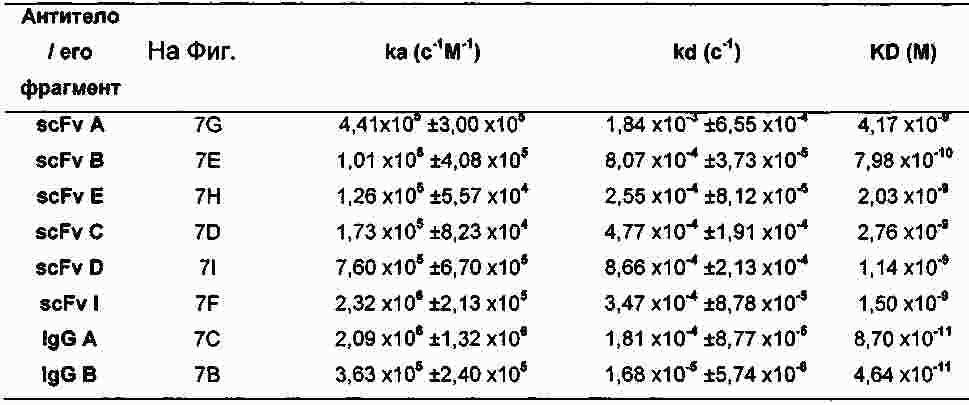
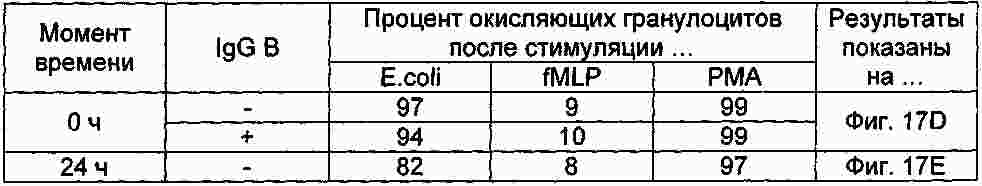
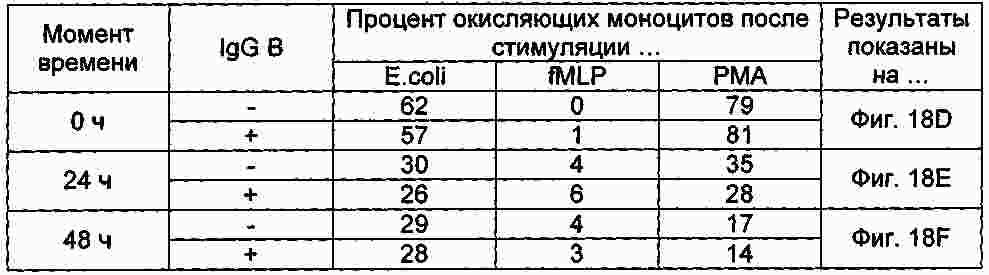
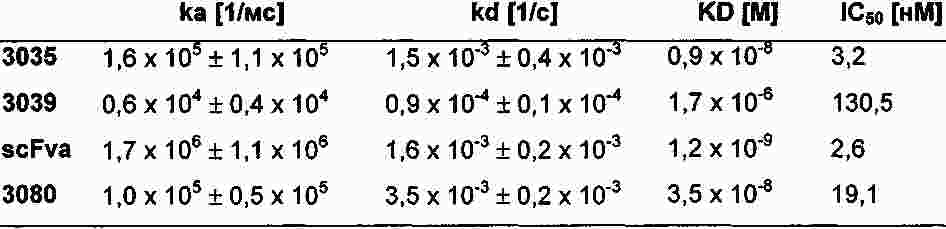|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000013162b*\id |
|
Термины запроса в документе Реферат Настоящее изобретение относится к человеческому моноклональному антителу или его фрагменту, которые специфически связываются с GM-CSF приматов и нейтрализуют его. Формула [0001] Человеческое моноклональное антитело или его фрагмент, которые специфически связываются с GM-CSF (гранулоцитарно-макрофагальный колониестимулирующий фактор) приматов и нейтрализуют его. [0002] Человеческое моноклональное антитело или его фрагмент по п.1, где указанный примат представляет собой человека или примата, не являющегося человеком. [0003] Человеческое моноклональное антитело или его фрагмент по п.2, где указанный примат, не являющийся человеком, представляет собой макака-крабоеда, макака-резуса или гиббона. [0004] Человеческое моноклональное антитело по любому из пп.1-3, где указанное антитело представляет собой IgG. [0005] Человеческое моноклональное антитело по п.4, где указанный IgG представляет собой IgG1 или IgG4. [0006] Фрагмент человеческого моноклонального антитела по любому из пп.1-3, где указанный фрагмент представляет собой scFv, однодоменное антитело, Fv, VHH антитело, диатело, тандемное диатело, Fab, Fab' или F(ab)2. [0007] Человеческое моноклональное антитело или его фрагмент по любому из пп.1-6, где указанное человеческое моноклональное антитело или его фрагмент специфически связываются с эпитопом, предпочтительно с прерывистым эпитопом, GM-CSF примата, содержащим аминокислоты 23-27 (RRLLN) и/или аминокислоты 65-77 (GLR/QGSLTKLKGPL). [0008] Человеческое моноклональное антитело или его фрагмент по п.7, где указанный прерывистый эпитоп дополнительно содержит аминокислоты 28-31 (LSRD). [0009] Человеческое моноклональное антитело или его фрагмент по п.7 или 8, где указанный прерывистый эпитоп дополнительно содержит аминокислоты 32-33 (ТА) и/или аминокислоты 21-22 (ЕА). [0010] Человеческое моноклональное антитело или его фрагмент по любому из пп.1-9, где указанное человеческое моноклональное антитело или его фрагмент содержат в вариабельной области его тяжелой цепи область CDR3, содержащую аминокислотную последовательность, выбранную из группы, состоящей из последовательностей, определенных в любой из SEQ ID NO:1-13 или 56. [0011] Человеческое моноклональное антитело или его фрагмент по п.10, где любая из указанных последовательностей CDR3 вариабельной области тяжелой цепи существует в вариабельной области тяжелой цепи вместе с последовательностью CDR1 вариабельной области тяжелой цепи, определенной в SEQ ID NO:14, и с последовательностью CDR2 вариабельной области тяжелой цепи, определенной в SEQ ID NO: 15. [0012] Человеческое моноклональное антитело или его фрагмент по п.10 или 11, где указанное человеческое моноклональное антитело или его фрагмент содержат в вариабельной области его легкой цепи область CDR1, содержащую аминокислотную последовательность, определенную в SEQ ID NO:16, область CDR2, содержащую аминокислотную последовательность, определенную в SEQ ID NO: 17, и область CDR3, содержащую аминокислотную последовательность, определенную в SEQ ID NO: 18. [0013] Человеческое моноклональное антитело или его фрагмент по п.10 или 11, где указанное человеческое моноклональное антитело или его фрагмент дополнительно содержат в вариабельной области его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO:19, 54 или 55. [0014] Человеческое моноклональное антитело или его фрагмент по любому из пп.10-13, где указанное человеческое моноклональное антитело или его фрагмент содержат в вариабельной области его тяжелой цепи аминокислотную последовательность, как представлено в любой из SEQ ID NO:20-33, 52 или 53. [0015] Человеческое моноклональное антитело или его фрагмент по любому из пп.1-14, содержащее в вариабельной области его легкой цепи область CDR1, содержащую аминокислотную последовательность, как представлено в SEQ ID NO:16, область CDR2, имеющую аминокислотную последовательность, как представлено в SEQ ID NO: 17, и область CDR3, имеющую аминокислотную последовательность, как представлено в SEQ ID NO: 18; и содержащее в вариабельной области его тяжелой цепи область CDR1, содержащую аминокислотную последовательность, как представлено в SEQ ID NO: 14, область CDR2, имеющую аминокислотную последовательность, как представлено в SEQ ID NO: 15, и область CDR3, имеющую аминокислотную последовательность, как представлено в любой из SEQ ID NO: 1-13 или 56. [0016] Человеческое моноклональное антитело по любому из пп.10-15, содержащее аминокислотную последовательность легкой цепи, как представлено в SEQ ID NO:34, и аминокислотную последовательность тяжелой цепи, как представлено в любой из SEQ ID NO: 35-48. [0017] Человеческое моноклональное антитело или его фрагмент по любому из пп.10-16, где указанное человеческое моноклональное антитело или его фрагмент содержат аминокислотную последовательность, имеющую по меньшей мере 70%-ную гомологию с соответствующей аминокислотной последовательностью, как представлено в любой из SEQ ID NO:1-48 и/или 52-56. [0018] Полинуклеотидная молекула, имеющая нуклеотидную последовательность, кодирующую аминокислотную последовательность, как представлено в любой из SEQ ID NO:1-48 и/или 52-56, или нуклеотидную последовательность, демонстрирующую по меньшей мере 70%-ную гомологию с ней, [0019] Фармацевтическая композиция, содержащая человеческое моноклональное антитело или его фрагмент по любому из пп.1-17. [0020] Фармацевтическая композиция, содержащая полинуклеотидную молекулу по п.18. [0021] Применение человеческого моноклонального антитела или его фрагмента по любому из пп.1-17 или полинуклеотидной молекулы по п.18 в изготовлении лекарственного средства, возможно содержащего один или более чем один дополнительный противовоспалительный агент, для лечения воспалительных заболеваний. [0022] Применение по п.21, где указанные воспалительные заболевания выбраны из группы, состоящей из ревматоидного артрита (RA) (включая RA, который является устойчивым к лечению нейтрализаторами TNF (фактор некроза опухолей)-альфа), астмы, рассеянного склероза (MS), хронического обструктивного заболевания легких (COPD), острого респираторного дистресс-синдрома (ARDS), идиопатического легочного фиброза (IPF), воспалительного заболевания кишечника (IBD), увеита, дегенерации желтого пятна, колита, псориаза, Уоллеровской дегенерации, антифосфолипидного синдрома (APS), острого коронарного синдрома, рестеноза, атеросклероза, рецидивирующего полихондрита (RP), острого или хронического гепатита, неудачного приживления ортопедических имплантатов, гломерулонефрита, волчанки или аутоиммунных расстройств. [0023] Применение человеческого моноклонального антитела или его фрагмента по любому из пп.1-17 или полинуклеотидной молекулы по п.18 в изготовлении лекарственного средства, возможно содержащего один или более чем один дополнительный противораковый агент, для лечения опухолевого заболевания или другого состояния с замедленным апоптозом клеток, повышенным выживанием клеток или пролиферацией. [0024] Применение по п.23, где указанное опухолевое заболевание представляет собой рак. [0025] Применение по п.24, где указанное раковое заболевание представляет собой лейкоз, множественную миелому, карциному желудка или карциному кожи. Полный текст патента Настоящее изобретение относится к антителам и их фрагментам, которые нейтрализуют активность человеческого гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF). Данное изобретение дополнительно относится к фармацевтическим композициям, содержащим такие антитела и их фрагменты, а также к применениям таких антител и их фрагментов для получения лекарственных средств для лечения различных состояний. Гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), первоначально описанный как сильнодействующий стимул роста и дифференцировки клеток-предшественников гранулоцитов и макрофагов in vitro, представляет собой гликопротеин с молекулярной массой приблизительно 23 кДа со структурой пучка из четырех альфа-спиралей, который связывается с гетеродимерным рецептором, состоящим из субъединиц, принадлежащих к семейству цитокиновых рецепторов типа 1. Среди прочего он стимулирует созревание макрофагов, нейтрофилов, гранулоцитов, эозинофилов и антигенпредставляющих дендритных клеток для увеличения их функциональной способности в борьбе с инфекциями. Эксперименты по генетической абляции, т.е. эксперименты с сайленсингом или нокаутом интересующего гена, здесь GM-CSF, у мышей показали, что GM-CSF необходим для поддержания функциональной активности некоторых популяций макрофагов, таких как популяции, которые участвуют в очистке поверхностно-активного соединения в легких и в реакции на определенные типы инфекции или в иммунных ответах. В то время как GM-CSF имеет мощные стимулирующие активности in vitro на клетки-предшественники нейтрофилов, эозинофилов, макрофагов и, в меньшей степени, на эритроидные клетки и мегакариоциты, результаты, полученные in vivo на мышах с нокаутом генов, говорят о том, что основная физиологическая роль GM-CSF заключается в поддержании или стимуляции функциональной активности зрелых макрофагов и гранулоцитов и в стимуляции презентации антигена иммунной системе. Он осуществляет последнее благодаря его прямым эффектам на продукцию дендритных клеток и макрофагов, а также посредством повышения экспрессии главного комплекса гистосовместимости класса II и Fc-рецепторов на макрофагах и дендритных клетках. GM-CSF стимулирует функциональные активности нейтрофилов, эозинофилов и моноцитов-макрофагов. Они включают повышение активности хемотаксиса, увеличенную экспрессию клеточных адгезивных молекул, повышенную адгезию к поверхностям и повышенную фагоцитарную активность, а также ингибирование и задержку апоптоза этих клеток. Нейтрофилы представляют собой первую линию защиты против вмешательств. Программируемая смерть нейтрофилов задерживается провоспалительными стимулами, включающими GM-CSF, для обеспечения правильного рассасывания воспаления во времени и в пространстве. GM-CSF также стимулирует способность этих клеток опосредовать антитело-зависимую клеточную цитотоксичность и внутриклеточно убивать микроорганизмы и оказывает "примирующий" эффект на эти клетки для усиления их реакции на последующие стимулы в отношении окислительного взрыва (продукция супероксид-аниона), дегрануляции и высвобождения противомикробных агентов и хемотаксиса. Кроме того, GM-CSF стимулирует высвобождение из этих клеток вторичных цитокинов и медиаторов, включающих IL-1, G-CSF, М-CSF и лейкотриены из нейтрофилов, а также IL-1, TNF, IL-6, G-CSF, M-CSF и простагландины из макрофагов. Из вышеуказанного очевидно, что GM-CSF играет ключевую роль в активации и поддержании популяций клеток, необходимых для предупреждения инфекции. Однако в некоторых случаях активация этих популяций клеток может быть нежелательной. Например, активация вышеупомянутых клеточных линий, когда патоген отсутствует, во многих случаях приводит к острым и/или хроническим воспалительным состояниям, которые в крайних случаях могут угрожать жизни. Аналогично, сверхэкспрессия GM-CSF может приводить к избытку иммунной активации, приводящему к воспалению. В таких случаях может быть желательной нейтрализация активности GM-CSF с тем, чтобы симптомы этих воспалительных состояний устранялись или, по меньшей мере, ослаблялись. Примеры такой нейтрализующей активности существуют в предшествующем уровне техники. Например, обнаружили, что нейтрализующее антитело против GM-CSF способствовало увеличению скорости апоптоза эозинофилов в образцах периферической крови (Kankaanranta et al. (2000) Journal of Allergy and Clinical Immunology 106, 77-83). Так как повышенная выживаемость эозинофилов коррелирует с астмой, то ожидают, что усиление апоптоза эозинофилов будет ослаблять астматические симптомы. При хронических воспалительных заболеваниях, таких как астма, ревматоидный артрит и рассеянный склероз, уровни GM-CSF возрастают локально и в некоторых случаях системно, и они коррелировали с воспалительным процессом при этих заболеваниях. Целью данного изобретения, следовательно, является улучшение способов нейтрализации повышенной и/или нежелательной активности GM-CSF, известной ранее в предшествующем уровне техники. Соответственно, один аспект данного изобретения относится к человеческому моноклональному антителу или его фрагменту, которые специфически связываются с GM-CSF приматов и нейтрализуют его. Термин "специфически связывает" или родственные выражения, такие как "специфическое связывание", "связывающий специфически", "специфический связыватель" и т.д. в том виде, как они здесь используются, относятся к способности человеческого моноклонального антитела или его фрагмента различать GM-CSF приматов и любое число других потенциальных антигенов, отличных от GM-CSF приматов, в такой степени, что из пула множества различных антигенов, служащих в качестве потенциальных партнеров связывания, связывается только GM-CSF приматов, или связывается в значительной степени. В контексте данного изобретения GM-CSF приматов "значительно" связывается, когда из пула множества, в равной степени доступных различных антигенов, служащих в качестве потенциальных партнеров связывания, GM-CSF приматов связывается по меньшей мере в 10 раз, предпочтительно в 50 раз, наиболее предпочтительно в 100 или более раз чаще (в кинетическом смысле), чем любой другой антиген, отличный от GM-CSF приматов. Такие кинетические измерения можно провести на установке BIAcore. Термины "нейтрализация", "нейтрализатор", "нейтрализующий" и их грамматически родственные варианты в том виде, как они здесь используются, относятся к частичному или полному ослаблению биологического(их) эффекта(ов) GM-CSF. Такое частичное или полное ослабление биологического(их) эффекта(ов) GM-CSF происходит из-за модификации, прерывания и/или отмены GM-CSF-опосредованной трансдукции сигнала, что проявляется, например, во внутриклеточной передаче сигнала, пролиферации клеток или в высвобождении растворимых веществ, в повышающей или понижающей регуляции внутриклеточной активации генов, например, той, которая приводит к экспрессии поверхностных рецепторов для лигандов, отличных от GM-CSF. Как понятно специалисту в данной области, существует множество способов определения того, подлежит ли агент, например рассматриваемое антитело или его фрагмент, классификации в качестве нейтрализатора. В качестве примера это может быть осуществлено с помощью стандартного анализа in vitro, который обычно проводят следующим образом. В первом эксперименте по пролиферации клеточную линию, для которой известно, что степень ее пролиферации зависит от активности GM-CSF, инкубируют в серии образцов с варьирующими концентрациями GM-CSF, после чего измеряют степень пролиферации данной клеточной линии. Из этого измерения определяют концентрацию GM-CSF, обеспечивающую полумаксимальную пролиферацию данных клеток. Затем проводят второй эксперимент по пролиферации с использованием в каждой из серий образцов такого же количества клеток, которое использовалось в первом эксперименте по пролиферации, определенной выше концентрации GM-CSF и, на этот раз, варьирующих концентраций антитела или его фрагмента, которое, как предполагают, является нейтрализатором GM-CSF. Вновь измеряют пролиферацию клеток для определения концентрации антитела или его фрагмента, достаточной для осуществления полумаксимального ингибирования роста. Если полученный график ингибирования роста относительно концентрации антитела (или его фрагмента) имеет сигмоидальную форму, приводящую к уменьшению клеточной пролиферации при увеличении концентрации антитела (или его фрагмента), тогда обеспечивается некоторая степень антителозависимого ингибирования роста, т.е. активность GM-CSF была до некоторой степени нейтрализована. В таком случае антитело или его фрагмент можно рассматривать как "нейтрализатор" в контексте настоящего изобретения. Одним примером клеточной линии, для которой известно, что уровень ее пролиферации зависит от активности GM-CSF, является клеточная линия TF-1, как описано в Kitamura, Т. et al. (1989). J. Cell. Physiol. 140, 323-34. Как понятно среднему специалисту в данной области, степень клеточной пролиферации является не единственным параметром, с помощью которого можно установить нейтрализующую способность. Например, для идентификации предполагаемого нейтрализатора GM-CSF можно использовать измерение уровня сигнальных молекул (например, цитокинов), уровень секреции которых зависит от GM-CSF. Другие примеры клеточных линий, которые могут быть использованы для определения того, является ли рассматриваемое антитело или его фрагмент нейтрализатором активности GM-CSF приматов, включают AML-193 (Lange, В. et al. (1987). Blood 70, 192-9); GF-D8 (Rambaldi, A. et al. (1993). Blood 81, 1376-83); GM/SO (Oez, S. et al. (1990). Experimental Hematology 18, 1108-11); MO7E (Avanzi, G.C. et al. (1990). Journal of Cellular Physiology 145, 458-64); TALL-103 (Valtieri, M. et al. (1987). Journal of Immunology 138, 4042-50); UT-7 (Komatsu, N. et al. (1991). Cancer Research 51, 341-8). Человеческое антитело или его фрагмент по данному изобретению являются моноклональными. Следует понимать, что термин "моноклональное" в том виде, как он здесь используется, имеет значение, обычно приписываемое ему в данной области, а именно антитело (или его соответствующий фрагмент), происходящее из одного клона антитело-продуцирующей клетки, такой как В-клетка, и распознающее один эпитоп на связываемом антигене. Особенно сложно получить человеческие антитела, которые являются моноклональными. В отличие от слияний мышиных В-клеток с бессмертными клеточными линиями, слияния В-клеток человека с бессмертными клеточными линиями являются нежизнеспособными. Таким образом, человеческое моноклональное антитело по данному изобретению является результатом преодоления значительных технических трудностей, которые, как общеизвестно, существуют в области технологии антител. Моноклональная природа антитела делает его особенно хорошо приспособленным для применения в качестве терапевтического агента, поскольку такое антитело будет существовать в виде единственной гомогенной молекулярной частицы, которая может быть хорошо охарактеризована и воспроизводимо получена и очищена. Эти факторы приводят к получению продукта, биологическая активность которого может быть предсказана с высоким уровнем точности, что очень важно, если собираются получить одобрение регулирующего органа для терапевтического введения такой молекулы человеку. Особенно важным является то, чтобы моноклональное антитело (или соответствующий фрагмент) по изобретению представляли собой человеческое антитело (или соответствующий фрагмент). При рассмотрении агента, представляющего собой антитело, предназначенного для терапевтического введения человеку, весьма полезно, чтобы это антитело имело человеческое происхождение. После введения пациенту, являющемуся человеком, человеческое антитело или его фрагмент скорее всего не будет вызывать сильной иммуногенной реакции иммунной системы пациента, т.е. не будет распознаваться как "чужеродный", т.е. нечеловеческий белок. Это означает, что антитела хозяина, т.е. пациента, не будут генерироваться против терапевтического антитела, что, в противном случае, блокировало бы активность терапевтического антитела и/или ускоряло бы элиминацию терапевтического антитела из организма пациента, предотвращая, таким образом, оказание им желательного терапевтического эффекта. Следует понимать, что термин "человеческое антитело" в том виде, как он здесь используется, означает, что антитело по данному изобретению или его фрагмент включает аминокислотную(ые) последовательность(и), которые содержатся в спектре человеческих антител зародышевой линии. Поэтому с целью определения здесь антитело или его фрагмент можно считать человеческим, если оно состоит из такой(их) (а) аминокислотной(ых) последовательности(ей) зародышевой линии человека, т.