|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000013109b*\id |
|
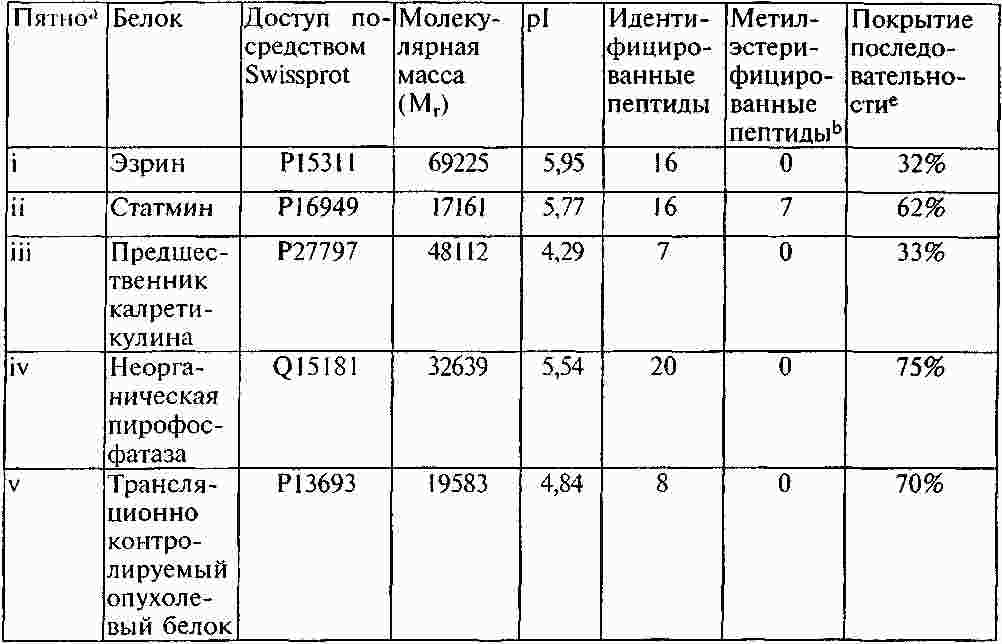
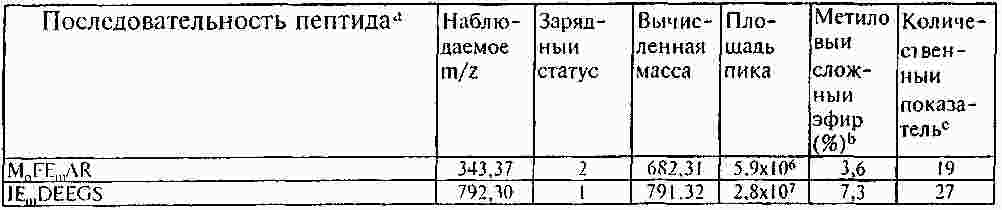
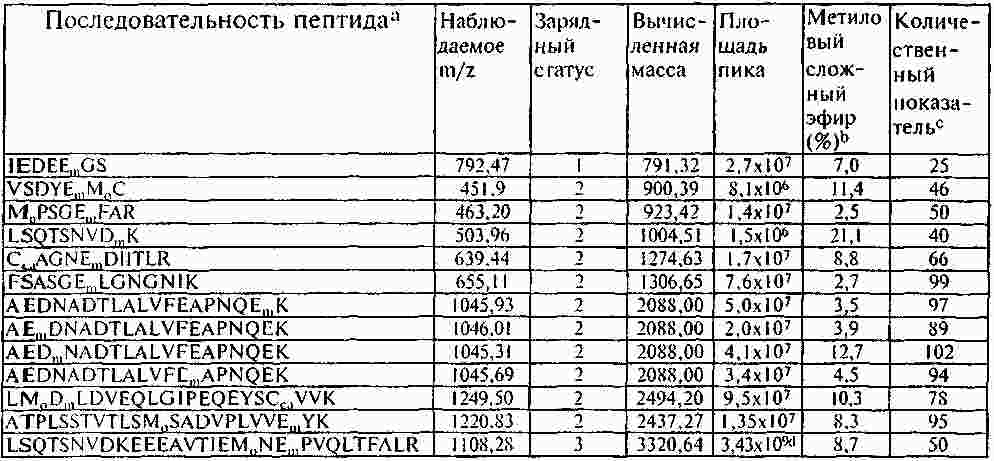
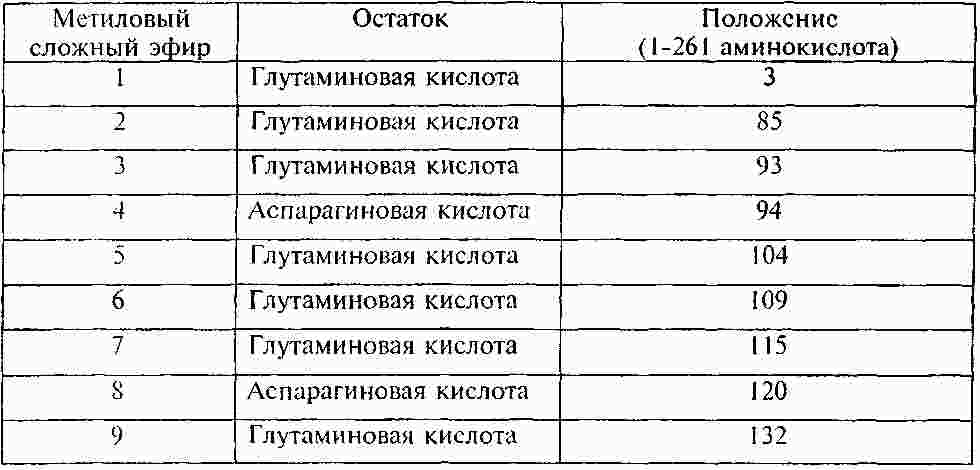
Термины запроса в документе Реферат Раскрывают способы и композиции для выявления присутствия csPCNA изоформы посредством идентифицирования одной или нескольких посттрансляционных модификаций. Раскрывают способы идентифицирования csPCNA изоформы через посредство посттрансляционных модификаций, включая уровни метилэстерификации. Формула [0001] Способ выявления раковой изоформы ядерного антигена пролиферирующих клеток (csPCNA) в биологическом образце, включающий [0002] Способ по п.1, отличающийся тем, что биологическим образцом является биологическая жидкость организма. [0003] Способ по п.1, отличающийся тем, что биологическая жидкость организма выбрана из группы, включающей кровь, плазму, лимфу, сыворотку, плевральный выпот, цереброспинальную жидкость, слюну, мокроту, мочу, желудочный сок, сок поджелудочной железы, асцитическую жидкость, синовиальную жидкость, молоко и сперму. [0004] Способ по п.1, отличающийся тем, что биологическим образцом является образец ткани. [0005] Способ по п.1, отличающийся тем, что ткань выбрана из группы, включающей ткань молочной железы, ткань предстательной железы, ткань легких, ткань толстой кишки, эпителиальную ткань, соединительную ткань, ткань шейки матки, ткань пищевода, ткань головного мозга, ткань вилочковой железы, ткань щитовидной железы, ткань поджелудочной железы, ткань яичек, ткань яичника, ткань кишечника, ткань мочевого пузыря, ткань желудка, ткань саркомы мягких тканей, ткань остеосаркомы, лейкозную ткань, ткань лимфомы, ткань карциномы, ткань аденокарциномы, ткань плаценты, волокнистую соединительную ткань, ткань половых клеток и их экстракты. [0006] Способ по п.1, отличающийся тем, что метиловый сложный эфир присутствует на аспарагиновой кислоте. [0007] Способ по п.1, отличающийся тем, что метиловый сложный эфир присутствует на глутаминовой кислоте. [0008] Способ по п.1, отличающийся тем, что метиловый сложный эфир присутствует на одном или нескольких из 16 остатков аспарагиновой или глутаминовой кислот на csPCNA изоформе. [0009] Способ по п.8, отличающийся тем, что метиловый сложный эфир присутствует на одном или нескольких из 16 остатков аспарагиновой или глутаминовой кислот, соответствующих аминокислотным положениям 3, 85, 93, 94, 104, 109, 115, 120, 132, 143, 174, 189, 201, 238, 255 и 259 csPCNA изоформы. [0010] Способ по п.8, отличающийся тем, что метиловый сложный эфир присутствует на одном или нескольких из 16 остатков аспарагиновой или глутаминовой кислот, соответствующих пептидам с модифицированными аминокислотными остатками, выбранными из числа: [0011] Способ по п.1, отличающийся тем, что выявление csPCNA изоформы осуществляется посредством масс-спектрометрического анализа. [0012] Способ по п.11, отличающийся тем, что масс-спектрометрическим анализом является жидкостной хроматографический (LC) масс-спектрометрический (MS) анализ. [0013] Способ по п.11, отличающийся тем, что масс-спектрометрический анализ csPCNA пептида выявляет 14 Да смещение массы по сравнению с соответствующим немодифицированным пептидом. [0014] Способ диагностирования либо прогнозирования злокачественного новообразования, включающий [0015] Способ по п.14, отличающийся тем, что посттрансляционной модификацией является метилэстерификация. [0016] Способ по п.15, отличающийся тем, что метилэстерификация присутствует на одном или нескольких из 16 остатков аспарагиновой либо глутаминовой кислот на csPCNA изоформе. [0017] Способ по п.14, отличающийся тем, что посттрансляционная модификация включает определение уровней метилэстерификации csPCNA изоформы и nmPCNA изоформы. [0018] Способ по п.16, отличающийся тем, что метиловый сложный эфир присутствует на одном или нескольких из 16 остатков аспарагиновой или глутаминовой кислот, соответствующих аминокислотным положениям 3, 85, 93, 94, 104, 109, 115, 120, 132, 143, 174, 189, 201, 238, 255 и 259 csPCNA изоформы. [0019] Модифицированный пептид ядерного антигена пролиферирующих клеток (PCNA), включающий аминокислотную последовательность, выбранную из группы, включающей: [0020] Модифицированный пептид по п.19, где пептиды являются посттрансляционно модифицированными. [0021] Модифицированный пептид по п.19, где пептиды подвергают стадии протеолитического гидролиза. [0022] Модифицированный пептид по п.19, где пептиды являются синтетическими. Полный текст патента Область техники, к которой относится настоящее изобретение Настоящее изобретение относится к выявлению злокачественных клеток, включающему применение модификаций ракового белка. Одним из наименее изученных и наиболее сложных болезненных процессов является трансформация, происходящая в процессе озлокачествления клетки. В этот процесс вовлекаются как генетические мутации, так и трансформации системы белкового обмена, вследствие чего клетка получает возможность выхода из-под нормального контроля, предотвращающего несоответствующее деление клеток. Раковые клетки имеют некоторые общие свойства. Пролиферация большинства раковых клеток происходит за рамками нормальных контролей клеточных циклов, у них появляются морфологические изменения и различные биохимические нарушения клеточных процессов. Рак, как правило, диагностируется, когда опухоль становится хорошо видимой после первого появления изменений в клетках. Многие раки диагностируются после гистологического исследования биоптата на предмет морфологических отклонений от нормы, признаков пролиферации клеток и генетических отклонений. Эффективное лечение злокачественного новообразования часто зависит от способности надежного выявления присутствия опухолевых клеток на ранних стадиях заболевания, с тем, чтобы эффективное лечение могло начинаться на стадии, когда заболевание является наиболее восприимчивым к такому лечению. Таким образом, необходимо уметь надежно выявлять потенциально злокачественную клетку, которая в своем развитии еще не достигла гистологической стадии, распознаваемой как раковая, но дальнейшее развитие которой может привести к состоянию злокачественности. Существует также необходимость в быстром, минимально инвазивном способе, с помощью которого можно было бы надежно выявлять или лечить злокачественные клетки или потенциально злокачественные клетки. Ядерный антиген пролиферирующих клеток (PCNA) представляет собой ядерный белок массой 29 кДа. Этот белок, благодаря экспрессии в клетке в S-фазе и G2-фазе клеточного цикла, оказывается хорошим маркером пролиферации клеток. Было показано также его участие во многих молекулярных путях, несущих ответственность за жизнь и смерть клетки. Периодическое появление этого белка в ядрах S-фазы позволяет предположить его вовлечение в процесс репликации ДНК. Позднее PCNA был идентифицирован как вспомогательный фактор ДНК-полимеразы в клетках млекопитающих и неотъемлемый фактор для репликации ДНК вируса SV40 (вакуолизирующий обезьяний вирус, ОВ-40) in vitro. В дополнение к своей роли в качестве скользящего белка-фиксатора ДНК и вспомогательного фактора ДНК-полимеразы в клетках млекопитающих, PCNA взаимодействует с рядом других белков, вовлеченных в транскрипцию, в стадии клеточного цикла, реконструкцию хроматина, рекомбинацию, апоптоз и другие формы репарации ДНК. Наряду с многоаспектностью действия, многие партнеры связывания PCNA объединяются своим вкладом в точный механизм