu/Research/TB-HIV_Lab/epimatrix/epimatrix.html) и BIMAS, (URL bimas.dcrt.nih.gov/; SYFPEITHI на сайте URL syfpeithi.bmi-heidelberg.com/). В предпочтительном воплощении иммуноген STEAP-1 содержит одну или несколько аминокислотных последовательностей, идентифицированных с помощью хорошо известных в данной области способов, таких как последовательности, приведенные в табл. V-XVIII и XXII-LI, или пептид из 8, 9, 10 или 11 аминокислот, определенный как мотив/супермотив HLA класса I (например, табл. IV(А), IV(D) или IV(Е)), и/или пептид, состоящий по меньшей мере из 9 аминокислот, который содержит мотив/супермотив HLA класса II (например, табл. IV(В) или IV(С)). Как известно в данной области, бороздка HLA класса I имеет, по существу, закрытые концы, поэтому только пептиды определенного размера могут плотно прилегать к бороздке и связываться с ней, как правило, эпитопы HLA класса I имеют длину 8, 9, 10 или 11 аминокислот. Наоборот, связывающая бороздка HLA класса II имеет, по существу, открытые концы; следовательно, с молекулой HLA класса II может связываться пептид, состоящий из 9 или более аминокислот. Вследствие различия в связывающих бороздках HLA классов I и II мотивы HLA класса I специфичны к длине, т.е. второе положение в мотиве класса I соответствует второй аминокислоте в направлении от аминоконца к карбоксильному концу пептида. Аминокислотные положения в мотиве класса II соответствуют только друг другу, но не всему пептиду, т.е. к амино- и/или карбоксильному концу мотив-несущей последовательности могут быть присоединены другие аминокислоты. Длина эпитопов HLA класса II может составлять 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислот или более. В данной области известен широкий ряд способов получения иммунного ответа у млекопитающего (например, первая стадия при получении гибридом). Способы получения иммунного ответа у млекопитающего включают в себя воздействие иммуногенного эпитопа, содержащегося в белке (например, в белке STEAP-1), на иммунную систему млекопитающего так, что генерируется иммунный ответ. Типичное воплощение включает в себя способ индуцирования у хозяина иммунного ответа на STEAP-1 путем приведения в контакт хозяина с достаточным количеством, по меньшей мере одного, эпитопа STEAP-1 или его аналога, специфичного для В-клеток или цитотоксичных Т-клеток; и по меньшей мере один периодический интервал после повторного приведения в контакт хозяина с эпитопом STEAP-1 или его аналога, специфичным для В-клеток или цитотоксичных Т-клеток. Конкретное воплощение относится к способу индуцирования иммунного ответа против белка, родственного STEAP-1, или искусственного полиэпитопного пептида, который включает в себя введение человеку или другому млекопитающему иммуногена STEAP-1 (например, белка STEAP-1 или его пептидного фрагмента, гибридного белка STEAP-1, его аналога и др.) в составе вакцинного препарата. Обычно такие вакцинные препараты дополнительно содержат подходящий адъювант (см., например, патент США № 6146635) или универсальный хелперный эпитоп, такой как пептид PADRE ™ (Epimmune Inc., San Diego, CA; см., например, Alexander et al., J. Immunol. 2000; 164(3): 1625-1633; Alexander et al., Immunity, 1994, 1(9): 751-761 и Alexander et al., Immunol. Res. 1998, 18(2): 79-92). Альтернативный способ включает в себя индуцирование у субъекта иммунного ответа против иммуногена STEAP-1 путем введения in vivo в мышцу или кожу организма субъекта молекулы ДНК, которая содержит последовательность ДНК, кодирующую иммуноген STEAP-1, последовательность ДНК, функционально связанную с регуляторной последовательностью, которая контролирует экспрессию последовательности ДНК, где молекула ДНК поглощается клетками, затем последовательность ДНК экспрессируется в клетках и индуцируется иммунный ответ против иммуногена (см., например, патент США № 5962428). Также можно ввести средство, способствующее действию генетической вакцины, такое как анионные липиды; сапорины; лектины; эстрогенные соединения; гидроксилированные низшие алкилы; диметилсульфоксид; и мочевину. Кроме того, чтобы индуцировать ответ на целевой антиген, можно ввести антиидиотипическое антитело, которое имитирует STEAP-1. Вакцины на основе нуклеиновых кислот. Вакцинные композиции согласно изобретению включают в себя композиции на основе нуклеиновых кислот. Пациенту можно вводить ДНК или РНК, которые кодируют белок (белки) согласно изобретению. Методы генетической иммунизации можно использовать для индуцирования гуморального и клеточного иммунных ответов против раковых клеток, экспрессирующих STEAP-1, с профилактической или терапевтической целью. Конструкции, которые содержат ДНК, кодирующую STEAP-1-родственный белок/иммуноген, а также подходящие регуляторные последовательности, можно вводить непосредственно в мышцу или кожу субъекта так, чтобы клетки мышцы или кожи поглощали конструкцию и экспрессировали кодируемый ею белок STEAP-1/иммуноген. Альтернативно, вакцина содержит белок, родственный STEAP-1. Экспрессия иммуногенного STEAP-1-родственного белка приводит к индуцированию профилактических или терапевтических гуморального и клеточного ответов против клеток, несущих белок STEAP-1. Можно использовать разные, известные в данной области методы профилактической и терапевтической генетической иммунизации (информацию и ссылки, позволяющие сделать обзор по данной теме, можно найти на Интернет-сайте genweb.com). Доставка с использованием нуклеиновых кислот описана, например, в Wolff et. al., Science, 247: 1465 (1990), а также в патентах США № 5580859; 5589466; 5804566; 5739118; 5736524; 5679647; WO 98/04720. Примеры способов доставки с использованием ДНК включают в себя "метод оголенной ДНК", облегченную (опосредованную бупивикаином, полимерами, пептидами) доставку, метод с применением катионных липидных комплексов и метод с применением частиц ("метод генной пушки") или доставку под давлением (см., например, патент США № 5922687). С целью терапевтической или профилактической иммунизации белки согласно изобретению можно экспрессировать с помощью вирусных или бактериальных векторов. Вирусные системы доставки генов, которые можно использовать при осуществлении согласно изобретению, включают в себя, без ограничения, вирус коровьей оспы, вирус птичьей оспы, вирус канареечной оспы, аденовирус, вирус гриппа, полиовирус, адено-ассоциированный вирус, лентивирус и вирус sindbis (см., например, Restifo, 1996, Curr. Opin. Immunol. 8: 658-663; Tsang et al. J. Natl. Cancer Inst. 87: 982-990 (1995)). Для индуцирования противоопухолевого ответа также можно использовать невирусные системы доставки путем введения пациенту оголенной ДНК, кодирующей белок, родственный STEAP-1 (например, внутримышечно или внутрикожно). Вирус коровьей оспы используют, например, для экспрессии нуклеотидных последовательностей, кодирующих пептиды согласно изобретению. После введения хозяину рекомбинантный вирус коровьей оспы обеспечивает экспрессию иммуногенного пептида белка и посредством этого индуцирует у хозяина иммунный ответ. Векторы на основе вируса коровьей оспы и способы их применения в методах иммунизации описаны, например, в патенте США № 4722848. В качестве вектора также можно использовать BCG (Bacille Calmette Guerin). Векторы на основе BCG описаны в Stover et al., Nature, 351: 456-460 (1991). Из настоящего описания специалистам в данной области будет очевидно, что для введения с терапевтическими целями или для иммунизации против пептидов согласно изобретению можно использовать широкий ряд других векторов, таких как векторы на основе адено- и адено-ассоциированных вирусов, ретровирусные векторы, векторы на основе обезвреженного вируса сибирской язвы и т.п. Таким образом, для доставки молекулы нуклеиновой кислоты, родственной STEAP-1, используют системы генной доставки. В одном воплощении используют полноразмерную кДНК человеческого STEAP-1. В другом воплощении используют молекулы нуклеиновых кислот STEAP-1, кодирующие эпитопы, специфичные для цитотоксичных Т-лимфоцитов (CTL) и/или антител. Ex vivo вакцины. Для индуцирования иммунного ответа можно использовать различные ex vivo стратегии. Один из подходов включает в себя применение антигенпрезентирующих клеток (АРС), таких как дендритные клетки (DC), для предоставления антигена STEAP-1 иммунной системе пациента. Дендритные клетки экспрессируют молекулы МНС классов I и II, костимулятор В7 и IL-12 и, следовательно, являются высокоспециализированными антигенпрезентирующими клетками. При раке простаты аутологичные дендритные клетки, сенсибилизированные пептидами простат-специфического мембранного антигена (PSMA), используют в фазе I клинических испытний для стимуляции иммунной системы пациентов с раком простаты (Tjoa et al., 1996, Prostate, 28: 65-69; Murphy et al., 1996, Prostate, 29: 371-380). Таким образом, дендритные клетки можно использовать для презентирования пептидов STEAP-1 Т-клеткам в контексте молекул МНС классов I или II. В одном воплощении аутологичные дендритные клетки сенсибилизируют пептидами STEAP-1, способными связываться с молекулами МНС класса I и/или класса II. В другом воплощении дендритные клетки сенсибилизируют полноразмерным белком STEAP-1. Следующее воплощение включает в себя достижение с помощью методов генной инженерии сверхэкспрессии гена STEAP-1 в дендритных клетках с использованием векторов, известных в данной области, таких как векторы на основе аденовируса (Arthur et al., 1997, Cancer Gene Ther. 4: 17-25), ретровируса (Henderson et al., 1996, Cancer Res. 56: 3763-3770), лентивируса, адено-ассоциированного вируса, а также путем трансфекции ДНК (Ribas et al., 1997, Cancer Res. 57: 2865-2869), или трансфекции опухолевой РНК (Ashley et al., 1997, J. Exp. Med. 186: 1177-1182). Клетки, экспрессирующие STEAP-1, также можно изменить методами генной инженерии, чтобы они экспрессировали иммунные модуляторы, такие как GM-CSF, и использовать их в качестве иммунизирующих средств. Х.В. STEAP-1 в качестве мишени для терапии антителами. STEAP-1 является привлекательной мишенью для терапевтических стратегий с применением антител. В данной области известен ряд стратегий с применением антител, направленных как на внеклеточные, так и на внутриклеточные молекулы (см., например, комплемент- и ADCC-опосредованное уничтожение, а также применение интрател). Поскольку STEAP-1 экспрессируется раковыми клетками различного происхождения, но не соответствующими нормальными клетками, системное введение STEAP-1-иммунореактивных композиций обеспечивает превосходную чувствительность, не вызывая токсических, неспецифичных или связанных с другими органами эффектов, обусловленных связыванием иммунореактивной композиции органами и тканями, не являющимися мишенями. Антитела, специфически взаимодействующие с доменами STEAP-1, можно использовать для системного лечения злокачественных заболеваний, сопровождающихся экспрессией STEAP-1, либо в виде конъюгатов с токсином или терапевтическим средством, либо в виде отдельных антител, способных ингибировать клеточную пролиферацию или функцию. Пациенту можно вводить антитела STEAP-1 с тем, чтобы они связывались со STEAP-1 и модулировали его функции, такие как взаимодействие с партнером по связыванию, и, как следствие, опосредовали деструкцию опухолевых клеток и/или ингибировали рост опухолевых клеток. Механизмы, посредством которых антитела оказывают терапевтический эффект, могут включать в себя комплемент-опосредованный цитолиз, антитело-зависимую клеточную цитотоксичность, модуляцию физиологической функции STEAP-1, ингибирование связывания лиганда или путей передачи сигнала, модуляцию дифференциации опухолевых клеток, изменение профилей факторов опухолевого ангиогенеза и/или апоптоз. Примеры препаратов на основе антител включают в себя Rituxan ® для неходжкинской лимфомы, Herceptin ® для метастазирующего рака молочной железы и Erbitux ® для рака ободочной и прямой кишки. Специалистам в данной области следует понимать, что можно использовать антитела, специфичные к иммуногенным молекулам и связывающиеся с такими молекулами, например с иммуногенным участком последовательности STEAP-1, изображенной на фиг. 2 или 3. Кроме того, специалистам в данной области известно, что получение конъюгатов антител с цитотоксичными средствами является рутинной практикой (см., например, Slevers et al. Blood, 93: 11, 3678-3684 (June 1, 1999)). Если цитотоксичное и/или терапевтическое средство доставляется непосредственно к клеткам, например, путем использования конъюгатов такого средства с антителами, специфичными к молекуле, экспрессирующейся данными клетками (например, STEAP-1), то цитотоксичное средство оказывает свой известный биологический эффект (т.е. цитотоксичный) непосредственно на указанные клетки. В данной области известен широкий ряд композиций и способов, связанных с использованием конъюгатов антитело-цитотоксичное средство для уничтожения клеток. В случае злокачественных заболеваний типичные способы включают в себя введение животному, несущему опухоль, биологически эффективного количества конъюгата, содержащего выбранное цитотоксичное и/или терапевтическое средство, связанное с направляющим агентом (например, с антителом против STEAP-1), который связывается с экспрессируемым маркером (например, STEAP-1), доступным для связывания или локализованным на поверхности клеток. Типичное воплощение относится к способу доставки цитотоксичного и/или терапевтического средства к клетке, экспрессирующей STEAP-1, который включает в себя получение конъюгата цитотоксичного средства с антителом, которое иммуноспецифично связывается с эпитопом STEAP-1, и обеспечение воздействия полученного конъюгата антитело-средство на клетку. Другое иллюстративное воплощение относится к способу лечения субъекта, предположительно страдающего от метастазирующего злокачественного заболевания, который включает в себя стадию парентерального введения указанному субъекту фармацевтической композиции, содержащей терапевтически эффективное количество антитела, конъюгированного с цитотоксическим и/или терапевтическим средством. Противораковую иммунотерапию с использованием антител против STEAP-1 можно проводить разными способами, успешно применяющимися для лечения других типов рака, включающих в себя, без ограничения, рак толстой кишки (Arlen et al., 1998, Crit. Rev. Immunol. 18: 133-138), множественную миелому (Ozaki et al., 1997, Blood. 90: 3179-3186, Tsunenari et al., 1997, Blood 90: 2437-2444), рак желудка (Kasprzyk et al., 1992, Cancer Res. 52: 2771-2776), В-клеточную лимфому (Funakoshi et al., 1996, J. Immunother. Emphasis Tumor Immunol. 19: 93-101), лейкоз (Zhong et al., 1996, Leuk. Res. 20: 581-589), рак толстой и прямой кишки (Moun et al., 1994, Cancer Res. 54: 6160-6166; Velders et al., 1995, Cancer Res. 55: 4398-4403) и рак молочной железы (Shepard et al., 1991, J. Clin. Immunol. 11: 117-127). Некоторые терапевтические способы включают в себя получение конъюгатов свободного антитела с токсином или радиоизотопом, например получение конъюгатов Y 91 или I 131 с антителами против CD20 (например, Zevalin ™, IDEC Pharmaceuticals Corp. или Bexxar ™, Coulter Pharmaceuticals) соответственно, тогда как другие способы включают в себя совместное введение антител и других терапевтических средств, таких как Herceptin ™ (trastuzuMab), с паклитакселом (Genentech, Inc.). Антитела можно конъюгировать с терапевтическим средством. При лечении рака простаты, например, антитела STEAP-1 можно вводить в сочетании с лучевой терапией, химиотерапией или антигормональной терапией. Кроме того, антитела можно конъюгировать с токсином, таким как каличемицин (например, Mylotarg ™, Wyeth-Ayerst, Madison, NJ, каппа-цепь рекомбинантного гуманизированного антитела IgG 4 , конъюгированная с противоопухолевым антибиотиком каличемицином), майтанзиноид (например, активируемое опухолью пролекарство на основе таксана, ТАР, platform, ImmunoGen, Cambridge, MA, также см., например, патент США № 5416064) или ауристатин Е (Seattle Genetics). Хотя терапию антителами против STEAP-1 можно проводить на всех стадиях рака, она в особенности подходит для лечения запущенных или метастазирующих злокачественных заболеваний. Терапия с применением антител согласно изобретению показана для пациентов, перенесших один или несколько циклов химиотерапии. Альтернативно, для пациентов, которые не получали химиотерапии, лечение антителами согласно изобретению можно объединить с химиотерапией или лучевой терапией. Кроме того, терапия антителами позволяет уменьшить дозы химиотерапевтических средств, что особенно важно для пациентов, которые плохо переносят токсичность химиотерапевтических средств. В Fan et al. (Cancer Res. 53: 4637-4642, 1993), Prewett et al. (International J. of Onco. 9: 217-224, 1996) и Hancock et al. (Cancer Res. 51: 4575-4580, 1991) описано применение разных антител вместе с химиотерапевтическими средствами. Хотя терапию антителами против STEAP-1 можно проводить на всех стадиях рака, она в особенности подходит для лечения запущенных или метастазирующих злокачественных заболеваний. Терапия с применением антител согласно изобретению показана для пациентов, перенесших один или несколько циклов химиотерапии. Альтернативно, для пациентов, которые не получали химиотерапии, лечение антителами согласно изобретению можно объединить с химиотерапией или лучевой терапией. Кроме того, терапия антителами позволяет уменьшить дозы химиотерапевтических средств, что особенно важно для пациентов, которые плохо переносят токсичность химиотерапевтических средств. У раковых пациентов можно определить наличие и уровень экспрессии STEAP-1 предпочтительно с помощью иммуногистохимических анализов опухолевой ткани, количественной визуализации STEAP-1, или других методов, позволяющих надежно определить наличие и степень экспрессии STEAP-1. Для данной цели предпочтительно использовать иммуногистохимический анализ биопсийных образцов опухолевой ткани или образцов, полученных при хирургической операции. Методы иммуногистохимических анализов опухолевых тканей хорошо известны в данной области. Моноклональные антитела против STEAP-1, используемые для лечения рака простаты и других злокачественных заболеваний, включают в себя антитела, которые инициируют мощный иммунный ответ против опухоли, а также антитела, которые сами по себе обладают цитотоксичной активностью. При этом моноклональные антитела против STEAP-1 (Mab) могут вызывать лизис опухолевых клеток по механизму либо комплемент-опосредованной, либо антитело-зависимой клеточной цитотоксичности (ADCC), причем для реализации обоих указанных механизмов требуется присутствие в молекуле иммуноглобулина интактного Fc-фрагмента, обеспечивающего взаимодействие с Fc-рецепторными участками эффекторных клеток в присутствии белков комплемента. Кроме того, Mab против STEAP-1, которые оказывают непосредственное биологическое действие на рост опухоли, можно использовать для лечения злокачественных заболеваний, сопровождающихся экспрессией STEAP-1. Механизмы непосредственного цитотоксического действия Mab включают в себя ингибирование клеточного роста, модуляцию дифференциации клеток, модуляцию профилей факторов опухолевого ангиогенеза и индукцию апоптоза. Механизм(ы) противоопухолевого действия конкретного Mab против STEAP-1 определяют с помощью ряда широко известных в данной области анализов in vitro, позволяющих оценить гибель клеток, таких как ADCC, ADMMC, комплемент-опосредованный лизис клеток и др. У некоторых пациентов применение мышиных или других нечеловеческих моноклональных антител, или человеческих/мышиных химерных Mab, может индуцировать иммунные ответы против нечеловеческих антител с интенсивностью от средней до высокой. Такие иммунные ответы могут приводить к выведению антитела из кровотока и к уменьшению эффективности антитела. В большинсте тяжелых случаев при таком иммунном ответе может происходить обширное образование иммунных комплексов, которое может вызвать почечную недостаточность. Соответственно, в терапевтических способах согласно изобретению предпочтительно используют либо полностью человеческие, либо гуманизированные моноклональные антитела, которые специфически связываются с целевым антигеном STEAP-1 с высоким сродством, но не вызывают антигенных реакций у пациента или вызывают их в незначительной степени. Терапевтические способы согласно изобретению включают в себя введение отдельных Mab против STEAP-1, а также сочетаний и смесей разных Mab. Такие смеси Mab могут иметь некоторые преимущества, поскольку они могут содержать Mab против разных эпитопов, действующие через разные механизмы, или смесь Mab, обладающих непосредственной токсичностью, и Mab, являющихся иммунными эффекторами. Такие Mab в сочетании могут оказывать синергические терапевтические эффекты. Кроме того, Mab против STEAP-1 можно вводить наряду с применением других терапевтических средств, включающих в себя, без ограничения, разные химиотерапевтические средства, андрогенные блокаторы, иммунные модуляторы (например, IL-2, GM-CSF), хирургическое лечение или лучевую терапию. Mab против STEAP-1 вводят либо в "свободной", или неконъюгированной, форме, либо в виде конъюгата с терапевтическим средством (средствами). Композиции антител против STEAP-1 вводят с помощью способов, позволяющих доставлять антитела к опухолевым клеткам. Способы введения включают в себя, без ограничения, внутривенный, внутрибрюшинный, внутримышечный, внутриопухолевый, внутрикожный и т.п. Лечение обычно включает в себя повторяющееся введение препаратов антител против STEAP-1 с использованием подходящего способа введения, такого как внутривенное введение (в.в.), при котором вводимая доза обычно составляет 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или 25 мг/кг массы тела. Как правило, дозы в интервале 10-1000 мг Mab в неделю являются эффективными и хорошо переносятся. Клинические эксперименты с использованием Mab Herceptin ™ для лечения метастазирующего рака молочной железы показали, что подходящий режим дозирования включает в себя в.в. введение начальной ударной дозы, составляющей приблизительно 4 мг препарата Mab против STEAP-1/кг массы тела пациента, и затем еженедельное в.в. введение дозы, составляющей приблизительно 2 мг/кг. Предпочтительно начальную ударную дозу вводят в течение 90-минутной или более продолжительной инфузии. Периодическую поддерживающую дозу вводят в течение 30-минутной или более продолжительной инфузии при условии, что пациент хорошо перенес начальную дозу. Специалистам в данной области известно, что выбор идеального для каждого конкретного случая режима дозирования зависит от разных факторов. Такие факторы включают в себя, например, сродство связывания и период полужизни используемых Ab или Mab, степень экспрессии STEAP-1 у пациента, уровень сбрасываемого антигена STEAP-1 в крови, желательный уровень стационарной концентрации антитела, частота введения препарата, влияние химиотерапевтических или других средств, используемых в сочетании со способом лечения согласно изобретению, а также состояние здоровья конкретного пациента. Чтобы облегчить выбор наиболее эффективного режима дозирования и др., у пациента необязательно определяют уровень STEAP-1 в конкретном образце (например, уровни антигена STEAP-1 и/или STEAP-1-экспрессирующих клеток в кровотоке). Такие анализы также проводят с целью мониторинга на протяжении всего лечения и используют для оценки эффективности лечения в сочетании со значениями других параметров (например, с результатами цитологического анализа мочи и/или уровня ImmunoCyt при лечении рака мочевого пузыря или по аналогии - уровня PSA в сыворотке при лечении рака простаты). Антиидиотипические антитела против STEAP-1 также можно использовать в противораковой терапии в качестве вакцины для индуцирования иммунного ответа на клетки, экспрессирующие белок, родственный STEAP-1. В частности, в данной области хорошо известны способы получения антиидиотипических антител; данную методологию можно легко адаптировать для получения антиидиотипических антител против STEAP-1, которые имитируют эпитоп, находящийся в белке, родственном STEAP-1 (см., например, Wagner et al., 1997, Hybridoma, 16: 33-40; Foon et al., 1995, J. Clin. Invest. 96: 334-342; Herlyn et al., 1996, Cancer Immunol. Immunother. 43: 65-76). Такие антиидиотипические антитела могут быть использованы в вакцинных стратегиях противораковой терапии. Х.С. STEAP-1 как мишень для клеточных иммунных ответов. Следующими воплощениями согласно изобретению являются вакцины и способы получения вакцин, содержащих иммунологически эффективное количество одного или нескольких HLA-связывающих пептидов, описанных в данном документе. Вакцины в соответствии с данным изобретением охватывают композиции, содержащие один или несколько из заявляемых пептидов. Пептид может присутствовать в вакцине один. Альтернативно, пептид может существовать в виде гомополимера, содержащего несколько копий одного и того же пептида, или в виде гетерополимера, состоящего из разных пептидов. Полимеры имеют преимущество, заключающееся в повышенной иммунологической реакционноспособности, и, если для получения полимера используют разные пептиды, иммунный ответ дополняется способностью полимера индуцировать образование антител и/или CTL, которые взаимодействуют с другими антигенными детерминантами патогенного организма или связанного с опухолью пептида. Композиция может представлять собой природное окружение антигена или она может быть получена, например, рекомбинантными или синтетическими методами. Вакцины согласно изобретению могут содержать хорошо известные в данной области носители, которые включают в себя, например, тироглобулин, альбумины, такие как человеческий сывороточный альбумин, столбнячный токсин, полиаминокислоты, такие как поли-L-лизин, поли-L-глутаминовая кислота, вирус гриппа, капсидный белок вируса гепатита В и т.п. Вакцины могут содержать физиологически допустимый (т.е. приемлемый) разбавитель, такой как вода, или физиологический раствор, предпочтительно забуференный фосфатом физиологический раствор. Кроме того, вакцины обычно содержат адъювант. Адъюванты, такие как неполный адъювант Фрейнда, фосфат алюминия, гидроксид алюминия или квасцы, представляют собой вещества, хорошо известные в данной области. Кроме того, как описано в данном документе, CTL-ответы можно инициировать с помощью конъюгатов пептидов согласно изобретению с липидами, такими как трипальмитоил-3-глицерилцистеинилсерилсерин (P 3 CSS). Было также обнаружено, что адъюванты, такие как синтетические олигонуклеотиды, содержащие цитозин-фосфоротиолированный-гуанин (CpG), усиливают CTL-ответы в 10-100 раз (см., например, Davila and Celis, J. Immunol. 165: 539-547 (2000)). После иммунизации пептидной композицией согласно изобретению путем инъекции, аэрозольного введения, перорального введения, чрескожного введения, введения через слизистую оболочку, внутриплеврального введения, интратекального введения или других способов введения иммунная система хозяина отвечает на вакцину, продуцируя большие количества CTL и/или HTL, специфичных к целевому антигену. В результате хозяин становится, по меньшей мере, частично иммунным по отношению к клеткам с более поздним развитием, в которых происходит экспрессия или сверхэкспрессия антигена STEAP-1, или испытывает, по меньшей мере, некоторое улучшение, если антиген является ассоциированным с опухолью. В некоторых воплощениях может быть желательно объединить пептидные компоненты класса I с компонентами, которые индуцируют или облегчают нейтрализацию антител и/или ответы хелперных Т-клеток, направленные на целевой антиген. Предпочтительное воплощение такой композиции содержит эпитопы класса I и класса II в соответствии с данным изобретением. Альтернативное воплощение такой композиции включает в себя эпитоп класса I и/или класса II в соответствии с данным изобретением, наряду с перекрестно взаимодействующим эпитопом HTL, таким как молекула PADRE ™ (Epimmune, San Diego, CA) (описанная, например, в патенте США № 5736142). Вакцина согласно изобретению также может содержать антигенпрезентирующие клетки (АРС), такие как дендритные клетки (DC), в качестве носителя, презентирующего пептиды согласно изобретению. Вакцинные композиции могут быть получены in vitro, после мобилизации и сбора дендритных клеток, при этом загрузка дендритных клеток происходит in vitro. Например, дендритные клетки трансфицируют мини-геном согласно изобретению или сенсибилизируют пептидами. Затем дендритные клетки вводят пациенту, чтобы вызвать иммунный ответ in vivo. Вакцинные композиции на основе либо ДНК, либо пептида также можно вводить in vivo в сочетании с мобилизацией дендритных клеток, при этом загрузка дендритных клеток происходит in vivo. При выборе совокупности эпитопов, подходящих для включения в состав полиэпитопной композиции, входящей в состав вакцины, или при выборе отдельных эпитопов, подходящих для включения в состав вакцины, и/или кодируемых нуклеиновыми кислотами, такими как мини-ген, желательно использовать нижеследующие принципы. Желательно также при осуществлении выбора сопоставлять нижеследующие принципы друг с другом. Несколько эпитопов, предназначенных для включения в определенную вакцинную композицию, необязательно могут быть смежными в последовательности нативного антигена, из которого они получены. 1) Выбирают эпитопы, которые при введении имитируют иммунные ответы, которые по результатам наблюдения коррелируют с уничтожением опухоли. В случае HLA класса I такие эпитопы включают в себя 3-4 эпитопа, относящихся по меньшей мере к одному опухолеспецифическому антигену (ТАА). Это справедливо также для HLA класса II; снова выбирают 3-4 эпитопа, относящихся по меньшей мере к одному ТАА (см., например, Rosenberg et al., Science, 278: 1447-1450). Эпитопы из одного ТАА можно использовать в сочетании с эпитопами из одного или нескольких других ТАА с получением вакцины, направленной на опухоли с разным характером экспрессии часто экспрессируемых ТАА. 2) Выбирают эпитопы, имеющие желательное сродство связывания, которое, как установлено, коррелирует с иммуногенностью: для HLA класса I IC 50 составляет 500 нМ или меньше, часто 200 нМ или меньше; а для HLA класса II IC 50 составляет 1000 нМ или меньше. 3) Выбирают пептиды, несущие супермотивы, или совокупность пептидов, несущих аллель-специфические мотивы, которые имеют достаточно широкий популяционный охват. Например, предпочтительно они имеют по меньшей мере 80% популяционный охват. Для оценки широты или величины популяционного охвата можно использовать анализ Монте-Карло, известный в данной области метод статистических испытаний. 4) При выборе эпитопов из связанных с раком антигенов зачастую полезно выбирать аналоги, поскольку у пациента может развиться толерантность к нативному эпитопу. 5) Особое значение имеют эпитопы, называемые эпитопами "футлярного" типа. "Футлярные" эпитопы встречаются там, где по меньшей мере два эпитопа перекрываются в некоторой пептидной последовательности. Футлярная пептидная последовательность может включать в себя эпитопы В-клеток, HLA класса I и/или HLA класса II. При выборе футлярных эпитопов основной целью является выбор наибольшего числа эпитопов на последовательность. Следует избегать выбора пептида, который имеет несколько большую длину, чем аминоконец аминоконцевого эпитопа и карбоксильный конец карбоксильного концевого эпитопа в пептиде. При выборе полиэпитопной последовательности, такой как последовательность, содержащая футлярные эпитопы, желательно провести скрининг данной последовательности, чтобы удостовериться, что она не имеет патологических или других вредных биологических свойств. 6) При создании полиэпитопного белка или при создании мини-гена желательно получить наименьший пептид, охватывающий представляющие интерес эпитопы. Данный принцип подобен, если не идентичен, принципу, используемому при выборе пептида, содержащего футлярные эпитопы. Однако в случае искусственного полиэпитопного пептида минимизацию размера следует соотносить с необходимостью внедрения каких-либо спейсерных последовательностей между эпитопами в полиэпитопном белке. Спейсерные аминокислотные остатки, например, можно ввести, чтобы избежать соединенных эпитопов (эпитопов, распознаваемых иммунной системой, не присутствующих в целевом антигене, а образованных в результате созданного человеком непосредственного соседства эпитопов), или для облегчения расхождения эпитопов и, как результат, увеличения презентации эпитопа. Образования соединенных эпитопов обычно избегают, потому что реципиент может генерировать иммунный ответ на такие неприродные эпитопы. Особое значение имеет соединенный эпитоп, являющийся "доминантным эпитопом". Доминантный эпитоп может вызывать такой интенсивный ответ, при котором иммунные ответы на другие эпитопы уменьшаются или подавляются. 7) Если присутствуют последовательности нескольких вариантов одного целевого белка, потенциальные пептидные эпитопы можно выбирать на основе их консервативности. Например, показатель консервативности может указывать, что полноразмерная последовательность связывающего пептида HLA класса I или все 9-мономерное ядро связывающего пептида класса II обладают указанным процентом консервативности последовательностей, определенным для конкретного белкового антигена. Х.С.1. Вакцины на основе мини-генов. Известен ряд способов, обеспечивающих одновременную доставку нескольких эпитопов. Нуклеиновые кислоты, кодирующие пептиды согласно изобретению, являются особенно важным воплощением согласно изобретению. Эпитопы для включения в мини-ген выбирают в соответствии с принципами, описанными в предыдущем разделе. В предпочтительных способах введения нуклеиновых кислот согласно изобретению используют конструкции мини-генов, кодирующие пептиды, содержащие один или несколько эпитопов согласно изобретению. Применение полиэпитопных мини-генов описано ниже и в Ishioka et al., J. Immunol. 162: 3915-3925, 1999; Ann, L. and Whitton, J.L., J. Virol. 71: 2292, 1997; Thomson, S.A. et al., J. Immunol. 157: 822, 1996; Whitton, J.L. et al., J. Virol. 67: 348, 1993; Hanke, R. et al., Vaccine, 16: 426, 1998. Например, с помощью методов генной инженерии можно получить полиэпитопную ДНК-плазмиду, кодирующую эпитопы STEAP-1, несущие супермотив и/или мотив, универсальный эпитоп хелперных Т-клеток PADRE ™ или несколько эпитопов HTL из STEAP-1 (см., например, табл. V-XVIII и XXII-LI), и последовательность транслокационного сигнала эндоплазматического ретикулума. Вакцина также может содержать эпитопы из других ТАА. Иммуногенность полиэпитопного мини-гена у трансгенных мышей можно подтвердить, определив величину ответов против тестируемых эпитопов, связанных с индукцией CTL. Кроме того, иммуногенность ДНК-кодируемых эпитопов in vivo может коррелировать с ответами специфических линий CTL in vitro против клеток-мишеней, трансфицированных ДНК-плазмидой. Таким образом, данные эксперименты демонстрируют, что присутствие мини-гена может приводить к 1) генерации ответа CTL и 2) экспрессии кодируемых им эпитопов клетками, распознаваемыми индуцированными CTL. Например, чтобы создать последовательность ДНК, кодирующую выбранные эпитопы (мини-ген) и пригодную для экспрессии в человеческих клетках, аминокислотные последовательности эпитопов можно подвергнуть обратной трансляции. При выборе кодона для каждой аминокислоты можно руководствоваться таблицей используемости человеческих кодонов. Данные последовательности ДНК, кодирующие эпитопы, могут непосредственно примыкать друг к другу, тогда при трансляции образуется непрерывная полипептидная последовательность. Чтобы оптимизировать экспрессию и/или иммуногенность, в мини-ген можно включить дополнительные элементы. Примеры аминокислотных последовательностей, которые можно подвергнуть обратной трансляции и ввести в последовательность мини-гена, включают в себя: эпитопы HLA класса I, эпитопы HLA класса II, эпитопы антител, последовательность сигнала убиквитинирования и/или сигнала, направленного на эндоплазматический ретикулум. Кроме того, презентацию HLA CTL и эпитопов HTL можно улучшить путем включения в состав мини-гена синтетических (например, полиаланин) или природных фланкирующих последовательностей так, чтобы они примыкали к эпитопам CTL или HTL; эпитопы, входящие в состав этих более крупных пептидов, также включены в объем согласно изобретению. Последовательность мини-гена можно превратить в последовательность ДНК путем сборки олигонуклеотидов, кодирующих плюс- и минус-цепи мини-гена. Можно синтезировать перекрывающиеся олигонуклеотиды (30-100 оснований в длину), которые затем можно фосфорилировать, очистить и подвергнуть отжигу в подходящих условиях с помощью хорошо известных методов. Концы олигонуклеотидов можно соединять, например, с помощью ДНК-лигазы Т4. Затем полученный синтетический мини-ген, кодирующий эпитопный полипептид, можно клонировать в подходящем векторе экспрессии. Чтобы обеспечить экспрессию в клетках-мишенях, в состав вектора предпочтительно включают стандартные регуляторные последовательности, хорошо известные специалистам в данной области. Желательно, чтобы в векторе присутствовали следующие элементы: промотор с участком клонирования для вставки мини-гена, расположенным ниже по ходу считывания; сигнал полиаденилирования для эффективной терминации транскрипции; репликатор Е.coli; и селектируемый маркер Е.coli (например, ген устойчивости к ампициллину или канамицину). Для данной цели можно использовать ряд промоторов, например промотор человеческого цитомегаловируса (hCMV). Другие подходящие последовательности промоторов можно найти, например, в патентах США № 5580859 и 5589466. Для оптимизации экспрессии и иммуногенности мини-гена можно использовать другие модификации вектора. В некоторых случаях, если для эффективной экспрессии гена требуются интроны, в транскрибируемый участок мини-гена можно включить один или несколько синтетических или природных интронов. Последовательности, стабилизирующие мРНК, и последовательности, обеспечивающие репликацию в клетках млекопитающих, также могут вносить вклад в увеличение экспрессии мини-гена. После выбора вектора экспрессии мини-ген клонируют в полилинкерном участке, расположенном ниже промотора. Данную плазмиду трансформируют в подходящем штамме Е.coli и получают ДНК с помощью стандартных методов. Ориентацию и последовательность ДНК мини-гена, а также присутствие в векторе других элементов подтверждают с помощью рестрикционной карты и анализа последовательности ДНК. Бактериальные клетки, несущие правильную плазмиду, хранят в виде банка базовых клеток и банка рабочих клеток. Кроме того, в иммуногенность ДНК-вакцин вносят вклад иммуностимуляторные последовательности (ISSs или CpGs). Если нужно повысить иммуногенность, данные последовательности вводят в состав вектора вне кодирующей последовательности мини-гена. В некоторых воплощениях можно использовать бицистронный вектор экспрессии, который обеспечивает продукцию как кодируемых мини-геном эпитопов, так и второго белка (используемого для увеличеня или уменьшения иммуногенности). Примеры белков или полипептидов, которые при совместной экспрессии могут существенно усилить иммунный ответ, включают в себя цитокины (например, IL-2, IL-12, GM-CSF), индуцируемые цитокинами молекулы (например, LeIF), костимуляторные молекулы или в случае HTL-ответов связывающие белки pan-DR (PADRE ™, Epimmune, San Diego, CA). Эпитопы хелперных лимфоцитов (HTL) можно соединить с внутриклеточными направляющими сигналами и экспрессировать отдельно от эпитопов CTL; это позволяет направить эпитопы HTL и CTL в разные компартменты клетки. При необходимости это может обеспечить более эффективный вход эпитопов HTL в путь, опосредованный HLA класса II, и посредством этого повысить индукцию HTL. В противоположность индукции HTL или CTL при некоторых заболеваниях благоприятный эффект может иметь специфическое уменьшение иммунного ответа под влиянием совместной экспрессии иммуносупрессивных молекул (например, TGF- β). Плазмидную ДНК в терапевтических количествах можно получить, например, путем ферментации в Е.coli с последующей очисткой. Аликвоты из банка рабочих клеток вносят в питательную среду и выращивают до насыщения в колбах для шейкера или в биореакторе, используя известные в данной области методы. Плазмидную ДНК можно очистить с помощью стандартных методов биоразделения, таких как твердофазное разделение на анионообменных смолах, поставляемых QIAGEN, Inc. (Valencia, California). При необходимости сверхспиральные ДНК можно выделить из открытых циклических и линейных форм с помощью гель-электрофореза или других методов. Очищенную плазмидную ДНК для инъекций можно получить с использованием ряда композиций. Самая простая из них представляет собой раствор лиофилизованной ДНК в стерильном физиологическом растворе, забуференном фосфатом (PBS). Данный препарат, известный как "оголенная ДНК", в настоящее время используется в клинических испытаниях для внутримышечного введения (в.