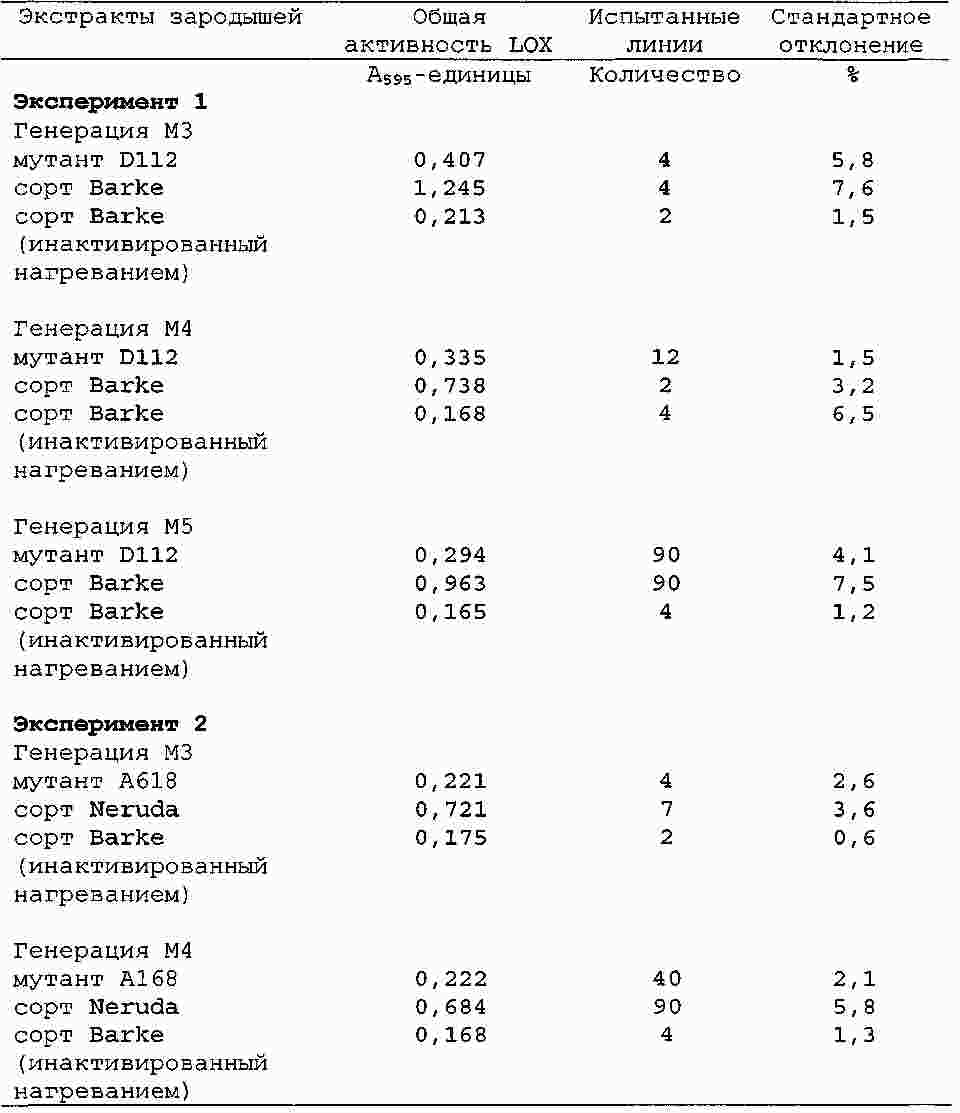
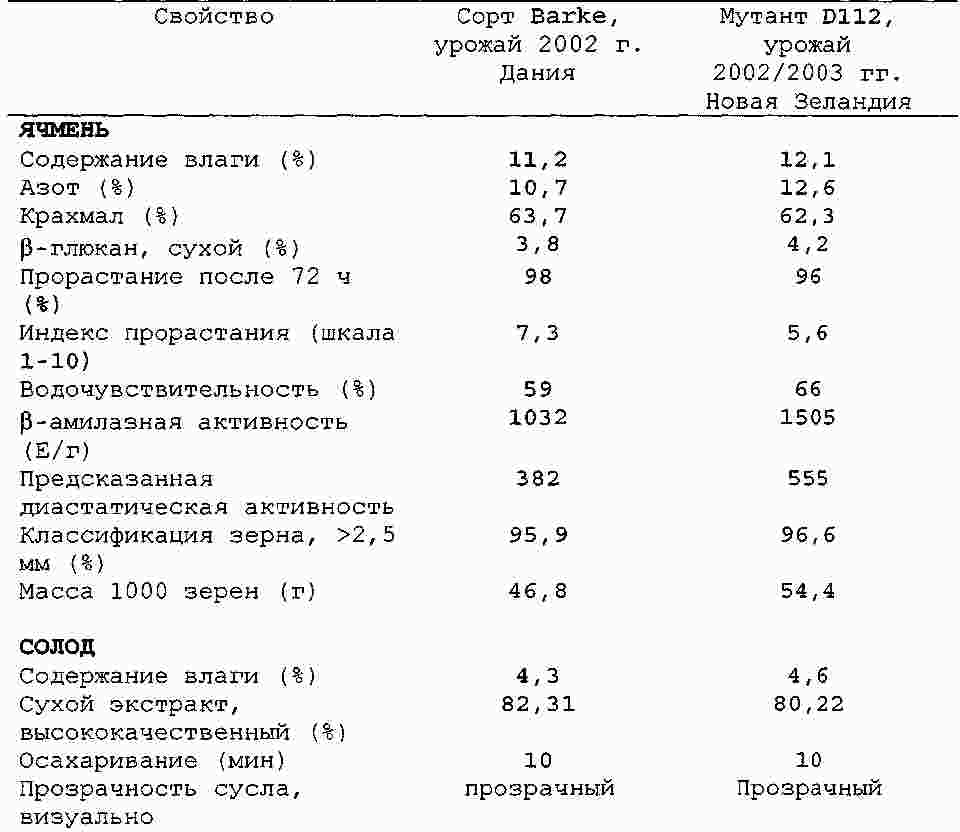
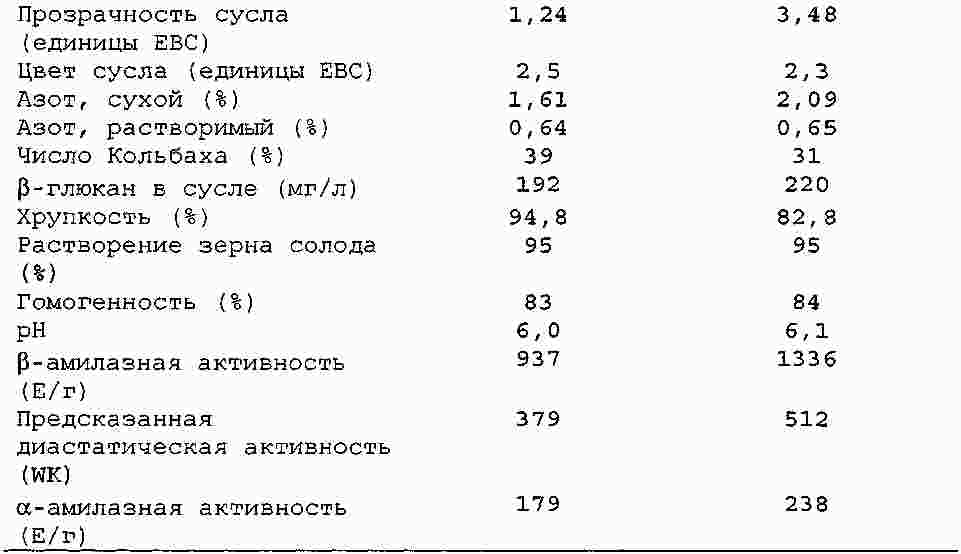
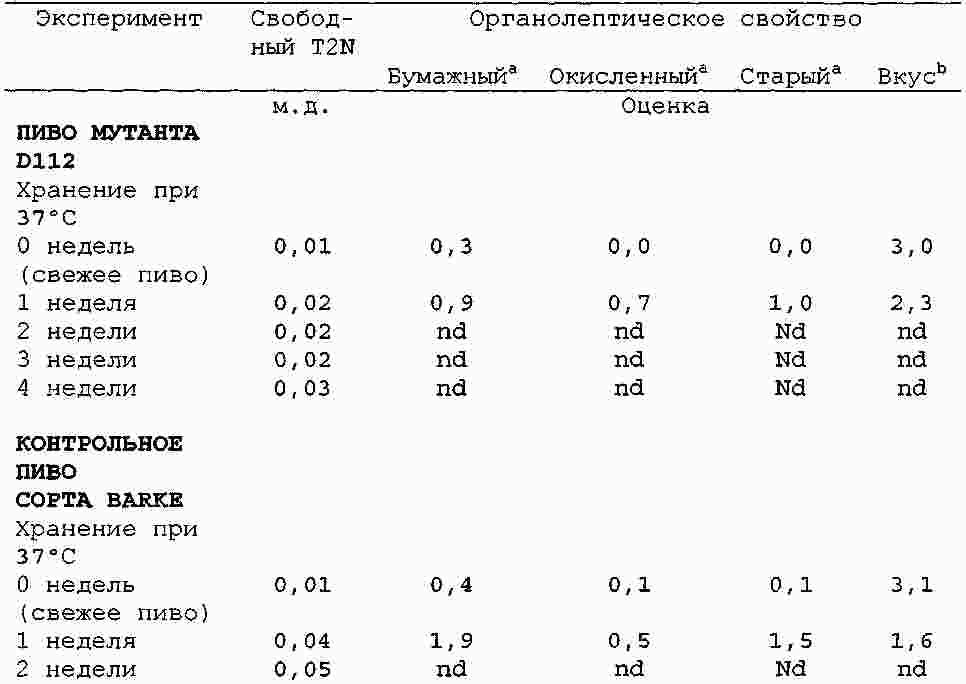
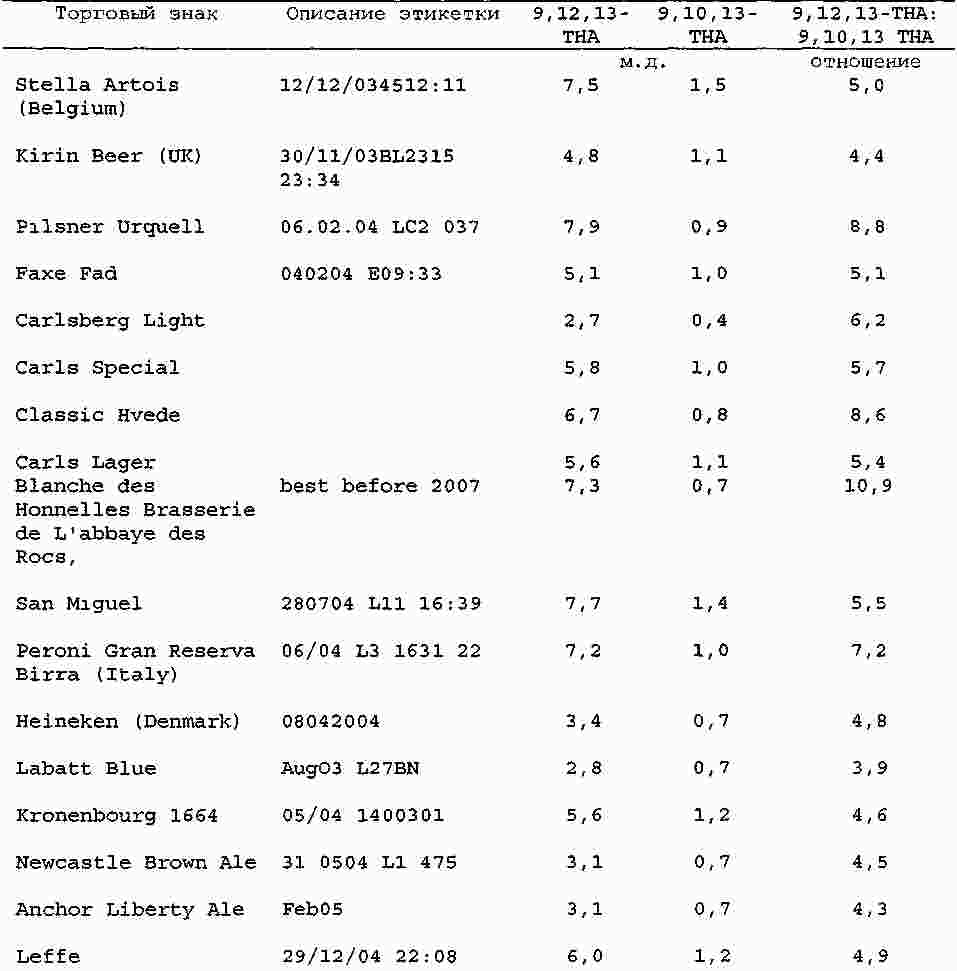
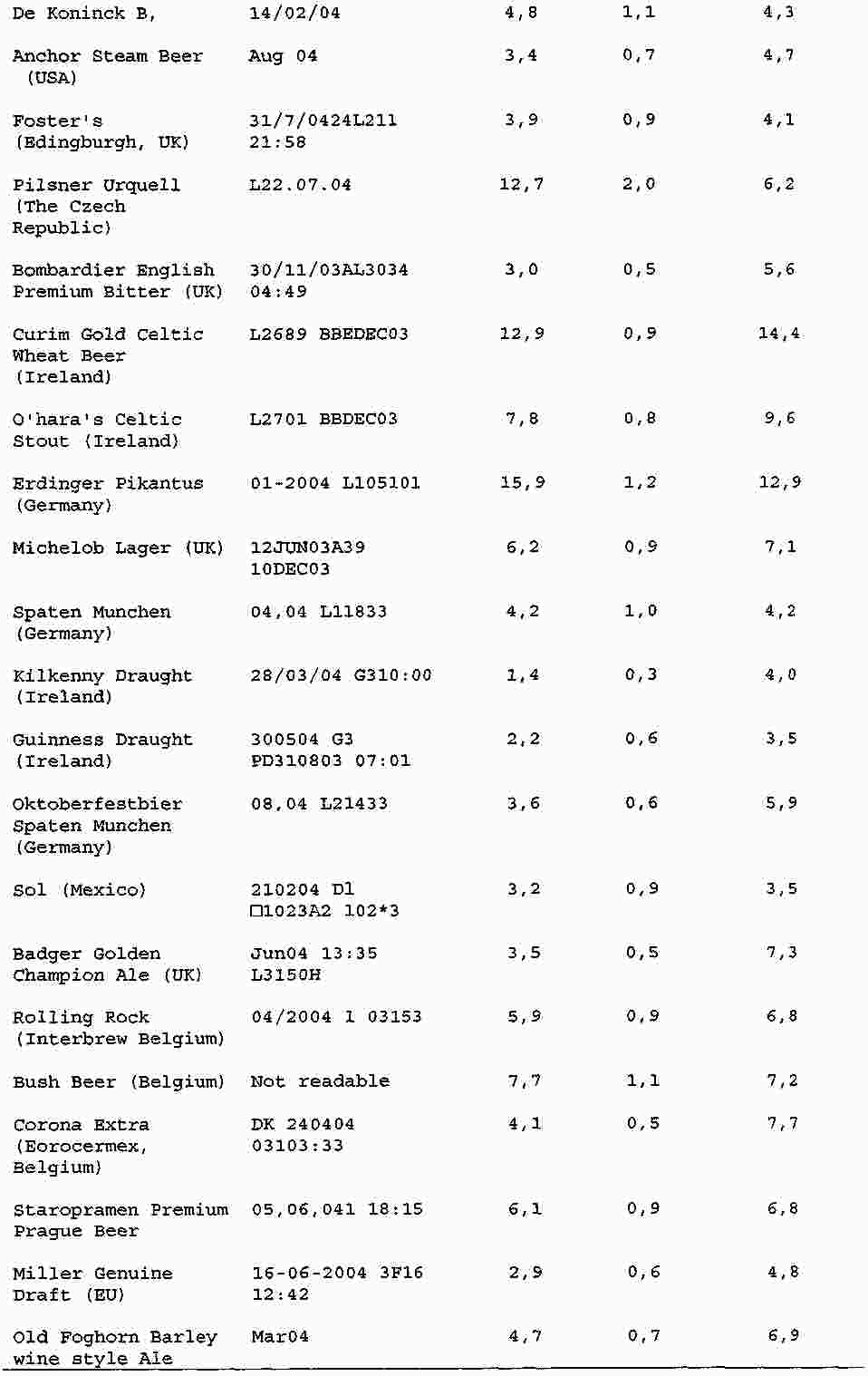
|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000014705b*\id |
|
Термины запроса в документе Реферат Согласно данному изобретению обеспечен нуль-LOX-1-ячмень и продукты растительного происхождения, полученные из него, такие как солод, приготовленный с использованием зерен ячменя, дефектных в синтезе превращающего жирные кислоты фермента липоксигеназы-1. Указанный фермент является ответственным за основную активность, связанную с превращением линолевой кислоты в 9-гидропероксиоктадекадиеновую кислоту, метаболит липоксигеназного пути, которая посредством дополнительных ферментативных и спонтанных реакций может приводить к появлению транс-2-ноненаля. Данное изобретение позволяет пивоварам производить пиво, лишенное детектируемых транс-2-ноненаль-специфических привкусов, даже после продолжительного хранения этого напитка. Формула [0001] Растение ячменя или часть указанного растения, содержащие мутантный белок липоксигеназы LOX-1, у которого отсутствует весь или по меньшей мере часть С-концевого домена, соответствующего аминокислотам 520-862 LOX-1 растения ячменя дикого типа (SEQ ID NO:3 или 7), где указанное растение проявляет нулевую LOX-1-активность. [0002] Растение ячменя или его часть по п.1, где указанной частью растения ячменя является зерно (зерна). [0003] Растение ячменя или его часть по п.1 или 2, где ген, кодирующий LOX-1, указанного растения содержит преждевременный нонсенс-кодон (бессмысленный, терминирующий трансляцию кодон). [0004] Растение ячменя или его часть по п.3, где ген, кодирующий LOX-1, указанного растения содержит нонсенс-кодон, соответствующий основаниям 3572-3574 SEQ ID NO:2. [0005] Растение ячменя или его часть по п.1 или 2, где ген, кодирующий LOX-1, указанного растения содержит по меньшей мере одну мутацию в сайте сплайсинга. [0006] Растение ячменя или его часть по п.5, где ген, кодирующий LOX-1, указанного растения содержит мутацию сайта сплайсинга, причем указанная мутация соответствует основанию 2311 SEQ ID NO:6. [0007] Растение ячменя или его часть по любому из пп.1-6, где указанное растение характеризуется: [0008] Растение ячменя или его часть по любому из пп.1-7, где указанная часть растения ячменя представляет собой регенерируемые клетки. [0009] Солодовая композиция, содержащая обработанное растение ячменя или его часть, где указанным растением ячменя является растение ячменя по любому из пп.1-8. [0010] Композиция сусла, полученная с использованием растения ячменя или его части по любому из пп.1-8 или с использованием солодовой композиции, приготовленной из указанного растения ячменя или его части, или их смеси. [0011] Композиция сусла по п.10, где указанная композиция получена с использованием одного или нескольких добавленных ферментов. [0012] Напиток, имеющий стабильные органолептические качества, где указанный напиток получен переработкой растения ячменя или его части по любому из пп.1-8. [0013] Напиток по п.12, где указанный напиток является пивом. [0014] Напиток по п.12, где указанный напиток приготовлен с использованием солода, приготовленного из зерен указанного растения ячменя. [0015] Напиток по любому из пп.12-14, где указанный напиток приготовлен из композиции сусла, приготовленного из растения ячменя или его части или из солодовой композиции, приготовленной из указанного растения ячменя или его части. [0016] Напиток по п.12, где указанный напиток приготовлен из не подвергнутых солодоращению (несоложеных) растений ячменя или их частей. [0017] Напиток по любому из пп.12-16, где указанный напиток является неферментированным напитком. [0018] Применение растения ячменя или его части по любому из пп.1-8 для получения пищевой композиции, кормовой композиции, ароматной композиции сырьевого материала или любой их комбинации. [0019] Пищевая композиция, кормовая композиция или ароматная композиция сырьевого материала, содержащая растение ячменя или его часть по любому из пп.1-8. [0020] Способ получения напитка, имеющего стабильные органолептические качества, где указанный способ предусматривает стадии: [0021] Способ по п.20, где стадия (i) предусматривает приготовление солодовой композиции из зерен указанного растения ячменя или его части. [0022] Способ по любому из пп.20 и 21, где этот способ дополнительно предусматривает инкубирование с ингибитором LOX. [0023] Способ по п.20, где переработка композиции в напитки предусматривает стадию затирания. [0024] Способ по п.20, где ингибитор LOX добавляют во время указанной стадии затирания. [0025] Способ получения солодовой композиции по п.9 с нулевой активностью LOX-1, где указанный способ предусматривает стадии: Полный текст патента 1. Область техники, к которой относится изобретение Данное изобретение относится к биотехнологии растений и описывает ячмень и солод, дефектные в синтезе фермента липоксигеназы (LOX) LOX-1, обеспечивая таким образом новый сырьевой материал для промышленного применения. Например, указанный сырьевой материал может быть использован для производства нового и особого пива со стабильным вкусом, не имеющего или имеющего незначительные количества придающего привкус соединения транс-2-ноненаля (T2N). Указанный T2N образуется при последовательном действии ферментов LOX-пути, где LOX-1 представляет первичную активность, сообщающую диоксигенирование линолевой кислоты с образованием 9-гидропероксиоктадекадиеновой кислоты (9-HPODE). Ячмень и продукты растительного происхождения данного изобретения не обнаруживают или обнаруживают незначительные количества 9-HPODE. Кроме того, данное изобретение относится к напиткам, получаемым с использованием указанного ячменя и/или солода. Одной из задач этого исследования, связанной с современным производством пива, является определение молекулярных факторов для качества и стабильности пива. Большую часть пива получают на основе ячменя (Hordeum vulgare, L.). Он является однодольным культурным растением, выращиваемым во многих частях света не только вследствие его экономической важности в качестве источника промышленных продуктов, таких как пиво, но также в качестве источника корма для животных. В настоящее время Соединенные Штаты являются одним из ведущих производителей ячменя для солодоращения, с приблизительно 13% мирового урожая; Канада, Австралия и Европа, вместе отвечают за 70% этой продукции (Bios Intern., 2001). Селекционеры ячменя непрерывно пытаются развить стабильные высокоурожайные сорта, которые являются агрономически надежными. Для выполнения этой задачи попытки включали неспецифический мутагенез посредством химической обработки или облучения для модификации представляющих интерес признаков, например, для изменения экспрессии специфических генов, которые могут иметь вредные действия на рост растений и продуктивность сельскохозяйственной культуры в целом - но также на признаки, придающие дополнительное качество продукту, произведенному из этой сельскохозяйственной культуры. Хорошо установлено, что азид натрия, NaN 3 , является полезным химикалием для мутагенеза ячменя. Конкретно, NaN 3 -вызываемый мутагенез использовали для индуцирования генетических изменений в ячмене для генерирования мутантов, блокированных в синтезе антоцианинов и проантоцианинов (von Wettstein et al., 1977; von Wettstein et al., 1985; Jende-Strid, 1991; Jende-Strid, 1993; Olsen et al., 1993). Второй пример относится к зернам ячменя, мутагенизированным NaN 3 , для скрининга на высокие уровни свободного фосфата с целью идентификации низкофитатных мутантов (Rasmussen and Hatzak, 1998); были идентифицированы в целом 10 мутантов из 2000 подвергнутых скринингу зерен. Хотя основным недостатком в генетике ячменя была невозможность специфического исследования функции генов посредством обратной генетики, прямые генетические скрининги - например, после NaN 3 -индуцированного мутагенеза - продолжаются в отношении улучшений, которые связаны с питательными параметрами и параметрами качества продуктов ячменя и солода. За исключением суммарного и общего предсказания, селекционер не может предсказывать результат новых линий растений при развитии в традиционном процессе селекции. Эта непредсказуемость обусловлена в основном отсутствием контроля на клеточном уровне, более конкретно, на уровне ядерной ДНК сложность которого является огромной. Ряд других факторов влияют на результат процесса селекции растений, например климат и качество почвы в географическом местоположении размножения растений. В результате, разные селекционеры ячменя, которые используют традиционные способы, никогда не будут получать растения с идентичными признаками. В традиционном процессе селекции наиболее трудной задачей является идентификация растений, которые являются генетически лучшими, не только в отношении представляющего интерес признака, но также в отношении физиологических результатов, имеющих значение для роста растений. Этот способ селекции является особенно трудным, когда другие ставящие в тупик признаки маскируют представляющий интерес признак. Когда существующие в настоящее время процедуры селекции включают определение последовательности ДНК мутированного гена, это происходит на поздней стадии программы селекции - т.е. после характеристики мутанта, например, как было недавно описано для скрининга химически индуцированных мутаций в Arabidopsis и других растениях (Colbert et al., 2001). До сих пор создание мутаций генов обозначаемых потерей функции в масштабе всего генома сообщались для дрожжей Saccharomyces cerevisiae (Giaever et al., 2002). Для растения Arabidopsis, 21700 из ~29454 предсказанных генов инактивировались встраиванием Т-ДНК-последовательностей Agrobacterium (Alonso et al., 2003). До сих пор не является необычным, что традиционный процесс селекции от первого мутагенеза или скрещивания до выхода на рынок растений или семян занимает > 10 лет. Конкретно, было бы превосходным обеспечение селекционера растений способами детектирования мутаций в генах, связанных с представляющим интерес признаком. Такие усовершенствования могли бы повысить предсказуемость в программах селекции, в частности, когда селекция мутантов направлена на мутанты, имеющие нонсенс-мутации ("бессмысленные" мутации) в кодирующей белок части представляющего интерес гена. В других случаях это могло бы быть также предпочтительным для ранней идентификации мутаций ДНК, например, для прекращения дальнейшей селекции линий, характеризующихся мутациями промоторов в представляющем интерес гене, или в случае, когда другие мутации ДНК влияют на экспрессию - просто потому, что факторы окружающей среды и физиологические факторы могли бы вызывать обращение признака, индуцируемого данным мутагеном. Таким образом, существует потребность в нахождении альтернативных путей детектирования представляющих интерес мутаций в программе селекции. Это должно делать весь процесс селекции более быстрым и представляющим больший интерес в экономическом смысле с максимизацией количества зерна, продуцируемого на сельскохозяйственной земле. Большая часть продуцируемого ячменя содержит сорта для получения солода, зерна которых превращают в солод посредством процессов контролируемого замачивания, проращивания и сушки ячменя. Небольшую часть этого солода используют в качестве ингредиентов в пищевой промышленности, в то время как большую часть этого солода используют затем в качестве основного ингредиента в получении солодовых напитков, включая, но не ограничиваясь ими, пиво и виски. В пивоваренном заводе измельченный солод подвергают процессу затирания солода, предусматривающему ступенчатое увеличение температуры водной суспензии солода, которое вызывает ступенчатую, ферментативную деградацию и экстракцию, например, полимеров зерен крахмала и β-глюкана. После фильтрования этот водный затор кипятят с высушенными шишками хмеля с получением сусла. Затем указанное сусло ферментируют с дрожжами с получением пивного продукта, который - после созревания - разливают в бутылки. Сусло может быть также использовано для получения неферментированных солодовых напитков. Вкусовые качества и стабильность вкуса напитка являются важным фактором, относящимся к композиции ячменя и солода. Это обусловлено тем, что природные вкусовые молекулы, происходящие из указанного ячменя и солода - генерируемые действием ферментов, экстрагированных из указанного ячменя и солода могут придавать нежелательные вкусовые качества конечному продукту (Drost et al., 1990). В этом отношении, образование летучего соединения, придающего привкус картона, представляет, по-видимому, особый биохимический, а также экономический интерес. В 1970 году молекула, ответственная за привкус картона, была выделена и идентифицирована как T2N, алкеналь, содержащий девять атомов углерода (С 9 ) (Jamieson and Gheluwe, 1970). Поскольку порог вкусового ощущения для T2N у человека является крайне низким, определенным ранее как равный приблизительно 0,7 нМ или 0,1 м.д. (Meilgaard, 1975), продукты даже с незначительными уровнями этого альдегида рассматриваются как состарившиеся вследствие несвойственного вкуса продукта. Кроме того, выделение T2N в результате разложения T2N-аддуктов во время хранения пива может вызывать порчу продукта (Nyborg et al., 1999). Исследования с радиоактивным мечением ткани растения показали, что ноненали образуются из C 18 жирной кислоты, линолевой кислоты, тогда как гексанали и нонадиенали образуются из С 18 жирной кислоты, линоленовой кислоты (Grosch and Schwarz, 1971; Phillips and Galliard, 1978). Эти и многочисленные последующие наблюдения - например, суммированные Tijet et al. (2001), Noorderraeer et al. (2001) и Matsui et al. (2003) - были интерпретированы как доказательство того, что T2N образуется последовательным действием специфических для LOX-пути ферментов, причем действие LOX представляет собой раннюю ферментативную стадию. В соответствии с этим наблюдением, Kurodo et al. (2003) обнаружили, что солод содержит термостабильный ферментативный фактор, который является необходимым для превращения продуктов, произведенных LOX, в T2N. Зерно ячменя содержит три фермента LOX, известных как LOX-1, LOX-2 и LOX-3 (Van Mechelen et al., 1999). В то время как LOX-1 катализирует образование 9-HPODE - предшественника T2N, а также тригидроксиоктадеценовых кислот (сокращаемых как "ТНОЕ" или просто "THA") - из линолевой кислоты, LOX-2 катализирует превращение линолевой кислоты в 13-HPODE, которая метаболизируется далее до гексаналя (фиг. 1B), C 6 альдегида с уровнем порога вкусового ощущения около 0,4 м.д. (Meilgaard, supra). Хотя специфичность продукта LOX-3 остается невыясненной, очень низкий уровень экспрессии соответствующего гена, как показано van Mechelen et al. (supra), позволяет предположить, что его вклад в образование T2N является незначительным. Продолжается исследование для определения, является ли активность LOX единственным ферментативным источником для генерирования гидропероксидных предшественников линолевой кислоты, связанных с образованием T2N-специфических привкусов, или способствует ли этому также процесс самоокисления жирных кислот. Следует отметить, что C 18 гидропероксиды могут дополнительно превращаться более чем 7 различными семействами ферментов растений и животных, причем все эти реакции вместе называют LOX-путем (Feussner and Wasternack, 2002); этот путь называют также путем оксилипинов. Оксилипины, как указывает их название, является оксигенированными произведенными из липидов молекулами, которые происходят из оксигенирования ненасыщенных жирных кислот посредством LOX-реакции и включают также любые молекулы, произведенные из таких оксигенированных молекул. Зерна ячменя и растения ячменя, имеющие белок LOX-1, характеризующийся уменьшенной активностью, были описаны в РСТ-заявке PCT/IB01/00207, опубликованной в виде WO 02/053721A1 Douma et al. Однако указанная заявка не описывает генерирование и анализ зерен ячменя с неактивным ферментом LOX-1. Известны несколько примеров мутированных растений, которые синтезируют низкие уровни LOX. Например, в начале 1980-х годов были идентифицированы три линии сои, недостаточные по одному из трех ферментов LOX в зрелых семенах сои: (i) LOX-1. Хотя молекулярная основа нуль-мутации LOX-1 остается неопределенной, она соответствует отсутствию соответствующей зрелой мРНК (Hildebrandt and Hymowitz, 1982; Start et al., 1986); (ii) LOX-2. Транскрипты для мутированного гена были детектированы, и наблюдали изменение единственного основания, которое заменяет гистидиновый лиганд железа активного центра, приводя к нестабильности фермента (Davies and Nielsen, 1986; Wang et al., 1994); (iii) LOX-3. Нуль-мутанты LOX-3 не обнаруживали детектируемых уровней соответствующего транскрипта, возможно, вследствие цис-действующих элементов в промоторе гена (Kitamura et al., 1983; Wang et al., 1995). Было обнаружено, что в семенах гороха линия null-LOX-2 несет дефект, приводящий к отсутствию большей части белка LOX-2 (Forster et al., 1999). Поскольку эта линия обнаруживала большое уменьшение количества мРНК для LOX-2, было сделано предположение, что эта мутация вызывала разительное уменьшение стабильности мРНК. В рисе скрининг иммуноблота экстрактов выявил присутствие двух природных сортов Daw Dam и CI-115, в каждом отсутствовал один из трех ферментов LOX (Ramezanzadeh et al., 1999). Было определено, что количество гексаналя, пентаналя и пентанола в нормальном рисе со всеми тремя LOX значительно индуцировалось во время хранения, в то время как в Daw Dam и CI-115 оно уменьшалось в диапазоне 6б%-80%. Несмотря на то, что эти результаты позволяют предположить, что отсутствие ферментов LOX в зернах риса ослабляют окилительную порчу, молекулярные определяющие факторы, которые придают свойства отсутствия LOX Daw Dam и CI-115, остаются невыясненными. Было показано, что как опосредованная антисмысловой супрессией, так и опосредованная косупрессией трансгенная элиминация генов LOX применима для выяснения функции специфических ферментов LOX и их соответствующих продуктов в передаче сигналов защиты растений. В Arabidopsis, например, элиминация фермента LOX приводила к уменьшению индуцированного появлением раны накопления жасмоновой кислоты (Bell et al., 1995). А результаты опосредованной антисмысловой супрессией элиминации гена, кодирующего LOX, показали участие соответствующего фермента в признаке несовместимости растения табака, устойчивого к грибному патогену (Ranee et al., 1998). Третий пример, в котором трансгенные подходы использовали для выяснения функций LOX, относится к роли LOX картофеля, обозначаемого LOX-H1, в росте и развитии растений картофеля (Leon et al., 2002). Показано, что элиминация LOX-H1 приводила к заметному уменьшению летучих алифатических С 6 альдегидов, соединений, участвующих в реакциях защиты растений и действующих либо в качестве молекул передачи сигналов для индуцированной появлением раны экспрессии гена, либо в качестве антимикробных веществ. Дополнительное исследование показало, что трансгенные растения картофеля с элиминированной экспрессией гена фермента LOX обнаруживали отклоняющееся от нормы развитие клубней (Kolomiets et al., 2001). Однако специфические оксилипины, которые ответственны за фенотип клубней, не были идентифицированы. В другом исследовании опосредованная антисмысловой супрессией элиминация LOX-H3 картофеля подавляла индуцированную реакцию защиты растения, одновременно с более высоким урожаем клубней (Royo et al., 1999). Вместе эти результаты позволяют предположить, что экспрессия генов, кодирующих ферменты LOX, является важной в развитии растений, причем возможно, что некоторые ферменты LOX играют защитную роль против патогенов, тогда как другие ферменты LOX генерируют продукты, которые могут действовать как регуляторы развития клеток. Важно также отметить, что плоды томата с уменьшенными на 2-20% уровнями двух ферментов LOX не обнаруживали значимых изменений во вкусовых летучих компонентах в сравнении с плодами дикого типа (Griffits et al., 1999). Это открытие позволяет предположить, что либо очень низкие уровни LOX являются достаточными для генерирования альдегидов и спиртов, либо другие ферменты LOX являются активными в генерировании этих соединений. Происходит растущее осознание вреда окислительных ферментов для пищевой промышленности и промышленности напитков вследствие их действия на важные аспекты, связанные со вкусом и цветом производимых из растений продуктов. В этом отношении LOX представляют интерес вследствие их способности индуцировать образование свободных радикалов, которые затем могут атаковать другие компоненты, такие как витамины, красители, фенольные вещества, белки и т.д. Примечательно, что, как считается, некоторые из свободных радикалов играют роль в самоокислении свободных жирных кислот. Некоторые образующие свободные радикалы вещества могут противостоять термальной обработке и, следовательно, остаются достаточно активными в переработанных пищевых продуктах для инициации изменений качества во время хранения такого продукта. Антиоксиданты широко используются в качестве ингибиторов LOX, некоторые из которых ингибируют также самоокисление субстратов LOX. Однако ингибиторы LOX, применимые в качестве улучшающей вкус добавки для напитков, не были идентифицированы. Роль ферментов LOX связана также с проблемами, лежащими вне области производства пива, такими как LOX-катализируемое генерирование гидропероксижирных кислот, которые ингибируют образование микотоксинов в растениях, чувствительных к грибковому загрязнению, например, как описано в Патенте США № 5942661, выданном Keller. Хотя роль ферментов LOX в защите растений и реакциях на появление ран остается менее ясной, эти ферменты индуцируются после появления раны и заражения патогенами (Bell and Mullet, 1991; Bell and Mullet, 1993; Melan et al., 1993; Sarvitz and Siedow, 1996). Роль ферментов LOX в появлении ран и защите растений могла бы заключаться в продуцировании реакционноспособных гидропероксидов жирных кислот против патогенов (Rogers et al., 1988). Альтернативно, LOX могут индуцироваться стрессами для продуцирования сигнальных молекул, таких как метилжасмонат (Bell et al., supra). Были описаны также стратегии, в которых 13-HPODE, продуцируемая действием фермента LOX, действует в качестве субстрата для гидропероксидпревращающих ферментов для продуцирования связанных с привкусом альдегидов (Noordermeer et al., 2002; Husson and Belin, 2002). Сходные способы описаны в многочисленных патентах, например в патенте США № 6150245, выданном Hausler et al., и в Патенте США № 6274358, выданном Holtz et al. Было также показано, что ферменты LOX способствуют некоторым полезным эффектам в отношении хлебопечения (Casey, 1997). Кроме того, в Патенте США № 6355862 В1, выданном Handa and Kausch, описано, что качество фрукта может быть повышено ингибированием образования LOX, приводя к увеличению срока годности этого продукта. Таким образом, существует неудовлетворенная потребность в растениях ячменя, по существу, не имеющих LOX-активности, так как напитки, приготовленные из таких растений, будут иметь очень низкие уровни T2N. Кроме того, данное изобретение утверждает, что напитки, приготовленные из таких растений, будут иметь очень низкие уровни 9,12,13-ТНОЕ. Более того, такие растения могут быть применимы для других целей. Неожиданно, данное изобретение раскрывает способы получения растений ячменя, не имеющих LOX-1-активности или имеющих очень низкую LOX-1-активность. В частности, данное изобретение описывает нуль-мутации в гене LOX-1. Перспективные преимущества данного изобретения включают полную элиминацию T2N из соответствующего ответвления пути LOX, и, следовательно, данное изобретение обеспечивает наилучший путь контроля уровней T2N в зерне ячменя; и пиво, полученное из этого зерна, проявляет исключительную стабильность вкуса после продолжительного хранения, даже при повышенных температурах. Интересно, что данное изобретение обеспечивает также способы раннего детектирования мутаций, и, следовательно, данным изобретением были устранены недостатки поздней характеристики мутантов. Это позволяет использовать новую привлекательную процедуру для генерирования улучшенных сортов ячменя для получения солода введением последовательного использования характеризации фенотипа и определения последовательности ДНК генов-мишеней в популяции мутанта раньше по времени в процессе селекции. Выделенные растения могут быть улучшены дополнительно использованием различных способов селекции растений. Данное изобретение решает существующие проблемы, ограничения и недостатки, связанные с присутствием активного фермента LOX-1 в ячмене. Во-первых, данное изобретение обеспечивает новый, эффективный способ скрининга, который значительно уменьшает время и трудоемкость скрининга химически мутагенизированного ячменя. Во-вторых, данное изобретение включает новые нуль-LOX-1-ячмени, например, применимые в производстве пива со стабильным вкусом. Теоретической предпосылкой для LOX-мутантов растений, как описано выше, являются растения, имеющие уменьшенные уровни активности LOX. В отличие от этого, данное изобретение преодолевает ограничения и недостатки, связанные с низкой или остаточной активностью LOX, обеспечением путей эффективного генерирования нуль-LOX-1-растений ячменя. Конкретные отличия включают следующее: (i) в противоположность растениям ячменя, описанным в РСТ-заявке PCT/IBO1/00207, опубликованной в виде WO 02/053721A1, Douma et al., растения данного изобретения, по существу, не содержат активности LOX-1, предпочтительно все растения являются истинными нуль-LOX-1-растениями - т.е. растения обнаруживают полную потерю функции LOX-1-белка; (ii) описанный здесь признак истинный нуль-LOX-1-признак может быть идентифицирован скринингом на присутствие нонсенс-мутации (бессмысленной мутации) в соответствующем гене. Таким образом, растения ячменя, гомозиготные в отношении этого признака, могли бы быть полностью блокированы в синтезе активного фермента, независимо от условий выращивания или воздействий окружающей среды. Это является идеальным качеством в области селекции растений и контрастирует с результатом возможного молекулярного сценария в LOX-мутантах сои предшествующего уровня техники, где биотические и абиотические условия могли влиять на изменения в физиологическом состоянии клеток, вызывая стабилизацию мРНК с последующей трансляцией LOX; (iii) если признак релевантности в мутантах LOX сои и риса включал уменьшенные уровни имеющего интенсивный запах соединения гексаналя в главном пищевом продукте, данное изобретение относится к низким уровням соединения T2N со специфическим вкусом в напитке, а также к более низким уровням 9,12,13-ТХОЕ в напитке; (iv) LOX-мутанты сои и риса затронуты в молекулах пути LOX ниже по ходу процесса от 13-HPODE, тогда как нуль-LOX-1-признак относится к ветви пути LOX, которая содержит молекулы ниже по ходу процесса от 9-HPODE; (v) в то время как мутанты сои содержат индуцированные облучением мутации в генах LOX, a Daw Dam и Cl-115 представляют отобранные природно встречающиеся сорта линий селекции риса, мутации в растениях ячменя, имеющих нуль-LOX-1-признак, были индуцированы химически с использованием NaN 3 . Таким образом, целью данного изобретения является обеспечение растений ячменя, их частей или фрагментов, содержащих менее 5%, предпочтительно менее 1% активности LOX-1 растения ячменя дикого типа. Второй целью данного изобретения является обеспечение зерен из растения ячменя, содержащих менее 5%, предпочтительно менее 1% активности LOX-1 растения ячменя дикого типа. Третьей целью данного изобретения является обеспечение композиций, содержащих растение ячменя или его части или фрагменты, содержащих менее 5%, предпочтительно менее 1% активности LOX-1 растения ячменя дикого типа. Следующей целью данного изобретения является обеспечение композиций солода, содержащих обработанное растение ячменя, содержащее менее 5%, предпочтительно менее 1% активности LOX-1 растения ячменя дикого типа. Композиции солода могут быть предпочтительно чистыми композициями солода. Однако композиции солода могут быть также, например, смесями ячменя и солода. Целью данного изобретения является также обеспечение напитков, имеющих стабильные органолептические качества, причем указанные напитки производят с использованием растения ячменя данного изобретения или его части. В частности, предпочтительно, указанные напитки производят с использованием композиции солода, такой как чистая композиция солода или смесь ячменя и солода, описанные здесь выше. В предпочтительном варианте осуществления данного изобретения указанные напитки состоят из пива. Дополнительной целью данного изобретения является обеспечение напитка, имеющего стабильные органолептические качества, причем указанный напиток производят с использованием растения ячменя, в котором отношение 9,12,13-тригидроксиоктадеценовой кислоты (здесь сокращаемой как 9,12,13-ТНОЕ или просто как 9,12,13-THA) к 9,10,13-тригидроксиоктадеценовой кислоте (здесь сокращаемой как 9,10,13-ТНОЕ или просто как 9,10,13-THA) в указанном напитке равно максимально 1,8. Предпочтительно указанный напиток представляет собой пиво. Кроме того, целью данного изобретения является обеспечение композиций, например, пищевых композиций, кормовых композиций или ароматных композиций сырьевых материалов, которые содержат растение ячменя или его части в соответствии с данным изобретением. Кроме того, целью данного изобретения является обеспечение способов экспрессии рекомбинантного белка в растении ячменя в соответствии с данным изобретением, причем указанный способ предусматривает трансформацию указанного растения последовательностью нуклеиновой кислоты, содержащей, в виде функционально связанных компонентов, промотор, экспрессируемый в растениях ячменя или их частях, ДНК-последовательность, кодирующую указанный рекомбинантный белок, и район терминации транскрипции, экспрессирующей посредством этого указанный рекомбинантный белок в указанном растении ячменя. Далее, целью данного изобретения является обеспечение способов уменьшения уровней белка в растении ячменя данного изобретения, причем указанный способ предусматривает трансформацию указанного растения последовательностью нуклеиновой кислоты, содержащей, в виде функционально связанных компонентов, промотор, экспрессируемый в растениях ячменя или их частях, ДНК-последовательность, кодирующую указанный рекомбинантный белок, и район терминации транскрипции, причем экспрессия указанной ДНК-последовательности уменьшает экспрессию гена, кодирующего указанный белок посредством антисмысловой супрессии или косупрессии или интерференции РНК. Дополнительной целью данного изобретения является обеспечение способов получения растения ячменя, содержащего менее 5%, предпочтительно менее 1% активности LOX-1 растения ячменя дикого типа, предусматривающих стадии: (i) определения активности LOX-1 в зернах ячменя дикого типа или их частях; и (ii) мутагенеза растений ячменя, и/или зерен ячменя, и/или зародышей ячменя с получением посредством этого генерации МО ячменя; и (iii) отбора указанных мутагенизированных растений ячменя, зерен и/или зародышей в течение по меньшей мере 2 генераций с получением посредством этого растений ячменя генерации Мх, где х обозначает целое число ≥2; и (iv) получения зерен или их частей из указанных растений ячменя Мх; и (v) определения активности LOX-1 в указанных зернах или их частях; и (vi) отбора растений, в которых активность LOX-1 мутагенезированных зерен или их частей равна менее чем 5% активности LOX-1 зерен дикого типа или их части; с получением посредством этого растения ячменя, содержащего менее чем 5% активности LOX-1 растения ячменя дикого типа. Далее, целью данного изобретения является обеспечение способов получения напитка, имеющего стабильные органолептические качества, предусматривающих стадии: (i) обеспечения солодовой композиции в соответствии с данным изобретением; (ii) переработки указанной солодовой композиции в напиток; с получением посредством этого напитка со стабильными органолептическими качествами. Дополнительной целью данного изобретения является обеспечение способов получения солодовой композиции с низкой активностью LOX-1, предусматривающих стадии: (i) обеспечения зерен в соответствии с данным изобретением; (ii) замачивания указанных зерен; (iii) проращивания замоченных зерен при заданных условиях; (iv) обработки пророщенных зерен нагреванием; с получением посредством этого солодовой композиции с низкой активностью или без активности LOX-1. В одном предпочтительном варианте осуществления данное изобретение основано на непредсказанном результате функциональных исследований мутанта ячменя D112 (далее называемого также "мутантом D112" или "ячменем D112"), который обнаруживал полную потерю функции в отношении основного 9-HPODE-образующего фермента LOX-1. Неожиданным открытием было обнаружение распределения 10%:90% 9-HPODE:13-HPODE в биохимических анализах, предназначенных для определения профиля продукта после LOX-катализируемого превращения линолевой кислоты. При условии крайне низкого порога вкусового ощущения T2N, еще более неожиданным было то, что деградация остаточной 9-HPODE и т.