наследования клеточных функций каждым новым поколением клеток. PCNA может выступать в роли ведущего фактора, координирующего процессинг хромосом. Известно также, что PCNA взаимодействует с другими факторами, например FEN-1 (флэп-эндонуклеаза-1), ДНК-лигазой и ДНК-метилтрансферазой. В дополнение к этому, было показано также, что PCNA играет основную роль в многочисленных путях репарации ДНК. Взаимодействие с белками, например белком распознавания ошибочного спаривания оснований, Msh2, и эндонуклеазой эксцизионной репарации ДНК, XPG (белок группы G xeroderma pigmentosa (пигментозная ксеродерма)), предполагает причастность PCNA к процессам, отличающимся от синтеза ДНК. В основе взаимодействия с многочисленными партнерами лежат, как правило, механизмы, позволяющие PCNA избирательно взаимодействовать упорядоченным и энергетически выгодным способом. Ключи к разгадке механизма функционирования PCNA были первоначально обнаружены посредством исследования ДНК-синтезомы, мультибелкового репликационного комплекса ДНК, присутствующего в клетках млекопитающих. В ходе исследований, посвященных изучению синтетической активности ДНК-синтезомы, была идентифицирована повышенная частота ошибок в злокачественных клетках по сравнению с незлокачественными клетками. Эти результаты позволяют предположить, что в злокачественных клетках произошло структурное изменение одного или нескольких компонентов ДНК-синтезомы. Анализ PCNA, неотъемлемого компонента ДНК-синтезомы, посредством двумерного электрофоретического блотирования из полиакриламидного геля, выявил две различные изоформы с совершенно разными изоэлектрическими точками (pI). Одна из изоформ PCNA демонстрировала существенно основную pI и присутствовала как в злокачественных, так и в незлокачественных клетках. Другая изоформа имела кислую pI и обнаруживалась исключительно в злокачественных клетках. Эта изоформа, благодаря ее присутствию лишь в злокачественных клетках, была названа раковой изоформой или csPCNA, и посттрансляционное изменение, несущее ответственность за измененный, по результатам 2D-PAGE (двумерный электрофорез в полиакриламидном геле), профиль перемещения PCNA, остается неопределенным. Данные некоторых исследований с мечением PCNA позволили предположить, что перемещение PCNA, вероятнее всего, не было обусловлено такими изменениями, например, как фосфорилирование, ацетилирование, гликозилированис или сиализирование. Был проведен ряд давших противоречивые результаты исследований, в ходе которых пытались идентифицировать пострансляционные модификации PCNA. Сообщалось, например, что фосфорилирование PCNA оказывает воздействие на связывание последнего с участками синтеза ДНК. В другом же исследовании утверждалось, что PCNA, в конечном счете, не фосфорилировался, а ацетилировался. В дополнение к этим исследованиям, анализ дрожжевого PCNA показал, что он является мишенью убиквитинирования в ответ на повреждение ДНК и сумоилирования при отсутствии повреждения. Вследствие разнородных и противоречивых сведений относительно структуры PCNA, трудно установить, какие модификации, если они присутствуют, несут ответственность за появление и функционирование csPCNA изоформы. Таким образом, для разработки диагностических методов, а также для создания терапевтических средств, основанных на взаимодействии csPCNA с его партнерами, желательной является идентификация соответствующих посттрансляционных модификаций csPCNA. Злокачественные раковые клетки экспрессируют изоформу PCNA, которую называют раковым ядерным антигеном пролиферирующих клеток (csPCNA), а незлокачественные клетки экспрессируют изоформу, которую называют незлокачественным ядерным антигеном пролиферирующих клеток (nmPCNA) Для диагностирования и лечения раков необходимы эффективные композиции и способы различения двух упомянутых изоформ. Идентифицируются новые посттрансляционные модификации csPCNA. Идентифицируется метилэстерификация csPCNA. Для анализа csPCNA изоформы и идентификации метилэстерификаций, присутствующих на csPCNA, прибегли к 2D-PAGE/жидкостной хроматографии-тандемной масс-спектрометрии (LC-MS/MS). Было определено местоположение метилэстерифицированных модификаций на конкретных остатках глутаминовой и аспарагиновой кислот. В приведенном описании раскрывают структурный анализ одной кислой изоформы PCNA (csPCNA), выделенной из клеток рака молочной железы, посредством двумерного электрофореза (2D-PAGE) и жидкостной хроматографии-тандемной масс-спектрометрии (LC-MS/MS). Метиловые сложные эфиры находятся на 16 определенных остатках глутаминовой и аспарагиновой кислоты csPCNA. Метилэстерификация csPCNA представляет собой новый тип посттрансляционной модификации в клетках млекопитающих, которая имеет отношение к определению некоторых из разнообразных функций PCNA. Способ выявления раковой изоформы ядерного антигена пролиферирующих клеток (csPCNA) в биологическом образце включает стадии выявления посттрансляционной модификации, включающей метиловый сложный эфир на одном или нескольких аминокислотных остатках csPCNA изоформы в образце, подозреваемом на содержание csPCNA изоформы; и определения присутствия csPCNA изоформы посредством сравнения уровней содержания метиловых сложных эфиров на csPCNA изоформе с незлокачественной изоформой PCNA. Некоторые биологические образцы включают биологические жидкости организма, например кровь, плазму, лимфу, сыворотку, плевральный выпот, цереброспинальную жидкость, слюну, мокроту, мочу, желудочный сок, сок поджелудочной железы, асцитическую жидкость, синовиальную жидкость, молоко и сперму. Биологический образец включает также образец ткани, например ткани, полученной из молочной железы, предстательной железы, легких, толстой кишки, эпителиальной ткани, соединительной ткани, ткани шейки матки, пищевода, головного мозга, вилочковой железы, щитовидной железы, поджелудочной железы, яичек, яичника, кишечника, мочевого пузыря, желудка, саркомы мягких тканей, остеосаркомы, лейкозной ткани, ткани лимфомы, карциномы, аденокарциномы, плаценты, волокнистой соединительной ткани, ткани половых клеток и их экстракты. Пригодным для анализа является любой биологический образец, который может содержать csPCNA. Согласно одному из аспектов, метиловый сложный эфир присутствует на csPCNA изоформе на аспарагиновой кислоте либо на глутаминовой кислоте или на их комбинации. Метиловый сложный эфир присутствует на csPCNA изоформе на одном или нескольких из 16 остатков аспарагиновой или глутаминовой кислот. Метиловый сложный эфир присутствует на одном или нескольких из 16 остатков аспарагиновой или глутаминовой кислот, соответствующих аминокислотным положениям 3, 85, 93, 94, 104, 109, 115, 120, 132, 143, 174, 189, 201, 238, 255 и 259 csPCNA изоформы. csPCNA пептиды, включающие 16 модифицированных остатков аспарагиновой или глутаминовой кислот, выглядят следующим образом: где E m представляет метилэстерифицированный остаток глутаминовой кислоты, a D m представляет метилэстерифицированный остаток аспарагиновой кислоты. Согласно одному из аспектов выявление csPCNA изоформы осуществляют посредством масс-спектрометрического анализа, например, посредством жидкостного хроматографического (LC)-масс-спектрометрического (MS) анализа. Применим любой приемлемый способ выявления метиловых сложных эфиров либо метилэстерифицированных аминокислотных остатков. Следствием масс-спектрометрического анализа csPCNA пептида является выявление 14 Да смещения массы по сравнению с соответствующим немодифицированным пептидом. Способ диагностирования