м.). Для максимизации иммунотерапевтических эффектов вакцин на основе ДНК мини-генов может быть желательным использование альтернативного способа получения очищенной плазмиды ДНК. Ряд способов уже описан и могут стать доступными новые способы. В композиции также можно использовать катионные липиды, гликолипиды и сливающиеся липосомы (см., например, WO 93/24640; Mannino & Gould-Fogerite, BioTechniques, 6(7): 682 (1988); патент США № 5279833; WO 91/06309 и Felgner, et al., Proc. Natl. Acad. Sci. USA, 84: 7413 (1987). Кроме того, можно получить комплексы очищенной плазмидной ДНК с пептидами и соединениями, называемыми в совокупности защитными, интерактивными, неконденсирующими соединениями (PINC), которые позволяют модифицировать такие характеристики, как стабильность, внутримышечная дисперсия или способность достигать конкретные органы или типы клеток. Сенсибилизацию целевых клеток можно использовать в качестве функционального анализа экспрессии кодируемых мини-геном эпиопов CTL и их презентации HLA класса I. Например, плазмидную ДНК вводят в линию клеток млекопитающих, подходящую для использования в качестве мишени в стандартном анализе высвобождения хрома из CTL. Выбор метода трансфекции зависит от конечной композиции. Для введения "оголенной" ДНК можно использовать электропорацию, где катионные липиды позволяют осуществлять непосредственную трансфекцию in vitro. Можно одновременно трансфицировать плазмиду, экспрессирующую зеленый флуоресцентный белок (GFP), позволяющий осуществить селекцию трансфицированных клеток с помощью сортинга клеток с активацией флуоресценции (FACS). Затем данные клетки метят хромом-51 ( 51 Cr) и используют в качестве клеток-мишеней для эпитоп-специфичных линий CTL; цитолиз, детектируемый по высвобождению 51 Cr, указывает как на продукцию кодируемых мини-геном эпитопов CTL, так и на их презентацию HLA. Экспрессию эпитопов HTL можно оценить аналоговым способом с использованием результатов анализов активности HTL. Иммуногенность in vivo является вторым способом функционального тестирования композиций, содержащих ДНК мини-генов. Трансгенных мышей, экспрессирующих соответствующие человеческие белки HLA, иммунизируют продуктом ДНК. Доза и способ введения зависят от состава композиции (например, в.м. введение используют для ДНК в PBS, а внутрибрюшинное (в.б.) введение используют для комплексов ДНК с липидами). Через 21 день после иммунизации спленоциты собирают и повторно стимулируют в течение одной недели в присутствии пептидов, кодирующих каждый тестируемый эпитоп. Затем в случае эффекторных клеток CTL анализируют цитолиз нагруженных пептидом 51 Cr-меченных клеток-мишеней с помощью стандартных методов. Лизис клеток-мишеней, сенсибилизированных HLA, содержащим пептидные эпитопы, соответствующие эпитопам, кодируемым мини-геном, демонстрирует влияние ДНК-вакцины на индукцию CTL in vivo. Иммуногенность эпитопов HTL у трансгенных мышей подтверждают аналогичным способом. Альтернативно, нуклеиновые кислоты можно вводить с помощью баллистической системы доставки, как описано, например, в патенте США № 5204253. Данный метод включает в себя введение частиц, содержащих только ДНК. В следующем альтернативном воплощении вводят ДНК, прилипшую к частицам, таким как частицы золота. Мини-гены также можно доставлять с помощью других бактериальных или вирусных систем доставки, хорошо известных в данной области, например экспрессионная конструкция, кодирующая эпитопы согласно изобретению, может входить в состав вирусного вектора, такого как вектор на основе вируса коровьей оспы. Х.С.2. Сочетание пептидов CTL с хелперными пептидами. Вакцинные композиции, содержащие пептиды CTL согласно изобретению, можно модифицировать, например, с получением аналогов, обладающих желательными свойствами, такими как увеличенный период полужизни в сыворотке, расширенный популяционный охват или повышенная иммуногенность. Например, способность пептида индуцировать активность CTL, можно повысить путем присоединения пептида к последовательности, которая содержит по меньшей мере один эпитоп, способный индуцировать ответ, опосредованный хелперными Т-клетками. Хотя пептид CTL можно непосредственно присоединить к пептиду Т-хелпера, зачастую они соединены в конъюгатах эпитоп CTL/эпитоп HTL через спейсерную молекулу. Спейсер обычно представляет собой относительно маленькие нейтральные молекулы, такие как аминокислоты или имитаторы аминокислот, которые в физиологических условиях являются практически незаряженными. Спейсеры обычно выбирают, например, из Ala, Gly или других неполярных аминокислот или нейтральных полярных аминокислот. Следует понимать, что необязательно присутствующий спейсер может состоять из одинаковых или разных остатков и, следовательно, может представлять собой гетеро- или гомоолигомер. В случае присутствия спейсер обычно содержит по меньшей мере один или два остатка, чаще от трех до шести остатков, иногда 10 или более остатков. Пептидный эпитоп CTL может быть связан с пептидным эпитопом Т-хелпера либо непосредственно или через спейсер, либо по амино- или карбоксиконцу пептида CTL. Аминоконец иммуногенного пептида или Т-хелперного пептида может быть ацилирован. В некоторых воплощениях Т-хелперный пептид представляет собой пептид, который распознается Т-хелперными клетками, присутствующими у большей части генетически различающихся популяций. Выбирают пептиды, которые связываются со многими, большинством или со всеми молекулами HLA класса II. Примеры аминокислотных последовательностей, связывающихся со многими молекулами HLA класса II, включают в себя последовательности из таких антигенов, как столбнячный токсин, положения 830-843 QYIKANSKFIGITE (SEQ ID NO: 64), околоспорозоидный белок (CS) Plasmodium falciparum, положения 378-398 DIEKKIAKMEKASSVFNVVNS (SEQ ID NO: 65) и белок Streptococcus размером 18 кДа, положения 116-131 GAVDSILGGVATYGAA (SEQ ID NO: 66). Другие примеры включают в себя петиды, несущие супермотив DR 1-4-7 или любой из мотивов DR3. Альтернативно, можно получить синтетические пептиды, способные стимулировать Т-хелперные лимфоциты, в форме, несколько ограниченной HLA, с использованием аминокислотных последовательностей, не обнаруженных в природе (см., например, публикацию РСТ WO 95/07707). Указанные синтетические соединения, называемые Pan-DR-связывающие эпитопы (например, PADRE ™, Epimmune, Inc., San Diego, CA), наиболее предпочтительно конструируют так, чтобы они связывали большинство молекул HLA-DR (человеческие HLA класса II). Например, было обнаружено, что pan-DR-связывающий эпитопный пептид, имеющий формулу XKXVAAWTLKAAX (SEQ ID NO: 67), где X обозначает любой остаток из циклогексилаланина, фенилаланина или тирозина; А обозначает либо D-аланин, либо L-аланин, связывается с большинством аллелей HLA-DR и стимулирует ответ Т-хелперных лимфоцитов у большинства субъектов, независимо от типа HLA. В альтернативном pan-DR-связывающем эпитопе все аминокислоты представляют собой природные L-аминокислоты и могут быть представлены в виде нуклеиновых кислот, кодирующих эпитоп. Пептидные эпитопы HTL также можно модифицировать с целью изменения их биологических свойств. Например, в их состав можно ввести D-аминокислоты для увеличения устойчивости к протеазам и, следовательно, увеличения периода полужизни в сыворотке или их можно конъюгировать с другими молекулами, такими как липиды, белки, углеводы и т.п., для увеличения их биологической активности. Например, Т-хелперный пептид можно конъюгировать с одной или несколькими цепями пальмитиновой кислоты по аминоконцу или по карбоксильному концу. X.С.3. Сочетания пептидов CTL с агентами, сенсибилизирующими Т-клетки. В некоторых воплощениях желательно включать в состав фармацевтических композиций согласно изобретению по меньшей мере один компонент, который сенсибилизирует В- или Т-лимфоциты. Известно, что липиды способны сенсибилизировать CTL in vivo. Например, остатки пальмитиновой кислоты можно присоединить к ε- и α-аминогруппам остатка лизина и полученное производное можно присоединить, например, с использованием одного или нескольких связующих остатков, таких как Gly, Gly-Gly, Ser, Ser-Ser и т.п., к иммуногенному пептиду. Затем липидированный пептид можно вводить, либо непосредственно в мицелле или частице, заключенной в липосому, либо в виде эмульсии в адъюванте, например в неполном адъюванте Фрейнда. В предпочтительном воплощении особенно эффективная иммуногенная композиция содержит пальмитиновую кислоту, присоединенную к ε-и α-аминогруппам Lys, который через мостик, например Ser-Ser, соединен с аминоконцом иммуногенного пептида. В качестве другого примера липида, инициирующего ответы, опосредованные CTL, для сенсибилизации вирус-специфических CTL можно использовать липобелки Е.coli, такие как трипальмитоил-S-глицерилцистеинилсерилсерин (P 3 CSS), ковалентно связанные с соответствующим пептидом (см., например, Deres, et al., Nature, 342: 561, 1989). Пептиды согласно изобретению можно присоединить к P 3 CSS, например, и полученный липопептид можно ввести субъекту для индуцирования специфического иммунного ответа на целевой антиген. Более того, поскольку эпитопы, конъюгированные с P 3 CSS, также могут инициировать образование нейтрализующих антител, для более эффективной индукции как гуморального, так и клеточного ответов, две указанные композиции можно объединить. X.С.4. Вакцинные композиции, содержащие DC, сенсибилизированные пептидами CTL и/или HTL. Воплощение вакцинной композиции в соответствии с данным изобретением включает в себя введение ex vivo смеси эпитоп-несущих пептидов в РВМС или в выделенные из них DC, полученные из крови пациента. Можно использовать фармацевтическое средство, облегчающее сбор DC, такое как Progenipoietin ™ (Pharmacia-Monsanto, St. Louis, МО) или GM-CSF/IL-4. После сенсибилизации пептидами и перед повторным вливанием пациентам DC промывают, чтобы удалить несвязанные пептиды. В данном воплощении вакцина содержит сенсибилизированные пептидами DC, которые презентируют на поверхности, продуцированные в результате сенсибилизации пептидные эпитопы, в комплексе с молекулами HLA. DC можно сенсибилизировать ex vivo смесью пептидов, некоторые из которых стимулируют ответы CTL на STEAP-1. Для усиления ответа CTL в состав композиции необязательно вводят пептид хелперной Т-клетки (HTL), такой как природный или искусственный пептид, несколько ограниченный HLA класса II. Таким образом, вакцину согласно изобретению используют для лечения рака, сопровождающегося экспрессией или сверхэкспрессией STEAP-1. X.D. Адоптивная иммунотерапия. Антигенные STEAP-1-родственные пептиды используют для индуцирования как CTL-, так и HTL-ответов vivo. Полученные CTL или HTL клетки можно использовать для лечения опухолей у пациентов, которые не отвечают на другие традиционные формы терапии или не отвечают на терапевтический вакцинный пептид или вакцинную нуклеиновую кислоту согласно изобретению. CTL- или HTL-ответы ex vivo на конкретный антиген индуцируют путем инкубации культуры ткани пациента или генетически совместимых клеток-предшественников CTL или HTL вместе с источником антигенпрезентирующих клеток (АРС), таких как дендритные клетки, и соответствующим иммуногенным пептидом. По прошествии подходящего времени инкубации (обычно приблизительно 7-28 дней), в течение которого клетки-предшественники активируются и развиваются в эффекторные клетки, клетки вводят обратно пациенту, где они разрушают (CTL) свои специфические клетки-мишени (например, опухолевые клетки) или облегчают их разрушение (HTL). В качестве антигенпрезентирующих клеток также можно использовать трансфицированные дендритные клетки. X.Е. Введение вакцин с терапевтической или профилактической целью. Фармацевтические и вакцинные композиции согласно изобретению обычно используют для лечения и/или профилактики злокачественного заболевания, сопровождающегося экспрессией или сверхэкспрессией STEAP-1. При терапевтическом применении композиции, содержащие пептиды и/или нуклеиновые кислоты, вводят пациенту в количестве, достаточном для индуцирования эффективного В-клеточного, CTL- и/или HTL-ответа на антиген, а также для лечения или, по меньшей мере, частичного устранения или замедления симптомов и/или осложнений. Количество, достаточное для осуществления данной цели, называют "терапевтически эффективной дозой". Количество, эффективное для данного применения, зависит, например, от вида вводимой композиции, способа введения, стадии и тяжести заболевания, подлежащего лечению, массы тела и общего состояния здоровья пациента, а также от оценки врача, назначающего лечение. Что касается фармацевтических композиций, иммуногенные пептиды согласно изобретению или кодирующие их молекулы ДНК, как правило, вводят субъекту, уже несущему опухоль, которая сопровождается экспрессией STEAP-1. Пептиды или кодирующие их ДНК можно вводить индивидуально или в виде гибридов одной или нескольких пептидных последовательностей. Пациентов можно лечить иммуногенными пептидами отдельно или, по обстоятельствам, в сочетании с другими способами лечения, такими как хирургия. Что касается терапевтического применения, введение, как правило, следует начинать при первом диагнозе STEAP-1-ассоциированного злокачественного заболевания. Затем вводят поддерживающие дозы до тех пор, пока, по меньшей мере, не начнут ослабевать симптомы, и в течение некоторого времени после этого. Воплощение вакцинной композиции (т.е. включающее в себя, без ограничения, такие воплощения, как смеси пептидов, полиэпитопные полипептиды, мини-гены или ТАА-специфичные CTL или сенсибилизированные дендритные клетки), доставляемое пациенту, может варьировать в зависимости от стадии заболевания или состояния здоровья пациента. Например, в случае пациента с опухолью, сопровождающейся экспрессией STEAP-1, вакцина, содержащая STEAP-1-специфичные CTL, может быть более эффективной в отношении уничтожения опухолевых клеток пациента с запущенным заболеванием, чем альтернативные воплощения. Как правило, важно выбрать количество пептидного эпитопа, доставляемого с помощью определенного способа введения, достаточное для эффективной стимуляции Т-клеточного ответа; в соответствии с данным воплощением согласно изобретению также можно вводить композиции, которые стимулируют ответы хелперных Т-клеток. Разовая доза при начальной терапевтической иммунизации обычно находится в интервале, где низшее значение составляет приблизительно 1, 5, 50, 500 или 1000 мкг, а высшее значение составляет приблизительно 10000, 20000, 30000 или 50000 мкг. Значения дозы для человека массой 70 кг, как правило, варьируют приблизительно от 500 до 50000 мкг. Поддерживающие дозы в интервале приблизительно от 1,0 до 50000 мкг пептида, которые вводят в течение нескольких недель или месяцев, можно назначать в зависимости от ответа и состояния пациента, определяемых путем измерения специфической активности CTL и HTL, полученных из крови пациента. Введение следует продолжать до тех пор, пока, по меньшей мере, клинические симптомы или лабораторные анализы не укажут, что неоплазия устранена или уменьшена и в течение некоторого периода после этого. Дозы, способы введения и дозировочные режимы подбирают в соответствии с известными в данной области методами. В некоторых воплощениях пептиды и композиции согласно изобретению используются для лечения тяжелых болезненных состояний, т.е. в ситуациях, когда существует или может существовать угроза для жизни. В таких случаях, поскольку используются минимальные количества чужеродных веществ, а пептиды, входящие в состав предпочтительных композиций согласно изобретению, являются относительно нетоксичными, данные пептидные композиции можно вводить в количествах, значительно превышающих указанные дозы, что зачастую желательно с точки зрения лечащего врача. Вакцинные композиции согласно изобретению также можно использовать только в качестве профилактических средств. Как правило, при начальной профилактической иммунизации разовая доза находится в интервале, где низшее значение составляет приблизительно 1, 5, 50, 500 или 1000 мкг, а высшее значение составляет приблизительно 10000, 20000, 30000 или 50000 мкг. Значения дозы для человека массой 70 кг, как правило, варьируют приблизительно от 500 до 50000 мкг. Поддерживающие дозы в интервале приблизительно от 1,0 до 50000 мкг пептида вводят через определенные интервалы времени, приблизительно от четырех недель до шести месяцев, после первоначального введения вакцины. Иммуногенность вакцины можно определить путем измерения специфической активности CTL и HTL, полученных из образца крови пациента. Для терапевтического лечения можно использовать фармацевтические композиции для парентерального, локального, перорального, назального, интратекального или местного (например, в виде крема или мази для местного применения) введения. Предпочтительно фармацевтические композиции вводят парентерально, например внутривенно, подкожно, внутрикожно или внутримышечно. Следовательно, данное изобретение предлагает композиции для парентерального введения, которые содержат раствор или суспензию иммуногенных пептидов в подходящем носителе, предпочтительно в водном носителе. Можно использовать ряд водных носителей, например воду, забуференную воду, 0,8% физиологический раствор, 0,3% раствор глицина, гиаулуроновой кислоты и т.п. Данные композиции можно стерилизовать традиционными, хорошо известными методами, в том числе фильтрованием. Полученные водные растворы можно фасовать как есть или в виде лиофилизованных препаратов, которые перед введением объединяют со стерильным раствором. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, необходимые для приближения композиции к физиологическим условиям, такие как средства, используемые для доведения рН, и забуферивающие средства, средства, обеспечивающие нужную тоничность, увлажняющие средства, консерванты и т.п., например ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция, монолаурат сорбитана, олеат триэтаноламина и др. Концентрация пептидов согласно изобретению в фармацевтических композициях может широко варьировать, например от менее чем приблизительно 0,1 мас.%, обычно по меньшей мере приблизительно 2 мас.%, до 20-50 мас.% или более, ее выбор, в первую очередь, зависит от объема жидкости, вязкости и т.п. в соответствии с выбранным способом введения. Человеческая стандартная лекарственная форма композиции обычно включена в фармацевтическую композицию, которая содержит разовую дозу приемлемого для человека носителя, в одном воплощении водного носителя, ее вводят в объеме/количестве, обычно используемом специалистами в данной области для введения таких композиций людям (см., например, Remington's Pharmaceutical Sciences, 17 th Edition, A. Gennaro, Editor, Mack Publishing Co., Easton, Pennsylvania, 1985). Например, для пациента массой 70 кг доза пептида при начальной иммунизации может составлять приблизительно от 1 до 50000 мкг, как правило, 100-5000 мкг. Например, в случае нуклеиновых кислот начальную иммунизацию можно проводить путем в.м. (или п.к., или в.к.) введения вектора экспрессии в виде оголенной нуклеиновой кислоты в количестве 0,5-5 мг в разные участки. Нуклеиновую кислоту (от 0,1 до 1000 мкг) также можно вводить методом генной пушки. По истечении 3-4-недельного инкубационного периода вводят поддерживающую дозу. В качестве поддерживающего препарата можно вводить рекомбинантный вирус птичьей оспы в дозе от 5 ×10 7 до 5 ×10 9 бое (бляшкообразующих единиц). В случае антител лечение обычно включает в себя повторяющееся введение препарата антител против STEAP-1 посредством приемлемого способа введения, такого как внутривенное введение (в.в.), в дозе, варьирующей приблизительно от 0,1 до 10 мг/кг массы тела. Как правило, дозы в интервале 10-500 мг Mab в неделю являются эффективными и хорошо переносятся. Приемлемый дозировочный режим включает в себя в.в. введение начальной ударной дозы препарата Mab против STEAP-1, составляющей приблизительно 4 мг/кг массы тела пациента, и последующее еженедельное в. в. введение дозы, составляющей приблизительно 2 мг/кг. Специалистам в данной области известно, что выбор идеальной дозы для каждого конкретного случая зависит от разных факторов. Такие факторы включают в себя, например, период полужизни композиции, сродство связывания Ab, иммуногенность вещества, степень экспрессии STEAP-1 у пациента, степень сбрасывания антигена STEAP-1 в кровоток, желательный уровень равновесной концентрации, частота введения, влияние химиотерапевтических или других средств, используемых в сочетании со способом лечения согласно изобретению, а также состояние здоровья конкретного пациента. Неограничивающие предпочтительные человеческие разовые дозы составляют, например, 500 мкг-1 мг, 1-50 мг, 50-100 мг, 100-200 мг, 200-300 мг, 400-500 мг, 500-600 мг, 600-700 мг, 700-800 мг, 800-900 мг, 900 мг-1 г или 1-700 мг. В некоторых воплощениях доза составляет 2-5 мг/кг массы тела, например, с последующим еженедельным введением дозы 1-3 мг/кг; 0,5 мг, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 мг/кг массы тела, например, с последующим еженедельным введением дозы в течение двух, трех или четырех недель; 0,5-10 мг/кг массы тела, например, с еженедельным введением дозы в течение двух, трех или четырех недель; 225, 250, 275, 300, 325, 350, 375, 400 мг/м 2 площади тела еженедельно; 1-600 мг/м 2 площади тела еженедельно; 225-400 мг/м 2 площади тела еженедельно; указанные дозы могут сопровождаться еженедельным введением доз в течение 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более недель. В одном воплощении стандартные лекарственные формы для применения человеком содержат полинуклеотиды в подходящем интервале доз или в эффективном количестве, обеспечивающем какой-либо терапевтический эффект. Рядовому специалисту в данной области известно, что терапевтический эффект зависит от ряда факторов, включающих в себя последовательность полинуклеотида, молекулярную массу полинуклеотида и способ введения. Дозы обычно выбирает врач или другой специалист здравоохранения в соответствии с рядом параметров, известных в данной области, таких как тяжесть симптомов, история болезни пациента и т.п. Как правило, дозировочный интервал для полинуклеотида размером приблизительно 20 оснований может иметь нижнюю границу, независимо выбранную, например, из таких значений, как приблизительно 0,1, 0,25, 0,5, 1, 2, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400 или 500 мг/кг, и верхнюю границу, независимо выбранную, например, из таких значений, как приблизительно 60, 80, 100, 200, 300, 400, 500, 750, 1000, 1500, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000 или 10000 мг/кг, при условии, что верхняя граница превышает нижнюю. Например, доза может находиться в любом из следующих приблизительных интервалов: 0,1-100, 0,1-50, 0,1-25, 0,1-10, 1-500, 100-400, 200-300, 1-100, 100-200, 300-400, 400-500, 500-1000, 500-5000 или 500-10000 мг/кг. Как правило, при парентеральных способах введения требуются более высокие дозы полинуклеотида, чем при более направленном введении непосредственно в больную ткань, как в случае более длинных полинуклеотидов. В одном воплощении стандартные лекарственные формы для применения человеком содержат Т-клетки в подходящем интервале доз или в эффективном количестве, обеспечивающем какой-либо терапевтический эффект. Рядовому специалисту в данной области известно, что терапевтический эффект зависит от ряда факторов. Дозы обычно выбирает врач или другой специалист здравоохранения в соответствии с рядом параметров, известных в данной области, таких как тяжесть симптомов, история болезни пациента и т.п. Доза может составлять приблизительно от 10 4 до 10 6 клеток, приблизительно от 10 6 до 10 8 клеток, приблизительно от 10 8 до 10 11 клеток или приблизительно от 10 8 до 5 ×10 10 клеток. Доза также может составлять приблизительно от 10 6 до 10 10 клеток/м 2 или приблизительно от 10 6 до 10 8 клеток/м 2 . Белок (белки) согласно изобретению и/или нуклеиновые кислоты, кодирующие данный белок (белки), также можно вводить с использованием липосом, которые могут служить для 1) направления белка (белков) к конкретной ткани, такой как лимфоидная ткань; 2) избирательного направления к больным клеткам или 3) увеличения периода полужизни пептидной композиции. Препараты липосом включают в себя эмульсии, пенки, мицеллы, нерастворимые монослои, жидкие кристаллы, дисперсии фосфолипидов, ламеллярные слои и т.п. В данных препаратах доставляемый пептид включен в липосому, один или в сочетании с молекулой, которая связывается с рецептором, распространенным среди лимфоидных клеток, такой как моноклональные антитела, связывающиеся с антигеном CD45, или в сочетании с другими терапевтическими или иммуногенными композициями. Так, липосомы, либо содержащие внутри, либо экспонирующие целевой пептид согласно изобретению, можно направить к лимфоидным клеткам, куда липосомы доставляют пептидные композиции. Липосомы для применения в соответствии с данным изобретением получают из стандартных везикулообразующих липидов, которые обычно включают в себя нейтральные и отрицательно заряженные фосфолипиды и стерин, такой как холестерин. Как правило, выбор липидов зависит от таких факторов, как, например, размер липосом, устойчивость к воздействию кислот и стабильность липосом в кровотоке. Для получения липосом можно использовать разные способы, например, описанные в Szoka, et al., Ann. Rev. Biophys. Bioeng. 9: 467 (1980) и в патентах США № 4235871, 4501728, 4837028 и 5019369. Чтобы направить липосомы к клеткам иммунной системы, в их состав можно ввести лиганд, включающий в себя, например, антитела или их фрагменты, специфичные к поверхностным детерминантам целевых клеток иммунной системы. Суспензию липосом, содержащую пептид, можно вводить внутривенно, локально, местно и т.п. в дозе, которая варьирует в зависимости от способа введения, типа доставляемого пептида, стадии заболевания, подлежащего лечению, и т.п. Для получения твердых композиций можно использовать традиционные нетоксичные твердые носители, которые включают в себя такие носители фармацевтической категории, как, например, маннит, лактоза, крахмал, стеарат магния, сахарин натрия, тальк, целлюлоза, глюкоза, сахароза, карбонат магния и т.п. Фармацевтически приемлемую нетоксичную композицию для перорального введения получают с использованием любых традиционно использующихся наполнителей, таких как перечисленные ранее носители, такая композиция обычно содержит 10-95% активного ингредиента, включающего в себя один или несколько пептидов согласно изобретению, более предпочтительно 25-75%. Иммуногенные пептиды для аэрозольного введения предпочтительно предоставляют в мелкоизмельченном виде вместе с поверхностно-активным веществом и пропеллентом. При этом доля пептидов обычно составляет приблизительно 0,01-20 мас.%, предпочтительно приблизительно 1-10 мас.%. Разумеется, поверхностно-активное вещество должно быть нетоксичным и предпочтительно растворимым в пропелленте. Типичными примерами таких веществ являются сложные эфиры или частичные сложные эфиры жирных кислот, содержащих приблизительно от 6 до 22 атомов углерода, таких как капроновая, октановая, лауриновая, пальмитиновая, стеариновая, линолевая, линоленовая, олестериновая и олеиновая кислоты, и алифатического многоатомного спирта или его циклического ангидрида. Можно использовать смешанные сложные эфиры, такие как смешанные или природные глицериды. Масса поверхностно-активного вещества может составлять приблизительно 0,1-20% по отношению к массе композиции, предпочтительно приблизительно 0,25-5%. Остальную часть композиции обычно составляет пропеллент. При желании в состав композиции для интраназальной доставки также может быть включен носитель, такой как, например, лецитин. XI. Диагностические и прогностические воплощения STEAP-1. Как указано в данном описании, полинуклеотиды STEAP-1, полипептиды, реакционноспособные цитотоксичные Т-клетки (CTL), реакционноспособные хелперные Т-клетки (HTL) и антитела против полипептидов используют в хорошо известных диагностических, прогностических и терапевтических анализах, позволяющих диагностировать состояния, связанные с нарушением регуляции клеточного роста, такие как рак, в частности злокачественные заболевания, перечисленные в табл. I (см., например, специфический характер тканевой экспрессии, а также сверхэкспрессию при некоторых злокачественных заболеваниях, как описано, например, в примере, озаглавленном "Анализ экспрессии STEAP-1 в нормальных тканях и в образцах, полученных от пациентов"). STEAP-1 можно сравнить с антигеном, ассоциированным с простатой PSA, исконным маркером, используемым врачами-терапевтами в течение многих лет для идентификации и мониторинга рака простаты (см., например, Merrill et al., J. Urol. 163(2): 503-5120 (2000); Polascik et al., J. Urol. Aug; 162(2): 293-306 (1999) и Fortier et al., J. Nat. Cancer Inst. 91(19): 1635-1640 (1999)). Для данной цели также используют ряд других диагностических маркеров, в том числе р53 и K-ras (см., например, Tulchinsky et al., Int. J. Mol. Med. 1999 Jul., 4(1): 99-102 и Minimoto et al., Cancer Detect Prev. 2000, 24(1): 1-12). Следовательно, данное описание полинуклеотидов STEAP-1 и полипептидов (а также полинуклеотидных зондов STEAP-1 и антител против STEAP-1, используемых для идентификации присутствия данных молекул), а также их свойств позволит опытным специалистам использовать данные молекулы в способах, аналогичных используемым, например, в ряде диагностических анализов, направленных на диагностику состояний, связанных с раком. Типичные воплощения диагностических способов, в которых используются полинуклеотиды STEAP-1, полипептиды STEAP-1, специфичные к STEAP-1 Т-клетки и антитела против STEAP-1, аналогичны традиционным способам диагностических анализов, в которых используются, например, полинуклеотиды PSA, полипептиды PSA, специфичные к PSA Т-клетки и антитела против PSA. Например, так же, как и полинуклеотиды PSA, которые используются в качестве зондов (например, в анализе методом нозерн-блоттинга, см., например, Sharief et al., Biochem. Mol. Biol. Int. 33(3): 567-74 (1994)) и праймеров (например, в анализе ПЦР, см., например, Okegawa et al., J. Urol. 163(4): 1189-1190 (2000)) при определении присутствия и/или уровня мРНК PSA в методах мониторинга сверхэкспрессии PSA или метастаз рака простаты, описанные в данном документе полинуклеотиды STEAP-1 можно использовать аналогично для детектирования сверхэкспрессии STEAP-1 или метастаз рака простаты и других злокачественных заболеваний, сопровождающихся экспрессией данного гена. Альтернативно, так же, как и полипептиды PSA, которые используются для стимуляции образования антител, специфичных к PSA, которые затем можно использовать для определения присутствия и/или уровня белков PSA в методах мониторинга сверхэкспрессии белка PSA (см., например, Stephan et al., Urology, 55(4): 560-3 (2000)) или метастаз клеток опухоли простаты (см., например, Alanen et al., Pathol. Res. Pract. 192(3): 233-7 (1996)), описанные в данном документе полипептиды STEAP-1 можно использовать для стимуляции образования антител, применяющихся для детектирования сверхэкспрессии STEAP-1 или метастаз клеток опухоли простаты, а также клеток других раковых опухолей, экспрессирующих данный ген. А именно, поскольку метастазирование включает в себя перемещение раковых клеток из органа первоначальной опухоли (такого как легкое, предстательная железа и др.) в другие области организма (такие как лимфатические узлы), анализы биологического образца на присутствие клеток, экспрессирующих полинуклеотиды и/или полипептиды STEAP-1, можно использовать для обнаружения метастаз. Так, присутствие STEAP-1-экспрессирующих клеток в биологическом образце, который обычно их не содержит (лимфатический узел), например обнаружение экспрессии STEAP-1 в LAPC4 и LAPC9, ксенотрансплантатах, выделенных из лимфатического узла и костных метастазов соответственно, указывает на наличие метастаз. Альтернативно, полинуклеотиды и/или полипептиды STEAP-1 можно использовать для диагностики рака, например, если обнаруживается, что клетки в биологическом образце, которые обычно не экспрессируют STEAP-1 или экспрессируют его на другом уровне, экспрессируют STEAP-1 или экспрессируют его на повышенном уровне (см., например, уровни экспрессии STEAP-1 при злокачественных заболеваниях, перечисленных в табл. I, a также в образцах пациентов и др., приведенные на прилагающихся чертежах). С помощью таких анализов специалисты могут получить дополнительные доказательства метастазирования путем тестирования биологического образца на присутствие другого тканеспецифического маркера (в дополнение к STEAP-1), такого как PSA, PSCA и др. (см., например, Alanen et al., Pathol. Res. Pract. 192(3): 237 (1996)). Детектирование присутствия полипептида STEAP-1 в срезе ткани с помощью иммуногистохимического анализа может указывать на изменение состояния некоторых клеток в данной ткани. В данной области хорошо известно, что способность антитела локализовывать полипептид, экспрессирующийся в раковых клетках, используется для диагностики заболевания, стадии заболевания, развития и/или агрессивности опухоли. С помощью такого антитела также можно детектировать изменение распределения полипептида в раковых клетках по сравнению с соответствующей немалигнизированной тканью. Полипептид STEAP-1 и иммуногенные композиции также используются для детектирования изменения внутриклеточной локализации при болезненных состояниях. Изменение состояния клеток из нормального на болезненное сопровождается изменениями в клеточной морфологии и зачастую связано с изменениями во внутриклеточной локализации/внутриклеточном распределении белка. Например, клеточные мембранные белки, которые в нормальных клетках экспрессируются в поляризованном виде, при возникновении болезненного состояния могут измениться, что приводит к распределению белка в неполяризованном виде на всей поверхности клетки. Изменение внутриклеточной локализации белка в болезненном состоянии было продемонстрировано для белков MUC1 и HER2 с помощью иммуногистохимического анализа. Нормальные эпителиальные клетки имеют обычное поверхностное распределение MUC1 при небольшой локализации гликопротеина на поверхности ядра, тогда как злокачественные клетки часто демонстрируют аполярный характер окрашивания (Diaz et al., The Breast Journal, 7: 40-45 (2001); Zhang et al., Clinical Cancer Research, 4: 2669-2676 (1998): Cao, et al., The Journal of Histochemistry and Cytochemistry, 45: 1547-1557 (1997)). Кроме того, клетки нормального эпителия молочной железы либо являются отрицательными по белку HER2, либо имеют базолатеральное распределение данного белка, тогда как в злокачественных клетках экспрессирующийся белок распределяется по всей клеточной поверхности (De Potter, et al., International Journal of Cancer, 44: 969-974 (1989); McCormick, et al., 117: 935-943 (2002)). Альтернативно, в больном состоянии распределение белка может измениться с исключительно поверхностной локализации на диффузную цитоплазматическую экспрессию. Примером такого белка может служить MUC1 (Diaz, et al., The Breast Journal, 7: 40-45 (2001)). Изменение локализации/распределения белка в клетке, определяемое гистохимическими методами, также может предоставить информацию, имеющую значение для выбора способов лечения. Последнее применение иллюстрируется случаем, где белок в нормальной ткани находится внутри клетки, а в малигнизированной ткани - на поверхности клетки; локализация белка на поверхности клеток делает клетки доступными для антител, применяющихся в диагностических и терапевтических способах. Если такое изменение локализации белка происходит в случае STEAP-1, белок STEAP-1 и связанные с ним иммунные ответы также можно использовать в указанных способах. Соответственно, очень важно определить, действительно ли изменение внутриклеточной локализации белка, наблюдающееся для 24Р4С12, делает белок STEAP-1 и связанные с ним иммунные ответы очень полезными. Применение композиций STEAP-1 позволяет специалистам в данной области решать важные диагностические и терапевтические проблемы. Иммуногистохимические реагенты, специфичные к STEAP-1, также можно использовать для детектирования метастаз опухолей, экспрессирующих STEAP-1, в том случае, если полипептид STEAP-1 обнаруживается в тканях, в которых он обычно не продуцируется. Таким образом, полипептиды STEAP-1 и антитела, полученные путем индуцирования иммунных ответов на указанные полипептиды, можно использовать для достижения разных целей, включающих в себя диагностические, прогностические, профилактические и/или терапевтические задачи, известные специалистам в данной области. Полинуклеотидные фрагменты и варианты STEAP-1 применяются аналогично полинуклеотидным фрагментам и полинуклеотидным вариантам PSA, используемым специалистами в данной области в способах мониторинга PSA. В частности, полинуклеотиды PSA обычно используются в способах мониторинга PSA в виде зондов или праймеров, которые состоят из фрагментов последовательности кДНК PSA. Например, праймеры, используемые в полимеразной цепной реакции для амплификации полинуклеотида PSA, должны содержать фрагмент, размер которого меньше полной последовательности PSA. Для проведения ПЦР опытные специалисты обычно получают ряд полинуклеотидных фрагментов, которые можно использовать в качестве праймеров для амплификации разных фрагментов представляющего интерес полинуклеотида или для оптимизации реакций амплификации (см., например, Caetano-Anolles, G. Biotechniques, 25(3): 472-476, 478-480 (1998); Robertson et al., Methods Mol. Biol. 98: 121-154 (1998)). Применение таких фрагментов дополнительно иллюстрируется в примере, озаглавленном "Анализ экспрессии STEAP-1 в нормальных тканях и в образцах, полученных от пациентов", где полинуклеотидный фрагмент STEAP-1 используют в качестве зонда при определении экспрессии молекул РНК STEAP-1 в раковых клетках. Кроме того, в качестве праймеров и зондов для соответствующих мРНК в методах ПЦР и нозерн-блоттинга используют вариантные нуклеотидные последовательности (см., например, Sawai et al., Fetal Diagn. Ther. 1996. Nov.