е. если аминокислотная(ые) последовательность(и) рассматриваемого антитела или его фрагмента идентична(ы) экспрессируемой(ым) аминокислотной(ым) последовательности(ям) человеческой зародышевой линии. Антитело или его фрагмент также можно считать человеческими, если они состоят из (а) последовательности(ей), которая(ые) отклоняется(ются) от ближайшей к ней(им) последовательности(ей) человеческой зародышевой линии не более чем ожидалось бы из-за импринтинга соматической гипермутации. Кроме того, антитела многих млекопитающих, не являющихся человеком, например, грызунов, таких как мыши и крысы, содержат аминокислотные последовательности VH CDR3, которые, как можно ожидать, также существуют в спектре экспрессируемых человеческих антител. Любая такая(ие) последовательность(и) человеческого или нечеловеческого происхождения, которая(ые), как можно ожидать, существует(ют) в экспрессируемом человеческом спектре, также считалась(ись) бы "человеческой(ими)" для целей настоящего изобретения. Согласно одному воплощению изобретения GM-CSF приматов представляет собой GM-CSF человека (Homo sapiens) или GM-CSF приматов, не являющихся человеком. Особенно предпочтительные варианты GM-CSF приматов, не являющихся человеком, включают в себя GM-CSF обезьяны гиббона (Nomascus concolor, также известный как западный черный гиббон) и GM-CSF обезьян семейства макак, например GM-CSF макака-резуса (Масаса mulatta) и GM-CSF макака-крабоеда (Масаса fascicularis). Согласно этому воплощению изобретения человеческое моноклональное антитело или его фрагмент демонстрируют перекрестную реактивность как между человеком, так и по меньшей мере одним из видов обезьян, упомянутых выше. Это является особенно полезным для молекулы антитела, которая предназначена для терапевтического введения человеческим субъектам, так как такое антитело в норме должно будет пройти через множество тестов перед тем, как будет одобрено регулирующими органами, из которых некоторые ранние тесты включают вид животных, не являющихся человеком. При проведении таких тестов обычно желательно использовать в качестве видов, не являющихся человеком, виды, имеющие высокую степень генетического сходства с человеком, так как результаты, полученные таким образом, обычно будут в значительной степени предсказывать соответствующие результаты, которые можно ожидать при введении той же молекулы человеку. Однако такая прогнозирующая способность, основанная на тестах с животными, по меньшей мере, частично зависит от сравнимости молекулы и является очень высокой, когда из-за перекрестной реактивности между видами одна и та же терапевтическая молекула может быть введена человеку и животным моделям. Как и в этом воплощении изобретения, когда молекула антитела перекрестно реагирует с одним и тем же антигеном у человека и у другого близкородственного вида, тесты с использованием одной и той же молекулы антитела можно проводить и у человека, и у этого близкородственного вида, например у одного из видов обезьян, упомянутых выше. Это увеличивает как эффективность самих тестов, так и прогнозирующую способность, обеспечиваемую такими тестами, относительно поведения таких антител у человека, основного вида, представляющего интерес с терапевтической точки зрения. Согласно другому воплощению изобретения человеческое моноклональное антитело может представлять собой IgG антитело. Как хорошо известно в данной области, IgG содержит не только вариабельные области антитела, ответственные за высокоизбирательное распознавание и связывание антигена, но также и константные области тяжелых и легких полипептидных цепей антитела, обычно присутствующие в эндогенно продуцируемых антителах, и в некоторых случаях даже декорирование по одному или более чем одному сайту углеводами. Такое гликозилирование обычно является отличительным признаком IgG-формата, и части этих константных областей составляют так называемую Fc-область целого антитела, которая, как известно, индуцирует различные эффекторные функции in vivo. Кроме того, Fc-область опосредует связывание IgG с Fc-рецептором и, следовательно, пролонгирование периода полувыведения in vivo, а также облегчение "хоминга" IgG к местам с повышенным присутствием Fc-рецепторов, например, к воспаленной ткани. Преимущественно, IgG антитело представляет собой IgG1 антитело или IgG4 антитело, форматы, которые являются предпочтительными, так как их механизм действия in vivo является особенно хорошо изученным и охарактеризованным. Это особенно верно для IgG1 антител. Согласно другому воплощению изобретения фрагмент человеческого моноклонального антитела может представлять собой scFv, однодоменное антитело, Fv, антитело VHH, диатело, тандемное диатело, Fab, Fab' или F(ab)2. Эти форматы обычно можно разделить на два подкласса: на те фрагменты, которые состоят из одной полипептидной цепи, и на те, которые содержат по меньшей мере две полипептидные цепи. Члены первого подкласса включают в себя scFv (содержащий одну VH-область и одну VL-область, соединенные в одну полипептидную цепь через полипептидный линкер); однодоменное антитело (содержащее одну вариабельную область антитела), такое как антитело VHH (содержащее одну VH-область). Члены последнего подкласса включают в себя Fv (содержащий одну VH-область и одну VL-область в виде отдельных полипептидных цепей, которые не связаны ковалентно друг с другом); диатело (включающее две ковалентно несвязанные полипептидные цепи, каждая из которых содержит две вариабельные области антитела: обычно одну VH и одну VL на полипептидную цепь, где эти две полипептидные цепи организованы в конформацию "голова-к-хвосту", так что образуется молекула бивалентного антитела); тандемное диатело (биспецифические одноцепочечные Fv антитела, содержащие четыре ковалентно связанные вариабельные VH- и VL-области иммуноглобулина двух разных специфичностей, образующие гомодимер, который в два раза больше диатела, описанного выше); Fab (содержащий в качестве одной полипептидной цепи целую легкую цепь антитела, которая сама включает VL-область и всю константную область легкой цепи и, в качестве другой полипептидной цепи, часть тяжелой цепи антитела, содержащей всю VH-область и часть константной области тяжелой цепи, где две указанные полипептидные цепи связаны между собой через межцепьевую дисульфидную связь); Fab' (в виде Fab, описанного выше, за исключением дополнительных восстановленных дисульфидных связей, имеющихся на тяжелой цепи антитела); и F(ab)2 (содержащий две молекулы Fab', где каждая молекула Fab' связана с другой соответствующей молекулой Fab' через межцепьевые дисульфидные связи). В общем, фрагменты антител описанных выше типов обеспечивают большую гибкость в приспособлении, например, фармакокинетических свойств антитела, желательного для терапевтического введения, к конкретным имеющимся потребностям. Например, может быть желательным уменьшение размера вводимого антитела для того, чтобы увеличить степень проникновения в ткань, когда известно, что ткани, которые лечат, являются недостаточно васкуляризованными (например, суставы). При некоторых обстоятельствах также может быть желательным увеличение скорости, с которой терапевтическое антитело элиминируется из организма, где указанная скорость обычно ускоряется при уменьшении размера вводимого антитела. Согласно другому воплощению изобретения указанное человеческое моноклональное антитело или его фрагмент могут быть представлены в моновалентной моноспецифической; поливалентной моноспецифической, конкретно в бивалентной моноспецифической; или в поливалентной полиспецифической, конкретно в бивалентной биспецифической формах. В общем, поливалентное моноспецифическое, конкретно бивалентное моноспецифическое антитело, такое как целый IgG человека, как описано выше, может иметь терапевтическое преимущество в том, что нейтрализация, осуществляемая таким антителом, потенцируется эффектами авидности, т.е. связыванием одного и того же антитела со многими молекулами одного и того же антигена, здесь GM-CSF приматов. Несколько форм моновалентных моноспецифических фрагментов антитела по изобретению было описано выше (например, scFv, Fv, VHH или однодоменное антитело). Поливалентные полиспецифические, в частности бивалентные биспецифические, формы человеческого моноклонального антитела против GM-CSF приматов по изобретению могут включать целый IgG, в котором один связывающий домен связывается с GM-CSF приматов, тогда как другой связывающий домен связывается с другим антигеном, отличным от GM-CSF приматов. Другая поливалентная полиспецифическая, в частности бивалентная биспецифическая, форма может представлять собой преимущественно человеческое одноцепочечное биспецифическое антитело, т.е. конструкцию рекомбинантного человеческого антитела, содержащую две структурные единицы scFv, как описано выше, соединенные в одну непрерывную полипептидную цепь коротким полипептидным спейсером, вставленным между ними, как общеизвестно в данной области (см., например, WO 99/54440 относительно анти-CD19 х анти-CD3 биспецифического одноцепочечного антитела). Здесь один scFv участок биспецифического одноцепочечного антитела, содержащегося в биспецифическом одноцепочечном антителе, будет специфически связывать GM-CSF приматов, как указано выше, тогда как другой соответствующий scFv участок этого биспецифического одноцепочечного антитела будет связывать другой антиген, который, как было определено, имеет терапевтическую пользу. Согласно другому воплощению может быть получено производное человеческого моноклонального антитела или его фрагмента, например, с органическим полимером, например, с одной или более чем одной молекулой полиэтиленгликоля ("PEG") и/или поливинилпирролидона ("PVP"). Как известно в данной области, получение такого производного может быть полезным в модуляции фармакодинамических свойств антител или их фрагментов. Особенно предпочтительными являются молекулы PEG, полученные в виде PEG-малеимида, обеспечивающего конъюгацию с антителом или его фрагментом сайт-специфическим образом через сульфгидрильную группу аминокислоты цистеина. Особенно предпочтительными из них являются 20 кДа и/или 40 кДа PEG-малеимид как в разветвленной форме, так и в прямоцепочечной форме. Особенно полезным может быть увеличение эффективной молекулярной массы меньших фрагментов человеческих антител против GM-CSF приматов, таких как scFv фрагменты, путем связывания последних с одной или более чем одной молекулой PEG, особенно с PEG-малеимидом. Согласно другому воплощению изобретения человеческое моноклональное антитело или его фрагмент специфически связывается с эпитопом, в частности с прерывистым эпитопом человеческого или не являющегося человеческим GM-CSF приматов, содержащим аминокислоты 23-27 (RRLLN) и/или аминокислоты 65-77 (GLR/QGSLTKLKGPL). Вариабельность в положении 67 в пределах отрезка аминокислотной последовательности 65-77, указанного выше, отражает гетерогенность в этом участке GM-CSF приматов, с одной стороны, между GM-CSF человека и гиббона (у которых в положении 67 находится R), а с другой стороны, между GM-CSF обезьян из семейства макак, например макака-крабоеда и макака-резуса (у которых в положении 67 находится Q). В том виде, как она здесь используется, нумерация человеческого и не являющегося человеческим GM-CSF приматов относится к нумерации зрелого GM-CSF, т.е. GM-CSF без его 17-аминокислотной сигнальной последовательности (общая длина зрелого GM-CSF и у человека, и у приматов, не являющихся человеком, описанных выше, составляет 127 аминокислот). Последовательность GM-CSF человека и GM-CSF гиббона является следующей: Последовательность GM-CSF у определенных членов семейства обезьян макак, таких как, например, макак-резус и макак-крабоед, является следующей: Минимальный эпитоп, преимущественно прерывистый эпитоп, связываемый человеческим моноклональным антителом по изобретению (или его фрагментом), как описано выше, указан в приведенной выше последовательности GM-CSF жирным шрифтом. Термин "прерывистый эпитоп" в том виде, как он здесь используется, следует понимать как по меньшей мере два несмежных отрезка аминокислотной последовательности в данной полипептидной цепи, здесь зрелого человеческого и не являющегося человеческим GM-CSF приматов, которые одновременно и специфически (как представлено выше) связываются антителом. Согласно этому определению такое одновременное специфическое связывание может представлять собой связывание полипептида GM-CSF в линейной форме. Здесь можно представить, что зрелый полипептид GM-CSF образует протяженную петлю, в одном участке которой две последовательности, указанные выше жирным шрифтом, выравниваются, например, более или менее параллельно и поблизости друг от друга. В этом состоянии они специфически и одновременно связываются фрагментом антитела по изобретению. Согласно этому определению одновременное специфическое связывание двух отрезков последовательности зрелого GM-CSF, указанных выше, также может принимать форму связывания антитела с конформационным эпитопом. Здесь зрелый GM-CSF уже образовал свою третичную конформацию, в которой он обычно существует in vivo (Sun, H.W., J. Bernhagen, et al. (1996). Proc. Natl. Acad. Sci. USA 93, 5191-6). В этой третичной конформации полипептидная цепь зрелого GM-CSF свернута таким образом, чтобы привести два отрезка последовательности, указанных выше, в пространственную близость, например, на наружной поверхности конкретной области зрелого, свернутого GM-CSF, где они затем распознаются благодаря их трехмерной конформации в контексте окружающих полипептидных последовательностей. В предпочтительном воплощении приведенный выше (прерывистый) эпитоп дополнительно содержит аминокислоты 28-31 (LSRD), указанные курсивом в приведенных выше последовательностях человеческого и не являющегося человеческим GM-CSF приматов. В особенно предпочтительном воплощении любой из указанных выше (прерывистых) эпитопов дополнительно содержит аминокислоты 32-33 (ТА) и/или аминокислоты 21-22 (ЕА), где каждый из этих отрезков подчеркнут в приведенных выше последовательностях человеческого и не являющегося человеческим GM-CSF приматов. Согласно другому воплощению изобретения человеческое моноклональное антитело или его фрагмент содержат в вариабельной области его тяжелой цепи область CDR3, содержащую аминокислотную последовательность, выбранную из группы, состоящей из последовательностей, определенных в любой из SEQ ID NO: 1-13 или 56. Предпочтительным является человеческое моноклональное антитело или его фрагмент, содержащие последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 1; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 2; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 3; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 4; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 5; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 6; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 7; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 8; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 9; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 10; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 11; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 12; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 13; или содержащее последовательность CDR1 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 14, последовательность CDR2 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 15, и последовательность CDR3 вариабельной области тяжелой цепи, как представлено в SEQ ID NO: 56. Еще более предпочтительно любая из 14 комбинаций последовательностей CDR1, CDR2 и CDR3, определенных выше, существует в человеческом моноклональном антителе или его фрагменте, дополнительно содержащем в вариабельной области его легкой цепи область CDR1, содержащую аминокислотную последовательность, определенную в SEQ ID NO: 16, область CDR2, содержащую аминокислотную последовательность, определенную в SEQ ID NO: 17, и область CDR3, содержащую аминокислотную последовательность, определенную в SEQ ID NO: 18. Согласно другому воплощению человеческое моноклональное антитело по изобретению или его фрагмент содержат в вариабельной области его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 19. Предпочтительным является человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 20; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 21; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 22; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 23; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 24; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 25; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 26; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 27; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 28; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 29; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 30; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 31; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 32; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 33; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 52; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 19, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 53. Согласно другому воплощению человеческое моноклональное антитело по изобретению или его фрагмент содержат в вариабельной области его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 54. Предпочтительным является человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 20; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 21; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 22; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 23; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 24; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 25; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 26; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 27; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 28; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 29; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 30; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 31; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 32; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 33; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 52; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 54, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 53. Согласно другому воплощению человеческое моноклональное антитело по изобретению или его фрагмент содержат в вариабельной области его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 55. Предпочтительным является человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 20; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 21; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 22; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 23; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 24; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 25; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 26; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 27; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 28; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 29; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 30; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 31; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 32; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 33; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 52; или человеческое моноклональное антитело или его фрагмент, вариабельная область легкой цепи которого содержит аминокислотную последовательность, как представлено в SEQ ID NO: 55, и вариабельная область тяжелой цепи содержит аминокислотную последовательность, как представлено в SEQ ID NO: 53. В предпочтительном воплощении предложено человеческое моноклональное антитело или его фрагмент, содержащее в вариабельной области его легкой цепи область CDR1, содержащую аминокислотную последовательность, как представлено в SEQ ID NO: 16, область CDR2, имеющую аминокислотную последовательность, как представлено в SEQ ID NO: 17, и область CDR3, имеющую аминокислотную последовательность, как представлено в SEQ ID NO: 18, и содержащее в вариабельной области его тяжелой цепи область CDR1, содержащую аминокислотную последовательность, как представлено в SEQ ID NO: 14, область CDR2, имеющую аминокислотную последовательность, как представлено в SEQ ID NO: 15, и область CDR3, имеющую аминокислотную последовательность, как представлено в любой из SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 56. В другом предпочтительном воплощении данное человеческое моноклональное антитело содержит в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 35; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 36; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 37; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 38; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 39; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 40; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 41; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 42; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 43; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 44; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 45; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 46; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 47; или в его легкой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 34, и в его тяжелой цепи аминокислотную последовательность, как представлено в SEQ ID NO: 48. Согласно предпочтительным воплощениям, приведенным выше, предложены молекулы человеческого моноклонального антитела и/или его фрагмента, которые являются особенно полезными в качестве нейтрализаторов активности GM-CSF приматов, особенно GM-CSF человека. Человеческие моноклональные антитела или их фрагменты согласно этим особенно предпочтительным воплощениям являются весьма полезными по нескольким причинам. Во-первых, они высокоспецифически распознают GM-CSF приматов, т.