п. в зернах мутанта D112 вызывала лишь очень низкое высвобождение T2N - значительно более низкое, чем порог вкусового ощущения T2N - во время старения продуктов пива, изготовленных из солода указанных зерен. Изучение этих результатов из анализов с использованием зерен дикого типа и нуль-LOX-1-зерен дало ясное доказательство того, что высокая активность LOX-1 может усиливать затхлый привкус картона T2N, подтверждая таким образом важную роль пути LOX в регуляции образования алкеналя. Этот вывод противоречит утверждению Liegeois et al. (supra), который позволил предположить, что активность LOX вносит только небольшую долю молекул-предшественников T2N. Признак нуль-LOX-l может быть введен в любое другое растение ячменя, например в установленные сорта ячменя, делая возможным получение напитков со стабильным вкусом с пролонгированным сроком годности. Это может быть, например, осуществлено традиционными способами селекции, хорошо известными специалистам в данной области. Этот подход будет независимым не только от географического района, где выращен полученный из мутанта D112 ячмень, но также будет независимым от местоположения, где производят и продают потребителям полученное из мутанта D112 пиво. Растения ячменя мутанта D112 или произведенные из них растения являются потенциально важным экономическим фактором для фермеров, выращивающих эту сельскохозяйственную культуру, и для пивоваренных заводов, которые используют их в качестве сырьевого материала для производства пива или производства других напитков на основе ячменя. Ожидается, что и другие применения, которые зависят от сырьевых материалов без образующих 9-HPODE/9-HPOTE активностей, также выиграют от этих свойств мутанта ячменя D112. В соответствии с одним вариантом осуществления данного изобретения обеспечены несколько новых мутантов ячменя, применяемых для получения солода, например мутанта D112 ячменя или мутанта А618 ячменя (называемого здесь также "мутантом А618" или "ячменем А618"). Таким образом, данное изобретение относится к зернам мутантов D112 или А618 ячменя, к растениям ячменя D112 или А618 и к способам получения растения ячменя из скрещивания мутантов D112 или А618 ячменя друг с другом или с другой линией ячменя. Кроме того, данное изобретение включает нуль-LOX-1-варианты, генерированные мутагенезом или трансформацией мутанта ячменя D112 или А618. Таким образом, все растения, полученные с использованием мутантов ячменя D112 или А618 - или их производного - в качестве исходного растения находятся в рамках данного изобретения. В другом аспекте данное изобретение обеспечивает регенерируемые клетки для применения в культуре ткани мутантного растения ячменя D112 или А618. Эта культура ткани будет предпочтительно применима для регенерации растений, имеющих характеристики предыдущих растений ячменя, в том числе морфологические и генетические характеристики. Регенерированными клетками в таких культурах ткани будут зародыши, протопласты, меристематические клетки, каллус, пыльца, пыльники и т.д. Понятно, что данное изобретение обеспечивает также растения ячменя, регенерированные из культур ткани данного изобретения. В предпочтительном варианте осуществления данное изобретение включает солод, полученный из зерен нуль-LOX-1-ячменя. Данное изобретение относится также к композициям сусла, приготовленным из растений нуль-LOX-1-ячменя или их частей или из композиций солода, приготовленных из таких растений ячменя. Далее, данное изобретение относится к напиткам, таким как пиво, произведенным с использованием либо зерен нуль-LOX-1-ячменя данного изобретения, либо солода, полученного из указанных зерен. Кроме того, данное изобретение относится к продукту растительного происхождения, полученному из растения нуль-LOX-ячменя или его частей. Такой продукт растительного происхождения может быть любым продуктом, полученным в результате переработки указанного растения ячменя или его частей. Предпочтительно, указанный продукт растительного происхождения выбран из группы, состоящей из солода, сусла, ферментированных напитков, таких как пиво, неферментированных напитков, пищевых продуктов, таких как ячменная мука и кормовые продукты. Целью данного изобретения является также обеспечение зерен нуль-LOX-1-ячменя, обнаруживающих такие уровни устойчивости к болезням, которые являются неотличимыми от растений ячменя дикого типа или даже имеют улучшенную устойчивость к болезням. Далее, данное изобретение включает зерна нуль-LOX-1-ячменя и солод, полученный из указанных зерен, причем как зерна, так и солод обнаруживают уменьшенные уровни микотоксинов. Данное изобретение включает также сорта нуль-LOX-1-ячменя с повышенной устойчивостью к болезням относительно растений дикого типа. Далее, описаны растения нуль-LOX-1-ячменя, имеющие уменьшенную устойчивость к болезням относительно растений дикого типа, при условии, что другие характеристики указанных растений обеспечивают преимущества, которые являются более важными, чем свойство уменьшенной устойчивости к болезням. Кроме того, данное изобретение обеспечивает зерна нуль-LOX-1-ячменя, применимые для получения ароматических веществ, происходящих из пути LOX, в том числе соединений ноты зелени. Кроме того, данное изобретение обеспечивает трансгенные растения нуль-LOX-1-мутантов ячменя D112 или А618, или произведенные из них растения, где введенный ген (введенные гены) придают такие признаки, как устойчивость к гербицидам, устойчивость к насекомым, устойчивость в отношении бактериальных, грибковых или вирусных болезней, повышенную пищевую ценность и промышленную применимость. Этот ген может быть эндогенным геном ячменя или, альтернативно, трансгеном, введенным способами генной инженерии. Наконец, данное изобретение обеспечивает способы уменьшения активности LOX-1 с использованием ингибиторов LOX-1. Продукты растений или продукты, полученные из растений, в том числе напитки и пиво, полученные указанными способами, могут иметь свойства, сходные со свойствами продуктов, полученных из нуль-LOX-1-ячменя в качестве сырьевого материала. Эти и другие признаки, аспекты и преимущества данного изобретения будут более понятными при ссылке на следующие определения, описания, примеры, прилагаемую формулу изобретения, а также сопутствующие списки последовательностей и рисунки. 3.1 Определения. В данном описании, фигурах и таблицах, которые следуют далее, используется ряд терминов. Для обеспечения описания и формулы изобретения, включающих такие термины во всем их объеме, приведены следующие определения. В применении здесь, формы единственного числа могут обозначать один или более, в зависимости от контекста, в котором они используются. Термин "агрономический признак" описывает фенотипический признак растения, который способствует производительности или экономической ценности указанного растения. Такие признаки включают устойчивость к болезням, устойчивость к насекомым, устойчивость к вирусам, устойчивость к нематодам, засухоустойчивость, солеустойчивость, урожайность, высоту растений, дни до созревания, сортировку семян по размерам (т.е. фракционирование семян по размерам), содержание азота семян и т.п. Под "антисмысловой нуклеотидной последовательностью" понимают последовательность, которая находится в обращенной ориентации относительно нормальной кодирующей 5' →3' ориентации этой нуклеотидной последовательности. В случае присутствия в клетке растения эта антисмысловая ДНК-последовательность предпочтительно предотвращает нормальную экспрессию нуклеотидной последовательности данного эндогенного гена и может нарушать продуцирование соответствующего нативного белка. Термин "ячмень" в ссылке на процесс получения пива, в частности, при использовании для описания процесса получения солода, обозначает зерна ячменя. Во всех других случаях, если нет особых указаний, "ячмень" обозначает растение ячменя (Hordeum vulgare, L.), в том числе любые сорта ячменя. Под "устойчивостью к болезням" подразумевают, что эти растения лишены симптомов болезни, которые являются результатом взаимодействий растение-патоген. Таким путем происходит предотвращение вызывания болезней растений патогенами и ассоциированных с конкретной болезнью симптомов или, альтернативно, симптомов болезни. Альтернативно, симптомы болезней, вызываемых патогенами, минимизируются или уменьшаются. "Зерновое" растение, определяемое в этой публикации, является членом семейства растений Gramineae, культивируемых прежде всего для получения их крахмалсодержащих семян. Зерновые растения включают, но не ограничиваются ими, ячмень (Hordeum), пшеницу (Triticum), рис (Oryza), кукурузу (Zea), рожь (Secale), овес (Avena), сорго (Sorghum) и Triticale, гибрид ржи и пшеницы. Под терминами "кодирующая" или "кодируемая", в контексте указанной нуклеиновой кислоты, подразумевают содержание информации для трансляции в определенный белок. Нуклеиновая кислота, кодирующая белок, может содержать некодирующие последовательности (например, интроны) в транслируемых районах этой нуклеиновой кислоты, или может быть лишена таких интронных нетранслируемых последовательностей (например, в кДНК). Информация, при помощи которой кодируется белок, указывается с использованием кодонов. В применении здесь, "экспрессия" в контексте нуклеиновых кислот должна пониматься как транскрипция и накопление смысловой мРНК или антисмысловой РНК, полученной из фрагмента нуклеиновой кислоты. Термин "экспрессия", используемый в контексте белков, относится к трансляции мРНК в полипептид. Под "ароматизирующими (вкусовыми) молекулами" подразумевают альдегиды и/или спирты, которые продуцируются и являются компонентами запаха и/или вкуса в растениях. В частности, ароматизирующие молекулы включают некоторые летучие спирты и альдегиды. Примеры ароматизирующих молекул, которые являются летучими, включают, но не ограничиваются ими, гексаналь, (3Z)-гексеналь, (2Е)-гексеналь, (2Е)-гексенол, (3Z)-ноненаль, (2Е)-ноненаль. Данное изобретение может быть использовано для модуляции ароматизирующих молекул в растениях. Термин "ген" обозначает сегмент ДНК, участвующий в продуцировании полипептидной цепи; он включает районы, предшествующие кодирующему району и следующие за кодирующим районом (промотор и терминатор). Эукариотические гены являются непрерывными, причем белки, кодируемые ими, состоят из экзонов, прерываемых интронами. После транскрипции в РНК эти интроны удаляются сплайсингом с образованием зрелой мессенджер-РНК (мРНК). "Сайты сплайсинга" между экзонами обычно определяются консенсусными последовательностями, действующими как сигналы сплайсинга для процесса сплайсинга, состоящего из делеции интрона из первичного РНК-транскрипта и соединения или слияния концов оставшейся РНК с каждой стороны вырезанного интрона. В некоторых случаях альтернативные или различные картины сплайсинга могут генерировать разные белки из одного и того же отрезка ДНК. Нативный ген может называться эндогенным геном. "Сайленсинг генов" (отключение генов) является способом изменения экспрессии гена. Он относится к РНК-сайленсингу, который является посттранскрипционным механизмом сайленсинга генов, консервативным среди различных организмов. Этот способ включает посттранскрипционный сайленсинг генов (PTGS) и интерференцию РНК (RNAi). PTGS является феноменом сайленсинга генов эндогенных и экзогенных гомологичных генов. Хотя большинство примеров PTGS обнаруживаются на эффектах, вызываемых косупрессией конструкций или экспрессией трансгенов в антисмысловой ориентации, его наблюдали также в растениях традиционных программ селекции, например, мутация Lgc1 в рисе (Kusaba et al., 2003). Было обнаружено, что эта мутация подавляет экспрессию глютелина посредством РНК-сайленсинга, возможно, вследствие делеции 3,5 т.п.н. между двумя высокосходными генами для глютелина, которая образует инвертированный повтор хвост-к-хвосту, который может образовывать двухцепочечную молекулу РНК и, следовательно, эффективный индуктор РНК-сайленсинга. Вторая форма сайленсинга РНК известна как интерференция РНК (RNAi), где основной предпосылкой является способность двухцепочечной РНК специфически блокировать экспрессию ее гомологичного гена при инъекции в клетки или поглощении клетками (Goenczy et al., 2000). В применении здесь "гетерологичная" при ссылке на нуклеиновую кислоту обозначает нуклеиновую кислоту, которая происходит из чужеродного вида, или, если она происходит из того же самого вида, является, по существу, модифицированной из ее нативной формы в составе и/или геномном локусе преднамеренным вмешательством человека. Термин "проращивание" обозначает в данном контексте начало или возобновление роста зерна ячменя в различных композициях, таких как нормальная почва в таком виде, в каком она находится в природе. Проращивание может происходить в почве сосудов в камерах для выращивания и т.п. или, например, иметь место на увлажненной фильтровальной бумаге, помещенной в стандартные лабораторные чашки Петри. Проращивание обычно предусматривает увлажнение зерен, набухание зерен и индукцию роста зародыша. Факторы окружающей среды, влияющие на проращивание, включают влажность, температуру и уровень кислорода. Наблюдается развитие корней и побегов. "Ноты зелени" является термином, описывающим летучие вкусовые и ароматизирующие молекулы, присутствующие в многочисленных растениях, и характеризуемые в органолептических терминах как свежие и травянистые. Эти молекулы продуцируются растением путем деградации липидов и свободных жирных кислот, таких как линолевая и линоленовая кислоты. В применении здесь, термин "выделенный" означает, что данный материал выделен из его исходного окружения. Например, природно встречающийся полинуклеотид или полипептид, присутствующий в живом организме, не является выделенным, но тот же самый полинуклеотид или полипептид, отделенный от некоторых или всех сосуществующих веществ в природной системе, является выделенным. Такие полинуклеотиды могли бы быть частью вектора и/или такие полинуклеотиды или полипептиды могли бы быть частью композиции и все еще быть выделенными, так как такие вектор или композиция не является их природным окружением. Термин "зерно" определяется как содержащее зерновку злаков, также обозначаемую как внутреннее семя, нижнюю цветковую чешую и верхнюю цветковую чешую. В большинстве сортов ячменя нижняя цветковая чешуя и верхняя цветковая чешуя прикреплены к зерновке и являются частью зерна после обмолота. Однако встречаются также голозерные сорта ячменя. Термины "зерна" и "зерно" используют здесь взаимозаменяемо. "Созревание зерен" или "развитие зерна" обозначают период, начинающийся оплодотворением, в котором метаболизируемые запасы, например депонированные сахара, олигосахариды, крахмал, фенольные вещества, аминокислоты и белки откладываются, с направлением или без направления в вакуоли, в различные ткани в зернах (зерне), например, в эндосперме, тесте (семенной оболочке), алейроновом слое и щитке, приводя к увеличению зерен (зерна), наливу зерен (зерна) и завершению высыхания зерен (зерна). Термин "активность LOX-1" относится к ферментативной активности фермента LOX-1 ячменя. В частности, в контексте данного изобретения "активность LOX-1" является катализируемой ферментом диоксигенирования линолевой кислоты до 9-HPODE. Даже хотя фермент LOX-1 способен катализировать другие реакции, для цели определения активности LOX-1 в соответствии с данным изобретением, будет рассматриваться только образующая 9-HPODE активность. Фиг. 1В представляет биохимический путь, в котором линолевая кислота превращается в T2N. Термин "низкая LOX" (низкая липоксигеназная активность) относится к присутствию одной или нескольких мутаций в одном или нескольких эндогенных генах, вызывающих частичную потерю функции указанного фермента LOX, предпочтительно в отношении - но не только - ферментативной активности. Например, растения ячменя, описанные в РСТ-заявке PCT/IB01/00207, опубликованной в виде WO 02/053721A1, Douma et al., продуцируют мутированный фермент LOX-1, имеющий 10% остаточную активность в сравнении с соответствующим ферментом дикого типа. Термин "низкая LOX" при ссылке на растение относится к растению, имеющему частичную потерю функции указанного фермента LOX. "Получение солода" является особой формой проращивания зерен ячменя, имеющей место при контролируемых условиях окружающей среды, в том числе, но не только, ограниченных замочными танками и аппаратом для проращивания зерна солодовни. Согласно способу данного изобретения, получение солода (солодоращение) начинает происходить во время и/или после замачивания зерен ячменя. Процесс получения солода может быть остановлен высушиванием зерен ячменя. Понятно, что композиция солода, полученная из нуль-LOX-1-ячменя, содержит нуль-LOX-1-солод, такой как чистый нуль-LOX-1-солод, или любую смесь, содержащую нуль-LOX-1-солод. "Затирание солода" является инкубированием размолотого солода в воде. Затирание солода выполняют предпочтительно при конкретной температуре и в конкретном объеме воды. Температура и объем воды являются важными, так как они влияют на скорость уменьшения активности ферментов, происходящих из солода, и, следовательно, особенно на степень гидролиза крахмала, который может происходить. Затирание может иметь место в присутствии добавок, которые, как должно быть понятно, содержат любой источник углеводов, другой, чем солод, например, ячмень (в том числе нуль-LOX-1-ячмень), кукурузная или рисовая добавка, используемые в основном в качестве дополнительного источника экстракта. Требования в отношении обработки добавки в пивоварении зависят от состояния и типа используемой добавки и, в частности, от температуры клейстеризации или разжижения крахмала. Если температура разжижения крахмала выше температуры нормального осахаривания солода, то крахмал клейстеризуют и разжижают перед добавлением к затору. "Мутации" включают делеции, инсерции, трансверсии и точковые мутации в кодирующем и некодирующих районах гена. Делеции могут быть делециями всего гена или только части гена. Точковые мутации могут приводить к стоп-кодонам, мутациям со сдвигом рамки или заменам аминокислот. Соматическими мутациями являются мутации, которые имеют место только в определенных клетках или тканях растения и не являются наследуемыми следующей генерацией. Мутации зародышевой линии могут быть обнаружены в любой клетке растения и являются наследуемыми. Термин "нуль-LOX" относится к присутствию мутации в LOX-кодирующем гене, вызывающей полную потерю функции кодируемого фермента LOX. Мутации, которые генерируют кодоны преждевременной терминации (нонсенс-мутации) в гене, кодирующем LOX, представляют только один механизм, при помощи которого может быть получена полная потеря функции. Молекулярные подходы к получению полной потери функции фермента LOX предусматривают генерирование мутаций, которые вызывают полное отсутствие транскриптов для указанного фермента, или мутаций, которые полностью инактивируют кодируемый фермент. "Нуль-LOX" при ссылке на растение обозначает растение, имеющее полную потерю функции указанного фермента LOX. Термин "функционально связанные" является термином, используемым для ссылки на ассоциацию двух или более фрагментов нуклеиновых кислот на одном полинуклеотиде, так что на функцию одного влияет функция другого. Например, промотор функционально связан с кодирующей последовательностью, когда он способен влиять на экспрессию этой кодирующей последовательности, т.е. эта кодирующая последовательность находится под транскрипционным контролем этого промотора. Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в смысловой или антисмысловой ориентации. "ПЦР" или "полимеразная цепная реакция" хорошо известна специалистам в данной области как способ, используемый для амплификации конкретных ДНК-сегментов (патенты США с номерами 4683195 и 4800159, выданные Mullis et al.). Термин "растение" или "растительный материал" включает клетки растений, протопласты растений, культуры ткани клеток растений, из которых могут быть регенерированы растения ячменя, каллусы растений и клетки растений, которые являются интактными в растениях или частях растений, таких как зародыши, пыльца, яйцеклетки, цветки, зерна, листья, корни, кончики корней, пыльники, и любую часть или любой продукт растения. Термин "продукт растительного происхождения" обозначает продукт, полученный из переработки растения или части растения. Таким образом, указанный растительный продукт может быть, например, солодом, затором, ферментированным или неферментированным напитком, пищевым или кормовым продуктом. В применении здесь, "рекомбинантный" при ссылке на белок является белком, который происходит из чужеродного вида или, если он происходит из того же самого вида, модифицирован из его нативной формы в составе преднамеренным вмешательством человека. "РНК-транскрипт" обозначает продукт, происходящий из катализируемой РНК-полимеразой транскрипции ДНК-последовательности. Когда этот РНК-транскрипт является точной комплементарной копией ДНК-последовательности, его называют первичным транскриптом. Когда РНК-последовательность произведена из посттрансляционного процессинга первичного транскрипта, ее называют зрелой РНК. "Мессенджер-РНК" или "мРНК" обозначает РНК, которая не содержит интронов и которая может быть транслирована в белки клеткой. "кДНК" обозначает ДНК, которая комплементарна мРНК-матрице и произведена из мРНК-матрицы. Эта кДНК может быть одноцепочечной или может быть превращена в двухцепочечную форму с использованием, например, фрагмента Кленова ДНК-полимеразы I. "Смысловая РНК" является РНК-транскриптом, который включает мРНК и таким образом может быть транслирован в полипептид клеткой. "Антисмысловая РНК" обозначает РНК-транскрипт, который комплементарен всему транскрипту-мишени или части первичного транскрипта-мишени или мРНК-мишени и который блокирует экспрессию гена-мишени. Комплементарность антисмысловой РНК может быть любой частью конкретной нуклеотидной последовательности, т.е. в 5'-некодирующей последовательности, 3'-некодирующей последовательности, в интронах или в кодирующей белок последовательности. "Функциональная РНК" обозначает смысловую РНК, антисмысловую РНК или другую РНК, которая не может быть транслирована в белок, но все-таки влияет на клеточные процессы. Если нет других указаний, "T2N" обозначает свободную форму T2N. Термин "Т2N-потенциал" описывает химические вещества, которые способны высвобождать T2N или могут быть превращены в T2N, в одной или нескольких реакциях. Т2N-потенциал может быть измерен в виде концентрации T2N в растворе, например в сусле или пиве, после инкубирования (например, в течение 2 ч) при повышенной температуре (например, 100 °С) и низкой кислотности (например, рН 4,0). Эта обработка пробы вызывает высвобождение T2N из T2N-потенциала, например, из "T2N-аддуктов", причем этот термин используют для описания T2N, конъюгированного с одним или несколькими веществами, включая, но не ограничеваясь ими, белком (белками), сульфитом, клеточными остатками, клеточными стенками, или т.п. T2N-аддукты, сами по себе не ощущаются людьми как привкусы. Однако T2N, высвобождаемый из указанных T2N-аддуктов, например, нагреванием или кислотой, может приводить к возникновению привкуса. "Культура ткани" обозначает композицию, содержащую выделенные клетки того же самого или отличающегося типа, или собрание таких клеток, организованных в части растения, например, протопласты, каллусы, зародыши, пыльцу, пыльники и т.п. "Трансформация" обозначает введение ДНК в организм таким образом, что эта ДНК сохраняется, либо в виде внехромосомного элемента (без интеграции и стабильного наследования), либо в виде хромосомной интегрированной ДНК (с генетически стабильным наследованием). Если нет других указаний, способом, используемым здесь для трансформации Е. coli, является CaCl 2 -способ (Sambrook and Russel, supra). Для трансформации ячменя может выполняться Agrobacterium-опосредованная трансформация, в основном, как описано Tingay et al. (1997) и Wang et al. (2001), за исключением того, что другой сорт, такой как сорт Golden Promise, может быть использован в качестве хозяина. "Трансген" является геном, который был трансформирован в геном с использованием процедуры трансформации. В применении здесь, "трансгенная" включает ссылку на клетку, которая была модифицирована введением гетерологичной нуклеиновой кислоты, или на клетку, полученную из модифицированной таким образом клетки. Так, например, трансгенные клетки экспрессируют гены, которые не обнаруживаются в идентичной форме в нативной форме этой клетки, или экспрессируют нативные гены, которые в другом случае аномально экспрессируются, слабо экспрессируются или вообще не экспрессируются как результат преднамеренного вмешательства человека. Термин "трансгенное" в отношении растений, в частности, растений ячменя, в данном контексте не включает изменение клетки способами традиционной селекции растений, например, NaN 3 -произведенным мутагенезом, и природно встречающимися событиями, такими как события, встречающиеся без преднамеренного вмешательства человека. Термин "растение ячменя дикого типа" относится к обычному растению ячменя, предпочтительно этот термин относится к растению ячменя, из которого были получены растения ячменя данного изобретения, т.