-Dec. 11(6): 407-13 и Current Protocols In Molecular Biology. Vol. 2. Unit 2. Frederick M. Ausubel et al. eds., 1995). Для данной цели используют полинуклеотидные фрагменты и варианты, способные связываться с целевой полинуклеотидной последовательностью (например, с полинуклеотидом STEAP-1, изображенным на фиг. 2, или с его вариантом) в условиях высокой жесткости. Далее, полипептиды PSA, содержащие эпитоп, который может распознаваться антителом или Т-клеткой, специфически связывающейся с данным эпитопом, используют в способах мониторинга PSA. Фрагменты и аналоги полипептидов STEAP-1 можно использовать аналогичным способом. В данной области для получения антител (например, антител или Т-клеток, специфичных к PSA) практикующие специалисты обычно используют фрагменты или варианты полипептидов с использованием разных систем, таких как гибридные белки (см., например, Current Protocols In Molecular Biology. Vol. 2. Unit 16. Frederick M. Ausubel et al. eds., 1995). В данном контексте каждый эпитоп (эпитопы) должен иметь конфигурацию, обеспечивающую взаимодействие с антителом или Т-клеткой. Обычно специалисты в данной области получают разные полипептидные фрагменты, которые можно использовать для инициации иммунных ответов, специфичных для разных участков представляющего интерес полипептида (см., например, патенты США № 5840501 и 5939533). Например, иногда предпочтительно использовать полипептид, содержащий один из описанных в данном документе мотивов STEAP-1 или мотив-несущую субпоследовательность, которые специалист в данной области может легко идентифицировать, используя известные мотивы. В данном контексте используют полипептидные фрагменты, варианты или аналоги, которые еще содержат эпитоп, способный индуцировать образование антител или Т-клеток, специфичных к целевой полипептидной последовательности (например, к полипептиду STEAP-1, изображенному на фиг. 3). Как указано в данном описании, полинуклеотиды и полипептиды STEAP-1 (а также полинуклеотидные зонды STEAP-1 и антитела против STEAP-1 или Т-клетки, специфичные к STEAP-1, используемые для детектирования присутствия данных молекул) обладают специфическими свойствами, которые позволяют использовать их для диагностики злокачественных заболеваний, таких как приведенные в табл. I. Диагностические анализы, которые позволяют определять присутствие генных продуктов STEAP-1 и посредством этого диагностировать наличие или появление болезненного состояния, описанного в данном документе, такого как рак простаты, используют для выявления пациентов, нуждающихся в профилактических мерах, или для их последующего мониторинга, как и диагностические анализы PSA, которые успешно применяются в настоящее время. Более того, указанные вещества удовлетворяют существующей в данной области потребности в молекулах, имеющих подобные или комплементарые PSA характеристики, в тех случаях, когда, например, окончательный диагноз метастаз простатического происхождения не может быть поставлен на основе анализа одного PSA (см., например, Alanen et al., Pathol. Res. Pract. 192(3): 233-237 (1996)), следовательно, такие вещества, как полинуклеотиды и полипептиды STEAP-1 (а также полинуклеотидные зонды STEAP-1 и антитела против STEAP-1, используемые для детектирования присутствия данных молекул), необходимы для подтверждения метастаз простатического происхождения. Наконец, помимо применения в диагностических анализах, полинуклеотиды STEAP-1, раскрытые в данном описании, имеют ряд других применений, например, для идентификации связанных с онкогенезом хромосомных нарушений в участке хромосомы, к которому относят STEAP-1 (см. приведенный ниже пример под заголовком "Хромосомное картирование STEAP-1"). Кроме того, помимо применения в диагностических анализах, STEAP-1-родственные белки и полинуклеотиды, раскрытые в данном описании, имеют ряд других применений, например, в судебно-медицинских анализах тканей неизвестного происхождения (см., например, Takahama K. Forensic Sci. Int. 1996, Jun. 28; 80(1-2): 63-9). Кроме того, STEAP-1-родственные белки или полинуклеотиды согласно изобретению можно использовать для лечения патологического состояния, характеризующегося сверхэкспрессией STEAP-1. Например, аминокислотную или нуклеотидную последовательность, приведенную на фиг. 2 или 3, или их фрагменты, можно использовать для индуцирования иммунного ответа на антиген STEAP-1. Антитела или другие молекулы, которые взаимодействуют со STEAP-1, можно использовать для модуляции функции данной молекулы и получения терапевтического эффекта, опосредованного данной функцией. XII. Ингибирование белковой функции STEAP-1. Данное изобретение включает в себя различные способы и композиции, направленные на ингибирование связывания STEAP-1 с его партнером по связыванию или его ассоциации с другим белком (белками), а также способы ингибирования функции STEAP-1. XII.А. Ингибирование STEAP-1 внутриклеточными антителами. В одном способе рекомбинантный вектор, кодирующий одноцепочечные антитела, которые специфически связываются со STEAP-1, вводят в STEAP-1-экспрессирующие клетки с помощью методов, использующихся для переноса генов. Соответственно, кодируемое вектором одноцепочечное антитело против STEAP-1 экспрессируется внутри клетки, связывается с белком STEAP-1 и посредством этого ингибирует его функцию. Генно-инженерные методы получения таких внутриклеточных одноцепочечных антител хорошо известны. Такие внутриклеточные антитела, также известные как "интратела", направляются к конкретному компартменту клетки, обеспечивая контроль над областью фокусирования ингибиторной активности лекарственного средства. Указанная технология успешно применяется в данной области (обзор можно найти в Richardson and Marasco, 1995, TIBTECH, vol. 13). Показано, что интратела действительно нивелируют экспрессию клеточных поверхностных рецепторов, которые обычно присутствуют в большом количестве (см., например, Richardson et al., 1995, Proc. Natl. Acad. Sci. USA 92: 3137-3141; Beerli et al., 1994, J. Biol. Chem. 289: 23931-23936; Deshane et al., 1994, Gene Ther. 1: 332-337). Одноцепочечные антитела включают в себя вариабельные домены тяжелой и легкой цепей, соединенные гибким линкерным полипептидом, экспрессирующиеся в одном полипептиде. Необязательно, одноцепочечные антитела экспрессируются в виде одноцепочечного фрагмента вариабельного участка, соединенного с константным участком легкой цепи. Чтобы точно направить интратело к целевому внутриклеточному компартменту, в состав рекомбинантных полинуклеотидных векторов, кодирующих такие одноцепочечные антитела, вводят с помощью методов генной инженерии хорошо известные сигналы внутриклеточного переноса. Например, в состав антител, направляемых к эндоплазматическому ретикулуму (ER), включают лидерный пептид и, необязательно, С-концевой сигнал удерживания ER, такой как аминокислотный мотив KDEL. В состав интрател, активность которых реализуется в клеточных ядрах, вводят сигнал ядерной локализации. Чтобы привязать интратело к цитозольной стороне клеточной мембраны, в его состав вводят липидные фрагменты. Функция интрател также может реализовываться в цитозоле. Например, цитозольные интратела используют для изоляции факторов, находящихся в цитозоле, и предотвращения посредством этого их транспортировки к месту назначения. В одном воплощении интратела используют для улавливания STEAP-1 в ядрах, препятствуя таким образом проявлению его активности в данном компартменте клетки. Чтобы достичь нужной направленности, в такие интратела STEAP-1 с помощью методов генной инженерии вводят сигналы направления в ядро. Полученные указанными способами интратела STEAP-1 специфически связываются с конкретным доменом STEAP-1. В другом воплощении с помощью цитозольных интрател, которые специфически связываются с белком STEAP-1, предотвращают проникновение STEAP-1 в ядра и, посредством этого, реализацию биологической активности STEAP-1 в ядрах (например, предотвращают образование транскрипционных комплексов STEAP-1 с другими факторами). Чтобы осуществить экспрессию таких интрател в конкретных клетках, для регуляции транскрипции используют подходящий опухолеспецифический промотор и/или энхансер. Например, чтобы осуществить экспрессию интратела именно в простате, можно использовать промотор и/или промотор/энхансер PSA (см., например, патент США № 5919652, выданный 6 июля 1999 г.). XII.В. Ингибирование STEAP-1 рекомбинантными белками. В другом способе рекомбинантные молекулы связываются со STEAP-1 и посредством этого ингибируют функцию STEAP-1. Например, указанные рекомбинантные молекулы предотвращают доступ STEAP-1 к партнеру (партнерам) по связыванию, или ингибируют связывание STEAP-1 с партнером (партнерами) по связыванию, или предотвращают или ингибируют образование ассоциации STEAP-1 с другим белком (белками). Такие рекомбинантные молекулы могут содержать, например, реакционноспособную часть (части) STEAP-1-специфичной молекулы антитела. В конкретном воплощении STEAP-1-связывающий домен партнера STEAP-1 по связыванию входит в состав полученного рекомбинантными методами димерного гибридного белка, в результате чего гибридный белок содержит два домена, связывающих лиганд STEAP-1, соединенных с фрагментом Fc человеческого IgG, такого как человеческий IgG 1 . Такой фрагмент IgG может содержать, например, домены С Н 2 и С Н 3 и шарнирный участок, но не домен С Н 1. Такие димерные гибридные белки вводят в растворимой форме пациентам, страдающим от рака, связанного с экспрессией STEAP-1, после чего данные белки специфически связываются со STEAP-1 и блокируют взаимодействие STEAP-1 с партнером по связыванию. Димерные гибридные белки можно также объединить в мультимерные белки с помощью известных методов соединения антител. XII.С. Ингибирование транскрипции или трансляции STEAP-1. Настоящее изобретение также включает в себя разные способы и композиции, направленные на ингибирование транскрипции гена STEAP-1. Аналогично, данное изобретение предлагает способы и композиции, направленные на ингибирование трансляции мРНК STEAP-1 с образованием белка. Один способ ингибирования транскрипции гена STEAP-1 включает в себя приведение в контакт гена STEAP-1 с антисмысловым полинуклеотидом STEAP-1. Другой способ ингибирования трансляции мРНК STEAP-1 включает в себя приведение в контакт мРНК STEAP-1 с антисмысловым полинуклеотидом. В следующем способе используют STEAP-1-специфичный рибозим, который отщепляет транскрипт STEAP-1 и в результате этого ингибирует трансляцию. Такие способы с использованием антисмысловых последовательностей и рибозимов также могут быть направлены на регуляторные участки гена STEAP-1, такие как промоторные и/или энхансерные элементы STEAP-1. Подобным образом, для ингибирования транскрипции мРНК STEAP-1 можно использовать белки, способные ингибировать фактор транскрипции гена STEAP-1. Различные полинуклеотиды и композиции, используемые в приведенных выше сособах, описаны выше. Применение антисмысловых молекул и рибозимов для ингибирования транскрипции и трансляции широко известно в данной области. Другие факторы, ингибирующие транскрипцию STEAP-1 путем нарушения активации транскрипции STEAP-1, также можно использовать для лечения злокачественных заболеваний, сопровождающихся экспрессией STEAP-1. Подобным образом, для лечения злокачественных заболеваний, сопровождающихся экспрессией STEAP-1, можно использовать факторы, препятствующие процессингу STEAP-1. Способы лечения рака, в которых используются такие факторы, также входят в объем согласно изобретению. XII.D. Общие принципы терапевтических стратегий. Способы переноса гена и генной терапии можно использовать для доставки терапевтических полинуклеотидных молекул к опухолевым клеткам, синтезирующим STEAP-1 (т.е. антисмысловых молекул, рибозимов, полинуклеотидов, кодирующих интратела, и других молекул, ингибирующих STEAP-1). В данной области известны разные способы генной терапии. Рекомбинантные векторы, кодирующие антисмысловые полинуклеотиды STEAP-1, рибозимы, факторы, препятствующие транскрипции STEAP-1, и т.д., можно доставить к целевым опухолевым клеткам с помощью таких способов генной терапии. Вышеуказанные терапевтические способы можно объединить с любым из широкого ряда хирургических, химиотерапевтических и традиционных способов лечения. Терапевтические способы согласно изобретению позволяют использовать пониженные дозы химиотерапевтических средств (или других лекарственных препаратов) и/или уменьшить частоту введения, это является преимуществом для всех пациентов, но особенно для тех, которые плохо переносят токсичность химиотерапевтических средств. Противоопухолевую активность конкретной композиции (например, содержащей антисмысловую молекулу, рибозим или интратело) или сочетания таких композиций можно оценить с помощью разных аналитических систем in vitro и in vivo. Анализы in vitro, используемые для определения терапевтической активности, включают в себя анализы клеточного роста, анализы с применением мягкого агара и другие анализы, позволяющие определить опухоль-стимулирующую активность, анализы связывания, позволяющие определить степень ингибирования связывания STEAP-1 с партнером по связыванию под действием терапевтической композиции, и др. Эффект терапевтической композиции STEAP-1 in vivo можно оценить на подходящей животной модели. Например, можно использовать модели ксеногенного рака простаты, где эксплантаты рака простаты человека или пересаженную ткань ксенотрансплантата вводят иммунокомпромиссным животным, таким как голые мыши или мыши SCID (Klein et al., 1997, Nature Medicine, 3: 402-408). Например, в патентной заявке РСТ WO 98/16628 и патенте США № 6107540 описаны разные модели ксенотрансплантатов человеческого рака простаты, способные воспроизводить развитие первичных опухолей, микрометастаз и образование остеобластических метастаз, характерных для поздней стадии заболевания. Эффективность препаратов можно прогнозировать с помощью анализов ингибирования образования опухоли, регрессии опухоли или метастаз и т.п. Для тестирования терапевтических композиций можно использовать анализы in vivo, которые позволяют оценить индукцию апоптоза. В одном воплощении ксенотрансплантаты из мышей, несущих опухоль, обработанных терапевтической композицией, можно анализировать на присутствие апоптотических очагов по сравнению с необработанными контрольными мышами, несущими ксенотрансплантат. Относительное количество апоптотических очагов, обнаруженных в опухолях обработанных мышей, соответствует терапевтической эффективности композиции. Терапевтические композиции, используемые при осуществлении вышеописанных способов, могут быть получены в виде фармацевтических композиций, содержащих носитель, подходящий для желательного способа доставки. Подходящие носители включают в себя любое вещество, которое при объединении с терапевтической композицией сохраняет противоопухолевую функцию терапевтической композиции и, как правило, не вызывает у пациента иммунного ответа. Примеры таких носителей включают в себя, без ограничения, любые стандартные фармацевтические носители, такие как забуференный фосфатом стерильный физиологический раствор, бактериостатическая вода и т.п. (общее описание можно найти в Remington's Pharmaceutical Sciences, 16 th Edition, A. Osal., Ed., 1980). Терапевтические композиции можно солюбилизировать и вводить любыми способами, обеспечивающими доставку терапевтической композиции к участку опухоли. Потенциально эффективные способы введения включают в себя, без ограничения, внутривенное, парентеральное, внутрибрюшинное, внутримышечное, внутриопухолевое, внутрикожное, внутриорганное, ортотопическое и т.п. введение. Предпочтительная композиция для внутривенного введения включает в себя раствор терапевтической композиции в обработанной консервантами бактериостатической воде, стерильной не обработанной консервантами воде и/или может находиться в поливинилхлоридных или полиэтиленовых пакетах в виде раствора в 0,9% стерильном растворе хлорида натрия для инъекций, USP. Терапевтические белковые препараты можно лиофилизировать и хранить в виде стерильных порошков, предпочтительно в вакууме, которые перед введением растворяют в бактериостатической воде (содержащей, например, бензиловый спирт в качестве консерванта) или в стерильной воде. Дозы и режимы введения лекарственных средств для лечения злокачественных заболеваний с использованием вышеописанных способов зависят от выбранного способа лечения, от конкретного типа рака, а также от ряда других факторов, известных в данной области. XIII. Идентификация. Характеристика и применение модуляторов STEAP-1. Способы идентификации и применения модуляторов. В одном воплощении проводят скрининг для идентификации модуляторов, которые индуцируют или подавляют конкретный профиль экспрессии, подавляют или индуцируют специфические метаболические пути и предпочтительно генерируют связанный с их активностью фенотип. В другом воплощении, идентифицировав конкретные гены с дифференциальной экспрессией в зависимости от состояния, проводят скрининг для идентификации модуляторов, которые изменяют, либо увеличивая, либо уменьшая, экспрессию отдельных генов. В следующем воплощении проводят скрининг для идентификации модуляторов, которые изменяют биологическую функцию продукта гена с особым характером экспрессии. Опять же, идентифицировав значение гена для конкретного состояния, проводят скрининг для идентификации средств, которые связываются с продуктом гена и/или модулируют его биологическую активность. Кроме того, проводят скрининги генов, индуцируемых в ответ на средство-кандидат. После идентификации модулятора (либо модулятора, который снижает экспрессию в раковой ткани до нормального уровня, либо модулятора ракового гена, который обеспечивает экспрессию гена на уровне, наблюдающемся в нормальной ткани) проводят скрининг для идентификации генов, которые специфично модулируются в ответ на данное средство. При сравнении профилей экспрессии в нормальной ткани и в раковой ткани, обработанной лекарственным средством, обнаружены гены, которые не экспрессируются в нормальной ткани или в раковой ткани, но экспрессируются в ткани, обработанной лекарственным средством, и наоборот. Специфичные к данному средству последовательности идентифицируют и используют в способах, описанных в данном документе для раковых генов или белков. В частности, указанные последовательности и кодируемые ими белки используют для получения или идентификации клеток, обработанных лекарственным средством. Кроме того, получают антитела против белков, индуцируемых данным средством, и используют их для направления новых терапевтических средств к раковой ткани, подлежащей лечению. Идентификация модуляторов и скрининговые анализы. Анализы, связанные с экспрессией генов. Белки, нуклеиновые кислоты и антитела согласно изобретению используют в скрининговых анализах. Связанные с раком белки, антитела, нуклеиновые кислоты, модифицированные белки и клетки, содержащие данные последовательности, используют в таких скрининговых анализах, как анализ влияния лекарственных средств-кандидатов на "профиль экспрессии генов", профиль экспрессии полипептидов или на биологическую функцию. В одном воплощении анализы профилей экспрессии проводят предпочтительно в сочетании с методами скрининга высокой пропускной способности, позволяющими осуществлять мониторинг профилей экспрессии генов после лечения средством-кандидатом (например, Davis, G.F., et al., J. Biol. Screen, 7: 69 (2002); Zlokarnik, et al., Science, 279: 84-8 (1998); Heid, Genome Res, 6: 986-94, 1996). В скрининговых анализах используют раковые белки, антитела, нуклеиновые кислоты, модифицированные белки и клетки, содержащие нативные или модифицированные раковые белки или гены. То есть настоящее изобретение включает в себя способы скрининга композиций, которые модулируют раковый фенотип или физиологическую функцию ракового белка согласно изобретению. Скрининг проводят непосредственно по гену или путем оценки влияния средств-кандидатов на "профиль экспрессии гена" или на биологическую функцию. В одном воплощении анализ на профили экспрессии предпочтительно используют в сочетании с методами скрининга высокой пропускной способности, позволяющими осуществлять мониторинг после лечения средством-кандидатом, см. Zlokamik, приведено выше. Проводят ряд анализов, направленных на гены и белки согласно изобретению. При проведении анализов используют определенный уровень нуклеиновой кислоты или белка. То есть проводят скрининг тестируемых соединений на способность модулировать экспрессию гена, который был идентифицирован как ген, участвующий в повышающей регуляции злокачественного заболевания, или на способность связывать раковый белок согласно изобретению. Термин "модуляция" в данном контексте включает в себя повышение или снижение экспрессии гена. Предпочтительная степень модуляции зависит от исходного изменения экспрессии гена в опухолевой ткани по сравнению с нормальной и составляет по меньшей мере 10%, предпочтительно 50%, более предпочтительно 100-300% и в некоторых воплощениях 300-1000% или выше. Так, если экспрессия гена в ракой ткани в 4 раза выше, чем в нормальной, желательно достичь приблизительно 4-кратного уменьшения экспрессии; аналогично, при 10-кратном уменьшении экспрессии в ракой ткани по сравнению с нормальной желательно, чтобы тестируемое соединение уменьшило экспрессию в 10 раз. Модуляторы, которые усиливают экспрессию гена при злокачественном заболевании, тоже могут быть полезными, например, для модели повышающей регуляции в дальнейших анализах. Величину экспрессии гена определяют с помощью нуклеотидных зондов и количественного определения уровня экспрессии гена или, альтернативно, регистрируют генный продукт, например, посредством применения антител против ракового белка и стандартных иммунологических анализов. Для количественной оценки экспрессии также используют протеомики и методы разделения. Мониторинг экспрессии для идентификации соединений, которые модифицируют экспрессию гена. В одном воплощении мониторинг экспрессии гена, т.е. получение профиля экспрессии, проводят одновременно для ряда образцов. Такие профили обычно получают для одного или нескольких генов, приведенных на фиг. 2. В таком воплощении, например, зонды, полученные из раковых нуклеиновых кислот, присоединяют к биочипам, чтобы детектировать раковые последовательности в конкретной клетке и определять их количество. Альтернативно, можно использовать ПЦР. Например, в соответствующие лунки титрационного микропланшета помещают праймеры. Затем каждую лунку анализируют методом ПЦР. Для идентификации соединений, которые модифицируют экспрессию одной или нескольких последовательностей, ассоциированных с раком, например полинуклеотидной последовательности, приведенной на фиг. 2, проводят мониторинг экспрессии. Как правило, тестируемый модулятор добавляют к клеткам до анализа. Кроме того, скрининг также проводят для идентификации средств, которые модулируют злокачественное заболевание, модулируют раковые белки согласно изобретению, связываются с раковым белком согласно изобретению или препятствуют связыванию ракового белка согласно изобретению с антителом или другим партнером по связыванию. В одном воплощении способы скрининга с высокой пропускной способностью включают в себя использование библиотек, содержащих большое количество потенциальных терапевтических соединений (соединений-кандидатов). Затем такие "комбинаторные химические библиотеки" подвергают скринингу с помощью одного или нескольких анализов, чтобы идентифицировать компоненты библиотеки (отдельные химические соединения или подклассы соединений), которые обладают желательной специфической активностью. Идентифицированные таким образом соединения можно использовать в качестве традиционных "ведущих соединений" для скрининга или в качестве терапевтических средств. В некоторых воплощениях комбинаторные библиотеки потенциальных модуляторов подвергают скринингу на способность связывать раковый полипептид или модулировать его активность. Как правило, новые химические соединения с нужными свойствами получают путем идентификации химического соединения (называемого "ведущее соединение"), обладающего желательным свойством или активностью, например ингибирующей активностью, создания вариантов ведущего соединения и анализа свойств и активности данных вариантов. Зачастую для таких анализов используют способы скрининга с высокой пропускной способностью. Как указано выше, для тестирования модуляторов-кандидатов (например, белков, нуклеиновых кислот маленьких молекул) обычно используют мониторинг экспрессии генов. После добавления средства-кандидата клетки оставляют инкубироваться в течение некоторого периода времени, затем образец, содержащий целевую анализируемую последовательность, добавляют, например, к биочипу. При необходимости целевую последовательность получают с помощью известных методов. Например, образец обрабатывают известными буферами для лизиса, чтобы лизировать клетки, подвергают электропорации и др., используя, в зависимости от обстоятельств, очистку и/или амплификацию, например, методом ПЦР. Например, проводят транскрипцию in vitro, используя метки, ковалентно присоединенные к нуклеотидам. Обычно нуклеиновые кислоты метят биотин-FITC или РЕ, или су3, или су5. Чтобы обеспечить детектирование специфического связывания целевой последовательности с зондом, целевую последовательность можно пометить, например, флуоресцентным, хемилюминесцентным, химическим или радиоактивным сигналом. В качестве метки также можно использовать фермент, такой как щелочная фосфатаза или пероксидаза хрена, который в присутствии субстрата продуцирует детектируемый продукт. Альтернативно, в качестве метки можно использовать меченое соединение или маленькую молекулу, такую как ингибитор фермента, которая связывается с ферментом, но не вступает в реакции, катализируемые ферментом, или не изменяется под действием фермента. Метка также может представлять собой фрагмент молекулы или целое соединение, например эпитопный фрагмент или биотин, который специфически связывается со стретавидином. В случае использования биотина стрептавидин метят, как описано выше, с целью получения детектируемого сигнала от связанной целевой последовательности. Несвязанный меченый стрептавидин обычно удаляют перед анализом. Специалистам в данной области известно, что к указанным выше анализам относятся анализы с применением прямой гибридизации или "сэндвич-анализы", включающие в себя использование нескольких зондов, общее описание таких анализов можно найти в патентах США № 5681702; 5597909; 5545730; 5594117; 5591584; 5571670; 5580731; 5571670; 5591584; 5624802; 5635352; 5594118; 5359100; 5124246 и 5681697. В данном воплощении, как правило, целевую нуклеиновую кислоту получают по описанному выше способу и добавляют к биочипу, содержащему множество нуклеотидных зондов, в условиях, которые обеспечивают образование комплекса из гибридизованных молекул. В настоящем изобретении используют ряд условий гибридизации, включающих в себя указанные выше условия высокой, средней и низкой жесткости. Анализы проводят в условиях такой жесткости, которая обеспечивает образование гибридизационного комплекса меченого зонда только в присутствии целевой последовательности. Жесткость можно регулировать путем изменения шагового параметра, который представляет собой термодинамическую переменную и включает в себя, без ограничения, температуру, концентрацию формамида, концентрацию соли, концентрацию хаотропной соли, рН, концентрацию органического растворителя и др. Данные параметры также можно использовать для регуляции неспецифического связывания, как описано в патенте США № 5681697. Так, чтобы уменьшить неспецифическое связывание, желательно проводить некоторые стадии в условиях более высокой жесткости. Описанные в данном документе реакции можно проводить разными способами. Компоненты, участвующие в реакции, можно добавлять одновременно или последовательно в разном порядке, предпочтительные воплощения данных способов описаны ниже. Кроме того, реакционная смесь может содержать ряд других реагентов. Данные реагенты включают в себя соли, буферы, нейтральные белки, например альбумин, детергенты и др., которые используют для обеспечения оптимальной гибридизации и детектирования и/или уменьшения неспецифических или фоновых взаимодействий. По обстоятельствам, в зависимости от способа получения образца и чистоты целевой последовательности можно использовать реагенты, которые иным образом улучшают эффективность анализа, такие как ингибиторы протеаз, ингибиторы нуклеаз, противомикробные средства и др. Путем анализа полученных результатов определяют уровни экспрессии отдельных генов и изменение экспрессии при изменении состояния клеток с получением профиля экспрессии генов. Анализы биологической активности. Данное изобретение предлагает способы идентификации или скрининга соединения, которое модулирует активность связанного с раком гена или белка согласно изобретению. Указанные способы включают в себя добавление тестируемого соединения, как указано выше, к клетке, содержащей раковый белок согласно изобретению. Данные клетки содержат рекомбинантную нуклеиновую кислоту, кодирующую раковый белок согласно изобретению. В другом воплощении библиотеку средств-кандидатов анализируют на нескольких клетках. В одном аспекте анализы проводят в присутствии или в отсутствие либо при предварительном или последующем добавлении физиологических сигналов, например гормонов, антител, пептидов, антигенов, цитокинов, факторов роста, потенциалов действия, фармакологических средств, включающих в себя химиотерапевтические средства, радиоактивные средства, канцерогенные средства или другие клетки (т.е. межклеточные контакты). В другом примере анализ проводят на разных стадиях клеточного цикла. В данном способе идентифицируют соединения, которые модулируют гены или белки согласно изобретению. Соединения, обладающие фармакологической активностью, могут повышать активность ракового белка согласно изобретению или препятствовать реализации такой активности. После идентификации оценивают подобные структуры, чтобы выявить важные структурные признаки соединения. В одном воплощении предлагается способ модулирования (например, ингибирования) деления раковых клеток; данный способ включает в себя введение модулятора злокачественного заболевания. В другом воплощении предлагается способ модулирования (например, ингибирования) злокачественного заболевания; данный способ включает в себя введение модулятора злокачественного заболевания. В следующем воплощении предлагаются способы лечения раковых клеток или субъектов, страдающих от рака; данный способ включает в себя введение модулятора злокачественного заболевания. В одном воплощении предлагается способ модулирования статуса клетки, которая экспрессирует ген согласно изобретению. Используемый в данном описании термин "статус" включает в себя такие принятые в данной области параметры, как рост, пролиферация, жизнеспособность, функционирование, апоптоз, старение, локализация, ферментативная активность клетки, передача сигнала и др. В одном воплощении раковый ингибитор представляет собой антитело, как указано выше. В другом воплощении раковый ингибитор представляет собой антисмысловую молекулу. Как указано в настоящем описании, специалистам в данной области известен ряд анализов клеточного роста, пролиферации и метастазирования. Скрининг с высокой пропускной способностью для идентификации модуляторов. При проведении скрининга с высокой пропускной способностью можно применять анализы, используемые для идентификации подходящих модуляторов. Так, предпочтительные анализы позволяют детектировать повышение или ингибирование транскрипции ракового гена, ингибирование или увеличение экспрессии полипептида и ингибирование или увеличение активности полипептида. В одном воплощении модуляторы, участвующие в скрининге с высокой пропускной способностью, представляют собой белки, обычно природные белки или фрагменты природных белов. Так, например, можно использовать клеточные экстракты, содержащие белки, или белковые клеточные экстракты, подвергнутые случайному или направленному расщеплению ферментами. В данном способе для скрининга, используемого в способах согласно изобретению, получают библиотеки белков. В данном воплощении можно использовать библиотеки бактериальных, грибковых, вирусных белков и белков млекопитающих, причем последние являются предпочтительными, а человеческие белки являются особенно предпочтительными. Особенно полезные тестируемые соединения имеют функцию, связанную с классом белков, к которому принадлежит мишень, например, они представляют собой субстраты ферментов или лиганды рецепторов. Применение методов выращивания на мягком агаре и образования колоний для идентификации и характеристики модуляторов. Нормальные клетки растут после присоединения к твердому субстрату. Если клетки трансформируются, они утрачивают данное свойство фенотипа и растут неприсоединенными к субстрату. Например, трансформированные клетки могут расти в перемешиваемой суспензионной культуре или при суспендировании в полутвердой среде, такой как полутвердый или мягкий агар. После трансфицирования геном-супрессором опухоли трансформированные клетки могут восстановить нормальный фенотип и потребность в твердом субстрате для присоединения и роста. Анализы роста на мягком агаре или образования колоний используют для идентификации модуляторов раковых последовательностей, которые после экспрессии в клетках-хозяевах ингибируют аномальную пролиферацию и трансформацию клеток. Модулятор уменьшает или устраняет способность клеток расти суспендированными в твердой или полутвердой среде, такой как агар. Способы проведения анализов роста на мягком агаре или образования колоний в суспензии описаны в Freshney, Culture of Animal Cells a Manual of Basic Technique (3 rd ed., 1994). Описание таких способов также можно найти в методическом разделе Garkavtsev et al. (1996), приведено выше. Идентификация и характеристика модуляторов с помощью анализа контактного ингибирования и ограничения роста плотностью клеток. Рост нормальных клеток обычно имеет плоский и организованный характер, пока клетки не начнут контактировать друг с другом. Когда клетки начинают соприкасаться друг с другом, вступает в силу контактное ингибирование и рост прекращается. Однако у трансформированных клеток контактное ингибирование отсутствует, они продолжают расти и достигают высокой плотности в очаге дезорганизованного роста. Таким образом, трансформированные клетки достигают более высокой плотности насыщения, чем нормальные клетки. Морфологически это проявляется в образовании дезориентированного монослоя клеток или очага клеток. Альтернативно, степень мечения ( 3 Н)-тимидином при плотности насыщения, используемая для измерения ограничения роста плотностью, подобно МТТ или анализу с использованием аламара синего (Alamar blue), позволяет оценить способность клеток к пролиферации, а также способность модуляторов оказывать влияние на пролиферацию. См. Freshney (1994), приведено выше. После трансфицирования генами-супрессорами опухоли трансформированные клетки могут восстановить нормальный фенотип, способность к контактному ингибированию и начинают расти до более низкой плотности. Анализ степени мечения ( 3 Н)-тимидином при плотности насыщения является предпочтительным способом измерения ограничения роста плотностью. Трансформированные клетки-хозяева трансфицируют ассоциированной с раком последовательностью и выращивают в течение 24 ч при плотности насыщения в условиях неограничивающей среды. Процент клеток, меченных ( 3 Н) -тимидином, определяют по включенной радиоактивности, измеряемой в cpm (число импульсов в минуту). Независимый от контакта рост используют для идентификации модуляторов раковых последовательностей, вызывающих аномальную пролиферацию и трансформацию клеток. Модулятор уменьшает или устраняет независимый от контакта рост и возвращает клеткам нормальный фенотип. Идентификация и характеристика модуляторов путем анализа зависимости от факторов роста или сыворотки. Трансформированные клетки имеют более низкую зависимость от сыворотки, чем их нормальные аналоги (см., например, Temin, J. Natl. Cancer Inst. 37: 167-175 (1966); Eagle et al., J. Exp. Med. 131: 836-879 (1970)); Freshney, приведено выше. Отчасти это является следствием высвобождения разных факторов роста трансформированными клетками. Можно сравнить степень зависимости от факторов роста или от сыворотки у трансформированных клеток-хозяев и у контрольных клеток. Например, степень зависимости клетки от факторов роста или от сыворотки определяют в способах, используемых для идентификации и характеристики соединений, модулирующих ассоциированные с раком последовательности согласно изобретению. Идентификация и характеристика модуляторов путем измерения уровней опухолеспецифических маркеров. Опухолевые клетки высвобождают более высокие количества некоторых факторов (далее называемых "опухолеспецифические маркеры"), чем их нормальные аналоги. Например, активатор плазминогена (РА) высвобождается из человеческой глиомы на более высоком уровне, чем из нормальных клеток мозга (см., например, Gullino, Angiogenesis, Tumor Vascularization, and Potential Interference with Tumor Growth, in Biological Responses in Cancer, p. 178-184 (Mihich (ed.), 1985)). Подобным образом, фактор опухолевого ангиогенеза (TAF) высвобождается из опухолевых клеток на более высоком уровне, чем из их нормальных аналогов. См., например, Folkman, Angiogenesis and Cancer, Sem Cancer Biol. (1992)), а также высвобождение bFGF из опухолей эндотелиальной ткани (Ensoli, В. et al.). Разные методы, позволяющие измерять высвобождение данных факторов, описаны в Freshney (1994), приведено выше. Описание таких методов также можно найти в Unkless et al., J. Biol. Chem. 249: 4295-4305 (1974); Strickland & Beers, J. Biol. Chem. 251: 5694-5702 (1976); Whur et al., Br. J. Cancer. 