либо прогнозирования злокачественного новообразования, где упомянутый способ включает стадии выявления раковой изоформы ядерного антигена пролиферирующих клеток (csPCNA) в биологическом образце посредством идентифицирования статуса посттрансляционной модификации csPCNA изоформы, который отличает csPCNA изоформу от незлокачественной изоформы PCNA (nmPCNA); и диагностирования злокачественного новообразования на основе выявления csPCNA в биологическом образце. Для выявления злокачественного новообразования применяют посттрансляционную модификацию, например, метилэстерификацию, которую выявляют на csPCNA, и уровни метилэстерификации csPCNA по сравнению с nmPCNA. Модифицированный пептид ядерного антигена пролиферирующих клеток (PCNA) содержит аминокислотную последовательность, выбранную из числа нижеприведенных: где E m представляет метилэстерифицированный остаток глутаминовой кислоты, a D m представляет метилэстерифицированный остаток аспарагиновой кислоты. Согласно одному из аспектов модифицированные пептиды csPCNA или PCNA подвергаются посттрансляционной модификации. Согласно другому аспекту модифицированные пептиды csPCNA или PCNA подвергаются стадии расщепления протеазами. Согласно другому аспекту модифицированные пептиды csPCNA или nmPCNA являются синтетическими. Согласно одному из аспектов способ выявления раковой изоформы ядерного антигена пролиферирующих клеток (csPCNA) в биологическом образце включает определение общих уровней метилэстерификации csPCNA и nmPCNA, а также определение, на основе уровней метилэстерификации, того, что биологический образец содержит csPCNA. Дополнительные отличительные особенности настоящего изобретения будут очевидны специалистам в данной области техники после рассмотрения приведенного ниже подробного описания вариантов осуществления, представляющих в качестве примера наилучший способ осуществления предмета настоящего изобретения в том виде, в котором он представляется на современном этапе. Фиг. 1 демонстрирует идентификацию csPCNA после проведения 2D-PAGE посредством определения характеристик пептида с помощью LC-MS/MS. (А) представляет репрезентативный окрашеный кумасси синим 2D-PAGE гель ядерного экстракта клеток линии MDA MB 468 (2 мг). Для триптического протеолиза были выбраны многочисленные пятна, идентифицированные посредством LC-MS/MS, csPCNA идентифицирована (стрелка черного цвета) на кислой стороне геля с приблизительной pI 4,6. При объемном представлении 2D-PAGE выявляются два пятна, соответствующие csPCNA, пятно присутствия в умеренном количестве (стрелка черного цвета) и более кислое пятно присутствия в незначительном количестве (стрелка белого цвета), выявленные с помощью программы двумерного анализа. Пятна (i-v) белков, отличающихся от PCNA, которые были идентифицированы в процессе анализа, представлены в табл. I. (В) представляет хроматограмму основных пиков пептидов, полученных посредством триптического расщепления csPCNA. (С) представляет масс-спектр csPCNA пептида, который элюируется на 43,3 мин. Анализ тандемного масс-спектра иона с отношением массы к заряду (m/z) 1038,7 (D) идентифицирует его как двухзарядный пептид, соответствующий остаткам 92-110 человеческого PCNA. (E) - масс-спектр, представляющий элюирование 1045,4 m/z иона на 43,74 мин. Разность масс однозарядных ионов вдвое превосходит разность двухзарядного иона и 1045,4 m/z иона, демонстрируя, тем самым, +14 Да смещение массы. (F) показывает, что тандемный масс-спектр 1045,4 m/z иона почти идентичен тандемному масс-спектру в части D, за исключением того, что все y-ионы смещены на 14 Да. Смещение массы только b 18 -иона определяет местоположение дополнительных 14 Да на глутаминовой кислоте в положении 109 csPCNA. Фиг. 2 показывает создание карт смещенных пептидных последовательностей csPCNA. (А) Репрезентативные хроматограммы основных пиков, показывающие элюирование пептидов, полученных посредством расщепления csPCNA трипсином (синий цвет), трипсином/CNBr (зеленый цвет), GluC (красный цвет) и AspN (черный цвет). (В) Карты последовательностей csPCNA, идентифицированные посредством LC-MS/MS. Обозначены места расположения метиловых сложных эфиров (X m ). Фиг. 3 показывает, что С-конечный хвост PCNA метилэстерифицирован на многих участках. (А) Элюирование трех различных пептидов, полученных с С-конечного хвоста csPCNA. Хроматограмма основных пиков csPCNA, расщепленного трипсином, в сравнении с хроматограммами отдельных ионов пептида IEDEEGS (778,5 m/z) и метилэстерифицированных пептидов IEDEEGS (792,5). Метилэстерифицированные пептиды обладают повышенной гидрофобностью и в обратнофазовом градиенте элюируются позднее. (В) Истолкование тандемных масс-спектров немодифицированного пептида показывает фрагментацию однозарядного пептида с последовательностью IEDEEGS. Нейтральная потеря воды (-18 кДа) наблюдается, как правило, со многими фрагментными ионами. В дополнение к этому, при 373,9 m/z наблюдается фрагментный ион, представляющий нейтральную потерю воды и СО-группы из y 4 -иона. (С) Тандемный масс-спектр 792,3 m/z иона из пептида, первым элюированого на вышеприведенной хроматограмме. В спектре могут быть идентифицированы b 3-6 -ионы, все со смещением на 14 Да, но y 3 и y 5 -ионы не смещены, что позволяет предположить, что модификация находится на лейцине либо на глутаминовой кислоте N-конца. (D) Тандемный масс-спектр 792,3 m/z иона из пептида, элюированного вторым при разделении. b 5 и b 6 -ионы содержат 14 Да смещение, подобное упомянутому выше, в то время как b 3 и b 4 -ионы подобного смещения не содержат, что соответствует нахождению метилового сложного эфира в пептиде на глутаминовой кислоте в положении 5. На фиг. 4 представлен иммуноблот PCNA. На фиг. 5 представлена аминокислотная последовательность PCNA (1-262 аминокислоты). Белок ядерного антигена пролиферирующих клеток (PCNA) в раковых клетках изменяется. PCNA представляет собой белок, имеющий молекулярную массу 28 кДа, с электрофоретической подвижностью, эквивалентной электрофоретической подвижности белка массой 36 кДа. PCNA является вспомогательным фактором, необходимым для ДНК-полимеразы δ для опосредования высокоэффективной репликационной активности ДНК. ДНК-синтезома, очищенная из клетки злокачественного новообразования, содержит по крайней мере две формы PCNA. Две разновидности PCNA существенно различаются по общему заряду. Так, на двумерном полиакриламидном геле можно различить кислую, злокачественную или раковую форму PCNA, csPCNA, и основную, незлокачественную или нормальную форму PCNA, nmPCNA. Кислая форма PCNA (csPCNA) экспрессируется в линиях клеток злокачественных новообразований, например HeLa (человеческий рак шейки матки), Hs578T (рак молочной железы), HL-60 (человеческий промиелолейкоз), FM3A (рак молочной железы мышей), PC 10 (рак предстательной железы), LNCaP (рак предстательной железы), LN99 (рак предстательной железы), MD-MB468 (рак молочной железы людей), MCF-7 (рак молочной железы), KGE 90 (рак пищевода-толстой кишки), KYE 350 (рак пищевода-толстой кишки), SW 48 (рак пищевода-толстой кишки) и Т 98 (злокачественная глиома). Кислая форма PCNA (csPCNA) также экспрессируется в клетках злокачественных новообразований, полученных из опухолей молочной железы людей, опухолей предстательной железы, опухолей головного мозга, опухолей желудочно-кишечного тракта и пищевода-толстой кишки людей, опухолей молочной железы мышей и при хроническом миелолейкозе людей. Кислая форма PCNA (csPCNA) не выявляется в линиях незлокачественных клеток, например