е. в смеси GM-CSF приматов с другими колониестимулирующими факторами приматов (например, с G-CSF или M-CSF) связывающие молекулы согласно этим особенно предпочтительным воплощениям являются в высокой степени избирательными к GM-CSF приматов, тогда как другие колониестимулирующие факторы в той же среде не распознаются. Это означает, что ожидается, что человеческое моноклональное антитело или его фрагмент согласно этим воплощениям при введении человеку будут специфически связываться и нейтрализовать только желательную мишень, тогда как другие нежелательные мишени не связываются и не нейтрализуются. В конечном счете, это приводит к высокой степени предсказуемости относительно терапевтического способа действия in vivo. Во-вторых, связыватели согласно этим особенно предпочтительным воплощениям связываются с GM-CSF приматов с чрезвычайно высокой аффинностью. Для молекул этого класса наблюдали значения K D от примерно 4 ×10 -9 М до таких низких значений, как примерно 0,04 ×10 -9 М, причем последнее соответствует примерно 40 пМ. Поскольку кинетика скорости ассоциации (on-rate) таких молекул в водных средах главным образом контролируется диффузией и, следовательно, не может улучшаться в большей степени, чем позволяют локальные условия диффузии при физиологических условиях, низкая K D возникает главным образом в результате кинетики скорости диссоциации (k off ), которая для наивысшей аффинности связывателя антитела составляет приблизительно 10 -5 с -1 . Это означает, что как только образуется комплекс между человеческим моноклональным антителом или его фрагментом согласно любому из этих воплощений изобретения, с одной стороны, и GM-CSF приматов, с другой стороны, он распадается с трудом или, по меньшей мере, не быстро. Для связывающих молекул, предназначенных быть нейтрализаторами биологической активности, эти характеристики являются весьма полезными, так как желательный нейтрализующий эффект обычно длится только при условии, что молекула, биологическая активность которой подлежит нейтрализации (здесь GM-CSF приматов), остается связанной с нейтрализующей связывающей молекулой. Так что нейтрализующая молекула, которая остается связанной с ее заданной мишенью в течение длительного периода времени, будет продолжать осуществлять нейтрализацию в течение соответствующего длительного периода времени. Высокая аффинность связывания человеческих моноклональных антител или их фрагментов с GM-CSF приматов имеет дополнительное преимущество. Обычно антитела или их фрагменты будут элиминироваться из кровотока пациента в зависимости от их размера, причем меньшие молекулы экскретируются и элиминируются раньше, чем более крупные молекулы. Поскольку комплекс двух полипептидов - антитела или фрагмента антитела и связанного GM-CSF - очевидно больше, чем одно антитело, низкая k off , упомянутая выше, оказывает эффект в том, что терапевтический нейтрализатор экскретируется и элиминируется из организма человека более медленно, чем это было бы в том случае, если бы он не был связан с GM-CSF. Таким образом, увеличивается не только величина нейтрализующей активности, но также и ее продолжительность in vivo. Наконец, нейтрализующая активность, определенная для связывателей согласно этим особенно предпочтительным воплощениям, является неожиданно высокой. Как будет описано более подробно здесь ниже, нейтрализующую активность измеряли in vitro с использованием анализа ингибирования роста TF-1 (Kitamura, T. et al. (1989). J. Cell. Physiol. 140, 323-34). В качестве показателя нейтрализующего потенциала измеряли значения IC 50 , где IC 50 представляет собой концентрацию человеческого моноклонального антитела или его фрагмента согласно любому из этих воплощений изобретения, необходимую для вызывания полумаксимального ингибирования пролиферации клеток TF-1. Для человеческих моноклональных антител или их фрагментов согласно любому из этих воплощений изобретения было определено значение IC 50 приблизительно 3 ×10 -10 М или примерно 0,3 нМ. Следовательно, связывающие молекулы согласно любому из этих воплощений изобретения являются высокоэффективными нейтрализаторами активности GM-CSF приматов. В заключение, в таком случае человеческое моноклональное антитело или его фрагмент согласно любому из приведенных выше воплощений изобретения демонстрирует высокую степень распознавания желательного антигена, связывает этот антиген чрезвычайно прочно и в течение длительного периода времени и демонстрирует весьма мощную нейтрализующую активность в течение длительного периода времени, пока оно остается связанным. В то же самое время длительная персистенция комплекса связыватель-антиген замедляет элиминацию этого связывателя из организма, удлиняя тем самым продолжительность желательного терапевтического эффекта in vivo. Согласно другому аспекту изобретения предложено человеческое моноклональное антитело или его фрагмент, содержащие аминокислотную последовательность, имеющую по меньшей мере 70%-ную гомологию с аминокислотной последовательностью, как представлено в любой из SEQ ID NO: 1-48 и/или 52-56. Гомологию можно определить стандартными программами выравнивания последовательностей, такими как Vector NTI (InforMax ™, Maryland, USA). Такие программы сравнивают выравненные последовательности по принципу аминокислота за аминокислотой и в них можно установить разные уровни строгости для сравнения (например, идентичная аминокислота, консервативная аминокислотная замена и т.д.). В том виде, как здесь используется данный термин, две рассматриваемые аминокислоты считаются "консервативными заменами" друг друга, если они принадлежат к одному и тому же химическому классу, т.е. к классу кислых, неполярных, незаряженных полярных и основных аминокислот. В качестве неограничивающего примера две разные аминокислоты, принадлежащие к классу неполярных аминокислот, считаются "консервативными заменами" друг друга, даже если эти две аминокислоты не являются идентичными, тогда как неполярная аминокислота, с одной стороны, и основная аминокислота, с другой стороны, не считаются "консервативными заменами" друг друга. В табл. 3.1 в "Molecular Biology of the Cell", 4 th Edition (2002), by Alberts, Johnson, Lewis, Raff, Roberts and Walter, аминокислоты сгруппированы в четырех основных группах: кислые, неполярные, незаряженные полярные и основные. В целях настоящего изобретения такое группирование можно использовать для определения того, является ли конкретная аминокислота консервативной заменой другой рассматриваемой аминокислоты или нет. Согласно другому аспекту изобретения предложена полинуклеотидная молекула, имеющая нуклеотидную последовательность, кодирующую аминокислотную последовательность, как представлено в любой из SEQ ID NO: 1-48 и/или 52-56, или нуклеотидную последовательность, демонстрирующую по меньшей мере 70%-ную гомологию с ними, где гомологию можно определить путем сравнения нуклеотидной последовательности, кодирующей аминокислотную последовательность любой из SEQ ID NO: 1-48 и/или 52-56 с рассматриваемой нуклеотидной последовательностью путем выравнивания последовательностей (как описано выше для аминокислотных последовательностей), где нуклеотид в рассматриваемой последовательности считается гомологичным, если он является либо идентичным соответствующему нуклеотиду в нуклеотидной последовательности, кодирующей соответствующую аминокислотную последовательность из любой из SEQ ID NO: 1-48 и/или 52-56, или если одно или более чем одно отклонение(я) в нуклеотидах рассматриваемой последовательности от соответствующего одного или более чем одного нуклеотида(ов) в нуклеотидной последовательности, кодирующей аминокислотную последовательность из любой из SEQ ID NO: 1-48 и/или 52-56, приводит к образованию нуклеотидного триплета, который при трансляции дает аминокислоту, которая либо идентична (благодаря вырожденному триплету) соответствующей аминокислоте в соответствующей аминокислотной последовательности из любой из SEQ ID NO: 1-48 и/или 52-56, либо является консервативной заменой соответствующей аминокислоты в соответствующей аминокислотной последовательности из любой из SEQ ID NO: 1-48 и/или 52-56. Здесь термин "консервативная замена" следует понимать, как описано выше. Согласно другому аспекту изобретения предложена фармацевтическая композиция, содержащая человеческое моноклональное антитело или его фрагмент, или полинуклеотидную молекулу, имеющую нуклеотидную последовательность, кодирующую аминокислотную последовательность, как представлено в любой из SEQ ID NO: 1-48 и/или 52-56, или кодирующую аминокислотную последовательность, содержащую аминокислотную последовательность, имеющую по меньшей мере 70%-ную гомологию с любой из SEQ ID NO: 1-48 и/или 52-56, где "гомологию" следует понимать, как объяснено в данном описании выше. Согласно данному изобретению термин "фармацевтическая композиция" относится к композиции для введения пациенту, преимущественно пациенту, являющемуся человеком. В предпочтительном воплощении данная фармацевтическая композиция содержит композицию для парентерального, трансдермального, интралюминального, внутриартериального, интратекального и/или интраназального введения или путем прямой инъекции в ткань. Конкретно предусматривается, что указанную фармацевтическую композицию вводят пациенту посредством инфузии или инъекции. Введение подходящих композиций можно осуществлять разными путями, например, внутривенным, интраперитонеальным, подкожным, внутримышечным, местным или интрадермальным введением. Фармацевтическая композиция по настоящему изобретению может дополнительно содержать фармацевтически приемлемый носитель. Примеры подходящих фармацевтических носителей хорошо известны в данной области и включают в себя забуференные фосфатом физиологические растворы, воду, эмульсии, такие как эмульсии типа масло в воде, разные типы увлажнителей, стерильные растворы, липосомы и т.д. Композиции, содержащие такие носители, можно приготавливать в виде препаратов с помощью общеизвестных традиционных способов. Эти фармацевтические композиции можно вводить субъекту в подходящей дозе. Схема дозирования определяется лечащим врачом и клиническими факторами. Как хорошо известно в области медицины, дозировки для любого пациента зависят от многих факторов, включая размер тела пациента, площадь поверхности тела, возраст, конкретное соединение, подлежащее введению, пол, время и путь введения, общее состояние здоровья и другие лекарственные средства, которые вводятся одновременно. Препараты для парентерального введения включают в себя стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъецируемые органические эфиры, такие как этилолеат. Водные носители включают в себя воду, спиртовые/водные растворы, эмульсии или суспензии, включающие физиологический раствор и забуференные среды. Парентеральные наполнители включают в себя раствор хлорида натрия, декстрозу Рингера, декстрозу и хлорид натрия, лактат Рингера, или нелетучие масла. Внутривенные наполнители включают в себя жидкие и питательные наполнители, электролитные наполнители (такие как наполнители на основе декстрозы Рингера) и т.п. Также могут присутствовать консерванты и другие добавки, такие как, например, противомикробные агенты, антиоксиданты, хелатирующие агенты, инертные газы и т.п. Кроме того, фармацевтическая композиция по настоящему изобретению может содержать белковые носители, подобные, например, сывороточному альбумину или иммуноглобулину, предпочтительно человеческого происхождения. Предусматривается, что фармацевтическая композиция по данному изобретению, помимо человеческого моноклонального антитела или его фрагментов (как описано в этом изобретении), может содержать дополнительные биологически активные агенты, в зависимости от предполагаемого применения данной фармацевтической композиции. Такие агенты могут представлять собой лекарственные средства, действующие на желудочно-кишечную систему, лекарственные средства, действующие как цитостатики, лекарственные средства, предотвращающие гиперурикемию, лекарственные средства, ингибирующие иммунореакции (например, кортикостероиды), лекарственные средства, модулирующие воспалительную реакцию, лекарственные средства, действующие на систему кровообращения и/или такие агенты, как цитокины, известные в данной области. Согласно другому аспекту изобретения предложено применение человеческого моноклонального антитела или его фрагмента, как описано здесь выше, или полинуклеотидной молекулы, содержащей нуклеотидную последовательность, кодирующую аминокислотную последовательность, как представлено в любой из SEQ ID NO: 1-48 и/или 52-56, или кодирующую аминокислотную последовательность, содержащую аминокислотную последовательность, имеющую по меньшей мере 70%-ную гомологию с любой из SEQ ID NO: 1-48 и/или 52-56, где "гомологию" следует понимать так, как объяснено в данном описании выше, в изготовлении лекарственного средства, возможно содержащего один или более чем один противовоспалительный агент, для лечения воспалительных заболеваний. Воспалительные заболевания преимущественно выбраны из группы, состоящей из ревматоидного артрита (RA) (включая RA, который является устойчивым к лечению нейтрализаторами TNF-альфа), астмы, рассеянного склероза (MS), хронического обструктивного заболевания легких (COPD), острого респираторного дистресс-синдрома (ARDS), болезни Крона, идиопатического легочного фиброза (IPF), воспалительного заболевания кишечника (IBD), увеита, дегенерации желтого пятна, колита, псориаза, Уоллеровской дегенерации, антифосфолипидного синдрома (APS), острого коронарного синдрома, рестеноза, атеросклероза, рецидивирующего полихондрита (RP), острого или хронического гепатита, неудачного приживления ортопедических имплантатов, гломерулонефрита, волчанки или аутоиммунных расстройств. Особый интерес представляет собой применение человеческого моноклонального антитела или его фрагмента согласно данному изобретению для получения лекарственного средства для лечения RA (включая RA, который является устойчивым к лечению нейтрализаторами TNF-альфа), астмы, MS и/или болезни Крона. Что касается RA, астмы и/или MS, т.е. две популярные теории относительно патогенеза ревматоидного артрита (RA). Первая принимает, что Т-клетка, через взаимодействие с еще неидентифицированным антигеном, является первичной клеткой, ответственной за инициацию заболевания, а также за развитие хронического воспалительного процесса. Эта теория основана на известной связи RA с главными антигенами гистосовместимости класса II, большим количеством CD4+ Т-клеток и с использованием деформированного гена рецептора Т-клеток в синовиальной оболочке, пораженной RA. Известно, что GM-CSF усиливает антиген-презентирующую функцию через увеличение экспрессии поверхностных молекул МНС класса II, и GM-CSF продуцируется Т-клетками, что указывает на предполагаемую роль GM-CSF в развитии заболевания согласно гипотезе, основанной на Т-клетках. Вторая теория принимает, что, хотя Т-клетки могут играть важную роль в инициации данного заболевания, хроническое воспаление может долго поддерживаться макрофагами и фибробластами независимым от Т-клеток образом. Эта теория основана на относительном отсутствии фенотипов активированных Т-клеток при хроническом RA и на преобладании фенотипов активированных макрофагов и фибробластов. GM-CSF является мощным стимулятором макрофагов и стимулирует пролиферацию моноцитов и макрофагов. Известно, что GM-CSF, продуцируемый главным образом "эффекторными клетками" (макрофагами) и клетками соединительной ткани (фибробластами), экспрессируется в большом количестве в синовиальной оболочке, пораженной RA, и в синовиальной жидкости, как измерено с помощью ELISA или исследований мРНК. Согласно "макрофагально-фибробластной теории" RA, эти два типа клеток, по-видимому, в значительной степени отвечают за создание состояния длительно поддерживающегося воспаления, при котором участие Т-клеток может больше не быть критическим. При этом сценарии активированный макрофаг непрерывно секретирует IL-1 и TNF, которые поддерживают синовиальный фибробласт в активированном состоянии. Фибробласт, в свою очередь, секретирует большие количества: а) цитокинов - IL6, IL8 и GM-CSF; б) простагландинов и в) протеаз. GM-CSF подвергается действию обратной связи для стимуляции созревания вновь рекрутированных моноцитов до макрофагов. IL8 и IL6 способствуют рекрутингу и/или активации других клеточных популяций, тогда как простагландины и протеазы непосредственно действуют при эрозии и разрушении близлежащих соединительных тканей, таких как кости и хрящи. Что касается болезни Крона, то рекомбинантный человеческий гранулоцитарно-макрофагальный колониестимулирующий фактор (rGM-CSF) из дрожжей продемонстрировал эффективность при лечении болезни Крона, тяжесть которой варьировала от умеренной до тяжелой формы (Dieckgraefe B.K., Korzenik J.R. (2002). Lancet 360, 1478-80). С этого времени в нескольких обзорных статьях были высказаны предположения о терапевтическом эффекте этого мощного провоспалительного цитокина в данном заболевании, которое, как считают, имеет воспалительную природу. Возможные объяснения способа действия rGM-CSF включали компонент иммунонедостаточности при болезни Крона, деформацию Th 2 и экспансию дендритных клеток, стимулирующих дифференцировку регуляторных Т-клеток (Wilk N.J., Viney J.L. (2002). Curr. Opin. Invest. Drugs 3, 1291-6; Folwaczny С. et al. (2003). Eur. J. Gastroenterol. Hepatol. 15, 621-6). Авторы изобретения считают, что, по-видимому, существует более простой способ действия, который в то же время больше согласуется с известной ролью GM-CSF в других провоспалительных заболеваниях. GM-CSF является одним из самых мощных известных адъювантов, вследствие чего данный цитокин используют для совместного введения во многих продолжающихся в настоящее время исследованиях по вакцинации. В то же время GM-CSF является высокоиммуногенным (Ragnhammar P. et al. (1994). Blood 84, 4078-87). В совсем недавнем исследовании (Rini В. et al. (2005) Cytokine 29, 56-66) было показано, что ежесуточное подкожное лечение с использованием rGM-CSF из дрожжей, которое осуществляли при исследовании болезни Крона (Dieckgraefe B.K., Korzenik J.R. (2002). Lancet 360, 1478-80), в течение трех месяцев приводило у 87% (13/15) пациентов с раком простаты к развитию антител против данного цитокина. У шестидесяти процентов пациентов (9/15) появлялись (поликлональные) антитела, нейтрализующие GM-CSF. В исследовании болезни Крона не исследовалась возможность нейтрализующей реакции на GM-CSF, а также не определялись уровни GM-CSF в сыворотке при терапии. В пределах объема данного воплощения изобретения предполагают, что пациенты с болезнью Крона, которых лечили rGM-CSF, не реагировали непосредственно только на иммуностимулирующую активность данного цитокина, но также, по меньшей мере частично, клинически реагировали на антительный ответ, нейтрализующий как введенный, так и эндогенный GM-CSF, который, как известно, сверхпродуцируется при болезни Крона (Agnholt J. et al. (2004) Eur. J. Gastroenterol. Hepatol. 16, 649-55). Тогда нейтрализующие антитела против GM-CSF могут иметь аналогичную терапевтическую активность при болезни Крона, как и rGM-CSF, и их следует рассматривать в качестве альтернативного терапевтического подхода, как это предусмотрено выше. В другом аспекте изобретения предложено применение человеческого моноклонального антитела или его фрагмента, как описано выше, или полинуклеотидной молекулы, содержащей нуклеотидную последовательность, кодирующую аминокислотную последовательность, как представлено в любой из SEQ ID NO: 1-48 и/или 52-56, или кодирующую аминокислотную последовательность, включающую аминокислотную последовательность, имеющую по меньшей мере 70%-ную гомологию с любой из SEQ ID NO: 1-48 и/или 52-56, где "гомологию" следует понимать, как объяснено выше, в изготовлении лекарственного средства, возможно содержащего один или более чем один дополнительный противораковый агент, для лечения опухолевого заболевания или другого состояния с замедленным апоптозом клеток, повышенным выживанием или пролиферацией клеток. Предпочтительное опухолевое заболевание представляет собой рак, из которого особенно предпочтительными являются лейкоз, множественная миелома, карцинома желудка или карцинома кожи. Olver и др. ((2002) Cancer Chemother Pharmacol. 