42: 305-312 (1980); Gullino, Angiogenesis, Tumor Vascularization, and Potential Interference with Tumor Growth, in Biological Responses in Cancer, p. 178-184 (Mihich (ed.), 1985); Freshney, Anticancer Res. 5: 111-130 (1985). Например, уровни опухолеспецифических маркеров определяют в способах, используемых для идентификации и характеристики соединений, модулирующих ассоциированные с раком последовательности согласно изобретению. Идентификация и характеристика модуляторов путем измерения способности клеток внедряться в матригель Степень внедрения в матригель или в компонент клеточного матрикса можно использовать для идентификации и характеристики соединений, модулирующих ассоциированные с раком последовательности. Существует положительная корреляция между степенью злокачественности опухолевых клеток и степенью их проникновения в матригель или какой-нибудь другой компонент клеточного матрикса. В данном анализе онкогенные клетки используют в качестве клеток-хозяев. Экспрессия гена-супрессора опухоли в данных клетках-хояевах уменьшает инвазивность клеток-хозяев. Можно использовать методы, описанные в Cancer Res. 1999; 59: 6010; Freshney (1994), приведено выше. Коротко говоря, уровень проникновения клеток-хозяев определяют путем использования фильтров, покрытых матригелем или каким-нибудь другим компонентом клеточного матрикса. Инвазивность определяют как проникновение в гель или через удаленную сторону фильтра и рассчитывают гистологически путем определения числа клеток и пройденной дистанции или путем предварительного мечения клеток 125 I и подсчетом радиоактивности на удаленной стороне фильтра или на дне чашки. См., например, Freshney (1994), приведено выше. Идентификация и характеристика модуляторов путем измерения опухолевого роста in vivo Влияние ассоциированных с раком последовательностей на рост клеток анализируют на трансгенных организмах или организмах с подавленным иммунитетом. Трансгенные организмы получают с помощью ряда известных в данной области способов. Например, можно получить нокаутные трансгенные организмы, например млекопитающих, таких как мыши, у которых раковый ген выключают или вставляют. Нокаутных трансгенных мышей получают путем вставки маркерного гена или другого гетерологичного гена в участок эндогенного ракового гена мышиного генома посредством гомологичной рекомбинации. Таких мышей можно получить путем замены эндогенного ракового гена на мутировавшую версию ракового гена или путем мутации эндогенного ракового гена, например, под воздействием канцерогенов. Чтобы получить трансгенных химерных животных, например мышей, конструкцию ДНК вводят в ядра эмбриональных стволовых клеток. Клетки, содержащие повреждения генома, полученные рекомбинантными методами, вводят мышиному эмбриону-хозяину, который снова имплантируют самке-реципиенту. Некоторые из данных эмбрионов развиваются у химерных мышей, у которых некоторые гаметы образуются из мутантной клеточной линии. Следовательно, путем селекции можно получить новую линию химерных мышей, содержащих искусственное повреждение гена (см., например, Capecchi et al., Science, 244: 1288 (1989)). Химерных мышей можно получить по способу, описанному в патенте США № 6365797, выданном 2 апреля 2002 г.; в патенте США № 6107540, выданном 22 августа 2000 г.; Hogan et al., Manipulating the Mouse Embryo: A laboratory Manual, Cold Spring Harbor Laboratory (1988) и Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, Robertson, ed., IRL Press, Washington, D.C. (1987). Альтернативно, можно использовать разных животных с подавленной или дефектной иммунной системой. Например, в качестве хозяина можно использовать генетически бестимусную "голую" мышь (см., например, Giovanella et al., J. Natl. Cancer Inst. 52: 921 (1974)), мышь SCID, мышь с удаленной вилочковой железой или облученную мышь (см., например, Bradley et al., Br. J. Cancer, 38: 263 (1978); Selby et al., Br. J. Cancer, 41: 52 (1980)). Опухолевые клетки (обычно около 10 6 клеток), пересаженные изогенным хозяевам, продуцируют инвазивные опухоли с высокой частотой, тогда как нормальные клетки такого же происхождения не продуцируют опухоль. Хозяевам с развившимися инвазивными опухолями подкожно или в ортотопическую область вводят клетки, экспрессирующие ассоциированные с раком последовательности. Затем мышей разделяют на группы, включающие в себя контрольные группы, и вводят им (экспериментальным группам) тестируемое средство (например, модулятор). Через определенный промежуток времени, предпочтительно через 4-8 недель, измеряют рост опухоли (например, по объему, или по двум наибольшим размерам, или по массе) и сравнивают его с контролем. Говорят, что рост опухоли ингибируется, если происходит статистически значимое уменьшение объема опухоли (определяемое, например, с использованием Т-теста Стьюдента). Идентификация и характеристика модуляторов с помощью анализов in vitro. Идентифицировать соединения, обладающие моделирующей активностью, можно с помощью анализов in vitro. Например, в начале раковый полипептид приводят в контакт с потенциальным модулятором и затем инкубируют его в течение подходящего периода времени, например от 0,5 до 48 ч. В одном воплощении, уровни ракового полипептида определяют in vitro путем измерения уровня белка или мРНК. Уровень белка измеряют с помощью иммунологических анализов, таких как вестерн-блоттинг, ELISA и т.п., с использованием антитела, которое селективно связывается с раковым полипептидом или его фрагментом. Количество мРНК предпочтительно определяют методом амплификации, например с использованием ПЦР, LCR, или гибридизации, например нозерн-гибридизации, защиты РНКазы, дот-блоттинга. Как указано в данном документе, для определения уровня белка или мРНК используют непосредственно или косвенно меченые агенты, например флуоресцентно или радиоактивно меченые нуклеиновые кислоты, радиоактивно или ферментативно меченые антитела и т.п. Альтернативно, можно использовать систему репортерного гена, содержащую промотор ракового белка, функционально связанный с репортерным геном, таким как ген люциферазы, зеленого флуоресцентного белка, CAT или P-gal. Репортерную конструкцию' обычно трансфицируют в клетку. После обработки потенциальным модулятором измеряют величину транскрипции, трансляции или активности репортерного гена с помощью стандартных методов, известных специалистам в данной области (Davis G.F., приведено выше; Gonzalez, J. & Negulescu, P. Curr. Opin. Biotechnol. 1998, 9: 624). Как указано выше, скрининги in vitro проводят по отдельным генам и генным продуктам. То есть после идентификации конкретного гена с дифференциальной экспрессией в зависимости от состояния проводят скрининг модуляторов экспрессии гена или генного продукта. В одном воплощении проводят скрининг модуляторов экспрессии конкретного гена (генов). Как правило, определяют экспрессию только одного или нескольких генов. В другом воплощении при проведении скринингов вначале выявляют соединения, которые связываются с белками, экспрессия которых меняется в зависимосит от состояния клетки. Затем определяют способность данных соединений модулировать активность, связанную с изменяющейся экспрессией. Кроме того, после идентификации исходных соединений-кандидатов можно подвергнуть скринингу их варианты, чтобы более полно осветить взаимоотношения структура-активность. Идентификация и характеристика модуляторов путем анализа связывания. В анализах связывания в соответствии с данным изобретением обычно используют очищенный или выделенный генный продукт. Например, получают антитела против белка согласно изобретению и с помощью иммунологических анализов определяют количество и/или локализацию белка. Альтернативно, в таких анализах используют клетки, содержащие раковые белки. Таким образом, данные способы включают в себя объединение ракового белка согласно изобретению с соединением-кандидатом, таким как лиганд, и определение связывания соединения с раковым белком согласно изобретению. В предпочтительных воплощениях используют человеческий раковый белок; также можно разработать и использовать животные модели человеческого злокачественного заболевания. Кроме того, специалистам в данной области известно, что можно использовать другие аналогичные белки млекопитающих. Более того, в некоторых воплощениях используют варианты или производные раковых белков. Как правило, раковый белок согласно изобретению, или лиганд, связан с нерастворимой подложкой и не способен диффундировать. Подложкой, например, может быть поверхность, принимающая выделенный образец (планшет для микротитрования, чип и др.). Нерастворимые носители могут быть изготовлены из любого вещества, с которым могут связываться композиции, которое легко отделяется от растворимой фракции и в других аспектах совместимо со способом скрининга в целом. Поверхность таких носителей может быть сплошной или пористой и может иметь любую подходящую форму. Примеры подходящих нерастворимых подложек включают в себя планшеты для микротитрования, чипы, мембраны и шарики. Обычно их изготавливают из стекла, пластика (например, полистирола), полисахарида, нейлона, нитроцеллюлозы, Teflon ™ и др. Особенно подходящими являются планшеты для микротитрования и чипы, поскольку они позволяют одновременно проводить большое число анализов с использованием маленьких количеств реагентов и образцов. Конкретный характер связывания композиции с носителем не имеет большого значения при условии, что он совместим с реагентами и способами согласно изобретению в целом, позволяет сохранять активность композиции и не сопровождается диффузией. Предпочтительные способы связывания включают в себя применение антител, которые при присоединении белка к подложке не вызывают стерической блокады ни центра связывания лиганда, ни последовательности активации, непосредственное присоединение к "липким" или ионным подложкам, химическую сшивку, синтез белка или агента на поверхности и др. После присоединения белка или лиганда/связывающего агента к носителю избыток несвязанного вещества удаляют промыванием. Затем поверхности, принимающие образец, можно блокировать путем инкубации с бычьим сывороточным альбумином (БСА), казеином или другим нетоксичным белком или фрагментом. После присоединения ракового белка к подложке к аналитической смеси добавляют тестируемое соединение. Альтернативно, к подложке присоединяют вещество, связывающее соединение-кандидат и затем добавляют белок согласно изобретению. Связывающие вещества включают в себя специфические антитела, неприродные связывающие средства, идентифицированные при скрининге химических библиотек, пептидных аналогов и др. Особый интерес представляют анализы, использующиеся для идентификации средств, обладающих низкой токсичностью для клеток человека. Для данной цели можно использовать широкий ряд анализов, включающих в себя анализы пролиферации, анализы цАМФ, анализы связывания меченых in vitro белков с другими белками, анализы сдвига злектрофоретической подвижности, иммунологические анализы связывания белков, функциональные анализы (анализы фосфорилирования и др.) и т.п. Определение связывания тестируемого соединения (лиганда, связывающего агента, модулятора и др.) с раковым белком согласно изобретению можно осуществить разными способами. Тестируемое соединение можно пометить и непосредственно определить связывание, например, путем присоединения всего ракового белка согласно изобретению или его фрагмента к твердой подложке с последующим добавлением меченого соединения-кандидата (например, содержащего флуоресцентную метку), отмыванием избытка реагента и определением присутствия метки на твердой подложке. В зависимости от обстоятельств можно использовать разные стадии блокирования и промывания. В некоторых воплощениях метят только один из компонентов, например, белок согласно изобретению или лиганды. Альтернативно, можно метить несколько компонентов разными метками, например белки - 125 I, а соединение - флуорофором. Также можно использовать реагенты, обеспечивающие близкие взаимодействия, например гашение или перенос энергии. Идентификация и характеристика модуляторов путем анализа конкурентного связывания. В одном воплощении связывание "тестируемого соединения" определяют путем анализа конкурентного связывания с "соединением-конкурентом". Соединение-конкурент представляет собой связывающий фрагмент, который связывается с целевой молекулой (например, раковым белком согласно изобретению). Соединения-конкуренты включают в себя такие соединения, как антитела, пептиды, партнеры по связыванию, лиганды и др. В определенных условиях связывания соединение-конкурент вытесняет тестируемое соединение. В одном воплощении тестируемое соединение метят. Либо тестируемое соединение, либо соединение-конкурент, либо оба соединения инкубируют с белком в течение периода времени, достаточного для того, чтобы произошло связывание. Инкубации проводят при температуре, которая обеспечивает оптимальную активность, как правило, от 4 до 40 °С. Периоды инкубации обычно оптимизируют, например, чтобы обеспечить быстрый скрининг с высокой пропускной способностью; как правило, достаточным является период от 0 до 1 ч. Избыток реагента обычно удаляют или отмывают. Затем добавляют второй компонент и, чтобы определить связывание, детектируют присутствие или отсутствие меченого компонента. В одном воплощении вначале добавляют соединение-конкурент, затем тестируемое соединение. Замещение соединения-конкурента указывает на то, что тестируемое соединение способно связываться с раковым белком и, возможно, модулировать его активность. В данном воплощении любой компонент может быть меченым. Так, например, если пометить соединение-конкурент, присутствие метки в растворе после отмывания тестируемого соединения указывает на то, что тестируемое соединение замещает соединение-конкурент. Альтернативно, если пометить тестируемое соединение, присутствие метки на подложке будет указывать на замещение. В альтернативном воплощении вначале добавляют тестируемое соединение, затем после инкубации и промывания добавляют соединение-конкурент. Отсутствие связывания с конкурентом указывает на то, что тестируемое соединение связывается с раковым белком с более высоким сродством, чем соединение-конкурент. Таким образом, если меченым является тестируемое соединение, присутствие метки на подложке указывает на отсутствие связывания соединения-конкурента, а также на то, что тестируемое соединение связывается с раковым белком согласно изобретению и, следовательно, возможно, модулирует его активность. Соответственно, методы конкурентного связывания включают в себя дифференциальный скрининг для идентификации средств, способных модулировать активность раковых белков согласно изобретению. В данном воплощении способы включают в себя объединение ракового белка и соединения-конкурента в первом образце. Второй образец содержит тестируемое соединение, раковый белок и соединение-конкурент. Связывание конкурента определяют для обоих образцов, и изменение связывания в одном образце по сравнению с другим или разница в связывании в двух образцах указывает на присутствие средства, способного связываться с раковым белком и, возможно, модулировать его активность. То есть если связывание конкурента в первом образце отличается от связывания конкурента во втором образце, значит, средство способно связываться с раковым белком. Альтернативно, дифференциальный скрининг используют для идентификации лекарственных средств-кандидатов, которые связываются с нативным раковым белком, но не способны связываться с модифицированными раковыми белками. Например, модель структуры ракового белка используют в рациональном дизайне лекарственного средства для синтеза соединений, которые взаимодействуют с конкретным участком, соединений, которые обычно не связываются с белками, у которых этот участок модифицирован. Более того, скрининг лекарственных средств на способность повышать или понижать активность раковых белков позволяет идентифицировать лекарственные средства-кандидаты, которые влияют на активность нативного ракового белка. В данных анализах можно использовать положительные и отрицательные контроли. Чтобы получить статистически значимые результаты, контрольные и тестируемые образцы предпочтительно используют по меньшей мере с тройными повторами. Инкубацию всех образцов проводят в течение периода времени, достаточного для того, чтобы произошло связывание соединения с белком. После инкубации образцы отмывают от неспецифически связанных веществ и определяют количество связанного, обычно меченого, соединения. Например, при использовании радиоактивной метки количество связанного соединения можно определить путем измерения количества радиоактивности на сцинтилляционном счетчике. В скрининговых анализах можно использовать ряд других реагентов. Такие реагенты включают в себя такие соединения, как соли, нейтральные белки, например альбумин, детергенты и др., которые используют для обеспечения оптимального белок-белкового связывания и/или уменьшения неспецифических или фоновых взаимодействий. Кроме того, можно использовать реагенты, которые иным образом улучшают эффективность анализа, такие как ингибиторы протеаз, ингибиторы нуклеаз, противомикробные средства и др. Компоненты смеси добавляют в таком порядке, который обеспечивает необходимое связывание. Применение полинуклеотидов для понижающей регуляции или ингибирования белка согласно изобретению. Полинуклеотидные модуляторы рака можно вводить в клетку, содержащую целевую нуклеотидную последовательность, путем образования конъюгата с лигандсвязывающей молекулой, как описано в WO 91/04753. Подходящие лигандсвязывающие молекулы включают в себя, без ограничения, клеточные поверхностные рецепторы, факторы роста, другие цитокины или другие лиганды, которые связываются с клеточными поверхностными рецепторами. Предпочтительно конъюгирование лигандсвязывающей молекулы, по существу, не препятствует способности данной молекулы связываться с соответствующей молекулой или рецептором или блокировать вход смыслового или антисмыслового олигонуклеотида или его конъюгированной версии в клетку. Альтернативно, полинуклеотидный модулятор рака можно ввести в клетку, содержащую целевую нуклеотидную последовательность, например, путем образования комплекса полинуклеотид-липид, как описано в WO 90/10448. Понятно, что антисмысловые молекулы или модели с нокаутным и выключенным геном помимо способов лечения также можно использовать в описанных выше скрининговых анализах. Ингибиторные и антисмысловые нуклеотиды. В некоторых воплощениях активность ассоциированного с раком белка подвергается понижающей регуляции или полностью ингибируется путем применения антисмыслового полинуклеотида или ингибиторных маленьких ядерных РНК (snPHK), т.е. нуклеиновых кислот, комплементарных и предпочтительно специфически гибридизующихся с нуклеотидной последовательностью, кодирующей, например, раковый белок согласно изобретению, мРНК или ее субпоследовательность. Связывание антисмыслового полинуклеотида с мРНК уменьшает трансляцию и/или стабильность мРНК. В контексте согласно изобретению антисмысловые полинуклеотиды могут содержать природные нуклеотиды или синтетические, полученные из природных нуклеотидов или их близких гомологов. Антисмысловые полинуклеотиды также могут содержать измененные углеводные фрагменты или измененные связи между остатками сахаров. Примерами таких измененных нуклеотидов являются фосфоротиоат и другие серосодержащие виды, известные в данной области. Аналоги входят в объем согласно изобретению при условии, что они гибридизуются с нуклеотидами согласно изобретению. См., например, Isis Pharmaceuticals, Carlsbad, СА; Sequitor, Inc., Natick, MA. Такие антисмысловые полинуклеотиды можно легко получить рекомбинантными методами или путем синтеза in vitro. Оборудование для такого синтеза продается несколькими поставщиками, в том числе Applied Biosystems. Способы получения других олигонуклеотидов, таких как фосфоротиоаты и алкилированные производные, хорошо известны специалистам в данной области. В данном описании термин "антисмысловые молекулы" включает в себя антисмысловые или смысловые олигонуклеотиды. Смысловые олигонуклеотиды, например, можно использовать для блокирования транскрипции путем связывания с антисмысловой цепью. Антисмысловые и смысловые олигонуклеотиды включают в себя последовательность одноцепочечной нуклеиновой кислоты (либо РНК, либо ДНК), способной связываться с целевыми последовательностями мРНК (смысловой) или ДНК (антисмысловой), кодирующими раковые молекулы. В соответствии с настоящим изобретением антисмысловые или смысловые олигонуклеотиды включают в себя фрагмент, содержащий, как правило, по меньшей мере приблизительно 12 нуклеотидов, предпочтительно приблизительно от 12 до 30 нуклеотидов. Способы получения антисмысловых или смысловых олигонуклеотидов на основе последовательности кДНК, кодирующей целевой белок, описаны, например, в Stein & Cohen (Cancer Res. 48: 2659 (1988) и van der Krol et al. (BioTechniques, 6: 958 (1988)). Рибозимы. Помимо антисмысловых полинуклеотидов для ингибирования ассоциированных с раком нуклеотидных последовательностей, можно использовать рибозимы. Рибозим представляет собой молекулу РНК, которая каталитически расщепляет другие молекулы РНК. Описаны разные виды рибозимов, в том числе рибозимы группы I, молотоголовые (hammerhead) рибозимы, шпилечные (hairpin) рибозимы, РНКаза Р и рибозимы с урезанной головой (axhead) (общий обзор свойств разных рибозимов можно найти, например, в Castanotto et al., Adv. in Pharmacology, 25: 289-317 (1994)). Общие свойства шпилечных рибозимов описаны, например, в Hampel et al., Nucl. Acids Res. 18: 299-304 (1990); в Европейской патентной заявке № 0360257 и в патенте США № 5254678. Способы получения рибозимов хорошо известны специалистам в данной области (см., например, WO 94/26877; Ojwang et al., Proc. Natl. Acad. Sci. USA, 90: 6340-6344 (1993); Yamada et al., Human Gene Therapy, 1: 39-45 (1994); Leavitt et al., Proc. Natl. Acad. Sci. USA, 92: 699-703 (1995); Leavitt et al., Human Gene Therapy, 5: 1151-120 (1994) и Yamada et al., Virology, 205: 121-126 (1994)). Применение модуляторов в фенотипическом скрининге. В одном воплощении тестируемое соединение вводят в популяцию раковых клеток, которые имеют связанный с раком профиль экспрессии. Под "введением" или "приведением в контакт" в данном описании подразумевают добавление модулятора к клеткам таким способом, который обеспечивает действие модулятора на клетку, либо внутриклеточное действие после поглощения, либо действие на клеточную поверхность. В некоторых воплощениях нуклеиновую кислоту, кодирующую белковый агент (т.е. пептид), помещают в вирусную конструкцию, например в аденовирусную или ретровирусную, и добавляют к клетке так, чтобы обеспечить экспрессию пептидного агента, см., например, РСТ US97/01019. Также можно использовать регуляторные системы для генной терапии. После введения модулятора клетки при необходимости промывают и оставляют инкубироваться предпочтительно в физиологических условиях в течение определенного периода времени. Затем клетки собирают и определяют профиль экспрессии генов. Так, например, для скрининга средств, которые модулируют, например, индуцируют или подавляют раковый фенотип, используют раковую ткань. Изменение экспрессии по меньшей мере одного гена, предпочтительно нескольких, в общем профиле экспрессии указывает на то, что тестируемое средство влияет на раковую активность. Подобным образом, изменение биологической функции или сигнального пути указывает на наличие модулирующей активности. После определения такого влияния на раковый фенотип предпринимают скрининг новых лекарственных средств, которые изменяют фенотип. В данном подходе лекарственное средство может быть неизвестным и может не присутствовать в исходном скрининге экспрессии гена/белка, уровень транскрипта целевого белка также может не меняться. Ингибирующая функция модулятора служит суррогатным маркером. Как указано выше, скрининги проводят по генам или генным продуктам. То есть после идентификации конкретного гена с дифференциальной экспрессией, в зависимости от состояния, проводят скрининг модуляторов по экспрессии либо гена, либо генного продукта. Применение модуляторов для воздействия на пептиды согласно изобретению. Определение активности ракового полипептида или ракового фенотипа проводят с помощью ряда анализов. Например, влияние модуляторов на функцию ракового полипептида (полипептидов) измеряют путем определения описанных выше параметров. Для оценки влияния тестируемого соединения на полипептиды согласно изобретению используют физиологическое изменение, вызываемое изменением активности полипептида. Если функциональные анализы проводят на интактных клетках или животных, можно оценивать разные эффекты, например, в случае рака, связанного с солидными опухолями, это - рост опухоли, метастазирование опухоли, реваскуляризация, высвобождение гормонов, изменение транскрипции как известных, так и неохарактеризованных генных маркеров (например, определяемое методом нозерн-блоттинга), изменение клеточного метаболизма, например рост клеток или изменение рН, а также изменение уровня внутриклеточных вторичных мессенджеров, таких как cGNIP. Способы идентификации и характеристики ассоциированных с раком последовательностей. Экспрессия разных генных последовательностей коррелирует с наличием злокачественного заболевания. Соответственно, определяют нарушения, связанные с мутантными или вариантными раковыми генами. В одном воплощении данное изобретение предлагает способы идентификации клеток, содержащих варианты раковых генов, например определение присутствия всей или части последовательности по меньшей мере одного ракового гена в клетке. Такое определение можно проводить с помощью разных методов секвенирования. Данное изобретение включает в себя способы идентификации ракового генотипа у субъекта, например определение присутствия у субъекта всей или части последовательности по меньшей мере одного гена согласно изобретению. Данный анализ проводят с использованием по меньшей мере одной ткани субъекта, например ткани, приведенной в табл. I, но можно использовать несколько тканей или несколько разных образцов одной ткани. Данный способ может включать в себя сравнение последовательности секвенированного гена с последовательностью известного ракового гена, т.е. гена дикого типа, для отнесения к определенному семейству, а также для выявления гомологии, мутаций или вариантов. Затем последовательность всего или части гена можно сравнить с последовательностью известного ракового гена, чтобы определить, существуют ли какие-нибудь различия. Такое сравнение можно осуществить с помощью ряда известных программ для определения гомологии, таких как BLAST, Bestfit и др. Как указано в данном описании, различие в последовательностях ракового гена пациента и известного ракового гена коррелирует с болезненным состоянием или с предрасположенностью к болезненному состоянию. В предпочтительном воплощении раковые гены используют в качестве зондов для определения числа копий ракового гена в геноме. Раковые гены также используют в качестве зондов для определения хромосомной локализации раковых генов. Информацию о хромосомной локализации можно использовать для диагностики и прогнозирования, особенно если в локусе ракового гена обнаружены хромосомные нарушения, такие как транслокации и т.п. XIV. иРНК и терапевтическое применение коротких интерферирующих РНК (киРНК). Настоящее изобретение также направлено на олигонуклеотиды киРНК, в частности на двухцепочечные РНК, содержащие, по меньшей мере, фрагмент участка, кодирующего STEAP-1, или 5"-участки UTR, а также на олигонуклеотиды, комплементарные последовательности STEAP-1, или на антисмысловые олигонуклеотиды, специфичные для последовательности STEAP-1. В одном воплощении такие олигонуклеотиды используют для освещения функции STEAP-1 или для скрининга с целью выявления модуляторов функции или экспрессии STEAP-1. В другом воплощении экспрессию гена STEAP-1 уменьшают путем трансфекции киРНК, что приводит к значительному снижению пролиферативной способности трансформированных раковых клеток, которые эндогенно экспрессируют антиген; у клеток, обработанных специфическими киРНК STEAP-1, наблюдается пониженная выживаемость, определяемая, например, по метаболическому считыванию клеточной жизнеспособности, которая коррелирует с пониженной способностью к пролиферации. Таким образом, композиции на основе киРНК STEAP-1 содержат киРНК (двухцепочечная РНК), которая соответствует последовательности ORF нуклеиновой кислоты, кодирующей белок STEAP-1 или его субпоследовательности; как правило, указанные субпоследовательности состоят из 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 или более непрерывных нуклеотидов РНК в длину и содержат последовательности, которые являются комплементарными и некомплементарными по меньшей мере части последовательности, кодирующей мРНК. В предпочтительном воплощении субпоследовательности содержат 19-25 нуклеотидов в длину, наиболее предпочтительно 21-23 нуклеотидов в длину. Подавление с использованием РНК является новым подходом к получению молчащих генов in vitro и in vivo, следовательно, короткие двухцепочечные РНК (киРНК) являются ценными терапевтическими средствами. Способность киРНК приводить к молчанию конкретных генов в настоящее время используется у животных моделей заболевания, а также у людей. Например, доказано, что введение мыши путем гидродинамической инфузии раствора киРНК против конкретной мишени имеет терапевтический эффект. В новаторской работе, проведенной Song E. et al., показано, что один тип природных нуклеиновых кислот, а именно, короткие интерферирующие РНК (киРНК), можно использовать в качестве терапевтических средств без какой-либо дополнительной химической модификации (Song, E., et al. "RNA interference targeting Fas protects mice from fulminant hepatitis". Nat. Med. 9(3): 347-51 (2003)). В данной работе впервые доказывается in vivo, что инфузия киРНК животному может облегчать симптомы заболевания. В данном случае авторы делают мышам инъекции киРНК, направленной против белка FAS (рецептор клеточной гибели, который при воспалительном ответе приходит в гиперактивное состояние и вызывает гибель гепатоцитов и других клеток). На следующий день животным дают антитело, специфичное к Fas. Контрольные мыши умирают от острой печеночной недостаточности в течение нескольких дней, тогда как более 80% мышей, получивших киРНК, выживают без симптомов серьезного заболевания. Приблизительно от 80 до 90% клеток печени таких мышей содержат оголенные олигонуклеотиды киРНК. Кроме того, молекулы РНК, уже функционирующие в течение 10 дней, теряют активность через 3 недели. Для лечения людей киРНК доставляют с помощью эффективных систем, обеспечивающих длительную активность иРНК. Основным требованием при клиническом применении является доставка киРНК к нужным клеткам. По-видимому, гепатоциты являются наиболее восприимчивыми к экзогенной РНК. В настоящее время мишени, находящиеся в печени, привлекают внимание исследователей, поскольку печень является органом, который могут легко достигать молекулы нуклеиновых кислот и вирусные векторы. Однако мишенями также могут служить другие ткани и органы. В терапии для улучшения введения киРНК используют композиции, содержащие киРНК вместе с соединениями, которые обеспечивают прохождение через клеточные мембраны. Другое воплощение относится к химически модифицированным синтетическим киРНК, которые являются устойчивыми к нуклеазам и стабильными в сыворотке и соответственно имеют повышенную продолжительность действия. Таким образом, технология с использованием киРНК представляет собой способ лечения злокачественных заболеваний человека путем доставки молекул киРНК, направленных против STEAP-1, субъектам, страдающим от злокачественных заболеваний, таких как перечисленные в табл. I. Такое введение киРНК приводит к снижению роста раковых клеток, экспрессирующих STEAP-1, и обеспечивает противоопухолевую терапию, уменьшая заболеваемость и/или смертность, связанные со злокачественными заболеваниями. Анализы in vitro или in vivo показали высокую эффективность данного способа устранения генного продукта. Эффективность in vitro можно легко продемонстрировать путем введения киРНК в культуру клеток (как описано выше) или в биопсийные образцы раковых пациентов, если анализы in vitro используют для детектирования пониженной экспрессии белка STEAP-1. XV. Наборы/готовые изделия. Наборы для описанных в данном документе лабораторных, прогностических, диагностических и терапевтических применений входят в объем согласно изобретению. Такие наборы могут содержать носитель, упаковку или контейнер, разделенный на отсеки, которые могут вмещать один или несколько таких контейнеров, как флаконы, пробирки и т.п., причем каждый отдельный элемент контейнера предназначен для использования в описанном способе, а также этикетку или вкладыш с инструкциями по применению, такому как применение, описанное в данном документе. Например, контейнер (контейнеры) содержит зонд, который может быть помечен детектируемой меткой. Такой зонд может представлять собой антитело или полинуклеотид, специфичный для белка, или гена, или транскрипта согласно изобретению соответственно. Если набор предназначен для детектирования целевой нуклеиновой кислоты методом гибридизации, в его состав могут входить контейнеры, содержащие нуклеотид(ы) для амплификации последовательности целевой нуклеиновой кислоты. Наборы могут включать в себя контейнер, содержащий репортерное соединение, такое как биотинилированный белок, например авидин или стрептавидин, связанный с репортерной молекулой, такой как ферментная, флуоресцентная или радиоактивная метка; такое репортерное соединение можно использовать, например, вместе с нуклеиновой кислотой или антителом. Набор может содержать полноразмерные или частичные аминокислотные последовательности, приведенные на фиг. 2 или 3, или их аналоги, или молекулы нуклеиновых кислот, кодирующих такие аминокислотные последовательности. Набор согласно изобретению обычно содержит описанный выше контейнер и один или несколько связанных с ним других контейнеров, которые содержат средства, необходимые с коммерческой и пользовательской точки зрения, и включающие в себя буферы, разбавители, фильтры, иглы, шприцы; этикетки для носителя, упаковки, контейнера, флакона и/или пробирки с описанием содержимого и/или инструкциями по применению, а также вкладыши в упаковку с инструкциями по применению. На контейнере или вместе с ним может присутствовать этикетка, указывающая, что композиция предназначена для специфического терапевтического или нетерапевтического применения, такого как прогностическое, профилактическое, диагностическое или лабораторное применение, этикетка также может указывать для in vivo или in vitro применения, такого как описанное в данном документе, предназначена данная композиция. Входящие в состав набора вкладыш(и) или этикетка(и) могут содержать инструкции или другую информацию. Этикетка может находиться на контейнере или она может быть связана с ним. Этикетка находится на контейнере, если буквы, числа или другие символы, образующие этикетку, отформованы или выгравированы на самом контейнере; этикетка может быть связана с контейнером, если она присутствует в таре или носителе, который также содержит контейнер, например в виде вкладыша в упаковку. Этикетка может указывать, что композиция предназначена для диагностики, лечения, профилактики или прогнозирования такого состояния, как неоплазия ткани, приведенной в табл. I. Термины "набор" и "готовое изделие" можно использовать как синонимы. В другом воплощении согласно изобретению предлагается готовое изделие (готовые изделия), которое включает в себя композиции, содержащие аминокислотную последовательность (аминокислотные последовательности), маленькую молекулу (маленькие молекулы), нуклеотидную последовательность (нуклеотидные последовательности) и/или антитело (антитела), т.