е. исходные растения. В одном предпочтительном варианте осуществления данного изобретения, "растение ячменя дикого типа" выбрано из группы, состоящей из сортов Celeste, Lux, Prectige, Saloon и Neruda. Более предпочтительно, "растение ячменя дикого типа" является сортом Barke. Сорта ячменя дикого типа или их семена обычно являются доступными, например, из обычных компаний, продающих семена. Данное изобретение может быть более понятным из следующего подробного описания и сопутствующего списка последовательностей (суммированного в табл. 9), который образует часть этой заявки. Указанная таблица дает список нуклеиновых кислот и полипептидов, которые описаны здесь, обозначение кДНК-клонов, которые содержат фрагменты нуклеиновых кислот, кодирующие полипептиды, представляющие все полипептиды или существенную часть этих полипептидов, и соответствующий идентификатор [SEQ ID NO:]. Описания последовательностей и прилагаемый к ним Список последовательностей подчиняются правилам, контролирующим описания нуклеотидных и/или аминокислотных последовательностей в заявках на патент. Список последовательностей содержит однобуквенный код для характеристик нуклеотидных и аминокислотных последовательностей, как определено в соответствии со стандартизованными рекомендациями (Cornish-Bowden, 1985; IUPAC-IUB Joint Commission on Biochemical Nomenclature, 1984), которые включены здесь в качестве ссылки. Символы и формат, используемые для данных по нуклеотидным и аминокислотным последовательностям, согласуются с правилами, определяющими описания последовательностей в заявках на патент. Фиг. 1 разделен на три схемы А, В и С. Фиг. 1А показывает, как могут быть размножены NaN 3 -мутагенизированные зерна ячменя. Зерна генерации МО выращивают до растений, которые развивают зерна генерации M1. Они могут быть посеяны и развиты в растения M1, которые производят новые зерна генерации М2. Затем выращивают растения М2 и получают зерна генерации М3, которые могут быть собраны и использованы для скрининг-анализов. М3 могут быть также посеяны и цветки соответствующих растений могут быть использованы для скрещиваний для получения растений генерации М4. Фиг. 1В является упрощенным представлением того, как действует биохимический путь LOX для деградации линолевой кислоты, давая в конечном счете T2N. Фиг. 1С иллюстрирует, каким образом линолевая кислота может быть превращена в соответствующую 9-гидропероксикислоту (9-HPODE) под действием LOX-1, с последующими дополнительными ферментативными превращениями эпоксиалкогольсинтазой и эпоксидгидролазой в 9,12,13-тригидрокси-10-октадеценовую кислоту (9,12,13-ТНОЕ). Фиг. 2 является графическим сравнением общих активностей LOX, измеренных в экстрактах зародышей сорта Barke, мутанта D112, и в контрольной пробе, содержащей инактивированный нагреванием экстракт зародышей сорта Barke. Фиг. 3 изображает сравнение при помощи диаграммы общих активностей LOX, измеренных в экстрактах зародышей мутанта А618, сорта Neruda и в контрольной пробе, содержащей инактивированный нагреванием экстракт зародышей сорта Barke. Фиг. 4 показывает общие активности LOX, измеренные в зернах 12 индивидуальных линий потомства М4 мутанта D112. Активности контрольных проб, состоящих из экстрактов зерен сорта Barke и инактивированных нагреванием экстрактов зерен сорта Barke, включены в сравнение. Фиг. 5 суммирует результаты анализов общей активности LOX в 90 индивидуальных экстрактах зерен линий потомства М5 мутанта D112. Активности контрольных экстрактов зерен сорта Barke и инактивированных нагреванием экстрактов зерен сорта Barke включены в сравнение. Фиг. 6 суммирует сравнение общих активностей LOX, измеренных в 40 индивидуальных экстрактах зерен линий потомства М4 мутанта А618. Активности контрольных проб с экстрактами зерен сорта Barke и инактивированных нагреванием экстрактов Barke включены в сравнение. Фиг. 7 состоит из двух отдельных иммуноблотов, показывающих, что иммунореактивный белок LOX-1 является недетектируемым в экстрактах зерен мутанта D112, генерации M3. Каждый иммуноблот зондировали антителом к LOX-1 ячменя, и пробы состояли из экстрактов клеток Е. coli, экспрессирующих рекомбинантный LOX-1 (дорожка 1), экстрактов зерен сорта Vintage (дорожка 2), мутантной линии G (дорожка 3 и дорожка 7), сорта Barke (дорожка 6 и дорожка 8) и отдельных линий мутанта D112, генерации M3 (дорожки 4-5 и 9-16). Указано положение иммунореактивного белка LOX-1. Фиг. 8 показывает два отдельных иммуноблота, детализирующих отсутствие LOX-1 в экстрактах зерен мутанта А618, генераций M3 и М4. Каждый иммуноблот зондировали антителом к LOX-1 ячменя, и пробы состояли из экстрактов зерен мутантной линии G (дорожка 1), сорта Neruda (дорожка 6 и дорожка 16). Экстракты случайно выбранных зерен M3 и М4, которые не были подвергнуты процедуре отбора LOX, разделяли в дорожках 2-5 и 8-12, соответственно; все из этих экстрактов содержали иммунореактивный белок LOX-1. Нуль-LOX-1-фенотип экстракта зерен мутанта А618, генерации М3 (дорожка 7) наследовался в отдельных линиях потомства М4 мутанта А618-82 (дорожки 8-12). Указано положение иммунореактивного белка LOX-1. Фиг. 9 иллюстрирует в виде схемы генетику программы обратного скрещивания для мутанта D112 сорта Prestige. Признак LOX-1 дикого типа обозначен NN, тогда как признак нуль-LOX-1-мутанта обозначен nn. Растения, имеющие подчеркнутые фенотипы, подвергают скрещиваниям. Фиг. 10 иллюстрирует семь отдельных иммуноблотов, каждый из которых зондирован антителом к LOX-1 ячменя. Эти иммуноблоты показывают присутствие или отсутствие иммунореактивного белка LOX-1 в зернах отдельных растений первой генерации обратного скрещивания мутанта D112 с сортом Prestige (дорожки 1-6 и дорожки 9-14), и присутствие или отсутствие иммунореактивного белка LOX-1 в зернах второй генерации обратного скрещивания мутанта D112 с сортом Prestige (дорожки 17-22, дорожки 25-30, дорожки 33-38, дорожки 41-45 и дорожки 48-52). Контрольные экстракты зерен мутанта D112, лишенного иммунореактивного LOX-1 (дорожки 7, 15, 23, 31, 39, 46, 53), и сорта Prestige, содержащего иммунореактивный LOX-1 (дорожки 8, 16, 24, 32, 40, 47, 54), использовали в качестве контролей. Указано положение иммунореактивного белка LOX-1. Фиг. 11 является упрощенным схематическим видом процесса производства пива без применения добавок, но включающего замачивание зерна ячменя (1), получение солода (2), печную сушку (3), размол высушенного солода (4), затирание (5), фильтрование (6), кипячение сусла в присутствии добавленного хмеля (7), ферментацию в присутствии дрожжей (8), созревание пива (9), фильтрование пива (10), упаковку, включающую - но не ограничивающуюся ими - упаковку в бутылки, банки и т.п. (11) и этикетирование (прикрепление этикетки) (12). Отдельные процессы могут быть сгруппированы в секции, включающие получение солода (1-3), получение сусла (4-7), ферментацию (8-9) и получение готового к употреблению пива (10-12). Фиг. 12 сконцентрирована на характеристиках пива, приготовленного с использованием солода, полученного из ячменя нуль-LOX-1-мутанта D112. Фиг. 12А иллюстрирует накопление свободного T2N во время форсированного созревания в течение 4 недель при 37 °С. Альдегид измеряли в пиве, полученном из солода нуль-LOX-1-мутанта D112 Фиг. 13 изображает хроматограммы ВЭЖХ-анализов, используемых для анализа на образование 9- и 13-HPODE в тканях ячменя. Уровни HPODE анализировали измерением оптической плотности при 234 нм, причем эти результаты приводятся в миллиединицах оптической плотности (mAU). Пики профилей элюции, которые соответствуют 9-HPODE и 13-HPODE, указаны стрелками. Фиг. 13А показывает хроматограмму стандартов 9-HPODE и 13-HPODE. Фиг. 13В является хроматограммой HPODE, образованных в экстрактах, полученных из зрелых зародышей сорта Barke. Фиг. 13С является хроматограммой HPODE, образованных в экстрактах, полученных из зрелых зародышей зерен с низкой LOX. Фиг. 13D является хроматограммой HPODE, образованных в экстрактах зрелых зародышей нуль-LOX-1-мутанта D112. Фиг. 14 изображает хроматограммы ВЭЖХ-анализов, используемых для анализа на образование 9- и 13-HPODE в солоде. Уровни HPODE анализировали измерением оптической плотности при 234 нм, причем эти результаты приводятся в миллиединицах оптической плотности (mAU). Пики профилей элюции, которые соответствуют 9-HPODE и 13-HPODE, указаны стрелками. Фиг. 14А показывает хроматограмму стандартов 9-HPODE и 13-HPODE. Пики хроматограммы, соответствующие 9-HPODE и 13-HPODE, указаны стрелками. Фиг. 14В является хроматограммой HPODE, образованных в экстрактах солода из сорта Barke. Фиг. 14С является хроматограммой HPODE, образованных в экстрактах солода из ячменя с низкой LOX. Фиг. 14D является хроматограммой HPODE, образованных в экстрактах солода из нуль-LOX-1-мутанта D112. Фиг. 15 является картой, показывающей организацию гена LOX-1 ячменя, охватывающего стартовый кодон (ATG) и стоп-кодон (ТАА). Этот схематический рисунок последовательности длиной 4165 п.н. показывает 7 экзонов (черные блоки) и 6 интронов (линии). Положение мутаций, идентифицированных в гене LOX-1, т.е. специфических для мутантной линии G (низкая LOX), мутанта А618 и мутанта D112, указаны стрелками. Фиг. 16 суммирует предсказанные молекулярные различия, связанные с геном LOX-1 растений ячменя дикого типа, мутанта А618 и мутанта D112. Эта информация перечислена в столбцах, обозначенных "Результат", "Длина в аминокислотах" и "Масса в кДа", предсказанных из ДНК-последовательности. Фиг. 17 представляет пути, использованные для выполнения анализа мутантов при помощи ОТ-ПЦР, и подтверждение транскриптов, связанных с геном ячменя, кодирующим LOX-1. В А схематически показан принцип ОТ-ПЦР-детектирования специфического транскрипта для гена, кодирующего LOX-1, в развивающихся зародышах сорта Vintage и мутантной линии G с низкой LOX-1. Праймеры состояли из FL821 [SEQ ID NO:11] и FL852 [SEQ ID NO:12], которые отжигаются в экзонах, фланкирующих интрон 5 83 п.н.; указаны различия продукта ПЦР с использованием в качестве матриц геномной ДНК или мРНК. В В показан результат анализа ОТ-ПЦР в агарозном геле, где внимание было сфокусировано на детектировании специфического транскрипта, связанного с геном, кодирующим LOX-1, в развивающихся зародышах ячменя, сорта Vintage и мутантной линии G. Дорожки 1 и 5 содержали маркерные фрагменты, а дорожки 2, 3 и 4 содержали продукты ПЦР, полученные из тканей зародышей сорта Vintage после 20, 40 и 60 дней после зацветания (DAF), соответственно. Дорожки 6, 7 и 8 содержат продукты, полученные из тканей зародышей мутантной линии G после 20, 40 и 60 дней после зацветания (DAF), соответственно. В С дорожки 1-5 показывают результат эксперимента, сходного с экспериментом, детально описанным для дорожек 1-5 в В, тогда как дорожки 6, 7 и 8 содержали продукты, полученные из ОТ-ПЦР-детектирования транскрипта, специфического для зародышей мутанта D112, гена LOX-1 после 20, 40 и 60 DAF, соответственно. В D показана электроферограмма, которая была получена из реакции секвенирования ОТ-ПЦР-фрагмента, специфического для гена LOX-1. Анализ последовательности выявил, что РНК-мишень ОТ-ПЦР не содержала ДНК. Черный треугольник указывает на точку сплайсинга, свидетельствующую о правильном сплайсинге этого транскрипта. Фиг. 18 детализирует результаты детектирования с использованием SNP мутанта D112 ячменя. Этот анализ основан на генерировании специфической картины ПЦР-фрагментов с использованием двух наборов ПЦР-реакций на пробу, как схематически иллюстрировано в А (набор праймеров 1 состоит из праймера FL820 [SEQ ID NO:13] и праймера FL823 [SEQ ID NO:15], а набор праймеров 2 состоит из праймера FL820 [SEQ ID NO:13] и праймера FL825 [SEQ ID NO:14]. В В показан результат анализа ПЦР-картины элитного селекционного материала. Геномную ДНК растений подвергали ПЦР-анализам. Результаты показаны в дорожках 2-3 (растение 1), 4-5 (растение 2), 6-7 (растение 3), 8-9 (растение 4), 10-11 (растение 5), 12-13 (растение 6), 14-15 (растение 7), 16-17 (растение 8) и 18-19 (растение 9), с использованием комбинации 1 праймеров (дорожки с четными номерами) или комбинацию праймеров 2 (дорожки с нечетными номерами). Сравнение картины распределения полос (бэндинга) с картиной, показанной в А, выявил, что растения 1, 2, 4, 5, 7 и 8 были гомозиготными мутантами, тогда как генотип растений 3, 6 и 9 мог быть классифицирован как гомозиготный дикий тип. Маркерную ДНК разделяли в дорожках 1 и 20. Фиг. 19 демонстрирует принцип многократного анализа SNP проб ячменя, содержащих материал мутанта G или мутанта D112. Этот анализ использовал многократные ПЦР-реакции, так что длина амплифицированного фрагмента могла быть связана с генотипом добавленного материала. Амплификация фрагмента 370 п.н. указывает на то, что проба солода содержала материал, полученный из мутантной линии G, тогда как амплификация фрагмента 166 п.н. указывает на присутствие материала, полученного из мутанта D112. Панель А является схематической иллюстрацией, детализирующей специфические пары праймеров, каждая из которых имеет один праймер, который содержит последовательность, которая является специфической для представляющего интерес мутанта (звездочка; для нуклеотида с номером 2279 мутантной линии G в геномном клоне для LOX-1, и для положения 3574 мутанта D112). Комбинацию праймеров FL918 [SEQ ID NO:16] и FL920 [SEQ ID NO:17] использовали для детектирования мутации, специфической для мутантной линии G, тогда как FL820 [SEQ ID NO:13] и FL823 [SEQ ID NO:15] использовали для детектирования изменения основания, специфического для мутанта D112. В В показано, как относительные количества мутант-специфического материала (дорожки 2-7: мутантная линия G; дорожки 8-13: мутант D112) в пробах могут усиливать синтез специфического ПЦР-фрагмента (дорожки 2 и 8: без добавленного материала мутанта; дорожки 3 и 9: 20% добавленного материала мутанта; дорожки 4 и 10: 40% добавленного материала мутанта; дорожки 5 и 11: 60% добавленного материала мутанта; дорожки 6 и 12: 80% добавленного материала мутанта; дорожки 7 и 13: 100% материала мутанта). Дорожка 1 состояла из маркерных фрагментов. Фиг. 20 представляет результат электрофореза в ДСН-ПААГ аффинно очищенного, His-меченого фермента LOX-1 из клеток Е. coli, трансформированных векторной плазмидой pET19b (дорожки 2-5), экспрессионной плазмидой pETL1 (дорожки 6-10) и экспрессионной плазмидой pETL2 (дорожки 11-15). Анализировали белки из фракций, содержащих несвязанные белки (дорожки 2, 6, 11); первой промывки (дорожки 3, 7, 12); второй промывки (дорожки 4, 8, 13); первого элюирования (дорожки 5, 9, 14); и второго элюирования (дорожки 10 и 15). Верхняя стрелка указывает положение рекомбинантого LOX-1 (соответствующего LOX-1 дикого типа), тогда как нижняя стрелка указывает положение укороченного, рекомбинантного LOX-1 (соответствующего LOX-1 в мутанте ячменя D112). Дорожка 1 содержала разделенные маркерные белки. Фиг. 21 иллюcтрирует инсерты плазмид для трансформации ячменя. В А иллюстсрирована экспрессионная кассета, состоящая из промотора убиквитина-1 и интрона 1 кукурузы (вместе обозначаемых как промотор UBI), регулирующих конститутивную экспрессию гена bar (BAR), который кодирует селектируемую маркерную фосфинотрицинацетилтрансферазу. Терминация транскрипции обеспечивается терминаторной последовательностью NOS (N). В В иллюстрирована экспрессионная кассета, состоящая из вышеупомянутого промотора UBI, регулирующего здесь конститутивную экспрессию кДНК-последовательности ячменя для LOX-1 в смысловой или антисмысловой ориентации. В С иллюстрирована экспрессионная кассета, состоящая из промотора UBI, регулирующего конститутивную экспрессию интронсодержащей шпилечной конструкции, где последовательность интрона 1 (Int) гена десатуразы жирных кислот FAD2 Arabidopsis фланкирована смысловым плечом ( →) и антисмысловым плечом ( ←) фрагмента приблизительно 200 п.н. гена LOX-1. Терминация транскрипции обеспечивается терминаторной последовательностью NOS (N). Для генерирования растений ячменя, проявляющих косупрессию гена LOX-1, использовали смеси плазмид, которые содержат равные количества экспрессионных плазмид, содержащих инсерты, подробно описанные в А и В. Для генерирования растений ячменя, проявляющих полный сайленсинг гена LOX-1, использовали смеси, которые содержат равные количества экспрессионных плазмид, содержащих инсерты, описанные в А и С. Фиг. 22 детализирует экспериментальные результаты, касающиеся ингибиторов, которые уменьшают активность LOX-1. В А изображено электрофоретическое разделение белков в 10% электрофорезе в ДСН-ПААГ, причем отдельные дорожки иллюстрируют результат ступенчатой очистки His-меченого LOX-1 из клеток Е. coli (сравните пример 18). Белки в неочищенных экстрактах трансформантов с векторной pET19b и плазмидой pETL1 показаны в дорожке 1 и дорожке 2, соответственно, тогда как дорожки 3-5 содержат разделенные белки промывных растворов 2, 3 и 4. Аликвоты проб 3 мкл из 1 мл элюатов аффинной колонки разделяли в дорожке 6 (элюат 1), дорожке 7 (элюат 2), дорожке 8 (элюат 3) и дорожке 9 (элюат А). Горизонтальная стрелка указывает положение рекомбинантного LOX-1. Аликвоты LOX-1 из элюата 2 использовали для ингибиторных исследований, как суммировано в В. Здесь, остаточную активность LOX-1 измеряли после инкубирования с 5 мкл LOX-1 (элюата 2) в присутствии ингибитора, либо NDGA Фиг. 23 представляет суммарные данные, детализирующие уровни T2N в пробах сусла, полученного из затираний без добавленного ингибитора (белые столбцы) или в присутствии 0,5 мМ ингибитора LOX-1 октилгаллата (черные столбцы). Пробы, взятые после затирания (при 37 °С) или после кипячения (прокипяченое сусло), содержали сусло затираний с солодом ячменя сорта Barke или нуль-LOX-1-мутанта ячменя D112. Для целей ясности описания, и не в качестве ограничения, подробное описание данного изобретения подразделено на следующие подразделы. (i) Растение ячменя. (ii) Получение нуль-LOX-1-ячменя. (iii) Композиция. (iv) Химический мутагенез. (v) Отбор мутантов ячменя. (vi) Селекция растений. (vii) Скрещивания ячменя. (viii) Ферменты LOX. (ix) Продукты LOX-пути. (х) T2N-потенциал. (xi) Устойчивость к болезням. (xii) Микотоксины. (xiii) Ароматические (вкусовые) вещества. (xiv) Гетерологичная экспрессия генов, кодирующих LOX. (xv) Ингибиторы LOX. 6.1. Растение ячменя. "Дикий ячмень", Hordeum vulgare ssp. spontaneum, считается прародителем культивируемых в настоящее время форм ячменя. Давно было признано, что эксплуатация этого зернового растения обеспечивает ключ к объяснению начала культивирования зерновых культур в Fertile Crescent. Тот факт, что люди могли собирать это зерно на протяжении продолжительного летнего сезона, сделали его предварительно адаптированным кандидатом для окультуривания. Ранние окультуренные растения были, вероятно, генетически очень различающимися, что подтверждается исследованием дикого ячменя из Израиля, Турции и Ирана (Nevo, 1992). Было обнаружено, что популяции дикого ячменя значительно отличаются в их аллельном содержании. Было обнаружено, что из 127 аллелей при 27 общих локусах 65 аллелей являются уникальными, т.е. они встречаются только в одной стране. Считалось, что переход ячменя из дикого в культивированное состояние совпал с радикальным изменением частоты аллелей в многочисленных локусах. Редкие аллели и новые мутационные события положительно отбирались фермерами, которые быстро установили новые признаки в окультуренных популяциях растений, названных "местными сортами (ландрасами) ячменя". Они являются генетически более близкородственными относительно современных сортов, чем дикий ячмень, и представляют источник применимых аллелей для дальнейших попыток селекции (Ellis et al., 1998). До позднего девятнадцатого столетия местные сорта ячменя существовали в виде высоко гетерогенных смесей инбредных линий и гибридных сегрегатов, в том числе нескольких растений, полученных случайными скрещиваниями в более ранних генерациях. Большинство местных сортов были вытеснены в передовых агрономических хозяйствах сортами чистых линий. Промежуточные или высокие уровни генетического разнообразия характеризуют оставшиеся местные сорта (ландрасы). Сначала сорта "современного ячменя" представляли сорта, полученные селекцией из местных сортов. Позднее их получали из последовательных циклов скрещиваний между установленными чистыми линиями, такими как линии различного географического происхождения. Со временем это приводило к заметному сужению генетической основы во многих, возможно, во всех передовых агрономических хозяйствах. В сравнении с местными сортами, современные сорта ячменя имеют многочисленные улучшенные свойства, (Nevo, 1992 и von Bothmer et al., 2003), например (без ограничения): (i) Покрытые и голые зерна; (ii) Покой семян; (iii) Устойчивость к болезням; (iv) Доли лизина и других аминокислот; (v) Содержание белка; (vi) Содержание азота; (vii) Состав углеводов; (viii) Распределения гордеина. Таким образом, в одном варианте осуществления данного изобретения растение ячменя является современным сортом ячменя, модифицированным таким образом, что оно содержит менее 1% активности LOX-1 соответствующего растения ячменя дикого типа. Таким образом, в этом варианте осуществления растение ячменя предпочтительно не является местным (примитивным) сортом ячменя. Данное изобретение относится к растению ячменя и его частям, содержащим менее 5%, предпочтительно менее 4%, более предпочтительно менее 3%, даже более предпочтительно менее 2%, еще более предпочтительно менее 1% активности LOX-1 растения ячменя дикого типа. Растения ячменя данного изобретения, содержащие менее 1% активности LOX-1, называют здесь также "растениями нуль-LOX-1-ячменя". Растение ячменя может находиться в любой подходящей форме. Например, растение ячменя в соответствии с данным изобретением может быть жизнеспособным растением ячменя, высушенным растением, гомогенизированным растением, таким как размолотые зерна ячменя. Это растение может быть зрелым растением, зародышем, проросшим зерном, осоложенным зерном или т.п. Части растений ячменя могут быть любой подходящей частью растения, такой как зерна, зародыши, листья стебли, корни, цветки или их фракции. Фракции могут быть, например, частью зерна, зародыша, листьев, стебля, корня или цветка. Части растения ячменя могут быть также фракцией гомогената или размолотого растения ячменя или зерна. В одном варианте осуществления данного изобретения, частями растений ячменя могут быть клетки указанного растения ячменя, предпочтительно жизнеспособные клетки, которые могут быть размножены в культурах тканей in vitro. В предпочтительном варианте осуществления данного изобретения, растения нуль-LOX-1-ячменя содержат менее 5%, предпочтительно менее 3%, более предпочтительно менее 1%, предпочтительно менее, 0,5%, даже более предпочтительно менее 0,1% активности растения ячменя дикого типа. Эта активность может быть определена любым подходящим способом, но предпочтительно эту активность определяют с использованием способа примера 1, описанного здесь ниже. В очень предпочтительном варианте осуществления данного изобретения, растения нуль-LOX-1-ячменя, по существу, не имеют активности LOX-1, более предпочтительно вообще не имеют активности LOX-1. "По существу не имеющие активности LOX-1" означает отсутствие детектируемой активности при использовании анализа на активность LOX-1, описанного здесь ниже. Почти отсутствующая активность LOX-1 нуль-LOX-1-ячменя может быть, например, результатом того, что указанный ячмень содержит белок LOX-1 с нарушенной функцией, такой, как мутантный белок LOX-1. Однако нуль-LOX-1-ячмень содержит лишь малое количество белка LOX-1 или, более предпочтительно, не содержит белка LOX-1, например, содержит менее 5%, предпочтительно менее 3%, более предпочтительно менее 1%, предпочтительно менее 0,5%, более предпочтительно менее 0,1% белка LOX-1 в сравнении с растением ячменя дикого типа. Более предпочтительно, нуль-LOX-1-ячмень, по существу, не содержит белка LOX-1, наиболее предпочтительно вообще не содержит белка LOX-1. "По существу, не содержит белка LOX-1" означает отсутствие детектируемого белка LOX-1. Белок LOX-1 может быть детектирован любыми подходящими средствами, известными специалисту в данной области, однако, предпочтительно этот белок детектируют способами, в которых белок LOX-1 детектируют специфическими антителами к LOX-1. Указанными способами могут быть, например, Вестерн-блоттинг или ELISA. Указанные специфические антитела могут быть моноклональными или поликлональными, но предпочтительно указанные антитела являются поликлональными, узнающими несколько различных эпитопов в белке LOX-1. Белок LOX-1 может быть также детектирован опосредованно, например, способами, определяющими активность LOX-1, способами, детектирующими мутации в гене, кодирующем LOX-1, или способами, детектирующими экспрессию гена LOX-1. Мутации в гене, кодирующем LOX-1, могут быть, например, детектированы секвенированием указанного гена. Экспрессия гена LOX-1 может быть, например, детектирована Нозерн-блоттингом или ОТ-ПЦР. В одном предпочтительном варианте осуществления данного изобретения, белок LOX-1 детектируют с использованием способов, описанных в примере 4 данной публикации. Термин белок LOX-1 включает полноразмерный белок LOX-1 ячменя, представленный в [SEQ ID NO:3] или [SEQ ID NO:7], или его функциональный гомолог. Активный центр LOX-1 расположен в С-концевой части LOX-1. В частности, район, охватывающий аминокислотные остатки 520-862 этого белка, является важным для активности LOX-1. Таким образом, в одном варианте осуществления нуль-LOX-1-ячмень предпочтительно содержит ген, кодирующий мутантную форму LOX-1, лишенную нескольких аминокислот или всех аминокислот 520-862 LOX-1. Указанный мутантный LOX-1 может быть также лишен других аминокислотных остатков, которые присутствуют в LOX-1 дикого типа. Таким образом, нуль-LOX-1-ячмень может содержать укороченную форму LOX-1, которая не является функциональной, такую как укороченная на N-конце или на С-конце форма. Предпочтительно указанная укороченная форма содержит не более чем 800, более предпочтительно не более чем 750, даже более предпочтительно не более чем 700, еще более предпочтительно не более чем 690, даже еще более предпочтительно не более чем 680 и еще более предпочтительно не более чем 670 последовательных аминокислот LOX-1, например, не более чем 665, например, не более чем 650, например, не более чем 600, например, не более чем 550, например, не более чем 500, например, не более чем 450, например, не более чем 425, например, не более чем 399 последовательных аминокислот LOX-1 [SEQ ID NO:3]. Предпочтительно, указанная укороченная форма содержит только укороченный на N-конце фрагмент LOX-1. Таким образом, предпочтительно указанная укороченная форма содержит максимально 800, более предпочтительно максимально 750, даже более предпочтительно максимально 700 и еще более предпочтительно максимально 690, даже более предпочтительно максимально 680, еще более предпочтительно максимально 670, даже более предпочтительно максимально 665 N-концевых аминокислот [SEQ ID NO:3], например, не более чем 665, например, не более чем 650, например, не более чем 600, например, не более чем 550, например, максимально 425, например, максимально 399 N-концевых аминокислот [SEQ ID NO:3]. В одном очень предпочтительном варианте осуществления эта укороченная форма состоит из аминокислот 1-665 [SEQ ID NO:3]. В предпочтительном варианте осуществления данного изобретения растение ячменя содержит ген, транскрибируемый в мРНК, кодирующую LOX-1, где указанная мРНК содержит терминирующий нонсенс-кодон или стоп-кодон слева от стоп-кодона мРНК LOX-1 дикого типа. Такой терминирующий нонсенс-кодон обозначается здесь как преждевременный терминирующий кодон. Предпочтительно, все гены, транскрибируемые в мРНК, кодирующую LOX-1 указанного растения, содержат преждевременный терминирующий кодон, расположенный при максимально 800, более предпочтительно максимально 750, даже более предпочтительно максимально 700, еще более предпочтительно максимально 690, даже более предпочтительно максимально 680, еще более предпочтительно максимально 670, даже более предпочтительно максимально 665 кодонах справа от инициирующего (стартового) кодона. Последовательность геномной ДНК дикого типа, кодирующей LOX-1, приведена в [SEQ ID NO:1] или [SEQ ID NO:5]. В одном предпочтительном варианте растение ячменя содержит ген, кодирующий LOX-1, где пре-мРНК транскрибированная из указанного гена, содержит рибонуклеиновую кислотную последовательность соответствующую [SEQ ID NO:2]. В другом предпочтительном варианте осуществления данного изобретения растение ячменя содержит ген, кодирующий мутантный LOX-1, причем указанный ген содержит по меньшей мере одну, например, 1, например, 2, например, 3, например, 4, например, 5 мутаций по меньшей мере в одном, например, 1, например, 2, например, 3 сайтах сплайсинга. Предпочтительно, указанная мутация (указанные мутации) приводит (приводят) к тому, что указанный по меньшей мере один сайт сплайсинга является нефункциональным. мРНК, транскрибированная из такого гена, будет, следовательно, аномальной вследствие отклоняющегося от нормы сплайсинга. Таким образом, предпочтительно, мРНК, транскрибированная из гена LOX-1 растения нуль-LOX-1-ячменя в соответствии с данным изобретением, не кодирует белок или кодирует белок, содержащий только N-конец LOX-1. Указанный белок может содержать другие последовательности, кодируемые аномальной мРНК, которые не происходят из гена LOX-1. В этом контексте, N-конец LOX-1 содержит аминокислоту 1 к аминокислоте N, где N является целым числом в диапазоне 3-800, более предпочтительно в диапазоне 2-750, еще более предпочтительно в диапазоне 2-700, даже более предпочтительно в диапазоне 2-650, еще более предпочтительно в диапазоне 2-600, даже более предпочтительно в диапазоне 2-550, еще более предпочтительно в диапазоне 2-500, еще более предпочтительно в диапазоне 2-450, даже более предпочтительно в диапазоне 2-400, еще более предпочтительно в диапазоне 2-378. В одном варианте осуществления данного изобретения растение ячменя содержит ген, кодирующий мутантный LOX-1, причем указанный ген имеет мутацию в сайте сплайсинга, приводящую к мРНК, кодирующей белок, состоящий из аминокислот 1-378 [SEQ ID NO:3], а также дополнительную аминокислотную последовательность, не происходящую из LOX-1. Предпочтительно, указанный мутантный LOX-1 состоит из последовательности, описанной в [SEQ ID NO:8]. В одном очень предпочтительном варианте осуществления данного изобретения ген, кодирующий мутантный LOX-1 