линии клеток молочной железы Hs578Bst и MCF-10A, или в образцах незлокачественной сыворотки или ткани, например молочной железы. Вариант определения характеристик пептида посредством LC-MS/MS применили для секвенирования csPCNA изоформы, обнаруженной в злокачественных клетках. Была идентифицирована посттрансляционная модификация нового типа, присутствующая на многочисленных остатках csPCNA. Эта модификация, метилэстерификация, присутствовала в csPCNA на 16 различных остатках аспарагиновой и глутаминовой кислот. Немодифицированная аминокислотная последовательность PCNA представлена на фиг. 5. Эти метиловые сложные эфиры первоначально были идентифицированы как 14 Да смещения массы пептида и их местоположение, с помощью тандемной масс-спектрометрии, было определено на остатках глутаминовой либо аспарагиновой кислот. Относительное количественное определение метилэстерифицированных пептидов показало, что csPCNA белки в злокачественных клетках включают несколько молекул, содержащих один либо несколько метиловых сложных эфиров, которые находятся на многочисленных остатках в упомянутом белке. Следствием метилэстерификации определенных остатков являются, вероятно, дискретные конформационные изменения белка и эти изменения могут усиливать и/или разрушать белок-белковые взаимодействия. Влияние метилэстерификации на функции белков млекопитающих плохо изучено. Многие прошлые исследования метилэстерификации белков млекопитающих сосредоточивались на старении белка и репарации остатков изоаспартила посредством фермента белок-изоаспартат-метилтрансферазы (PIMT). Большая часть метиловых сложных эфиров, присутствующих на csPCNA, обнаруживается, однако, на остатках глутаминовой кислоты, а не на остатках аспарагиновой кислоты, что позволяет выдвинуть предположение о том, что модификация осуществляется альтернативным путем. Возможно, что метилэстерификация PCNA изменяет его конформацию и фактически скрывает и/или открывает определенные места связывания белка и определяет его функцию. Был также осуществлен анализ последовательностей рекомбинантного PCNA посредством LC-MS/MS и было обнаружено свидетельство метилэстерификации. Метилэстерификация, обнаруженная на PCNA, может, таким образом, стабилизировать специфический конформационный статус в противном случае неупорядоченного белка. В дополнение к этому, вычисление электростатического потенциала PCNA показывает, что наружная поверхность тримера PCNA имеет сильный отрицательный потенциал и избыток остатков глутаминовой и аспарагиновой кислот. Метилирование этих остатков могло бы, таким образом, изменить этот потенциал и фактически изменить топологию поверхности упомянутого белка. Раскрывается определение характеристик пептида специфической изоформы, csPCNA, отделенной с помощью 2D-PAGE, посредством LC-MS/MS. Применяя комбинацию протеолитических подходов, были получены карты смещенных пептидных последовательностей csPCNA, идентифицирующие 100% последовательность csPCNA белка посредством LC-MS/MS. Была идентифицирована новая посттрансляционная модификация, присутствующая на csPCNA. Метиловые сложные эфиры в csPCNA изоформе появились на по крайней мере одном из 16 специфических остатков аспарагиновой и/или глутаминовой кислот. Эти модификации могут изменять структуру тримера PCNA и, вследствие этого, усиливать конформационные изменения, которые могут относиться к упорядоченным белок-белковым взаимодействиям. Биологическим образцом может быть образец биологической жидкости организма, который может включать кровь, плазму, лимфу, сыворотку, плевральный выпот, цереброспинальную жидкость, слюну, мокроту, мочу, сперму, слезы, синовиальную жидкость или же любую биологическую жидкость организма, которая может проверяться на присутствие csPCNA изоформы. Согласно альтернативному варианту биологическим образцом может быть образец ткани, где клетки упомянутого образца ткани могут подозреваться в злокачественности. Например, срезы ткани либо клеточные культуры могут наноситься на стеклянные или пластиковые предметные стекла и контактировать с антителами согласно стандартным иммуноцитохимическим методикам. Пригодными являются также тканевые экстракты либо концентраты клеток или клеточные экстракты. Согласно другому варианту осуществления предлагается способ диагностирования злокачественного новообразования. Упомянутый способ включает стадию выявления csPCNA в биологическом образце, полученном от человека или, в частности, от больного, подозреваемого на наличие злокачественного заболевания, где стадия выявления csPCNA включает выявление уровней посттрансляционной модификации, включающей раскрытые в настоящем описании метиловые сложные эфиры. Согласно другому варианту осуществления предлагается способ, помогающий при диагностировании злокачественного новообразования. Упомянутый способ включает стадию выявления посттрансляционных модификаций csPCNA в злокачественных клетках образца ткани, в сравнении с PCNA в нормальных клетках, где клетки образца ткани подозреваются на наличие злокачественности и где стадия выявления csPCNA включает выявление метиловых сложных эфиров на csPCNA. Следует понимать, что злокачественные клетки не ограничиваются злокачественными клетками в тканях, например, молочной железы, простаты, крови, головного мозга, поджелудочной железы, гладкой или поперечно-полосатой мышцы, печени, селезенки, вилочковой железы, легких, яичника, кожи, сердца, соединительной ткани, почек, мочевого пузыря, кишечника, желудка, надпочечника, лимфатического узла либо шейки матки или в линиях клеток, например, Hs578T, MCF-7, MDA-MB468, HeLa, HL60, FM3A, ВТ-474, MDA-MB-453, T98, LNCaP, LN 99, PC 10, SK-OV-3, MKN-7, KGE 90, KYE 350 или SW 48. Согласно другому варианту осуществления предлагается способ, помогающий при прогнозировании развития злокачественного новообразования. Упомянутый способ включает выявление csPCNA в образце ткани посредством выявления раскрываемых в настоящем описании посттрансляционных модификаций, где клетки образца ткани могут подозреваться на злокачественность, и соотнесение уровней csPCNA с развитием конкретного злокачественного заболевания. В дополнение к этому выявление и анализ посттрансляционных модификаций csPCNA может применяться для прогнозирования результатов потенциального выживания больного, у которого происходит развитие злокачественного новообразования. Следует понимать, что к числу заболеваний, которые могут диагностироваться либо прогнозироваться с помощью упомянутых антител, относятся (но без ограничения) такие злокачественные новообразования, как различные формы глиобластомы, глиомы, астроцитомы, менингиомы, нейробластомы, ретинобластомы, меланомы, рак толстой кишки, рак легких, аденокарциномы, рак шейки матки, рак яичников, рак мочевого пузыря, лимфобластомы, лейкоз, остеосаркомы, рак молочной железы, гепатомы, нефромы, рак надпочечника, рак предстательной железы, рак пищевода. Если раковая клетка экспрессирует csPCNA изоформу, раскрытые в настоящем описании методики способны выявить упомянутую csPCNA изоформу. Способы выявления, включающие выявление раскрываемых в настоящем описании посттрансляционных модификаций, могут также выявлять злокачественность некоторых инвазивных и неинвазивных типов опухолей в ткани молочной железы, к числу которых относятся киста протоков, изменение эпителия ацинуса молочной железы, склеротическое поражение лимфатических узлов, гиперплазия эпителия протоков, неатипичный внутрипротоковый папилломатоз, изменения столбчатых эпителиоцитов, радиальное