е. вещества, используемые для диагностики, прогнозирования, профилактики и/или лечения неоплазии тканей, приведенных в табл. I. Готовое изделие обычно содержит по меньшей мере один контейнер и по меньшей мере одну этикетку. Подходящие контейнеры включают в себя, например, бутылки, флаконы, шприцы и пробирки. Контейнеры могут быть изготовлены из разных материалов, таких как стекло, металл или пластик. Контейнер может содержать аминокислотную последовательность (аминокислотные последовательности), маленькую молекулу (маленькие молекулы), нуклеотидную последовательность (нуклеотидные последовательности), популяцию (популяции) клеток и/или антитело (антитела). В одном воплощении контейнер содержит полинуклеотид для применения в анализе профиля экспрессии мРНК в клетке вместе с реагентами, используемыми для данной цели. В другом воплощении контейнер содержит антитело, его связывающий фрагмент или специфический связывающий белок для применения в анализе экспрессии белка STEAP-1 в клетках и тканях или для соответственных лабораторных, прогностических, диагностических, профилактических и терапевтических применений; указания и/или инструкции по таким применениям могут присутствовать на контейнере или вместе с ним как реагенты и другие композиции или средства, используемые для данных целей. В другом воплощении контейнер содержит вещества, вызывающие клеточный или гуморальный иммунный ответ, вместе с прилагающимися указаниями и/или инструкциями. В другом воплощении контейнер содержит вещества для адоптивной иммунотерапии, такие как цитотоксичные Т-клетки (CTL) или хелперные Т-клетки (HTL), вместе с прилагающимися указаниями и/или инструкциями; контейнер также может содержать реагенты и другие композиции или средства, используемые для данных целей. Альтернативно, контейнер может содержать композицию, эффективную для лечения, диагностики, прогнозирования или профилактики состояния, а также стерильное входное отверстие (например, контейнер может представлять собой пакет, содержащий раствор для внутривенного вливания, или флакон, имеющий пробку, которую можно проколоть иглой для подкожных инъекций). Активный ингредиент композиции может представлять собой антитело, способное специфически связывать STEAP-1 и модулировать функцию STEAP-1. Готовое изделие может дополнительно включать в себя второй контейнер, содержащий фармацевтически приемлемый буфер, такой как забуференный фосфатом физиологический раствор, раствор Рингера и/или раствор декстрозы. Он может также содержать другие средства, необходимые с точки зрения производителя и потребителя, которые включают в себя другие буферы, разбавители, фильтры, мешалки, иглы, шприцы и/или вкладыши в упаковку с указаниями и/или инструкциями по применению. Различные аспекты согласно изобретению далее описываются и иллюстрируются с помощью нескольких приведенных ниже примеров, ни один из которых не предназначен для ограничения объема согласно изобретению. Пример 1. Выделение фрагмента кДНК гена STEAP-1 методом SSH. Материалы и методы. Ксенотрансплантаты LAPC. Ксенотрансплантаты LAPC получают от Dr. Charles Sawyers (UCLA) и используют по описанному способу (Klein et al., 1997, Nature Med. 3: 402-408; Craft et al., 1999, Cancer Res. 59: 5030-5036). Андрогензависимые и андрогеннезависимые ксенотрансплантаты LAPC-4 (LAPC-4 AD и AI соответственно) и ксенотрансплантаты LAPC-9 (LAPC-9 AD и AI соответственно) выращивают у интактных самцов мышей SCID или у кастрированных самцов соответственно и пересаживают в виде маленьких кусочков ткани самцам-реципиентам. Ксенотрансплантаты LAPC-4 AI получают из опухолей LAPC-4 AD, а ксенотрансплантаты LAPC-9 AI получают из опухолей LAPC-9 AD. Чтобы получить ксенотрансплантаты AI, самцов мышей, несущих опухоли LAPC AD, кастрируют и выдерживают в течение 2-3 месяцев. После того как опухоли LAPC снова вырастают, их выделяют и пересаживают кастрированным самцам или самкам мышей SCID. Ксенотрансплантаты LAPC-4 AD выращивают внутри голени следующим образом. Выращенную подкожно опухолевую ткань ксенотрансплантата LAPC-4 AD измельчают с получением кусочков размером 1-2 мм 3 в среде 1X Iscoves, затем измельченную ткань центрифугируют при 1,3 тыс. об/мин в течение 4 мин, супернатант ресуспендируют в 10 мл охлажденной на льду среде 1X Iscoves и центрифугируют при 1,3 тыс. об/мин в течение 4 мин. Затем осадок ресуспендируют в среде 1X Iscoves, содержащей 1% проназы Е, и инкубируют 20 мин при комнатной температуре при умеренном встряхивании, после чего инкубируют на льду в течение 2-4 мин. Фильтрат центрифугируют при 1,3 тыс. об/мин в течение 4 мин, супернатант удаляют отсасыванием, а осадок снова суспендируют в 10 мл среды Iscoves и центрифугируют, чтобы удалить проназу. Затем скопления клеток помещают в среду PrEGM и выращивают в течение ночи. После этого клетки собирают, фильтруют, промывают 2 × RPMI и считают. Приблизительно 50000 клеток смешивают на льду с равным объемом охлажденного на льду матригеля и хирургическим путем вводят в проксимальный метафиз большеберцовой кости мышей SCID с помощью иглы 27 калибра. Через 10-12 недель извлекают опухоли LAPC-4, растущие в костном мозге. Клеточные линии и ткани. Человеческие клеточные линии (например, HeLa) получают от АТСС и держат в среде DMEM, содержащей 5% фетальной телячьей сыворотки. Человеческие ткани для анализов РНК и белка получают из Центра хранения человеческих тканей (HTRC - Human Tissue Resource Center) UCLA (Los Angeles, CA) и от QualTek, Inc. (Santa Barbara, CA). Выделение РНК. Чтобы выделить общую РНК, опухолевую ткань и клеточные линии гомогенизируют в реагенте Trizol (Life Technologies, Gibco BRL), используемом в количестве 10 мл/г ткани или 10 мл/10 8 клеток. Из общей РНК выделяют поли-А с помощью мини- и миди-наборов для выделения РНК Qiagen's Oligotex. Общую РНК и мРНК количественно определяют с помощью спектрофотометрического анализа (O.D. 260/280 нм) и анализируют методом гель-электрофореза. Олигонуклеотиды. Используют нижеследующие нуклеотиды, очищенные методом ВЭЖХ. DPNCDN (праймер для синтеза кДНК): Адаптор 1: Адаптор 2: Праймер 1 для ПЦР: Футлярный праймер (NP) 1: Футлярный праймер (NP) 2: Супрессивная вычитательная гибридизация. Супрессивную вычитательную гибридизацию (SSH) используют для идентификации кДНК, соответствующих генам, которые подвергаются повышающей регуляции при андрогензависимом раке простаты по сравнению с доброкачественной гиперплазией простаты (ВРН). Двухцепочечные кДНК, соответствующие ксенотрансплантату LAPC-4 AD (тестер (tester)) и ткани ВРН (драйвер (driver)) синтезируют из 2 мкг смеси поли(А)+РНК, выделенной из ксенотрансплантата и ткани ВРН, как описано выше, с помощью набора для вычитательной селекции кДНК CLONTECH, с использованием 1 нг олигонуклеотида RSACDN в качестве праймера. Одно- и двухцепочечный синтез проводят по способу, описанному в инструкции по применению данного набора (CLONTECH Protocol No. PT1117-1, Catalog No. K1804-1). Полученную кДНК расщепляют Rsa I в течение 3 ч при 37 °С. Расщепленную кДНК экстрагируют смесью фенол/хлороформ (1:1) и осаждают этанолом. Драйверную кДНК (ВРН) получают путем объединения в соотношении 4:1 Rsa I-расщепленной кДНК из ВРН с расщепленной кДНК из мышиной печени, чтобы подтвердить, что мышиные гены вычитаются из тестерной кДНК (LAPC-4 AD). Тестерную кДНК (LAPC-4 AD) получают путем разбавления 1 мкл Rsa I-расщепленной кДНК LAPC-4 AD (400 нг) в 5 мкл воды. Затем разбавленную кДНК (2 мкл, 160 нг) лигируют с 2 мкл адаптора 1 и адаптора 2 (10 мкМ) в отдельных реакциях лигирования в общем объеме 10 мкл при 16 °С в течение ночи с использованием 400 ед. ДНК-лигазы Т4 (CLONTECH). Лигирование останавливают путем добавления 1 мкл 0,2 М EDTA и нагревания при 72 °С в течение 5 мин. Первую гибридизацию проводят путем добавления 1,5 мкл (600 нг) драйверной кДНК к каждой из двух пробирок, содержащих 1,5 мкл (20 нг) тестерной кДНК, лигированной с адаптором 1 и адаптором 2. На образцы, имеющие конечный объем 4 мкл, наносят слой минерального масла, после чего содержащуюся в образцах ДНК подвергают денатурации в термоблоке для проведения реакций MJ Research при 98 °С в течение 1,5 мин и затем оставляют гибридизоваться в течение 8 ч при 68 °С. Затем две гибридизационные смеси смешивают вместе с 1 мкл свежеденатурированной драйверной кДНК и оставляют гибридизоваться в течение ночи при 68 °С. После второй гибридизации смесь разбавляют в 200 мкл 20 мМ Hepes, pH 8,3, 50 мМ NaCl, 0,2 мМ EDTA, нагревают при 70 °С в течение 7 мин и хранят при -20 °С. Амплификация методом ПЦР, клонирование и секвенирование генных фрагментов, полученных с помощью SSH. Чтобы амплифицировать генные фрагменты, полученные с помощью реакций SSH, проводят две реакции ПЦР1. В первой реакции ПЦР1 мкл разбавленной конечной гибридизационной смеси добавляют к 1 мкл праймера ПЦР1 (10 мкМ), 0,5 мкл смеси dNTP (10 мкМ), 2,5 мкл 10 × реакционного буфера (CLONTECH) и 0,5 мкл 50 × предпочтительной смеси кДНК-полимераз (CLONTECH) в конечном объеме 25 мкл. ПЦР1 проводят с использованием следующих условий: 75 °С в течение 5 мин, 94 °С в течение 25 с, затем 27 циклов, включающих в себя 94 °С в течение 10 с, 66 °С в течение 30 с, 72 °С в течение 1,5 мин. В каждом эксперименте проводят пять отдельных реакций ПЦР. Продукты объединяют и разбавляют водой в соотношении 1:10. Для проведения второй реакции ПЦР1 мкл объединенной и разбавленной реакционной смеси после первой ПЦР добавляют к реакционной смеси, аналогичной используемой для ПЦР1, за исключение того, что вместо праймера ПЦР1 используют праймеры NP1 и NP2 (10 мкМ). ПЦР2 проводят, используя 10-12 циклов по 94 °С в течение 10 с, 68 °С в течение 30 с, 72 °С в течение 1,5 мин. Продукты ПЦР анализируют методом электрофореза в 2% агарозном геле. Продукты ПЦР вставляют в вектор pCR2.1, используя набор для клонирования векторов Т/А (Invitrogen). Трансформированные Е.coli подвергают селекции по синему/белому окрашиванию и по устойчивости к ампициллину. Белые колонии отбирают, помещают в 96-луночные планшеты и выращивают в жидкой культуре в течение ночи. Чтобы идентифицировать вставки, проводят ПЦР-амплификацию в 1 мл бактериальной культуры с использованием условий ПЦР1 и NP1 и NP2 в качестве праймеров. Продукты ПЦР анализируют методом электрофореза в 2% агарозном геле. Бактериальные клоны хранят в 20% глицерине в 96-луночном формате. Плазмидную ДНК выделяют, секвенируют и анализируют на гомологию с использованием баз данных GenBank, dbEST и NCICGAP. Анализ экспрессии методом ОТ-ПЦР. Одноцепочечные кДНК получают с помощью системы Superscript Preamplification, Gibco-BRL с использованием 1 мкг мРНК в качестве матрицы и олигонуклеотидов (dT) 12-18 в качестве праймеров. Согласно инструкции производителя реакцию проводят путем инкубации с обратной транскриптазой в течение 50 мин при 42 °С с последующей обработкой РНКазой Н при 37 °С в течение 20 мин. После завершения реакции и перед нормализацией объем увеличивают путем добавления 200 мкл воды. Одноцепочечные кДНК из 16 разных нормальных тканей человека получают от CLONTECH. Нормализацию одноцепочечных кДНК из нескольких тканей проводят с использованием праймеров 5'atatcgccgcgctcgtcgtcgacaa3' (SEQ ID NO: 76) и 5'agccacacgcagctcattgtagaagg3' (SEQ ID NO: 77) по амплификации β-актина. Одноцепочечную кДНК (5 мкл) амплифицируют в общем объеме 50 мкл, содержащем 0,4 мкМ праймеров, 0,2 мкМ каждого из dNTP, 1 × буфера для ПЦР (CLONTECH, 10 мМ Tris-HCl, 1,5 мМ MgCl 2 , 50 мМ KCl, рН 8,3) и 1 × ДНК-полимеразы Кленова (CLONTECH). После 18, 20 и 22 циклов отбирают 5 мкл реакционной смеси ПЦР и анализируют методом электрофореза в агарозном геле. ПЦР проводят с использованием термоблока для проведения реакций MJ Research в следующих условиях: начальную денатурацию проводят при 94 °С в течение 15 с, затем проводят 18, 20 и 22 циклов, включающих в себя 94 °С в течение 15 c, 65 °С в течение 2 мин, 72 °С в течение 5 с. Конечное удлинение при 12 °С проводят в течение 2 мин. После электрофореза в агарозном геле визульно сравнивают интенсивность окрашивания полос β-актина, соответствующих 283 п.о., из разных тканей. Рассчитывают коэффициент разбавления одноцепочечных кДНК, получая равную интенсивность полос β-актина для всех тканей после 22 циклов ПЦР. Для достижения равной интенсивности полос во всех тканях после 22 циклов ПЦР требуется три цикла нормализации. Чтобы определить уровень экспрессии гена STEAP-1, 5 мкл нормализованной одноцепочечной кДНК анализируют методом ПЦР, используя 25, 30 и 35 циклов амплификации и следующие пары праймеров: Полуколичественный анализ экспрессии проводят путем сравнения продуктов ПЦР после такого числа циклов, которое дает слабую интенсивность окрашивания полос. Результаты. Проводят несколько экспериментов SSH по способу, описанному в приведенном выше разделе "Материалы и методы", и выделяют ряд клонов-кандидатов генных фрагментов. Все клоны-кандидаты секвенируют и подвергают анализу на гомологию со всеми последовательностями из основной общедоступной базы данных генов и базы данных EST, позволяющему получить информацию по идентичности соответствующего гена и выбрать конкретный ген для анализа на дифференциальную экспрессию. Как правило, анализу на дифференциальную экспрессию методом ОТ-ПЦР и/или нозерн-блоттинга подвергают фрагменты генов, которые не имеют гомологии ни с одной известной последовательностью, присутствующей в используемых базах данных, и, следовательно, считаются новыми генами, а также фрагменты генов, обладающие гомологией с ранее секвенированными экспрессирующимися маркерными последовательностями (EST). Один из клонов кДНК, обозначаемый STEAP-1, имеет 436 п.о. в длину и обладает гомологией с последовательностью EST из базы данных опухолевых генов NCI-CGAP. Используя данную кДНК, выделяют полноразмерную кДНК, кодирующую ген STEAP-1, которую также называют STEAP-1. Нуклеотидная последовательность кДНК STEAP-1 соответствует нуклеотидным остаткам 150-585 в последовательности кДНК STEAP-1, приведенной на фиг. 1. Другой клон, обозначаемый 28Р3Е1, имеет 561 п.о. в длину и обладает гомологией с рядом последовательностей EST из базы данных опухолевых генов NCICGAP или из других баз данных. Часть последовательности 28Р3Е1 (356 п.о.) идентична EST, полученной из человеческой фетальной ткани. После получения и секвенирования полноразмерной кДНК STEAP-1 выясняется, что данный клон также соответствует STEAP-1 (более конкретно, остаткам от 622 до 3'-конца нуклеотидной последовательности STEAP-1, приведенной на фиг. 1). Пример 2. Выделение полноразмерной кДНК, кодирующей STEAP-1. Фрагмент гена STEAP-1 размером 436 п.о. (см. пример, озаглавленный "Выделение фрагмента кДНК гена STEAP-1 методом SSH") используют для выделения других кДНК, кодирующих ген 8P1D4/STEAP-1. Коротко говоря, библиотеку кДНК нормальной простаты человека (CLONTECH) подвергают скринингу с использованием меченого зонда, полученного из кДНК STEAP-1 размером 436 п.о. Один из положительных клонов, клон 10, содержит 1195 п.о. в длину и кодирует белок из 339 аминокислот, причем данная нуклеотидная и кодируемая ею аминокислотная последовательности не обладают гомологией ни с одним известным человеческим геном или белком (гомология с крысиным белком, ассоциированным с нарушением функции почек, описана в международной заявке WO 98/53071). Кодируемый белок содержит по меньшей мере 6 предсказанных трансмембранных мотивов, предполагающих экспонирование на поверхности клетки. Данные структурные признаки обусловливают аббревиатуру "STEAP", которая расшифровывается как "шесть трансмембранных эпителиальных антигенов простаты - Six Transmembrane Epithelial Antigen of the Prostate". После идентификации других белков "STEAP" продукт гена STEAP-1 обозначают как "STEAP-1". кДНК STEAP-1 и кодируемые ею аминокислотные последовательности приведены на фиг. 2A-2Q. Клон 10 кДНК STEAP-1 помещен на хранение в Американскую коллекцию типовых культур ("АТСС") (10801 University Blvd., Manassas, VA 20110-2209 USA) как плазмидный клон STEAP-1 10.1 26 августа 1998 г. и имеет номер доступа АТСС 98849. Клон кДНК STEAP-1 можно выделить из плазмиды путем двойного расщепления EcoRI/XbaI (EcoRI по 5'-концу, XbaI по 3'-концу). Пример 3. Хромосомное картирование STEAP-1. Хромосомная локализация гена может быть связана с его участием в патогенезе. Известно несколько способов хромосомного картирования, в том числе гибридизация in situ с использованием флуоресцентной метки (FISH), картирование с применением панелей человеческих/хомячковых гибридов с использованием радиоактивной метки (RH) (Walter et al., 1994; Nature Genetics. 7: 22; Research Genetics, Huntsville Al), картирование с применением панелей гибридов соматических клеток человека и грызунов, например, доступных от института Coriell (Camden, New Jersey), а также исследование генома и установление гомологии с помощью BLAST с секвенированными и картированными геномными клонами (NCBI, Bethesda, Maryland). Определение положения STEAP-1 на хромосоме 7q21 с использованием последовательности STEAP-1 и программного обеспечения NCBI BLAST (которое можно найти в глобальной сети на сайте (.ncbi.nlm.nih.gov/genome/seq/page.cgi?F=HsBlast.html & &ORG=Hs)). Пример 4. Анализ экспрессии STEAP-1. Экспрессия STEAP-1 в образцах пациентов с раком желудка приведена на фиг. 14A-14E. На фиг. 14A изображены результаты анализа РНК, экстрагированной из нормального желудка (N) и из 10 разных образцов пациентов с раком желудка (Т). При проведении нозерн-блоттинга на полосу наносят 10 мкг общей РНК, используя в качестве зонда последовательность STEAP-1. Результаты показывают, что в тканях опухоли желудка происходит интенсивная экспрессия STEAP-1 размером приблизительно 1,6 т.о. В нижнем секторе представлено окрашивание блоттинга бромидом этидия, демонстрирующее качество образцов РНК. На фиг. 14B показано, что STEAP-1 экспрессируется в тканях пациентов с раком прямой кишки. РНК экстрагируют из нормальной прямой кишки (N), из опухолей пациентов с раком прямой кишки (Т) и из метастаз рака прямой кишки (М). При проведении нозерн-блоттинга на полосу наносят 10 мкг общей РНК, используя в качестве зонда последовательность STEAP-1. Результаты показывают, что в тканях пациентов с раком прямой кишки происходит интенсивная экспрессия STEAP-1. В нижнем секторе представлено окрашивание блоттинга бромидом этидия, демонстрирующее качество образцов РНК. Анализ экспрессии STEAP-1 методом ОТ-ПЦР показывает, что STEAP-1 интенсивно экспрессируется в эндотелиальных клетках пупочной вены человека (HUVEC) (фиг. 14C). Одноцепочечную кДНК получают из клеток HUVEC, ксенотрансплантатов рака простаты LAPC-4 AD и LAPC-9 AD, а также из тканей мозга человека. Нормализацию проводят методом ПЦР с использованием праймеров для актина и GAPDH. При проведении полуколичественной ПЦР используют праймеры для STEAP-1 и 27 и 30 циклов амплификации. В качестве контроля проводят ПЦР с использованием праймеров для актина. Результаты показывают, что в клетках HUVEC происходит интенсивная экспрессия STEAP-1, подобная экспрессии, детектируемой в тканях ксенотрансплантатов рака простаты. Экспрессия STEAP-1 в клетках HUVEC показывает, что использование STEAP-1 в качестве мишени может также сделать мишенью эндотелиальные клетки новообразованных сосудов опухоли. На фиг. 14D приведены результаты электрофореза продуктов ОТ-ПЦР в агарозном геле. На фиг. 14E приведены результаты количественного анализа продуктов ПЦР с помощью программного обеспечения Alphalmager. Результаты показывают, что среди всех тестируемых нормальных тканей интенсивная экспрессия STEAP-1 наблюдается в нормальной простате. Повышающая регуляция экспрессии STEAP-1 обнаружена в пулах рака простаты, рака мочевого пузыря, рака почки, рака прямой кишки, рака легкого, рака яичника и рака молочной железы. Интенсивная экспрессия STEAP-1 обнаружена в пулах раковых метастаз, ксенотрансплантатов рака простаты и метастаз рака простаты в лимфатическом узле. Экспрессия STEAP-1 в образцах пациентов с лимфомой показана на фиг. 14F. Одноцепочечную кДНК получают из ряда образцов пациентов, страдающих от лимфомы. Нормализацию проводят методом ПЦР с использованием праймеров для актина. Полуколичественную ПЦР с праймерами для STEAP-1 проводят, используя 26 и 30 циклов амплификации. Образцы анализируют на агарозном геле и с помощью программного обеспечения Alphalmager определяют количество продуктов ПЦР. Экспрессию считают сильной или средней, если сигнал детектируется после 26 или 30 циклов амплификации соответственно, и считают, что экспрессия отсутствует, если сигнал не детектирутся даже после 30 циклов амплификации. Результаты показывают, что экспрессия STEAP-1 наблюдается в 8 из 11 анализируемых опухолевых образцов (72,7%). Пример 5. Сплайсинг-варианты STEAP-1. Транскрипционные варианты представляют собой варианты зрелой мРНК, кодируемые одним геном, которые образуются в результате альтернативной транскрипции или альтернативного сплайсинга. Альтернативными транскриптами являются транскрипты, образующиеся в результате транскрипции одного гена, но начинающейся в разных точках. Сплайсинг-варианты представляют собой варианты мРНК, образующиеся в результате альтернативного сплайсинга одного транскрипта. У эукариотов исходная РНК, образующаяся при транскрипции полиэкзонного гена из геномной ДНК, подвергается сплайсингу с получением функциональной мРНК, которая содержит только экзоны и используется для трансляции в аминокислотную последовательность. Соответственно, один ген может иметь от нуля до нескольких альтернативных транскриптов и каждый транскрипт может иметь от нуля до нескольких сплайсинг-вариантов. Каждый транскрипционный вариант имеет уникальный экзонный состав и может содержать кодирующие и/или некодирующие (5'- или 3'-концевые) фрагменты, отличные от содержащихся в исходном транскрипте. Транскрипционные варианты могут кодировать одинаковые или разные белки с одинаковыми или подобными функциями или белки с разными функциями, причем данные варианты могут экспрессироваться в одной ткани в одно и то же время, или в разных тканях в одно и то же время, или в одной ткани в разное время, или в разных тканях в разное время. Белки, кодируемые транскрипционными вариантами, могут иметь одинаковую или разную клеточную или внеклеточную локализацию, например они могут секретироваться или находиться внутри клетки. Транскрипционные варианты идентифицируют с помощью ряда известных в данной области методов. Например, альтернативные транскрипты и сплайсинг-варианты идентифицируют путем экспериментов с полноразмерным клонированием или путем применения полноразмерного транскрипта и последовательностей EST. Вначале все человеческие EST группируют в кластеры, все члены которых прямо или косвенно идентичны друг другу. Затем EST в одном кластере дополнительно подразделяют на подкластеры и собирают в консенсусную последовательность. Последовательность исходного гена сравнивают с консенсусной последовательностью (последовательностями) других полноразмерных последовательностей. Каждая консенсусная последовательность представляет собой потенциальный сплайсинг-вариант данного гена (см., например, Kan, Z., et al., Gene structure prediction and alternative splicing analysis using genomically aligned ESTs, Genome Research, 2001 May, 11(5): 889-900). Даже если идентифицирован вариант, который не является полноразмерным клоном, данный фрагмент варианта можно использовать для получения антитела и для дальнейшего клонирования полноразмерного сплайсинг-варианта с помощью известных в данной области методов. Более того, в данной области известны компьютерные программы, которые позволяют идентифицировать транскрипционные варианты на основе геномных последовательностей. Программы для идентификации транскрипционных вариантов на основе геномных последовательностей включают в себя FgenesH (A. Salamov and V. Solovyev, "Ab initio gene finding in Drosophila genomic DNA". Genome Research. 2000 April; 10(4): 516-22); Grail (URL compbio.ornl.gov/Grail-bin/EmptyGrailForm) и GenScan (URL genes.mit.edu/GENSCAN.html). Общее описание способов идентификации сплайсинг-вариантов можно найти, например, в Southan, С. A genomic perspective on human proteases, FEBS Lett. 2001, Jun. 8; 498(2-3): 214-8; de Souza, S.J., et al., Identification of human chromosome 22 transcribed sequences with ORF expressed sequence tags, Proc. Natl. Acad. Sci. USA. 2000, Nov. 7; 97(23): 12690-3. Параметры транскрипционного варианта можно подтвердить с помощью ряда известных в данной области методов, таких как полноразмерное клонирование, протеомный анализ, анализ на основе ПЦР, анализ 5' RACE и др. (см., например, протеомный анализ: Brennan, S.О., et al., Albumin banks peninsula: a new termination variant characterized by electrospray mass spectrometry, Biochem Biophys Acta. 1999, Aug. 17; 1433(1-2): 321-6; Ferranti P., et al., Differential splicing of premessenger RNA produces multiple forms of mature caprine alpha (s1)-casein, Eur. J. Biochem. 1997, Oct. 1; 249(1): 1-7. Анализ на основе ПЦР: Wellmann S., et al., Specific reverse transcription-PCR quantification of vascular endothelial growth factor (VEGF) splice variants by Light Cycler technology, Clin Chem. 2001 Apr.; 47(4): 654-60; Jia, H.P., et al., Discovery of new human beta-defensins using a genomics-based approach, Gene. 2001, Jan. 24; 263(1-2): 211-8. Анализ на основе ПЦР и 5' RACE: Brigle, K.Е., et al., Organization of the murine reduced folate carrier gene and identification of variant splice forms, Biochem Biophys Acta. 1997, Aug. 7; 1353(2): 191-8). В данной области известно, что при злокачественных заболеваниях происходят изменения геномных участков. Если при конкретном злокачественном заболевании происходит изменение геномного участка, к которому относят ген, также изменяются альтернативные транскрипты или сплайсинг-варианты данного гена. В данном описании указано, что STEAP-1 имеет особый профиль экспрессии, связанный с злокачественным заболеванием. Альтернативные транскрипты и сплайсинг-варианты STEAP-1 также могут участвовать в развитии злокачественного заболевания одной или разных тканей и, следовательно, служить опухолеспецифическими маркерами/антигенами. Экзонный состав исходного транскрипта, обозначаемого STEAP-1 v. 1, приведен в табл. LIII. С использованием полноразмерного гена и последовательностей EST идентифицируют два транскрипционных варианта, которые обозначают STEAP-1 v. 2 и v. 3. По сравнению со STEAP-1 v. 1, при сплайсинге транскрипционного варианта STEAP-1 v. 2 не происходит отщепления интрона 4, присутствующего в STEAP-1 v. 1, а при сплайсинге варианта STEAP-1 v. 3 происходит отщепление одного дополнительного экзона из интрона 4 STEAP-1 v. 1, как изображено на фиг. 11. Теоретически, каждое другое сочетание экзонов в пространстве, например экзонов 2 и 3, представляет собой потенциальный сплайсинг-вариант. На фиг. 11 показано схематическое сравнение первичных последовательностей экзонов транскрипционных вариантов. Пример 6. Однонуклеотидный полиморфизм STEAP-1. Однонуклеотидный полиморфизм (SNP) представляет собой изменение одной пары оснований в конкретном положении нуклеотидной последовательности. Для любой определенной точки генома существует четыре возможных варианта пар нуклеотидных оснований: А/Т, C/G, G/C и Т/А. Генотип относится к специфической последовательности пар оснований в одном или нескольких положениях генома субъекта. Гаплотип относится к последовательности пар оснований более чем одного положения на одной молекуле ДНК (или на одной хромосоме у высших организмов), зачастую в рамках одного гена или в рамках нескольких тесно связанных генов. SNP, который относится к кДНК, называют cSNP. Указанные cSNP могут изменять аминокислоты белка, кодируемого геном, и посредством этого влиять на функцию белка. Некоторые SNP вызывают наследственные заболевания; другие вносят вклад в количественные вариации фенотипа субъектов и реакции субъектов на факторы окружающей среды, такие как диета и лекарственные средства. Следовательно, SNP и/или сочетания аллелей (называемые гаплотипами) могут иметь разные применения, включающие в себя диагностику наследственных заболеваний, определение реакции на лекарственные средства и их дозировки, идентификацию генов, ответственных за болезни, и анализ генетических взаимоотношений субъектов (P. Nowotny, J.М. Kwon and A.M. Goate, "SNP analysis to dissect human traits", Curr. Opin. Neurobiol. 2001 Oct.; 11(5): 637-641; M. Pirmohamed and B.K. Park, "Genetic susceptibility to adverse drug reactions", Trends Pharmacol. Sci. 2001 Jun.; 22(6): 298-305; J.H. Riley, C.J. Allan, E. Lai and A. Roses, "The use of single nucleotide polymorphisms in the isolation of common disease genes", Pharmacogenomics, 2000 Feb.; 1(1): 39-47; R. Judson, J.C. Stephens and A. Windemuth, "The predictive power of haplotypes in clinical response", Pharmacogenomics, 2000 Feb.; 1(1): 15-26). SNP идентифицируют с помощью ряда известных в данной области методов (P. Bean, "The promising voyage of SNP target discovery", Am. Clin. Lab. 2001 Oct.-Nov.; 20(9): 18-20; K.M. Weiss, "In search of human variation", Genome Res. 1998 Jul.; 8(7): 691-697; M.M. She, "Enabling large-scale pharmacogenetic studies by high-throughput mutation detection and genotyping technologies", Clin. Chem. 2001 Feb.; 47(2): 164-172). Например, SNP идентифицируют путем секвенирования фрагментов ДНК, которые обладают полиморфизмом по данным анализов, проводимых с применением геля, таких как анализ полиморфизма длины фрагментов рестрикции (RFLP) и денатурирующий электрофорез в градиентном геле (DGGE). Они могут быть обнаружены путем непосредственного секвенирования образцов ДНК, взятых от разных субъектов, или путем сравнения последовательностей разных образцов ДНК. В связи с быстрым накоплением сведений о последовательностях в общедоступных и закрытых базах данных SNP можно обнаружить путем сравнения последовательностей с помощью компьютерных программ (Z. Gu, L. Hillier and P.Y. Kwok, "Single nucleotide polymorphism hunting in cyberspace," Hum. Mutat. 1998; 12(4): 221-225). Для подтверждения SNP и определения генотипа или гаплотипа субъекта можно использовать ряд методов, включающих в себя непосредственное секвенирование и анализ с применением микрочипов высокой пропускной способности (P.Y. Kwok, "Methods for genotyping single nucleotide polymorphisms, "Annu. Rev. Genomics Hum. Genet. 2001; 2: 235-258; M. Kokoris, K. Dix, K. Moynihan, J. Mathis, B. Erwin, P. Grass, B. Hines and A. Duesterhoeft, "High-throughput SNP genotyping with the Masscode system", Mol. Diagn. 2000 Dec.; 5(4): 329-340). С помощью указанных выше способов было идентифицировано 14 SNP в транскрипте из клона GTH9, обозначаемого STEAP-1 v. 2, в положениях 602 (C/G), 386 (С/Т), 1087 (T/G), 1447 (Т/С), 1621 (А/Т), 1625 (G/T), 1716 (С/А), 2358 (С/Т), 2646 (T/G), 2859 (T/G), 2908 (А/Т), 3006 (G/C), 3107 (С/Т) и 3180 (А/Т). Транскрипты или белки с альтернативными аллелями обозначают как варианты STEAP-1 v. 4, v. 5, v. 6, v. 7, v. 8, v. 9, v. 10, v. 11, v. 12, v. 13, v. 14, v. 15, v. 16 и v. 17 соответственно. На фиг. 10 показано схематичное сравнение первичных последовательностей вариантов SNP. На фиг. 12 показано схематичное сравнение первичных последовательностей белковых вариантов, соответствующих нуклеотидным вариантам. Указанные аллели SNP, хотя показаны здесь отдельно, могут встречаться в разных сочетаниях (гаплотипах) и в любом из транскрипционных вариантов (таких как STEAP-1 v. 1 и v. 3), последовательность которых содержит SNP. Например, первые два SNP также обнаружены в STEAP-1 v. 3 в таких же положениях, а в STEAP-1 v. 1 в положениях 572 и 356 соответственно. Пример 7. Получение рекомбинантного STEAP-1 в прокариотических системах. Чтобы экспрессировать рекомбинантный STEAP-1 и варианты STEAP-1 в прокариотических клетках, полноразмерные или частичные последовательности кДНК STEAP-1 и варианта STEAP-1 клонируют в любом из ряда векторов, известных в данной области. Можно использовать полноразмерную кДНК или любой ее фрагмент, кодирующий 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или более смежных аминокислот белка STEAP-1, его вариантов или аналогов. А. Конструкции для транскрипции и трансляции in vitro. pCRII. Чтобы получить смысловые и антисмысловые РНК-зонды STEAP-1 для анализов РНК in situ, создают конструкции pCRII (Invitrogen, Carlsbad CA), кодирующие полноразмерные кДНК STEAP-1 или их фрагменты. Вектор pCRII содержит промоторы Sp6 и Т7, фланкирующие вставку и управляющие транскрипцией РНК STEAP-1 для применения в качестве зондов в анализах РНК методом гибридизации in situ. Данные зонды используются для анализа экспрессии STEAP-1 на уровне РНК в клетках и тканях. Чтобы синтезировать белок STEAP-1, транскрибируемую РНК STEAP-1, представляющую участок кДНК гена STEAP-1, кодирующий аминокислотную последовательность, используют в системах трансляции in vitro, таких как TnT ™ Coupled Reticulolysate System (Promega, Corp., Madison, WI). В. Бактериальные конструкции. Конструкции pGEX. Чтобы получить в бактериальных клетках рекомбинантные белки STEAP-1, гибридизованные с белком глутатион-S-трансферазой (GST), полноразмерные кДНК или варианты STEAP-1 или их фрагменты клонируют в GST-гибридном векторе семейства pGEX (Amersham Pharmacia Biotech, Piscataway, NJ). Полученные конструкции позволяют контролировать экспрессию последовательностей рекомбинантного белка STEAP-1, гибридизованного с GST по аминоконцу и с эпитопом из шести остатков гистидина (6 ×His) по карбоксильному концу. Маркеры GST и 6 ×His позволяют проводить выделение рекомбинантного гибридного белка из индуцированных бактерий с использованием носителя, обладающего подходящей аффинностью, и обеспечивают распознавание гибридного белка антителами против GST и His. Маркер 6 ×His получают путем добавления 6 гистидиновых кодонов к праймеру клонирования по 3'-концу, например открытой рамки считывания (ORF). Можно также добавить участок протеолитического расщепления, такой как участок распознавания PreScission ™ в pGEX-6P-1, позволяющий отщепить GST от белка, родственного STEAP-1. Ген устойчивости к ампициллину и участок начала репликации pBR322 позволяют осуществлять селекцию и поддержание плазмид pGEX в Е.coli. Конструкции pMAL. Чтобы получить в бактериальных клетках рекомбинантные белки STEAP-1, гибридизованные с белком, связывающим мальтозу (MBP), полноразмерные или частичные кДНК STEAP-1, кодирующие белковые последовательности, гибридизуют с геном МВР путем клонирования в векторах pMAL-c2X и pMAL-p2X (New England Biolabs, Beverly, MA). Полученные конструкции позволяют контролировать экспрессию последовательностей рекомбинантного белка STEAP-1, гибридизованного с МВР по аминоконцу и с эпитопом 6 ×His по карбоксильному концу. Маркеры МВР и 6 ×His позволяют проводить выделение рекомбинантного гибридного белка из индуцированных бактерий с использованием носителя, обладающего подходящей аффинностью, и обеспечивают распознавание гибридного белка антителами против МВР и His. Маркерный эпитоп 6 ×His получают путем добавления 6 гистидиновых кодонов к 3'-концу праймера клонирования. Участок распознавания фактора Ха позволяет отщеплять маркер pMAL от STEAP-1. Векторы pMAL-c2X и pMAL-p2X оптимизируют, чтобы обеспечить экспрессию рекомбинантного белка в цитоплазме или периплазме соответственно. При экспрессии в периплазме увеличивается укладка белков посредством дисульфидных связей. Конструкции рЕТ. Чтобы экспрессировать STEAP-1 в бактериальных клетках, полноразмерные или частичные кДНК STEAP-1, кодирующие белковые последовательности, клонируют в векторах семейства рЕТ (Novagen, Madison, WI). Данные векторы позволяют строго контролировать экспрессию рекомбинантного белка STEAP-1 в бактериях, который необязательно может быть гибридизован с белками, повышающими растворимость, такими как NusA и тиоредоксин (Trx), и маркерными эпитопами, такими как 6 ×His и S-Tag ™, которые позволяют проводить выделение и детектирование рекомбинантного белка. Например, с использованием гибридной системы рЕТ NusA 43.1 получают конструкции, обеспечивающие экспрессию участков белка STEAP-1 в виде аминоконцевых гибридов с NusA. С. Дрожжевые конструкции. Конструкции pESC. Чтобы экспрессировать STEAP-1 в дрожжах вида Saccharomyces cerevisiae с целью получения рекомбинантного белка и проведения функциональных исследований, полноразмерные или частичные кДНК STEAP-1, кодирующие белковые последовательности, клонируют в векторах семейства pESC, каждый из которых содержит 1 из 4 селектируемых маркеров, His3, TRP1, LEU2 и URA3 (Stratagene, La Jolla, CA). Данные векторы позволяют контролировать экспрессию одного или двух разных генов из одной плазмиды или клонировать последовательности, содержащие либо маркерный эпитоп Flag ™, либо маркерный эпитоп Мус, в одной дрожжевой клетке. Данную систему можно использовать для подтверждения белок-белковых взаимодействий STEAP-1. Кроме того, экспрессия в дрожжевых клетках сопровождается посттрансляционными модификациями, такими как гликозилирование и фосфорилирование, подобными наблюдающимся при экспрессии в эукариотических клетках. Конструкции pESP. Чтобы экспрессировать STEAP-1 в дрожжах вида Saccharomyces pombe, полноразмерные или частичные кДНК STEAP-1, кодирующие белковые последовательности, клонируют в векторах семейства pESP. Данные векторы позволяют контролировать на высоком уровне экспрессию последовательности белка STEAP-1, гибридизованному либо по аминоконцу, либо по карбоксиконцу с GST, который помогает проводить выделение рекомбинантного белка. Маркерный эпитоп Flag ™ позволяет детектировать рекомбинантный белок с помощью антитела против Flag ™. Пример 8. Получение рекомбинантного STEAP-1 в системах высших эукариотов. А. Конструкции млекопитающих. Чтобы экспрессировать рекомбинантный STEAP-1 в эукариотических клетках, полноразмерные или частичные последовательности кДНК STEAP-1 клонируют в любом из ряда векторов экспрессии, известных в данной области. С использованием данных конструкций можно экспрессировать один или несколько из участков STEAP-1, включающих в себя участок STEAP-1 v. 1, v. 4, содержащий аминокислоты 1-339, участок v. 2, содержащий аминокислоты 1-258, участок v.3, содержащий аминокислоты 1-282, или любые 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50 или больше смежных аминокислот из STEAP-1, его вариантов или аналогов. В некоторых воплощениях экспрессируется участок конкретного варианта STEAP-1, кодирующий в определенном положении аминокислоту, отличающуюся от присутствующих в данном положении аминокислот любых других вариантов. В других воплощениях экспрессируется участок варианта STEAP-1, который полностью или частично относится к последовательности, уникальной для данного варианта. Указанные конструкции можно трансфицировать в любые клетки млекопитающих, например в клетки 293Т. Лизаты трансфицированных клеток 293Т можно анализировать с помощью описанной выше поликлональной сыворотки против STEAP-1. Конструкции pcDNA4/HisMax. Чтобы экспрессировать STEAP-1 в клетках млекопитающих, ORF STEAP-1 или ее фрагменты клонируют в векторе pcDNA4/HisMax, версия A (Invitrogen, Carlsbad, CA). Экспрессия белка находится под контролем промотора цитомегаловируса (CMV) и энхансера трансляции SP16. Рекомбинантный белок содержит эпитоп Xpress ™ и эпитоп из шести гистидиновых остатков (6 ×His), присоединенные к аминоконцу. Вектор pcDNA4/HisMax также содержит сигнал полиаденилирования бычьего гормона роста (BGH) и последовательность терминации транскрипции для повышения стабильности мРНК наряду с точкой начала эписомной репликации SV40 и участком, обеспечивающим "спасение" вектора в клеточных линиях, экспрессирующих супер-Т-антиген. Ген устойчивости к зеоцину позволяет осуществлять селекцию клеток млекопитающих, экспрессирующих белок, а ген устойчивости к ампициллину и точка начала репликации ColE1 позволяют осуществлять селекцию и поддержание плазмиды в Е.coli. Конструкции pcDNA3.1/MycHis. Чтобы экспрессировать STEAP-1 в клетках млекопитающих, ORF STEAP-1 или ее фрагменты вместе с консенсусным участком инициации