растения нуль-LOX-1-ячменя содержит нонсенс-мутацию, причем указанная мутация соответствует замене G →А в положении 3574 [SEQ ID NO:1]. Более предпочтительно, растение нуль-LOX-1-ячменя является растением, обозначенным D112, имеющим номер депозита Американской Коллекции Типовых Культур (АТСС) РТА-5487. В другом очень предпочтительном варианте осуществления данного изобретения ген, кодирующий LOX-1 растения нуль-LOX-1-ячменя, содержит нефункциональный сайт сплайсинга интрона 3. Таким образом, мРНК LOX-1 указанного растения кодирует белок, содержащий аминокислоты 1-378 LOX-1 и дополнительные аминокислоты из интрона 3, содержащиеся в [SEQ ID NO:8]. Более предпочтительно, растение нуль-LOX-1-ячменя является растением, обозначенным А618, имеющим номер депозита Американской Коллекции Типовых Культур (АТСС) РТА-5584. Растения ячменя данного изобретения могут быть также потомством растения нуль-LOX-1-ячменя. Таким образом, это растение ячменя может быть потомком растения, обозначенного D112, имеющего номер депозита Американской Коллекции Типовых Культур (АТСС) РТА-5487, или растения, обозначенного А618, имеющего номер депозита Американской Коллекции Типовых Культур (АТСС) РТА-5584. Растение ячменя по данному изобретению может быть получено любым подходящим способом, известным специалисту в данной области, предпочтительно одним из способов, описанных здесь ниже (см., например, Раздел 6.2 "Получение нуль-LOX-1-ячменя"). В одном варианте осуществления данного изобретения, предпочтительно, растение нуль-LOX-1-ячменя в соответствии с данным изобретением имеет физиологию роста растения и развития семян, сравнимую с ячменем дикого типа. Таким образом, предпочтительно, растение нуль-LOX-1-ячменя является сходным с ячменем дикого типа в отношении высоты растения, количества побегов на одно растение, начала цветения и/или количества зерен на один колос. Один из аспектов данного изобретения обеспечивает растение нуль-LOX-1-ячменя, причем указанное растение характеризуется: (i) наличием повышенной устойчивости к болезням; или (ii) наличием пониженного потенциала в отношении продуцирования микотоксинов; или (iii) содержанием регенерируемых клеток для применения в культуре ткани; или (iv) любой комбинацией признаков (i)-(iii). В одном варианте осуществления данного изобретения это растение является растением нуль-LOX-1-ячменя, при условии, что указанное растение ячменя не несет мутацию G в донорном сайте сплайсинга интрона 5. В этом варианте осуществления данное изобретение относится также к растительным продуктам, таким как солод, сусло, ферментированные или неферментированные напитки, пиво, пищевые и кормовые продукты, приготовленные из растения нуль-LOX-1-ячменя или его части, при условии, что указанное растение ячменя не несет мутацию G в донорном сайте сплайсинга интрона 5. Указанный G соответствует, например, G в положении 2968 SEQ ID NO:1. Получен ли растительный продукт из растения ячменя с указанной мутацией, может быть определено выделением ДНК из указанного растительного продукта и идентификацией присутствия или отсутствия указанной мутации традиционными способами, известными специалисту в данной области. ДНК может быть, например, выделена из сусла, пива или другого напитка лиофилизацией, ресуспендированием в водном буфере, экстракцией смесью хлороформ/изоамиловый спирт с последующим осаждением спиртом. Например, мутация G в донорном сайте сплайсинга интрона 5 может быть идентифицирована способом, сходным со способом, описанным в WO2004/085652, Hirota et al. 6.2. Получение нуль-LOX-1-ячменя. Растение ячменя в соответствии с данным изобретением может быть получено любым подходящим способом, известным специалисту в данной области. Предпочтительно растения ячменя данного изобретения получают способом, предусматривающим стадии мутагенеза растений ячменя или их частей, например, зерен ячменя, с последующим скринингом и отбором растений ячменя в отношении растений с меньшей, чем 5%, активностью LOX-1. Представляет интерес то, что данное изобретение в одном аспекте относится к новому и очень эффективному способу скрининга, значительно превосходящему способ скрининга, описанный, например, в WO 02/053721, Douma et al. Этот новый способ скрининга позволяет воспроизводимо идентифицировать растения ячменя без активности LOX-1 или с очень малой активностью LOX-1. Этот новый способ скрининга включает получение зерен или их частей, таких как зародыши, из мутагенизированного ячменя и определение активности LOX-1 в указанных зернах или их частях. Таким образом, целью данного изобретения является обеспечение способов получения растения ячменя, содержащего менее чем 5%, активности LOX-1 растения ячменя дикого типа, предусматривающих стадии: (i) определения активности LOX-1 в зернах ячменя дикого типа или их частях; и (ii) мутагенеза растений ячменя, и/или клеток ячменя, и/или ткани ячменя, и/или зерен ячменя и/или зародышей ячменя с получением посредством этого генерации МО ячменя; и (iii) отбора указанных мутагенизированных растений, зерен и/или зародышей ячменя в течение по меньшей мере 2 генераций с получением посредством этого генерации Мх растений ячменя, где х обозначает целое число ≥2; и (iv) получения зерен или их частей из указанных растений ячменя Мх; и (v) определения активности LOX-1 в указанных зернах или их частях; и (vi) отбора растений, в которых активность LOX-1 мутагенезированных зерен или их частей равна менее чем 5% активности LOX-1 зерен дикого типа или их части; с получением посредством этого растения ячменя, содержащего менее чем 5% активности LOX-1 растения ячменя дикого типа. Стадия (ii) в приведенном выше перечне может включать мутагенез живого материала, выбранного из группы, состоящей из растения ячменя, клеток ячменя, ткани ячменя, зерен ячменя и зародышей ячменя, предпочтительно выбранных из группы, состоящей из растений ячменя, зерен ячменя и зародышей ячменя, более предпочтительно зерен ячменя. Предпочтительно, активность LOX-1 мутагенизированных зерен определяют с использованием того же самого типа материала, который использовали для определения активности LOX-1 зерен ячменя дикого типа, т.е. предпочтительно, зерна ячменя или их части стадии (i) являются зернами ячменя или их частями того же типа, что и упомянутые в стадии (iv). Например, если активность LOX-1 ячменя дикого типа определяют в зародышах ячменя дикого типа, предпочтительно, стадия (iv) предусматривает определение активности LOX-1 в зародышах мутагенизированных растений ячменя. Мутагенез может быть выполнен любым подходящим способом. В одном варианте осуществления, мутагенез выполняют инкубированием растения ячменя или его части, например, зерен ячменя или индивидуальных клеток из ячменя, с мутагенизирующим агентом. Указанный агент известен специалисту в данной области, например, им может быть, но не только, азид натрия (NaN 3 ), этилметансульфонат (EMS), азидоглицерин (AG, 3-азидо-1,2-пропандиол), метилнитрозомочевина (MNU) и малеиновый гидразид (МН). В другом варианте осуществления мутагенез может быть выполнен облучением, например УФ, растения ячменя или его части, такой как зерно. В предпочтительных вариантах осуществления данного изобретения, мутагенез выполняют в соответствии с любым из способов, описанных здесь ниже в Разделе 6.4 "Химический мутагенез". Неограничивающий пример подходящего протокола мутагенеза приведен в примере 1. Предпочтительно мутагенез выполняют таким образом, что ожидаемая частота желаемых мутантов равна по меньшей мере 0,5, например, в диапазоне 0,5-5, например, в диапазоне 0,9-2,3 на 10000 зерен, при скрининге генерации М3 ячменя. В предпочтительном варианте осуществления, мутагенез выполняют на зернах ячменя. Мутагенизированные зерна называют генерацией МО (см. также фиг. 1А). После мутагенеза отбирают растения ячменя или их части, которые содержат менее чем 5%, предпочтительно менее чем 1% активности LOX-1. Отбор может выполняться в соответствии с любым подходящим способом, известным специалисту в данной области. Предпочтительно отбор предусматривает получение пробы из растения ячменя, например, из зерна ячменя, определение активности LOX-1 в указанной пробе и отбор растений, в которых указанная проба имеет менее чем 5% или предпочтительно менее чем 1% активности LOX-1 растений ячменя дикого типа. Эта проба может быть взята из любой подходящей части указанного растения. Однако предпочтительно эту пробу берут из зерна, более предпочтительно эту пробу берут из ткани зародыша зерна, еще более предпочтительно эта проба состоит из ткани зародыша зерна. Обычно эта проба должна быть гомогенизирована с использованием любого подходящего способа перед определением активности LOX-1. В предпочтительном варианте осуществления эту пробу берут из зерен генерации Мх, где х обозначает целое число ≥2, предпочтительно х обозначает целое число в диапазоне 2-10, более предпочтительно в диапазоне 3-8. В очень предпочтительном варианте осуществления, активность LOX-1 определяют на зернах МЗ или пробе, полученной из зерен. В этом варианте осуществления предпочтительно выращивать мутагенизированные зерна ячменя (генерации МО) для получения растений ячменя, которые скрещивают для получения зерен M1. Эту процедуру повторяют, пока не будут доступны зерна М3 (см. также фиг. 1А). Определение активности LOX-1 может проводиться с использованием любого подходящего анализа, предпочтительно одного из способов, описанных здесь ниже. В частности, предпочтительно этот анализ отслеживает диоксигенирование линолевой кислоты до 9-HPODE посредством LOX-1. Таким образом, обычно этот анализ будет предусматривать стадии: (i) обеспечения пробы, полученной из зерна ячменя или его части; и (ii) обеспечения линолевой кислоты; и (iii) инкубирования указанной пробы с указанной линолевой кислотой; и (iv) детектирования диоксигенирования линолевой кислоты до 9-HPODE. Детектирование может выполняться непосредственно или опосредованно. Любой подходящий способ может быть использован с данным изобретением. В одном варианте осуществления данного изобретения детектируют гидропероксиды линолевой кислоты. Гидропероксиды линолевой кислоты могут быть детектированы сопряжением деградации указанных гидропероксидов линолевой кислоты с окислительной реакцией, которая развивает детектируемое соединение. Например, это может быть выполнено, как описано в примере 1. В другом варианте осуществления 9-HPODE детектируют непосредственно, например, спектрофотометрическими способами, такими как ВЭЖХ, как описано в примере 9. В одном варианте осуществления данного изобретения активность LOX-1 определяют просто определением количества 9-HPODE в пробе из зерна ячменя. Это может быть выполнено любым подходящим способом, известным специалисту в данной области, например, как описано в примере 9. Важно, при каком рН выполняют определение активности LOX-1. Предпочтительно указанное определение выполняют при рН, который делает возможной высокую активность LOX-1, но только низкую активность LOX-2. Таким образом, определение активности LOX предпочтительно выполняют при рН в диапазоне 3-6,5, например, в диаразоне 3-4, например, в диапазоне 4-5, например, в диапазоне 5-6, например, в диапазоне 6-6,5. Предпочтительно рН равен приблизительно 3, например, приблизительно 3,5, например, приблизительно 4, например, приблизительно 4,5, например, приблизительно 5, например, приблизительно 5,5, например, приблизительно 6, например, приблизительно 6,5, например, приблизительно 7. Предпочтительно также, указанную пробу готовят при подходящем рН, таком как рН в диапазоне 3-6,5, например, в диапазоне 3-4, например, в диапазоне 4-5, например, в диапазоне 5-6, например, в диапазоне 6-6,5. Предпочтительно, рН равен приблизительно 3, например, приблизительно 3,5, например, приблизительно 4, например, приблизительно 4,5, например, приблизительно 5, например, приблизительно 5,5, например, приблизительно 6, например, приблизительно 6,5, например, приблизительно 7. Предпочтительные способы отбора растений ячменя в соответствии с данным изобретением описаны ниже в разделе 6.5 "Отбор мутантов ячменя". Предпочтительный пример способа определения активности LOX-1 приведен в примере 1. Процедура отбора может быть адаптирована к процедурам анализа в микротитрационных планшетах или к другим известным, повторяемым, высокопроизводительным форматам анализа, которые делают возможным быстрый скрининг многих проб. Предпочтительно по меньшей мере 5000, например, по меньшей мере 7500, например, по меньшей мере 10000, например, по меньшей мере 15000 мутагенизированных растений ячменя анализируют на активность LOX-1. После отбора пригодных растений ячменя с менее чем 5% активностью LOX-1, могут быть выполнены один или несколько дополнительных скринингов. Например, выбранные мутанты могут быть дополнительно размножены и следующие генерации могут быть опять подвергнуты скринингу на активность LOX-1. После отбора пригодных растений ячменя они могут быть подвергнуты селекции, например, традиционной селекции. Способы селекции описаны здесь ниже (Раздел 6.6 "Селекция растений" и Раздел 6.7 "Скрещивания растений"). Однако растение ячменя в соответствии с данным изобретением может быть получено другими способами, например способами, приводящими к уменьшенной транскрипции и/или трансляции LOX-1. Таким образом, может быть получено растение нуль-LOX-1-ячменя трансформацией растения ячменя последовательностью нуклеиновой кислоты, содержащей, в виде функционально связанных компонентов, промотор, экспрессируемый в растениях ячменя, ДНК-последовательность и район терминации транскрипции, причем экспрессия указанной ДНК-последовательности уменьшает экспрессию гена, кодирующего LOX-1 посредством: (i) антисмыслового сайленсинга; или (ii) сайленсинга косупрессией; или (iii) интерференции РНК. В одном варианте осуществления растение ячменя получают способом, предусматривающим трансформацию растения ячменя последовательностью нуклеиновой кислоты, способной уменьшать транскрипцию или трансляцию гена, кодирующего LOX-1, например, последовательностью нуклеиновой кислоты, содержащей антисмысловые последовательности LOX-1. Указанные последовательности могут быть, например, антисмысловой последовательностью [SEQ ID NO:1], или ее фрагментом. Эта антисмысловая последовательность должна быть функционально связана с промоторной последовательностью из гена, экспрессируемого в растениях ячменя. Неограничивающий пример такого способа описан в примере 16 здесь ниже. Растение ячменя может быть трансформировано любым применимым способом, например, опосредованным Agrobacterium tumefaciens переносом или поглощением ДНК опосредованным бомбардировкой частицами. В рамках данного изобретения находится также получение растения нуль-LOX-1-ячменя способом, предусматривающим стадии: (i) мутагенеза растений ячменя, и/или зерен ячменя, и/или зародышей ячменя; и (ii) определения присутствия или отсутствия мутации в гене для LOX-1, где указанная мутация приводит к гену для LOX-1, кодирующему полипептидную форму LOX-1, содержащую менее чем 700 смежных аминокислот последовательности, представленной в SEQ ID NO:3, предпочтительно указанный полипептид является N-концевым фрагментом LOX-1, содержащим максимально 700 N-концевых аминокислот [SEQ ID NO:3], (iii) отбора растений, несущих указанную мутацию, с получением посредством этого растения ячменя, содержащего менее чем 5% активности LOX-1 растения ячменя дикого типа. Более предпочтительно указанная мутация может приводить к гену LOX-1, кодирующему любой из описанных здесь выше N-концевых фрагментов LOX-1. Указанная мутация может быть детектирована с использованием любого подходящего способа, например секвенирования гена LOX-1 или анализа полиморфизма единичного нуклеотида (SNP). Один пример, показывающий, как может быть выполнен анализ SNP, описан в примере 13 и примере 14 здесь ниже. После получения растения нуль-LOX-1-ячменя с конкретной мутацией в гене LOX-1 (такой как любая из вышеупомянутых мутаций) могут быть генерированы дополнительные растения ячменя с той же самой мутацией с использованием традиционных способов селекции, хорошо известных специалисту в данной области. Например, указанное растение нуль-LOX-1-ячменя может быть подвергнуто обратному скрещиванию в генетический фон другого сорта ячменя. Фиг. 9 описывает пример схемы такого обратного скрещивания. 6.3. Композиция. Данное изобретение относится также к композициям, содержащим описанные выше растения ячменя или их части, или композициям, полученным из указанных растений ячменя или их частей. Поскольку указанные растения ячменя имеют менее 5%, предпочтительно менее 1% активности LOX-1, композиции, содержащие указанные растения ячменя или их части, или полученные из указанных растений ячменя или их частей, будут обычно содержать очень низкие уровни T2N и T2N-потенциала. Примеры применимых композиций, содержащих нуль-LOX-1-ячменя или полученных из нуль-LOX-1-ячменя, описаны здесь ниже. Предпочтительно, указанные композиции имеют: (i) менее чем 30%, предпочтительно менее чем 20%, более предпочтительно менее чем 10%, даже более предпочтительно менее чем 2%, например, менее чем 1% T2N; и/или (ii) менее чем 30%, предпочтительно менее чем 20%, более предпочтительно менее чем 10%, даже более предпочтительно менее чем 2%, например, менее чем 1% T2N-потенциала; в сравнении с подобной композицией, содержащей растения ячменя дикого типа или полученной из растений ячменя дикого типа. Данное изобретение относится в одном аспекте к зернам ячменя, содержащим менее чем 1% активности LOX-1, в сравнении с зернами дикого типа. Предпочтительно, эти зерна не содержат активности LOX-1. Данное изобретение относится также к композициям, содержащим указанные зерна, и композициям, полученным из указанных зерен. В одном аспекте, данное изобретение относится к солодовым композициям, полученным из нуль-LOX-1-зерен солодоращением. Под термином "солодоращение" следует понимать проращивание замоченных зерен ячменя, происходящее в контролируемых условиях окружающей среды (например, как иллюстрировано на фиг. 11, стадиях 2 и 3). Солодоращение является процессом контролируемого замачивания и проращивания с последующей сушкой зерна ячменя. Эта последовательность событий является важной для синтеза многочисленных ферментов, которые вызывают модификацию зерна, процесс, который преимущественно деполимеризует мертвые клеточные стенки эндосперма и мобилизует питательные вещества зерна. В последующем процессе сушки вкус и цвет образуются вследствие химических реакций потемнения. Хотя солод используют прежде всего для производства пива, он может быть также использован в других промышленных процессах, например в качестве источника ферментов в хлебопекарной промышленности, или в качестве вкусового и красящего агента в пищевой промышленности, например, в виде солода или в виде солодовой муки или опосредованно в виде солодового сиропа и т.д. В одном аспекте данное изобретение относится к способам получения указанной солодовой композиции. Эти способы предпочтительно предусматривают стадии: (i) обеспечения зерен нуль-LOX-1-ячменя; (ii) замачивания указанных зерен; (iii) проращивания замоченных зерен при заданных условиях; (iv) сушки указанных пророщенных зерен; с получением посредством этого солодовой композиции с низкой активностью или без активности LOX-1. Например, этот солод может быть получен любым из способов, описанных в Hoseney (1994). Однако с данным изобретением может быть также использован любой другой подходящий способ, например, способы получения специальных типов солода, в том числе способы обжаривания солода. Один неограничивающий пример описан в примере 6. В другом аспекте данное изобретение относится к композициям сусла, полученным из солодовых композиций, приготовленных из нуль-LOX-1-зерен. Указанный солод может быть приготовлен только из нуль-LOX-1-зерен или из смесей, которые содержат и другие зерна. Данное изобретение относится также к композициям сусла, полученным с использованием нуль-LOX-1-ячменя или его частей, отдельно или в виде смеси с другими компонентами. Указанное сусло может быть первым, и/или вторым, и/или последующим суслом. Обычно композиция сусла будет иметь высокое содержание аминного азота и ферментируемых углеводов, в основном мальтозы. На фиг. 11, стадии 4-6 иллюстрируют обычный способ получения сусла из солода. Обычно сусло готовят инкубированием солода с водой, т.е. затиранием. Во время затирания композиция солод/вода может быть дополнена дополнительными богатыми углеводами композициями, например, добавками ячменя, кукурузы или риса. Не подвергнутые солодоращению (несоложеные) зерновые добавки обычно не содержат активных ферментов и, следовательно, используют ферменты солода или экзогенные ферменты для обеспечения ферментов, необходимых для превращений сахаров. Обычно первой стадией в процессе производства сусла является размол солода, чтобы вода могла получить доступ к частицам зерен в фазе затирания, которая является, по существу, продлением процесса солодоращения с ферментативной деполимеризацией субстратов. Во время затирания размолотый солод инкубируют с жидкой фракцией, такой как вода. Температуру либо поддерживают на постоянном уровне (изотермическое затирание), либо постепенно увеличивают. В любом случае растворимые вещества, образуемые в солодоращении и затирании, экстрагируются в указанную жидкую фракцию перед ее разделением фильтрованием на сусло и оставшиеся твердые частицы, называемые дробиной или бардой. Это сусло может также называться первым суслом. После фильтрования получают второе сусло. Дополнительные сусла могут быть приготовлены повторением этой процедуры. Неограничивающие примеры подходящих процедур для приготовления сусла описаны в Hoseney (supra). Композиция сусла может быть также приготовлена инкубированием растений нуль-LOX-1-ячменя или их частей, таких как не подвергнутых солодоращению (несоложеные) нуль-LOX-1-растения или их части, с одним или несколькими подходящими ферментами, такими как композиции ферментов или композиции смесей ферментов, например, Ultraflo или Cereflo (Novozymes). Композиция сусла может быть также приготовлена с использованием смеси солода и не подвергнутых солодоращению (несоложеных) растений ячменя или их частей, необязательно с добавлением одного или нескольких подходящих ферментов во время указанного приготовления. Данное изобретение относится также к пищевым композициям, кормовым композициям и ароматным композициям сырьевого материала, которые содержат растения нуль-LOX-1-ячменя или их части. Пищевыми композициями могут быть, например, но не только, подвергнутые или не подвергнутые солодоращению зерна ячменя, ячменная мука, ячменный хлеб, ячменная каша, зерновые смеси, содержащие ячмень, полезные для здоровья продукты, такие как содержащие ячмень напитки, ячменные сиропы и плющеные, размолотые или экструдированные ячменные композиции. Кормовые композиции включают, например, композиции, содержащие зерна ячменя, и/или мучки. Ароматные композиции сырьевого материала описаны здесь ниже. Данное изобретение относится также к смесям композиций данного изобретения. Например, данное изобретение в одном аспекте относится к композиции, приготовленной смешиванием (i) композиции, содержащей растение ячменя или его часть, содержащие менее чем 5% активности LOX-1 растения ячменя дикого типа, и (ii) солодовой композиции, полученной из нуль-LOX-1-зерен. В предпочтительном аспекте данное изобретение относится к напиткам, более предпочтительно к полученным из солода напиткам, даже более предпочтительно к алкогольным напиткам, таким как пиво, имеющим стабильные органолептические качества, причем указанный напиток получают с использованием нуль-LOX-1-ячменя или его частей. Таким образом, в одном предпочтительном варианте осуществления данного изобретения, этот напиток получают ферментацией нуль-LOX-1-ячменя или его частей или его экстрактов, например, ферментацией сусла, приготовленного из солода, полученного из нуль-LOX-1-ячменя, отдельно или в комбинации с другими ингредиентами. Однако в других вариантах осуществления данного изобретения этот напиток является неферментированным напитком, например суслом. В данное изобретение включено также, что указанный напиток может быть получен из не подвергнутых солодоращению (несоложеных) растений ячменя или их частей. Однако предпочтительно, указанный напиток получают из солодовой композиции, приготовленной из зерен нуль-LOX-1-ячменя. Более предпочтительно указанный напиток является пивом. Он может быть любым типом пива, известным специалисту в данной области. В одном варианте осуществления это пиво является лагерным (светлым) пивом. Это пиво предпочтительно варят с использованием солодовой композиции, содержащей пророщенный нуль-LOX-1-ячмень. Однако эта солодовая композиция может также содержать другие компоненты, например, другие пророщенные или непророщенные злаки, такие как ячмень дикого типа, нуль-LOX-1-ячмень, пшеница и/или рожь, или непророщенные сырьевые материалы, содержащие сахара, или композиции, полученные из соложеных или несоложеных сырьевых материалов, например, композиции в виде сиропов. "Органолептические качества" обозначают качества, вызывающие обонятельные и вкусовые ощущения. Их анализирует, например, квалифицированная дегустационная комиссия. Предпочтительно, указанная квалифицированная дегустационная комиссия специально обучена узнавать альдегидные привкусы, такие как привкус T2N. Обычно эта дегустационная комиссия состоит из 3-30 членов, например, из 5-15 членов. Эта дегустационная комиссия может оценивать присутствие различных вкусовых качеств, таких как привкусы, такие как бумажный привкус, окисленный привкус, привкус старого пива и хлебный привкус. Способ определения "органолептических качеств" напитка описан здесь в примере 6 ниже. В предпочтительных вариантах осуществления, стабильные органолептические качества являются по меньшей мере частично результатом низкого продуцирования T2N или T2N-потенциала. Таким образом, одной целью данного изобретения является обеспечение напитков, приготовленных с использованием растения ячменя, (таких как пиво), предпочтительно содержащих менее чем 50%, предпочтительно менее чем 40%, более предпочтительно менее чем 35%, например, менее чем 30%, например, менее чем 20%, например, менее чем 10%, например, предпочтительно менее чем 5%, например, менее чем 2%, например, менее чем 1% T2N и/или T2N-потенциала, в сравнении с напитком, приготовленным из ячменя дикого типа, после хранения в течение по меньшей мере 1 недели, предпочтительно по меньшей мере 2 недель, более предпочтительно по меньшей мере 3 недель, даже более предпочтительно в течение по меньшей мере 4 недель, например, в диапазоне 1-3 месяцев, например, в диапазоне 3-6 месяцев, например, в диапазоне 6-12 месяцев, например, в течение более, чем 1 года. Хранение осуществляют при температуре в диапазоне 15-40 °С, предпочтительно в диапазоне 30-37 °С, более предпочтительно при 37 °С. Напитки данного изобретения предпочтительно содержат максимально 0,07, предпочтительно максимально 0,06, более предпочтительно максимально 0,05, даже более предпочтительно максимально 0,04, например, максимально 0,03 м.д. свободного T2N, после хранения в течение по меньшей мере 1 недели, предпочтительно по меньшей мере 2 недель, более предпочтительно по меньшей мере 3 недель, даже более предпочтительно в течение по меньшей мере 4 недель, например, в диапазоне 1-3 месяцев, например, в диапазоне 3-6 месяцев, например, в диапазоне 6-12 месяцев, например в течение более чем 1 года, после хранения при температуре в диапазоне 15-40 °С, предпочтительно в диапазоне 30-37 °С, более предпочтительно при 37 °С. Предпочтительно, этот напиток содержит также сульфит в диапазоне 1-10 м.д. (миллионных долей), более предпочтительно в диапазоне 2-8 м.д., более предпочтительно в диапазоне 3-7 м.д., еще более предпочтительно в диапазоне 4-6 м.д. сульфита. В одном предпочтительном варианте осуществления напитки в соответствии с данным изобретением содержат максимально 0,04, более предпочтительно максимально 0,03, например, максимально 0,025 м.д. свободного T2N, после хранения в течение 2 недель при 37 °С. В другом предпочтительном варианте осуществления данного изобретения напитки в соответствии с данным изобретением содержат максимально 0,07, предпочтительно максимально 0,06, более предпочтительно максимально 0,05, даже более предпочтительно максимально 0,04, например, максимально 0,03 м.д. свободного T2N, после хранения в течение 4 недель при 37 °С в присутствии сульфита в диапазоне 4-6 м.д. Предпочтительно, напитки в соответствии с данным изобретением имеют меньший бумажный привкус в сравнении с подобным напитком, полученным из другого ячменя, чем нуль-LOX-1-ячмень, после хранения в течение по меньшей мере 10 месяцев при температуре в диапазоне 15-25 °С, например, при приблизительно 20 °С. Предпочтительно, указанный бумажный привкус является меньшим чем 90%, более предпочтительно меньшим чем 80%, например, меньшим чем 70%, согласно оценке квалифицированной дегустационной комиссией. В одном варианте осуществления данное изобретение относится к напиткам, таким как пиво, с низкими уровнями определенных тригидроксиоктадеценовых кислот, в частности, к напиткам с низкими уровнями 9,12,13-ТНОЕ. Тригидроксиоктадеценовые кислоты имеют горький вкус (Baur and Grosch, 1977 и Baur et al., 1977) и, следовательно, являются нежелательными. Таким образом, желательно, чтобы уровень 9,12,13-ТНОЕ был как можно более низким, предпочтительно более низким чем 1,3 м.д., например, более низким чем 1 м.