50, 171-8) подкожно применяли антагонист GM-CSF E21R у пациентов с солидными опухолями, для которых известно, что в них экспрессируются рецепторы GM-CSF, что приводило лишь к временному снижению уровней PSA в сыворотке. Кроме того, применение этого антагониста GM-CSF при остром миелоидном лейкозе ("AML") не выявило клинической активности (Jakupovic et al. (2004) Blood 103, 3230-2.). Более того, применение моноклональных антител против GM-CSF у пациентов с AML не выявило антилейкозного эффекта, несмотря на достаточные сывороточные уровни и биологическую активность данного антитела in vivo (Bouabdallah et al. (1998) Leuk Lymphoma 30, 539-49). Поэтому авторы изобретения заключили, что у пациентов с AML лечение антителами против GM-CSF является неэффективным. Данное изобретение теперь будет описано более подробно в следующих неограничивающих примерах и графических материалах, обзор которых приведен ниже. Фиг. 1. Интенсивность поглощения (прямо пропорциональна силе связывания) для целого ряда молекул scFv против rhGM-CSF, полученных после четырех или пяти раундов "пэннинга" в фаговом дисплее, как определено с помощью ELISA. Фиг. 2. Средняя интенсивность флуоресценции (обратно пропорциональна силе нейтрализации) для целого ряда scFv против rhGM-CSF и других тестируемых молекул, как определено с помощью анализа на основе проточной цитометрии. Фиг. 3. Результаты анализа ингибирования пролиферации TF-1, проведенного с использованием молекулы scFv 5-306 против rhGM-CSF. Фиг. 4. Интенсивность поглощения (прямо пропорциональна силе связывания) для целого ряда человеческих молекул scFv против rhGM-CSF, полученных после четырех или пяти раундов "пэннинга" в фаговом дисплее, как определено с помощью ELISA. Фиг. 5. Результаты анализа ингибирования пролиферации TF-1, проведенного с использованием различных репрезентативных удачных человеческих scFv против rhGM-CSF. Фиг. 6. Специфичность связывания человеческих моноклональных антител против GM-CSF человека и других колониестимулирующих факторов человека. Фиг. 7. Измерения поверхностного плазмонного резонанса, характеризующего кинетическое связывание человеческих моноклональных антител и их фрагментов против GM-CSF. Фиг. 8А. Выравнивание последовательностей GM-CSF приматов, не являющихся человеком, и GM-CSF человека. Фиг. 8В. Радиограмма пептидных пятен, демонстрирующая связывание фрагмента человеческого моноклонального антитела против GM-CSF с GM-CSF человека. Фиг. 9. Качественные результаты анализа ингибирования пролиферации TF-1, проведенного с использованием различных репрезентативных scFv-фрагментов человеческих антител против rhGM-CSF. Фиг. 10. Количественные результаты анализа ингибирования пролиферации TF-1, проведенного с использованием различных репрезентативных человеческих IgG и соответствующих scFv-фрагментов против rhGM-CSF. Фиг. 11. Количественные результаты анализа продукции IL8, проведенного с использованием различных репрезентативных scFv-фрагментов человеческих антител против rhGM-CSF. Фиг. 12. Количественные результаты анализа ингибирования пролиферации TF-1, проведенного с использованием различных репрезентативных человеческих IgG и соответствующих scFv-фрагментов против macGM-CSF (GM-CSF макака). Фиг. 13. Результаты сравнительного исследования связывания, показывающие селективность связывания антитела IgG В против GM-CSF с рекомбинантным GM-CSF человека и GM-CSF из разных видов, не являющихся приматами. Фиг. 14. Результаты анализа зависимости нейтрализующего потенциала антитела IgG В против GM-CSF от гликозилирования GM-CSF. Фиг. 15. Результаты исследования эффекта антитела IgG В против GM-CSF на GM-CSF-опосредованное выживание эозинофилов. Фиг. 16 Результаты исследования эффекта антитела IgG В против GM-CSF на GM-CSF-опосредованную активацию эозинофилов. Фиг. 17. Результаты токсикологического исследования ex vivo с использованием антитела IgG В против GM-CSF, измеренного на основе фагоцитоза (А-С) и окислительного взрыва (D-F) гранулоцитами. Фиг. 18. Результаты исследования токсикологии ex vivo с использованием антитела IgG В против GM-CSF, измеренной на основе фагоцитоза (А-С) и окислительного взрыва (D-F) моноцитами. Пример 1. Получение рекомбинантного человеческого ("rh") GM-CSF антигена, используемого для получения нейтрализующих человеческих антител и их фрагментов. Пример 1.1. Клонирование, экспрессия и очистка rhGM-CSF антигена. Ген, кодирующий GM-CSF антиген человека, субклонировали в вектор pET22b(+) (Novagene, USA) из экспрессирующего вектора pORF-hGM-CSF (Novagen, USA) через ПЦР-введенные сайты распознавания рестрикционными ферментами NdeI и XhoI. hGM-CSF кодирующий ген в рЕТ22b(+) подвергали слиянию с лидерной последовательностью pelB, и он подходит для экспрессии в периплазме Е.coli. Продуцирование и очистку белка осуществляли, как описано производителем. Кратко, Е.coli BL21DE3 трансформировали экспрессирующей плазмидой и выращивали при 37 °С в селективной среде до оптической плотности 0,5-0,8 при 600 нм. Продуцирование белка индуцировали путем добавления IPTG до 1 мМ и снижения температуры до 25 °С. Получение периплазмы осуществляли с помощью осмотического шока, используя 20%-ный раствор сахарозы для избирательного разрушения клеточной стенки при сохранении интактной клеточной мембраны. Нативный hGM-CSF содержит два дисульфидных мостика, и экспрессия в окислительной периплазме Е.coli обеспечивает образование этих функционально важных дисульфидных мостиков. Рекомбинантный человеческий GM-CSF ("rhGM-CSF") очищали в процессе двухступенчатой очистки посредством аффинной хроматографии с иммобилизованным металлом (IMAC) и гель-фильтрации. Для хроматографии использовали систему FPLC Äkta ® и программу Unicorn ®. Все реактивы имели качество, подходящее для использования в исследованиях, и были приобретены у Sigma (Deisenhofen) или Merck (Darmstadt). IMAC осуществляли с использованием колонки Qiagen Ni-NTA Superflow согласно протоколу производителя. Колонку уравновешивали буфером А2 (20 мМ фосфата натрия, pH 7,2, 0,4 М NaCl) и препарат периплазмы ("РРР") (100 мл) наносили на колонку (2 мл) при скорости протока 2 мл/мин. Колонку промывали 5 объемами колонки 5%-ного буфера В2 (20 мМ фосфата натрия, pH 7,2, 0,4 М NaCl, 0,5 М имидазола) для удаления несвязавшегося образца. Связанный белок элюировали с использованием 100%-ного буфера В2 в 5 объемах колонки. Элюированные белковые фракции объединяли для дальнейшей очистки. Гель-фильтрацию осуществляли на колонке Superdex 200 Prep Grade (Pharmacia), уравновешенной PBS (Gibco). Образцы элюированного белка (скорость протока 1 мл/мин) подвергали стандартному ПААГ-ДСН (электрофорез в полиакриламидном геле с додецилсульфатом натрия) и Вестерн-блоттингу для определения. Перед очисткой колонку калибровали для определения молекулярной массы (набор маркеров молекулярной массы, Sigma MW GF-200). Концентрации белка определяли, измеряя OD при 280 нм, и рассчитывали с использованием специфического для последовательности коэффициента молекулярной экстинкции. Пример 1.2. Биотинилирование rhGM-CSF антигена. Для селекции фаговой библиотеки rhGM-CSF антиген, продуцируемый в E.coli (см. выше), биотинилировали. Биотинилирование осуществляли в PBS, содержащем 5% ДМСО (Sigma), с пятикратным молярным избытком EZ-Link Sulfo NHS-LC-LC-Biotin (Pierce) в течение 1 ч при комнатной температуре в смесителе для образцов (Dynal). Для разделения свободного биотина и биотинилированного rhGM-CSF антигена осуществляли анионообменную хроматографию (Resource Q, Amersham Biosciences) согласно стандартным протоколам. При обоих подходах (обозначенных А и В, описанных ниже) хроматография давала два пика элюции. В случае А первый элюированный пик вновь фракционировали посредством второй стадии анионообменной хроматографии (такие же условия, как описано выше) на два пика элюции. Затем полученные фракции последовательно разбавляли (разведения 1:2; исходная концентрация 6 мкг/мл, определенная из высоты пика), вносили их в 96-луночные планшеты для ELISA и осуществляли определение. Определение осуществляли, используя (А) антитело М500-А против GM-CSF человека (Sigma, 2,5 мкг/мл в PBS/1% БСА (бычий сывороточный альбумин)), определяемое поликлональным антителом козы, специфическим в отношении Fab2 мыши, конъюгированным с пероксидазой хрена (Dianova, 1 мкг/мл PBS/1% БСА), и (Б) материнское антитело (1 мкг/мл PBS/1% БСА), определяемое поликлональным антителом козы против антитела крысы, конъюгированным с пероксидазой хрена (Dianova, 1 мкг/мл PBS/1% БСА). Успешное биотинилирование продемонстрировали с помощью аналогичного эксперимента с использованием ELISA, который осуществляли с использованием стрептавидина, конъюгированного с пероксидазой хрена (Dako, 1 мкг/мл PBS/1% БСА). Сигнал проявляли путем добавления раствора субстрата OPD (Sigma) и определяли при длине волны 492 нм (эталонная длина волны 620 нм). Для оценки степени биотинилирования вышеупомянутую ELISA осуществляли с использованием фракций непосредственно от анионообменной хроматографии или после стадии инкубации с 6,7 ×10 7 магнитных стрептавидиновых шариков (Dynabeads M-280-Streptavidin, Dynal) с осторожным перемешиванием в течение 30 мин. Полученный супернатант вносили в лунки 96-луночных планшетов для ELISA и осуществляли определение, как описано выше. Результаты ELISA показали, что второй элюированный пик содержал биотинилированный rhGM-CSF и что примерно 95% элюированного rhGM-CSF было конъюгированным. Концентрации оценивали, используя в качестве стандарта исходный материал, и оказалось, что они составляли примерно 20 мкг/мл. Сохраняющуюся биоактивность биотин-меченого rhGM-CSF подтверждали в анализах пролиферации TF-1 согласно протоколам, описанным ниже, при характеристике одноцепочечных антител (scFv). Пример 1.3. Мечение rhGM-CSF антигена флуоресцеином. Для исследований связывания на клетках TF-1 рекомбинантный GM-CSF антиген человека, продуцируемый в Е.coli (см. пример 1.2 выше), конъюгировали с N-сукцинимидиловым эфиром флуоресцеин-5(6)-карбоксиамидокапроновой кислоты (Fluka, флуоресцеин-NHS). Стадию конъюгации осуществляли в боратном буфере (0,05 М борная кислота, 0,1 М NaCl, pH 8,5), содержащем 17,5% ДМСО, с пятикратным молярным избытком флуоресцеина-NHS в течение 1 ч при комнатной температуре в смесителе для образцов. Затем осуществляли гель-фильтрацию (Sephadex G25, Amersham Biosciences) для диссоциации флуоресцеин-меченого rhGM-CSF антигена из свободного флуоресцеин-NHS. В результате гель-фильтрации получали два пика, которые измеряли при длине волны 485 нм (эталонная длина волны 535 нм), тогда как первичный пик представляет собой FITC-меченый rhGM-CSF. Степень мечения определяли путем нахождения соотношения F/P конъюгата ([мг/мл]=(A 280 -0,35 ×А 493 ) ×1,08; F/P=(А 493 /73,000) ×(15,000/[мг/мл])). Определенная концентрация составляла 0,041 мг/мл с отношением F/P, равным 1,2. Пример 2. Получение и селекция человеческих нейтрализующих антител и их фрагментов против GM-CSF. Пример 2.1. Клонирование материнской VH из гибридомы НВ-9569. Термин "материнская" V-область в том виде, как он используется во всех вышеупомянутых примерах, означает, что рассматриваемая V-область происходит из молекулы целого иммуноглобулина. Термин "удачная" (hit) в том виде, как он используется во всех вышеупомянутых примерах, означает молекулу, о которой известно, что она связывает интересующий антиген, но данное связывание не было количественно оценено. "Удачной" является молекула на ранней стадии определения характеристик, для которой возможно уже было осуществелено продуцирование в малом масштабе. Такая молекула находится на стадии подтверждения характеристик. Термин "потенциальная" (lead) молекула в том виде, как он используется во всех вышеупомянутых примерах, означает молекулу, потенциалы связывания и нейтрализации которой были количественно охарактеризованы. Продуцирование "потенциальной" молекулы уже имело место в крупном масштабе. В следующих примерах описан один из возможных способов получения полностью человеческого моноклонального антитела - нейтрализатора GM-CSF и получения его фрагментов. Целью этого эксперимента является выделение и субклонирование гена, кодирующего VH в материнском mAb, продуцируемом линией гибридомных клеток НВ-9569. Гибридому НВ-9569 получали из АТСС (Американская коллекция типовых культур, США). Гибридомные клетки культивировали в полной ростовой среде АТСС: среда RPMI 1640, содержащая 2 мМ L-глутамина, и бикарботат натрия, содержание которого доведено до 1,5 г/л, 4,5 г/л глюкозы, 10 мМ HEPES и 1,0 мМ пирувата натрия, и дополненная 0,05 мМ 2-меркаптоэтанола, 10% фетальной коровьей сыворотки, 37 °С с 5% СО 2 . Для получения общей РНК использовали 1 ×10 7 клеток и РНК получали, как описано в руководстве к продукту Qiagen Omni-Skript Kit (Qiagen, Германия). кДНК синтезировали согласно стандартным способам (Sambrook, Cold Spring Harbor Laboratory Press 1989, Second Edition). Для выделения ДНК V-области тяжелой цепи осуществляли ОТ-ПЦР (полимеразная цепная реакция с использованием обратной транскриптазы) с использованием набора праймеров MHALT1 R.V: GCC GAA TTC CAC CAT GGR ATG SAG CTG KGT MAT SCT CTT и rlgG2a/b расы GSP: CAC ACC GCT GGA CAG GGC TCC AGA GTT CC. Для амплификации использовали следующую программу ПЦР: денатурацию при 94 °С в течение 15 с, отжиг праймеров при 52 °С в течение 50 с и удлинение праймеров при 72 °С в течение 90 с осуществляли в течение 40 раундов с последующим конечным удлинением при 72 °С в течение 10 мин. Затем выделяли ДНК V-фрагментов тяжелой цепи согласно стандартным протоколам. ДНК V-фрагмента тяжелой цепи клонировали в PCR script-CAM (Stratagene), как описано производителем. Последовательности идентифицировали секвенированием. Пример 2.2. Селекция VL человека. Целью этого эксперимента является селекция VL человека, которая может образовать пару с материнской VH, клонированной, как описано выше. Пример 2.2.1. Выделение РНК из селектированных IqD-положительных В-клеток. У пяти здоровых человеческих доноров брали 100 мл крови. Выделяли мононуклеарные клетки периферической крови (РВМС) в градиенте фикола согласно стандартным способам. Для селекции IgD-положительных клеток 1 мл антимышиных IgG-шариков (CELLection ™ Pan Mouse IgG Kit; DYNAL) покрывали 20 мкг мышиного антитела против человеческого IgD (PharMingen). Добавляли к шарикам приблизительно 2,5 ×10 7 РВМС и инкубировали при 4 °С в течение 15 мин. После промывки четыре раза 1 мл среды RPMI (BioChrom) IgD-положительные клетки высвобождали из шариков путем добавления 8 мкл буфера для высвобождения (DNase) и переносили в чистые пробирки. Этим способом смогли получить от 0,9 ×10 5 до 3,7 ×10 6 IgD-положительных клеток. Общую РНК выделяли из IgD-положительных клеток с использованием RNeasy ® Midi Kit (QIAGEN), следуя инструкциям производителя. кДНК синтезировали согласно стандартным способам (Sambrook, Cold Spring Harbor Laboratory Press 1989, Second Edition). Пример 2.2.2. ПЦР-амплификация вариабельных областей легкой цепи (VL-областей). Для выделения ДНК V-области легкой цепи осуществляли ОТ-ПЦР с использованием наборов праймеров. РНК из IgD-положительных В-клеток транскрибировали в кДНК (как описано выше) и использовали в качестве матричной ДНК в ПЦР-реакциях. В каждой ПЦР-реакции один 5'-праймер объединяли с одним 3'-праймером. Число разных ПЦР-реакций определялось числом возможных комбинаций 5'- и 3'-праймеров. Для амплификации использовали следующую ПЦР-программу: денатурацию при 94 °С в течение 15 с, отжиг праймеров при 52 °С в течение 50 с и удлинение праймеров при 72 °С в течение 90 с осуществляли в течение 40 раундов с последующим конечным удлинением при 72 °С в течение 10 мин. Затем выделяли ДНК V-фрагментов легкой цепи согласно стандартным протоколам. Пример 2.2.3. Конструирование библиотеки - клонирование пула VL человека. Библиотеку фагового дисплея обычно конструировали на основе стандартных методик, как раскрыто, например, в "Phage Display: A Laboratory Manual"; Ed. Barbas, Burton, Scott & Silverman; Cold Spring Harbor Laboratory Press, 2001. Праймеры, выбранные для ПЦР-амплификации, приводили к образованию сайтов распознавания 5'-SacI и 3'-SpeI для V-фрагментов легкой цепи. Ставили две реакции лигирования, причем каждая из них включала 400 нг фрагментов каппа-легкой цепи (расщепленных SacI-SpeI) и 1400 нг плазмиды pBluescript KS+ (расщепленной SacI-SpeI; большой фрагмент). Каждый из двух образующихся пулов V-легкой цепи антитела затем трансформировали в 300 мкл электрокомпетентных клеток Escherichia coli XL1 Blue путем электропорации (2,5 кВ, просвет кюветы 0,2 см, 25 мФ, 200 Ом, прибор Gene-Pulser Biorad), что приводило к получению библиотеки размером 5,8 ×10 8 независимых клонов. ДНК-фрагменты каппа (легкая цепь) из разных ПЦР-амплификаций взвешивали для каждого лигирования следующим образом: каждый 5'-праймер определяет специфическую группу. В этих группах 3'-праймеры определяют подгруппы. Данные подгруппы взвешивали 1:2:1:1, что соответствует праймерам 3'-hu-Vk-J1-SpeI-BsiWI:3'-hu-Vk-J2/4-SpeI-BsiWI:3'-hu-Vk-J3-SpeI-BsiWI:3'-hu-Vk-J5-SpeI-BsiWI. Данные группы взвешивали согласно их распределению в зародышевой линии 1:1:1:0,2:0,2, что соответствует праймерам 5'-huVK1-Sac-2001:5'-huVK3-Sac-2001:5'-huVK2/4-Sac-2001:5'-huVK5-Sac-2001:5'-huVK6-Sac-2001. После электропорации данный образец инкубировали в бульоне SOC (Fluka) для экспрессии фенотипа. Каждую культуру затем инкубировали в 500 мл селективной среды SB, содержащей 50 мкг/мл карбенициллина и 2% мас./об., глюкозы, в течение ночи. На следующие сутки клетки собирали центрифугированием и получение плазмиды осуществляли с использованием имеющегося в продаже набора для получения плазмид (Qiagen). Пример 2.2.4. Конструирование библиотеки антител: VL человека - материнская VH. ПЦР осуществляли для амплификации материнской VH из вектора, содержащего материнскую VH, описанную выше в примере 2.1. Для амплификации следовали протоколу ПЦР согласно стандартным методикам с использованием 5'-праймера MVH8 (5'-GAG GTT CAG CTC GAG CAG TCT GGA GCT-3') и 3'-праймера 3'-MuVHBstEII (5'-TGA GGA GAC GGT GAC CGT GGT CCC TTG GCC CCA G-3'). После очистки продукта амплификации, имеющего размер приблизительно в 350 п.н., из аналитического агарозного геля, фрагмент ДНК вырезали рестрикционными ферментами BstEII и XhoI. Фагмиду pComb3H5BHis (этот вектор описан в диссертации Dr. Ralf Lutterb üse) расщепляли соответствующим образом и большой фрагмент лигировали с вышеупомянутым фрагментом. После трансформации в E.coli XL1 blue одиночный клон культивировали в 100 мл среды SB (содержащей 50 мкг/мл карбенициллина) и плазмиду получали согласно стандартным протоколам. Успешное клонирование подтверждали секвенированием вставки (Sequiserve, Мюнхен). Этот вектор pComb3H5BHis/материнская VH разрезали рестрикционными ферментами SacI и SpeI. Выделяли большой фрагмент вектора. Плазмидную ДНК, содержащую VK-библиотеку из примера 2.2.3, вырезали рестрикционными ферментами SacI и SpeI. Выделяли полосу малого фрагмента VK (приблизительно 350 п.н.). 1200 нг фрагмента данного вектора лигировали с 400 нг фрагментов VK и трансформировали в 300 мкл электрокомпетентных клеток Е.coli XL1 Blue путем электропорации (2,5 кВ, просвет кюветы 0,2 см, 25 мФ, 200 Ом), что приводило к получению общей библиотеки scFv размером 2,8 ×10 8 независимых клонов. После экспрессии фенотипа и медленной адаптации к карбенициллину, библиотеку антител переносили в селективную среду с SB-карбенициллином (50 мкг/мл). Библиотеку антител затем инфицировали инфекционной дозой 1 ×10 12 частиц хелперного фага VCSM13, что приводило к продуцированию и секреции нитевидного фага М13, где каждая фаговая частица содержала одноцепочечную pComb3H5BHis-ДНК, кодирующую наполовину человеческий scFv-фрагмент, и экспонировала соответствующий scFv белок в виде трансляционного слияния с оболочечным белком III фага. Пример 2.2.5. Селекция VL человека с помощью фагового дисплея. Фаговые частицы, несущие репертуар scFv, собирали из культурального супернатанта осаждением с помощью PEG8000/NaCl и центрифугированием. Затем приблизительно от 1 ×10 11 до 1 ×10 12 scFv фаговых частиц ресуспендировали в 0,4 мл PBS/0,1% БСА и инкубировали с рекомбинантным биотинилированным растворимым rhGM-CSF (продуцируемым в Е.coli, как описано выше в примере 1) в течение 2 ч при слабом перемешивании в общем объеме 0,5 мл (концентрации антигена: раунды 1-3: 100 нм; раунд 4: 10 нм; раунд 5: 1 нМ). Затем добавляли 6,7 ×10 7 стрептавидиновых магнитных шариков (Dynabeads M-280-Streptavidin, Dynal) и осуществляли дополнительную инкубацию при слабом перемешивании в течение 30 мин. scFv фаг, который специфически не связывался с антигеном-мишенью, удаляли на стадиях промывки с помощью PBS/0,1% БСА. Для этой цели собирали магнитом комплексы биотинилированный антиген стрептавидиновый шарик (с потенциальными связывателями scFv) и ресуспендировали в 1 мл раствора для промывки (одна стадия промывки). Эту процедуру промывки повторяли вплоть до четырех раз в дополнительных раундах. После промывки связывающие структурные единицы элюировали с использованием HCl-глицина, pH 2,2. После нейтрализации с помощью 2 М Трис, pH 12, данный элюат использовали для инфицирования свежей неинфицированной культуры Е.coli XL1 Blue. Для элюции оставшихся структурных единиц с сильным связыванием эту стадию повторяли с использованием HCl-глицина, pH 1,0. Этот второй элюат вновь нейтрализовали и использовали для инфицирования свежей неинфицированной культуры Е.coli XL1 Blue. Обе инфицированные культуры Е.coli затем смешивали и клетки, которые были успешно трансдуцированы фагмидной копией, кодирующей фрагмент человеческого scFv, вновь подвергали селекции на устойчивость к карбенициллину и затем инфицировали хелперным фагом VCSM13 для начала второго раунда дисплея антител и селекции in vitro. Из культур Е.coli выделяли плазмидную ДНК, соответствующую 4 и 5 раундам "пэннинга". Для продукции растворимого белка scFv VL-ДНК фрагменты вырезали из плазмид (SacI-SpeI) и клонировали через те же сайты рестрикции в плазмиде pComb3H5BFlag/His с материнской VH, отличающейся от исходной pComb3H5BHis/материнской VH тем, что экспрессируемая конструкция (например, scFv) включает Flag-метку (TGDYKDDDDK) между scFv и His6-меткой и дополнительные фаговые белки делетированы. После лигирования каждый пул (разные раунды "пэннинга") плазмидной ДНК трансформировали в 100 мкл компетентных к тепловому шоку клеток E.coli XL1 Blue и переносили в чашки на LB-агар с карбенициллином. Одиночные колонии собирали в 100 мкл LB-карбенициллина (50 мкг/мл). 