склеротизирующее поражение (радиальный рубец), аденома сосков молочной железы, внутрипротоковая папиллома, фиброаденома, лактационная папиллома, атипичная гиперплазия эпителия протоков, атипичная гиперплазия долек молочной железы, внутриэпителиальный рак протоков, подразделяемый на ядерные подклассы 1, 2 и 3, внутриэпителиальный рак долек молочной железы, плеоморфный внутриэпителиальный рак долек молочной железы, интрамаммарная липома, гамартома молочной железы, зернистоклеточная миобластома, интрамаммарный жировой некроз, псевдоангиоматозная гиперплазия стромы (PASH), злокачественная меланома, охватывающая молочную железу, злокачественная лимфома, охватывающая молочную железу, листовидная опухоль доброкачественного, пограничного и злокачественного подклассов и саркома молочной железы. Согласно другому варианту осуществления, раскрываемые в настоящем описании способы применяют для определения стадии злокачественности опухолей посредством сравнения уровней csPCNA в опухоли с течением времени для отслеживания развития ракового заболевания либо реакции больного на лечение. Упомянутые способы могут использоваться также для выявления злокачественных клеток, высвободившихся из опухоли и присутствующих в кровотоке больного, посредством использования упомянутых антител для анализа пробы крови на присутствие csPCNA изоформы. Биологический образец может быть получен от больного человека либо больного животного. Термин "антитело" включает моноклональные антитела (в том числе непроцессированные моноклональные антитела), поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител до тех пор, пока они проявляют желаемую биологическую активность либо специфичность. Могут быть получены антитела, специфически распознающие посттрансляционно модифицированную nmPCNA или csPCNA. Термин "модифицированный белок или пептид" означает присутствие одной или нескольких посттрансляционных модификаций, присутствующих на csPCNA либо nmPCNA белках или пептидах, полученных из этих белков. Упомянутый термин относится также к синтетическим, выделенным и очищенным пептидам csPCNA и nmPCNA, которые содержат одну либо несколько посттрансляционных модификаций. Упомянутый термин относится также к протеолитически расщепленным пептидам csPCNA и PCNA, и пептидам csPCNA и PCNA, фрагментированным любыми известными способами, которые содержат одну либо несколько посттрансляционных модификаций. Все примененные химикаты были получены от компании Sigma-Aldrich (St. Louis, штат Миссури) либо компании Fisher Scientific (Hampton, штат Нью-Гемпшир), если не указано иное. Вода спектрометрической чистоты и ацетонитрил были получены от компании Honeywell Burdick and Jackson (Morristown, штат Нью-Джерси). csPCNA и nmPCNA, присутствующие в образцах, полученных от людей, анализируют на наличие посттрансляционных модификаций с помощью любого стандартного метода. Пригодным методом, например, является масс-спектрометрический анализ. Масс-спектрометрический анализ может сочетаться с другими методами. Клетки рака молочной железы линий MDA MB 468 и MCF 7 были получены из АТСС (Американская коллекция типовых культур) и сохранялись на DMEM (модифицированная по способу Дульбекко среда Игла) (компания MediaTech, Herndon, штат Вирджиния), содержащей 10% FCS (фетальная телячья сыворотка) (компания BioWhittaker, Walkersville, штат Мериленд), антибиотики и противогрибковые препараты (компания Invitrogen, Carlsbad, штат Калифорния). Клетки выращивали на 100-мм чашках для культивирования культуры клеток до 60-70% слияния, промывали и соскребали резиновым скребком в имеющий температуру тающего льда PBS (забуференный фосфатом физиологический раствор) перед осаждением центрифугированием. Клетки фракционировали до получения ядерного экстракта, как описано у Малкас (Malkas) и другие, Biochemistry, 1990, 29, 6362-6374. Вкратце, клетки гомогенизировали с помощью гомогенизатора Dounce ™ с последующим осаждением ядер центрифугированием. После этого ядерные белки экстрагировали 150 мМ раствором KCl в течение 2 ч при температуре 4 °С и мембраны осаждали ультрацентрифугированием. Ядерные экстракты сохраняли при температуре -80 °С до применения. Изоэлектрическое фокусирование осуществляли с помощью IEF кюветы (кювета для изоэлсктрического фокусирования) и 17 см полосок с иммобилизованным градиентом рН (Ready Strip IPG strips) (компания Bio-Rad, Hercules, штат Калифорния) при рН 4-7. Образцы белка обессоливали с помощью колонок с вращающейся лентой для обессоливания белка (компания Pierce, Rockford, штат Иллинойс) и лиофилизировали на скоростной вакуумной сушилке (компания ATR Biotech, Laurel, штат Мериленд). Лиофилизированные образцы ресуспендировали в буфере для повторной гидратации (9 М раствор мочевины, 4% раствор CHAPS (3-[(3-хлорамидопропил)диметиламмонио]-1-пропансульфонат), 0,2% раствор Bio-Lytes (компания Bio-Rad, Hercules, штат Калифорния), 2 мМ раствор трибутилфосфина, 0,001% раствор бромфенолового синего) и подвергали пассивной повторной гидратации на IPG полосках в течение 12 ч при температуре 20 °С. Изоэлектрическое фокусирование осуществляли в соответствии с инструкциями производителя (компания Bio-Rad, Hercules, штат Калифорния). Перед проведением SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия), IPG полоски восстанавливали посредством инкубирования в буфере (6 М раствор мочевины, 0,375 М раствор трис-буфера, 2% раствор SDS (додецилсульфат натрия), 20% раствор глицерина, рН 8,8), содержащем 20 мг/мл DTT (дитиотреитол) и алкилировали в том же самом буфере, содержащем йодоацетамид вместо DTT. SDS-PAGE проводили на 12% полиакриламидных гелях (20 см × 20 см) с помощью аппарата Protean XL (компания Bio-Rad, Hercules, штат Калифорния) с постоянным током в течение приблизительно 5 ч. Гели фиксировали 10% раствором уксусной кислоты/50% раствором метанола в воде и окрашивали Gel Code Blue (компания Pierce, Rockford, штат Иллинойс) в течение ночи. Визуализацию осуществляли с помощью сканирующего денситометра GS710 (компания Bio-Rad, Hercules, штат Калифорния) и анализ гелей осуществляли с помощью программы Phoretix Evolution 2D (компания NonLinear Dynamics, Inc., Durham, штат Северная Каролина). Отбор пятен осуществляли вручную с помощью 1 мм устройства для отбора столбиков геля (компания The Gel Company, San Francisco, штат Калифорния). Расщепление белков осуществляли с помощью трипсина (компания Promega, Madison, штат Висконсин), бромциана (CNBr) (компания Sigma-Aldrich, St. Louis, штат Миссури), GluC либо AspN (компания Roche, Indianapolis, штат Индиана), как описывалось ранее с некоторыми модификациями (Розенфельд (Rosenfeld) и другие, Anal. Biochem., 1992, 203, 173-179; фон Монфор (van Montfort) и другие, Biochim. Biophys. Acta, 2002 1555, 111-115). Вкратце, пятна на геле обесцвечивали 25 мМ раствором бикарбоната аммония (АВС)/50% раствором ацетонитрила, с последующей дегидратацией 100% раствором ацетонитрила и высушиванием в скоростной вакуумной сушилке (компания ATR Biotech, Laurel, штат Мериленд). После этого пятна повторно гидратировали в растворе протеазы (10 мкг/мл трипсина, GluC или AspN) в 25 мМ растворе ABC. Через 10 мин при температуре 4 °С избыточный раствор протеазы удаляли, заменяли свежим 25 мМ раствором ABC и инкубировали в течение ночи при температуре 37 °С. Пептиды экстрагировали из геля посредством озвучивания ультразвуком в присутствии 25 мМ раствора АВС/50% раствора ацетонитрила (один раз) и 5% раствора муравьиной кислоты/50% раствора ацетонитрила (дважды). Пептидные