д. Однако общая концентрация 9,12,13-ТНОЕ в полученном из солода напитке (таком как пиво) также зависит от количества солода, используемого для приготовления указанного конкретного напитка. Таким образом, обычно крепкое пиво будет содержать больше 9,12,13-ТНОЕ, чем более легкое пиво, и более высокий общий уровень 9,12,13-ТНОЕ будет приемлемым в более крепком пиве. Таким образом, предпочтительно, напиток в соответствии с данным изобретением содержит более низкий уровень 9, 12,13-ТНОЕ, чем обычное пиво подобного рода. Такой напиток может быть получен с использованием нуль-LOX-1-ячменя для приготовления указанного напитка. Таким образом, предпочтительные напитки в соответствии с данным изобретением содержат низкий показатель 9,12,13-ТНОЕ в сравнении с внутренним стандартом, который вносит поправку на количество солода, использованного в приготовлении указанного напитка. Указанным стандартом может быть, например, другая тригидроксиоктадеценовая кислота. Для качества такого напитка, как пиво, важно, чтобы отношение различных тригидроксиоктадеценовых кислот (THA) поддерживалось в пределах конкретного диапазона. Неожиданно, кроме низких уровней T2N, продукта LOX-1-пути (см. фиг. 1В), напитки, приготовленные из нуль-LOX-1-ячменя в соответствии с данным изобретением, имеют также очень низкий уровень 9,12,13-ТНОЕ (см. фиг. 1С) и, соответственно, очень низкое отношение 9,12,13-THA к 9,10,13-THA. Таким образом, в одном аспекте данное изобретение относится к напиткам, таким как пиво, имеющим стабильные органолептические качества, где указанный напиток производят с использованием растения ячменя или его частей, предпочтительно нуль-LOX-1-ячменя, и где отношение 9,12,13-THA к 9,10,13-THA в указанном напитке равно максимально 1,8, предпочтительно максимально 1,7, более предпочтительно максимально 1,6, еще более предпочтительно максимально 1,5, даже еще более предпочтительно максимально 1,4. Таким образом, очень предпочтительно, чтобы указанное отношение находилось в диапазоне 0-1,8, предпочтительно в диапазоне 0-1,6, например, в диапазоне 1,4. В одном варианте осуществления указанное отношение равно приблизительно 1,3. Количество 9,12,13-ТНОЕ и 9,10,13-ТНОЕ в напитке может быть определено стандартными способами, например газовой хроматографией-масс-спектрометрией, например, как описано в Hamber, 1991. Предпочтительно, указанные THA являются оксилипинами превращения линолевой кислоты. Представляет интерес то, что напитки с такими отношениями THA могут быть получены с использованием растений ячменя в соответствии с данным изобретением. Предпочтительно указанные напитки получают без использования другого ячменя, чем нуль-LOX-1-ячмень, так что не готовят другой солод, чем солод, приготовленный из нуль-LOX-1-ячменя. В одном предпочтительном варианте осуществления данного изобретения этот напиток содержит: (i) отношение 9,12,13-THA к 9,10,13-THA, описанное выше; и (ii) уровень свободного T2N после хранения, описанный выше. В одном варианте осуществления данное изобретение относится к напитку, такому как пиво, с улучшенной устойчивостью пены в сравнении с подобным обычным напитком. Такие напитки могут быть, например, приготовлены из нуль-LOX-1-ячменя или его частей, например, из солода. Стойкость пены может быть определена, например, как описано в Brautechnische Analysenmetoden, 2002. Данное изобретение относится также к способам получения указанного напитка. Эти способы предпочтительно предусматривают стадии: (i) обеспечения солодовой композиции, содержащей пророщенные нуль-LOX-1-зерна; (ii) переработку указанной солодовой композиции в напиток; с получением посредством этого напитка со стабильными органолептическими качествами. В одном предпочтительном варианте осуществления этим напитком является пиво. В этом случае, стадия переработки предпочтительно предусматривает приготовление сусла из указанной солодовой композиции, например, любым из описанных здесь выше способов, и ферментирование указанного сусла. В общих чертах, алкогольные напитки, такие как пиво, могут быть произведены из соложеных и/или несоложеных зерен ячменя. Солод, наряду с хмелем и дрожжами, способствует вкусу и цвету пива. Кроме того, солод используется в качестве источника ферментируемого сахара и ферментов. Схематическое представление общего процесса производства пива показано на фиг. 11, тогда как подробные описания примеров подходящих способов солодоращения и пивоварения могут быть найдены, например, в недавней публикации Hoseney (supra). Многочисленные, регулярно обновляемые способы для анализов продуктов ячменя, солода и пива являются доступными [например, но не только, American Association of Cereal Chemist (1995); American Society of Brewing Chemists (1992); European Brewery Convention (1998); Institute of Brewing (1997)]. Общепризнанным является то, что многие конкретные процедуры используются для конкретного пивоваренного завода, причем большинство существенных вариаций связано с предпочтениями местных потребителей. Любой такой способ получения пива может быть использован с данным изобретением. Один неограничивающий пример описан в примере 6. Солодовая композиция для указанного напитка, например пива, солодовых напитков или неферментированного сусла, может быть, например, получена любым из описанных здесь выше способов. Сусло может быть получено из указанной солодовой композиции, как описано выше. Первая стадия производства пива из сусла включает предпочтительно кипячение указанного сусла. Во время кипячения могут быть добавлены другие ингредиенты, например, хмель, которые обеспечивают типичные горькие и ароматические характеристики пива. Кипячение сусла вызывает также агрегацию между полифенолами и денатурированными белками, которые в основном осаждаются во время последующей фазы охлаждения сусла. После охлаждения сусло переносят в ферментационные чаны, содержащие дрожжи. Предпочтительно, этими дрожжами являются пивные дрожжи, Saccharomyces carlsbergensis. Это сусло ферментируют в течение любого подходящего периода времени, обычно в течение диапазона 1-100 дней. Во время ферментации в течение нескольких дней сахар превращается в спирт и СО 2 вместе с развитием некоторых ароматных (вкусовых) веществ. Затем это пиво может обрабатываться дополнительно. Обычно его охлаждают. Оно может быть также отфильтровано и/или выдержано - в этом процессе развивается менее дрожжевой приятный аромат и вкус. Наконец, это пиво может быть пастеризовано или отфильтровано перед его упаковкой (например, в бутылки или банки). Несмотря на успехи, которые были достигнуты в области производства пива, было бы полезным уменьшение уровней T2N, его предшественников и T2N-потенциала в пиве. Таким образом, все еще существует необходимость новых сырьевых материалов, в частности ячменя и солода, которые способствуют уменьшению привкусов готового пива. Таким образом, одной целью данного изобретения является обеспечение такого пива и солода. 6.4. Химический мутагенез. В одном аспекте данное изобретение основано, по меньшей мере частично, на использовании химического мутагенеза зерен ячменя, способе, который, как известно, вводит мутации случайным образом. Мутагенез ячменя может выполняться с использованием любого мутагенизирующего химикалия, однако, предпочтительно его выполняют обработкой зерен NaN 3 , проращиванием выживших зерен и затем анализом растений-потомков. Генерация растений, выращиваемых из мутагенизированных зерен, называемая М0, содержит гетерозиготные химеры для любой конкретной мутации. Потомство, собранное после самоопыления, называют генерацией M1, и оно расщепляется как на гетерозиготы, так и на гомозиготы в отношении конкретной мутации (срав. фиг. 1А и фиг. 9). Обработка зерен NaN 3 не является эквивалентной обработке отдельной клетки, так как зерна после обработки будут содержать некоторое количество немутантных клеток и разнообразие клеток, имеющих ДНК-мутации. Поскольку мутации в линии дифференцировки, которые не приводят к зародышевой линии, будут потеряны, задачей является нацеливание мутагена на те немногие клетки, которые развиваются в репродуктивные ткани, которые способствуют развитию генерации M1. Для оценки общей эффективности мутации выполняли подсчет химер-альбиносов и растений-альбиносов в генерации М0 и генерации M1, соответственно. Подсчет количества мутантов как функции выживающих растений дает оценку эффективности мутации, тогда как подсчет количества мутантов как функции обработанных семян измеряет комбинацию как эффективности мутации, так и убивания зерен. Следует отметить, что клетки имеют механизмы гарантии качества фактически в каждой стадии экспрессии генов, возможно, для сдерживания эффектов повреждающих мутаций. Одним хорошо исследованным примером в эукариотах является опосредованное нонсенс-кодоном (терминирующим кодоном) разложение мРНК, называемое NMD, которое предотвращает синтез потенциально вредных, преждевременно укороченных белков (Maquat and Carmichael, 2001). В NMD, терминирующий кодон идентифицируют как преждевременный по его положению относительно расположенных справа (по ходу транскрипции) дестабилизирующих элементов. В Saccharomyces cerevisiae, они являются рыхло расположенными мРНК-последовательностями, а в клетках млекопитающих они являются белковыми комплексами, которые расположены на экзон-экзонных границах во время сплайсинга пре-мРНК. Каким образом деградация антисмысловых мРНК и белков, которые они продуцируют, координированы в поле деятельности для будущего исследования. Мутации, которые генерируют кодоны преждевременной терминации (нонсенс-кодоны) (РТС), увеличивают иногда уровни альтернативно сплайсированных транскриптов, которые пропускают неблагоприятные мутации, потенциально спасая посредством этого функцию белка (Mendell and Dietz, 2001). Поскольку считается, что трансляция и сплайсинг РНК происходят в разных клеточных компарментах, было парадоксальным обнаружение нонсенс-кодон-специфического повышающего регуляторного механизма мРНК, который действует независимо от разрушения мРНК усилителем сплайсинга в клетках млекопитающих (Wang et al., 2002). Однако такие механизмы не наблюдались ни в растениях ячменя данного изобретения, ни в других растениях. NMD, РТС и т.п. представляют особенный интерес в контексте селекции растений, так как такие феномены повышают количество зерен или зерновок, подлежащих скринингу для идентификации нового представляющего интерес признака. 6.5. Отбор мутантов ячменя. Одним аспектом данного изобретения является обеспечение условий скрининга на активность LOX-1, в которых активность из LOX-2 является уменьшенной. Эти способы основаны на неожиданном открытии, что характер ткани ячменя, подлежащей скринингу, и условия реакции могут усиливать активность LOX, происходящую из фермента LOX-1, и уменьшать активность из фермента LOX-2. Хотя скрининг на мутанты с низкой LOX-1, подробно описанный в РСТ-заявке PCT/IB01/00207, опубликованной в виде WO 02/053721А1, Douma et al., использовал белковый экстракт кончиков листьев ячменя, с определением ферментной активности при рН 7,5, эта публикация подробно описывает выгодные параметры скрининга, позволяющие воспроизводимо идентифицировать нуль-LOX-1-мутанты ячменя. Во-первых, при скрининге на активность LOX-1 важно использовать специфические ткани растений ячменя. Предпочтительно эта ткань включает зерно ячменя, более предпочтительно зародыши или зерна ячменя. Обычно, скрининг будет выполняться на экстракте указанной ткани, т.е. на экстракте зерен ячменя или зародышей ячменя. Более предпочтительно, экстракты для определения активности LOX-1 содержат гомогенизированную ткань зародышей или сухие зерна ячменя или более предпочтительно состоят из гомогенизированной ткани зародышей или сухих зерен ячменя. Таким путем, только незначительная активность, полученная из LOX-2, будет участвовать в определении активности. Во-вторых, анализы на активность LOX-1 выполняют при рН, который предпочтительно инактивирует ферменты алленоксидсинтазы, что позволяет получать HPODE с хорошим выходом. 6.6. Селекция растений. В одном варианте осуществления данного изобретения целью является обеспечение агрономически полезных растений ячменя, содержащих нуль-LOX-1-признак. Создание сельскохозяйственной культуры можно рассматривать как продолжительный процесс, который только начинается с введения нового признака. С точки зрения перспективы селекционера растений эта стадия почти всегда приводит к растению, которое имеет менее желательный общий профиль агрономических признаков, чем общий профиль агрономических признаков существующих коммерческих сортов. Наряду с нуль-LOX-1-признаком, имеются другие важные факторы, которые должны учитываться, например выход зерна, размер зерна и другие параметры, которые имеют отношение к эффективности солодоращения. Поскольку было показано, что многие - если не все - из таких признаков находятся под генетическим контролем, было бы очень желательным обеспечение современных, гомозиготных, дающих высокие урожаи сорта, которые происходят из скрещиваний с растениями нуль-LOX-1-ячменя, которые описаны в данной публикации. Зерна таких растений ячменя обеспечивают новый, превосходный сырьевой материал, не имеющий способности или имеющий только незначительную способность превращения линолевой кислоты в 9-HPODE. Таким образом, селекционер, работающий с ячменем, должен отбирать и развивать растения ячменя, имеющие признаки, которые приводят к лучшим сортам с потерей функции LOX-1. Альтернативно, селекционеры, работающие с ячменем, могут использовать растения данного изобретения для дополнительного мутагенеза для генерирования новых сортов, происходящих из нуль-LOX-1-ячменя. Растения ячменя в соответствии с данным изобретением, могут подвергнуты селекции в соответствии с любой подходящей схемой. 6.7. Скрещивания ячменя. Другой целью данного изобретения является обеспечение агрономически элитных растений ячменя, содержащих нуль-LOX-1-признак. Таким образом, данное изобретение направлено также на способы получения нового растения нуль-LOX-1-ячменя скрещиванием первого родительского растения ячменя со вторым родительским растением ячменя, где как первое, так и второе родительское растение является нуль-LOX-1-ячменем. Дополнительно, как первое, так и второе родительские растения ячменя могут происходить из сорта нуль-LOX-1-ячменя. Таким образом, любые такие способы, использующие сорт нуль-LOX-1-ячменя, являются частью данного изобретения: самоопыление, обратные скрещивания, скрещивания с популяциями и т.п. Все растения, продуцируемые с использованием сорта нуль-LOX-1-ячменя в качестве родителя, находятся в объеме этого изобретения, в том числе растения, полученные из сортов, произведенных из сорта нуль-LOX-1-ячменя. Нуль-LOX-1-ячмень может быть также использован для генетической трансформации в случаях, в которых экзогенную ДНК вводят в нуль-LOX-1-растение или ткань растения и экспрессируют в нуль-LOX-1-растении или ткани растения. Способы обратного скрещивания могут быть использованы с данным изобретением для введения нуль-LOX-1-признака мутированного растения ячменя в другой сорт, например, сорт Scarlett или сорт Jersey, оба из которых являются современными, высокоурожайными применимыми для солодоращения сортами ячменя. В стандартном протоколе обратного скрещивания исходный представляющий интерес сорт (повторяющийся родитель, т.е. родительская форма, с которой гибрид скрещивается вновь) скрещивают со вторым сортом (неповторяющимся родителем, т.е. родительской формой, с которой гибрид не скрещивается вновь), который несет единственный представляющий интерес ген, который должен быть перенесен. Затем полученное нуль-LOX-1-потомство из этого скрещивания опять скрещивают с повторяющимся родителем (т.е. родительской формой, с которой гибрид скрещивается вновь), причем этот процесс повторяют, пока не получают растение ячменя, в котором, по существу, все характеристики, определяемые повторяющимся родителем, восстанавливаются в превращенном растении, наряду с перенесенной генетической установкой для нуль-LOX-1-признака неповторяющегося родителя. Затем эту последнюю генерацию обратного скрещивания самоопыляют с получением чистого отобранного на нуль-LOX-1-признак потомства (см. фиг. 9). Наличие подходящего повторяющегося родителя является важным для успешной процедуры обратного скрещивания, задача которого заключается во введении признака нуль-LOX-1 в исходный сорт. Для выполнения этого генетический фон повторяющегося сорта модифицируют генетическим фоном для признака нуль-LOX-l из неповторяющегося родителя, при сохранении по существу всего остального генетического фона из исходного сорта. Хотя способы обратного скрещивания упрощаются, когда переносимое свойство является доминантным аллелем, было возможным и обратное скрещивание для переноса рецессивного признака нуль-LOX-l, но в этом случае было необходимо введение биохимического анализа для оценки, была ли перенесено это желаемое свойство. Путь для ускорения процесса селекции растений предусматривает начальное размножение генерируемых мутантов с использованием культуры ткани и способов регенерации. Таким образом, другой аспект данного изобретения состоит в обеспечении клеток, которые после роста и дифференцировки образуют растения ячменя, имеющие нуль-LOX-1-признак. Например, селекция может включать традиционные скрещивания, получение фертильных, произведенных из пыльников растений или использование культуры микроспор. 6.8. Ферменты LOX. Важным объектом данного изобретения является обеспечение растений ячменя, которые лишены способности синтезировать активный фермент LOX-1. LOX являются большими мономерными белками с единственным не имеющим структуры гема железосодержащим фактором. Исследование Банка Данных Белков в http://www.rcsb.org/pdb выявило, что структура нескольких ферментов LOX была определена рентгеновской кристаллографией. Эти белки имеют общую организацию складок и доменов, причем каждый имеет меньший N-концевой восьмицепочечный β-бочковидный домен и больший С-концевой домен, состоящий в основном из длинных α-спиралей. Атом железа расположен в С-концевом домене, где он координирован с остатками гистидина и, уникально, с карбоксильным концом этого полипептида, которым может быть изолейцин. Несколько каналов идут от поверхности этого белка к близкому с сайтом железа району, и они предположительно предоставляют доступ для субстратов, полиненасыщенных жирных кислот и молекулярного кислорода, к активному центру. Поскольку связывание LOX липосомой и липидным тельцом липидного тельца огурца зависит от присутствия N-концевого β-бочковидного домена (May et al., 2000), a LOX-1 сои связывается с бислойными мембранами в процессе, который усиливается ионами кальция (Tatulian and Steczko, 1998), можно предположить, что ферменты LOX связываются с липидными бислойными мембранами и что это является, вероятно, функцией N-концевого домена. Способы определения активности LOX-1, а также выделения, характеристики и количественного определения непосредственных и следующих за ними продуктов катализа LOX являются легко доступными для специалистов в данной области. 6.9. Продукты LOX-пути. В различных вариантах осуществления данное изобретение относится к растениям ячменя или их продуктам, блокированным в способности образовывать алкеналь T2N. Ферменты LOX катализируют диоксигенирование полиненасыщенных жирных кислот цис-1,цис-4-пентадиеновой системой. В растениях С 18 полиненасыщенные кислоты линолевая кислота (18:2 49,12 ) и α-линоленовая кислота (18:2 49,12,15 ) являются главными субстратами LOX. Этот липоксигеназный путь метаболизма жирных кислот инициируется присоединением молекулярного кислорода в положении С-9 или С-13 ацильной цепи с образованием соответствующих гидропероксидов 9- и 13-линолевой или линоленовой кислоты. С линолевой кислотой в качестве субстрата могут быть образованы 9- или 13-гидропероксиоктадекадиеновые кислоты (HPODE), тогда как 9- или 13-гидропероксиоктадекатриеновые кислоты (НРОТЕ) являются продуктами, когда субстратом является α-линоленовая кислота. В ответвлении гидропероксидлиазы LOX-пути как 9-, так и 13-гидропероксиды могут затем расщепляться на оксокислоты и альдегиды с короткой цепью (см. фиг. 1В). Следует отметить, что 9-HPODE могут быть дополнительно метаболизированы в 9,12,13-ТНОЕ (см. фиг. 1С), причем ТНОЕ имеют горький вкус (Baur et al., 1977; Baur and Drosch, 1977). Таким образом, растения с инактивированным LOX-1 будут образовывать ТНОЕ в соотношениях, отличающихся от соотношений, наблюдаемых в растениях дикого типа. Авторы осознают, что данное изобретение включает влияние на образование последующих метаболитов LOX-1-катализа, которые продуцируются не в качестве прямого продукта LOX-катализируемой реакции, а в качестве продукта последующей реакции из ряда реакций, включающих продукт LOX-1-катализа. Эти реакции включают спонтанную, индуцируемую факторами или катализируемую ферментом изомеризацию. Таким образом, экспрессия гидропероксидлиазы (HPL) могла бы модулировать образование этих последующих метаболитов. При допущении, что самоокисление линолевой кислоты может генерировать молекулы предшественника, связанные с образованием T2N, можно дополнительно уменьшить уровень этого алкеналя. Конкретно, ожидается, что отрицательная регуляция генов, кодирующих Δ9-десатуразу (которая превращает стеариновую кислоту в олеиновую кислоту) или Δ12-десатуразу (которая превращает олеиновую кислоту в линолевую кислоту), изменяет относительные доли C 18 жирных кислот (стеариновой, олеиновой и линолевой кислот) уменьшением уровней жирных кислот ниже по ходу реакции после соответствующего фермента и увеличением уровней промежуточного состоящего из жирных кислот субстрата. Примеры, в которых селективный отбор с использованием природных вариантов или индуцированных мутаций применяли для развития диапазона улучшенных масел в масличных сельскохозяйственных культурах, включают, но не ограничиваются ими, сою с высоким содержанием стеариновой кислоты (HS) (Graef et al., 1985), рапс с высоким содержанием олеиновой кислоты (НО) (Auld et al., 1992), а также подсолнечник с высоким содержанием HS и НО, описанный Osorio et al. (1995) и Soldatov (1976), соответственно. Особый интерес представляет то, что данное изобретение включает получение альдегидов, которые не являются прямыми продуктами действия LOX-1, но продуцируются под действием ферментов LOX-пути или изомеризацией альдегидов, например, изомеризацией (3Z)-ноненаля в (2Е)-ноненаль, как показано на фиг. 1В. Авторы осознают также, что данное изобретение включает получение таких спиртов, которые соответствуют альдегидам, продуцируемым ферментами LOX-пути, и которые соответствуют альдегидам, образуемым в результате указанной изомеризации. Указанные спирты обычно образуются под действием ферментов, являющихся членами семейства альдо-кеторедуктаз (Srivastava et al., 1999), например, посредством ферментативного превращения (2Е)-ноненаля в (2Е)-ноненол. 6.10. T2N-потенциал. Следующей целью данного изобретения является уменьшение или элиминация молекул, связанных с образованием T2N, в том числе образованием предшественников T2N и аддуктов альдегидов. Хотя несколько химических реакций, связанных с потерей свежести пива, остаются невыясненными, основными причинами развития затхлого привкуса в пивных продуктах считаются окислительные процессы (Narziss, 1986; Ohtsu et al. 1986). Как подробно описано в разделе 2 ("Уровень техники"), хорошо известно, что основной молекулой, содействующей затхлому привкусу, является T2N. Когда этот альдегид образуется в процессе приготовления пива в стадии производства перед ферментацией, он может участвовать в образовании аддуктов посредством связывания, например, с аминокислотами и белками (Noel and Collin, 1995), но, возможно, также нуклеиновыми кислотами, глутатионом или т.п., и затем быть защищен от восстановления или окисления ферментирующими дрожжами (Lermusieau et al., 1999). Однако аддукты T2N могут быть также образованы с сульфитом во время ферментации, делая неактивным альдегидный привкус (Nyborg et al., supra). Большинство аддуктов T2N переносятся в готовое пиво, где свободный T2N высвобождается (Liegeols et al., 2002), причем важными факторами в этом процессе являются кислотность и температура. Аддукты T2N составляют часть T2N-потенциала, меры деградации аддуктов T2N до свободного T2N при определенных условиях реакции, например, при инкубировании при 100 °С, рН 4,0 в течение 2 ч. Квалифицированный специалист знает, каким образом использовать этот T2N-потенциал в качестве индикатора того, как пиво будет высвобождать T2N во время хранения, например, как описано Drost et al. (supra). Зерна ячменя данного изобретения ограничены в LOX-1-катализируемом образовании 9-HPODE, молекулы, которая в норме функционирует в качестве предшественника ветви LOX-пути, которая производит T2N. Таким образом, пиво, полученное с использованием зерен нуль-LOX-1-ячменя, будет обладать не только очень низким уровнем T2N, но также очень низким уровнем Т2N-потенциала. В объеме данного изобретения находятся зерна нуль-LOX-1-ячменя, дающие пивные продукты, которые полностью лишены T2N или содержат незначительные уровни T2N-потенциала, в том числе аддуктов T2N. Таким образом, по существу, нет развития T2N-специфических привкусов или имеется только незначительное развитие T2N-специфических привкусов во время хранения пива, полученного с использованием нуль-LOX-1-ячменя. 6.11. Устойчивость к болезням. Данное изобретение относится дополнительно к ячменю, устойчивому к болезням. Считается, что LOX растений участвуют в развитии активных механизмов устойчивости к болезням, вместе известных как гиперчувствительная реакция (HR), форма программированной смерти клеток (Rusterucci et al., 1999). В HR за событием инфицирования следует быстрая смерть клеток растения, локализованных около места инфицирования, и это приводит к образованию некротического повреждения. Таким путем распространение патогена ограничивается и предотвращается дальнейшее повреждение остальной части органа растения. В нескольких системах растение-патоген, HR связана с экспрессией LOX, специфических для генерирования 9-HPODE и 9-НРОТЕ (Rusterucci et al., supra; Jalloul et al., 2002), возможно, вследствие того, что массивное продуцирование гидропероксижирных кислот приводит к некрозу тканей. Ген, кодирующий LOX-1, экспрессируется прежде всего в зернах ячменя, в то время как многочисленные дополнительные ферменты LOX экспрессируются в листьях этих растений. Таким образом, ответвления LOX-пути, приводящие к образованию 9-HPODE, 13-HPODE, 9-НРОТЕ и 13-НРОТЕ, являются функциональными в листьях ячменя, и различные наборы оксилипинов отражают отдельные события инфицирования и появления ран. Подобный молекулярный сценарий был описан для листьев картофеля (Weber et al., 1999). Природно встречающиеся летучие альдегиды ингибируют рост определенных патогенов на растении, и природная устойчивость некоторых растений к конкретному патогену может быть приписана генерированию летучих альдегидов (Croft et al., 1993; Blee and Joyard, 1996; Vancanneyt et al., 2001). Таким образом, измененный относительно растений дикого типа профиль оксилипинов растений нуль-LOX-1-ячменя может предотвращать, уменьшать, ослаблять или элиминировать присутствие патогена, продукта патогена или продукта взаимодействия растение-патоген. Одним неограничивающим примером патогена является Aspergillus (см. здесь ниже). Таким образом, в одном варианте осуществления данное изобретение относится к растению нуль-LOX-1-ячменя, обнаруживающему повышенную устойчивость к болезням. 6.12 Микотоксины. Данное изобретение описывает также применение растений ячменя с уменьшенной чувствительностью к колонизации Aspergillus. Aspergillus является причиняющим беспокойство агентом колонизации зерен ячменя, часто вызывающим загрязнение канцерогенными микотоксинами афлатоксином и стеригматоцистином. Поскольку на образование афлатоксина этим грибом влияют высокие уровни 9-HPODE, 13-HPODE, 9-НРОТЕ и 13-НРОТЕ, Патент США с номером 5942 661, выданный Keller, заявляет трансгенные сельскохозяйственные растения, которые продуцируют указанные гидропероксижирные кислоты в количествах, достаточных для ингибирования образования грибных микотоксинов. Кроме того, указанный Патент США, а также данные Burow et al. (2000) показывают, что 13-HPODE ингибирует образование афлатоксина, в то время как 9-HPODE повышает образование афлатоксина. Поскольку нуль-LOX-1-зерна лишены активного фермента LOX-1, указанные зерна содержат слегка более высокие уровни 13-HPODE, чем растения дикого типа, но также более низкие уровни 9-HPODE относительно ткани его генетически не модифицированного родительского растения. Таким образом, нуль-LOX-1-зерна могут отражать колонизацию Aspergillus или проявлять пониженные уровни микотоксина после загрязнения Aspergillus, в сравнении с зернами дикого типа. Таким образом, данное изобретение относится к растениям ячменя с уменьшенными уровнями микотоксинов в сравнении с растениями ячменя дикого типа. 6.13. Ароматные (вкусовые) вещества. Аспектом данного изобретения является также применение нуль-LOX-1-ячменя для получения ароматных (вкусовых) веществ и соединений ноты зелени. До сих пор большинство попыток исследований, связанных с различными ответвлениями LOX-пути в ячмене, фокусировались на аспектах генерирования жасмоновой кислоты из 13-НРОТЕ (Turner et al., 2002) и на описанной выше устойчивости к болезням. Меньше внимания уделялось гидропероксижирным кислотам для альтернативных, коммерческих целей. Однако примечательно, что полное отсутствие активного LOX-1 в зернах нуль-LOX-1-ячменя должно, как ожидается, обогащать указанные зерна 13-HPODE и 13-НРОТЕ. На основе этого нового свойства, вероятными являются новые приложения в отношении промышленной применимости этой культуры ячменя, например, в получении короткоцепочечных алифатических альдегидов и спиртов (например, соединений ноты зелени гексаналя/гексеналя и гексанола/гексенола). Несколько аспектов, связанных с получением соединений ноты зелени описаны в патентах, включающих, но не ограничивающихся ими, патенты США с номерами 6008034, 6150145 и 6274358, которые обсуждаются. Хотя патент США № 6008034, выданный Hausler et al., раскрывает применение специфической гидропероксидлиазы для получения соединений ноты зелени, патент США № 6150145, выданный Hausler et al., и патент США № 6274358, выданный Holtz et al., описывают применение стандартного растительного материала для такого процесса. Использование нуль-LOX-1-зерен для получения соединений ноты зелени предусматривает использование нового сырьевого материала для указанного получения. Новый сырьевой материал, полученный из зерен нуль-LOX-1-ячменя данного изобретения, не может рассматриваться как стандартный растительный материал, так как он произведен из зерен, которые были отобраны в соответствии с протоколом мутагенеза, подробно описанным в Разделах 6.4-6.7 данной публикации. Промышленная применимость зерен нуль-LOX-1-ячменя рассматривается вне объема формул изобретений, приведенных в патентах, описанных в предыдущем абзаце, прежде всего потому, что новый сырьевой материал, получаемый из нуль-LOX-1-зерен данного изобретения, будет значительно улучшать обычные ограничения, вызываемые LOX-1-катализируемым генерированием 9-HPODE и 9-НРОТЕ, двух гидропероксижирных кислот, которые не могут действовать в качестве молекул-предшественников для ферментативного генерирования соединений ноты зелени цис-3-гексеналя и цис-3-гексенола. 