10 мкл этой клеточной суспензии типично инкубировали в 5 мл среды SB, дополненной карбенициллином до концентрации 50 мкг/мл и MgCl 2 до конечной концентрации 20 мм, в течение приблизительно 6 ч при 37 °С при перемешивании. Затем добавляли IPTG до конечной концентрации 1 мМ и продолжали инкубацию в течение ночи на шейкере при 30 °С. Клетки центрифугировали до осадка и этот осадок типично ресуспендировали в 0,5 мл PBS. С помощью четырех раундов замораживания при -70 °С и оттаивания при 37 °С наружную мембрану бактерий разрушали осмотическим шоком и растворимые периплазматические белки, включающие scFv, высвобождали в супернатант. После удаления интактных клеток и клеточного дебриса дополнительным центрифугированием (5 мин при 10000 ×g), супернатант (т.е. РРР), содержащий scFv, собирали и далее исследовали. RhGM-CSF антиген (Leukine Liquid, Immunex) иммобилизовали на планшетах для ELISA в течение ночи при 4 °С (50 мкл 1 мкг антигена/мл PBS на лунку). После промывки лунок один раз PBS и блокирования с помощью 3% БСА в PBS в течение 1 ч при комнатной температуре, в лунки добавляли 100 мкл РРР, содержащего scFv, и типично инкубировали в течение 1 ч при комнатной температуре. После трех промывок PBS/0,05% Tween 20, определение scFv-фрагментов, связанных с иммобилизованным антигеном, осуществляли с использованием М2 против flag (1 мкг/мл PBS/1% БСА) и определяли с помощью специфического поликлонального антитела козы против Fab2 мыши, конъюгированного с пероксидазой хрена (Dianova, 1 мкг/мл PBS/1% БСА). Сигнал проявляли путем добавления раствора субстрата 2,2'-азиноди[3-этилбензтиазолин-6-сульфоновой кислоты] ("ABTS") и определяли при длине волны 405 нм согласно стандартным протоколам. Из 20 протестированных клонов (10, полученных после 4 раундов, и 10, полученных после 5 раундов "пэннинга") 5 лизатов показывали сильные сигналы в ELISA на рекомбинантном антигене по сравнению с PBS в качестве отрицательного контроля. Результаты ELISA показаны на фиг. 1, на которой разные протестированные молекулы scFv расположены вдоль оси X, а ось Y показывает интенсивность измеренного поглощения, причем более сильное поглощение указывает на более прочное связывание. Отрицательный контроль с PBS показан на оси X слева. Молекулы scFv, демонстрирующие существенное связывание, отмечены над соответствующим столбиком, показывающим интенсивность поглощения, либо ромбиком, либо звездочкой. Ромбик или звездочка на фиг. 1 представляют собой две разные последовательности, т.е. scFv, столбик интенсивности поглощения которого показан ромбиком, представлял собой одну последовательность, тогда как все scFv, столбики интенсивности поглощения которых показаны звездочками, имеют одинаковую общую последовательность. Пять ELISA-положительных клонов подвергали секвенированию ДНК. Секвенирование осуществляли в Sequiserve (Мюнхен). Всего четыре клона имели последовательность ДНК, соответствующую scFv 5-306, тогда как другая последовательность (4-301) была идентифицирована только один раз. Доминантная последовательность, соответствующая scFv 5-306, а также последовательность 4-301 имели человеческое происхождение и демонстрировали очень близкую гомологию с последовательностью Vk1-O12 зародышевой линии человека. Пример 2.2.6. Характеристика удачных конструкций scFv, полученных в результате селекции huVL. Целью следующих экспериментов была характеристика удачных scFv, полученных вышеописанными способами. Пример 2.2.6.1. Экспрессия и очистка в малом масштабе удачных scFv (полученных, как описано выше) в Е.coli. Чтобы получить РРР, клетки выращивали в среде SB, дополненной 20 мМ MgCl 2 и 50 мкг/мл карбенициллина, и повторно растворяли в 1 мл PBS после сбора. Наружную мембрану бактерий разрушали температурным шоком (четыре раунда замораживания при -70 °С и оттаивания при 37 °С) и растворимые периплазматические белки, включающие scFv, высвобождались в супернатант. После удаления интактных клеток и клеточного дебриса центрифугированием супернатанты, содержащие scFv, собирали и исследовали далее. Для дальнейшей очистки к соответствующему РРР добавляли 25 мкл 20 мМ NaH 2 PO 4 , 400 мМ NaCl, 250 мМ имидазола, pH 7,0. РРР очищали с помощью колонок Ni-NTA Spin (Qiagen), как рекомендовано в руководстве по применению. Кратко, соответствующий раствор РРР добавляли к предварительно уравновешенной колонке для связывания со смолой. Колонки Spin дважды промывали 20 мМ NaH 2 PO 4 , 400 мМ NaCl, 20 мМ имидазола, pH 7,0. Связанный белок дважды элюировали в 200 мкл 20 мМ NaH 2 PO 4 , 400 мМ NaCl, 250 мМ имидазола, pH 7,0. Очищенные белки scFv дополнительно анализировали в отношении силы связывания (кинетика скорости диссоциации) и способностей к нейтрализации (ингибирование GM-CSF-зависимой пролиферации TF-1), как описано в следующих примерах. Несмотря на отсутствие разделения или различия между разными возможными конформациями scFv, эта грубая очистка РРР дает белок scFv 80% чистоты, как оценено вестерн-блот-анализом (данные не показаны). Пример 2.2.6.2. Ингибирование FITC-меченого rhGM-CSF. Целью этого эксперимента является демонстрация того, что идентифицированные клоны scFv способны ингибировать связывание rhGM-CSF с рецепторным комплексом GM-CSF, экспонированным на поверхности клеток TF-1. Ожидалось, что нейтрализующие конструкции scFv будут конкурировать за эпитоп связывания с рецептором на молекуле rhGM-CSF, делая невозможным связывание rhGM-CSF с рецепторным комплексом GM-CSF. В той степени, в которой связывание rhGM-CSF с его рецептором ингибируется описанным выше способом, ожидается, что будет наблюдаться снижение интенсивности флуоресцентного окрашивания клеток TF-1 флуоресцеин-меченым rhGM-CSF (rhGM-CSF-FITC) в анализе на основе проточной цитометрии. Следующее описывает эффективность такого анализа, основанного на проточной цитометрии. Конечную концентрацию 0,4 мкг/мл конъюгата rhGM-CSF-FITC в PBS инкубировали с 10 мкг/мл материнского антитела или неразведенного периплазматического экстракта scFv, который очищали на колонке NiNTA Spin. Образцы белка оставляли уравновешиваться при 25 °С в течение 1 ч перед добавлением клеточной суспензии TF-1. Клетки TF-1 культивировали в среде RPMI 1640 (Gibco; не содержит L-глутамин, феноловый красный), 10% инактивированной нагреванием FCS, в отсутствие rhGM-CSF в течение ночи. Использовали конечную концентрацию 2 ×10 6 клеток/мл и 150 мкл клеточной суспензии на образец. Клетки собирали центрифугированием при 500 ×g при 4 °С в течение 3 мин и дважды промывали буфером FACS. Промытые клетки ресуспендировали в 100 мкл предварительно уравновешенного образца белка, содержащего rhGM-CSF-FITC и соответствующее материнское антитело или scFv. Образцы инкубировали при 4 °С в течение 60 мин. После двух дополнительных промывок клетки ресуспендировали в 150 мкл ледяного буфера FACS и затем анализировали с помощью проточной цитометрии. Результаты показаны на фиг. 2. Конкретно, на фиг. 2 показан график, на котором разные тестируемые молекулы расположены вдоль оси X и на котором средняя интенсивность флуоресценции ("MFI") показана на оси Y. Как можно видеть на фиг. 2, с материнским антителом наблюдали очевидную потерю интенсивности флуоресценции клеток TF-1 (второй слева вдоль оси X). Конкурентное связывание молекулы scFv, обозначенной 5-306, с rhGM-CSF можно было отслеживать по потере флуоресцентного окрашивания клеток TF-1, тогда как молекула scFv 4-301 едва показала какой-либо эффект. Поскольку эти результаты говорят о том, что молекула scFv, обозначенная 5-306, может быть многообещающим нейтрализатором GM-CSF, дальнейший анализ нейтрализующей активности был ограничен scFv 5-306. Пример 2.2.6.3. Крупномасштабная экспрессия и очистка потенциального scFv. Крупномасштабное продуцирование и очистку белка осуществляли следующим образом. Кратко, Е.coli BL21DE3 трансформировали экспрессирующей плазмидой и выращивали при 37 °С в 1 л селективной среды до оптической плотности 0,5-0,8 при 600 нм. Продуцирование белка индуцировали путем добавления IPTG до концентрации 1 мМ и культуры инкубировали в течение еще 16 ч с перемешиванием при температуре 25 °С. Клетки собирали центрифугированием при 5000 ×g в течение 10 мин и ресуспендировали в 100 мл 1 ×PBS. Периплазматические белки экстрагировали оптимальным последовательным замораживанием в этаноле/сухом льду и оттаиванием в водяной бане при 37 °С в течение четырех раундов. Наконец, данный экстракт центрифугировали при 10000 ×g в течение 20 мин. scFv 5-306 выделяли в процессе двухступенчатой очистки, включающем аффинную хроматографию с иммобилизованным металлом (IMAC) и гель-фильтрацию. Все потенциальные (scFv) очищали согласно этому способу. Для хроматографии использовали систему Äkta ® FPLC (Pharmacia) и программное обеспечение Unicorn ®. Все реагенты имели аналитическую степень чистоты и были приобретены у Sigma (Deisenhofen) или Merck (Darmstadt). IMAC осуществляли с использованием колонки Qiagen Ni-NTA Superflow согласно протоколу производителя. Колонку уравновешивали буфером А2 (20 мМ фосфата натрия, pH 7,2, 0,4 М NaCl) и РРР (100 мл) наносили на колонку (2 мл) при скорости протока 2 мл/мин. Колонку промывали 5 объемами колонки 5%-ного буфера В2 (20 мМ фосфата натрия, pH 7,2, 0,4 М NaCl, 0,5 М имидазола) для удаления несвязавшегося образца. Связанный белок элюировали, используя 100%-ный буфер В2 в 5 объемах колонки. Фракции элюированного белка объединяли для дальнейшей очистки. Гель-фильтрацию осуществляли на колонке HiLoadTM 16/60 Superdex 75 Prep Grade (Pharmacia), уравновешенной PBS (Gibco). Образцы элюированного белка (скорость протока 1 мл/мин) подвергали стандартному ПААГ-ДСН и вестерн-блоттингу для определения. Перед очисткой колонку калибровали для определения молекулярной массы (набор маркеров молекулярной массы, Sigma MW GF-200). Зависимое от размера разделение на колонке Superdex 75 Prep Grade приводило к получению четко различимых пиков фракций мономера и ассоциативного димера потенциальных scFv. Концентрации белка определяли, измеряя оптическую плотность при 280 нм, и рассчитывали, используя коэффициент молекулярной экстинкции, специфической для последовательности, соответствующего потенциального scFv. Пример 2.2.6.4. Ингибирование rhGM-CSF-зависимой пролиферации клеток TF-1 потенциальным scFv. Целью этого эксперимента является получение количественной информации относительно нейтрализующей активности наполовину человеческого scFv 5-306 с использованием rhGM-CSF-зависимой клеточной линии TF-1 (DSMZ АСС 334). Клетки TF-1 культивировали в среде RPMI 1640 (Gibco; не содержит L-глутамин, феноловый красный), 10% инактивированной нагреванием FCS, в присутствии 2,5 нг/мл rhGM-CSF, как описано дистрибьютером (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Германия). Клетки выращивали до плотности клеток 0,5 ×10 6 клеток/мл. Для анализа пролиферации клетки TF-1 собирали центрифугированием при 300 ×g в течение 4 мин и промывали 1 ×PBS (Dulbecco's, Gibco). Клетки ресуспендировали в конечной концентрации 1 ×10 5 клеток/мл в RPMI 1640, 10% FCS и использовали 90 мкл клеточной суспензии на лунку плоскодонного планшета для клеточных культур Microtest (0,9 ×10 4 клеток/лунку). Для стимуляции пролиферации клеток TF-1 использовали конечную концентрацию rhGM-CSF 0,3 нг/мл. Для нейтрализации hGM-CSF-зависимой пролиферации добавляли 10 мкл очищенного scFv к 100 мкл TF-1 и раствору rhGM-CSF в серии разведений, варьирующих от приблизительно 100 мкг/мл до 100 пг/мл. Образцы инкубировали при 37 °С в 5% СО 2 в течение 72 ч. Через 72 ч определяли пролиферативный статус клеток TF-1, добавляя WST-1 и наблюдая за колориметрическим изменением с помощью ELISA-ридера при 450 нм. Данные аппроксимировали для полумаксимального ингибирования пролиферации (IC 50 ), используя построение нелинейной кривой регрессии с помощью программы Prism. Можно было наблюдать очевидный дозозависимый эффект scFv 5-306 в отношении ингибирования пролиферации, и он был сравнимым для мономерной и димерной конформационных форм. Путем аппроксимации полумакисмального ингибирования пролиферации определили значение IC 50 7,3 нМ для мономерной формы и 3,5 нМ - для димерной формы. Результаты показаны на фиг. 3. Пример 2.3. Конструирование библиотек антител и селекция человеческих VH с помощью фагового дисплея. Целью следующих экспериментов является селекция набора человеческих VH-областей, которые образовывали бы пары с человеческой VL-областью scFv 5-605, выбранного, как описано выше. Пример 2.3.1. Выделение РНК из мононуклеарных клеток периферической крови (РВМС). От пяти здоровых доноров брали по 100 мл крови. Мононуклеарные клетки периферической крови (РВМС) выделяли с помощью градиента фиколла согласно стандартным способам. Общую РНК выделяли из РВМС, используя RNeasy ® Midi Kit (QIAGEN), следуя инструкциям производителя. кДНК синтезировали согласно стандартным способам (Sambrook, Cold Spring Harbor Laboratory Press 1989, Second Edition). Пример 2.3.2. ПЦР-амплификация вариабельных областей тяжелых цепей (VH-областей). Конструировали библиотеку VH и называли ее Lib 134-VH. Эта библиотека VH состоит из репертуара FR1-CDR2-FR2-CDR2-FR3 человека из ПЦР-амплифицированных VH-областей описанного выше пула РВМС, связанных функциональным образом с VH CDR3 материнского антитела, после которой следует человеческая последовательность FR4 зародышевой линии. Для выделения человеческих матричных VH-областей осуществляли ОТ-ПЦР с использованием набора 5'-VH-специфических праймеров и набора из двух 3'-VH-специфических праймеров Для каждой ПЦР-реакции один 5'-праймер объединяли с одним 3'-праймером, причем число разных ПЦР-реакций определялось числом возможных комбинаций 5'- и 3'-праймеров. В качестве источника VH-генов использовали только кДНК РВМС (как описано выше) четырех доноров. Для амплификации использовали следующую программу ПЦР: денатурацию при 94 °С в течение 15 с, отжиг праймеров при 52 °С в течение 50 с и удлинение праймеров при 72 °С в течение 60 с осуществляли в течение 40 раундов с последующим конечным удлинением при 72 °С в течение 10 мин. Выделяли продукты амплификации с размером приблизительно 350 п.н. согласно стандартным способам. Для выделения Lib 134-VH-областей осуществляли ОТ-ПЦР в две стадии. Во-первых, человеческие VH-сегменты тяжелой цепи (FR1-CDR1-FR2-CDR2-FR3) амплифицировали с помощью ПЦР из выделенных матричных VH-фрагментов, используя такой же набор 5'-VH-специфических праймеров, как описано выше и набор 3'-специфических праймеров для человеческих VH-подсемейств 1, 3 и 4, совместимых в самой концевой области FR3. Использовали следующие комбинации праймеров: Для каждой ПЦР-реакции один 5'-праймер объединяли с 3'-праймером; число разных ПЦР-реакций определялось числом возможных комбинаций 5'- и 3'-праймеров. Для амплификации использовали следующую программу ПЦР: денатурацию при 94 °С в течение 15 с, отжиг праймеров при 52 °С в течение 50 с и удлинение праймеров при 72 °С в течение 90 с осуществляли в течение 40 раундов с последующим конечным удлинением при 72 °С в течение 10 мин. С помощью этой стадии ПЦР и соответствующей последовательности 3'-праймера человеческие VH-области удлиняли на часть материнской VH CDR3, которая в свою очередь представляет собой затравочный сайт для 3'-праймера второй стадии ПЦР. Эти VH-(FR1-CDR1-FR2-CDR2-FR3) ДНК-фрагменты затем использовали в качестве матрицы во второй ПЦР-реакции, снова используя соответствующий 5'VH-специфический праймер и универсальный 3'-праймер, соответствующий универсальному 3'-концу амплифицированных фрагментов ДНК Для амплификации использовали следующую программу ПЦР: денатурацию при 94 °С в течение 15 с, отжиг праймеров при 52 °С в течение 50 с и удлинение праймеров при 72 °С в течение 60 с осуществляли в течение 40 раундов с последующим конечным удлинением при 72 °С в течение 10 мин. V-фрагменты ДНК выделяли согласно стандартным протоколам. Пример 2.3.3. Конструирование библиотеки - клонирование пула VH человека. Во втором раунде вышеуказанного способа выбирали человеческую VL scFv 5-306, идентифицированного в первой, предыдущей процедуре селекции, и затем объединяли с библиотекой человеческих VH-фрагментов, описанных в примере 2.3.2, с целью получения человеческого scFv. Библиотеку фагового дисплея обычно конструировали на основе стандартных методик, как, например, раскрыто в "Phage Display: A Laboratory Manual"; Ed. Barbas, Burton, Scott & Silverman; Cold Spring Harbor laboratory Press, 2001. ДНК-фрагменты тяжелой цепи из разных ПЦР-амплификаций взвешивали для каждого лигирования следующим образом: а:б:в:г:д:е=3:1:3:1:1:1, где а-е имеют следующие значения: Проводили одну реакцию лигирования, включающую 400 нг пула человеческих Lib 134-VH фрагментов (расщепленных XhoI- BstEII) и 1200 нг плазмиды pComb3H5BHis/5-306 VL (ДНК, кодирующую VL-область scFv 5-306, клонировали через сайты рестрикции SacI и SpeI в pComb3H5BHis согласно стандартным методикам). Полученный пул VH человеческих антител затем трансформировали в 300 мкл электрокомпетентных клеток Escherichia coli XL1 Blue путем электропорации (2,5 кВ, просвет кюветы 0,2 см, 25 мФ, 200 Ом, прибор Gene-Pulser Biorad), что приводило в целом к размеру библиотеки 1,6 ×10 8 независимых клонов. После электропорации реакционную среду инкубировали в бульоне SOC (Fluka) для экспрессии фенотипа. Затем каждую культуру инкубировали в 500 мл селективной среды SB, содержащей 50 мкг/мл карбенициллина и 2% об./об. глюкозы, в течение ночи. На следующие сутки клетки из культур собирали центрифугированием и осуществляли получение плазмид с использованием имеющегося в продаже набора для получения плазмид (Qiagen) для сохранения библиотеки ДНК. 1,5 мкг этого плазмидного пула, кодирующего соответствующий пул scFv, затем вводили путем электропорации в Е.coli XL1 Blue (2,5 кВ, просвет кюветы 0,2 см, 25 мФ, 200 Ом, прибор Gene-Pulser Biorad), что приводило в целом к размеру библиотеки 2,4 ×10 9 независимых клонов. После экспрессии фенотипа и медленной адаптации к карбенициллину библиотеку антител переносили в селективную среду SB-карбенициллин (50 мкг/мл). Данную библиотеку антител затем инфицировали инфекционной дозой 1 ×10 12 частиц хелперного фага VCSM13, что приводило к продуцированию и секреции нитевидного фага М13, где каждая частица фага содержала одноцепочечную pComb3H5BHis-ДНК, кодирующую человеческий scFv-фрагмент, и экспонировала соответствующий белок scFv в виде трансляционного слияния с оболочечным белком III фага. Пример 2.3.4. Селекция VH человека с помощью фагового дисплея. Полученную фаговую библиотеку, несущую клонированный репертуар scFv, собирали из культурального супернатанта путем осаждения с помощью PEG8000/NaCl и центрифугирования. Приблизительно от 1 ×10 11 до 1 ×10 12 фаговых частиц с scFv ресуспендировали в 0,4 мл PBS/0,1% БСА и инкубировали с рекомбинантным биотинилированным растворимым rhGM-CSF (материал Е.coli, как описано в примере 1) в течение 1 ч при слабом перемешивании в общем объеме 0,5 мл. Затем добавляли 6,7 ×10 7 магнитных шариков, покрытых стрептавидином (Dynabeads M-280-Streptavidin, Dynal), и инкубировали далее при слабом перемешивании в течение 30 мин. scFv фаг, который специфически не связывается с антигеном-мишенью, устраняли стадиями промывки с PBS/0,1% БСА. Для этой цели комплексы биотинилированный антиген - стрептавидиновый шарик (с потенциальными связывателями scFv) собирали магнитом и ресуспендировали в 1 мл раствора для промывки (одна стадия промывки). Эту процедуру