трансляции Козака клонируют в векторе pcDNA3.1/MycHis, версия A (Invitrogen, Carlsbad, CA). Экспрессия белка находится под контролем промотора цитомегаловируса (CMV). Рекомбинантные белки содержат эпитоп myc и эпитоп 6 ×His, присоединенные к карбоксиконцу. Вектор pcDNA3.1/MycHis также содержит сигнал полиаденилирования бычьего гормона роста (BGH) и последовательность терминации транскрипции для повышения стабильности мРНК, наряду с точкой начала эписомной репликации SV40 и участком, обеспечивающим "спасение" вектора в клеточных линиях, экспрессирующих супер-Т-антиген. Используют ген устойчивости к неомицину, который позволяет осуществлять селекцию клеток млекопитающих, экспрессирующих белок, а также ген устойчивости к ампициллину и точку начала репликации ColE1, позволяющие осуществлять селекцию и поддержание плазмиды в Е.coli. Конструкция pcDNA3.1/CT-GFP-TOPO. Чтобы экспрессировать STEAP-1 в клетках млекопитающих и обеспечить детектирование рекомбинантных белков с помощью флуоресцентного анализа, ORF STEAP-1 или ее фрагменты вместе с консенсусным участком инициации трансляции Козака клонируют в векторе pcDNA3.1/CT-GFP-TOPO (Invitrogen, CA). Экспрессия белка находится под контролем промотора цитомегаловируса (CMV). Рекомбинантные белки содержат зеленый флуоресцентный белок (GFP), присоединенный к карбоксильному концу, который позволяет проводить неинвазивное детектирование in vivo и исследования в области клеточной биологии. Вектор pcDNA3.1CT-GFP-TOPO также содержит сигнал полиаденилирования бычьего гормона роста (BGH) и последовательность терминации транскрипции для повышения стабильности мРНК, наряду с точкой начала эписомной репликации SV40 и участком, обеспечивающим "спасение" вектора в клеточных линиях, экспрессирующих супер-Т-антиген. Ген устойчивости к неомицину позволяет осуществлять селекцию клеток млекопитающих, экспрессирующих белок, а ген устойчивости к ампициллину и точка начала репликации ColE1 позволяют осуществлять селекцию и поддержание плазмиды в Е.coli. Другие конструкции, содержащие гибрид с GFP по аминоконцу, получают на основе вектора pcDNA3.1/NT-GFP-TOPO, кодирующего полноразмерный белок STEAP-1. pAPtag. ORF STEAP-1 или ее фрагменты, клонируют в векторе pAPtag-5 (GenHunter Corp. Nashville, TN). Данная конструкция кодирует гибрид, в котором к карбоксиконцу белка STEAP-1 присоединена щелочная фосфатаза, а к аминоконцу - сигнальная последовательность IgGK. Также получают конструкции, кодирующие гибриды, в которых щелочная фосфатаза вместе с аминоконцевой сигнальной последовательностью IgGK присоединена к аминоконцу белка STEAP-1. Полученные рекомбинантные белки STEAP-1 можно оптимизировать для секреции в среду трансфицированных клеток млекопитающих и использовать для идентификации белков, таких как лиганды или рецепторы, которые взаимодействуют с белками STEAP-1. Экспрессия белка находится под контролем промотора CMV, и, кроме того, рекомбинантные белки содержат эпитопы myc и 6 ×His, присоединенные к карбоксильному концу, которые облегчают детектирование и очистку. Присутствующий в векторе ген устойчивости к зеоцину позволяет осуществлять селекцию клеток млекопитающих, экспрессирующих рекомбинантный белок, а ген устойчивости к ампициллину позволяет осуществлять селекцию плазмиды в Е.coli. ptag-5. ORF STEAP-1 или ее фрагменты также клонируют в векторе ptag-5. Данный вектор подобен вектору pAPtag, но не кодирует гибрид щелочной фосфатазы. Данная конструкция кодирует белок STEAP-1, содержащий на аминоконце сигнальную последовательность IgGK, а на карбоксильном конце - маркерные эпитопы myc и 6 ×His, облегчающие детектирование и аффинную очистку. Полученный рекомбинантный белок STEAP-1 можно оптимизировать для секреции в среду трансфицированных клеток млекопитающих и использовать в качестве иммуногена или лиганда для идентификации белков, таких как лиганды или рецепторы, которые взаимодействуют с белками STEAP-1. Экспрессия белка находится под контролем промотора CMV. Присутствующий в векторе ген устойчивости к зеоцину позволяет осуществлять селекцию клеток млекопитающих, экспрессирующих белок, а ген устойчивости к ампициллину позволяет осуществлять селекцию плазмиды в Е.coli. psecFc: ORF STEAP-1 или ее фрагменты также клонируют в векторе psecFc. Вектор psecFc получают путем клонирования участка Fc (шарнирный участок, участки С Н 2, С Н 3) человеческого иммуноглобулина G1 (IgG) в векторе psectag-2 (Invitrogen, California). Данная конструкция кодирует гибрид, содержащий белок STEAP-1, присоединенный к нему по карбоксильному концу Fc-участок IgG 1 и сигнальную последовательность IgGK, присоединенную по N-концу. Также используют гибриды STEAP-1 с участком Fc мышиного IgG 1 . Полученные рекомбинантные белки STEAP-1 можно оптимизировать для секреции в среду трансфицированных клеток млекопитающих и использовать в качестве иммуногенов или для идентификации белков, таких как лиганды или рецепторы, которые взаимодействуют с белком STEAP-1. Экспрессия белка находится под контролем промотора CMV. Присутствующий в векторе ген устойчивости к гидромицину позволяет осуществлять селекцию клеток млекопитающих, экспрессирующих рекомбинантный белок, а ген устойчивости к ампициллину позволяет осуществлять селекцию плазмиды в Е.coli. Конструкции pSR α. Чтобы получить линии клеток млекопитающих, которые конститутивно экспрессируют STEAP-1, ORF STEAP-1 или ее фрагменты клонируют в конструкциях pSR α. Амфотропные и экотропные ретровирусы получают путем трансфекции конструкций pSR α в линию клеток с дефектом упаковки 293Т-10А1 или путем совместной трансфекции pSR α и хелперной плазмиды (в которой удалена последовательность с дефектом упаковки) в клетки 293 соответственно. Ретровирус используют для инфицирования ряда клеточных линий млекопитающих с интеграцией клонируемого гена, STEAP-1, в линии клеток-хозяев. Экспрессия белка находится под контролем длинного концевого повтора (LTR). Присутствующий в векторе ген устойчивости к неомицину позволяет осуществлять селекцию клеток млекопитающих, экспрессирующих белок, а ген устойчивости к ампициллину и точка начала репликации ColE1 позволяют осуществлять селекцию и поддержание плазмиды в Е.coli. Затем ретровирусные векторы используют для инфицирования и получения разных клеточных линий с применением, например, клеток РС3, NIH 3Т3, TsuPr1, 293 или RAT1. Можно получить другие конструкции pSR α, кодирующие белки, в которых к карбоксильному концу последовательностей STEAP-1 присоединяют маркерный эпитоп, такой как маркер Flag ™, чтобы обеспечить детектирование с помощью антител против Flag. Например, последовательность Flag ™ 5'-gat tac aag gat gac gac gat aag-3' (SEQ ID NO: 80) добавляют к праймеру клонирования по 3'-концу ORF. Другие конструкции pSR α кодируют гибридные белки, содержащие полноразмерные белки STEAP-1 и присоединенные к ним по амино- и карбоксильным концам GFP и myc/6 ×His. Другие вирусные векторы. Для вирус-опосредованной доставки и экспрессии STEAP-1 получают другие конструкции. Высокий титр вируса, приводящий к высокому уровню экспрессии STEAP-1, достигается при использовании таких вирусных систем доставки, как аденовирусные векторы и векторы на основе ампликона герпеса. Кодирующие последовательности STEAP-1 или их фрагменты амплифицируют методом ПЦР и субклонируют в челночном векторе AdEasy (Stratagene). Рекомбинацию и упаковку вируса проводят в соответствии с инструкциями производителя по получению аденовирусных векторов. Альтернативно, чтобы получить векторы на основе вируса герпеса, кодирующие последовательности STEAP-1 или их фрагменты клонируют в векторе HSV-1 (Imgenex). Затем вирусные векторы используют для инфицирования разных клеточных линий, таких как клетки РС3, NIH 3Т3, 293 или RAT1. Регулируемые системы экспрессии. Чтобы осуществить контролируемую экспрессию STEAP-1 в клетках млекопитающих, последовательности, кодирующие STEAP-1 или их фрагменты клонируют в регулируемых системах экспрессии млекопитающих, таких как система T-Rex (Invitrogen), система GeneSwitch (Invitrogen) и строго регулируемая система Ecdysone (Sratagene). Данные системы позволяют исследовать зависимость эффектов рекомбинантного STEAP-1 от времени и концентрации. Затем указанные векторы используют для регуляции экспрессии STEAP-1 в разных клеточных линиях, таких как клетки РС3, NIH 3Т3, 293 или RAT1. В. Бакуловирусные системы экспрессии. Чтобы получить рекомбинантные белки STEAP-1 с использованием бакуловирусной системы экспрессии, ORF STEAP-1 или ее фрагменты клонируют в бакуловирусном векторе переноса pBlueBac 4.5 (Invitrogen), который кодирует белок, содержащий His-маркер на N-конце. А именно, чтобы получить рекомбинантный бакуловирус, pBlueBac-STEAP-1 трансфицируют вместе с хелперной плазмидой pBac-N-Blue (Invitrogen) в клетки насекомых SF9 (Spodoptera frugiperda) (подробное описание данного способа можно найти в инструкции по эксплуатации Invitrogen). Затем бакуловирус собирают из клеточного супернатанта и очищают методом бляшкообразования. Затем рекомбинантный белок STEAP-1 получают путем инфицирования клеток насекомых HighFive очищенным бакуловирусом (Invitrogen). Рекомбинантный белок STEAP-1 можно детектировать с помощью антител против STEAP-1, антител против His-маркера. Белок STEAP-1 можно выделить и использовать в различных клеточных анализах или в качестве иммуногена для получения поликлональных и моноклональных антител, специфичных для STEAP-1. Пример 9. Профили антигенности и вторичная структура. На фиг. 5A-9A и 5B-9B графически изображены пять аминокислотных профилей вариантов STEAP-1 1 и 3 соответственно, анализ которых осуществлен с помощью web-сайта ProtScale, расположенного в глобальной сети на сервере молекулярной биологии ExPasy по адресу (URL.expasy.ch/cgi-bin/protscale.pi). Данные профили включают в себя следующие характеристики: фиг. 5A и 5B, гидрофильность (Норр Т.P., Woods K.R., 1981. Proc. Natl. Acad. Sci. U.S.A. 78: 3824-3828); фиг. 6A и 6B, гидропатичность, (Kyte J., Doolittle R.F., 1982. J. Mol. Biol. 157: 105-132); фиг. 7A и 7B, процент доступных остатков (Janin J., 1979, Nature, 277: 491-492); фиг. 8A и 8B, средняя жесткость (Bhaskaran R., and Ponnuswamy P.K., 1988. Int. J. Pept. Protein Res. 32: 242-255); фиг. 9A и 9B, бета-складка (Deleage, G., Roux B. 1987, Protein Engineering, 1: 289-294); а для идентификации антигенных участков белка STEAP-1 также можно использовать другие характеристики, известные в данной области, например, доступные на web-сайте ProtScale. Каждый из вышеуказанных аминокислотных профилей STEAP-1 получают с использованием нижеследующих аналитических параметров ProtScale: 1) размер составляет окна 9; 2) масса границ окна составляет 100% по сравнению с центром окна и 3) значения аминокислотного профиля нормализуют так, чтобы они находились в интервале от 0 до 1. Профили гидрофильности (фиг. 5A и 5B), гидропатичности (фиг. 6A и 6B) и процента доступных остатков (фиг. 7A и 7B) используют для определения участков гидрофильных аминокислот (т.е. имеющих значение выше 0,5 по профилям гидрофильности и процента доступных остатков и меньше 0,5 по профилю гидропатичности). Такие участки, скорее всего, экспонируются в водную среду, присутствуют на поверхности белка и, следовательно, доступны для иммунного распознавания, например, антителами. С помощью профилей средней жесткости (фиг. 8A и 8B) и бета-складки (фиг. 9A и 9B) определяют участки аминокислот (т.е. имеющие значение выше 0,5 по профилю бета-складки и по профилю средней жесткости), подвижность которых не ограничена вторичными структурами, такими как бета-слои и альфа-спирали. Такие участки, скорее всего, экспонируются на поверхности белка и, следовательно, доступны для иммунного распознавания, например, антителами. Антигенные последовательности белка STEAP-1 и вариантных белков, определенные, например, по профилям, приведенным на фиг. 5A и 5B, 6A и 6B, 7A и 7B, 8A и 8B и/или 9A и 9B, используют для получения иммуногенов, либо пептидов, либо кодирующих их нуклеиновых кислот, которые, в свою очередь, используют для получения терапевтических и диагностических антител против STEAP-1. Иммуноген может представлять собой последовательность, состоящую из любых 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50 или более смежных аминокислот белковых вариантов STEAP-1, приведенных на фиг. 2 и 3, или он может представлять собой нуклеиновую кислоту, кодирующую такую аминокислотную последовательность. В частности, пептидные иммуногены согласно изобретению могут включать в себя пептидный участок, содержащий по меньшей мере 5 аминокислот последовательности, приведенной на фиг. 2 или 3, с любым целочисленным приращением, который содержит аминокислотное положение, имеющее значение больше 0,5 по профилю гидрофильности, приведенному на фиг. 5A и 5B; пептидный участок, содержащий по меньшей мере 5 аминокислот последовательности, приведенной на фиг. 2 или 3, с любым целочисленным приращением, который содержит аминокислотное положение, имеющее значение меньше 0,5 по профилю гидрофобности, приведенному на фиг. 6A и 6B; пептидный участок, содержащий по меньшей мере 5 аминокислот последовательности, приведенной на фиг. 2 или 3, с любым целочисленным приращением, который содержит аминокислотное положение, имеющее значение больше 0,5 по профилю процента доступных остатков, приведенному на фиг. 7A и 7B; пептидный участок, содержащий по меньшей мере 5 аминокислот последовательности, приведенной на фиг. 2 или 3, с любым целочисленным приращением, который содержит аминокислотное положение, имеющее значение больше 0,5 по профилю средней жесткости, приведенному на фиг. 8A и 8B; и пептидный участок, содержащий по меньшей мере 5 аминокислот последовательности, приведенной на фиг. 2 или 3, с любым целочисленным приращением, который содержит аминокислотное положение, имеющее значение больше 0,5 по профилю бета-складки, приведенному на фиг. 9A и 9B. Иммуногены согласно изобретению также могут включать в себя нуклеиновые кислоты, кодирующие указанные выше пептиды. Все иммуногены согласно изобретению, как пептиды, так и нуклеиновые кислоты, могут быть представлены в виде человеческой стандартной лекарственной формы или они могут входить в состав композиции, которая содержит фармацевтический наполнитель, совместимый с физиологией человека. Вторичные структуры варианта 1 и варианта 3 STEAP-1, а именно наличие и местоположение альфа-спиралей, развернутых цепей и статистических клубков, предсказывают на основе первичных аминокислотных последовательностей с использованием метода HNN - иерархических нейронных сетей (Guermeur, 1997, http://pbil.ibcp.fr/cgibin/npsa_automat.pl?page=npsa_nn.html), который можно найти на сервере молекулярной биологии ExPasy, расположенном в глобальной сети по адресу (.expasy.ch/tools/). Результаты данного анализа показывают, что вариант 1 STEAP-1 на 64,60% состоит из альфа-спиралей, на 4,72% - из развернутых цепей и на 30,68% - из статистических клубков (фиг. 13A). Вариант 2 STEAP-1 на 62,79% состоит из альфа-спиралей, на 3,10% - из развернутых цепей и на 34,11% - из статистических клубков (фиг. 13B). Вариант 3 STEAP-1 на 58,87% состоит из альфа-спиралей, на 5,32% - из развернутых цепей и на 35,82% - из статистических клубков (фиг. 13C). Анализ возможного присутствия трансмембранных доменов в вариантах STEAP-1 проводят с использованием ряда алгоритмов прогнозирования трансмембранных доменов, которые можно найти на сервере молекулярной биологии ExPasy, расположенном в глобальной сети по адресу (.expasy.ch/tools/). На графическом изображении результатов анализа варианта 1 отражено присутствие и расположение 6 трансмембранных доменов, предсказанных с помощью программы TMpred (фиг. 13D) и программы ТМНММ (фиг. 13E). Также показаны результаты анализа варианта 2, отражающие присутствие и расположение 4 трансмембранных доменов, предсказанных с помощью программы TMpred (фиг. 13F), и 3 трансмембранных доменов, предсказанных с помощью программы ТМНММ (фиг. 13G). Анализ варианта 3 позволяет предсказать присутствие 4 трансмембранных доменов с помощью программы TMpred (фиг. 13H) и 3 трансмембранных доменов с помощью программы ТМНММ (фиг. 13I). Результаты применения каждой программы, а именно аминокислоты, составляющие трансмембранные домены, приведены в табл. XX. Пример 10. Получение поликлональных антител STEAP-1. Образование поликлональных антител можно индуцировать у млекопитающего, например, путем одной или нескольких инъекций иммунизирующего средства, при необходимости, вместе с адъювантом. Как правило, иммунизирующее средство и/или адъювант вводят млекопитающему путем нескольких подкожных или внутрибрюшинных инъекций. Помимо вариантов полноразмерного белка STEAP-1, для иммунизации используют иммуногены, созданные с помощью компьютерных алгоритмов, которые по результатам анализа аминокислотной последовательности обладают антигенными свойствами и доступны для распознавания иммунной системой иммунизированного хозяина (см. пример, озаглавленный "Профили антигенности и вторичная структура"). Предположительно такие участки должны быть гидрофильными, гибкими, иметь бета-складчатую конформацию и экспонироваться на поверхности белка (аминокислотные профили, указывающие на такие участки вариантов 1 и 3 белка STEAP-1, приведены, например, на фиг. 5A и 5B, 6A и 6B, 7A и 7B, 8A и 8B и/или 9A и 9B). Например, для получения поликлональных антител от новозеландских белых кроликов или моноклональных антител по способу, описанному в примере, озаглавленном "Получение моноклональных антител (Mabs) STEAP-1", в качестве антигенов используют рекомбинантные бактериальные гибридные белки или пептиды, содержащие гидрофильные, гибкие, бета-складчатые участки белковых вариантов STEAP-1. Например, такие участки включают в себя, без ограничения, аминокислоты 1-40, аминокислоты 143-165, аминокислоты 180-220 вариантов 1, 2 и 3 STEAP-1, аминокислоты 312-339 варианта 1 STEAP-1 и аминокислоты 250-282 варианта 3 STEAP-1. Предпочтительно использовать конъюгат иммунизирующего средства с белком, являющимся иммуногенным для иммунизируемого млекопитающего. Примеры таких иммуногенных белков включают в себя, без ограничения, гемоцианин лимфы улитки (KLH), сывороточный альбумин, бычий тироглобулин и соевый ингибитор трипсина. В одном воплощении пептид, содержащий аминокислоты 250-282 варианта 3 STEAP-1, конъюгируют с KLH. Затем полученный конъюгат используют в качестве иммуногена. Альтернативно, иммунизирующее средство может содержать полноразмерные или частичные вариантные белки STEAP-1, их аналоги или гибриды. Например, аминокислотную последовательность варианта 1 STEAP-1 можно гибридизовать, используя методы рекомбинантных ДНК, с одним из ряда белков-партнеров по гибридизации, хорошо известных в данной области, таких как глутатион-S-трансфераза (GST) и His-меченые гибридные белки. В другом воплощении аминокислоты 250-282 варианта 1 STEAP-1 гибридизуют с GST, используя рекомбинантные методы и вектор экспрессии pGEX, экспрессируют, очищают и используют для иммунизации кролика. Такие гибридные белки выделяют из индуцированных бактерий с помощью носителя, обладающего подходящей аффинностью. Также можно использовать другие рекомбинантные бактериальные гибридные белки, которые включают в себя белок, связывающий мальтозу, LacZ, тиоредоксин, NusA или константный участок иммуноглобулина (см. раздел, озаглавленный "Получение STEAP-1 в прокариотических системах" и Current Protocols In Molecular Biology, Volume 2, Unit 16, Frederick M. Ausubul et al. eds., 1995; Linsley, P.S., Brady, W., Urnes, M., Grosmaire, L., Damle, N., and Ledbetter, L. (1991), J. Exp. Med. 174, 561-566). Помимо бактериальных гибридных белков также используют белковые антигены, экспрессирующиеся в клетках млекопитающих. Данные антигены экспрессируют из векторов экспрессии млекопитающих, таких как tag-5 и Fc-гибридные векторы (см. раздел, озаглавленный "Продукция рекомбинантного STEAP-1 в зукариотических системах"), и сохраняют посттрансляционные модификации, такие как гликозилирование, присутствующие в нативном белке. В одном воплощении аминокислоты 185-218 варианта 1 STEAP-1 клонируют в векторе секреции млекопитающих tag-5 и экспрессируют в клетках 293Т. Рекомбинантный белок выделяют из супернатантов тканевой культуры клеток 293Т, экспрессирующих рекомбинантный вектор, методом металлохелатной хроматографии. Затем очищенный белок tag-5-вариант 1 STEAP-1 используют в качестве иммуногена. При осуществлении схемы иммунизации полезно смешивать антиген с адъювантами, которые усиливают иммунный ответ животного-хозяина, или эмульгировать антиген в таких адъювантах. Примеры адъювантов включают в себя, без ограничения, полный адъювант Фрейнда (CFA) и адъювант MPL-TDM (монофосфорил липида А, синтетический дикориномиколат трегалозы). В типичном способе иммунизации кроликам вначале вводят подкожно не более 200 мкг, обычно 100-200 мкг гибридного белка или пептида, конъюгированного с KLH и смешанного с полным адъювантом Фрейнда (CFA). Затем кроликам вводят подкожно каждые две недели не более 200 мкг, обычно 100-200 мкг иммуногена в неполном адъюванте Фрейнда (IFA). Приблизительно через 7-10 дней после каждой иммунизации берут контрольные пробы крови и определяют титр антисыворотки методом ELISA. Чтобы проанализировать реакционноспособность и специфичность иммунной сыворотки, такой как кроличья сыворотка, полученная путем иммунизации гибридным белком GST-вариант 1 STEAP-1, полноразмерную кДНК варианта 1 STEAP-1 клонируют в векторе экспрессии pcDNA3.1 myc-his (Invitrogen, см. пример, озаглавленный "Продукция рекомбинантного STEAP-1 в эукариотических системах"). После трансфекции конструкций в клетки 293Т клеточные лизаты анализируют на способность специфически взаимодействовать с денатурированным белком STEAP-1 методом вестерн-блоттинга с использованием антисыворотки против STEAP-1 и антител против His (Santa Cruz Biotechnologies, Santa Cruz, CA). Кроме того, иммунную сыворотку анализируют на способность специфически распознавать нативный белок с помощью методов флуоресцентной микроскопии, проточной цитометрии и иммунопреципитации против клеток 293Т и других клеток, экспрессирующих рекомбинантный STEAP-1. Методы вестерн-блоттинга, иммунопреципитации, флуоресцентной микроскопии и проточной цитометрии с использованием клеток, эндогенно экспрессирующих STEAP-1, также проводят для определения реакционноспособности и специфичности. Антисыворотку, полученную от кроликов, иммунизированных гибридами вариантных белков STEAP-1, такими как GST- и МВР-гибриды, очищают путем удаления антител, способных взаимодействовать с последовательностью партнера по гибридизации, пропуская сыворотку через аффинную колонку, содержащую партнер по гибридизации, либо в отдельности, либо в составе другого гибридного белка. Например, антисыворотку, полученную против гибридного белка GST-вариант 1 STEAP-1 вначале очищают путем пропускания через колонку, содержащую белок GST, ковалентно связанный с носителем AffiGel (BioRad, Hercules, Calif.). Затем антисыворотку очищают, пропуская через колонку, заполненную носителем AffiGel, ковалентно связанным с гибридным белком MBP-STEAP-1. Затем, чтобы выделить фракцию IgG, сыворотку очищают аффинной хроматографией с использованием белка G. Сыворотку, полученную от кроликов, иммунизированных другими His-мечеными антигенами и пептидами, а также сыворотку, истощенную по партнеру по гибридизации, подвергают аффинной очистке, пропуская через носитель для колонки, содержащий исходный белковый иммуноген или свободный пептид. Пример 11. Получение моноклональных антител (Mab) STEAP-1. В одном воплощении терапевтические Mab против вариантов STEAP-1 включают в себя Mab, которые взаимодействуют с эпитопами, характерными для каждого белкового варианта, или для последовательностей, общих для разных вариантов, и которые связывают, способствуют интернализации, нарушают или модулируют биологическую функцию вариантов STEAP-1, например нарушают взаимодействие с лигандами и партнерами по связыванию. Иммуногены, используемые для получения таких Mab, включают в себя иммуногены, кодирующие или содержащие внеклеточный домен или всю последовательность белкового варианта STEAP-1, участки, которые, как предсказано с помощью компьютерного анализа аминокислотной последовательности, содержат функциональные мотивы, и участки белковых вариантов STEAP-1, которые, как предсказано, являются антигенными (см., например, фиг. 5A-9A, 5B-9B и пример, озаглавленный "Профили антигенности и вторичная структура"). Иммуногены включают в себя пептиды, рекомбинантные бактериальные белки, tag-5-белки, экспрессируемые клетками млекопитающих, а также человеческие и мышиные IgG Fc-гибридные белки. Кроме того, для иммунизации мышей используют белок ptag-5, векторы ДНК, кодирующие ptag-5, клетки, экспрессирующие высокие уровни соответствующего варианта STEAP-1, такого как вариант 1 293T-STEAP-1 или 3Т3, RAT или 300.19-STEAP-1 вариант 1 мышиные пре-В клетки. Чтобы получить Mab против вариантов STEAP-1, мышей вначале иммунизируют путем введения внутрибрюшинно (в.б.) или в подошву ступни, как правило, 10-50 мкг белкового иммуногена или 10 7 STEAP-1-экспрессирующих клеток, смешанных с полным адъювантом Фрейнда. В качестве адъювантов также можно использовать Titermax (Sigma) и Immuneasy (Qiagen). Затем мышей иммунизируют путем в.б. введения каждые 2-4 недели, как правило, 10-50 мкг белкового иммуногена или 10 7 клеток, смешанных с неполным адъювантом Фрейнда. Альтернативно, для иммунизации можно использовать адъювант MPL-TDM. Помимо стратегий иммунизации с использованием вышеуказанных белков и клеток, используют схему иммунизации с применением ДНК, в которой вектор экспрессии млекопитающих, кодирующий последовательность варианта, вводят иммунизируемой мыши путем непосредственной инъекции плазмидной ДНК. Например, участок, кодирующий аминокислоты 185-218 варианта 1 STEAP-1, клонируют в векторе секреции млекопитающих tag-5 и рекомбинантный вектор используют в качестве иммуногена. В другом примере участок, кодирующий такие же аминокислоты, клонируют в векторе секреции Fc-гибрида, в котором последовательность варианта 1 STEAP-1 присоединена к аминоконцу лидерной последовательности IgK и к карбоксильному концу последовательности, кодирующей Fc-участок человеческого или мышиного IgG. Данный рекомбинантный вектор используют в качестве иммуногена. Схему иммунизации плазмидой используют в сочетании с очищенными белками, экспрессируемыми тем же вектором, и с клетками, экспрессирующими соответствующий вариант STEAP-1. В другом примере получают моноклональное антитело против варианта 3 STEAP-1, используя пептид, кодирующий аминокислоты 250-282. Получают конъюгат данного пептида с KLH и используют в качестве иммуногена. Иммунореактивные клоны идентифицируют методом ELISA с использованием свободного пептида. Реакционноспособность и специфичность моноклональных антител по отношению к варианту 1 полноразмерного белка STEAP-1 определяют методами Вестерн-блоттинга, иммунопреципитации и проточной цитометрии с использованием клеток, экспрессирующих как рекомбинантный, так и эндогенный вариант 1 STEAP-1. При осуществлении схемы иммунизации через 7-10 дней после инъекции берут контрольные пробы крови, чтобы определить титр и специфичность иммунного ответа. После достижения подходящих реакционноспособности и специфичности, определяемых методами ELISA, вестерн-блоттинга, иммунопреципитации, флуоресцентной микроскопии и проточной цитометрии, получают гибрид и гибридому с помощью общепринятых методов, хорошо известных в данной области (см., например, Harlow and Lane, 1988). Сродство связывания специфических моноклональных антител против варианта 1 STEAP-1 определяют с помощью стандартных методов. Измерение сродства позволяет количественно определить силу связывания антитела с эпитопом и помогает идентифицировать моноклональные антитела против варианта STEAP-1, являющиеся предпочтительными для диагностического или терапевтического применения, что оценивают специалисты в данной области. Система BIAcore (Uppsala, Sweden) является предпочтительным методом определения сродства связывания. В системе BIAcore для регистрации биомолекулярных взаимодействий в режиме реального времени используют метод поверхностного плазмонного резонанса (SPR, Welford K. 1991, Opt. Quant. Elect. 23: 1; Morton and Myszka, 1998, Methods in Enzymology 295: 268). Как правило, анализ BIAcore позволяет определять константы скорости ассоциации, константы скорости диссоциации, равновесные константы диссоциации и константы сродства. Чтобы получить моноклональные антитела, специфичные к другим вариантам STEAP-1, создают иммуногены, кодирующие аминокислотные последовательности, уникальные для данных вариантов. В одном воплощении синтезируют пептид, несущий аминокислоты, уникальные для вариантов STEAP-1, присоединяют его к KLH и полученный конъюгат используют в качестве иммуногена. В другом воплощении получают пептиды или бактериальные гибридные белки, которые содержат уникальную последовательность, образующуюся в результате альтернативного сплайсинга вариантов. Затем отбирают гибридомы, которые распознают антиген, характерный для соответствующего варианта, и, кроме того, распознают полноразмерный вариантный белок, экспрессирующийся в клетках. Такой отбор проводят с помощью описанных выше иммунологических анализов, таких как вестерн-блоттинг, иммунопреципитация и проточная цитометрия. В одном воплощении данное изобретение предлагает моноклональные антитела, обозначаемые Х92.1.30.1.1 (1) (a.k.a. М2/92.30) и Х120.545.1.1 (a.k.a. М2.120.545); М2/92.30 и М2/120.545 идентифицируют и демонстрируют, что они взаимодействуют и связываются с клеточным поверхностным STEAP-1 (см. фиг. 15 и 18). На фиг. 16 показано, что Mab M2/92.30 против STEAP-1 связывается с эндогенным клеточным поверхностным STEAP-1, экспрессирующимся в клетках мочевого пузыря и ксенотрансплантатов рака простаты. Кроме того, М2/92.30 взаимодействует и связывается с мышиным STEAP-1, как изображено на фиг. 17. Антитела, обозначенные Х92.1.30.1.1 (1) (a.k.a. M2/92.30) и Х120.545.1.1 (a.k.a. М2.120.545), отправлены (с помощью службы Федерал-Экспресс) в Американскую коллекцию типовых культур (АТСС), Р. О. Box 1549, Manassas, VA 20108, 6 февраля 2004 г., где им присвоены номера доступа РТА-5802 и РТА-5803 соответственно. Для клонирования антител М2/Х92.30 и М2/Х120.545 используют следующие способы. Клетки гибридомы лизируют реагентом Trizol (Life Technologies, Gibco BRL). Общую РНК очищают и количественно определяют. Одноцепочечные кДНК получают из общей РНК с использованием олиго(dT) 12-18 в качестве праймеров и системы Gibco-BRL Superscript Preamplification. Продукты ПЦР клонируют в векторе pCRScript (Stratagene, La Jolla). Некоторые клоны секвенируют и определяют вариабельные участки тяжелой ("V H ") и легкой ("V L ") цепей. Нуклеотидная и аминокислотная последовательности вариабельных участков тяжелой и легкой цепей М2/Х92.30 и М2/Х120.545 приведены на фиг. 19A-19D и 20A-20E. Пример 12. Характеристика антител STEAP-1. А. Клеточное поверхностное связывание. Способность антител STEAP-1 взамодействовать с белком, родственным STEAP-1, можно определить с помощью ряда хорошо известных методов, включающих в себя вестерн-блоттинг, иммунопреципитацию, ELISA и FACS, проводимые, в зависимости от обстоятельств, с использованием STEAP-1-родственных белков, STEAP-1-экспрессирующих клеток или их экстрактов. На фиг. 15 показаны результаты анализа FACS рекомбинантных клеток 3Т3 и RAT1, стабильно экспрессирующих либо STEAP-1, либо контрольный ген, окрашенных Mab M2/92.30 против STEAP (10 мкг/мл), где связанные с клеточной поверхностью Mab детектируют, используя в качестве вторичного реагента конъюгат козлиных антител против мышиных IgG с РЕ. Окрашенные клетки анализируют методом FACS. Сдвиг флуоресценции в клетках RAT1-STEAP-1 и 3T3-STEAP-1 по сравнению с контрольными клетками показывает, что Mab М2/92.30 специфически связываются со STEAP-1 на клеточной поверхности. Кроме того, проводят окрашивание клеток мочевого пузыря UGB1 и клеток рака простаты LAPC-9 с использованием 10 мкг/мл либо Mab М2/92.30, либо контрольных Mab против KLH. Связанные с поверхностью Mab М2/92.30 детектируют, используя в качестве вторичных Ab конъюгат козлиных антител против мышиных IgG с РЕ. Окрашенные клетки анализируют методом FACS. Полученные результаты показывают, что Mab М2/92.30 против STEAP-1 специфически связывается с эндогенным STEAP-1, экспрессирующимся на поверхности клеток ксенотрансплантатов рака мочевого пузыря и простаты (фиг. 21). Показано также, что М2/92.30 против STEAP-1 связывается с мышиным белком STEAP-1 (см. фиг. 17). В данном эксперименте клетки 293Т транзиторно трансфицируют либо pCDNA3.1, кодирующим кДНК мышиного STEAP-1, либо пустым вектором. Через 48 ч клетки собирают и окрашивают Mab М2/92.30 против STEAP-1 (10 мкг/мл), после чего связанные с клеточной поверхностью Mab детектируют, используя в качестве вторичного реагента конъюгат козлиных антител против мышиных IgG с РЕ. Затем клетки анализируют методом FACS. Результаты показывают, что Mab М2/92.30 против STEAP-1 специфически связывается с клетками 293Т, экспрессирующими STEAP-1. Показано также, что М2/120.545 против STEAP-1 специфически связывается со STEAP-1 (см. фиг. 18). Клетки 3Т3-neo (сектор А, закрашенная гистограмма) и 3T3-STEAP-1 (сектор А, незакрашенная гистограмма), а также клетки RAT1-neo (сектор В, закрашенная гистограмма) и RAT1-STEAP (сектор В, незакрашенная гистограмма) окрашивают Mab М2/120.545 (10 мкг/мл) и связанные с поверхностью Mab детектируют, используя в качестве вторичных Ab конъюгат козлиных антител против мышиных IgG с РЕ. Затем клетки анализируют методом FACS. Сдвиг флуоресценции в клетках 3T3-STEAP-1 и RAT1-STEAP-1 по сравнению с соответствующими neo-контролями показывает, что Mab M2/120.545 специфически связывается со STEAP-1 на клеточной поверхности. В секторе С показаны результаты анализа FACS, как описано выше, клеток LNCaP, окрашенных либо Mab M2/120.545, либо контрольными Mab против KLH. В секторе D показаны результаты флуоресцентной микроскопии клеток LNCaP, окрашенных М2/120.545, демонстрирующие интенсивную флуоресценцию на клеточной поверхности. Реакционноспособность и специфичность М2/92.30 и М2/120.545 также определяют методом иммунопреципитации. На фиг. 25 приведены результаты эксперимента, в котором клетки 3T3-STEAP-1 и 3Т3-neo лизируют в буфере RIPA (25 мМ Tris-Cl pH 7,4; 150 мМ NaCl, 0,5 мМ EDTA, 1% тритон Х-100, 0,5% дезоксихолевая кислота, 0,1% SDS и смесь ингибиторов протеаз). Клеточные лизаты предварительно осветляют с помощью гранул сефарозы, содержащих белок G, и затем инкубируют с 5 мкг Mab, либо М2/92.30, либо М2/120.545, в течение 2 ч при комнатной температуре. Добавляют гранулы, содержащие белок G, и смесь инкубируют еще 1 ч. Иммунные комплексы промывают и солюбилизируют в буфере для SDS-PAGE. Затем солюбилизированные образцы анализируют методами SDS-PAGE и вестерн-блоттинга, используя кроличьи pAb против STEAP. В качестве положительного контроля используют лизаты целых клеток 293Т, трансфицированных STEAP-1. Иммунореактивная полоса, соответствующая ~37 кДа, наблюдается только в образцах, полученных из клеток 3T3-STEAP-1, и указывает на специфичную иммунопреципитацию STEAP-1 под действием как Mab M2/92.30, так и Mab M2/120.545. В. Интернализация антитела против STEAP-1. Иммунотерапия, основанная на доставке токсинов к конкретным клеткам-мишеням с помощью моноклональных антител, относится к способам лечения злокачественных заболеваний. Общим принципом данных способов является доставка токсинов или противоопухолевых средств к раковым клеткам с помощью молекул, которые связываются с антигенами или рецепторами, экспрессирующимися только в клетках-мишенях или обладающими повышенным уровнем экспрессии в клетках-мишенях по сравнению с нормальными тканями. Иммунотоксины состоят из клеточно-селективных лигандов (как правило, моноклональных антител или цитокинов), ковалентно связанных с токсинами. Взаимодействие антитела или лиганда с рецепторами клеточной поверхности инициирует интернализацию. В определенных внутриклеточных полостях-компартментах токсиновый фрагмент отделяется и попадает в цитозоль, где он каталитически изменяет ключевые клеточные функции и приводит к гибели клетки. См., Frankel А.Е., Increased Sophistication of Immunotoxins, Clinical cancer research, 8: 942-944 (2002) и Allen T.M., Ligand-Targeted Therapeutics in Anti-cancer Therapy. Nature Reviews. 2: 750-760 (2002). Сапорин представляет собой белок, инактивирующий рибосомы (RIP), который катализирует in vitro депуринизацию конкретного аденинового остатка в больших рибосомальных РНК. Endo Y., et. al., Mechanism of Action of the Toxin Lectin Ricin on Eukaryotic Cells; The Site and Characteristics of the Modification in 28S RNA Caused by the Toxin, J. Biol. Chem. 262, 5908-5912 (1987). Как правило, он способен проникать в клетки только в составе комплекса с подходящей молекулой-носителем. Путем ковалентной конъюгации сапорина с моноклональными антителами, распознающими опухолевые антигены, получают иммунотоксины, которые обладают селективностью по отношению к раковым клеткам и способны к интернализации. См., Flavell, D.J., Sapoin Immunotoxins, Curr. Top. Microbiol. Immunol. 234: 51-61 (1998) и Flavell D.J., et. al., Therapy of Human T-cell Acute Lymphoblastic Leukemia with a Combination of Anti-CD7 and Anti-CD38-Saporin Immunotoxins is Significantly Better than Therapy with Each Individual Immunotoxins. Br. J. Cancer, 84: 571-578 (2001). Данные молекулы недавно поступили в I фазу клинических испытаний по лечению лейкоза и множественной миеломы. Foon K.A. Monoclonal Antibody Therapies for Lymohomas. Cancer J. 6: 273-278 (2000). Интернализация STEAP-1 с помощью М2/92.30 показана на фиг. 22. В данном эксперименте клетки PC3-STEAP-1 окрашивают при 4 °С с помощью Mab M2/120.545 (10 мкг/мл), промывают и инкубируют с конъюгатом козлиные антитела против мышиных IgG-РЕ в качестве вторичных Ab. Половину клеток инкубируют при 37 °С в течение 30 мин, а вторую половину оставляют при 4 °С. Затем клетки из каждой партии анализируют методом флуоресцентной микроскопии. На поверхности клеток, оставленных инкубироваться при 4 °С, наблюдается яркая "кольцеобразная" флуоресценция. На поверхности клеток, инкубировавшихся при 37 °С, "кольцеобразная" флуоресценция отсутствует, а появление точечной и агрегированной флуоресценции указывает на кэпирование и интернализацию. Интернализация STEAP-1 с помощью Mab M2/120.545 против STEAP-1 показана на фиг. 23. Клетки PC3-STEAP-1 окрашивают при 4 °С с помощью Mab M2/120.545 (10 мкг/мл), промывают и инкубируют с конъюгатом козлиные антитела против мышиных IgG-РЕ в качестве вторичных Ab. Половину клеток инкубируют при 37 °С в течение 30 мин, а вторую половину оставляют при 4 °С. Затем клетки из каждой партии анализируют методом флуоресцентной микроскопии. На поверхности клеток, оставленных инкубироваться при 4 °С, наблюдается яркая "кольцеобразная" флуоресценция. На поверхности клеток, инкубировавшихся при 37 °С, "кольцеобразная" флуоресценция отсутствует, а появление точечной и агрегированной флуоресценции указывает на кэпирование и интернализацию. В одном способе антитела-кандидаты, подходящие для доставки иммунотоксинов, выбирают путем уничтожения клеток с помощью вторичных антител, конъгированных с лекарственным средством или молекулой токсина. Вторичное конъюгированное антитело, переносимое на первичном антителе, позволяет детектировать интернализацию и перемещение первичного антитела в соответствующие внутриклеточные компартменты. После интернализации конъюгата сапорин отщепляется от направляющего средства и инактивирует рибосомы, приводя к гибели клеток-мишеней. Kohls M.D. and Lappi DA. Mab-ZAP: A Tool for Evaluating Antibody Efficacy for Use in an Immunotoxin. Bio Techniques. 