6.14. Гетерологичная экспрессия генов, кодирующих LOX. В различных вариантах осуществления данное изобретение относится к трансгенным растениям ячменя, имеющим нуль-LOX-1-признак. Ожидается, что будущие успехи в генной инженерии растений приведут к генерированию растений ячменя с подавленным синтезом LOX-1. Эта концепция была предложена в качестве средства контроля образования привкусов, но о результатах такого подхода не сообщалось (McElroy and Jacobsen, 1995). Изобретение, раскрытое в настоящей заявке, может быть использовано вместе с такими будущими усовершенствованиями для генерирования антисмысловых LOX-1-растений, имеющих антисмысловые конструкции, комплементарные по меньшей мере части мессенджер-РНК (мРНК) LOX-1, последовательности которых могут быть сконструированы. Антисмысловые нуклеотиды конструируют для гибридизации с соответствующей мРНК, например, аналогично тому, что описано для экспрессии последовательности протеинкиназы SnRK1 в трансгенном ячмене (Zhang et al., 2001). Модификации этих антисмысловых последовательностей могут выполняться, пока эти последовательности гибридизуются с соответствующей мРНК и препятствуют экспрессии соответствующей мРНК. Таким путем могут быть использованы антисмысловые конструкции, имеющие 70%, предпочтительно 80%, более предпочтительно 85% идентичность последовательности относительно соответствующих антисмысловых последовательностей. Кроме того, части этих антисмысловых нуклеотидов могут быть использованы для разрушения экспрессии гена-мишени. Обычно могут быть использованы последовательности из по меньшей мере 50 нуклеотидов, 100 нуклеотидов, 200 нуклеотидов или большего числа нуклеотидов. Таким образом, применимость этого изобретения не ограничивается только растениями, полученными традиционными способами мутагенеза. Хотя замена гена-мишени гомологичной рекомбинацией является очень легкой в дрожжах, ее эффективность в большинстве многоклеточных эукариот еще является ограниченной и пока не позволяет получать такие растения ячменя, а также получать разрушения генов в масштабе всего генома (Parinov and Sundaresan, 2000). Сайленсинг генов был недавно использован для исследования роли ~86% предсказанных генов генома Caenorhabditis elegans в нескольких разработанных процессах (Ashraft et al., 2003; Kamath et al., 2003). Для генерирования растений ячменя с полной потерей функции конкретного гена, такого как ген, кодирующий LOX-1, применение способа интерференции РНК (RNAi) имеет несколько недостатков. Они включают отсутствие стабильного наследования фенотипа, вариабельные уровни остаточной активности гена (Harmon, 2002; Bargman, 2001; Wesley et al., 2001) и невозможность одновременного сайленсинга нескольких неродственных генов (Kamath et al., 2000). Нуклеотидные последовательности данного изобретения могут быть также использованы в смысловой ориентации для подавления экспрессии эндогенных генов, кодирующих ферменты LOX в растениях. Способы подавления экспрессии генов в растениях с использованием нуклеотидных последовательностей в смысловой ориентации известны в данной области (см., например, патент США № 5283184, выданный Jorgensen и Napoli). Эти способы обычно включают трансформацию растений ДНК-конструкцией, содержащей промотор, который регулирует экспрессию в растении, функционально связанный по меньшей мере с частью нуклеотидной последовательности, которая соответствует транскрипту этого эндогенного гена. Обычно, такая нуклеотидная последовательность имеет существенную идентичность последовательности с последовательностью транскрипта эндогенного гена, предпочтительно большую, чем приблизительно 65% идентичность последовательности, более предпочтительно большую, чем 85% идентичность последовательности, наиболее предпочтительно большую, чем приблизительно 95% идентичность последовательности. Следует отметить, что различные аспекты, связанные с гетерологичной экспрессией генов, кодирующих ферменты LOX, описаны и раскрыты в Публикации заявки на патент США № 2003/0074693 Al, Cahoon et al. Хотя указанная заявка на патент описывает предыдущий уровень техники в отношении ферметов LOX ячменя и описывает многочисленные кодирующие LOX последовательности генов, ни один из генов ячменя, кодирующих LOX-1, LOX-2 и LOX-3, проявляющих достаточную степень идентичности, не включен в формулу изобретения, представленную в публикации заявки на патент США № 2003/0074693 A1, Cahoon et al. Хотя данное изобретение было подробно описано в предыдущем описании, это описание рассматривается как иллюстративное и неограничительное по характеру, причем должно быть понятно, что желательно, чтобы все изменения и модификации, которые находятся в пределах идеи данного изобретения, были защищены. Таким образом, будет очевидно, что некоторые изменения и модификации, такие как модификации и мутации единственного гена, сомаклональные варианты, вариантные особи, выбранные из больших популяций растений данного сорта, и т.п., могут быть использованы в объеме данного изобретения, как ограничиваемые только объемом прилагаемой формулы изобретения. Далее данное изобретение будет описано со ссылкой на следующие конкретные примеры; они приведены для дополнительной иллюстрации данного изобретения и не должны рассматриваться как ограничивающие его объем. 6.15. Ингибиторы LOX-1. Данное изобретение относится также к способам уменьшения или предотвращения активности LOX-1 ячменя. Несколько ингибиторов LOX могут быть выбраны из классов окислительно-восстановительных и не-окислительно-восстановительных ингибиторов, антиоксидантов, хелатирующих железо агентов, имидазолсодержащих соединений, производных бензопирана и т.п. Таким образом, данное изобретение в одном варианте осуществления относится к способу уменьшения активности LOX ячменя (предпочтительно LOX-1), предусматривающему стадии (i) обеспечения растения ячменя или его части или продукта растительного происхождения, полученного из ячменя, (ii) обеспечения ингибитора LOX, (iii) инкубирования указанного растения ячменя или его части или продукта растительного происхождения, полученного из ячменя, с указанным ингибитором LOX с уменьшением посредством этого активности LOX (предпочтительно LOX-1). В одном варианте осуществления указанным продуктом растительного происхождения является солод, и указанный ингибитор LOX добавляют к указанному солоду во время процесса затирания. Предпочтительно, это будет приводить к более низкому уровню T2N в сусле, полученном в указанном процессе затирания. Указанное растение ячменя или его часть или продукт растительного происхождения, полученный из ячменя, могут быть нуль-LOX-1-ячменем или его частью или продуктом растительного происхождения, полученным из нуль-LOX-1-ячменя. Однако с этими способами могут быть использованы предпочтительно другие ячмени. Среди различных ингибиторов LOX, окислительно-восстановительные ингибиторы LOX могут быть выбраны из производных катехолбутана, таких как любой из производных катехолбутана, описанных в Патентах США с номерами 5008294, Jordan et al., 4708964, Allen, и 4880637, Jordan, таких как нордигидрогуайяретовая кислота (NDGA), или один из ее энантиомеров. Антиоксидантный ингибитор LOX предпочтительно выбран из фенолов, флавоноидов и т.п. Антиоксидантный ингибитор LOX может быть также выбран из галлатов, в том числе может быть октилгаллатом. То, что соединение действительно является ингибитором LOX, может быть подтверждено анализом, описанным здесь в примере 18 ниже. Для промышленности октилгаллат является известным ингибитором липоксигеназы сои (На et al., 2004), и он представляет особенный интерес, так как он в настоящее время разрешен для применения в качестве антиоксидантной добавки в пище (Aruoma et al., 1993). Это свойство сделало его представляющим интерес для тестирования очищенного LOX-1 ячменя на активность в присутствии предположительного ингибитора октилгаллата. Интересно, что присутствие октилгаллата во время затирания приводит к более низким уровням T2N. Таким образом, одним вариантом осуществления данного изобретения является обеспечение ингибитора LOX-1, а также его применения, которые, после добавления к затору, будут давать уменьшенные уровни T2N. Приведенные здесь примеры иллюстрируют предпочтительные варианты осуществления данного изобретения и не должны рассматриваться как ограничивающие данное изобретение. Если нет других указаний, основные молекулярно-биологические способы выполняли для манипулирования нуклеиновыми кислотами и бактериями, как описано в Sambrook et al. (1989) и Sambrook and Russel (2001). Для целей ясности описания, но не в качестве ограничения, данный раздел примеров разделен на следующие темы. (i) Скрининг и отбор мутантов. (ii) Мутанты ячменя D112 и А618. (iii) Физиологические свойства мутантов ячменя D112 и А618. (iv) Мутанты D112 и А618 являются нуль-LOX-1-растениями. (v) Затирание. (vi) Получение солода и пива с использованием солода ячменя дикого типа и мутантного ячменя. (vii) Тригидоксиоктадеценовые кислоты в пиве из нуль-LOX-1-солода. (viii) Тригидоксиоктадеценовые кислоты в пиве. (ix) Биохимическая характеристика ферментативных продуктов из действия LOX в ячмене. (х) Ген для LOX-1 в мутанте ячменя D112 является мутированным. (xi) Ген для LOX-1 в мутанте ячменя А618 является мутированным. (xii) ОТ-ПЦР-детектирование транскриптов для LOX-1. (xiii) Генетическое детектирование мутантов ячменя, несущих мутацию D112. (xiv) Детектирование мутантов в пробах смешанного солода. (xv) Рекомбинантный LOX-1 мутанта D112 является неактивным. (xvi) Трансгенные растения ячменя. (xvii) Соединения ноты зелени. (xviii) Ингибиторы LOX-1. (xix) Затирание с октилгаллатом. Пример 1. Скрининг и отбор мутантов. Мутагенез ячменя. Зерна, собранные из растений ячменя сортов Barke, Celeste, Lux, Prestige, Saloon и Neruda, инкубировали по отдельности с мутагеном NaN 3 в соответствии с деталями, описанными Kleinhofs et al. (1978). Эту процедуру выбирали, так как известно, что она индуцирует точковые мутации в геномной ДНК ячменя и приводит к заменам аминокислот или укорочениям в белках, кодируемых этой мутагенизированной ДНК. В экспериментах по мутагенезу данной публикации было выбрано размножение мутированных зерен генерации M1 в полевых делянках посредством двух последовательных генераций, с получением в конечном счете высокой доли гомозиготных растений для целей скрининга (фиг. 1А). Ожидалось, что мутированные зерна полученной генерации М3 будут встречаться при частоте 0,9-2,3 на 10000 зерен (Kleinhofs et al., supra). Следует отметить, что зерна М2 не подвергали скринингу, так как они содержат относительно высокую долю гетерозиготных точковых мутаций. Скрининг. Целью было развитие процедуры быстрого высокопроизводительного скрининга для детектирования зерен мутанта ячменя М3, лишенных активности LOX-1, во избежание трудной процедуры скрининга с использованием кончиков листьев, которые, как известно, содержат несколько активностей LOX (описанных в РСТ-заявке PCT/IB01/00207, опубликованной в виде WO 02/053721A1, Douma et al.). В центре было определение активности LOX в зародыше, включающем ткань шитка, зрелых зерен ячменя. Обычно, условия анализа были сходными с условиями, описанными Anthon and Barrett (2001). Анализ был основан на LOX-катализируемом генерировании гидропероксидов линолевой кислоты, которые - в катализируемой гемоглобином реакции - окислительно связывают 3-метил-2-бензотиазолинон с 3-(диметиламино)бензойной кислотой, приводя к образованию синей окраски, которая может быть измерена спектрофотометрически. В практическом выражении, одна серия анализов инициировалась отдельной гомогенизацией тканей 96 зародышей ячменя, включающих ткани щитка, с получением композиций тонкоизмельченного порошка. Их переносили на планшеты для хранения на льду (ABgene), в которых каждая из 96 лунок на 1,2 мл содержала круглую стеклянную гранулу 5 мм и 200 мкл Н 2 О. Затем этот планшет инкубировали в течение 35 с в лабораторной мельнице ММ 300 (Retsch), электронно приспособленной для встряхивания при частоте 27 с -1 . Затем этот планшет центрифугировали при 4000 об/мин в центрифуге Allegra 6R (Beckman-Coulter) в течение 15 мин при 4 °С для осаждения нерастворимого материала и после этого выдерживали на льду в течение максимально 120 мин до последующей обработки. Этот 96-луночный планшет переносили в Лабораторную Автоматическую Рабочую Станцию Biomek 2000 (Beckman-Coulter), которая была запрограммирована для пипетирования в соответствии с анализом LOX, описанным Anthon and Barrett (supra). Наконец, 96 ×26 мкл экстрактов зародышей переносили в стандартный 96-луночный микротитрационный планшет (Nunc) с последующим добавлением 90 мкл Реагента А [12,5 мМ 3-(диметиламино)бензойная кислота, 0,625 мМ линолевая кислота (полученная, как описано подробно в примере 9)] и 90 мкл Реагента В (0,25 мМ 3-метил-2-бензотиазолингидразон, 0,125 мг/мл гемоглобина); Реагент А получали смешиванием сначала 155 мкл линолевой кислоты, соответствующей 134 мг свободной кислоты (Sigma, L-1376), и 257 мкл Твина-20, затем добавляли Н 2 О для получения объема 5 мл, с последующим добавлением 600 мкл 1 М NaOH, и, когда этот раствор становился прозрачным, его доводили до 20 мл дополнительным добавлением Н 2 О. В каждой из этих 96 лунок этого планшета измеряли А 595 с использованием спектрофотометра Fluorostar Galaxy (BMG Labtechnologies), причем образование окраски гидропероксидных продуктов является мерой суммарной активности присутствующих LOX [активности приведены, соответственно, в единицах А 595 (А 595 U)]. Идентификация потенциальных мутантов. Зерна ячменя сорта Barke (полученные из 2160 линий в целом), сорта Celeste (2867 линий), сорта Lux (2625 линий), сорта Prestige (1379 линий), сорта Saloon (1743 линии) и сорта Neruda (3780 линий) подвергали скринингу на активности LOX с целью идентификации зерен, сильно пониженных в указанной активности в сравнении с зернами дикого типа. В целом идентифицировали 90 потенциальных мутантов в генерации М3 [сравн. сорт Barke (12 линий), сорт Celeste (38 линий), сорт Lux (9 линий), сорт Prestige (16 линий), сорт Saloon (12 линий) и сорт Neruda (3 линии)]. Зерна из каждого из этих мутантов размножали до генерации М4, собирали и затем повторно подвергали скринингу на этот признак, связанный с очень низкой активностью LOX. Наконец, было показано, что только одна линия сорта Barke, названная мутантом D112, и одна линия сорта Neruda, названная мутантом А618, проявляли указанную очень низкую общую активность LOX. Подробные измерения активностей LOX выполняли с экстрактами зрелых, покоящихся зерен, в которых активность LOX была почти исключительно активностью LOX-1 (Schmitt and Van Mexhelen, 1997). Для зародышей сухих, зрелых зерен М3 мутанта D112 общая активность LOX-1 - определенная колориметрическим анализом LOX, как описано выше (срав. фиг. 2, таблицу 1) - была 0,407 ±5,8% А 595 U/зародыш, тогда как общая активность LOX для сорта Barke была 1,245 ±7,6% А 595 U/зародыш. Во второй серии экспериментов, было обнаружено, что активность LOX в экстрактах зародышей зрелых, покоящихся зерен генерации М3 мутанта А618 была 0,221 ±2,6% А 595 U/зародыш, тогда как 0,721 ±3,6% A 59 s U/зародыш было обнаружено в сорте Neruda дикого типа (фиг. 3; табл. 1). Пример 2. Мутанты ячменя D112 и А618. Анализы проводили для установления, были ли нуль-LOX-1-растения генераций М4 и М5 гомозиготными в отношении соответствующего фенотипа мутанта. Этот тип анализа был применим для определения рецессивного или доминантного характера представляющей интерес мутации в генерации М3. Другими словами, если растения генерации М3 были гетерозиготными в отношении доминантной мутации, то последующие генерации расщеплялись бы в отношении этого фенотипа. Общую активность LOX измеряли в зародышах генераций М3, М4 и М5 мутантов ячменя D112 и А618, и эти активности сравнивали с активностями зародышей из сорта Barke и сорта Neruda, соответственно. Определение активности LOX выполняли, как описано в примере 1 данного изобретения. Во всех этих экспериментах в качестве контрольных проб использовали стандартные экстракты из зародышей сорта Barke, а также инактивированные нагреванием стандартные экстракты из зародышей сорта Barke. Для зародышей генерации М4 зерна мутанта D112, средняя общая активность LOX была 0,334 ±1,5% A 595 U (n=12), а средняя общая активность LOX для зародышей генерации М5 мутанта D112 была 0,294 ±4,1% А 595 U (n=90). Для сравнения, зародыши генерации М4 и генерации М5 сорта Barke дикого типа давали 0,738 ±3,2% А 595 U (n=2) и 0,963 ±7,5% А 595 U (n=90), соответственно (срав. фиг. 4; фиг. 5; таблицу 1, эксперимент 1). Зародыши генерации М4 мутанта ячменя А618 давали среднюю активность LOX 0,222 ±2,1% А 595 U (n=4). Другие результаты этого эксперимента выявили, что активность LOX в зародышах сорта Neruda была 0,684 ±5,8% А 595 U (n=90). Эти результаты суммированы на фиг. 6 и в табл. 1, эксперимент 2. В целом, экспериментальные результаты подтвердили, что зерна генераций М4 и М5 мутанта D112 были гомозиготными в отношении генетического признака, специфического для очень низкой активности LOX. То же самое свойство было показано для зерен генерации М4 мутанта А618. Пример 3. Физиологические свойства мутантов ячменя D112 и А618. Размножение растений в оранжерее. Зерна сорта Barke и мутанта D112 (генераций М4 и М5) проращивали и выращивали в оранжерее при 20 ч света при 12 °С и при относительной влажности 65%. Характеристики роста мутанта D112 и растений сорта Barke дикого типа были сходными в отношении высоты растений, количества побегов на одно растение, начала зацветания и количества зерен на колос. Таким образом, можно сделать вывод, что мутант D112 не только имеет физиологию роста растения дикого типа, но имеет также нормальное развитие зерен. Зерна мутанта А618 генерации М4 сорта Neruda проращивали и выращивали в оранжерее при условиях свет/темнота 20 ч/4 ч при 12 °С и при относительной влажности 65%. При сравнении мутанта А618 и сорта Neruda не наблюдали различий в отношении высоты растений, количества побегов на одно растение, начала зацветания и количества зерен на колос. Однако дорсальная сторона зерен мутанта А618 отличалась от материнского сорта Neruda ненормальной подобной дырке структурой. В целом можно сделать вывод, что мутант А618 обнаруживает физиологию роста растений, сходную с физиологией роста растений дикого типа, но отклоняющееся от нормы развитие зерен. Агрономическая эффективность мутанта D112 в полевых условиях. Мутант 1122 и растения сорта Barke сравнивали в полевых испытаниях для идентификации возможных различий в отношении высоты, времени колошения, устойчивости к болезням, полегания, раскрытия колоса, времени созревания и урожая растений (см. табл. 2). Эти испытания выполняли в соответствии со стандартными процедурами для полевых испытаний. Таким образом, равные количества зерен мутанта D112 и сорта Barke высевали в делянках 7,88 м 2 в 2 местоположениях, каждое из которых имело 3 повторности. Характеристики агрономических результатов, с особым вниманием в отношении описанных выше свойств, тщательно наблюдали на протяжении всего вегетационного периода. Не наблюдали различий в отношении агрономических признаков ни для мутанта D112, ни для сорта Barke. Пример 4. Мутанты D112 и А618 являются нуль-LOX-1-растениями. Анализ белка. Следующие анализы выполняли для характеристики фенотипа мутантов D112 и А618. Вестерн-блот-анализы выполняли с экстрактами зародышей, удаленных из покоящихся зерен ячменя. Один зародыш экстрагировали в 300 мкл охлажденной на льду воды в ступке, экстракты переносили в микроцентрифужную пробирку и центрифугировали при 10000 ×g. Аликвоты проб, содержащие 10 мкл неочищенных экстрактов, разделяли электрофорезом в додецилсульфат натрия-полиакриламидном геле (ДСН-ПААГ) в соответствии с описаниями, обеспеченными Laemmli (1970). После этого разделенные белки переносили на нитроцеллюлозные мембраны полусухим блоттингом, как подробно описано Towbin et al. (1979). Этот блот зондировали разведением 1:500 LOX-1-специфического моноклонального антитела 5D2 (Hofman et al., 1996) с последующим инкубированием с козьим антимышиным антителом, конъюгированным со щелочной фосфатазой, и детектировали с субстратами щелочной фосфатазы нитрокрасителем тетразолиевым синим и 5-бром-4-хлор-3-индолилфосфатом, как описано Holtman et al. (supra). LOX-1 узнавался антителами 5D2 в экстрактах зародышей сорта Barke, и этот белок мигрировал при электрофорезе в ДСН-ПААГ сходно с миграцией LOX-1 из сорта Vintage. Иммунодетектируемый LOX-1 отсутствовал в пробах мутанта D112, но этот белок можно было детектировать в экстрактах сорта Barke, мутантной линии G и сорте Vintage. Вестерн-анализы сорта Barke и линий потомства мутанта D112 генерации М4 выявил, что белок LOX-1 присутствует в зернах из зародышей сорта Barke, но не в зернах любого из потомств мутанта D112 (фиг. 7), что подтверждало, что нуль-LOX-1-признак является генетически стабильным. Эти результаты были статистически значимыми, как было определено с использованием критерия хи-квадрат (р <3,84). Мутант А618 и сорт Neruda анализировали на белок LOX-1 в зародышах генераций М3 и М4, как описано выше. Белок LOX-1 мог быть детектирован в экстрактах зародышей сорта Neruda обеих генераций. Однако, очень слабую полосу LOX-1 наблюдали в исходном мутанте А618, а также в зародышах линий потомства (фиг. 8), возможно, вследствие перекрестной реакции с другими ферментами LOX. Обратное скрещивание. Повторяемое обратное скрещивание использовали для переноса нуль-LOX-1-фенотипа из мутанта D112 в повторяющегося родителя (срав. фиг. 9), в данной публикации сорт Prestige. Была разработана программа обратного скрещивания, показанная на фиг. 9, объединенная с отбором на представляющий интерес признак. Целью была прогрессирующая замена генома мутанта D112 геномом повторяющегося родителя. Таким путем, могут быть исключены другие потенциальные, нежелательные мутации, введенные в геном мутанта D112 во время обработки мутагенеза с использованием NaN 3 . В первом обратном скрещивании гомозиготного нуль-LOX-1-мутанта D112 (обозначенного как генотип nn) с сортом Prestige (обозначенным как генотип NN) ожидалось, что линии потомства содержат гетерозиготный фенотип (обозначенный как генотип Nn). Следует отметить, что фенотип низкого LOX, вследствие его рецессивного характера, мог бы избежать детектирования в линиях, которые являются гетерозиготными в отношении этой мутации. Ожидалось, что самоопыляющиеся растения-потомки дадут популяцию растений, которая могла бы расщепляться по Менделевскому типу, а именно, в соотношении 1NN:2Nn:1nn. Ожидалось, что гомозиготный генотип nn, содержащий нуль-LOX-1-генотип и происходящий из первого скрещивания, содержал 50% генетического фона сорта Prestige. Ожидалось, что после 10 раундов обратного скрещивания генетический фон повторяющегося родителя увеличится до ~99,9%. Растения ячменя сорта Prestige и нуль-LOX-1-мутанта D112 размножали в оранжерее на протяжении всей программы обратного скрещивания. Зерна подвергнутых обратному скрещиванию потомков анализировали на присутствие белка LOX-1 в экстрактах зародышей, как описано в примере 1. Ожидаемая частота нуль-LOX-1-фенотипа в расщепляющемся потомстве первой и второй генераций обратного скрещивания была 25% в отношении рецессивной мутации (фиг. 10). С использованием Вестерн-блот-анализа как основы для детектирования мутантных линий ячменя, лишенных полосы белка LOX-1, эта частота в первой генерации обратного скрещивания соответствовала 3 линиям из всех 12 обратно скрещенных линий. Во второй генерации обратного скрещивания 9 линий из всех 28 обратно скрещенных линий были лишены полосы белка LOX-1 в Вестерн-блот-анализе (фиг. 10). Поскольку фон повторяющегося родителя увеличивается до приблизительно 75% в потомстве второго скрещивания, совместное наследование мутированного гена для LOX-1 и соответствующего нуль-LOX-1-фенотипа обеспечивало подтверждение их генетического сцепления. Критерий хи-квадрат выявил, что наблюдаемые результаты могли быть отнесены к категории статистически значимых. Величина р была низкой ( <3,84), свойство, которое показало значимость для первой, второй, третьей и четвертой генерации скрещивания. Программа обратного скрещивания продемонстрировала, что мутантный аллель, сообщающий нуль-LOX-1-фенотип, может быть перенесен в альтернативный генетический фон и что он наследовался рецессивным монофакториальным образом согласно менделевскому расщеплению. Пример 5. Затирание. Получение сусла. Для испытания свойств новых сортов ячменя из них получали пробы солода 25-225 г (см. фиг. 11). С использованием лабораторной системы затирания, которая содержала наружный смеситель и водяную баню, снабженную термостатом, способным понижать температуру в виде хорошо определенного градиента, затирание выполняли в малом масштабе. Готовое сусло фильтровали с использованием бумажного фильтра. Кипячение сусла выполняли в лабораторном масштабе с использованием нагревательной рубашки и круглодонной колбы, соединенной с обратным холодильником. Пример 6. Получение солода и пива с использованием солода ячменя дикого типа и мутантного ячменя. Ячмень сорта Barke и мутант D112 размножали в поле в течение нескольких сезонов для получения достаточного зернового материала для солодоращения и пивоварения. Анализ готового пива на T2N, а также органолептический анализ показали улучшенную стабильность вкуса пива, сваренного с солодом мутанта D112. Солодоращение зерен, полученных из мутанта D112 и сорта Barke. Солодоращение выполняли в масштабе 20 кг в солодовне следующим образом: зерно мутанта D112 (собранное в 2003 году), зерно сорта Barke (собранное в 2002 году). Условия замачивания: 8 ч замоченное; 14 ч сухое; 8 ч замоченное; 10 ч сухое; 4 ч замоченное в замочной воде при 16 °С. Условия солодоращения: 12 ч при 18 °С; 24 ч при 16 °С; 24 ч при 14 °С; 60 ч при 12 °С; Условия сушки: 12 ч при 60 °С; 3 ч при 68 °С; 4 ч при 74 °С; 3 ч при 80 °С. Результаты анализов солодоращения с использованием проб солода, полученного из мутанта D112, и солода сорта Barke, сравниваются в табл. 3. Эти результаты показали, что солод мутанта D112 и сорта Barke удовлетворяли спецификациям солода, и подтвердили, что эти солоды были пригодны для пивоварения. Значимое уменьшение уровней T2N наблюдали в солоде из мутанта D112 при сравнении с солодом сорта Barke, соответствующее уменьшению ~64% (табл. 4). Пивоварение с солодом мутанта D112 и сорта Barke. Пивоварение выполняли в масштабе 50 л, и оно включало следующие стадии: (i) получение сусла; (ii) отделение сусла; (iii) кипячение сусла; (iv) ферментирование; (v) выдержка; (vi) фильтрование светлого пива; и (vii) розлив в бутылки. Сусло получали с использованием солода мутанта D112 или солода сорта Barke, причем последний использовали в качестве эталонной пробы. Для каждой варки использовали всего 13,5 кг солода. Затирание выполняли при 47 °С в течение 20 мин с последующим нагреванием в течение 18 мин, в котором температуру повышали с 48 °С до 67 °С; затем 30 минутная пауза при 67 °С; затем нагревание до 12 °С в течение 5 мин; затем 15-минутная пауза при 72 °С; нагревание до 78 °С в течение 6 мин; 5-минутная пауза при 78 °С. Стадии пивоварения фильтрование и кипячение сусла, отделение гидроциклонным сепаратором, ферментирование, выдерживание и упаковка в зеленые стеклянные бутылки выполняли в соответствии со спецификациями для стандартной практики пивоварения. Всего было розлито в бутылки 33 л пива. Стабильность вкуса и анализы T2N. Пиво получали с использованием солода мутанта D112 и сорта Barke, как описано выше. Свежерозлитое в бутылки пиво хранили при 5 °С и анализировали в пределах 2 месяцев от получения. Стабильность вкуса свежего и подвергнутого хранению пива оценивали в соответствии с двумя различными типами условий хранения. В одной экспериментальной серии пиво подвергали процессу форсированного старения при 37 °С в течение периода 1-4 недель. Уровни T2N проб пива определяли газовой хроматографией с масс-спектрометрическим детектированием после дериватизации карбонилов О-(2,3,4,5,6-пентафторбензил)гидроксиламином, по существу, как описано Gronquvist et al. (1993). Квалифицированная дегустационная комиссия по оценке вкуса пива давала общую балльную оценку вкуса этого пива. Следует отметить, что оба типа свежего пива содержали сходные уровни сульфита, а именно, 4 м.