промывки повторяли вплоть до четырех раз. После промывки связывающие структурные единицы элюировали с использованием HCl-глицина, pH 2,2, и после нейтрализации с использованием 2М Трис, pH 12, элюат использовали для инфицирования свежей неинфицированной культуры Е.coli XL1 Blue. Для элюции оставшихся сильно связывающих структурных единиц шарики непосредственно ресуспендировали в 200 мкл свежей культуры Е.coli XL1 Blue (ОП 600 ≥0,5) и инкубировали в течение 10 мин при слабом перемешивании. Затем смешивали обе культуры, и клетки, успешно трансдуцированные фагмидной копией, кодирующей scFv-фрагмент человека, вновь подвергали селекции на устойчивость к карбенициллину и затем инфицировали хелперным фагом VCMS13 для начала второго раунда дисплея антител и селекции in vitro. Для двух антител осуществляли в целом 4 раунда селекции. Концентрации антигена во время селекции снижали до конечных концентраций следующим образом: Выделяли плазмидную ДНК из культур Е.coli, соответствующих 3 или 4 раунду "пэннинга". Для получения растворимого scFv-белка фрагменты VH-VL-ДНК вырезали из плазмид (XhoI-SpeI) и клонировали через те же сайты рестрикции в плазмиде pComb3H5BFlag/His (без дополнительных фаговых белков, необходимых для фаговой инфекции). После лигирования каждый пул (разные раунды "пэннинга") плазмидной ДНК трансформировали в 100 мкл компетентных к тепловому шоку клеток Е.coli TG1 и переносили на чашки с карбенициллином и LB-агаром. Одиночные колонии отбирали и инокулировали в 120 мкл LB-карбенициллина (50 мкг/мл), 1% глюкозы в 96-луночных планшетах (Greiner). Лунки запечатывали полупроницаемой мембраной (Greiner), и планшеты инкубировали в течение ночи при 37 °С во встряхиваемом инкубаторе (основной планшет). Затем 10 мкл культур из основных планшетов переносили во второй 96-луночный планшет (рабочий планшет), содержащий 90 мкл LB-карбенициллина (50 мкг/мл), 0,1% глюкозы на лунку. После инкубации в течение 4 ч при 37 °С во встряхиваемом инкубаторе продуцирование scFv индуцировали путем добавления 20 мкл LB-карбенициллина, 6 мМ IPTG в каждую лунку. После другой стадии инкубации в течение ночи при 30 °С со встряхиванием, клетки лизировали при инкубации в течение 1 ч при комнатной температуре с помощью 40 мкл лизирующего буфера (400 мМ борной кислоты, 320 мМ NaCl, 4 мМ EDTA pH 8, 2,5 мг/мл лизоцима). Оставшиеся клетки и клеточный дебрис отделяли центрифугированием в течение 12 мин при 1900 ×g (Hettich). Супернатанты, содержащие молекулы scFv, затем тестировали на связывание в ELISA-анализах. Определение scFv-фрагментов, связанных с иммобилизованным антигеном rhGM-CSF (Лейкин), осуществляли с использованием анти-flag М2 (1 мкг/мл PBS/1% БСА), которые определяли специфическим поликлональным антителом козы против Fab2 мыши, конъюгированным с пероксидазой хрена (Dianova, 1 мкг/мл PBS/1% БСА). Сигнал проявляли путем добавления раствора субстрата ABTS и определяли при длине волны 405 нм. Из приблизительно 100 клонов, протестированных после третьего раунда селекции, 12 клонов показали сильное связывание с rhGM-CSF. Из приблизительно 160 клонов, протестированных после четвертого раунда, свыше 80% лизатов показало сильные сигналы в ELISA на рекомбинантном антигене по сравнению с PBS в качестве отрицательного контроля. Результаты от репрезентативных клонов показаны на фиг. 4, на которой эти репрезентативные клоны расположены вдоль оси X, а интенсивность поглощения указана по оси Y. Как можно видеть на фиг. 4, отрицательный контроль с PBS (второй справа по оси X) не показал существенного связывания, тогда как репрезентативные клоны scFv: scFv A, scFv 3035, scFv 3039, scFv 3080 и scFv 5-306 показали разные уровни силы связывания, определенные с помощью ELISA. Все лизаты тестировали без rhGM-CSF в параллельных экспериментах на неспецифическое связывание с блокирующим агентом. Никакого значимого определимого сигнала не наблюдали, что указывает на специфичность связывания с rhGM-CSF. Определяли последовательности ДНК более чем 13 ELISA-положительных клонов scFv. Всего идентифицировали шесть разных последовательностей. Все последовательности имели человеческое происхождение и были близкородственными с человеческой последовательностью зародышевой линии VH-11-02. Пример 2.3.5. Характеристика человеческих конструкций scFv, содержащих человеческие VL- и VH-области. Пример 2.3.5.1. Крупномасштабное продуцирование и очистка потенциальных конструкций scFv, полученных способом, описанным в примере 3. Потенциальные scFv выделяли и очищали, как описано в примере 2.2.6.3. Пример 2.3.5.2. Кинетический анализ связывания потенциальных scFv с помощью поверхностного плазмонного резонанса (SPR). Целью данного эксперимента является глубокая характеристика потенциальных scFv. Кинетику связывания (kd и ka) потенциальных scFv измеряли, инъецируя 10 мкл очищенного белка в серии разведений, варьирующих от 10 мкг/мл до 1 пг/мл очищенного scFv, и отслеживая диссоциацию при 25 °С в течение 100 с. Белок забуферивали в HBS-EP (0,01 М HEPES, pH 7,4, 0,15 М NaCl, 3 мМ EDTA, 0,005% поверхностно-активного вещества Р20). Данные аппроксимировали с использованием программы BIAevaluation ™, определяющей константу скорости для кинетики диссоциации и ассоциации с помощью уравнения связывания Ленгмюра 1:1 (формулы 1 и 2), где А представляет собой концентрацию инъецированного анализируемого вещества, и В представляет собой концентрацию лиганда. Кривые кинетики связывания определяли с использованием вплоть до 8 концентраций каждого проанализированного потенциального scFv. Независимая аппроксимация исходных данных дала константы скорости диссоциации и ассоциации, которые использовали для расчета равновесной константы диссоциации (KD, результаты показаны в табл. 1). Пример 2.3.5.3. Ингибирование rhGM-CSF-зависимой пролиферации клеток TF-1 потенциальными scFv. После подтверждения того, что у потенциальных scFv сохранялась сила специфического связывания, целью этого эксперимента был анализ специфичности взаимодействия потенциального scFv с антигеном rhGM-CSF. Ингибирование биологической функции rhGM-CSF антигена путем связывания scFv характеризовали в эксперименте по ингибированию пролиферации TF-1. Эксперименты по ингибированию пролиферации TF-1 осуществляли, как описано выше. Клетки ресуспендировали в конечной концентрации 1 ×10 5 клеток/мл в RPMI 1640, 10% FCS, и использовали 90 мкл клеточной суспензии на лунку (0,9 ×10 4 клеток/лунку). Для стимуляции пролиферации клеток TF-1 использовали конечную концентрацию rhGM-CSF 0,3 нг/мл. Для нейтрализации rhGM-CSF-зависимой пролиферации добавляли очищенный scFv в 1 ×PBS в серии разведений с конечными концентрациями белка, варьирующими от 100 мкг/мл до 10 пг/мл. 10 мкл диализованного и подвергнутого стерилизующей фильтрации белкового раствора (фильтр 0,22 мкм) добавляли к 100 мкл раствора TF-1 и rhGM-CSF. Образцы инкубировали при 37 °С в 5% СО 2 в течение 72 ч. Через 72 ч определяли пролиферативный статус клеток TF-1, добавляя WST-1 и отслеживая колориметрическое изменение с помощью ELISA-ридера при 450 нм (фиг. 5). Как можно видеть на фиг. 5, четко продемонстрирована активность, нейтрализующая GM-CSF человека. ScFv A продемонстрировал самую сильную нейтрализующую активность. Пример 2.4. Оптимизация характеристик связывания выбранных scFv. Предусматривалось, что биологическую активность нейтрализующего агента в отношении мономерного лиганда можно улучшить или даже оптимизировать путем увеличения силы связывания между нейтрализатором и лигандом, конкретно путем повышения скорости диссоциации нейтрализатора. Предпочтительно этого можно добиться мутированием последовательности соответствующей VH- или VL-области случайным образом путем (1) введения одной или более чем одной случайной мутации в пределах всей последовательности или путем (2) введения одиночных мутаций или множественных смежных мутаций (например, отрезков из пяти, шести, семи, восьми, девяти или десяти аминокислот) в области scFv, которые имеют высокую вероятность взаимодействия с антигеном. Соответствующие мутанты затем необходимо охарактеризовать в отношении любого увеличения активности или, перед характеристикой, их необходимо улучшить до предпочтительных качеств (например, более сильное связывание) с помощью подходящих способов селекции (например, фаговый дисплей). Пример 2.4.1. Увеличение аффинности мутированием VH CDR3 в одном или более чем одном положении. Для улучшения характеристик связывания фрагмента антитела, например молекулы scFv, с помощью одиночных точечных мутаций или коротких аминокислотных отрезков, необходимо сделать мишенями аминокислотные остатки, которые имеют очень высокую вероятность взаимодействия с соответствующим антигеном. При таком подходе нет необходимости подвергать скринингу более чем только ограниченное число мутантов, без снижения вероятности успеха. CDR3 тяжелой цепи антитела или его фрагмента обычно вносит существенный вклад в связывание антигена этим антителом или фрагментом антитела в целом. Поэтому предусматривалось, что многообещающим способом увеличения аффинности связывания антитела или фрагмента антитела может быть мутирование нуклеотидной последовательности, кодирующей VH CDR3. Существует целый ряд разных методологий для проведения такого направленного случайного мутагенеза, некоторые из которых описаны ниже в контексте того, как можно увеличить аффинность связывания молекул scFv, описанных выше. А) Для того чтобы сделать VH CDR3 мишенью, необходимо ввести в нуклеотидную последовательность в VH CDR3 подходящий сайт рестрикции, предпочтительно путем синтеза гена целой VH-области с модифицированной нуклеотидной последовательностью CDR3, сохраняя первоначальную аминокислотную последовательность (Entelechon, Германия). Посредством расщепления соответствующим ферментом рестрикции и добавления нуклеазы S1/ДНК-полимеразы I Кленова и dGTP с последующим добавлением мутантного олигомерного дуплекса можно провести направленный случайный мутагенез в одном или более чем одном положении аминокислот согласно Matteucci and Heyneker, Nucleic Acids Research 11: 3113 ff (1983). Мутировавшие VH затем объединяют с соответствующей VL (через подходящий линкер) в подходящем векторе для экспрессии scFv и трансформируют в клетки Е.coli. Одиночные колонии, экспрессирующие вариантные scFv, затем можно собрать и подвергнуть скринингу на улучшенные scFv, как описано для скрининга и характеристики удачных scFv и потенциальных scFv в предыдущих примерах. Б) Альтернативным способом является олигонуклеотид-опосредованный мутагенез с помощью способа с двойным праймером, как подробно описано у Sambrook, Fritsch, Maniatis (1989) "A laboratory manual". По существу, VH-область клонируют в вектор на основе М13 и выделяют одноцепочечную плазмиду. Праймер, способный к гибридизации с одноцепочечной плазмидной матрицей, содержащей рандомизированную последовательность, отжигают и удлиняют. После размножения соответствующего плазмидного пула в Е.coli мутировавшие VH можно собрать из пула плазмид и объединить с первоначальной VL (через подходящий линкер) в подходящем векторе для экспрессии scFv и трансформировать в клетки Е.coli. Одиночные колонии, экспрессирующие вариантные scFv, выбирают и подвергают скринингу на улучшенные scFv, как описано для скрининга и характеристики удачных scFv и потенциальных scFv в предыдущих примерах. В) Еще одной альтернативой является мутирование вплоть до шести или даже большего числа смежных аминокислот. Для этой цели можно сконструировать делеционный вариант нуклеотидной последовательности VH, имеющей делетированные CDR3 и FR4. Эту конструкцию используют в качестве матрицы для одно- или двухступенчатой ПЦР-амплификации, при которой подходящий 6'-праймер (гибридизующийся с 5'-концом последовательности VH и добавляющий подходящий сайт клонирования) объединяют с набором 3'-праймеров, которые гибридизуются на 3'-конце FR3-области в качестве матрицы и добавляют к амплифицируемому фрагменту CDR3 и FR4 область (с подходящим сайтом рестрикции). Этот набор 3'-праймеров содержит последовательность из одного или более чем одного триплета для вставки случайных кодонов в последовательность CDR3. Этот пул VH-областей, содержащих рандомизированные области CDR3, затем можно последовательно объединить с соответствующей VL (через подходящий линкер) в подходящем векторе для экспрессии scFv и трансформировать в клетки Е.coli. Одиночные колонии, экспрессирующие вариантные scFv, затем отбирают и подвергают скринингу на улучшенные scFv, как описано для скрининга и характеристики удачных scFv и потенциальных scFv в предыдущих примерах. Соответствующие пулы мутировавших scFv, которые имеют более высокое разнообразие (которые можно легко подвергнуть скринингу), можно клонировать в подходящий вектор для фагового дисплея, и улучшенные scFv затем можно подвергнуть селекции с помощью фагового дисплея на интересующем антигене, предпочтительно в условиях снижающихся концентраций антигена для выбора более высоких аффинностей. Селекции с помощью фагового дисплея проводят согласно стандартным протоколам, как описано здесь в другом месте. Любой из приведенных выше способов А-В можно объединять или проводить в повторяющихся циклах для дальнейшего улучшения и оптимизации уже модифицированных scFv. Пример 2.4.2. Увеличение аффинности путем случайного мутирования V-областей в пределах всей последовательности. Вместо мутирования специфических сайтов scFv, которые имеют высокую вероятность взаимодействия с соответствующим антигеном, можно осуществлять более прагматичный подход путем введения точечных мутаций в пределах всей последовательности VH и/или VL и затем проведения скрининга в отношении оптимизированных scFv или селекции и проведения скрининга оптимизированных scFv. В качестве примера, последовательность VH и/или VL можно подвергнуть мутагенезу с использованием штаммов-мутаторов Е.coli (как описано у Low et al. 260: 359 ff J. Mol. Biol. (1996)) или неправильного включения нуклеотидов ДНК-полимеразами, как подробно описано у Sambrook, Fritsch, Maniatis (1989) "A laboratory manual". Клонирование, экспрессию и селекцию оптимизированных вариантов scFv-молекул можно проводить с помощью фагового дисплея или часто используемой технологии рибосомного дисплея (как описано в ЕР 0975748А1). Оптимизированные версии экспрессируют в подходящих системах векторов/Е.coli для скрининга улучшенных кандидатов scFv. Подходящую методологию, как описано выше в примере 2.4, использовали для оптимизации репрезентативного потенциального scFv (scFv А), что приводило к получению класса фрагментов моноклональных человеческих нейтрализующих антител против GM-CSF, представленного молекулами scFv B-N. Характеристики этих молекул scFv будут выявлены и дополнительно описаны в следующих примерах. Получение молекул моноклонального IgG из выбранных молекул scFv описано в следующем примере. Пример 3. Клонирование и экспрессия эукариотами моноклональных антител из селектированных scFv. Хотя бактерии, как известно, экспрессируют функциональные Fab-фрагменты, они обычно не способны продуцировать полностью функциональные иммуноглобулины. Для продукции полностью функциональных антител необходимо использовать клетки млекопитающих, и поэтому VL-область scFv 5-306 и разные VH-области молекул scFv, селектированных в предыдущих примерах, субклонировали в экспрессирующие векторы млекопитающих (особенно VH-области scFv А и scFv В). Пример 3.1. Клонирование человеческой легкой цепи на основе scFv 5-306. Для получения подходящих терминальных сайтов рестрикции фрагмент ДНК, кодирующий VL-область scFv 5-306, повторно амплифицировали с помощью ПЦР, что давало V-каппа фрагменты с сайтом Bsu36I на 5'-конце и сайтом XhoI на 3'-конце. Этот фрагмент затем субклонировали в плазмиду BSPOLL Bsu36I и XhoI с использованием 5'-праймера (5'-ACGTCACCTTAGGTGTCCACTCCGATATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGC-3') и 3'-праймера (5'-CATGCACTCGAGCTTGGTCCCTCCGCCGAAAG-3'), добавляя, таким образом, лидерную последовательность млекопитающих и человеческую константную область Скаппа, и подтверждали секвенированием. Используя EcoRI и SalI, ДНК 5-306 VL-Скаппа вырезали из BSPOLL и субклонировали в эукариотический экспрессирующий вектор pEF-ADA, происходящий из экспрессирующего вектора pEF-DHFR (Mack et al. (1995) Proc. Natl. Acad. Sci. USA. 92, 7021-5), путем замещения кДНК, кодирующей мышиную дигидрофолатредуктазу (DHFR), на кДНК, кодирующую мышиную аденозиндезаминазу (ADA). Пример 3.2. Клонирование вариабельных доменов человеческой тяжелой цепи. Из разных человеческих VH-областей, селектированных в предыдущих примерах (особенно VH-областей scFv А и scFv В), вариабельную область повторно амплифицировали с помощью ПЦР, генерируя на обоих концах сайты рестрикции Bsu36I. Для всех конструкций использовали комбинацию двух праймеров: 5'-праймера VH-Bsu36I (5'-ACGTCACCTTAGGTGTCCACTCCCAGGTGCAGCTGGTCCAGTCTGGGGCTGAG GTGAAGAAGC-3') и 3'-праймера (5'-ACGTCACCTGAGGAGACGGTGACCATTGTCCCTTG-3'). Полученные фрагменты ДНК затем субклонировали с использованием этих сайтов рестрикции в эукариотический экспрессирующий вектор pEF-DHFR, уже содержащий эукариотическую лидерную последовательность и фрагмент ДНК, кодирующий константную область тяжелой цепи человеческого IgG1. Вариабельные области тяжелой цепи, таким образом, вставляли между лидерной последовательностью и константной областью тяжелой цепи. Правильные последовательности вариабельных областей подтверждали секвенированием. Пример 3.3. Превращение scFv-фрагментов в полные IqG человека. Плазмиду, кодирующую легкую цепь (VL 5-306/Скаппа), и плазмиду, кодирующую одну тяжелую цепь (VH/константная область человеческого IgG1), котрансфицировали в клетки НЕК согласно стандартным протоколам для временной экспрессии белка, и данные клетки культивировали для обеспечения экспрессии и продукции иммуноглобулинов в культуральную среду. Этим способом продуцировали IgG А, происходящий из scFv А, и IgG В, происходящий из scFv В. После соответствующего периода продукции собирали супернатанты и выделяли человеческие иммуноглобулины с помощью Протеин А-хроматографии согласно стандартным протоколам для очистки иммуноглобулинов. Очищенные иммуноглобулины затем использовали для дальнейших экспериментов по определению характеристик. Пример 3.4. Реконверсия IgG-специфичностей в scFv-фрагменты. VH-области из конструкций IgG (IgG А и IgG В, как описано выше) повторно клонировали в подходящий экспрессирующий вектор scFv согласно стандартным протоколам и функциональным образом связывали через гибкий линкер с VL-областью, происходящей из человеческой легкой цепи из примера 3.1. Эти конструкции продуцировались в растворимой форме в периплазме Е.coli, как описано выше. Характеристика этих scFv (scFv О, происходящего из IgG А, и scFv P, происходящего из IgG В) описана в следующих примерах. Пример 4. Оценка специфичности связывания человеческого моноклонального антитела против GM-CSF в отношении GM-CSF приматов и человека. Целью этого эксперимента была демонстрация того, что антитело, полученное, как представлено выше, специфически связывается с GM-CSF. Поэтому связывание такого антитела с разными рекомбинантными человеческими ("rh") колониестимулирующими факторами (rhG-CSF и rhM-CSF, Strathmann) сравнивали со связыванием того же антитела с rhGM-CSF с помощью ELISA. 50 мкл конкретного антигена (1 мкг/мл в PBS) наносили на планшет для ELISA (Nunc, Maxisorp) в течение 1 ч при комнатной температуре. После промывок 3 раза PBS/0,05% Tween 20 лунки блокировали 200 мкл PBS/3% обезжиренного сухого молока на лунку в течение 1,5 ч при комнатной температуре с последующей промывкой 3 раза PBS/0,05% Tween 20. 50 мкл/лунку серий человеческих антител (например, IgG А и IgG В), каждого с идентичными легкими цепями с последовательностью согласно SEQ ID NO: 34, но с разными тяжелыми цепями согласно SEQ ID NO: 35-48, добавляли в серии разведений, варьирующих от 1 мкг/мл до 0,5 нг/мл (в PBS/0,05% Tween 20/3% обезжиренного сухого молока), и инкубировали в течение 1 ч. После 3 промывок PBS/0,05% Tween 20, связанное антитело определяли с использованием 50 мкл антитела козы против человеческого IgG, конъюгированного с пероксидазой хрена (Dianova, разбавленное 1:1000 в PBS/0,05% Tween 20/3% обезжиренного сухого молока). Сигнал проявляли путем добавления 50 мкл/лунку раствора ABTS (Roche), и поглощение измеряли при 405 нм, используя длину волны 450 нм в качестве эталона. Имеющиеся в продаже антитела кролика (Strathmann Biotech AG), специфические, соответственно, к rhM-CSF и rhG-CSF, использовали в качестве положительных контролей для связывания этих антигенов, где указанное связывание определяли с использованием антител козы против антител кролика, конъюгированных с щелочной фосфатазой. Сигнал проявляли с помощью 50 мкл/лунку раствора pNpp (Sigma), и поглощение измеряли при 405 нм, используя длину волны 450 нм в качестве эталона. Результаты показаны на фиг. 6А, 6В и 6С для двух репрезентативных человеческих антител IgG А и IgG В. Как можно видеть на фиг. 6А, возрастающая концентрация титруемого антитела приводила к увеличению поглощения, что указывает на хорошее связывание обоих репрезентативных антител IgG А и IgG В с rhGM-CSF. На фиг. 6В показаны результаты связывания тех же двух репрезентативных антител с rhM-CSF. Как можно видеть на этой фигуре, увеличение концентраций кроличьего антитела против rhM-CSF приводило к увеличению поглощения, т.