экстракты смешивали и сушили в скоростной вакуумной сушилке. Последующее расщепление триптических гидролизатов (отобранных образцов) с помощью CNBr осуществляли посредством ресуспендирования высушенных пептидных экстрактов в 70% растворе муравьиной кислоты с добавлением CNBr до приблизительно 200 М избытка относительно пептида. Расщепление с помощью CNBr осуществляли при комнатной температуре в течение 4 ч с высушиванием в скоростной вакуумной сушилке. Все образцы ресуспендировали в 1% растворе муравьиной кислоты непосредственно перед проведением анализа. Высокоэффективное жидкостное хроматографирование (HPLC) нанопотоков осуществляли с помощью улавливающей колонки IntegraFrit (компания New Objective, Inc., Woburn, штат Массачусетс), заполненной в лаборатории материалом Magic C18Aq (5 мкм, 200 Å) (компания Michrom, Inc., Auburn, штат Калифорния) и 0,05 мм × 100 мм колонки из кварцевого стекла с оттянутым наконечником (компания PolymicroTechnologies, Phoenix, штат Аризона), заполненной в лаборатории материалом Magic C18Aq (5 мкм, 100 Å) и установленной на микрокрестовине, удерживаемой на месте с помощью изготовленной на заказ платформы для нанораспыливания (Гатлин (Gatlin) и другие, Anal. Biochem., 1998, 263, 93-101). Пептиды отделяли с помощью 2-50% линейного градиента ацетонитрила, содержащего 0,1% раствор муравьиной кислоты. Контрольно-измерительная аппаратура включала либо Surveyor HPLC и масс-спектрометр с ионной ловушкой LCQ Advantage (компания ThermoElectron, Waltham, штат Массачусетс), либо Ultimate HPLC (LC Packings) и LCQ DECA XP (компания ThermoElectron, Waltham, штат Массачусетс). Таблицы пиков получали с использованием необработанных данных с помощью программы BioWorks 3.1 (компания ThermoElectron, Waltham, штат Массачусетс). Поиск в базе данных Swissprot проводили с помощью механизма поиска Mascot (компания Matrix Science, Inc., Boston, штат Массачусетс). (Смотри также Кризи (Creasy), Коттрелл (Cottrell), Proteomics, 2002, 2, 1426-1434). Принимали 3 Да и 0,8 Да допуск на ион-предшественник и массу осколков, соответственно, и до двух упущенных сайтов расщепления. В качестве изменяющихся модификаций при первом просмотровом поиске в базе данных млекопитающих были выбраны карбамидометилцистеины и оксидированные метионины. В ходе последующего поиска с допуском на ошибку, трипсин был выбран в качестве фермента, а метиловые сложные эфиры глутамата и аспартата дополнительно принимали во внимание в качестве изменяющихся модификаций. Несмотря на то что в подробном описании и в приведенных ниже Примерах подробно, со ссылкой на конкретные варианты осуществления изобретения, описывается выявление посттрансляционных модификаций и их применение в отношении csPCNA изоформы, специалисту в данной области техники будет очевидна возможность внесения различных изменений и модификаций без отступления от сущности и объема изобретения. Все ссылки, приведенные в описании, включены в него в полном объеме в качестве ссылок. Следующие ниже примеры приведены лишь с иллюстративной целью и не предназначены для ограничения изобретения, описание которого в общих терминах было приведено выше. Пример 1. Двумерный (2d) имуноэлектрофорез в полиакриламидном геле и определение характеристик пептида csPCNA изоформы. Фракцию ядерного экстракта, выделенную из клеток рака молочной железы линии MDA MB 468, разделяли посредством 2D-PAGE (фиг. 1А). Многочисленные пятна, имеющие кажущуюся молекулярную массу около 36 кДа и изоэлектрические точки, равные или приближающиеся к 4,5 (кажущаяся молекулярная масса PCNA по результатам SDS-PAGE и вычисленная изоэлектрическая точка (pI)), вырезали из геля и подвергали трипсинизации без удаления геля. Полученные пептиды подвергали анализу посредством жидкостного хроматографирования (LC) нанопотоков и тандемной масс-спектрометрии с электрораспылением (LC-MS/MS) с помощью квадрупольного масс-спектрометра с ионной ловушкой. Белки, содержащиеся в каждом пятне, идентифицировали посредством поиска в базе тандемных масс-спектрометрических данных с помощью механизма поиска Mascot. Посредством подобного подхода было определено местоположение csPCNA на 2D-PAGE гелях. Этот подход обеспечил возможность проведения стандартной идентификации и анализа csPCNA с ограниченной манипуляцией образцами, что сводило до минимума потенциальную угрозу утраты посттрансляционных модификаций. На фиг. 1В представлена хроматограмма репрезентативных LC/MS основных пиков пептидов, полученных посредством трипсинизации без удаления геля пятен, содержащих csPCNA, вырезанных из 2D-PAGE гелей. В этом эксперименте были идентифицированы 25 пептидов, покрывающих 97% последовательности непроцессированного PCNA. Репрезентативный csPCNA пептид, идентифицированный в этом эксперименте, показан на фиг. 1С. Этот пептид элюировался на 43,3 мин и демонстрировал m/z 1038,7 (2+) (фиг. 1С). Индуцированная столкновениями диссоциация (CID) упомянутого пептида (Репсторф П. (Rocpstorff P.), Фольман Дж. (Fohlman J.), Biomecl. Mass Spectrom., 1984, 11, 601) выявила последовательность AEDNADTLALVFEAPNQEK, аминокислоты 92-110 человеческого PCNA (фиг. 1D). Пример 2. Метилэстерификация csPCNA. Этот пример демонстрирует, что метилэстерификация csPCNA изоформы присутствует на одной либо нескольких аминокислотах. LC-MS/MS данные идентифицировали многочисленные пептиды, демонстрирующие 14 Да смещение массы исходного пептида. Интересно отметить, что идентифицированные смещения массы позволяют предположить присутствие посттрансляционной модификации - метилэстерификации. Метилирование клеток млекопитающих рассматривается, как правило, как необратимая посттрансляционная модификация. Происходящее, главным образом, на аминогруппах остатков лизина и N-концах белков, метилирование идентифицировалось на таких белках, как р53 и гистоны Н3 и Н4. Проверка CID спектров на наличие пептидов с +14 Да смещением выявила нахождение метильных групп на остатках аспарагиновой и глутаминовой кислот, что соответствует метилэстерификации. Фиг. 1E показывает элюирование метилэстерифицированного пептида (1045,4 m/z [2+]) в этом эксперименте на 43,8 мин. Этот пептид элюировался вскоре после (через 25 с) немодифицированного 1038,7 m/z (2+) пептида (фиг. 1С), который все еще наблюдается в MS спектре. Фрагментирование 1045,4 m/z иона (фиг. 1F) выявило картину, весьма подобную 1038,7 m/z пептиду (фиг. 1D). Основным различием в этих фрагментационных спектрах является +14 смещение массы почти у всех ионов серии у 1045,4 m/z пептида, по сравнению с ионами серии у 1038,7 m/z пептида. При проверке высокомолекулярного участка 1045,4 CID спектра (фиг. 1F) идентифицировали два b-иона со значениями m/z 1943,3 и 1800,2. Упомянутый 1943,3 ион соответствует b 18 -иону и указывает на утрату немодифицированного лизина на С-конце. Этот ион также отличается от b 18 -иона (1928,4), наблюдавшегося в фрагментационном спектре 1038,7 m/z пептида (фиг. 1D), на 14 Да, что позволяет сделать дополнительное предположение о том, что упомянутый лизин немодифицирован и 14 Да смещение массы находится на другом остатке в упомянутом пептиде. b 17 -Ион (1800,2) на фиг. 1F, с другой стороны, близко напоминает b 17 -ион (1799,4) на фиг. 1D, указывая на утрату глутаминовой кислоты (129 Да) плюс метильной группы (14 Да) или на разницу в 143 Да между b 17 и b 18 -ионами 1045,4 m/z пептида. Небольшие пики сразу же слева от b 18 -ионов на фиг. 1D и фиг. 1F имеют значения m/z 1911,1 и 1925,4 соответственно, что соответствует нейтральной утрате b 