28(1): 162-165 (2000). Чтобы выбрать подходящее антитело-кандидат с помощью вышеуказанного способа, в качестве иммунотоксина используют конъюгат антитела против мышиного IgG с сапорином (Advanced Targeting Systems, San Diego, CA), позволяющий продемонстрировать, что мышиный STEAP-1 М2/120.545 проникает в клетки-мишени благодаря экспрессии STEAP-1 на поверхности клеток LNCaP. Анализ проводят следующим образом. Клетки LNCaP помещают в количестве 5000 клеток/90 мкл/лунку в 96-луночный планшет и инкубируют в течение ночи. Получают растворы конъюгатов вторичных антител с иммунотоксином (антитела против мышиных IgG-сапорин и антитела против козлиных IgG-сапорин) и антител против мышиных IgG в среде для культивирования клеток с конечной концентрацией 100 нг/мл. В каждую лунку добавляют 10 мкл полученных растворов. Добавляют первичное антитело в концентрации, варьирующей от 1 до 1000 нг/мл. Планшеты инкубируют в течение 72 ч, после чего методом МТТ определяют жизнеспособность клеток. Результаты, приведенные на фиг. 24, показывают, что клетки LNCaP погибают в присутствии конъюгата антитело против мышиных IgG-сапорин. При использовании только вторичных антител (против мышиных IgG) или конъюгатов неспецифичных вторичных антител (антител против козлиных IgG с сапорином) эффект не детектируется. При использовании только первичного антитела (М2/120.545) вплоть до концентрации 1 мкг/мл токсичность не наблюдается. Пример 13. Анализы связывания HLA класса I и класа II. Анализы связывания HLA класса I и класса II с использованием очищенных молекул HLA проводят в соответствии с раскрытыми способами (например, в публикациях РСТ WO 94/20127 и WO 94/03205; Sidney et al., Current Protocols in Immunology, 18.3.1 (1998); Sidney, et al., J. Immunol. 154: 247 (1995); Sette, et al., Mol. Immunol. 31: 813 (1994)). Коротко говоря, очищенные молекулы МНС (от 5 до 500 нМ) инкубируют в присутствии различных немеченых пептидных ингибиторов и 1-10 нМ меченных радиоактивным изотопом 125 I пептидных зондов по описанному способу. После инкубации комплексы МНС-пептид отделяют от несвязанного пептида путем гель-фильтрации и определяют фракции связанного пептида. Как правило, перед проведением эксперимента каждый препарат МНС титруют в присутствии фиксированного количества меченных радиоактивным изотопом пептидов, чтобы определить концентрацию молекул HLA, необходимую для связывания 10-20% общего количества радиоактивности. Все последующие анализы ингибирования и непосредственного связывания проводят, используя данные концентрации HLA. Поскольку в данных условиях [label] <[HLA], a IC 50 ≥[HLA], измеренные значения IC 50 являются приемлемыми приближениями фактических значений K D . Пептидные ингибиторы обычно используют в концентрациях, варьирующих от 120 мкг/мл до 1,2 нг/мл, и анализируют в двух-четырех независимых экспериментах. Чтобы сравнить результаты, полученные в разных экспериментах, для каждого пептида получают график относительного связывания путем деления IC 50 положительного контроля ингибирования на IC 50 каждого анализируемого пептида (как правило, немеченного пептида по сравнению с меченным радиоактивным изотопом пептидным зондом). Для формирования базы данных и сравнения результатов разных экспериментов, компилируют значения относительного связывания. Затем данные значения снова превращают в значения IC 50 , нМ, путем деления IC 50 , нМ, положительных контролей ингибирования на относительное связывание анализируемого пептида. Указанный способ компиляции данных является точным и позволяет получать согласующиеся результаты для сравнения пептидов, тестируемых в разные дни или с использованием разных партий очищенных МНС. Описанные выше анализы связывания можно использовать для тестирования пептидов, несущих супермотив HLA и/или мотив HLA (см. табл. IV). Пример 14. Конструирование полиэпитопных ДНК-плазмид, несущих "мини-ген". В данном примере описывается конструирование плазмиды экспрессии, несущей мини-ген. Разумеется, плазмиды, несущие мини-гены, могут содержать разные конфигурации эпитопов или аналогов эпитопов В-клеток, CTL и/или HTL, описанных в данном документе. Плазмида экспрессии, несущая мини-ген, как правило, содержит несколько пептидных эпитопов CTL и HTL. В настоящем примере пептидные эпитопы, несущие супермотивы HLA-A2, -A3, -В7, и пептидные эпитопы, несущие мотивы HLA-A1 и -А24, используют в сочетании с эпитопами, несущими супермотив DR, и/или с эпитопами DR3. Супермотив HLA класса I или мотив-несущие пептидные эпитопы STEAP-1 выбирают так, чтобы было представлено несколько супермотивов/мотивов, обеспечивающих широкий популяционный охват. Подобным образом, эпитопы HLA класса II выбирают из последовательности STEAP-1 так, чтобы обеспечить широкий популяционный охват, т.е. в состав конструкции, несущей мини-ген, включают и эпитопы, несущие супермотив HLA DR-1-4-7, и эпитопы, несущие мотив HLA DR-3. Затем выбранные эпитопы CTL и HTL вводят в состав мини-гена для экспрессии в векторе экспрессии. Такая конструкция может дополнительно включать в себя последовательности, направляющие эпитопы HTL в эндоплазматический ретикулум. Например, белок li может быть присоединен к одному или нескольким эпитопам HTL, как описано в данной области, где последовательность CLIP белка li удаляют и заменяют на последовательность эпитопа HLA класса II для того, чтобы направить эпитоп HLA класса II в эндоплазматический ретикулум, где данный эпитоп связывается с молекулами HLA класса II. В данном примере иллюстрируются способы, используемые для конструирования плазмиды экспрессии, несущей мини-ген. Для композиции мини-гена можно использовать и другие векторы экспрессии, известные специалистам в данной области. ДНК-плазмида данного примера, несущая мини-ген, содержит консенсусную последовательность Козака и консенсусную сигнальную последовательность легкой цепи каппа мышиного Ig, затем эпитопы CTL и/или HTL, выбранные в соответствии с описанными в данном документе принципами. Данная последовательность кодирует открытую рамку считывания, гибридизованную с маркерным эпитопом антитела против Мус и His, кодируемую вектором pcDNA3.1Myc-His. Синтезируют перекрывающиеся олигонуклеотиды, которые могут содержать в длину в среднем приблизительно 70 нуклеотидов, из которых 15 нуклеотидов перекрываются, и очищают их методом ВЭЖХ. Олигонуклеотиды кодируют выбранные пептидные эпитопы, а также содержат подходящие линкерные нуклеотиды, последовательность Козака и сигнальную последовательность. Конечный полиэпитопный мини-ген собирают путем удлинения перекрывающихся олигонуклеотидов в трех циклах реакций ПЦР. Используют машину для проведения ПЦР Perkin/Elmer 9600 и всего 30 циклов реакции с применением следующих условий: 95 °С в течение 15 с, температура отжига (на 5 ° ниже самой низкой рассчитанной Tm для каждой пары праймеров) в течение 30 с и 12 °С в течение 1 мин. Например, мини-ген получают следующим образом. В первой реакции ПЦР 5 мкг каждого из двух олигонуклеотидов подвергают отжигу и удлинению: в примере с использованием восьми олигонуклеотидов, т.е. четырех пар праймеров, олигонуклеотиды 1+2, 3+4, 5+6 и 7+8 объединяют в 100 мкл реакционных смесей, содержащих буфер для полимеразы Pfu (1 ×=10 мМ KCl, 10 мМ (NH 4 ) 2 SO 4 , 20 мМ Tris-хлорид, рН 8,75, 2 мМ MgSO 4 , 0,1% тритон Х-100, 100 мкг/мл БСА), 0,25 мМ каждого dNTP и 2,5 ед. полимеразы Pfu. Полноразмерные димерные продукты очищают в геле, две реакционные смеси, содержащие продукты 1+2 и 3+4, а также продукты 5+6 и 7+8, смешивают, подвергают отжигу и удлинению в течение 10 циклов. Затем смешивают половину двух реакционных смесей и проводят 5 циклов отжига и удлинения, после чего добавляют фланкирующие праймеры и амплифицируют полноразмерный продукт. Полноразмерный продукт очищают в геле, клонируют в векторе pCR-blunt (Invitrogen) и проводят скрининг отдельных клонов путем секвенирования. Пример 15. Плазмидная конструкция и степень ее иммуногенности. Степень иммуногенности плазмидной конструкции, например плазмидной конструкции, полученной по способу предыдущего примера, подтверждают in vitro путем определения презентации эпитопа АРС после трансдукции или трансфекции АРС конструкцией нуклеиновой кислоты, экспрессирующей эпитоп. Такое исследование позволяет определять "антигенность" с применением человеческих АРС. Данный анализ позволяет оценить способность АРС презентировать эпитоп, распознаваемый Т-клетками, путем количественного определения плотности комплексов эпитоп-HLA класса I на поверхности клеток. Количественное определение можно проводить путем непосредственного измерения количества пептида, элюируемого из АРС (см., например, Sijts et al., J. Immunol. 156: 683-692, 1996; Demotz et al., Nature, 342: 682-684, 1989); или количество комплексов пептид-HLA класса I можно определить путем количественного определения лизиса или высвобождения лимфокинов из больных или трансфицированных клеток с последующим определением концентрации пептида, необходимой для получения эквивалентных уровней лизиса или высвобождения лимфокинов (см., например, Kageyama et al., J. Immunol. 154: 567-576, 1995). Альтернативно, иммуногенность подтверждают путем in vivo введения мышам с последующей оценкой активности CTL и HTL in vitro с помощью анализов цитотоксичности и пролиферации соответственно, подробно описанных, например, в Alexander et al., Immunity, 1: 751-761, 1994. Например, чтобы подтвердить способность ДНК-конструкции, несущей мини-ген и кодирующей по меньшей мере один пептидный супермотив HLA-A2, индуцировать CTL in vivo, трансгенных мышей HLA-A2.1/K b , например, иммунизируют внутримышечно 100 мкг оголенной кДНК. Для сравнения уровня CTL, индуцированных в результате иммунизации кДНК, контрольную группу животных иммунизируют истинной пептидной композицией, которая содержит несколько эпитопов, синтезированных в виде одного полипептида, которые могли бы кодироваться анализируемым мини-геном. Спленоциты иммунизированных животных стимулируют дважды каждой из соответствующих композиций (пептидными эпитопами, кодируемыми мини-геном или полиэпитопным пептидом), затем анализируют на пептид-специфическую цитотоксичную активность по высвобождению 51 Cr. Результаты позволяют определить величину CTL-ответа, направленного против А2-ограниченного эпитопа, и оценить иммуногенность вакцины, несущей мини-ген, и полиэпитопной вакцины in vivo. В результате обнаружено, что мини-ген вызывает иммунные ответы, направленные против пептидных эпитопов супермотива HLA-А2, как и полиэпитопная пептидная вакцина. Чтобы оценить индукцию CTL под действием эпитопов мотива или супермотива HLA-А3 и HLA-B7, проводят подобный анализ с использованием других трансгенных мышиных моделей HLA-A3 и HLA-B7, в результате которого обнаруживают, что мини-ген вызывает соответствующие иммунные ответы, направленные против предоставленных эпитопов. Чтобы подтвердить способность мини-гена, кодирующего эпитоп класса II, индуцировать HTL in vivo, трансгенных мышей DR или в случае эпитопов, которые перекрестно взаимодействуют с соответствующей молекулой мышиного МНС, 1-A b -ограниченных мышей, например, иммунизируют внутримышечно 100 мкг плазмидной ДНК. Для сравнения уровня HTL, индуцированных в результате иммунизации ДНК, контрольную группу животных иммунизируют истинной пептидной композицией, эмульгированной в полном адъюванте Фрейнда. Т-клетки CD4+, т.е. HTL, выделяют из спленоцитов иммунизированных животных и стимулируют каждой из соответствующих композиций (пептиды, кодируемые мини-геном). Ответ HTL измеряют с использованием анализа пролиферации по включению 3 Н-тимидина (см., например, Alexander et al. Immunity, 1: 751-761, 1994). Результаты позволяют определить величину ответа HTL, соответствующую иммуногенности мини-гена in vivo. ДНК-мини-гены, полученные по способу, описанному в предыдущем примере, также можно использовать в качестве вакцины в сочетании с иммунизирующим средством по схеме примирование-повторная иммунизация. Иммунизирующее средсто может включать в себя рекомбинантный белок (например, Barnett et al., Aids Res. and Human Retroviruses 14, Supplement, 3: S299-S309, 1998) или рекомбинантную вакцину, например, экспрессирующую мини-ген или ДНК, кодирующую полноразмерный белок, представляющий интерес (см., например, Hanke et al., Vaccine, 16: 439-445, 1998; Sedegah et al., Proc. Natl. Acad. Sci. USA, 95: 7648-53, 1998; Hanke and McMichael, Immunol. Letters, 66: 177-181, 1999 и Robinson et al., Nature Med. 5: 526-34, 1999). Например, эффективность ДНК-мини-гена при иммунизации по схеме примирование-повторная иммунизация вначале оценивают у трансгенных мышей. В данном примере трансгенных мышей А2.1/K b иммунизируют в.м. 100 мкг ДНК-мини-гена, кодирующего иммуногенные пептиды, включающие в себя по меньшей мере один пептид, несущий супермотив HLA-A2. После инкубационного периода (варьирующего от 3 до 9 недель) мышей иммунизируют в.б. 10 7 бое/мышь рекомбинантного вируса коровьей оспы, экспрессирующего последовательность, идентичную последовательности, кодируемой ДНК-мини-геном. Контрольных мышей иммунизируют 100 мкг ДНК или рекомбинантного вируса коровьей оспы, не содержащих последовательности мини-гена, или ДНК, содержащей мини-ген, но не содержащей иммуноген коровьей оспы. После другого инкубационного периода, составляющего 2 недели, спленоциты мышей сразу анализируют на пептидспецифическую активность методом ELISPOT. Кроме того, спленоциты стимулируют in vitro А2-ограниченными пептидными эпитопами, кодируемыми мини-геном и рекомбинантным вирусом коровьей оспы, затем анализируют на пептидспецифичную активность методом альфа, бета и/или гамма IFN ELISA. Обнаружено, что мини-ген при иммунизации по схеме примирование-повторная иммунизация вызывает более сильный иммунный ответ против пептидов супермотива HLA-A2, чем одна ДНК. Чтобы оценить индукцию CTL под действием эпитопов мотива или супермотива HLA-A3 и HLA-B7, проводят подобный анализ с использованием трансгенных мышиных моделей HLA-A11 и HLA-B7. Применение схемы примирование-повторная иммунизация у людей описано ниже в примере, озаглавленном "Индукция ответов CTL с использованием схемы примирование-повторная иммунизация". Пример 16. Полиэпитопные вакцинные композиции, содержащие несколько антигенов. Чтобы создать вакцинную композицию для профилактики или лечения рака, сопровождающегося экспрессией STEAP-1 и других подобных антигенов, пептидные эпитопы STEAP-1 согласно изобретению используют в сочетании с эпитопами других опухолеспецифических антигенов-мишеней. Например, вакцинную композицию можно получить в виде одного полипептида, который содержит несколько эпитопов из STEAP-1, а также из опухолеспецифических антигенов, которые часто экспрессируются при злокачественном заболевании, связанном с экспрессией STEAP-1, или вакцинную композицию можно получить в виде смеси одного или нескольких отдельных эпитопов. Альтернативно, вакцину можно вводить в виде конструкции мини-гена или в виде дендритных клеток, нагруженных пептидными эпитопами in vitro. Пример 17. Применение пептидов для оценки иммунного ответа. Пептиды согласно изобретению можно использовать для анализа иммунного ответа по присутствию специфических антител, CTL или HTL, направленных на STEAP-1. Такой анализ можно проводить по способу, описанному Ogg et al., Science, 279: 2103-2106, 1998. В данном примере пептиды согласно изобретению используют в качестве реагента для диагностических или прогностических целей, но не в качестве иммуногена. В данном примере высокочувствительные тетрамерные комплексы ("тетрамеры") человеческого лейкоцитарного антигена используют для перекрестного анализа, например частоты встречаемости STEAP-1 HLA-A*0201-специфических CTL у HLA А*0201-положительных субъектов на разных стадиях заболевания или после иммунизации пептидом STEAP-1, содержащим мотив А*0201. Тетрамерные комплексы синтезируют по описанному в литературе способу (Musey et al., N. Engl. J. Med. 337: 1267, 1997). Коротко говоря, очищенную тяжелую цепь HLA (A*0201 в данном примере) и β2-микроглобулин синтезируют с использованием прокариотической системы экспрессии. Тяжелую цепь модифицируют путем делеции трансмембранно-цитозольного хвоста и добавления по СООН-концу последовательности, содержащей участок ферментативного биотинилирования BirA. Рефолдинг тяжелой цепи, β2-микроглобулина и пептида достигают путем разбавления. Продукт размером 45 кДа после рефолдинга выделяют методом жидкостной экспресс-хроматографии белков и затем биотинилируют под действием BirA в присутствии биотина (Sigma, St. Louis, Missouri), аденозин-5'-трифосфата и магния. Добавляют конъюгат стрептавидин-фикоэритрин в молярном соотношении 1:4 и концентрируют тетрамерный продукт до концентрации 1 мг/мл. Полученный продукт называют тетрамер-фикоэритрин. Чтобы проанализировать образцы крови пациента, приблизительно 1 миллион РВМС центрифугируют при 300g в течение 5 мин и ресуспендируют в 50 мкл холодного забуференного фосфатом физиологического раствора. Трехцветный анализ проводят с использованием тетрамер-фикоэритрина и трехцветного красителя против CD8 и CD38. РВМС инкубируют с тетрамером и антителами на льду в течение 30-60 мин, затем дважды промывают и фиксируют формальдегидом. Устанавливают границы измерения, включающие в себя значения > 99,98% от контрольных образцов. Контроли по тетрамеру включают в себя как А*0201-отрицательных субъектов, так и А*0201-положительных здоровых доноров. Затем методом проточной цитометрии определяют процент клеток, окрашенных тетрамером. Результаты позволяют определить в образце РВМС число клеток, содержащих эпитоп-ограниченные CTL, посредством этого величину иммунного ответа на эпитоп STEAP-1 и, следовательно, статус экспонирования STEAP-1 или экспонирования для вакцины, которая индуцирует защитный или терапевтический ответ. Пример 18. Индуцирование иммунных ответов с использованием схемы примирование-повторная иммунизация. Схему примирование-повторная иммунизация, подобную по своей основе схеме, используемой для подтверждения эффективности ДНК-вакцины у трансгенных мышей, такой как описанная выше в примере, озаглавленном "Плазмидная конструкция и степень ее иммуногенности", также можно использовать для введения вакцины людям. Такой режим вакцинирования может включать в себя первичное введение, например, оголенной ДНК, сопровождающееся повторной иммунизацией путем введения в адъюванте рекомбинантного вируса, кодирующего вакцину, или рекомбинантного белка/полипептида, или пептидной смеси. Например, для первичной иммунизации можно использовать вектор экспрессии, такой как полученный в примере, озаглавленном "Конструирование полиэпитопных ДНК-плазмид, несущих "мини-ген", в виде оголенной нуклеиновой кислоты, который вводят в.м. (или п.к., или в.к.) в количестве 0,5-5 мг в несколько участков. Нуклеиновую кислоту (от 0,1 до 1000 мкг) также можно вводить методом генной пушки. После периода инкубации, составляющего 3-4 недели, вводят повторную дозу антигена. При повторной иммунизации можно вводить рекомбинантный вирус птичьей оспы в дозе от 5 ×10 7 до 5 ×10 9 бое. Для повторной иммунизации также можно использовать альтернативный рекомбинантный вирус, такой как MVA, вирус канареечной оспы, аденовирус или адено-ассоциированный вирус либо полиэпитопный белок или смесь пептидов. Для оценки эффективности вакцины образцы крови пациента берут до иммунизации, а также через определенные интервалы после введения первичной вакцины и повторной дозы вакцины. Мононуклеарные клетки периферической крови выделяют из свежей гепаринизированной крови путем центрифугирования в градиенте плотности фикол-гипак, аликвоты помещают в среду для замораживания и хранят в замороженном состоянии. Образцы анализируют на активность CTL и HTL. Анализ полученных результатов показывает, что величина индуцируемого ответа достаточна для формирования терапевтического или защитного иммунитета против STEAP-1. Пример 19. Комплементарные полинуклеотиды. Последовательности, комплементарные STEAP-1-кодирующим последовательностям или их частям, используют для детектирования, уменьшения или ингибирования экспрессии природного STEAP-1. Описано применение олигонуклеотидов, содержащих приблизительно от 15 до 30 пар оснований, однако практически такой же способ можно использовать в применении к более мальньким или более крупным фрагментам последовательности. Подходящие олигонуклеотиды получают, например, с использованием программного обеспечения OLIGO 4.06 (National Biosciences) и кодирующей последовательности STEAP-1. Чтобы ингибировать транскрипцию, получают олигонуклеотид, комплементарный наиболее уникальной 5'-последовательности, и используют его для предотвращения связывания промотора с кодирующей последовательностью. Чтобы ингибировать трансляцию, получают комплементарный олигонуклеотид, предотвращающий связывание рибосом с транскриптом, кодирующим STEAP-1. Пример 20. Очистка природного или рекомбинантного STEAP-1 с помощью STEAP-1-специфических антител. Природный или рекомбинантный STEAP-1, по существу, очищают методом иммуноаффинной хроматографии с использованием антител, специфичных для STEAP-1. Иммуноаффинную колонку получают путем ковалентного присоединения антитела против STEAP-1 к активированной хроматографической смоле, такой как CNBr-активированная сефароза (Amersham Pharmacia Biotech). После присоединения смолу блокируют и промывают в соответствии с инструкциями производителя. Среду, содержащую STEAP-1, пропускают через иммуноаффинную колонку, после чего колонку промывают в условиях, обеспечивающих предпочтительную абсорбцию STEAP-1 (например, буферами с высокой ионной силой в присутствии детергента). Колонку элюируют в условиях, разрушающих связывание антитела со STEAP-1 (например, с использованием буфера, имеющего рН от 2 до 3, или содержащего высокую концентрацию агента, вызывающего диссоциацию комплексов, такого как мочевина или ион тиоцианата) и собирают GCR.P. Пример 21. Идентификация молекул, взаимодействующих со STEAP-1. STEAP-1 или его биологически активные фрагменты метят 121 I с использованием реактива Болтона-Хантера (см., например, Bolton et al. (1973), Biochem. J. 133: 529). Молекулы-кандидаты, предварительно размещенные в лунки многолуночного планшета, инкубируют с меченнм STEAP-1, промывают и определяют лунки, содержащие меченый комплекс STEAP-1. Результаты, полученные для разных концентраций STEAP-1, используют для расчета количества комплексов STEAP-1, сродства STEAP-1 к молекулам-кандидатам и степени ассоциации STEAP-1 с молекулами-кандидатами. Пример 22, Анализ инициации опухолевого роста под действием STEAP-1 in vivo. Влияние белка STEAP-1 на рост опухолевых клеток определяют in vivo путем оценки развития опухоли и роста клеток, экспрессирующих или не экспрессирующих STEAP-1. Например, мышам SCID подкожно вводят с каждой стороны 1 ×10 5 клеток либо 3Т3, либо линии рака простаты (например, клеток РС3), содержащих пустой вектор tkneo или STEAP-1. Можно использовать по меньшей мере две стратегии: (1) стратегия, в которой обеспечивают конститутивную экспрессию STEAP-1 под регуляцией такого промотора, как конститутивный промотор, полученный из геномов вирусов, таких как вирус полиомы, вирус птичьей оспы (UK 2211504, опубликованный 5 июля 1989 г.), аденовирус (такой как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и обезьяний вирус 40 (SV40), или из гетерологичных промоторов млекопитающих, таких как промотор актина или промотор иммуноглобулина, при условии, что такие промоторы совместимы с системами клетки-хозяина; и (2) стратегия, в которой обеспечивают регулируемую экспрессию, находящуюся под контролем индуцируемой векторной системы, такой как экдизон, тетрациклин и др., при условии, что такие промоторы совместимы с системами клетки-хозяина. Затем с помощью штангенциркуля измеряют объем опухоли в участке пальпируемой опухоли и через некоторое время определяют, не обладают ли STEAP-1-экспрессирующие клетки более высокой скоростью роста и не обладают ли опухоли, образованные STEAP-1-экспрессирующими клетками, признаками, характерными для изменения агрессивности (такими как повышенное метастазирование, васкуляризация, пониженная чувствительность к химиотерапевтическим средствам). Кроме того, мышам можно имплантировать 1 ×10 5 таких же клеток в гомологичную область, чтобы определить, влияет ли STEAP-1 на локальный рост в простате и влияет ли STEAP-1 на способность опухолей к метастазированию, особенно в лимфатические узлы и кости (Miki T. et al., Oncol Res. 2001; 12: 209; Fu X. et al., Int. J. Cancer. 1991, 49: 938). Влияние STEAP на образование и рост опухоли в кости можно оценить путем инъекции клеток опухоли простаты в большеберцовую кость. С помощью данного анализа также можно определить способность терапевтических композиций-кандидатов, таких как, например, интратела STEAP-1, антисмысловые молекулы STEAP-1 и рибозимы, оказывать ингибиторное действие на STEAP-1. Пример 23. Ингибирование роста опухолей in vivo, опосредованное моноклональными антителами STEAP-1. Значительный уровень экспрессии STEAP-1 в раковых тканях и поверхностная локализация STEAP-1, наряду с ограниченной экспрессией в нормальных тканях, делают STEAP-1 хорошей мишенью для терапии антителами. Аналогично, STEAP-1 может служить мишенью для иммунотерапии на основе Т-клеток. Терапевтическую эффективность Mab против STEAP-1 у мышиных моделей, несущих ксенотрансплантаты рака простаты человека, оценивают с использованием рекомбинантных клеточных линий, таких как РС3-STEAP-1 и 3T3-STEAP-1 (см., например, Kaighn, M.E., et al., Invest Urol., 1979, 17(1): 16-23), а также моделей, несущих ксенотрансплантаты простаты человека, таких как LAPC 9AD (Saffran et al. PNAS, 1999, 10: 1073-1078). Влияние антитела на рост опухоли и образование метастаз исследуют, например, на мышиных моделях, несущих ортотопические ксенотрансплантаты рака простаты. Антитела могут быть неконъюгированными, как описано в данном примере, или они могут быть конъюгированными с терапевтическим средством, такие конъюгаты известны в данной области. Mab против STEAP-1 ингибируют формирование ксенотрансплантатов легких и простаты. Mab против STEAP-1 также замедляют рост развитых ортотопических опухолей и увеличивают продолжительность жизни мышей, несущих опухоль. Полученные результаты указывают на возможность применения Mab против STEAP-1 для лечения рака простаты на стадиях локального и запущенного заболевания (см., например, Saffran, D., et al., PNAS, 10: 1073-1078 или сайт в глобальной сети URL pnas.org/cgi/doi/10.1073/pnas.051624698). Введение Mab против STEAP-1 приводит к замедлению роста развившейся ортотопической опухоли и к ингибированию метастаз в отдаленных участках, значительно увеличивая таким образом продолжительность жизни мышей, несущих опухоль. Полученные результаты показывают, что STEAP-1 является приемлемой мишенью для иммунотерапии и демонстрирует терапевтический потенциал Mab против STEAP-1 для лечения локального и метастазирующего рака простаты. В данном примере показано, что неконъюгированные моноклональные антитела STEAP-1 эффективно ингибируют рост ксенотрансплантатов опухоли простаты человека у мышей SCID; соответственно, сочетание таких эффективных моноклональных антител также является эффективным. Ингибирование опухоли с использованием нескольких неконъюгированных Mab STEAP-1. Материалы и методы. Моноклональные антитела STEAP-1. Моноклональные антитела против STEAP-1 получают по способу, описанному в примере, озаглавленном "Получение моноклональных антител (Mab) STEAP-1". Способность антител связываться со STEAP-1 определяют методами ELISA, вестерн-блоттинга, FACS и иммунопреципитации. Результаты эпитопного картирования Mab против STEAP-1, полученные с помощью методов ELISA и вестерн-блоттинга, свидетельствуют о том, что данные Mab распознают эпитопы на белке STEAP-1. Проводят иммуногистохимический анализ тканей и клеток рака простаты с использованием данных антител. Моноклональные антитела выделяют из асцитов или из супернатантов гибридомных тканевых культур путем хроматографии на сефарозе, содержащей G-белок, диализуют против PBS, стерилизуют фильтрацией и хранят при -20 °С. Определение белков проводят по методу Брэдфорда (Bio-Rad, Hercules, CA). Получают терапевтическое моноклональное антитело или смесь, содержащую отдельные моноклональные антитела, и используют для лечения мышей, получающих подкожные или ортотопические инъекции ксенотрансплантатов опухолей UM-UC3 и CaLu1. Клеточные линии и ксенотрансплантаты. Линии клеток рака простаты, РС3 и LNCaP, а также линию фибробластов NIH 3Т3 (Американская коллекция типовых культур) поддерживают в RPMI и DMEM соответственно, дополненных L-глутамином и 10% FBS. Популяции клеток PC3-STEAP-1 и 3T3-STEAP-1 получают путем переноса ретровирусного гена, как описано в Hubert, R.S., et al., Proc. Natl. Acad. Sci. USA, 1999, 96(25): 14523. Ксенотрансплантат LAPC-9, который экспрессирует рецептор андрогенов дикого типа и продуцирует специфический антиген простаты (PSA), пересаживают самцам мышей ICR с тяжелым комбинированным иммунодефицитом (SCID) (Taconic Farms) возрастом 6-8 недель путем п.к. имплантации с помощью трокара (Craft, N., et al., Nat. Med. 1999, 5: 280). Суспензионные культуры опухолевых клеток LAPC-9 получают по способу, описанному Craft, et al. Мышиные модели, несущие ксенотрансплантаты. Подкожные (п.к.) опухоли получают путем инъекции 1 ×10 6 раковых клеток, смешанных в соотношении 1:1 с матригелем (Collaborative Research), в правый бок самцов мышей SCID. Чтобы определить влияние антитела на формирование опухоли, инъекции антитела начинают в тот же день, что и инъекции раковых клеток. Контрольным мышам вводят либо очищенный мышиный IgG (ICN), либо PBS, либо очищенное моноклональное антитело, которое распознает другой антиген, не экспрессирующийся в клетках человека. В предварительных исследованиях не было обнаружено различия в воздействии мышиного IgG или PBS на рост опухоли. С помощью штангенциркуля определяют размеры опухолей и рассчитывают объем опухолей по формуле длина ×ширина ×высота. Мышей, у которых размер опухоли превышает в диаметре 1,5 см, умерщвляют. Ортотопические инъекции проводят под анестезией кетамином/ксилазином. При ортотопических исследованиях делают разрез стенки живота, открывая простату, и опухолевые клетки LAPC или РС3 (5 ×10 5 ), смешанные с матригелем, вводят в капсулу простаты в объеме 10 мкл. Рост опухолей отслеживают путем еженедельных пальпаций мышей и анализов крови на уровень PSA. Мышей разделяют на группы, которым в.б. вводят Mab против STEAP-1 или контрольные Mab. Mab против STEAP-1 ингибируют рост ксенотрансплантатов раковых опухолей, экспрессирующих STEAP-1. Влияние Mab против STEAP-1 на формирование опухоли анализируют путем применения ортотопических моделей LNCaP и LAPC-9. По сравнению с п.к. моделью опухоли ортотопическая модель, для получения которой опухолевые клетки вводят непосредственно в мышиную простату соответственно, приводит к локальному росту опухоли, развитию метастаз в удаленных участках, ухудшению здоровья мыши и последующей смерти (Saffran, D., et al., PNAS, приведено выше). Благодаря данным признакам ортотопическая модель лучше отражает развитие человеческого заболевания и позволяет отслеживать терапевтический эффект Mab на клинически релевантных конечных событиях. Соответственно, опухолевые клетки вводят в мышиную простату и через 2 дня мышей разделяют на две группы, которые обрабатывают либо а) 200-500 мкг Ab против STEAP-1, либо b) PBS три раза в неделю в течение 2-5 недель. Основным преимуществом ортотопических раковых моделей является то, что они позволяют исследовать развитие метастаз. Образование метастаз у мышей, несущих развившиеся ортотопические опухоли, исследуют с помощью анализа IHC на срезах легкого с использованием антитела против опухолеспецифического клеточного поверхностного белка, такого как антитело против CK20 в случае рака простаты (Lin S. et al., Cancer Detect Prev. 2001, 25: 202). Другим преимуществом моделей раковых ксенотрансплантатов является то, что они позволяют исследовать реваскуляризацию и ангиогенез. Опухолевый рост отчасти зависит от развития новых кровеносных сосудов. Хотя капиллярная система и развивающаяся кровеносная сеть по происхождению относятся к хозяину, инициация и архитектура реваскуляризации регулируются опухолью ксенотрансплантата (Davidoff A.M. et al., Clin. Cancer Res. 2001, 7: 2870; Solesvik О. et al., Eur. J. Cancer Clin. Oncol. 1984, 20: 1295). Влияние антител и маленьких молекул исследуют с помощью известных в данной области методов, таких как анализ опухолевых тканей и их микроокружения методом IHC. Мышам, несущим развившиеся ортотопические опухоли, вводят 1000 мкг либо Mab против STEAP-1, либо PBS в течение 4-недельного периода. Мышей из обеих групп оставляют для развития большой массы опухоли, чтобы обеспечить высокую вероятность образования метастаз в легких мышей. Затем мышей умерщвляют и анализируют их мочевые пузыри, печени, кости и легкие на присутствие опухолевых клеток методом IHC. Данные исследования демонстрируют широкую противоопухолевую эффективность антител против STEAP-1 при инициации и развитии рака простаты у мышиных моделей, несущих ксенотрансплантат. Антитела против STEAP-1 ингибируют образование опухолей, а также замедляют рост уже развившихся опухолей и увеличивают продолжительность жизни обработанных мышей. Более того, Mab против STEAP-1 сильно ингибируют распространение локальной опухоли простаты в отдаленные участки даже в случае большой массы опухоли. Таким образом, Mab против STEAP-1 оказывают эффективное влияние на большую часть клинически релевантных конечных событий (опухолевый рост), продолжительность жизни, состояние здоровья. Влияние Mab против STEAP-1 на рост ксенотрансплантатов рака простаты человека у мышей. Используют самцов мышей ICR-SCID возрастом 5-6 недель (Charles River Laboratory, Wilmington, MA). Мышей держат в контролируемых условиях в соответствии с инструкциями, приведенными в руководстве NIH по уходу за лабораторными животными и их применению. Для получения моделей, несущих ксенотрансплантат, используют андрогензависимую раковую опухоль простаты человека LAPC-9 AD. Исходные опухоли, обычно поддерживаемые в мышах SCID, вырезают в стерильных условиях, измельчают и обрабатывают проназой (Calbiochem, San Diego, CA). Полученные клеточные суспензии инкубируют в течение ночи при 37 °С, чтобы получить гомогенные суспензии отдельных клеток. М2/92.30 и М2/120.545 против STEAP-1 анализируют в двух разных дозах, 100 и 500 мкг. В качестве контролей используют PBS и Mab против KLH. В исследовании участвует 6 групп по 10 мышей в каждой. Mab вводят в.