е. к увеличению связывания этого контрольного антитела (закрашенные кружки), тогда как поглощения для двух репрезентативных антител, описанных выше (закрашенные квадратики и закрашенные треугольники), накладывались друг на друга в виде продолжения исходного поглощения, которое не увеличивается при увеличении концентрации тестируемого антитела. Полностью аналогичный результат виден как для контрольного антитела, так и для репрезентативных тестируемых IgG А и В на фиг. 6С, на которой показаны результаты связывания с rhG-CSF. Вместе взятые, эти данные, представленные на фиг. 6А, 6В и 6С, показывают, что два репрезентативных тестируемых антитела, IgG А и В, специфически связываются с rhGM-CSF, но не с другими колониестимулирующими факторами, такими как M-CSF и G-CSF. Такая специфичность связывания антигена является важной для многообещающего терапевтического антитела-антитела. Пример 5. Характеристика данных по связыванию для человеческих моноклональных антител и их фрагментов против GM-CSF. Было желательным создание качественного ранжирования разных членов, идентифицированных в ELISA в качестве положительных связывателей rhGM-CSF, как описано выше в примере 2. Данное ранжирование было предназначено для того, чтобы отражать кинетические (скорость диссоциации) и равновесные (аффинность) параметры различных репрезентативных антител-связывателей, идентифицированных таким образом. Для этой цели осуществляли поверхностный плазмонный резонанс (SPR) на установке BIAcore ™ 2000, BIAcore AB (Uppsala, Швеция) со скоростью протока 5 мкл/мин и HBS-EP (0,01 М HEPES, pH 7,4, 0,15 М NaCl, 3 мМ EDTA, 0,005% поверхностно-активного вещества Р20) в качестве проточного буфера при 25 °С. Рекомбинантный человеческий GM-CSF (Лейкин, Berlex, ниже альтернативно именуемый "антиген" или "rhGM-CSF"), продуцируемый в дрожжах, иммобилизовали в проточных ячейках 2-4 на сенсорном чипе СМ5. Поверхность чипа активировали путем впрыскивания 80 мкл 0,1 М гидроксисукцинимида натрия, 0,4 М N-этил-N'-(3-диметиламинопропил)карбодиимида (NHS/EDC). Антиген связывали путем впрыскивания вручную 10 мкг/мл rhGM-CSF в 0,01 М ацетата натрия, pH 4,7. В проточных ячейках 2-4 иммобилизовали разные плотности антигена, регулируя количество ручных впрыскиваний. Проточную ячейку 1 оставляли немодифицированной, тогда как проточную ячейку 2 покрывали наиболее высокой из возможных плотностью rhGM-CSF (800 RU). Проточную ячейку 3 покрывали 50% количества антигена, иммобилизованного на проточной ячейке 2, и проточную ячейку 4 покрывали самой низкой плотностью rhGM-CSF (типично 10%). Активированную поверхность сенсорного чипа блокировали путем впрыскивания 85 мкл 1 М этаноламина и оставляли чип уравновешиваться в течение ночи при постоянной скорости тока HBS-EP 5 мкл/мин. Пример 5.1. Качественное определение кинетических параметров связывания (скорость диссоциации) для scFv-фрагментов человеческих моноклональных антител против GM-CSF. Эксперименты с использованием BIAcore проводили, как изложено в предыдущем параграфе. Перед данным экспериментом растворы элюированного белка периплазматического препарата ("РРР") диализовали против PBS при 25 °С в течение 2 ч и разбавляли 1:1 в HBS-EP. Кинетики связывания членов заявленного класса измеряли, впрыскивая 10 мкл раствора очищенного периплазматического белка при 25 °С на сенсорный чип. Неспецифическую фоновую адсорбцию белка на поверхности немодифицированного сенсорного чипа (FC1) вычитали из ответного сигнала в проточных ячейках с иммобилизованным rhGM-CSF (FC2, FC3, FC4). Определяли относительный ответный сигнал (FC2-1, FC3-1, FC4-1), и специфическую скорость диссоциации отслеживали в течение 100 с. Результаты этих экспериментов показаны на фиг. 7А для серии репрезентативных scFv-фрагментов, которые ранее были идентифицированы как положительные связыватели rhGM-CSF в экспериментах с использованием ELISA. Репрезентативные scFv фрагменты антител, для которых данные BIAcore показаны на фиг. 7А, являются следующими: scFv A, scFv В, scFv С, scFv D, scFv E, scFv F, scFv G, scFv H, scFv I, scFv J, scFv K, scFv L, scFv M и scFv N. Обычно при интерпретации результатов BIAcore амплитуда пика связывания (RUmax) непосредственно кореллирует с концентрацией белка в инъецированном образце. Кинетическая скорость ассоциации (ka) зависит от концентрации и, из-за варьирующих концентраций белка в РРР, не может использоваться для качественного ранжирования членов заявленного класса. Кинетическая скорость диссоциации (kd) не зависит от концентрации белка и характеризует силу связывания соответствующих членов заявленного класса. Все идентифицированные члены заявленного класса демонстрируют специфическое связывание с иммобилизованным rhGM-CSF. Члены заявленного класса с наилучшей кажущейся скоростью диссоциации были идентифицированы и, после дополнительной корреляции данных SPR с данными по ингибированию, представлены для определения аффинности с помощью экспериментов по равновесному связыванию на BIAcore. При оценке фиг. 7А видны отдельные пики для каждого из репрезентативных scFv фрагментов A-N антител, верхние части каждого из которых демонстрируют характерную кривизну, которую можно экстраполировать для получения скорости диссоциации рассматриваемого scFv-фрагмента. Тогда с точки зрения качества можно заключить, что каждый из репрезентативных scFv-фрагментов хорошо связывается с человеческим GM-CSF. Пример 5.2. Количественное определение равновесных параметров связывания (аффинности) для определенных человеческих антител и их scFv-фрагментов против GM-CSF. Установив в примере 5.1 качественно, что целый ряд scFv-фрагментов антител против GM-CSF, которые ранее были протестированы в качестве положительных на связывание GM-CSF в ELISA, демонстрируют приемлемые кинетические скорости диссоциации при связывании с человеческим GM-CSF, затем было желательным получение количественного представления такого связывания для антител и их фрагментов, фокусируясь на равновесных характеристиках связывания с рекомбинантным человеческим GM-CSF. Как показано выше в примере 4, специфическое связывание с антигеном - здесь rhGM-CSF - является одной из характеристик и специфических атрибутов класса антител и их фрагментов против GM-CSF, как здесь заявлено. Кинетику связывания (скорость диссоциации (kd) и скорость ассоциации (ka)) определенных репрезентативных членов класса человеческих антител и их фрагментов против GM-CSF измеряли путем инъекции 10 мкл очищенного белка (например, антитела или его фрагмента) в серии разведений, варьирующих от 10 мкг/мл до 1 пг/мл очищенного белка, и отслеживанием диссоциации при 25 °С в течение 100 с. Очищенный белок забуферивали в HBS-EP. Данные аппроксимировали с использованием программы BIAevalution ™, определяя константу скорости для кинетики диссоциации и ассоциации с помощью уравнения связывания Ленгмюра 1:1 (см. формулы 1 и 2 ниже), где А представляет собой концентрацию инъецированного очищенного анализируемого белка, и В[0] представляет собой Rmax: Кривые кинетики связывания определяли с использованием вплоть до 8 концентраций каждого репрезентативного анализируемого человеческого антитела или его фрагмента против GM-CSF. Независимая аппроксимация исходных данных давала константы скорости диссоциации и ассоциации, которые использовали для расчета равновесной константы диссоциации (KD). Результаты, полученные для каждого репрезентативного человеческого антитела или его фрагмента против GM-CSF показаны на фиг. 7B-I. Конкретно, на фиг. 7В показаны данные по связыванию, полученные для репрезентативного IgG В; на фиг. 7С показаны данные по связыванию, полученные для репрезентативного IgG А; на фиг. 7D показаны данные по связыванию, полученные для репрезентативного scFv С; на фиг. 7Е показаны данные по связыванию, полученные для репрезентативного scFv I; на фиг. 7F показаны данные по связыванию, полученные для репрезентативного scFv В; на фиг. 7G показаны данные по связыванию, полученные для репрезентативного scFv А; на фиг. 7Н показаны данные по связыванию, полученные для репрезентативного scFv E, и на фиг. 71 показаны данные по связыванию, полученные для репрезентативного scFv D. Эти данные обобщены ниже в табл. 2. Пример 6. Требования для минимального эпитопа определенного репрезентативного фрагмента человеческого антитела против GM-CSF. Было желательным определить эпитоп, связываемый человеческими антителами и их фрагментами против GM-CSF, как описано и заявлено здесь. Для этой цели провели анализ пептидных пятен ("pepspot") с использованием scFv А в качестве репрезентативного члена этого класса молекул и GM-CSF человека в качестве антигена. В общем, "pepspot" эксперимент осуществляли следующим образом. Перекрывающиеся 13-мерные пептиды, происходящие из аминокислотной последовательности hGM-CSF (относительно последовательности GM-CSF человека и определенных других приматов, смотри выше, а также фиг. 8А, а также SEQ ID NO: 49-51), ковалентно связывали С-концом с целлюлозной мембраной Whatman 50, тогда как ацетилированный N-конец оставался свободным. Индивидуальные полученные 13-мерные пептиды (полученные JPT Peptide Technologies GmbH) показаны ниже в табл. 3. Длина перекрывающейся последовательности любых двух соответствующих пептидов была установлена на уровне 11 аминокислот. Согласно протоколу производителя, мембрану промывали абсолютным EtOH в течение 1 мин с последующей промывкой TBS и блокированием TBS/3% БСА в течение ночи. Как и в случае каждой последующей инкубации и промывки, блокирование осуществляли при комнатной температуре. После промывки 3 раза TBS/0,05% Tween 20 в течение 10 мин мембрану инкубировали с 1 мкг/мл scFv А в TBS/3% БСА в течение 2,5 ч с последующей промывкой, проведенной, как описано выше. Определение scFv осуществляли с использованием антитела против пента-His (Qiagen, 0,2 мкг/мл в TBS/3% БСА) с последующим использованием антитела козы против IgG мыши (Fc-гамма-специфического), конъюгированного с пероксидазой хрена (Dianova, 1:10000 в TBS/3% БСА), где инкубацию с каждым из этих соответствующих антител осуществляли в течение 1 ч. После промывки 3 раза TBS/0,05% Tween 20 в течение 10 мин сигнал проявляли повышенной хемилюминисценцией (SuperSignalWest Pico Luminol/Enhancer Solution и SuperSignalWest Pico StablePeroxide Solution; Pierce) и экспозицией на пленке BioMax (Kodak). Сильные сигналы связывания scFv А с отрезками человеческого GM-CSF определяли на отрезке пептидных пятен между пятном А и В, а также на пятне С (см. табл. 3 ниже и фиг. 8В). Как можно видеть на фиг. 8В, связывание с другими пятнами, по-видимому, имело меньшую силу. Отрезок пятен, охватывающий точки А и В, соответствует аминокислотным остаткам 15-35. Все 13-мерные пептиды, составляющие эту область, содержат один минимальный аминокислотный отрезок из аминокислот 23-27 (RRLLN). Пятно С соответствует аминокислотным остаткам 65-72 (GLRGSLTKLKGPL) GM-CSF человека. Эти данные подразумевают, что scFv А, по-видимому, распознает прерывистый эпитоп. Во вторичной структуре GM-CSF человека аминокислоты 15-35 расположены в спирали А, тогда как остатки, соответствующие пятну С, представляют собой часть петлевой структуры, локализованной между спиралями С и D. Трехмерная модель укладки данной молекулы выявляет тесную стерическую близость этих сайтов относительно друг друга. Минимальный мотив аминокислотной последовательности в пептидах пятен А-В соответствует остаткам 23-27 GM-CSF человека (RRLLN). Увеличение силы сигнала от пятна А к В можно объяснить лучшей доступностью эпитопа RRLLN в пептиде, соответствующем пятну В, чем в пептиде, соответствующем пятну А. В пептиде А данный эпитоп локализован непосредственно на С-конце, который связан с мембраной, тогда как в пептиде В он локализован на более доступном N-конце данного пептида. Пример 7. Эффективность нейтрализации определенных человеческих антител/фрагментов антител против GM-CSF человека. Пример 7.1. Качественная оценка нейтрализующего потенциала определенных репрезентативных человеческих антител и их фрагментов против GM-CSF человека. Целью этого эксперимента является получение качественной информации о нейтрализующей активности репрезентативных человеческих нейтрализующих антител и их фрагментов против GM-CSF. Для этого использовали человеческую GM-CSF-зависимую клеточную линию TF-1 (DSMZ, АСС 334). Скорость пролиферации этой клеточной линии зависит от присутствия GM-CSF человека, так что для определения, существует ли такая нейтрализующая активность на самом деле, можно использовать измерение роста клеток после инкубации клеток с человеческим GM-CSF в присутствии или в отсутствие антитела, у которого подозревают наличие GM-CSF-нейтрализующей активности. Клетки TF-1 культивировали в среде RPMI 1640 (Gibco; не содержит L-глутамин, феноловый красный), 10% инактивированной нагреванием FCS, в присутствии 2,5 нг/мл rhGM-CSF, как описано дистрибьютером (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Германия). Клетки выращивали до плотности клеток 0,5 ×10 6 клеток/мл. Для анализа пролиферации клетки TF-1 собирали центрифугированием при 300 ×g в течение 4 мин и промывали 1 ×PBS (среда Дульбекко, Gibco). Клетки ресуспендировали до конечной концентрации 1 ×10 5 клеток/мл в RPMI 1640, 10% FCS, и использовали 90 мкл клеточной суспензии лунку в плоскодонном планшете для культуры клеток Microtest (0,9 ×10 4 клеток/лунку). Для стимуляции пролиферации клеток TF-1 использовали конечную концентрацию 0,3 нг/мл rhGM-CSF. Для нейтрализации GM-CSF-зависимой пролиферации очищенный РРР соответствующих фрагментов человеческого антитела против GM-CSF диализовали против 1 ×PBS при 25 °С в течение 2 ч. 10 мкл диализовали, и белковый раствор, подвергнутый стерилизующей фильтрации (0,22 мкм фильтр), добавляли к 100 мкл раствора, содержащего TF-1 и rhGM-CSF. После инкубации в течение 72 ч при 37 °С в 5% CO 2 определяли пролиферативный статус клеток TF-1 с помощью колориметрического анализа, основанного на расщеплении тетразолиевых солей (WST-1, Roche) митохондриальной дегидрогеназой в жизнеспособных клетках. Формазановый краситель, образуемый метаболически активными клетками, количественно оценивали путем измерения его абсорбции с помощью ELISA-ридера при 450 нм. Ингибирование пролиферации клеток TF-1, зависимой от GM-CSF человека, тестируемыми репрезентативными фрагментами человеческого антитела против GM-CSF человека, варьировало по силе (фиг. 9). В то время как два таких фрагмента не имели нейтрализующего эффекта (scFv F и scFv L), пять конструкций (scFv J, scFv K, scFv M, scFv N и scFv H) показывали промежуточное ингибирование, и семь конструкций (scFv В, scFv I, scFv E, scFv D, scFv G, scFv C, scFv А) показывали сильное ингибирование GM-CSF-зависимой пролиферации клеток TF-1. Недостаток или более низкий уровень нейтрализующего эффекта мог быть обусловлен более низким уровнем экспрессии конкретных репрезентативных scFv или менее стабильным комплексом, образующимся между конкретным репрезентативным scFv и rhGM-CSF в течение 72-часового периода инкубации при 37 °С. Пример 7.2. Количественная оценка нейтрализующего потенциала определенных репрезентативных человеческих антител и их фрагментов против GM-CSF человека, измеренная с помощью пролиферации клеток. Выбранные репрезентативные молекулы scFv, которые, как было показано выше, показывают сильное ингибирование пролиферации TF-1, затем подвергали количественному анализу эффективности нейтрализации. С этой целью использовали ту же клеточную линию TF-1, зависимую от GM-CSF человека (DSMZ АСС 334). Клетки TF-1 культивировали и готовили для анализа пролиферации, как подробно описано выше в примере 7.1. Для стимуляции пролиферации клеток TF-1 использовали конечную концентрацию rhGM-CSF 0,3 нг/мл. Для нейтрализации GM-CSF-зависимой пролиферации к раствору, содержащему 100 мкл TF-1 и rhGM-CSF, добавляли в серии разведений 10 мкл очищенных образцов репрезентативных человеческих нейтрализующих моноклональных антител или их фрагментов против GM-CSF человека. Конечные концентрации белка варьировали от 10 мкг/мл до 10 пг/мл. Образцы инкубировали при 37 °С в 5% СО 2 в течение 72 ч. Через 72 ч определяли пролиферативный статус клеток TF-1, как описано выше в примере 7.1. Данные аппроксимировали для полумаксимального ингибирования пролиферации (IC 50 ), используя аппроксимацию нелинейной кривой регрессии с помощью программы Prism. Очевидный эффект нейтрализации GM-CSF, наблюдаемый в качественном эксперименте по ингибированию пролиферации, описанном в примере 7.1 выше, можно было подтвердить и количественно оценить. Все протестированные scFv-фрагменты человеческих моноклональных нейтрализующих антител против GM-CSF человека демонстрировали в этом эксперименте по ингибированию пролиферации константу полумаксимального ингибирования (IC 50 ) в наномолярном интервале. Можно было установить четкое ранжирование эффективности нейтрализации, как видно на фиг. 10А. Протестированные человеческие моноклональные нейтрализующие антитела IgG против GM-CSF человека демонстрируют значительно более высокую эффективность нейтрализации, чем их scFv аналоги. Константа полумаксимального ингибирования молекул IgG, полученных в этом эксперименте, находилась в субнаномолярном интервале. Как можно видеть на фиг. 10В, IC 50 , оцениваемая для IgG А, составляла 0,9 нМ, и IgG В имел IC 50 0,3 нМ. Для того чтобы проверить, соответствуют ли фрагменты антител scFv, полученные из IgG А и В (scFv О и Р соответственно) количественно по их нейтрализующему потенциалу фрагментам scFv А и В, выполнили аналогичные анализы нейтрализации TF-1, как описано выше, за исключением использования в качестве тестируемых молекул scFv О и Р. Данные результаты показаны на фиг. 10С и 10D для scFv P и О соответственно. Как можно увидеть на фиг. 10D, scFv О имеет такой же нейтрализующий потенциал, как и scFv A, что показывает, что реконверсия из IgG обратно в формат scFv возможна без потери биологической активности. Пример 7.3. Количественная оценка нейтрализующего потенциала определенных репрезентативных человеческих антител и их фрагментов против rhGM-CSF, измеренная с помощью снижения продукции IL-8. Этот эксперимент проводили для количественной оценки нейтрализующей активности репрезентативных человеческих антител и их фрагментов против GM-CSF человека путем измерения GM-CSF-зависимой продукции IL-8 клетками U-937. Антиген GM-CSF, используемый в вышеупомянутых экспериментах, представлял собой rhGM-CSF. Моноцитарные клетки U-937 культивировали в среде RPMI 1640 Gibco (не содержит L-глутамин, феноловый красный), дополненной 10% инактивированной нагреванием FCS, как описано дистрибьютером (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Германия). Клетки выращивали до плотности клеток 1 ×10 6 клеток/мл. При проведении анализа ингибирования, основанного на измерении продукции IL-8, клетки собирали центрифугированием при 300 ×g в течение 4 мин и ресуспендировали до конечной концентрации 1 ×10 6 клеток/мл в RPMI 1640, 10% FCS. 1,8 ×10 5 клеток/лунку (180 мкл клеточной суспензии) высевали в лунку плоскодонного планшета для культур клеток Microtest. Для стимуляции продукции IL-8 клетками U-937 использовали конечную концентрацию rhGM-CSF 1 нг/мл. К 180 мкл клеток U-937 и раствора rhGM-CSF в серии разведений добавляли 20 мкл очищенного scFv или IgG, что давало конечные концентрации белка, варьирующие от 10 мкг/мл до 10 пг/мл. После инкубации в течение 18 ч при 37 °С и 5% СО 2 , клетки осаждали центрифугированием культуральных планшетов при 600 ×g в течение 2 мин. Культуральные супернатанты собирали с помощью пипетки в новый планшет и анализировали для определения в них концентрации IL-8 с использованием набора OptEIA Human IL-8 ELISA (Becton Dickenson and Company). Определение с помощью ELISA осуществляли согласно инструкциям производителя. Кратко, 50 мкл иммобилизованного антитела, разведенного в 0,1 М карбоната натрия, pH 9,5, наносили на планшеты для микротестирования в течение ночи при 4 °С. После промывки 3 раза PBS/0,05% Tween 20, блокировали лунки 200 мкл PBS/ 10% FCS на лунку в течение 1 ч при комнатной температуре с последующими промывками 3 раза PBS/0,05% Tween 20. Затем в лунки добавляли 50 мкл образцов культурального супернатанта и инкубировали в течение 2 ч при комнатной температуре. Для последующей количественной оценки концентрации IL-8 осуществляли последовательное разведение стандарта IL-8, предоставленного производителем, в течение