д. и 5 м.д. сульфита для пива, полученного из солода мутанта D112 и сорта Barke, соответственно. Форсированное старение. Розлитое в бутылки пиво, полученное из солода сорта Barke, и розлитое в бутылки пиво, полученное из мутанта D112, испытывали и сравнивали в отношении конкретных результатов по развитию свободного T2N во время форсированного старения, как показано на фиг. 12А и в табл. 5. Видно, что эти типы пива могут быть отличены по явно выраженным различиям в кинетике развития T2N. Хотя эталонное пиво вело себя, как ожидалось, неожиданное и явно низкое развитие T2N наблюдали в пиве, полученном из мутанта D112, соответствующее 0,01 м.д. на протяжении 4 недель при 37 °С. Эксперимент по форсированному старению подчеркнул различие между двумя типами пива. Уже после 2,5 недель уровень T2N эталонного пива превышал порог вкусового ощущения, тогда как уровень T2N пива, полученного с использованием солода мутанта D112, становился равным концентрации T2N 0,025 м.д. после 2-3 недель инкубирования. Что касается стабильности вкуса и запаха, комиссия по качеству вкуса и запаха оценивала пиво, полученное с использованием солода нуль-LOX-1-мутанта ячменя D112. В центре внимания были пробы пива, которые были подвергнуты форсированному старению при 37 °С. Дегустационная комиссия нашла удовлетворительными профили вкуса и запаха для обоих типов свежего и подвергнутого форсированному старению пива (1 неделя при 37 °С). Однако балльные оценки в отношении бумажного привкуса были более высокими для эталонного пива, чем пива, полученного с использованием солода нуль-LOX-1-мутанта D112 (табл. 5), т.е. эталонное пиво имело более сильный указанный привкус. В целом, дегустационная комиссия предпочла пиво, полученное из солода нуль-LOX-1-мутанта D112 (балльная оценка одобрения вкуса и запаха, табл. 5). После инкубирования при 20 °С в течение 12 месяцев дегустационная комиссия из 10 дегустаторов пива, которые были специалистами и были обучены дегустации привкусов пива, сравнивала типы пива, полученные из солода нуль-LOX-1-мутанта D112 и контрольного солода. Все оценки - включающие такие вкусовые характеристики, как "бумажный", "окисленный", "подвергнутый старению", "хлебный", "карамельный", "пригорелый" и "сладкий" - выявила более высокие уровни специфических для старения привкусов в контрольных типах пива, чем в типах пива, полученных из нуль-LOX-1-солода (фиг. 12В). И с использованием балльной оценки по шкале от 0 до 5, где предпочтительными являются высокие величины, общая балльная оценка одобрения была оценена как 1,0 и 2,0 для контрольного пива и пива, полученного с солодом нуль-LOX-1-мутанта D112 ячменя, соответственно. В целом, улучшенная стабильность вкуса пива, полученного из солода мутанта D112 ячменя, является замечательной, прежде всего вследствие низких уровней T2N в пиве после хранения при 37 °С. Испытание пивоварения, которое имело в центре внимания применение солода нуль-LOX-1-ячменя, обеспечила доказательство того, что действие фермента LOX-1 ячменя во время процесса солодоращения и варки является ключевой причиной появления T2N, главного соединения привкуса в подвергнутом старению пиве. Пример 7. Тригидроксиоктадеценовые кислоты в пиве из нуль-LOX-1-солода. Специфические для пива тригидроксиоктадеценовые кислоты (THA; могут быть также сокращены как ТНОЕ), полученные из линолевой кислоты, были описаны 30 лет назад (Drost et al., 1974). С тех пор различные сообщения подтвердили, что общее содержание THA в пиве находится в диапазоне 5,7-11,4 мкг/мл (Hamberg, 19 91; и ссылки в этой работе). В то время как 9,12,13-THA в норме составляет 75-85% THA в пиве, 9,10,13-THA составляет обычно только 15-25%. Другие изомеры обнаруживаются в следовых количествах. В пиве, приготовленном из солода мутанта ячменя D112 (т.е. нуль-LOX-1-солода), концентрация 9,12,13-THA уменьшалась до 20% (т.е. почти в 5 раз) в сравнении с эталонным пивом, приготовленном из солода сорта Barke (табл. 6), т.е изомеры 9,12,13-THA и 9,10,13-THA присутствуют в почти равных количествах в пиве, полученном с использованием солода нуль-LOX-1-мутанта D112. Эти измерения проводили с использованием стандартных анализов ВЭЖХ-масс-спектрометрии. Пример 8. Тригидроксиоктадеценовые кислоты в пиве. Концентрация THA в большом разнообразии проб коммерчески доступного пива показана в табл. 7. Внимательное исследование этого результата на THA в пробах пива, как показано в табл. 7, обнаружило, что отношение 9,12,13-THA: 9,10,13-THA всегда превышало 3,5. В отличие от этого в пиве, полученном из D112, отношение 9,12,13-THA: 9,10,13-THA равно 1,3. Таким образом, пиво, полученное из нуль-LOX-1-ячменя, содержит значительно более низкое отношение, и определение отношения 9,12,13-THA: 9,10,13-THA обеспечивает инструмент для определения, получено ли пиво с использованием солода нуль-LOX-1-мутантов ячменя, например, солода мутанта ячменя D112. Следует отметить, что пиво, полученное из солода, полученного из нуль-LOX-1-мутанта ячменя D112, имеет значительно более низкий уровень общих 9,12,13-THA в сравнении с пивом, полученным из обычного солода. Пример 9. Биохимическая характеристика ферментативных продуктов из действия LOX в ячмене. Зрелые зерна ячменя дикого типа содержат две основные активности LOX, происходящие из ферментов LOX-1 и LOX-2. Эти ферменты катализируют диоксигенирование линолевой кислоты до гидропероксиоктадекадиеновых кислот (HPODE), причем фермент LOX-1 катализирует образование 9-HPODE, а фермент LOX-2 катализирует образование 13-HPODE. В зрелом зерне активность LOX заключена в зародыше. Для исследования, как действуют мутации в гене LOX-1 на образование HPODE, экстракты зародышей из сорта Barke и аналогичные экстракты из линии G ячменя (зерна с низкой LOX, РСТ-заявка PCT/IB01/00207, опубликованная в виде WO 02/053721A1, Douma et al.), а также экстракты зародышей нуль-LOX-1-мутанта D112 исследовали с использованием анализа жидкостной хроматографии высокого давления (ЖХВД). Зародыши ячменя. Препараты экстрактов неочищенного белка из зародышей получали сначала отсечением органов от зрелых зерен ячменя скальпелем для разреза между щитком и эндоспермом. Затем каждую пробу, состоящую из 4 зародышей, помещали между двумя кусочками бумаги для взвешивания и осторожно отбивали с получением гомогенной муки. Ее переносили в микроцентрифужную пробирку на 1,5 мл, добавляли 600 мкл 200 мМ содержащего молочную кислоту буфера, рН 4,5, и эту пробирку помещали на лед и выдерживали в течение 10 мин перед последующей гомогенизацией с использованием пластикового пестика (Kontes). Затем в каждую пробирку добавляли 600 мкл воды, и эти пробы центрифугировали в течение 2 мин при 20000 ×g. Аликвоту 100 мкл полученного супернатанта переносили в центрифужную пробирку на 15 мл [Cellstar (Cat. # 188271), купленную из Greiner Bio-One] для приготовления для анализа на продукты реакции после действия LOX. Готовили 2 мл 100 мМ натрий-фосфатного буфера, рН 6,5, содержащего 260 мкМ линолевую кислоту [этот субстрат готовили смешиванием 10 мл 100 мМ натрий-фосфатного буфера, рН 6,5, с 100 мкл 24 мМ исходного раствора линолевой кислоты. Последний раствор готовили сначала смешиванием 155 мкл линолевой кислоты (соответствующих 134 мг свободной кислоты; L-1376, Sigma) и 257 мкл Твина-20 с добавлением Н 2 О до объема 5 мл, с последующим добавлением 600 мкл 1 М NaOH, и, когда этот раствор становился прозрачным, конечный объем доводили до 20 мл Н 2 О]. После 15-минутного инкубирования на ротационном шейкере добавляли 2 мл этилацетата и содержимое пробы перемешивали интенсивным встряхиванием в течение 5 с для экстракции 9-HPODE и 13-HPODE. Затем эту пробу центрифугировали в течение 10 мин при 800 ×g и 1 мл этилацетата переносили в микроцентрифужную пробирку на 1,5 мл, в которой этилацетат выпаривали под током газообразного азота. Затем HPODE ресуспендировали в 300 мкл метанола и фильтровали через мембрану 0,45 мкм (Millex-HN filter, Millipore). Анализ содержания HPODE выполняли при помощи ВЭЖХ. В целом 15 мкл из каждой пробы инжектировали в ВЭЖХ-прибор (HP 1100 Series, Hewlett Packard), снабженный колонкой С184,6 ×250 мм Symmetry (Waters). Используемой подвижной фазой была смесь: вода : метанол : ацетонитрил : тетрагидрофуран : трифторуксусная кислота 16:12:12:10:0,5 (о:о:о:о:о). Скорость тока подвижной фазы была 1 мл/мин и давление, измеренное перед колонкой, было 140 бар. Разделение выполняли при 30 °С. Стандартная проба содержала смесь 9(S)-гидроперокси-10(Е),12(Z)-октадекадиеновой кислоты [(9(S)-HPODE] и 13(S)-гидроперокси-9(Z),11(Е)-октадекадиеновой кислоты [(13(S)-HPODE], как показано подробно на фиг. 13А. Анализы хроматограмм выявили, что в основном 9-HPODE образовывалась ферментами LOX, экстрагированными из зрелых зародышей ячменя сорта Barke (фиг. 13В), в то время как как 9-, так и 13-HPODE образовывались в экстрактах зрелых зародышей линии G с низкой LOX (фиг. 13С). Таким образом, экстракты зародышей мутанта D112 образовывали гораздо меньше 9-HPODE, чем экстракты зародышей линий ячменя дикого типа. Солод ячменя. Солод ячменя содержит две основные активности LOX, происходящие из LOX-1 и LOX-2. При этом LOX-1 катализирует образование 9-HPODE, а действие LOX-2 дает 13-HPODE. Для исследования действия мутаций в кодирующих LOX генах на образование HPODE в экстрактах мальта выполняли ВЭЖХ-анализы с экстрактами, полученными из солода, приготовленного из сорта Barke, линии G ячменя с низкой LOX и мутанта D112. Пробы экстракта неочищенного белка из солода готовили следующим образом. Одно зерно подвергнутого солодоращению (соложеного) ячменя помещали между двумя кусочками бумаги для взвешивания и осторожно отбивали с получением гомогенной муки. Все последующие манипуляции, смеси для инкубирования и способа ВЭЖХ-анализа были идентичными манипуляциям, описанным в предыдущем разделе этого примера, относящемся к измерениям продуктов LOX в экстрактах зародышей. Для ВЭЖХ-анализа использовали стандартную смесь 9(S)-гидроперокси-10(Е),12(Z)-октадекадиеновой кислоты [(9(S)-HPODE] и 13(S)-гидроперокси-9(Z),11(Е)-октадекадиеновой кислоты [(13(S)-HPODE], как показано на фиг. 14А. Анализ образующей HPODE активности в экстрактах подвергнутого солодоращению сорта Barke, линии с низкой LOX и D112 показал распределение 60:40 образующих 9- и 13-HPODE активностей в солоде сорта Barke (фиг. 14В), некоторую образующую 9-HPODE активность в солоде, полученном из ячменя с низкой LOX (фиг. 14С) и очень низкие уровни образования 9-HPODE в солоде мутанта D112 (фиг. 14D). Эти данные показывают явно выраженное более низкое образование 9-HPODE в экстрактах солода мутанта D112, чем в экстрактах солода, полученного из других линий ячменя. Пример 10. Ген для LOX-1 в мутанте D112 ячменя является мутированным. Нуклеотидные последовательности гена для LOX-1 в мутанте D112 [SEQ ID NO:2] и в сорте Barke [SEQ ID NO:1] были получены, и их сравнивали для определения молекулярной основы для нуль-LOX-1-фенотипа мутанта D112, который, как было обнаружено, отличается отсутствием соответствующего фермента LOX-1 в зерне. Геномную ДНК ячменя из мутанта D112 и сорта Barke дикого типа выделяли из тканей листьев проростков с использованием набора для выделения ДНК растений (Roche Applied Science) в соответствии с рекомендациями изготовителя. Последовательность 4224 п.н., фланкирующую кодирующий белок район для LOX-1 в геномной ДНК мутанта D112 и сорта Barke, амплифицировали при помощи ПЦР с использованием праймеров Основой для этих праймерных последовательностей была геномная последовательность гена для LOX-1 (van Mechelen et al., 1995; Rouster et al., 1997; схематическое изображение этой геномной последовательности, включающей стартовый (инициирующий) кодон и стоп-кодон (терминирующий кодон), показано на фиг. 15). ПЦР-реакции состояли из 100 нг геномной ДНК в объеме 20 мкл, содержащем 5 пмоль каждого праймера и 3,5 Е полимеразы Expand High Fidelity (Roche Applied Science). ПЦР-амплификации проводили в циклере MJ, с использованием следующих параметров проведения циклов: 2 мин при 96 °С для 1 цикла; 1 мин при 95 °С, 1 мин при 69 °С и 5 мин при 72 ° С в течение 30 циклов; 10 мин при 72 ° С для 1 цикла. Продукты ПЦР разделяли на 1,0% агарозных гелях. ДНК-фрагменты, соответствующие по длине этому амплифицированному району, очищали с использованием набора для экстракции гелей Qiaex II (Qiagen) и встраивали в плазмидный вектор pCR2.1-TOPO (Invitrogen). Нуклеотидную последовательность обеих цепей кодирующих районов определяли с использованием реакции терминации цепи дидезоксинуклеотидом со специфическими олигонуклеотидными праймерами и анализировали на секвенаторе ДНК MegaBACE 1000 (Amersham). Сравнения последовательностей выполняли с использованием пакета программ для анализа последовательностей Lasergene ver.5 (DNASRAR). В прямом сравнении между последовательностью для LOX-1 сорта Barke дикого типа [SEQ ID NO:1] и мутанта D112 [SEQ ID NO:2] нуклеотидная последовательность этого мутанта обнаруживала одну точковую мутацию в форме замены G →А в положении 3574 в экзоне 7 (фиг. 15; фиг. 16). Последовательность дикого типа для LOX-1 кодирует белок с длиной 862 остатка [SEQ ID NO:3] с предсказанной массой 96,4 кДа. В отличие от этого, мутация в положении 357 4 в соответствующей последовательности мутанта D112 вызывает введение преждевременного стоп-кодона. Предсказывается, что этот стоп-кодон (термирирующий кодон) в кодирующем LOX-1 гене мутанта D112 приводит к С-концевому укорочению из 197 аминокислот соответствующего белка и, следовательно кодированию белка 74,2 кДа, последовательность которого представлена в [SEQ ID NO:4]. Пример 11. Ген для LOX-1 в мутанте ячменя А618 является мутированным. Получение геномной ДНК, ПЦР-реакции и определение последовательности ДНК и анализы геномной ДНК мутанта ячменя А618 и сорта Neruda дикого типа были идентичными этим манипуляциям, описанным для мутанта D112 и сорта Barke в примере 10. Сравнение нуклеотидной последовательности для LOX-1 мутанта ячменя А618 [SEQ ID NO:6] с последовательностью родительского сорта Neruda [SEQ ID NO:5] показало, что последовательность этого мутанта имела одну точковую мутацию, соответствующую замене G →A в положении 2311 в этой геномной последовательности (фиг. 15; фиг. 16). Последовательность дикого типа для LOX-1 сорта Neruda кодирует белок с длиной 862 остатка [SEQ ID NO:7] с предсказанной массой 96,4 кДа. В отличие от этого, мутация в положении 2311 в соответствующей последовательности мутанта А168 мутирует донорный сайт интрона 3. Это вызывает ошибку сплайсинга в интроне 3, приводящую к преждевременному стоп-кодону в интроне 3 после трансляции 399 аминокислот. Стоп-кодон в рамке считывания в гене для LOX-1 мутанта А618 будет приводить также к укороченному транслируемому белку 44,5 кДа [SEQ ID NO:8]. Пример 12. ОТ-ПЦР-детектирование транскриптов для LOX-1. Растения ячменя сорта Vintage, мутантной линии G (с низкой LOX-1, РСТ-заявка PCT/IB01/00207, опубликованная в виде WO 02/053721А1, Douma et al.), сорта Barke и мутанта D112 выращивали в оранжерее во время весны 2002 года в Копенгагене, Дании. Колосья метили этикетками в день зацветания и собирали в дни 20, 40 и 60 после зацветания (DAF). Эти колосья хранили при -80 °С, пока все пробы не были собраны для одновременной обработки. В целом 10 зародышей на одну временную точку отсекали из развивающейся зерновки и РНК экстрагировали с использованием набора для выделения FastRNA, Green RNA (Qbiogene) с использованием рекомендаций изготовителя. Матрицы для ОТ-ПЦР-реакций состояли из 100 нг РНК вышеописанных зародышей. ОТ-ПЦР-реакции 20 мкл содержали 50 пмоль каждого праймера и 5 Е смеси ферментов для ОТ-ПЦР (Promega). ОТ-ПЦР-амплификации в циклере MJ: 45 мин при 48 °С для 1 цикла; 1 мин при 95 °С для 1 цикла, 1 мин при 94 °С, 1 мин при 65 °С и 1 мин при 72 °С в течение 30 циклов; и, наконец, 10 мин при 72 °С для 1 цикла. Прямой праймер, Полученный ПЦР-продукт включал район, соответствующий положениям нуклеотидов 3283-3659 в геномном клоне [SEQ ID NO:1]. Этот район содержал интрон 5 с длиной 83 п.н., который отсутствовал в матрице ОТ-ПЦР в не содержащем ДНК препарате РНК (фиг. 17А). Поскольку анализ последовательности ДНК подтвердил, что выделенный фрагмент был интегральной частью гена для LOX-1, и подтвердил отсутствие последовательности интрона 5, можно было исключить предположение, что неправильная амплификация давала фрагменты из гена ячменя для фермента LOX-2 (фиг. 17D). Таким образом, этот амплифицированный фрагмент представлял продукт амплификации РНК-транскрипта. Сравнительный ОТ-ПЦР-анализ РНК, очищенной из зародышей ячменя дней 20, 40 и 60 после зацветания (DAF) сорта Vintage и мутантной линии, G выявил, что уровни транскриптов для LOX-1 являются сходными для этих двух сортов на сходной стадии развития. Уровень транскрипта для LOX-1 постепенно увеличивался в период времени от 20 до 60 DAF (фиг. 17В). Однако в отличие от этого, заметное различие наблюдали, когда исследовали сходный набор данных для сорта Barke и мутанта D112. В этом случае, ОТ-ПЦР-эксперименты выявили, что LOX-1-транскрипты в мутанте D112 были значимо более низкими по относительному количеству в сравнении с LOX-1-тарнскриптами сорта Barke (фиг. 17С). В целом эти наблюдения могут объясняться потенциальной мутацией в районе промотора гена для LOX-1 в мутанте D112. Другие, еще неизвестные факторы могут участвовать в регуляции транскрипции гена для LOX-1 в мутанте D112. В этой связи, нельзя исключить, что стоп-кодон (терминирующий кодон) в транскрипте для LOX-1 мутанта D112 вызывает опосредованное антисмысловой последовательностью уменьшение мРНК (Isshiki et al., 2001). Пример 13. Генетическое детектирование мутантов ячменя, несущих мутацию D112. Современные стратегии селекции ячменя часто включают биотехнологические способы для ускорения процесса мутагенеза в целях организации серийного производства. Таким образом, может быть полезным проведение раннего скрининга растительного материала в отношении детектирования полиморфизмов отдельных нуклеотидов в представляющих интерес генах. С использованием этого способа с геномной ДНК и объединением его с высокопроизводительной системой можно сузить число линий селекции с 50% на стадии проростков. CAPS-анализы. Клонирование и секвенирование гена для LOX-1 линий потомства мутанта D112 показали, что эта мутация передается следующей генерации. Этот способ является трудоемким и не применим для практической селекции ячменя. Мутация, специфическая для линии G с низкой LOX, могла быть идентифицирована в селекционном материале с использованием анализа расщепленной амплифицированной полиморфной последовательности (CAPS-анализа), как описано в примере 4 РСТ-заявки PCT/IB01/00207, опубликованной в виде WO 02/053721А1, Douma et al. Однако природа этой мутации в гене для LOX-1 мутанта D112 не может быть использована для генерирования измененной рестрикционной карты в районе 60 п.н., содержащем эту мутацию. SNP-анализы. Альтернативным решением для этого является выполнение анализа, включающего определение полиморфизма единственного нуклеотида (SNP). SNP является точкой мутации с по меньшей мере двумя различными нуклеотидами, представленными в одном локусе. Этот анализ основан на комбинировании двух серий геномных ПЦР-реакций. Обе реакции содержат локус-специфический праймер и один из двух SNP-праймеров (по одному для каждого аллеля этой последовательности). Две серии ПЦР-реакций осуществляют на одну линию растений и результат ПЦР-реакции заключается в том, что SNP-праймер связывается либо с последовательностями мутанта, либо с аллелем дикого типа (фиг. 18А). В одном из нескольких способов, SNP-анализ может быть основан на идентификации мутантных линий посредством оценки картины распределения полос (бэндинга) после электрофореза ПЦР-продуктов. Геномную ДНК ячменя из 17 линий селекции и из сорта Barke дикого типа выделяли из тканей листьев проростков с использованием набора для выделения ДНК растений (Roche Applied Science) в соответствии с рекомендациями изготовителя. Олигонуклеотидными праймерами, используемыми для амплификации SNP гена LOX-1 дикого типа, были Для соответствующего гена мутанта D112 этими праймерами были Эти комбинации праймеров использовали в ПЦР-реакциях для амплификации ДНК-фрагментов 166 п.н., содержащих части кодирующих районов для LOX-1 мутанта D112 или сорта Barke (фиг. 18А). ПЦР-реакции состояли из 100 нг геномной ДНК в объеме 20 мкл, содержащем 25 пмоль праймера и 2,5 Е полимеразы FastStart Taq (Roche), с использованием в соответствии с инструкциями изготовителей. ПЦР-амплификации проводили в циклере MJ: 5 мин при 96 °С для 1 цикла; 1 мин при 95 °С, 1 мин при 70 °С, 1 мин при 72 °С течение 20 циклов; и, наконец, 10 мин при 72 °С для 1 цикла. Продукты ПЦР разделяли на 1,0% агарозных гелях. ДНК-фрагменты, соответствующие по длине этому амплифицированному району, очищали с использованием набора для экстракции гелей Qiaex II (Qiagen). Продукты ПЦР секвенировали с использованием реакции терминации цепи дидезоксинуклеотидом на генетическом анализаторе ABI Prism. Сравнения последовательностей выполняли с использованием пакета программ для анализа последовательностей Lasergene ver.5 (DNASRAR). Компилирование данных из скрининга в целом 17 линий селекции при помощи SNP-анализа, а также экспериментальных данных из прямого секвенирования продуктов ПЦР давало идентичные результаты. На основании этих экспериментов был сделан вывод о том, что способ SNP может быть использован для подтверждения, что исходный материал содержит последовательность гена, идентичную последовательности гена для LOX-1 мутанта ячменя D112 (фиг. 18В). Пример 14. Детектирование мутантов в пробах смешанного солода. Пивоваренная промышленность может использовать смеси ячменя и солода для получения пива, и это может маскировать нежелательные химические характеристики конкретной разновидности солода. Простое подтверждение применения конкретного материала семян может включать амплификацию мутантного гена с использованием ПЦР-анализа. SNP-анализ проб смешанного солода выполняли с использованием пробы смеси мутанта D112 и сорта Barke и пробы смеси мутантной линии G (РСТ-заявки PCT/IB01/00207, опубликованной в виде WO 02/053721A1, Douma et al.) и сорта Barke. Анализировали шесть проб ячменя, содержащих 0, 20, 40, 60, 80 и 100% зерен мутанта D112. В других сериях, анализировали шесть проб ячменя, содержащих 0, 20, 40, 60, 80 и 100% зерен мутантной линии G. ДНК выделяли из размолотых зерен с использованием набора для выделения ДНК Nucleon Phytopure (Amersham), в соответствии с рекомендациями изготовителя. Олигонуклеотидными праймерами, используемыми для амплификации SNP 166 п.н. гена LOX-1 мутанта D112 были Праймерами, используемыми для амплификации SNP 370 п.н. гена для LOX-1 мутантной линии G, были ПЦР выполняли в виде многократных реакций с использованием одновременно этих четырех праймеров (фиг. 19А). Каждая реакция содержала 100 нг геномной ДНК в объеме 20 мкл, содержащем 50 пмоль каждого праймера и 10 мкл раствора полимеразы RedTag (Sigma) в соответствии с инструкцией, обеспеченной поставщиком этого фермента. ПЦР-амплификации проводили в в циклере MJ: 1 мин при 95 °С для 1 цикла, 1 мин при 94 °С,1 мин при 66 °С, 30 с при 72 °С течение 25 циклов; и, наконец, 10 мин при 72 °С для 1 цикла. Продукты ПЦР разделяли на 1,0% агарозных гелях. ДНК-фрагменты, соответствующие по длине этому амплифицированному району, очищали с использованием набора для экстракции гелей Qiaex II (Qiagen) и встраивали в плазмидный вектор pCR2.1-TOPO (Invitrogen). Нуклеотидную последовательность обеих цепей плазмидных инсертов определяли с использованием реакции терминации цепи дидезоксинуклеотидом со специфическими олигонуклеотидными праймерами и анализировали на секвенаторе ДНК MegaBACE 1000 (Amersham). Сравнения последовательностей выполняли с использованием пакета программ для анализа последовательностей Lasergene ver.5 (DNASRAR). Этот анализ гелей, представленный на фиг. 19В, показал положительный анализ SNP для всех проб, полученных из смесей, содержащих зерна мутанта D112. Подобным образом, можно было идентифицировать пробы, содержащие материал из мутантной линии G. В целом, генетические анализы могут легко подтверждать применение смесей ячменя, содержащих мутантные LOX-1-растения либо мутантной линии G, либо мутанта D112. Пример 15. Рекомбинантный LOX-1 мутанта D112 является неактивным. Было показано, что LOX-1 мутанта D112 содержит преждевременный стоп-кодон (см. пример 19). Таким образом, ожидалось, что экспрессия in planta этого гена приводит к синтезу укороченной версии соответствующего фермента LOX, содержащего только первые 665 аминокислотных остатков, обнаруживаемых в LOX-1 дикого типа. Нуклеотидную последовательность этой укороченной версии LOX-1 экспрессировали в клетках Е. coli для подтверждения, что она является ферментативно неактивной и не может катализировать образование HPODE в клетках мутанта ячменя D112. Плазмиды для экспрессии LOX-1 дикого типа и мутантного LOX-1 в Е. coli. Полную открытую рамку считывания, кодирующую LOX-1, амплифицировали с использованием стандартного протокола ПЦР. Матрицей была кДНК ячменя (van Mechelen, 1999) и используемыми праймерами были: стартовый кодон отмечен жирным шрифтом; сайт NdeI подчеркнут] и комплементарная; стоп-кодон дикого типа отмечен жирным шрифтом; сайт EcoRI подчеркнут]. Амплифицированный ДНК-фрагмент 2597 получали и очищали. Этот ПЦР-фрагмент расщепляли NdeI-EcoRI и лигировали с большим NdeI-EcoRI-фрагментом вектора рЕТ19b (Novagen) с получением экспрессионной плазмиды pETL1, в которой ген для LOX-1 клонирован справа, в рамке считывания последовательности для имеющего 10 остатков His-конца. Анализы секвенирования ДНК подтвердили, что инсерт этой плазмиды содержал правильную последовательность. Следующие эксперимент включал конструирование плазмиды для экспрессии укороченной версии LOX-1. Цель состояла в изменении кодона 666 открытой рамки считывания для LOX-1 pTNL1 на стоп-кодон, так чтобы белковый синтез в Е. coli генерировал укороченную версию LOX-1. Для полного предотвращения прочитывания через этот стоп-кодон рибосомами в Е. coli конструировали экспрессионную плазмиду, в которой все кодоны справа от кодона для положения 665 LOX-1 в pETL1 удаляли и заменяли стоп-кодоном TGA. Использовали следующий протокол. Фрагмент 129 п.н. амплифицировали из pETL1 с использованием ПЦР в присутствии праймеров с отжигом слева от мутации в гене для LOX-1 мутанта D112; сайт BSlWI подчеркнут) и сайт EcoRI подчеркнут; жирные буквы указывают комплементарную последовательность введенного стоп-кодона). Эта амплификация вводила стоп-кодон и сайт EcoRI в этот фрагмент. Затем его расщепляли BslWI-EcoRI и лигировали с большим BslWI-EcoRI-фрагментом плазмиды pETL1. Полученная экспрессионная плазмида была названа pETL2, и правильная последовательность этого инсерта была подтверждена секвенированием ДНК. Трансформированные клетки Е. coll синтезируют рекомбинантные белки LOX. Клетки Е. coli BL21, купленные из Novagen, по отдельности трансформировали вектором рЕТ19b и экспрессионными плазмидами pETLl и pETL2 (описанными выше). Бактериальные клетки, несущие эти плазмиды, инокулировали в стандартную LB-среду (бульон Луриа-Бертани) и выращивали в течение 2 ч при 37 °С. После этого, добавляли 1 мМ IPTG для индукции экспрессии гетерологичных генов и культуры выращивали в течение ночи при 20 °С. Клетки собирали центрифугированием в течение 1 мин при 14000 ×g, с последующим ресуспендированием осадков клеток в растворе для денатурации, состоящем из 50 мМ натрий-фосфатного буфера, дополненного 6 М гидрохлоридом гуанидина, 0,3М NaCl и 10 мМ имидазолом. После обработки ультразвуком на льду лизированные клетки центрифугировали в течение 1 мин при 14000 ×g и супернатант смешивали с Ni-смолами (Novagen) с последующим 30-минутным инкубированием при 4 °С. Ni-смолы осаждали центрифугированием и промывали дважды раствором для денатурации, описанным выше. Наконец, His-меченые белки элюировали дважды из этих смол с использованием 50 мМ Na-фосфатного буфера, дополненного 0,3 М NaCl и 0,5 М имидазолом. Аликвоты фракционированных, элюированных проб разделяли электрофорезом на ДСН-ПААГ (фиг. 20). Отчетливые полосы ~100 кДа, соответствующие LOX-1, и ~66 кДа, соответствующая рассчитанной массе укороченного LOX-1, получали из клеток, несущих pETL1 и pETL2, соответственно. Клетки, несущие pET19b, не давали полос в элюированных фракциях. Укороченная версия LOX-1 является неактивной. Е. coli BL21, несущую pET19b, pETL1 и pETL2, инокулировали в стандартной среде LB и выращивали в течение 2 ч при 37 °С. После этого добавляли 1 мМ IPTG для индукции экспрессии гетерологичных генов и культуры выращивали в течение ночи при 20 °С. Клетки собирали центрифугированием в течение 1 мин при 14000 ×g. Клеточные лизаты получали ресуспендированием осадков клеток в смеси BugBuster и Benzonase (Novagen). Активность LOX измеряли в этих лизатах с использованием реагента для анализа липоксигеназы, содержащего 6,25 мМ 3-диметиламинобензойную кислоту, 0,3125 линолевую кислоту, 0,1 мМ 3-метил-2-бензотиазолингидразон и 0,05 мг/мл гемоглобина. 