б. 2 раза в неделю, всего 12 раз, начиная со дня инъекции опухолевых клеток. Размер опухоли измеряют штангенциркулем 2 раза в неделю. Объем опухоли рассчитывают по формуле W 2 ×L/2, где L - самый длинный размер; W - перпендикулярный ему размер. Концентрацию PSA в сыворотке каждого животного измеряют на 40-й день после начала обработки с помощью коммерческого набора ELISA. Чтобы учесть различия в росте опухоли и уровне PSA между группами, используют тест Крускала-Валлиса и U-тест Манна-Уитни. Все тесты являются двусторонними с α=0,05. Результаты эксперимента, приведенные на фиг. 26 и 27, показывают, что М2/92.30 и М2/120.545 против STEAP-1 значительно замедляют рост ксенотрансплантата опухоли простаты человека в зависимости от используемой дозы. Пример 24. Терапевтическое и диагностическое применение антител против STEAP-1 у людей. Моноклональные антитела против STEAP-1 можно безопасно и эффективно использовать в применении у людей для диагностических, профилактических, прогностических и/или терапевтических целей. При проведении вестерн-блоттинга и иммуногистохимического анализа раковых тканей и раковых ксенотрансплантатов с использованием Mab против STEAP-1 наблюдается сильное экстенсивное окрашивание в тканях карциномы при значительно более низком уровне или отсутствии окрашивания в нормальных тканях. Детектирование STEAP-1 при карциноме и при метастазирующем заболевании демонстрирует, что Mab можно использовать в качестве диагностического и/или прогностического маркера. Следовательно, антитела против STEAP-1 можно использовать в диагностических применениях, таких как иммуногистохимия биопсийных образцов почек для выявления злокачественного заболевания у предполагаемых пациентов. С помощью метода проточной цитометрии определяют, что Mab против STEAP-1 специфически связываются с клетками карциномы. Таким образом, антитела против STEAP-1 можно использовать для диагностики всего организма с помощью методов визуализации, таких как радиоиммуносцинтиграфия и радиоиммунотерапия (например, методы детектирования локализованных и метастазирующих злокачественных заболеваний, сопровождающихся экспрессией STEAP-1, описаны в Potamianos S., et. al. Anticancer Res, 20(2A): 925-948 (2000)). Сбрасывание или высвобождение внеклеточного домена STEAP-1 в межклеточную среду, например, как в случае щелочной фосфодиэстеразы В10 (Meerson, R., Hepatology, 27: 563-568 (1998)), позволяет осуществлять диагностическое детектирование STEAP-1 с помощью антител против STEAP-1 в сыворотке и/или образцах мочи предполагаемых пациентов. Антитела против STEAP-1, специфически связывающиеся со STEAP-1, используются в терапевтических применениях для лечения злокачественных заболеваний, сопровождающихся экспрессией STEAP-1. Антитела против STEAP-1 можно использовать в виде неконъюгированных молекул или в составе конъюгатов, которые содержат антитела, присоединенные к одному из разных терапевтических или визуализирующих средств, хорошо известных в данной области, таких как пролекарства, ферменты или радиоактивные изотопы. В предклинических исследованиях неконъюгированные и конъюгированные антитела против STEAP-1 анализируют на эффективность предотвращения образования и ингибирования роста опухоли у мышиных моделей SCID, несущих раковые ксенотрансплантаты, например модели рака почек AGS-K3 и AGS-K6 (см. пример, озаглавленный "Ингибирование опухолей мочевого пузыря и легких in vivo, опосредованное моноклональными антителами против STEAP-1"). Конъюгированные и неконъюгированные антитела против STEAP-1 используют в качестве терапевтических средств в клинических испытаниях в применении к человеку либо отдельно, либо в сочетании с другими способами лечения, как описано в нижеследующих примерах. Пример 25. Клинические испытания человеческих антител против STEAP-1 in vivo для лечения и диагностики карциномы человека. Антитела согласно изобретению, распознающие эпитоп STEAP-1, можно использовать для лечения некоторых опухолей, таких как перечисленные в табл. I. На основе ряда факторов, включающих в себя уровень экспрессии STEAP-1, такие опухоли, как перечисленные в табл. I, в настоящее время являются предпочтительными показаниями для применения указанных антител. В сучае каждого из данных показаний успешно применяются три клинических подхода. I) Дополнительная терапия. При дополнительной терапии пациентов лечат антителами против STEAP-1 в сочетании с химиотерапевтическим или противоопухолевым средством и/или лучевой терапией. Первичные раковые мишени, такие как перечисленные в табл. I, лечат по стандартным схемам путем добавления антител к стандартным первой и второй линиям терапии. Схемы лечения составляют так, чтобы достичь эффективности, оцениваемой по уменьшению массы опухоли, а также возможности уменьшить обычные дозы стандартной химиотерапии. Уменьшение доз позволяет проводить дополнительную и/или пролонгированную терапию в результате уменьшения связанной с дозой токсичностью химиотерапевтического средства. Антитела против STEAP-1 участвуют в клинических испытаниях нескольких дополнительных способов лечения в сочетании с химиотерапевтическими или противоопухолевыми средствами адриамицином (при запущенном раке простаты), цисплатином (при запущенном раке головы, шеи и легкого), таксолом (при раке молочной железы) и доксорубицином (при предклиническом состоянии). II) Монотерапия. При применении антител против STEAP-1 в монотерапии опухолей антитела вводят пациентам без химиотерапевтического или противоопухолевого средства. В одном воплощении монотерапию проводят в клинике в применении к пациентам, страдающим от экстенсивного метастазирующего заболевания на последней стадии. У пациентов наблюдается некоторая стабилизация заболевания. Испытания показывают, что данная терапия имеет эффект у пациентов с раковыми опухолями, не поддающимися лечению. III) Визуализирующее средство. После присоединения радионуклида (например, йода или иттрия ( 131 I, 90 Y) к антителам против STEAP-1 меченные радиоактивными изотопами антитела используют в качестве диагностического и/или визуализирующего средства. Меченые антитела, выполняющие такую роль, локализуются как на солидных опухолях, так и в участках метастазирующего поражения, содержащих клетки, экспрессирующие STEAP-1. При применении антител против STEAP-1 в качестве визуализирующих агентов данные антитела используют как вспомогательное средство при хирургическом лечении солидных опухолей как при дооперационном скрининге, так и при послеоперационном наблюдении с целью выявления оставшейся и/или рецидивирующей опухоли. В одном воплощении меченое ( 111 In) антитело против STEAP-1 используют в качестве визуализирующего средства в фазе I человеческих клинических испытаний у пациентов, страдающих от карциномы, сопровождающейся экспрессией STEAP-1 (по аналогии см., например, Divgi et al. J. Natl. Cancer Inst. 83: 97-104 (1991)). Пациентов наблюдают с помощью стандартной предшествующей и последующей гамма-камеры. Полученные результаты позволяют обнаружить первичные и метастазирующие поражения. Доза и способ введения. Как известно рядовому специалисту в данной области, дозировочный режим можно установить путем сравнения с аналогичными продуктами, применяющимися в клинике. Так, антитела против STEAP-1 можно вводить в дозах, варьирующих от 5 до 400 мг/м 2 , причем более низкие дозы используются, например, по соображениям безопасности. Сродство антитела против STEAP-1 по сравнению со сродством известного антитела к его мишени является параметром, используемым специалистами в данной области для определения аналогичных дозировочных режимов. Кроме того, полностью человеческие антитела против STEAP-1 выводятся более медленно, чем химерные антитела; соответственно, вводимая пациентам доза таких полностью человеческих антител против STEAP-1 может быть ниже, возможно, в интервале от 50 до 300 мг/м 2 , оставаясь при этом эффективной. Измерение дозы в мг/м 2 , в противоположность традиционному измерению дозы в мг/кг, основано на площади поверхности и является удобной мерой дозирования, которая разработана для учета пациентов всех размеров от детей до взрослых. Для доставки антител против STEAP-1 используют три разных способа доставки. Традиционная внутривенная доставка является стандартным способом доставки при многих опухолях. Однако в случае опухолей брюшной полости, таких как опухоли яичников, желчных протоков, других протоков и т.п., внутрибрюшинное введение может иметь преимущество, поскольку оно позволяет достичь высокой дозы антитела в районе опухоли, а также минимизировать выведение антитела. Подобным образом, некоторые солидные опухоли имеют сосудистую сеть, позволяющую осуществлять регионарную перфузию. Регионарная перфузия обеспечивает высокую концентрацию антитела в участке опухоли и минимизирует краткосрочное выведение антитела. План клинических испытаний (CDP). Общее представление. CDP помогает осуществить и разработать применение антител против STEAP-1 во всомогательной терапии, монотерапии и в качестве визуализирующего средства. Вначале проводят испытания, демонстрирующие безопасность, затем испытания, подтверждающие эффективность при многократном введении дозы. Для сравнения стандартной химиотерапии с сочетанием стандартной терапии и антител против STEAP-1 используют открытые исследования. Очевидно, что в качестве критерия, используемого при наборе пациентов, можно использовать уровни экспрессии STEAP-1 в опухолях пациентов, определяемые с помощью биопсии. При проведении терапии, основанной на инфузии любого белка или антитела, факторы опасности в первую очередь связаны с (i) синдромом высвобождения цитокинов, т.е. с гипертонией, лихорадкой, дрожью, ознобом; (ii) развитием иммунного ответа на вещество (т.е. образование у пациента человеческих антител против терапевтических антител, или ответ НАНА) и (iii) токсичностью для нормальных клеток, экспрессирующих STEAP-1. Для отслеживания каждого из указанных факторов опасности используют стандартные анализы и наблюдения. Обнаружено, что антитела против STEAP-1 являются безопасными для людей. Пример 26. Клинические испытания в применении к людям. Монотерапия человеческими антителами против STEAP-1. Вышеописанное испытание, связанное с дополнительной терапией, показывает, что антитела против STEAP-1 являются безопасными, а во II фазе клинических испытаний подтвержают эффективность монотерапии и устанавливают оптимальный дозировочный режим. Такое испытание проводят, используя такие же факторы опасности и итоговые анализы, как и в испытании, связанном с дополнительной терапией, за исключением того, что пациенты не получают химиотерапевтического средства одновременно с антителами против STEAP-1. Пример 27. Клинические испытания в применении к людям. Диагностическая визуализация антитела против STEAP-1. Опять же, поскольку в пределах описанных выше факторов опасности дополнительная терапия является безопасной, проводят клинические испытания, касающиеся применения антител против STEAP-1 в качестве диагностического визуализирующего средства. Для проведения испытания используют схемы, подобные описанным в данной области, например в Divgi et al. J. Natl. Cancer Inst. 83: 97-104 (1991). Обнаружено, что антитела можно безопасно и эффективно использовать в диагностических целях. Пример 28. Клинические испытания в применении к людям. Дополнительная терапия человеческими антителами против STEAP-1 в сочетании с химиотерапией, лучевой терапией и/или антигормональной терапией. Фазу I клинических испытаний предпринимают для определения безопасности шести внутривенных доз человеческого антитела против STEAP-1, используемого для лечения солидной опухоли, например рака тканей, перечисленных в табл. I. В данном исследовании оценивают безопасность однократных доз антител против STEAP-1, используемых в дополнение к введению противоопухолевого, или химиотерапевтического, или антигормонального средства, описанного в данном документе, включающего в себя, без ограничения, цисплатин, топотекан, доксорубицин, адриамицин, таксол, лупрон, Zoladex, Eulexin, Casodex, Anandron и т.п. Данное испытание включает в себя доставку шести однократных доз антитела против STEAP-1 с увеличением дозы антитела приблизительно от 25 до 275 мг/м 2 в течение курса лечения, проводимого по следующей схеме: Пациентов тщательно наблюдают в течение недели после каждого введения антитела и химиотерапии. В частности, у пациентов оценивают указанные выше факторы опасности: (i) синдром высвобождения цитокинов, т.е. гипертония, лихорадка, дрожь, озноб; (ii) развитие иммунного ответа на вещество (т.е. образование у пациента человеческих антител против терапевтических антител, или ответ НАНА) и (iii) токсичность для нормальных клеток, экспрессирующих STEAP-1. Для отслеживания каждого из указанных факторов опасности используют стандартные анализы и наблюдения. У пациентов также оценивают клинический результат, в особенности уменьшение массы опухоли, определяемое с помощью MRI или другого метода визуализации. Антитела против STEAP-1, которые по данным предыдущих испытаний являются безопасными и эффективными, используют во II фазе испытаний, где подтверждают их эффективность и устанавливают оптимальный дозировочный режим. Пример 29. Идентификация и подтверждение потенциальных путей передачи сигнала. Описано, что многие белки млекопитающих взаимодействуют с сигнальными молекулами и участвуют в регуляции сигнальных путей (J. Neurochem. 2001, 76: 217-223). В частности, фибронектин связан с сигнальным каскадом МАРК, регулирующим клеточный митогенез (Jiang F., Jia Y., Cohen I. Blood. 2002, 99:3579). В добавление к вышесказанному, белок STEAP-1 содержит несколько сайтов фосфорилирования (см. табл. XXI), указывающих на его связь с определенными сигнальными каскадами. С помощью методов иммунопреципитации и вестерн-блоттинга идентифицируют белки, связанные со STEAP-1 и опосредующие события передачи сигнала. STEAP-1 может регулировать несколько путей, которые, как известно, участвуют в биологии злокачественного заболевания и включают в себя фосфолипидные пути, такие как P13K, AKT и др., пути адгезии и миграции, в том числе FAK, Rho, Rac-1, катенин и др., а также каскады, связанные с митогенезом/выживанием, такие как ERK, р38 и др. (Cell Growth Differ. 2000, 11: 279; J Biol Chem. 1999, 274: 801; Oncogene. 2000, 19: 3003; J. Cell Biol. 1997, 138: 913). Чтобы определить, является ли экспрессия STEAP-1 достаточной для регуляции конкретных сигнальных путей, но не для проявления другой активности, в покоящихся клетках РС3, исследуют влияние данных генов на активацию каскада р38 МАРК в клеточной линии рака простаты РС3. Активация киназы р38 зависит от ее фосфорилирования по остаткам тирозина и серина. Фосфорилированное состояние р38 можно отличить от нефосфорилированного с помощью Mab против фосфо-р38. Данные фосфо-специфические Ab используют для исследования состояния фосфорилирования р38 в рекомбинантных клеточных линиях РС3. Клетки РСЗ трансфицируют геном устойчивости к неомицину отдельно или в сочетании с геном STEAP-1 в векторе pSR α. Клетки выращивают в течение ночи в 0,5% FBS, затем стимулируют 10% FBS в течение 5 мин в присутствии или в отсутствие 10 мкг/мг ингибитора MEK PD98058. Клеточные лизаты анализируют методами 12,5% SDS-PAGE и вестерн-блоттинга с использованием антител против фосфо-ERK (Cell Signaling) и антител против ERK (Zymed). Клетки NIH-3T3 трансфицируют геном устойчивости к неомицину отдельно или в сочетании с геном STEAP-1 в векторе pSR α. Клетки обрабатывают по описанному выше способу, но не добавляют ингибитор MEK. Кроме того, клетки NIH-3T3-neo обрабатывают 10 мг/мл салицилата Na. Экспрессия STEAP-1 индуцирует фосфорилирование ERK-1 и ERK-2 в сыворотке и ингибируется вышестоящим ингибитором киназы MEK PD98058. В другой серии экспериментов исследуют достаточность экспрессии STEAP-1 в клеточной линии рака простаты РС3 для активации митогенного пути MAPK, а именно каскада ERK. Активация ERK зависит от фосфорилирования тирозиновых и сериновых остатков. Фосфорилированное состояние ERK можно отличить от нефосфорилированного с помощью Mab против фосфо-ERK. Данные фосфоспецифические Ab используют для исследования состояния фосфорилирования ERK в рекомбинантных клеточных линиях РС3. Клетки РС3, экспрессирующие активированную форму Ras, используют в качестве положительного контроля. Полученные результаты показывают, что в то время как экспрессия контрольного гена neo не оказывает влияния на фосфорилирование ERK, экспрессия STEAP-1 в клетках РС3 является достаточной для индуцирования увеличения фосфорилирования ERK (фиг. 28). Данные результаты подтверждают методом вестерн-блоттинга с использованием антител против ERK и по активации пути ERK под действием STEAP-1. Поскольку FBS содержит несколько компонентов, которые могут вносить вклад в опосредованную рецептором активацию ERK, авторы исследуют влияние STEAP-1 в условиях низкого и оптимального уровней FBS. Клетки РС3, экспрессирующие neo или STEAP-1, выращивают в течение ночи в 0,1% или 10% FBS. Клетки анализируют методом вестерн-блоттинга с использованием антител против фосфо-ERK. Данный эксперимент показывает, что STEAP-1 индуцирует фосфорилирование ERK в 0,1% FBS и подтверждает, что экспрессия STEAP-1 является достаточной для индуцирования активации сигнального каскада ERK в отсутствие других стимулов. Чтобы подтвердить, что STEAP-1 непосредственно или косвенно активирует известные пути передачи сигнала в клетках, проводят анализы транскрипционного репортера на основе люциферазы (luc) в клетках, экспрессирующих отдельные гены. Данные транскрипционные репортеры содержат консенсусные участки связывания известных факторов транскрипции, которые лежат ниже хорошо охарактеризованных путей передачи сигнала. Репортеры и примеры ассоциированных факторов транскрипции, путей передачи сигнала и активирующих стимулов приведены ниже. 1. NFkB-luc, NFkB/Rel; Ik-kinase/SAPK; рост/апоптоз/стресс. 2. SRE-luc, SRF/TCF/ELK1; MAPK/SAPK; рост/дифференциация. 3. AP-1-luc, FOS/JUN; MAPK/SAPK/PKC; рост/апоптоз/стресс. 4. ARE-luc, рецептор андрогенов; стероиды/MAPK; рост/дифференциация/апоптоз. 5. p53-luc, p53; SAPK; рост/дифференциация/апоптоз. 6. CRE-luc, CREB/ATF2; PKA/p38; рост/апоптоз/стресс. 7. TCF-luc, TCF/Lef; катенин, адгезия/инвазия. Опосредованные геном эффекты можно анализировать на клетках, экспрессирующих мРНК. Плазмиды, содержащие люциферазный репортер, можно вводить путем липид-опосредованной трансфекции (TFX-50, Promega). Активность люциферазы как показатель относительной транскрипционной активности определяют путем инкубации клеточных экстрактов с субстратом люциферина и измерения люминесценции реакционной смеси с помощью люминометра. Сигнальные пути, активируемые STEAP-1, картируют и используют для идентификации и оценки терапевтических мишеней. Поскольку STEAP-1 участвует в передаче клеточного сигнала, его можно использовать в качестве мишени для диагностических, прогностических, профилактических и/или терапевтических целей. Пример 30. Участие STEAP-1 в транспорте маленьких молекул и межклеточных взаимодействиях. При образовании и развитии опухоли происходит нарушение регуляции межклеточных взаимодействий, которые нужны для поддержания целостности органа и гомеостаза. Межклеточные взаимодействия можно анализировать с помощью двух типов методов (J. Biol. Chem. 2000, 275: 25207). В первом анализе клетки, нагруженные флуоресцентным красителем, инкубируют в присутствии немеченых клеток-реципиентов, после чего клеточные популяции анализируют методом флуоресцентной микроскопии. Данный качественный анализ позволяет определить изменение распределения красителя между соседними клетками. Во второй аналитической системе донорные и реципиентные клеточные популяции обрабатывают, как описано выше, после чего проводят количественное измерение популяций реципиентных клеток методом FACS. С помощью двух указанных аналитических систем клетки, экспрессирующие STEAP-1, сравнивают с контрольными клетками, которые не экспрессируют STEAP-1, и в результате обнаруживают, что STEAP-1 усиливает межклеточные взаимодействия. На фиг. 29 показано, что STEAP-1 опосредует перенос маленьких молекул между соседними клетками и таким образом регулирует межклеточные взаимодействия в клетках рака простаты. В данном эксперименте реципиентные клетки РС3 метят конъюгатом декстран-техасовый красный в концентрации 1 мг/мл, а донорные клетки РС3 метят кальцеином AM (зеленый). Донорные (зеленые) и реципиентные (красные) клетки совместно культивируют при 37 °С, после чего с помощью микроскопии анализируют расположение техасового красного и кальцеина. Полученные результаты показывают, что, в то время как в контрольных клетках РС3 (экспрессия белка STEAP-1 не обнаруживается) перенос кальциина происходит на низком уровне, экспрессия STEAP-1 обеспечивает перенос маленьких молекул между клетками, и в результате совместной локализации красных и зеленых молекул изначально красные реципиентные клетки приобретают коричневатый цвет. Маленькие молекулы и/или антитела, которые модулируют межклеточные взаимодействия, опосредуемые STEAP-1, можно использовать для лечения злокачественных заболеваний, сопровождающихся экспрессией STEAP-1. На фиг. 30 показано, что для переноса маленьких молекул требуется, чтобы экспресссия STEAP-1 происходила как в донорных, так и в реципиентных популяциях. В данном эксперименте клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 в векторе pSR α. Реципиентные клетки метят конъюгатом декстран-техасовый красный в концентрации 1 мг/мл, а донорные клетки метят кальцеином AM в концентрации 2,5 мкг/мл. Донорные (зеленые) и реципиентные (красные) клетки совместно культивируют при 37 °С в течение 18-24 ч, после чего с помощью микроскопии анализируют расположение флуоресцентных красителей. Верхние секторы: оптическая микроскопия. Нижние секторы: УФ-флуоресценция. Левые секторы: в качестве доноров и реципиентов используют клетки РС3-neo. Центральные секторы: в качестве донорных клеток используют РС3-neo, а в качестве реципиентных клеток используют PC3-STEAP-1. Правые панели: в качестве доноров и реципиентов используют клетки PC3-STEAP-1. Межклеточные взаимодействия детектируют только в том случае, если STEAP-1 экспрессируется и на донорных, и на реципиентных клетках. Полученные результаты показывают, что при совместном культивировании контрольных клеток РС3 и клеток РС3 перенос кальцеина не наблюдается. Подобным образом, при совместной инкубации контрольных клеток РС3 и клеток PC3-STEAP-1 перенос кальцеина не наблюдается. Однако при совместном культивировании донорных клеток PC3-STEAP-1 и реципиентных клеток PC3-STEAP-1 происходит перенос маленьких молекул, определяемый по совместной локализации зеленого и красного красителей в одних и тех же клетках. Взятые вместе результаты, приведенные на фиг. 29 и 30, демонстрируют, что STEAP-1 опосредует перенос маленьких молекул и регулирует межклеточные взаимодействия путем образования каналов межклеточного взаимодействия, которые функционально подобны щелевидным контактам. Кроме того, подтверждают влияние М2/120.545 STEAP-1 на щелевой контакт (см. фиг. 31). В данном эксперименте клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 в векторе pSR α. Реципиентные клетки метят конъюгатом декстран-техасовый красный в концентрации 1 мг/мл, а донорные клетки метят кальцеином AM в концентрации 2,5 мкг/мл. Донорные (зеленые) и реципиентные (красные) клетки совместно культивируют при 37 °С в течение 18-24 ч, после чего с помощью микроскопии анализируют расположение флуоресцентных красителей. Во всех экспериментах в качестве доноров и реципиентов используют одинаковые клетки. Клетки инкубируют с указанными количествами Mab STEAP-1/120.545 в течение 10 мин перед внесением в планшеты, затем Mab оставляют в культуре на 24 ч до анализа. STEAP-1/120.545 уменьшает щелевой контакт, опосредованный STEAP-1, в зависимости от используемой дозы. Полученные результаты показывают, что STEAP-1/120.545 уменьшает щелевой контакт, опосредованный STEAP-1, в зависимости от используемой дозы. Таким образом, поскольку STEAP-1 участвует в межклеточных взаимодействиях и транспорте маленьких молекул, его можно использовать в качестве мишени для диагностических, прогностических, профилактических и/или терапевтических целей. Пример 31. Интерференция РНК (РНКи). Технологию интерференции РНК (РНКи) используют в ряде клеточных анализов, применяющихся в онкологии. РНКи представляет собой посттранскрипционный механизм "молчания" гена, активируемый двухцепочечными РНК (дцРНК). РНКи индуцирует деградацию специфической мРНК, приводящую к изменению экспрессии белка и, следовательно, функции гена. В клетках млекопитающих указанные дцРНК, называемые короткие интерферирующие РНК (киРНК), имеют правильный состав для активации пути РНКи, направленного на деградацию, особенно некоторых мРНК (см., Elbashir S.M., et. al., Duplexes of 21-nucleotide RNAs Mediate RNA interference in Cultured Mammalian Cells, Nature, 411(6836): 494-8 (2001)). Таким образом, технология РНКи успешно используется в клетках млекопитающих для выключения целевых генов. Утрата контроля за пролиферацией является отличительным признаком раковых клеток; следовательно, целесообразно определить значение STEAP-1 в процессах выживания/пролиферации клеток. Соответственно, для исследования функции антигена STEAP-1 используют РНКи. Чтобы получить киРНК для STEAP-1, используют алгоритмы, позволяющие предсказать структуру олигонуклеотидов, которые обладают критическими молекулярными параметрами (содержание G:C, температура плавления и др.) и обладают способностью значительно уменьшать уровни экспрессии белка STEAP-1 при введении в клетки. Соответственно, одной из целевых последовательностей киРНК STEAP-1 является: 5'-AAGCTCATTCTAGCGGGAAAT-3' (SEQ ID NO: 81). В соответствии с данным примером используют композиции киРНК STEAP-1, которые содержат киРНК (двухцепочечную короткую интерферирующую РНК), соответствующую нуклеотидной последовательности ORF белка STEAP-1 или ее субпоследовательностям. Так, используемые в данном способе киРНК, как правило, содержат 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 или более смежных нуклеотидов РНК в длину. Данные последовательности киРНК являются комплементарными и некомплементарными по меньшей мере части кодирующей последовательности мРНК. В предпочтительном воплощении субпоследовательности содержат 19-25 нуклеотидов в длину, наиболее предпочтительно 21-23 нуклеотидов в длину. В предпочтительных воплощениях указанные киРНК вызывают снижение уровня антигена STEAP-1 в клетках, экспрессирующих данный белок, и имеют функциональные эффекты, как описано ниже. Выбранные киРНК (олиго STEAP-1.b) анализируют на жизнеспособность/пролиферацию ряда клеточных линий методом MTS (позволяющим измерять метаболическую активность клеток). Колориметрические анализы с применением тетразолия (т.е. MTS) позволяют точно детектировать жизнеспособные клетки, поскольку живые клетки являются метаболически активными и, следовательно, в отличие от мертвых клеток могут восстанавливать соли тетразолия до окрашенных соединений формазана. Более того, указанный олиго STEAP-1.b вызывает снижение уровня антигена STEAP-1 в клетках, экспрессирующих данный белок, и имеет функциональные эффекты, как описано ниже с использованием нижеследующих способов. Трансфекция киРНК млекопитающих. За 1 день до трансфекции киРНК разные клеточные линии помещают в среду (RPMI 1640, содержащая 10% FBS в отсутствие антибиотиков) в количестве 2 ×10 3 клеток/лунку в объеме 80 мкл (в формате 96-луночного планшета), чтобы провести анализ жизнеспособности/MTS. В каждом эксперименте параллельно со специфическими олиго-киРНК STEAP-1 используют следующие контроли: а) клетки Mock, трансфицированные липофектамином 2000 (Invitrogen, Carlsbad, CA) и буфером для отжига (не содержащим киРНК); b) киРНК, специфичные для люциферазы-4 (целевая последовательность: 5'-AAGGGACGAAGACGAACACUUCTT-3') (SEQ ID NO: 82); и с) Eg5-специфические киРНК (целевая последовательность: 5'-AACTGAAGACCTGAAGACAATAA-3') (SEQ ID NO: 83). Конечная концентрация киРНК составляет 10 нМ, а липофектамина 2000 - 1 мкг/мл. Анализ проводят следующим способом. Вначале киРНК разбавляют OPTIMEM (не содержащая сыворотки среда для трансфекции, Invitrogen) до концентрации 0,1 мкМ (в 10 раз выше, чем концентрация в реакционной смеси) и инкубируют 5-10 мин при комнатной температуре (к.т.). Липофектамин 2000 разбавляют до концентрации 10 мкг/мл (в 10 раз выше, чем концентрация в реакционной смеси) для общего числа трансфекции и инкубируют 5-10 мин при к.т. Подходящее количество разбавленного раствора Липофектамин 2000 с 10-кратным превышением концентрации смешивают в соотношении 1:1 с разбавленным раствором киРНК с 10-кратным превышением концентрации и инкубируют при к.т. в течение 20-30" (раствор для трансфекции с 5-кратным превышением концентрации). Перед анализом 20 мкл раствора для трансфекции с 5-кратным превышением концентрации добавляют к соответствующим образцам и инкубируют при 37 °С в течение 96 ч. Анализ MTS. Анализ MTS представляет собой колориметрический метод определения числа жизнеспособных клеток в анализах пролиферации, цитотоксичности или чувствительности к химиотерапевтическим препаратам с применением соединения тетразолия [внутренняя соль 3-(4,5-диметилтиазол-2-ил)-5-(3-карбоксиметоксифенил)-2-(4-сульфофенил)-2Н-тетразолия; MTS (b)] и электрон-присоединяющего реагента (этосульфат феназина, PES). Анализы проводят путем добавления небольшого количества раствора реагента непосредственно в культуральные лунки, инкубации в течение 1-4 ч и регистрации поглощения при 490 нм с помощью планшет-ридера для 96-луночных планшет. Количество окрашенного формазанового продукта, определяемое по величине поглощения при 490 нм, прямо пропорционально митохондриальной активности и/или числу живых клеток в культуре. Чтобы выяснить функцию STEAP-1 в клетках, STEAP-1 "выключают" путем трансфекции клеточных линий, эндогенно экспрессирующих STEAP-1. Как изображено на фиг. 32, фосфорилирование и ERK-1, и ERK-2 индуцируется 10% сывороткой и ингибируется Mab M2/92.30 и киРНК STEAP-1. В данном эксперименте клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 и Mab в векторе pSR α. Для удаления РНКи клетки PC3-STEAP-1 стабильно трансфицируют вектором pPUR-U6-27-STEAP-1, содержащим киРНК STEAP-1. Клетки держат в условиях недостаточного количества питательных веществ в 0,1% FBS в течение 18 ч при 37 °С, затем помещают на лед, где держат в течение 10 мин в присутствии или в отсутствие 10 мкг/мл Mab M2/92.30, после чего в течение 3 мин нагревают до к.т. и стимулируют 10% FBS в течение 5 мин. Клетки лизируют в буфере RIPA, лизаты целых клеток анализируют методом 12,5% SDS-PAGE, а белки детектируют методом вестерн-блоттинга. Фосфо-ERK детектируют с помощью кроличьей антисыворотки (Cell Signaling), a ERK детектируют с помощью кроличьих антител против ERK (Zymed). STEAP-1 детектируют с помощью овечьих антител против STEAP-1, а актин детектируют с помощью Mab против актина (Santa Cruz). Кроме того, как изображенно на фиг. 33, специфические РНКи STEAP-1, стабильно экспрессирующиеся в клетках PC3-STEAP-1, уменьшают индуцированные STEAP-1 межклеточные взаимодействия. В данном эксперименте клетки РС3 трансфицируют геном устойчивости к неомицину отдельно или вместе с геном STEAP-1 в векторе pSR α. Для удаления РНКи клетки PC3-STEAP-1 стабильно трансфицируют вектором pPUR-U6-27-STEAP-1, содержащим киРНК STEAP-1, или пустым вектором. Реципиентные клетки метят конъюгатом декстран-техасовый красный в концентрации 1 мг/мл, а донорные клетки метят кальцеином AM в концентрации 2,5 мкг/мл. Донорные (зеленые) и реципиентные (красные) клетки совместно культивируют при 37 °С в течение 18-24 ч, после чего с помощью микроскопии анализируют расположение флуоресцентных красителей. Во всех экспериментах в качестве доноров и реципиентов используют одинаковые клетки. Другое воплощение согласно изобретению относится к методу анализа пролиферации клеток, экспрессирующих STEAP-1, путем измерения синтеза ДНК как маркера пролиферации. Используют меченные (например, 3 Н-тимидином) предшественники ДНК и количественно определяют их включение в ДНК. Включение меченого предшественника в ДНК прямо пропорционально числу клеточных делений в культуре. Другим способом измерения клеточной пролиферации являются клоногенные анализы. В данных анализах определенное число клеток помещают на подходящую подложку и считают число колоний, образовавшихся через определенный период инкубации после обработки киРНК. При оценке раковой мишени STEAP-1 в дополнение к анализу жизнеспособности/пролиферации клеток используют анализы, связанные с апоптозом и клеточным циклом. Отличительным биохимическим признаком апоптотического процесса является фрагментация геномной ДНК, необратимое событие, приводящее к гибели клетки. Фрагментированную ДНК в клетках можно обнаружить путем иммунологического детектирования комплексов гистона с фрагментами ДНК с помощью иммунологического анализа (например, ELISA для детектирования клеточной гибели), который позволяет определять обогащение цитоплазмы апоптотических клеток комплексами гистона с фрагментами ДНК (моно- и олигонуклеосомами). Данный анализ не требует предварительного мечения клеток и позволяет детектировать деградацию ДНК в клетках, которые не пролиферируют in vitro (например, свежевыделенные опухолевые клетки). Наиболее важную роль в запуске апоптотической гибели клеток играют эффекторные молекулы, называемые каспазами. Каспазы представляют собой протеазы, которые при активации расщепляют многочисленные субстраты по аспартатному остатку карбоксиконцевого участка, опосредуя наиболее ранние стадии апоптоза. Все каспазы синтезируются в виде проферментов, активация которых включает в себя расщепление по аспартатным участкам. В частности, по-видимому, каспаза 3 играет ключевую роль в инициации клеточных событий апоптоза. Анализы активации каспазы 3 позволяют детектировать ранние события апоптоза. После обработки РНКи детектирование методом вестерн-блоттинга активной каспазы 3 или продуктов протеолитического расщепления (например, PARP) в апоптотических клетках дополнительно подтверждает индукцию апоптоза. Поскольку клеточные механизмы, приводящие к апоптозу, являются сложными, каждый из них имеет свои преимущества и ограничения. Другие критерии/ключевые моменты, такие как клеточная морфология, конденсация хроматина, протекание мембраны, апоптотические тельца, помогают дополнительно подтвердить клеточную гибель как апоптотическую. Поскольку не все генные мишени, которые регулируют клеточный рост, являются антиапоптотическими, получают профиль содержания ДНК или профиль клеточного цикла путем измерения содержания ДНК в клетках с нарушенной проницаемостью мембраны. Ядра апоптотических клеток содержат меньше ДНК вследствие ее утечки в цитоплазму (популяция суб-G1). Кроме того, окрашивание ДНК (например, йодидом пропидия) также позволяет различить разные фазы клеточного цикла в популяции клеток, поскольку в фазах G0/G1, S и G2/M присутствует разное количество ДНК. С помощью данных анализов можно количественно определить субпопуляции. Что касается гена STEAP-1, исследования РНКи облегчают понимание вклада генного продукта в раковые пути. Такие активные молекулы РНКи можно использовать в скрининге Mab для идентификации соединений, обладающих противоопухолевой активностью. Кроме того, киРНК можно вводить раковым пациентам в качестве терапевтических средств, уменьшающих злокачественный рост при разных типах рака, включающих в себя перечисленные в табл. I. Поскольку STEAP-1 участвует в таких процессах, как выживание, клеточная пролиферация, онкогенез или апоптоз, его можно использовать в качестве мишени для диагностических, прогностических, профилактических и/или терапевтических целей. Пример 32. Модулирование функции STEAP-1. Транспорт ионов играет важную роль в регуляции клеточного роста, внутриклеточной проницаемости, транспортировке молекул и передаче сигналов (Minke В. Cell Mol. Neurobiol. 2001, 21: 629; Golovina et al., Am. J. Physiol Heart Circ Physiol. 2001, 280: H746), данные функции особенно характерны для неопластического состояния. Межклеточные взаимодействия регулируют гомеостаз, клеточную пролиферацию и гибель клеток (Evans W.H., Martin P.E. Mol. Membr. Biol. 2002, 19: 121; Carruba G, et al., Ann, NY, Acad. Sci. 2002, 963: 156), данные функции также особенно характерны для неопластического состояния. Ингибиторы функции STEAP-1 идентифицируют, используя контрольные клеточные линии и клеточные линии, экспрессирующие STEAP-1. Например, клетки РС3 и PC3-STEAP-1 инкубируют в присутствии и в отсутствие Mab или маленьких ингибиторных молекул. Эффект данных Mab или маленьких ингибиторных молекул исследуют с использованием описанных выше анализов ионных потоков, межклеточных взаимодействий, пролиферации и сигнальных путей. Передачу сигнала и биологический результат, опосредованный переносчиками, можно модулировать посредством разных механизмов, включающих в себя ингибирование связывания рецептора и лиганда, применение ионных антагонистов, белковые взаимодействия, регуляцию транспорта ионов и маленьких молекул и др. (Tang W. et al., Front Biosci, 2002, 7: 1583). Модуляторы (ингибиторы или усилители) функции STEAP-1 идентифицируют, используя контрольные клеточные линии и клеточные линии, экспрессирующие STEAP-1. Например, клетки РС3 и PC3-STEAP-1 инкубируют в присутствии и отсутствие Mab или маленьких модуляторных молекул. В качестве модуляторов функций STEAP-1, раскрытых в данном документе, в контексте согласно изобретению можно использовать блокаторы ионных каналов, которые включают в себя такие соединения, как амлодипин, азулен, лигидропиридины, тианины, нифедин, верапамил и их производные (Tanaka Y., Shigenobu K. Cardiovasc Drug Rev. 2001, 19: 297; Djuric D., Mitrovic V., Jakovljevic V. Arzineimittelforschung. 2002, 52: 365; Kourie J.I., Wood H.B. Prog. Biophys. Mol. Biol. 2000; 73: 91); и ингибиторы межклеточных взаимодействий, которые включают в себя такие соединения, как бета-глицирретиновая кислота, ретиноиды, ТРА (Krutovskikh V.A. et al., Oncogene. 2002, 21: 1989; Rudkin et al., J. Surg. Res. 2002, 103: 183; Ruch J. et al., J. Cell Biochem. 2001, 83: 163). Соответственно, эффект(ы) Mab или маленьких ингибиторных молекул исследуют с использованием анализов ионных потоков, межклеточных взаимодействий, пролиферации и сигнальных путей, описанных в приведенных выше примерах. Если Mab и маленькие молекулы модулируют, например ингибируют транспорт и онкогенную функцию STEAP-1, их используют для профилактических, прогностических, диагностических и/или терапевтических целей. В данном изобретении приводятся ссылки на данные, содержащиеся на разных web-сайтах, а также на разные публикации, патентные заявки и патенты (Web-сайты указываются по унифицированному указателю ресурса, или URL, адресам глобальной сети). Раскрытие каждой из указанных ссылок настоящим включается в данное описание в качестве ссылки во всей полноте. Объем согласно изобретению не следует ограничивать раскрытыми в данном описании воплощениями, которые предлагаются только для иллюстрации отдельных аспектов согласно изобретению, причем все фунционально эквивалентные воплощения входят в объем согласно изобретению. Различные модификации моделей и методов согласно изобретению, в добавление к описанным в данном документе, которые станут очевидными для специалистов в данной области из вышеприведенных описания и разъяснений, также входят в объем согласно изобретению. Такие модификации или другие воплощения могут быть осуществлены без отступления от объема и сущности согласно изобретению. Таблица 1 Таблица II Таблица III(A) Матрица аминокислотных замен адаптирована с помощью программного обеспечения GCG 9.0 BLOSUM62 (матрица с заменой блоков). Чем выше значение, тем больше вероятность обнаружения замены в родственных, природных белках (см. web-сайт URL ikp.unibe.ch/manual/blosum62.html) Таблица III(B) Таблица IV(А) Жирным шрифтом выделены предочтительные остатки, курсивом выделены менее предочтительные остатки. Считается, что пептид несет мотив, если он содержит первичные якорные фрагменты в каждом якорном положении мотива или супермотива. Таблица IV(В) Таблица IV(С) Курсивом выделены менее предпочтительные или "допустимые" остатки. Таблица IV(D) Курсивом выделены менее предпочтительные или "допустимые" остатки. Таблица IV(Е) Таблица IV(F) Таблица IV(G) Мотивы указывают остатки, определяющие особенности супертипов. Мотивы содержат остатки, которые, как определено на основании опубликованных данных, распознаются несколькими аллелями в супертипе. Остатки, заключенные в скобки, представляют собой дополнительные остатки, которые по прогнозам являются допустимыми для нескольких аллелей в супертипе. Таблица IV(H) Таблица IV(I) Таблицы V-XVIII приведены в патентной заявке США № 10/236878; поданной 6 сентября 2002 г., конкретное содержание которой полностью включено в данное описание в качестве ссылки. Таблица XIX Таблица XX Таблица XXI Таблицы XXII-LI приведены в патентной заявке США № 10/236878; поданной 6 сентября 2002 г., конкретное содержание которой полностью включено в данное описание в качестве ссылки. Таблица LII Таблица LIII |