18 -ионами Н 2 О либо NH 3 . Эти наблюдения, в сочетании со смещенными массами y-ионов, определяют местоположение метилового сложного эфира на остатке глутаминовой кислоты в положении 109 в непроцессированном csPCNA белке. Наряду с csPCNA, на присутствие метилэстерификации анализировались другие белки в геле. На фиг. 1А показано пять репрезентативных белковых пятен, а в табл. I представлена природа этих пятен и их метилэстерифицированный статус. Интересно отметить, что, как оказалось, один из пяти анализированных белков (статмин) был метилэстерифицирован, а также ацетилирован на N-конце. Метилэстерификация csPCNA не была результатом манипулирования образцами (например, фиксации геля уксусной кислотой и метанолом), поскольку метилэстерификация наблюдалась не на всех белках, присутствующих в геле, а лишь на небольшой подгруппе белков и постоянно на определенных остатках csPCNA. Пример 3. Идентификация метилэстерификации csPCNA. Наряду с AEDNADTLALVFEAPNQEK пептидом, идентифицированным на фиг. 1, 14 Да смещение массы постоянно демонстрировали многие другие csPCNA пептиды. Для успешной идентификации всех участков метилэстерификации, а также идентификации любых других потенциальных посттрансляционных модификаций, присутствующих на csPCNA изоформе, с помощью различных реактивов были получены смещенные пептиды, как показано на картах последовательностей csPCNA изоформы. Фиг. 2А показывает репрезентативные хроматограммы основных пиков LC-MS/MS экспериментов, полученные при раздельном расщеплении csPCNA только трипсином, трипсином с последующей обработкой бромцианом (CNBr), только AspN либо только GluC. Эти кривые основных пиков показывают различия профилей элюирования пептидов, полученных с помощью этих методов. Расщепление только трипсином, а также трипсином и CNBr, обеспечивало покрытие последовательности, а эксперименты с AspN и GluC обеспечили дополнительные подтверждающие данные. Расщепление с помощью CNBr, который обрезает С-конец до остатков метионина, уменьшало величину более крупных триптических пептидов, благодаря чему эти последовательности поддавались LC-MS/MS анализу с применением ионной ловушки. Это комбинированное расщепление было пригодно для определения характеристик триптического пептида DL1NEACWDI ×GVNLQSMDSSHVSLVQLTLR, состоящего из 33 остатков (остатки 21-53 PCNA). Расщепление с помощью AspN и GluC позволило получить подтверждающие данные, хотя оно и не обеспечивало полного покрытия последовательности. Это, вероятнее всего, обусловлено многими факторами. Например, GluC и AspN имеют меньшее количество сайтов расщепления на PCNA, по сравнению с трипсином, и, следовательно, образуют более крупные пептиды, которые труднее поддаются анализу. В дополнение к этому, эти протеазы распознают в белках остатки аспарагиновой и глутаминовой кислот, те самые остатки, которые, как было установлено в этом исследовании, подвергаются метилэстерификации. Следовательно, метилэстерификация могла бы потенциально влиять на расщепление протеазами с образованием более крупных пептидов и, в конечном счете, вести к худшему покрытию последовательности. Этот недостаток протеазного расщепления может применяться в качестве диагностического инструмента для выявления метилэстерификации. Пептиды, полученные с протонированным С-конечным остатком, например, триптические пептиды, дают, как правило, CID спектры хорошего качества. Комбинированные данные, полученные в результате многочисленных экспериментов по секвенированию с применением различных упомянутых подходов, постоянно идентифицировали 16 метилэстерифицированных остатков глутаминовой и аспарагиновой кислот на csPCNA из клеток рака молочной железы линий MDA MB468 и MCF7 (смотри табл. III). Полученные карты последовательностей csPCNA представлены на фиг. 2В вместе с любыми модификациями, идентифицированными LC-MS/MS. Наряду с метилэстерификацией, наблюдалось несколько других модификаций. Во всех экспериментах на csPCNA и других белках постоянно наблюдались оксидированные метионины и карбамидометилцистеины. В дополнение к этому, на продуктах гидролиза трипсином/CNBr наблюдались многочисленные "формилированные" серины, треонины и тирозины. Однако эти модификации, вероятнее всего, представляют собой химические модификации, являющиеся следствием манипулирования образцами и не являются "нативными" посттрансляционными модификациями. Пример 3А. Идентификация других посттрансляционных модификаций на PCNA и csPCNA. В дополнение к идентифицированию метиловых сложных эфиров на csPCNA, молекулу csPCNA в целом анализировали на присутствие других известных посттрансляционных модификаций. В отличие от предшествующих сообщений по другим формам PCNA, на csPCNA не идентифицировали убиквитинирования, сумоилирования, фосфорилирования или ацетилирования. Убиквитинирование и сумоилирование привело бы к значительному смещению массы PCNA на 2D-PAGE гелях, что не наблюдается в случае csPCNA изоформы. Кроме того, ни в одном LC-MS/MS эксперименте с csPCNA не были обнаружены пептиды, соответствующие убиквитинированию либо сумоилированию и ни один csPCNA пептид не демонстрировал смещения массы, соответствующего связыванию убиквитина либо сумоила, что достаточно убедительно указывает на то, что csPCNA в анализированных пятнах геля не были ни убиквитинированы, ни сумоилированы. И несмотря на то, что ранее сообщалось о фосфорилировании PCNA, в ходе этих анализов не было получено никаких данных, свидетельствующих о фосфорилировании каких-либо остатков csPCNA. Ни один из csPCNA пептидов, наблюдавшийся в этих анализах, не продемонстрировал +80 Да смещения массы и/или нейтральной потери Н 3 РО 4 (98 Да), что соответствует сообщениям, показывающим, что PCNA ацетилируется, а не фосфорилируется. В дополнение к этому, гели подвергали иммуноблоттированию на присутствие фосфосерина, фосфотреонина, фосфотирозина, ацетилированного лизина, а также поли(ADP)рибозы. Эти посттрансляционные модификации на PCNA выявлены не были. csPCNA изоформа в клетках рака молочной железы не фосфорилирована, ацетилирована, убиквитинирована либо сумоилирована, но вместо этого метилэстерифицирована. Подобно ацетилированию и фосфорилированию, метилэстерификация может изменять ее перемещение при проведении 2D-PAGE. Однако в отличие от ацетилирования и фосфорилирования, которые могли бы сместить молекулу к более кислой pI, метилэстерификация могла бы обеспечить разделение упомянутого белка при более основной pI. Можно, таким образом, утверждать, что небольшое количество кислой изоформы, наблюдаемой на фиг. 1А (стрелка белого цвета), может быть изоформой, которая содержит небольшое количество либо вовсе не содержит метиловых сложных эфиров, по сравнению с csPCNA изоформой. csPCNA изоформа имеет низкий уровень метилэстерификации, по сравнению с нормальной или незлокачественной формой PCNA (nmPCNA или просто PCNA). Незлокачественная или основная PCNA изоформа имеет более высокую степень метилэстерификации. Этот вывод основывается, отчасти, на том, что метилэстерификация модифицирует кислые остатки и могла бы сместить белок к более основной pI (вследствие утраты кислого заряда), a csPCNA изоформа располагается очень близко к своей расчетной pI 4,5. Ацетилирование, фосфорилирование и ADP-рибозилирование, однако, могли бы сместить упомянутый белок к более кислой pI ниже 4,5 (вследствие добавления кислого заряда). Таким образом, эти модификации, вероятно, не несут ответственности за смещение pI. Определение степени метилэстерификации PCNA и csPCNA позволяет различить