180 мкл этого реагента смешивали с 10 мкл соответствующего клеточного лизата и инкубировали 10 мин при комнатной температуре. Количество индамина, продуцированного во время этого инкубирования, определенное спектрофотометрически в виде оптической плотности при 595 нм, соответствует липоксигеназной активности клеточного лизата. В то время как клетки, трансформированные pETL1 (продуцирующие His-меченый фермент LOX-1), показали высокую активность LOX-1, клетки, продуцирующие мутантный, D112-специфический укороченный LOX-1, имели такую же активность LOX, что и контрольные клетки, трансформированные только вектором (табл. 8). Это показывает, что укороченный LOX-1 мутанта ячменя D112 является неактивным. Пример 16. Трансгенные растения ячменя. Плазмидные конструкции. Последовательности генов встраивают в полилинкерный район стандартных плазмидных векторов, таких как pUC18. Эти инсерты приведены на фиг. 21. В одной конструкции (фиг. 21А) промотор убиквитина-1 кукурузы (Christensen et al., 1992; Jensen et al., 1996) - включающий интрон 1 того же самого гена - управляет транскрипцией гена bar (White et al., 1990), который кодирует селектируемый маркер, фосфинотрицинацетилтрансферазу (PAT). Во второй конструкции, сконструированной для смысловой супрессии гена для LOX-1 (Dougherty and Parks, 1995), открытая рамка считывания для LOX-1 ячменя встроена сразу же справа от промотора убиквитина-1 кукурузы и интрона 1 (фиг. 21В). Конструкция для сайленсинга экспрессии гена ячменя для LOX-1 показана на фиг. 21С. Экспрессия этой конструкции в клетках ячменя вызывает полный сайленсинг указанного гена посредством образования интрон-сплайсированной шпилечной РНК, и эта конструкция сконструирована в соответствии с данными, подробно описанными на фиг. 1а публикации Smith et al. (2000). Конкретно, последовательность, названная "интроном 1", этой конструкции на фиг. 21С является идентичной последовательности интрона, показанного на фиг. 1а публикации Smirh et al. (supra). Смысловое и антисмысловое плечи этой конструкции на фиг. 21С представляют противоположные направления одного и того же фрагмента с длиной ~200 п.н., содержащего сегмент открытой рамки считывания, кодирующей LOX-1 ячменя, причем указанный сегмент рамки считывания расположен в любом месте в открытой рамке считывания для LOX-1. Альтернативно, последовательность с длиной 200 п.н. выбрана из последовательности справа от стоп-кодона гена ячменя, кодирующего LOX-1. Трансформация и регенерация трансгенных растений. Незрелые зародыши ячменя из выращенных в оранжерее донорных растений ячменя сорта Golden Promise, бомбардировали смесью плазмид, содержащих инсерты, показанные на фиг. 21А,В, для косупрессии гена ячменя, кодирующего LOX-1, и смесью плазмид, показанных на фиг. 21А,С, для сайленсинга указанного гена. Трансформацию, отбор трансформированных клеток и размножение трансгенных растений выполняют, как подробно описано Wan and Lemaux (1994) и Jensen et al. (supra). Трансгенные растения выращивают в течение нескольких генераций или опыляют отличающимся сортом ячменя с последующей идентификацией растений-потомков с желаемым фенотипом. Могут выращиваться две или более генераций для гарантии того, что экспрессия желаемой фенотипической характеристики стабильно сохраняется и наследуется. Семена собирают для испытания того, что была получена эта желаемая фенотипическая характеристика. Для испытания эффектов косупрессии или сайленсинга гена ячменя, кодирующего LOX-1, трансгенные зерна сначала анализируют на ферментативную активность, проявляемую LOX-1, как подробно описано в примере 1 данного изобретения. Трансгенные зерна, не имеющие активности LOX-1 или имеющие очень низкую активность LOX-1, испытывают после этого в экспериментах по солодоращению и пивоварению, как подробно описано для нуль-LOX-1-зерен в примере 5 и примере 6 данной публикации. Кроме того, экстракты трансгенных зерен анализируют для идентификации экстрактов, проявляющих отрицательное действие на рост гриба Aspergillus, с использованием способов, описанных в Патенте США № 5942661, выданном Keller. Пример 17. Соединения ноты зелени. Способ получения соединений ноты зелени предусматривает: (i) превращение зерен нуль-LOX-1-ячменя в тонкоизмельченную муку; (ii) суспендирование этой муки в воду или указанный буфер; (iii) инкубирование этой суспензии; альтернативно, реакции этой суспензии муки с (а) жирной кислотой (линолевой кислотой или линоленовой кислотой или их смесью); или (b) ферментом гидроксипероксидлиазой, имеющей специфичность в отношении 13-HPODE или 13-НРОТЕ или обеих; или (с) смесью, содержащей указанные жирные кислоты и указанный фермент; (iv) реакцию полученных альдегидов с алкогольдегидрогеназой; (v) очистку альдегидов или спиртов и получение применимого препарата вкусовой или ароматизирующей композиции. Пример 18. Ингибиторы LOX-1. Для этих анализов использовали свежеприготовленный раствор рекомбинантного LOX-1. В этом эксперименте 100 мл среды для выращивания АВ3 - дополненной 100 мкг/мл ампициллина - готовили с использованием рекомендации поставщика (Remel) и затем инокулировали 5 мл ночной культуры клеток Е. coli BL21 (DE3)pLysS, трансформированных плазмидой pETL1 (кодирующей His-меченый LOX-1; см. пример 15). Полученную бактериальную культуру размножали при 37 °С, пока плотность клеток не достигала OD 600 =0,8. Эту культуру инкубировали при 20 °С в течение 30 мин, затем дополняли 0,4 мМ IPTG для индукции экспрессии гетерологичного гена и инкубировали при 20 °С в течение ночи. Клетки этой культуры осаждали 15-минутным центрифугированием, ресуспендировали в 5 мл BugBuster HT (Novagen) и инкубировали в течение 20 мин при осторожном встряхивании для гидролиза нуклеиновых кислот. После этого остатки клеток удаляли центрифугированием, супернатант осветляли фильтрованием через фильтр 0,45 мкм и добавляли к равному объему буфера для связывания (50 мМ Na-фосфатный буфер, рН 7,5, дополненный 0,3 М NaCl, 10 мМ имидазолом). Полученный экстракт наносили на колонку HisTrap HP (Amersham Bioscience) в соответствии с рекомендациями изготовителя), промывали один раз промывочным буфером (идентичным буферу для связывания, за исключением того, что концентрация имидазола = 150 мМ) и связанный белок элюировали буфером для элюции (идентичным буферу для связывания, за исключением того, что концентрация имидазола = 500 мМ). Аликвоты по 3 мкл фракций 1 мл, содержащих белки промывок и элюций, анализировали электрофорезом в ДСН-ПААГ (фиг. 22А) и показали, что фракция 1 мм элюата 2 содержала ~0,7 мг рекомбинантного LOX-1. Затем очищенный LOX-1 использовали в анализах для определения, уменьшают ли выбранные ингибиторы LOX эти ферментативные активности. Сначала, исходный раствор линолевой кислоты (приготовленный, как подробно описано в примере 9) разбавляли до 1/10 его исходной концентрации, с получением раствора 2,4 мМ линолевой кислоты. Аликвоты 45 мкл этого раствора дополняли 5 мкл этанольных растворов, содержащих 0, 5, 12 и 24 мМ октилгаллат или NDGA (предположительные ингибиторы LOX-1). Аликвоты по 10 мкл смесей линолеат-ингибитор добавляли к 990 мкл 100 мМ Na-фосфатного буфера, рН 6,0, и инкубировали в течение 1 мин при 20 °С перед добавлением 5 мкл рекомбинантного LOX-1 (элюат 2, см. выше). После добавления LOX-1, А 254 регистрировали на протяжении периода времени 3 мин. Активность фермента LOX-1 определяли в виде наклона графика с А 254 в зависимости от времени. Эти результаты суммированы на фиг. 22В и показывают явно выраженное ингибирование LOX-1 при микромолярных концентрациях этих ингибиторов. Пример 19. Затирание с октилгаллатом, ингибитором LOX-1. Затирания в малом масштабе 100 мл, содержащих 25 г солода ячменя сорта Barke или 25 г солода нуль-LOX-1-мутанта D112, выполняли с использованием оборудования, сходного с оборудованием, описанным в примере 5. Затирание происходило при 37 °С в течение 15 мин, осахаривание при 68 °С в течение 30 мин, прекращение затирания при 77 °С в течение 10 мин с последующим конечным кипячением сусла в течение 60 мин; смещения температуры корректировали до 1 °С на минуту. Для испытания действия затирания в присутствии ингибитора LOX-1 затор с солодом ячменя сорта Barke дополняли 0,5 мМ октилгаллатом в начале затирания. Параллельные затирания включают эксперименты с солодом ячменя сорта Barke без добавленного октилгаллата, а также затирания с солодом нуль-LOX-1-мутанта ячменя D112 в присутствии или в отсутствие 0,5 мМ октилгаллата. Аликвоты проб всех четырех затираний собирали после фазы 15-минутного затирания, после фазы кипячения сусла с последующим определением уровней T2N, как описано в примере 6. Эти результаты суммированы на фиг. 23. Заметное уменьшение T2N наблюдали в пробах сусла затирания с солодом ячменя сорта Barke в присутствии октилгаллата, как в пробе, анализированной после начала затирания, так и в пробе кипяченого сусла. Также следует отметить, что концентрация T2N в прокипяченном сусле обоих типов солода достигала сходных уровней. В целом, дополнение затора ингибитором LOX-1 октилгаллатом во время начала затирания обеспечивает новый способ получения сусла, которое характеризуется уменьшенным уровнем T2N. а По шкале 0-9, где 0 обозначает отсутствие инфицирования или полегания, а 9 обозначает сильное инфицирование или полегание. b Относительный, средний урожай из трех повторностей в двух различных местоположениях. а Шкала оценки - 0: не присутствует; 1: слабый; 2: заметный; 3: средний; 4: сильный; 5: чрезвычайно сильный; предпочтительными являются низкие величины. b Шкала оценки 1-5; предпочтительными являются высокие величины. nd = не определяли. a Результаты приведены в виде средней величины из четырех отдельных измерений с указанными вариациями. Депонирование составляющих собственность Carlsberg A/S мутантов ячменя D112, описанных выше и перечисленных в прилагаемой формуле изобретения, было произведено Американской Коллекцией Типовых Культур (АТСС), 10801 University Boulevard, Manassas, Va.20110, USA. Датой депонирования для мутанта D112 была дата 11 сентября 2003 года, и депозит состоял из 2500 зерен, взятых из депозита в Carlsberg A/S, перед датой подачи этой заявки. Эти депонирования были произведены в соответствии с условиями Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры и в соответствии с правилами этого договора (Будапештского договора). Это гарантирует поддержание жизнеспособной культуры депозита в течение 30 лет от даты депонирования. Предполагается, что эти депозиты удовлетворяют всем требованиям 37 C.F.R #1.801-1.809, в том числе обеспечению указания о жизнеспособности этих образцов. Мутант D112 имеет номер доступа АТСС РТА-5487. Мутант А618 имеет номер доступа АТСС РТА-5584. Аликвоты этого депонированного материала могут быть получены из АТСС при указании номера доступа и при соблюдении стандартных ограничений, налагаемых АТСС. Однако должно быть понятно, что доступность депозита не является лицензией для применения на практике данного изобретения в качестве нарушения патентных прав, предоставляемых решением правительства. Во всей этой заявке делаются ссылки на различные публикации, патенты и заявки на патент. Описания этих публикаций в их полном виде включены тем самым в качестве ссылки в данную публикацию для более полного описания состояния области, к которой относится данное изобретение. Предыдущее описание данного изобретения является примерным описанием для целей иллюстрации и объяснения. Должно быть понятно, что могут быть произведены различные модификации без отклонения от идеи и объема данного изобретения. Таким образом, следующая далее формула изобретения должна интерпретироваться как включающая все подобные модификации. Патентные документы: US 4683195; Mullis, К.В. et al. US 4708964; Allen, L.M. US 4800159; Mullis, K.B. et al. US 4880637; Jordan, R.T. US 5008294; Jordan, R.T. et al. US 5283184; Jorgensen, R.A. and Napoli, C.A. US 5942661; Keller, N.P. US 6008034; Hausler et al. US 6150145; Hausler, A. et al. US 6274358; Holtz, R.B. et al. US 6355862 B1; Handa, AK. and Kausch, K.D; US patent application 2003/0074693 A1; Cahoon et al. WO 02/053721 Al; 12/2000; PCT Int'l Appl.; Douma, AC. et al. WO 2004/085652 A1; 10/2004; PCT Int'l Appl.; Hirota, N. et al. Другие публикации: Alonso, J.M. et al., "Genome-wide insertional mutagenesis of Arabidopsis thaliana. " Science 301, 653-657, 2003. American Association of Cereal Chemists, "Approved methods of the American Association of Cereal Chemists." ISBN 0-913250-86-4 (1995). American Society of Brewing Chemists, "Methods of analysis of the American Society of Brewing Chemists." ISBN 1-881696-01-4 (1992). Anthon, G.E. and Barrett, D.M., "Colorimetric method for the determination of lipoxygenase activity." J. Agric. Food Chem. 49:32-37, 2001. Aruoma, O.I. et al., "Evaluation of the antioxidant and prooxidant actions of gallic acid and its derivatives." J. Agric. Food Chem. 41:1880-1885, 1993. Ashrafi, K. et al., "Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes." Nature 421:268-272, 2003. Auld, D.L. et al., "Rapeseed mutants with reduced levels of polyunsaturated fatty acids and increased levels of oleic acid." Crop Sci. 32:657-662, 1992. Axelrod, B. et al., "Lipoxygenase from soybeans." Methods Enzymol. 71:442-451, 1981. Bargmann, C.I., "High-throughput reverse genetics: RNAi screens in Caenorhabditis elegans." Genome Biol. 2:Reviews1005.1-1005.3, 2001. Baur, C. and Grosch, W. "Investigation about the taste of di-, tri- and tetrahydroxy fatty acids." Z. Lebensm. Unters. Forsch. 165: 82-84, 1977a. Baur, C. et al. "Enzymatic oxidation of linoleic acid: Formation of bittertasting fatty acids." Z. Lebensm. Unters. Forsch. 164:171-176, 1977b. Bell, E. et al., "A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis." Proc. Natl. Acad. Sci. USA 92:8675-8679, 1995. Bell, E. and Mullet, J.E., "Lipoxygenase gene expression is modulated in plants by water deficit, wounding, and methyl jasmonate." Mol. Gen. Genet. 230:456-462, 1991. Bell, E. and Mullet, J.E., "Characterization of an Arabidopsis lipoxygenase gene responsive to methyl jasmonate and wounding." Plant Physiol. 103:1133-1137, 1993. Bios International, "Data." Bios Intern. 4:38-42, 2001. Blee, E. and Joyard, J., "Envelope membranes from spinach chloroplasts are a site of metabolism of fatty acid hydroperoxides." Plant Physiol. 110:445-454, 1996. Bohland, C. et al., "Differential induction of lipoxygenase isoforms in wheat upon treatment with rust fungus elicitor, chitin oligosaccharides, chitosan, and methyl jasmonate." Plant Physiol. 114:679-685, 1997. Brautechnische Analysenmetoden, Band II, Metodensammlung der Mitteleuropaischen Brautechnischen Analysenkommission. Section 2.19: "Schaum," pp. 118-125. Selbstverlag der MEBAK. ISBN 3-9805814-5-4 (2002). Burow, G.B. et al., "A peanut seed lipoxygenase responsive to Aspergillus colonization." Plant Mol. Biol. 42:689-701, 2000. Casey, R., "Lipoxygenases in the breadmaking process." In: "First European Symposium on Enzymes and Grain Processing." Angelino, S.A.G.F., van Hamer, R.J., Hartingsveldt, W., Heidekamp, F., van der Lugt, J.P., eds., pp.188-194. TNO Nutrition and Food Research Institute, 1997. ISBN 90-75202-04-0. Christensen, A.M. et al., "Maize polyubiquitin genes: Structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation." Plant Mol. Biol. 18:675-689, 1992. Colbert, T. et al., "High-throughput screening for induced point mutations." Plant Physiol. 126:480-484, 2001. Cornish-Bowden, A., "Nomenclature for incompletely specified bases in nucleic acid sequences: Recommendations 1984." Nucleic Acids Res. 13:3021-3030, 1985. Croft, K.P.C. et al., "Volatile products of the lipoxygenase pathway evolved from Phaseolus vulgaris (L.) leaves inoculated with Pseudomonas syringae pv phaseolicola." Plant Physiol. 101:13-24, 1993. Davies, C.S. and Nielsen, N.C., "Genetic analysis of null-allele for lipoxygenase-2 in soybean." Crop Sci. 26:460-463, 1986. Dougherty, W.G. and Parks, Т.О., "Transgenes and gene suppression: Telling us something new?" Curr. Opin. Cell Biol. 7:399-405, 1995. Drost, B.W. et al., "Role of individual compounds in beer staling." Tech. Q. MBAA 11:127-134, 1974. Drost, B.W. et al., "Flavor stability." J. Am. Soc. Brew. Chem. 48:124-131, 1990. Ellis, R.P. et al., "Barley domestication - Hordeum spontaneum, a source of new genes for crop improvement." Scottish Crop Research Institute, Annual Report 1998/99:97-100,1999. Also available at http://www.scri. sari.ac.uk/SCRI/upload/annualreportdocuments/ 99lndiv/14Barley.pdf. European Brewery Convention, "Analytica - EBC." ISBN 3-418-00759-7 (1998). Feussner, I. and Wasternack, C., "The lipoxygenase pathway." Annu. Rev. Plant Physiol. Plant Mol. Biol. 53:275-297, 2002. Forster, С. et al., "Molecular analysis of a null mutant for pea (Pisum sativum L.) seed lipoxygenase-2. " Plant Mol. Biol. 39:1209-1220, 1999. Gardner, H.W. and Grove, M.J., "Method to produce 9(S)-hydroperoxides of linoleic and linolenic acids by maize lipoxygenase." Lipids 36:529-533, 2001. Giaever, G. et al., "Functional profiling of the Saccharomyces cerevisiae genome." Nature 418:387-391, 2002. Goenczy, P. et al., "Functional genomic analysis of cell division in C. elegans using RNAi of genes on chromosome III." Nature 408:331-336, 2000. Graef, G.L. et al. "Fatty acid development in a soybean mutant with high stearic acid." J. Am. Oil Chem. Soc. 62:773-775, 1985. Griffiths, A. et al., "Fruit-specific lipoxygenase suppression in antisense-transgenic tomatoes." Postharvest Biol. Technol. 17:163-173, 1999. Groenqvist, A. et al., "Carbonyl compounds during beer production in beer." Proceedings of the 24th EBC Congress, Oslo, pp. 421-428, 1993. Grosch, W. and Schwartz, J.M., "Linoleic and linolenic acid as precursors of the cucumber flavor." Lipids 6:351-352, 1971. Ha, T.J. et al., "Lipoxygenase inhibitory activity of octyl gallate." J. Agric. Food Chem. 52:3177-3181, 2004. Hamberg, M., "Trihydroxyoctadecenoic acids in beer: Qualitative and quantitative analysis." J. Agric. Food Chem. 39:1568-1572, 1991. Hannon, G.J., "RNA interference." Nature 418:244-251, 2002. Hildebrand, D.F. and Hymowitz, Т., "Inheritance of lipoxygenase-1 activity in soybean seeds." Crop Sci. 22:851-853, 1982. Holtman, W.L. et al., "Differential expression of lipoxygenase isoenzymes in embryos of germinating barley. " Plant Physiol. 111:569-576, 1996. Husson, F. and Belin, J.M., "Purification of hydroperoxide lyase from green bell pepper (Capsicum annuum L.) fruits for the generation of С6-aldehydes in vitro." J. Agric. Food Chem. 50:1991-1995, 2002. Institute of Brewing, "Institute of Brewing. Methods of analysis." ISBN 0-900489-10-3 (1997). IUPAC-IUB Joint Commission on Biochemical Nomenclature, "Nomenclature andsymbolism for amino acids and peptides. Recommendations 1983." Biochem. J.219:345-373, 1984. Isshiki, M. et al., "Nonsense-mediated decay of mutant waxy mRNA in rice." Plant Physiol. 125:1388-1395, 2001. Jalloul, A. et al., "Lipid peroxoidation in cotton: Xanthomonas interactions and the role of lipoxygenases during the hypersensitive reaction." Plant J. 32:1-12, 2002. Jamieson, A.M. and Van Gheluwe, J.E.A., "Identification of a compound responsible for cardboard flavor in beer." Proc. Am. Soc. Brew. Chem. 29:192-197, 1970. Jende-Strid, В., "Gene-enzyme relations in the pathway of flavonoid biosynthesis in barley." Theor. Appl. Genet. 81:668-674, 1991. Jende-Strid, В., "Genetic control offlavonoid biosynthesis in barley." Hereditas 119:187-204, 1993. Jensen, L.G. et al., "Transgenic barley expressing a protein-engineered, thermostable (1,3-1,4)-|3-glucanase during germination. Proc. Natl. Acad. Sci. USA 93, 3487-3491, 1996. Kamath, R.S. et al., "Effectiveness of specific RNA-mediated interference through ingested double-stranded RNA in Caenorhabdtis elegans." Genome Biol. 2: Research0002.1-0002.10, 2000. Kamath, R.S. et al., "Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. "Nature 421:231-237, 2003. Kitamura et al., "Genetic analysis of a null-allele for lipoxygenase-3 in soybean seeds." Crop Sci. 23:924-927, 1983. Kleinhofs, A. et al., "Induction and selection of specific gene mutations in Hordeum and Pisutn." Mut. Res. 51:29-35, 1978. Kolomiets, M.V. et al., "Lipoxygenase is involved in the control of potato tuber development." Plant Cell 13:613-626, 2001. Kuroda et al., "Characterization of factors involved in the production of 2(E)-nonenal during mashing." Biosci. Biotechnol. Biochem. 67:691-697, 2003. Kusaba, M. et al., "Low glutelin contentl: A dominant mutation that suppresses the Glutelin multigene family via RNA silencing in rice." Plant Cell 15:1455-1467, 2003. Laemmli, U.K., "Cleavage of structural proteins during the assembly of the head of bacteriophage T4." Nature 227:680-685, 1970. Leon, J. et al., "Lipoxygenase H1 gene silencing reveals a specifc role in supplying fatty acid hydroperoxides for aliphatic aldehyde production." J. Biol. Chem. 277:416-423, 2002. Lermusieau, G. et al., "Nonoxidative mechanism for development of frans-2-nonenal in beer." J. Am. Soc. Brew. Chem. 57(1 ):29-33, 1999. Liegeois, C. et al., "Release of deuterated (E)-2-nonenal during beer aging from labeled precursors synthesized before boiling." J. Agric. Food Chem. 50:7634-7638, 2002. Maquat, L.E. and Carmichael, G.G., "Quality control of mRNAfunction." Cell 104:173-176, 2001. Matsui, K. et al., "Effects of overexpression of fatty acid 9-hydroperoxide lyase in tomatoes (Lycopersicon esculentum Mill.)-" J- Agric. Food Chem. 49:5418-5424, 2001. May, C. et al., "The N-terminal ( β-barrel structure of lipid body lipoxygenase mediates its binding to liposomes and lipid bodies." Eur. J. Biochem. 267:1100-1109, 2000. McElroy, D. and Jacobsen, J., "What's brewing in barley biotechnology?" Bio/Technology 13:245-249, 1995. Meilgaard, M.C., "Flavor chemistry of beer: Part II: Flavor and threshold of 239 aroma volatiles." Tech. Q. MBAA 12:151-167, 1975. Melan, M.A. et al., "An Arabidopsis lipoxygenase gene can be induced by pathogens, abscisic acid, and methyl jasraonate. Plant Physiol. 101:441-450, 1993. Mendell, J.T. and Dietz, H.C., "When the message goes awry: Disease-producing mutations that influence mRNA content and performance." Cell 107:411-414, 2002. Narziss, L, "Centenary Review: Technological factors of flavour stability." J. Inst. Brew. 92:346-353, 1986. Nevo, E., "Resources for Breeding of Wild Barley." In: "Barley: Genetics, Biochemistry, Molecular Biology and Biotechnology." Shewry, P.R., ed., pp. 3-18. CAB. International. ISBN 0-85198-725-7 (1992). Noel, S. and Collin, S., "Trans-2-nonenal degradation products during mashing." Eur. Brew. Conv. Proc. Congr. 25th, Brussels: 483-490, 1995. Noordermeer, M.A. et al., "Fatty acid hydroperoxide lyase: A plant cytochrome P450 enzyme involved in wound healing and pest resistance." ChemBioChem 2:494-504, 2001. Noordermeer, M.A. et al., "Development of a biocatalytic process for the production of C6-aldehydes from vegetable oils by soybean lipoxygenase and recombinant hydroperoxide lyase." J. Agric. Food Chem. 50:4270-4274, 2002. Norden, A.J. et al., "Variability in oil quality among peanut genotypes in the Florida breeding program." Peanut Sci. 14:7-11, 1987. Nyborg, M. et al., "Investigations of the protective mechanism of sulfite against beer staling and formation of adducts with trans-2-nonenal." J. Am. Soc. Brew. Chem. 57:24-28, 1999. Ohtsu, K. et al., "Flavor stability of packaged beer in relation to the oxidation of wort." Brew. Dig. 61(6):18-23, 1986. Olsen, O. et al., "Preferential generation of A-T-G-C transitions in the barley Ant18 gene." Proc. Natl. Acad. Sci. USA 90:8043-8047, 1993. Osorio, J. et al., "Mutant sunflower with high concentration of saturated fatty acids in the oil." Crop Sci. 35:739-742, 1995. Parinov, S. and Sundaresan, V., "Functional genomics in Arabidopsis: Large-scale insertional mutagenesis complements the genome sequencing project." Curr. Opin. Biotechnol. 11:157-161, 2000. Phillips, D.R. and Galliard, Т., "Flavour biogenesis, partial purification and properties of a fatty acid hydroperoxide cleaving enzyme from fruits of cucumber." Phytochemistry 17:355-358, 1978. Ramezanzadeh, F.M. et al., "Prevention of oxidative rancidity in rice bran during storage." J. Agric. Food Chem. 47:2997-3000, 1999. Ranee, I. et al., "The incompatible interaction between Phytophthora parasitica var. nicotianae race 0 and tobacco is suppressed in transgeniv plants expressing antisense lipoxygenase sequences." Proc. Natl. Acad. Sci. USA 95:6554-6559, 1998. Rasmussen, S.K. and Hatzak, F., "Identification of two low-phytate barley (Hordeum vulgare L) grain mutants by TLC and genetic analysis." Hereditas 129:107-112, 1998. Rogers, K.R. et al., "Lipid peroxidation is a consequence of elicitor activity." Plant Physiol. 86:547-553, 1988. Rouster, J. et al., "Identification of a methyl jasmonate-responsive region in the promoter of a lipoxygenase 1 gene expressed in barley grain." Plant J. 11:513-523, 1997. Royo, J. et al., "Antisense-mediated depletion of a potato lipoxygenase reduces wound induction of proteinase inhibitors and increases weight gain of insect pests." Proc. Natl. Acad. Sci. USA 96:1146-1151, 1999. Rusterucci, C. et al., "Involvement of lipoxygenase-dependent production of fatty acid hydroperoxides in the development of the hypersensitive cell death induced by cryptogein on tobacco leaves." J. Biol. Chem. 274:36446-36455, 1999. Sambrook, J. et al., "Molecular Cloning: A Laboratory Manual, 2nd Ed.", Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. ISBN 0-87969-309-6. Sambrook, J. and Russell, D.W., "Molecular Cloning. A Laboratory Manual, 3rd Ed.", Cold Spring Harbor Laboratory-Press, Cold Spring Harbor, New York, 2001. ISBN 0-87969-577-3. Saravitz, D.M. and Siedow, J.N., "The differential expression of wound-indueible lipoxygenase genes in soybean leaves." Plant Physiol. 110:287-299, 1996. Schmitt, N.F. and Van Mechelen, J.R., "Expression of lipoxygenase isoenzymes in developing barley grains." Plant Sci. 128:141-150, 1997. Soldatov, K.I., "Chemical mutagenesis in sunflower breeding." In: "Proceedings of the Vllth International Sunflower Conference, 27 June-3 July, 1976, Krasnodar, USSR, Vol. 1", pp 352-357. International Sunflower Association, Toowoomba, Australia, 1976. Srivastava, S. et al., "Structural and kinetic determinants of aldehyde reduction by aldose reductase." Biochemistry 38:42-54, 1999. Start, W.G. et al., "Two soybean seed lipoxygenase nulls accumulate reduced levels of lipoxygenase transcripts." Plant Mol. Biol. 7:11-23, 1986. Tatulian, S.A. et al., "Uncovering a calcium-regulated membrane-binding mechanism for soybean lipoxygenase-1." Biochemistry 37:15481-15490, 1998. Tijet, N. et al., "Biogenesis of volatile aldehydes from fatty acid hydroperoxides: Molecular cloning of a hydroperoxide lyase (CYP74C) with specificity for both the 9- and 13-hydroperoxides of linoleic and linolenic acids." Arch. Biochem. Biophys. 386:281-289, 2001. Tingay, S. et al., "Agrobacterium tumefaciens-mediated barley transformation." Plant J. 11:1369-1376, 1997. Towbin, H. et al., "Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications." Proc. Natl. Acad. Sci. USA 76:4350-4354, 1979. Turner, J.G. et al., "The jasmonate signal pathway." Plant Cell 14:5153-3164, 2002. Vancanneyt, G. et al., "Hydroperoxide lyase depletion in transgenic potato plants leads to an increase in aphid performance." Proc. Natl. Acad. Sci. USA 98:8139-8144, 2001. van Mechelen, J.R. et al., "Primary structure of a lipoxygenase from barley grain as deduced from its cDNA sequence." Bioche. Biophys. Acta 1254:221-225, 1995. van Mechelen, J.R. et al., "Molecular characterization of two lipoxygenases from barley." Plant Mol. Biol. 39:1283-1298, 1999. von Bothmer, R. et al., "Diversity in barley (Hordeum vulgare)" In: "Diversity in Barley (Hordeum vulgare)" von Bothmer, van Hintum, Knupffer, H., Sato, K., eds., pp. 129-136. ISBN 0-444-50587-7 (2003). Also available at http://www.genres.de/IGRREIHE/IGRREIHE/DDD/22-16.pdf. von Wettstein, D. et al., "Biochemical mutant in barley renders chemical stabilization of beer superfluous." Carlsberg Res. Commun. 42:341-351, 1977. von Wettstein, D. et al., "Proanthocyanidin-free barley for brewing: Progress in breeding for high yield and research tool in polyphenol chemistry." Tech. Q. MBAA 22:41-52, 1985. Wan, Y. and Lemaux, P.G., "Generation of large numbers of independently transformed fertile barley plants." Plant Physiol. 104:37-48, 1994. Wang, J. et al., "Alternatively spliced TCR mRNA induced by disruption of reading frame." Science 297:108-110, 2002. Wang, M.-B. et al., "Agrobacterium tumefaciens-mediated transformation of an elite Australian barley cultivar with virus resistance and reporter genes." Aust. J. Plant Physiol. 28:149-156, 2001. Wang, W.H. et al., "Molecular basis of a null mutation in soybean lipoxygenase 2: Substitution of glutamine for an iron-ligand histidine." Proc. Natl. Acad. Sci. USA 91:5828-5832, 1994. Wang, W.-H. et al., "Two single-base substitutions involved in altering in a paried-box of AAATAC in the promoter region of soybean lipoxygenase L-3 gene impair the promoter function in tobacco cells." Plant Sci. 109:67-73, 1995. Weber, H. et al., "Divinyl ether fatty acid synthesis in late blight-diseased potato leaves." Plant Cell 11:485-493, 1999. Wesley, S.V. et al., "Construct design for efficient, effective and high-throughput gene silencing in plants." Plant J. 27:581-590, 2001. White, J. et al., "A cassette containing the bar gene of Streptomyces hygroscopicus: A selectable marker for plant transformation. Nucleic Acids Res. 18:1062, 1990. Zhang, Y. et al., "Expression of antisense SnRKl protein kinase sequence causes abnormal pollen development and male sterility in transgenic barley." Plant J. 28:431-441, 2001. |