данной процедуры. После промывки 5 раз PBS/0,05% Tween 20, осуществляли определение с использованием 50 мкл рабочего детектора (определение Ab + Av-HRP), предоставленного в наборе OptEIA Human IL-8 ELISA. После 1 ч инкубации при комнатной температуре, лунки промывали еще 7 раз. Сигнал проявляли путем добавления раствора субстрата OPD (Sigma) и определяли при длине волны 490 нм (используя эталонную длину волны 620 нм). Строили стандартную кривую IL-8 для калибровки и концентрацию IL-8 в образцах культуральных супернатантов рассчитывали согласно этой калибровочной кривой. Данные аппроксимировали для полумаксимального ингибирования продукции IL-8 (IC 50 ) с использованием аппроксимации нелинейной кривой регрессии с помощью программы Prism. Все протестированные репрезентативные фрагменты человеческих моноклинальных нейтрализующих антител против rhGM-CSF показывали очевидное ингибирование GM-CSF-зависимой продукции IL-8 клетками U-937, как можно ясно видеть по снижению концентрации IL-8 при увеличении концентрации scFv на фиг. 11. Ранжирование по эффективности нейтрализации, наблюдаемое в этом эксперименте, согласуется с ранжированием, полученным при тестировании таких же молекул на их нейтрализующий эффект в эксперименте по ингибированию пролиферации TF-1, описанном выше. Следует отметить, что значения IC 50 , определенные в этом эксперименте, выше при сравнении со значениями, полученными для таких же молекул в предыдущем эксперименте по пролиферации TF-1. Это обусловлено более высокой концентрацией GM-CSF, необходимой для стимуляции продукции IL-8 клетками U-937, чем концентрация, необходимая для стимуляции TF-1. Пример 7.4. Количественная оценка нейтрализующего потенциала репрезентативных человеческих антител и их фрагментов против GM-CSF человека на рекомбинантном GM-CSF макак, измеренная с помощью пролиферации клеток. Целью этого эксперимента была демонстрация нейтрализующей способности соответствующих человеческих антител и их фрагментов против GM-CSF человека в отношении GM-CSF из нечеловекообразных приматов семейства макак ("macGM-CSF"). Для демонстрации нейтрализующего эффекта селектированных молекул scFv и IgG на macGM-CSF провели эксперимент по ингибированию пролиферации согласно протоколу, описанному в примерах 7.1 и 7.2, используя macGM-CSF вместо hGM-CSF. И hGM-CSF, и macGM-CSF стимулируют пролиферацию клеток TF-1 с такой же полумаксимальной эффективностью (ЕС 50 ). Конечную концентрацию 3 нг/мл macGM-CSF использовали для стимуляции пролиферации клеток TF-1 в эксперименте, тестирующем молекулы scFv, и 0,3 нг/мл rhGM-CSF использовали в эксперименте, тестирующем IgG В в качестве репрезентативного человеческого антитела против GM-CSF. Для того чтобы нейтрализовать пролиферацию клеток TF-1, 10 мкл очищенного человеческого антитела или его фрагмента против GM-CSF человека добавляли к 100 мкл раствора TF-1 и macGM-CSF в серии разведений. Конечные концентрации белка варьировали от 10 мкг/мл до 10 пг/мл. Образцы инкубировали при 37 °С в 5% СО 2 в течение 72 ч. Через 72 ч определяли пролиферативный статус клеток TF-1, как описано в примерах 7.1 и 7.2. Данные аппроксимировали для полумаксимального ингибирования пролиферации (IC 50 ) с использованием аппроксимации нелинейной кривой регрессии с помощью программы Prism. Как видно на фиг. 12А, определенные репрезентативные фрагменты человеческих моноклональных антител против GM-CSF человека также демонстрировали явный нейтрализующий потенциал в отношении macGM-CSF (scFv В, scFv E, scFv С, scFv I, scFv А). Кроме того, как можно видеть на фиг. 12В, возрастающие концентрации репрезентативного человеческого моноклонального антитела IgG В против GM-CSF человека явно приводили к снижению пролиферации TF-1, демонстрируя нейтрализующий потенциал этого антитела. Интересно, что значение IC 50 полученное в этом эксперименте для IgG В (0,3 нМ) с использованием macGM-CSF для индукции пролиферации клеток TF-1, равно значению, полученному в эксперименте с использованием hGM-CSF, демонстрируя явную перекрестную реактивность IgG В в отношении GM-CSF у этих видов. Пример 8. Перекрестная реактивность IgG В с GM-CSF из разных видов. Перекрестную реактивность IgG В с GM-CSF из разных видов, не являющихся человеком, исследовали для идентификации видов, подходящих для последующих исследований in vivo. В первой группе экспериментов тестировали связывание IgG В с имеющимися в продаже рекомбинантными GM-CSF человека (Лейкин ®, Berlex), свиньи, собаки, крысы (R &D Systems, Wiesbaden, Германия) и мыши (Strathmann Biotech, Hamburg, Германия) в ELISA-эксперименте. Конкретно, планшет для ELISA покрывали 1 мкг/мл GM-CSF из разных упомянутых видов. IgG В добавляли в серии разведений и определяли с использованием антитела против человеческого IgG1, конъюгированного с пероксидазой хрена. ELISA проявляли путем добавления раствора субстрата OPD (о-фенилендиамин, "OPD", желто-оранжевый при реакции с пероксидазой) (Roche, Германия) и осуществляли измерение при 490 нм. Как видно на фиг. 13, IgG В продемонстрировал надежное связывание с рекомбинантным человеческим GM-CSF, тогда как GM-CSF из других протестированных видов не распознавался. Свинья, собака, крыса или мышь, следовательно, не могут быть подходящими видами для тестирования in vivo. Однако как видно выше в примере 7.4, IgG В демонстрирует заметную перекрестную реактивность с macGM-CSF (из макака-крабоеда, Macaca fascicularis), что наводит на мысль о применимости по меньшей мере одного вида обезьян из семейства макак для исследований IgG В in vivo. Пример 9. Связывание IgG В с различно гликозилированными вариантами GM-CSF. Целью этого эксперимента было определение степени, до которой связывание IgG В с GM-CSF зависит от картины гликозилирования последнего. С этой целью серии разведений кондиционированной среды, содержащей природный hGM-CSF (человеческое гликозилирование), а также рекомбинантный hGM-CSF из Е.coli (нет гликозилирования) и дрожжей (дрожжевое гликозилирование), а также рекомбинантный GM-CSF макака тестировали на их способность индукцировать пролиферацию TF-1. Человеческий гликозилированный GM-CSF получали из культурального супернатанта клеток BEAS-2B, обработанных IL-1 β (человеческие клетки легкого, полученные от АТСС CRL-9609). Клетки BEAS-2B размножали в среде ВЕВМ, замещенной набором BEGM Bullet (Cambrex, Verviers, Belgium), но культивировали в RPMI 1640, 10% FCS, в присутствии 50 нг/мл IL-1 β (Strathmann Biotech, Hamburg, Германия) для индууции продукции GM-CSF. После 48-часовой инкубации при 37 °С, 5% СО 2 , культуральный супернатант анализировали на содержание в нем GM-CSF с использованием набора для ELISA OptEIA Human GM-CSF (BD Biosciences, Heidelberg, Германия) согласно инструкциям производителя. Рекомбинантный hGM-CSF из Е.coli продуцировался внутри, как представлено в примере 1.1 WO 2005/105844. Рекомбинантный hGM-CSF из дрожжей получали из коммерческого источника под торговым наименованием "Лейкин" (Berlex, USA). GM-CSF макака рекомбинантно продуцировали в клетках НЕК293. Серию разведений кондиционированной среды, содержащей природный hGM-CSF, а также рекомбинантный hGM-CSF из Е.coli и дрожжей и GM-CSF макака сначала тестировали на их способность индуцировать пролиферацию TF-1. Все три варианта гликозилирования GM-CSF человека и GM-CSF макака показывали очень сходные значения ЕС 50 для активации TF-1. Они составляли 10 пг/мл для hGM-CSF, продуцируемого Е.coli, 15 пг/мл для hGM-CSF, продуцируемого дрожжами, 36 пг/мл для hGM-CSF, продуцируемого клетками легкого человека и 11 пг/мл для GM-CSF макака, соответственно (фиг. 14А). Затем определяли нейтрализующую активность IgG В в присутствии 0,3 нг/мл рекомбинантного hGM-CSF или 0,2 нг/мл физиологического hGM-CSF. Через 72 ч пролиферативный статус клеток TF-1 в присутствии разных концентраций IgG В количественно оценивали с помощью колориметрической реакции (фиг. 14Б). Взятые вместе, эти данные, показанные на фиг. 14, демонстрируют, что GM-CSF-зависимая пролиферация клеток TF-1, ингибируемая IgG В в субнаномолярных концентрациях, по-видимому, не зависит от картины гликозилирования GM-CSF человека. Следовательно, картина гликозилирования GM-CSF человека не оказывает существенного влияния на способность IgG В нейтрализовать активность GM-CSF. Пример 10. Влияние IgG В на биологические активности GM-CSF на эозинофилах. Пример 10.1. Влияние IgG В на GM-CSF-опосредованное выживание эозинофилов. Одной из многих биологических активностей GM-CSF является продление выживания эозинофильных и нейтрофильных гранулоцитов. Так как воспалительные заболевания легких связаны с локальным накоплением эозинофилов, которые играют существенную роль в поддержании воспаления, протестировали эффективность IgG В в ингибировании GM-CSF-опосредованного выживания эозинофилов. Эозинофилы выделяли из периферической крови здоровых доноров путем элиминации CD16 + нейтрофилов из популяции гранулоцитов, полученной центрифугированием в градиенте плотности, и лизиса эритроцитов. Свежевыделенные эозинофилы периферической крови высевали с плотностью 5 ×10 4 клеток/лунку в RPMI 1640/ 10% FCS и пенициллин/стрептомицин в 96-луночные плоскодонные планшеты для микротестов. GM-CSF добавляли в серии разведений, варьирующих от 33 нг/мл до 10 пг/мл для мониторинга выживания эозинофилов, зависимого от концентрации. Для анализа ингибирующего потенциала IgG В на GM-CSF-зависимое выживание эозинофилов добавляли антитело в серии разведений, варьирующих от 10 мкг/мл до 0,1 нг/мл. Для воздействия на выживание эозинофилов использовали конечную концентрацию GM-CSF 0,1 нг/мл. После инкубации в течение 72 ч при 37 °С, 5% СО 2 , добавляли реагент WST-1. Полученную в результате колориметрическую реакцию, соответствующую части жизнеспособных клеток, количественно оценивали путем измерения поглощения при 450 нм. Данные анализировали и аппроксимировали для полумаксимального ингибирования выживания (IC 50 ), используя аппроксимацию нелинейной кривой регрессии с помощью программного пакета Prism. Как видно на фиг. 15А, определили полумаксимальную эффективную дозу (ЕС 50 ) rhGM-CSF, составляющую 0,02 нг/мл. Как видно на фиг. 15В, наблюдали мощный нейтрализующий эффект IgG В с полумаксимальным ингибированием выживания эозинофилов при концентрации антитела 0,13 нМ. Эти данные указывают на то, что IgG В эффективен при ингибировании GM-CSF-зависимого выживания эозинофилов дозозависимым образом. Пример 10.2. Влияние IgG В на GM-CSF-индуцированную активацию эозинофилов. Также было желательным исследовать влияние IgG В на GM-CSF-индуцированную активацию эозинофилов. Обнаружили, что экспрессия CD69 подвергается повышающей регуляции на периферических эозинофилах (CD16 - ), выделенных из крови человека после стимуляции в течение 20 ч или 3 суток (а) 0,1 нг/мл GM-CSF или (б) 0,1 нг/мл GM-CSF, IL-3 и IL-5, но не с (в) 0,1 нг/мл только IL-3 и IL-5 (фиг. 16А). Эозинофилы, культивируемые в присутствии только среды, не демонстрировали повышающей регуляции CD69. Следовательно, CD69 можно взять в качестве маркера активации эозинофилов GM-CSF, и уровень экспрессии CD69 отслеживали как показатель GM-CSF-зависимой активации эозинофилов. В оба момента времени (20 ч и 3 суток) IgG В (10 мкг/мл) практически полностью предотвращал GM-CSF-зависимую активацию эозинофилов, что видно по отсутствию экспрессии CD69 при проточной цитометрии. Эозинофилы выделяли, как описано выше в примере 10.1, и культивировали при плотности 5 ×10 5 клеток/лунку в RPMI 1640/10% FCS и пенициллин/стрептомицин в 48-луночном плоскодонном планшете для микротестов. Клетки инкубировали только со средой или в присутствии только 0,1 нг/мл GM-CSF или совместно с 0,1 нг/мл IL-3 и IL-5. 10 мкг/мл IgG В использовали для нейтрализации GM-CSF. После инкубации в течение 1 или 3 суток клетки анализировали на экспрессию CD69 с помощью проточной цитометрии. Определение CD69 с помощью проточной цитометрии. Экспрессию CD69 на эозинофилах определяли на установке FACS Calibur (Becton Dickinson). 10 5 клеток инкубировали с 5 мкл антитела против человеческого CD16, конъюгированного с FITC (клон 3G8, BD Biosciences), и антитела против человеческого CD69, конъюгированного с РЕ (клон FN50, BD Biosciences), с каждым в течение 1 ч при 4 °С. В качестве отрицательного контроля использовали нерелевантные, соответствующие по изотипу FITC- и РЕ-конъюгированные антитела. После инкубации клетки дважды промывали PBS, 1% FCS, 0,05% NaN 3 и ресуспендировали в 250 мкл PBS, 1% FCS, 0,05% NaN 3 . Непосредственно перед FACS анализом добавляли йодистый пропидий до конечной концентрации 1 мкг/мл для того, чтобы пометить мертвые клетки. Интерпретацию данных осуществляли с использованием программы CellQuestPro (BD Biosciences). Окрашиваемые йодистым пропидием (т.е. мертвые) клетки исключали из анализа экспрессии CD69. IgG В также снижал процентное содержание живых и активированных эозинофилов при мониторинге путем окрашивания йодистым пропидием клеток CD167CD69 + в присутствии 0,1 нг/мл GM-CSF. IgG В снижал процентное содержание активированных клеток от 35 до 8% через 1 сутки и от 43 до 3% через 3 суток после культивирования. В присутствии 0,1 нг/мл GM-CSF, IL-3 и IL-5 процентное содержание живых и активированных эозинофилов снижалось от 32 до 8% и от 48 до 11% после 1 и 3 суток соответственно. Даже несмотря на то что повышающая регуляция CD69 полностью ингибировалась IgG В, более высокие количества эозинофилов в состоянии покоя (CD16 - /CD69 + ) выживали в течение 3 суток в присутствии 0,1 нг/мл GM-CSF, IL-3 и IL-5 по сравнению с клетками, инкубируемыми со средой или только с GM-CSF (фиг. 16А, последний столбец). То же самое наблюдали для клеток, инкубируемых в присутствии 0,1 нг/мл IL-3 плюс IL-5. В экспериментах по нахождению дозы IgG В добавляли в серии разведений к эозинофилам, культивируемым в присутствии 0,1 нг/мл GM-CSF (фиг. 16В). Ингибирующий эффект IgG В на CD69-зависимую среднюю интенсивность флуоресценции (MFI) наблюдали с полумаксимальной концентрацией IgG В 0,22 нМ. При совместном рассмотрении эти данные указывают на то, что IgG В является эффективным нейтрализатором активности GM-CSF в биологическом контексте, весьма релевантном для воспалительных заболеваний дыхательных путей, например астмы. Пример 11. Предварительные исследования токсичности ex vivo с использованием IgG В. Как объяснено выше, нейтрализация активности GM-CSF может быть терапевтически полезной при целом ряде болезненных состояний. Однако в то же время GM-CSF играет важную роль в нормальной функции иммунной системы при борьбе с экзогенными патогенами, например, как это происходит при фагоцитозе нейтрофильными гранулоцитами и моноцитами. Эта природная функция нейтрофилов и моноцитов должна оставаться не подверженной воздействию в присутствии терапевтических количеств IgG В. Поэтому авторы изобретения исследовали два аспекта процесса фагоцитоза: 1) поглощение бактерий (фагоцитоз); и 2) активность окислительного взрыва (свидетельствующая о внутриклеточном киллинге). Эти исследования подробно описаны в следующих примерах. Пример 11.1. Поглощение бактерий (фагоцитоз). Определение фагоцитарной активности гранулоцитов и моноцитов в гепаринизированной цельной крови осуществляли с использованием набора Phagotest, произведенного Orpegen (Heidelberg, Германия). Этот тест основан на поглощении опсонизированной, флуоресцентно меченых клеток Е.coli фагоцитами. Эти клетки затем можно определить по зеленой флуоресценции при проточной цитометрии. 20 мкл флуоресцеин-меченых, опсонизированных клеток Е.coli добавляли к 100 мкл гепаринизированной цельной крови и инкубировали при 37 °С. В качестве отрицательного контроля осуществляли инкубацию при 0 °С. Через 10 мин процесс фагоцитоза останавливали путем охлаждения образцов на льду и добавления 100 мкл гасящего раствора (Orpegen). Этот раствор позволяет различать прикрепление и интернализацию бактерий путем гашения FITC-флуоресценции бактерий, связанных на поверхности, тогда как флуоресценция интернализованных частиц не подвергается воздействию. После трех стадий промывки 3 мл раствора для промывки (Orpegen) эритроциты лизировали. Оставшиеся лейкоциты один раз промывали 3 мл раствора для промывки (Orpegen). После добавления 200 мкл раствора для окрашивания ДНК, который обеспечивает исключение агрегированных бактерий или клеток, анализировали клетки с помощью проточной цитометрии. Процентное содержание клеток, осуществивших фагоцитоз, определяли посредством FITC-флуоресценции. Для определения влияния IgG В на фагоцитоз в три идентичных образца крови добавляли IgG В до конечной концентрации 10 мкг/мл. Затем эти три образца оставляли инкубироваться при 37 °С с IgG В в течение разных периодов времени перед добавлением Е.coli. В первый образец добавляли Е.coli немедленно, тогда как во второй и в третий образец добавляли Е.coli соответственно через 24 и 48 ч. Результаты, наблюдаемые для гранулоцитов: непосредственно после взятия крови свыше 98% гранулоцитов поглощали бактерии как в присутствии, так и в отсутствие IgG В (фиг. 17А). После инкубации образцов крови с IgG В в течение 24 ч определяли снижение до примерно 92% без IgG В, и до 90% в присутствии IgG В (фиг. 17В). Через 48 ч 81% гранулоцитов были положительными в отношении фагоцитоза в отсутствие и 89% - в присутствии IgG В (фиг. 17С). Результаты, наблюдаемые для моноцитов: независимо от того, присутствует IgG В или нет, непосредственно после взятия крови 98% моноцитов были фагоцитирующими (фиг. 18А). После 24 ч предынкубации с IgG В 90%, моноцитов были положительными (фиг. 18В). После 24 ч предынкубации без IgG В, такими были 92% моноцитов. Авторы изобретения обнаружили, что через 48 ч 81% моноцитов без IgG В и 89% с IgG В были положительными в отношении фагоцитоза (фиг. 18В). Пример 11.2. Окислительный взрыв. Определение активности окислительного взрыва гранулоцитов и моноцитов в гепаринизированной цельной крови осуществляли с использованием набора Phagoburst от Orpegen (Heidelberg, Германия). Этот анализ позволяет определять процентное содержание фагоцитирующих клеток, которые продуцируют реакционноспособные окислители, путем окисления субстрата дигидрородамина (DHR) 123 до флуоресцентного R 123. Клетки, демонстрирующие активность окислительного взрыва, можно идентифицировать при проточной цитометрии. Гепаринизированную кровь инкубировали с разными стимулами для индукции активности окислительного взрыва: с форбол-12-миристат-13-ацетатом ("РМА"), в качестве сильного стимула; с немечеными, опсонизированными клетками Е.coli в качестве промежуточного стимула и с хемотаксическим пептидом N-формил-MetLeuPhe (fMPL) в качестве слабого стимула. 100 мкл цельной крови инкубировали с этими стимулами при 37 °С. В качестве отрицательного контроля осуществляли инкубацию без стимуляции. После 10 мин инкубации добавляли раствор субстрата DHR 123 и осуществляли инкубацию в течение еще 10 мин. DHR 123 превращается окисляющими клетками в флуоресцентный R 123. После трех стадий промывки 3 мл раствора для промывки (Orpegen) эритроциты лизировали. Оставшиеся лейкоциты промывали один раз 3 мл раствора для промывки (Orpegen). После добавления 200 мкл раствора для окрашивания ДНК, что обеспечивает исключение агрегированных бактерий или клеток, анализировали клетки с помощью проточной цитометрии. Для определения влияния IgG В на окислительный взрыв, к трем идентичным образцам крови добавляли IgG В в конечной концентрации 10 мкг/мл. Каждый из этих трех образцов затем делили на три аликвоты и давали им инкубироваться при 37 °С в течение разного периода времени перед добавлением к отдельным аликвотам Е.coli, fMLP или РМА. Е.coli, fMLP или РМА добавляли к трем аликвотам первого образца немедленно, тогда как Е.coli, fMLP или РМА добавляли к трем аликвотам второго и третьего образца, соответственно, через 24 и 48 ч. Параллельные образцы крови, не содержащие IgG В, обрабатывали идентично тому, как это описано выше для контролей. Результаты показаны ниже в табл. 4, где "+" во втором столбце слева указывает на то, что IgG В присутствует в тестируемой аликвоте образца, и "-" во втором столбце слева указывает на контроль, не содержащий IgG В. Аналогичные результаты получали при использовании моноцитов вместо гранулоцитов. Данный эксперимент осуществляли аналогично эксперименту, описанному выше, и результаты показаны ниже в табл. 5, где "+" во втором столбце слева указывает на то, что IgG В присутствует в тестируемой аликвоте образца, и "-" во втором столбце слева указывает на контроль, не содержащий IgG В. Поэтому в целом можно заключить, что присутствие IgG В при физиологически релевантных температурах не оказывает вредного воздействия на фагоцитоз или окислительный киллинг бактерий как гранулоцитами, так и моноцитами. Тогда, в контексте in vivo, эти результаты говорят о том, что не ожидается, что терапевтическое введение IgG В окажет вредное воздействие на нормальные иммунные защитные механизмы пациента. |