злокачественную и незлокачественную изоформы (csPCNA и nmPCNA). Уровни метилэстерификации csPCNA и nmPCNA определяются и сравниваются с помощью раскрываемых в данном описании способов для диагностирования злокачественного образования. Например, в табл. II представлен метилэстерифицированный статус различных csPCNA пептидов и процент модифицированных в гетерогенной популяции. Таблица II может применяться как схема сравнения для определения различных уровней метилэстерификации при диагностировании злокачественного новообразования. Пример 4. Полуколичественная оценка метилэстерифицированных csPCNA пептидов. Дополнительно исследовалась идентификация метиловых сложных эфиров на csPCNA относительно pI упомянутой изоформы. Вычисленная pI PCNA равна приблизительно 4,5, a pI csPCNA, определенная после калибровки 2D-PAGE геля с применением изоэлектрических точек окружающих белков, была несколько выше, приблизительно 4,6. В противоположность этому, если бы 100% всех 16 кислых остатков, идентифицированных на фиг. 2, были бы метилэстерифицированы, изоэлектрическая точка упомянутого белка, вероятнее всего, сместилась бы в основную сторону несколько больше, чем на 0,1 единицы рН (например, 5,66). Могут существовать дополнительные остатки, которые модифицируются с образованием основной или nmPCNA изоформы. nmPCNA изоформа может также метилэстерифицироваться на иных и/или дополнительных остатках, по сравнению с csPCNA. Способы, раскрываемые в настоящем описании, позволяют специалисту в данной области техники определять уровни метилэстерификации csPCNA и nmPCNA. Относительное содержание метилэстерифицированных пептидов определяли и сравнивали с немодифицированными пептидами. Это осуществляли посредством измерения и сравнения площадей пиков каждого немодифицированного пептида и его метилэстерифицированного аналога. Сравнение площадей пиков выявило относительное содержание каждого метилового сложного эфира, идентифицированного в этом LC-MS/MS эксперимента, как показано в табл. II. Каждый из упомянутых пептидов, при сравнении площадей пиков, показывает лишь частичную метилэстерификацию ( <25%). Следовательно, csPCNA изоформа, вероятно, состоит из гетерогенной популяции молекул PCNA с одинаковой изоэлектрической точкой. Иными словами, одиночная молекула csPCNA, вероятно, демонстрирует один или несколько метиловых сложных эфиров, но не 16. Однако один или несколько метиловых сложных эфиров могут находиться на 16 различных остатках белка. Эта гетерогенность csPCNA иллюстрируется присутствием метилэстерификации на С-конечном пептиде csPCNA. Немодифицированный пептид, IEDEEGS (778 m/z), элюировался на 28,9 мин (фиг. 3А) и CID спектр этого пептида соответствует пептиду, содержащему немодифицированные кислые остатки (фиг. 3В). Интересно отметить, что на выбраной ионной хроматограмме метилэстерифицированной разновидности (792 m/z) этого пептида два пика идентифицируются с 2-3 мин увеличением времени удерживания. Это, вероятно, обусловлено повышенной гидрофобностью и потерей заряда, вызванной метилэстерификацией. Разделение этих двух пиков, таким образом, свидетельствовало о различии в структуре этих пептидов. Изучение CID спектров показало, что упомянутые пептиды метилэстерифицированы, но на разных остатках (фиг. 3С и фиг. 3D). Поскольку не наблюдалось ни одного пептида, содержащего метиловые сложные эфиры на обоих остатках, появление этих пептидов, с наибольшей вероятностью, явилось следствием анализа гетерогенной популяции csPCNA. Возможно, что наблюдаемая гетерогенность и низкий процент модифицированных разновидностей могут быть следствием высокой лабильности самих метилэстерифицированных модификаций. В некоторых сообщениях указывается на то, что метилэстерифицированные модификации белков являются недолговечными в нейтральных и основных растворах. Таким образом, белковые метиловые сложные эфиры, подобные обнаруженным на csPCNA, могут спонтанно гидролизоваться, оставляя в результате не модифицированный остаток и метанол. В дополнение к этому, основные и окисляющие условия SDS-PAGE также могут вести к утрате метиловых сложных эфиров из PCNA, и попытки разделения основной PCNA изоформы, разновидности PCNA с высоким уровнем метилэстерификации, до ее основной pI, демонстрируют спонтанную регрессию в сторону более кислой pI (фиг. 4). Высокий уровень метиловых сложных эфиров является, вероятно, причиной того, что PCNA фокусируется на основной pI, как показано на иммуноблоте на фиг. 4 (рН приблизительно 8,8-9,0). Фокусировка этой изоформы, однако, может не быть однородной (полосатой) и может происходить с более низкой интенсивностью. Основные значения рН, при которых происходит разделение этой изоформы, могут не обладать достаточной проводимостью для полного поддержания метилэстерификации упомянутого белка. Невозможность фокусировки в конкретной pI, наблюдаемая на этом геле (фиг. 4), обусловлена, вероятно, сопутствующей утратой одного или нескольких метиловых сложных эфиров вследствие фокусировки при основном значении рН. Происходит спонтанный гидролиз метиловых сложных эфиров, высвобождающий метанол и немодифицированную аминокислотную боковую цепь. Восстановление кислых боковых цепей этим "основным гидролизом" вызывает, вероятно, смещение pI PCNA с основной в сторону более кислой, о чем свидетельствует накопление PCNA на более кислой стороне геля (рН 7). Идентификация и анализ метилэстерификации могут осуществляться при условиях, которые сводят до минимума утрату метиловых сложных эфиров. Например, был описан способ кислого 2D-PAGE, при проведении которого создаются условия, способные сохранить белковые метиловые сложные эфиры (О'Коннор (O'Connor) и другие, Anal. Biochem., I985, 148, 79-86). Однако многие доступные протеазы, распознающие PCNA, активны при рН от нейтрального до кислого, и возможно, что некоторый значительный объем метилэстерификации может быть утрачен в процессе расщепления. В интактной клетке либо в экстрактах, фермент(-ы), ответственный(-е) за метилэстерификацию, может быть активными и может модифицировать остатки, утратившие метиловые сложные эфиры вследствие спонтанного гидролиза. Отделение PCNA от фермента(-ов), ответственного(-ых) за метилэстерификацию и инкубирование в условиях, поддерживающих гидролиз (например, рН выше 7,0) может привести к утрате одного либо нескольких метиловых сложных эфиров. Например, такая утрата метиловых сложных эфиров может быть сведена до минимального уровня путем поддержания слегка кислых условий в процессе манипулирования и анализа образцов. а: Местоположения белковых пятен идентифицированы на фиг. 1А. b: Пептиды четко демонстрируют +14 смещение массы на остатках аспарагиновой либо глутаминовой кислот. с: Покрытие последовательности вычисляется посредством деления количества идентифицированных аминокислотных остатков на общее количество остатков в белке. а: Присутствующие модификации пептидов: оксидированный метионин (M 0 ), карбамидометилцистеин (C ca ), метилэстерифицированная глутаминовая кислота (E m ) и метилэстерифицированная аспарагиновая кислота (D m ). b: Процентное содержание метилового сложного эфира вычисляли посредством деления площадей пиков метилэстерифицированных пептидов на объединенные площади пиков метилэстерифицированных и немодифицированных пептидов. с: Количественные показатели по механизму поиска Mascot представлены в виде -10log(P), где Р - вероятность того, что спаривание является случайным событием. d: Данные получены с помощью LCQ DECA ХР, по сравнению с LCQ Advantage. |