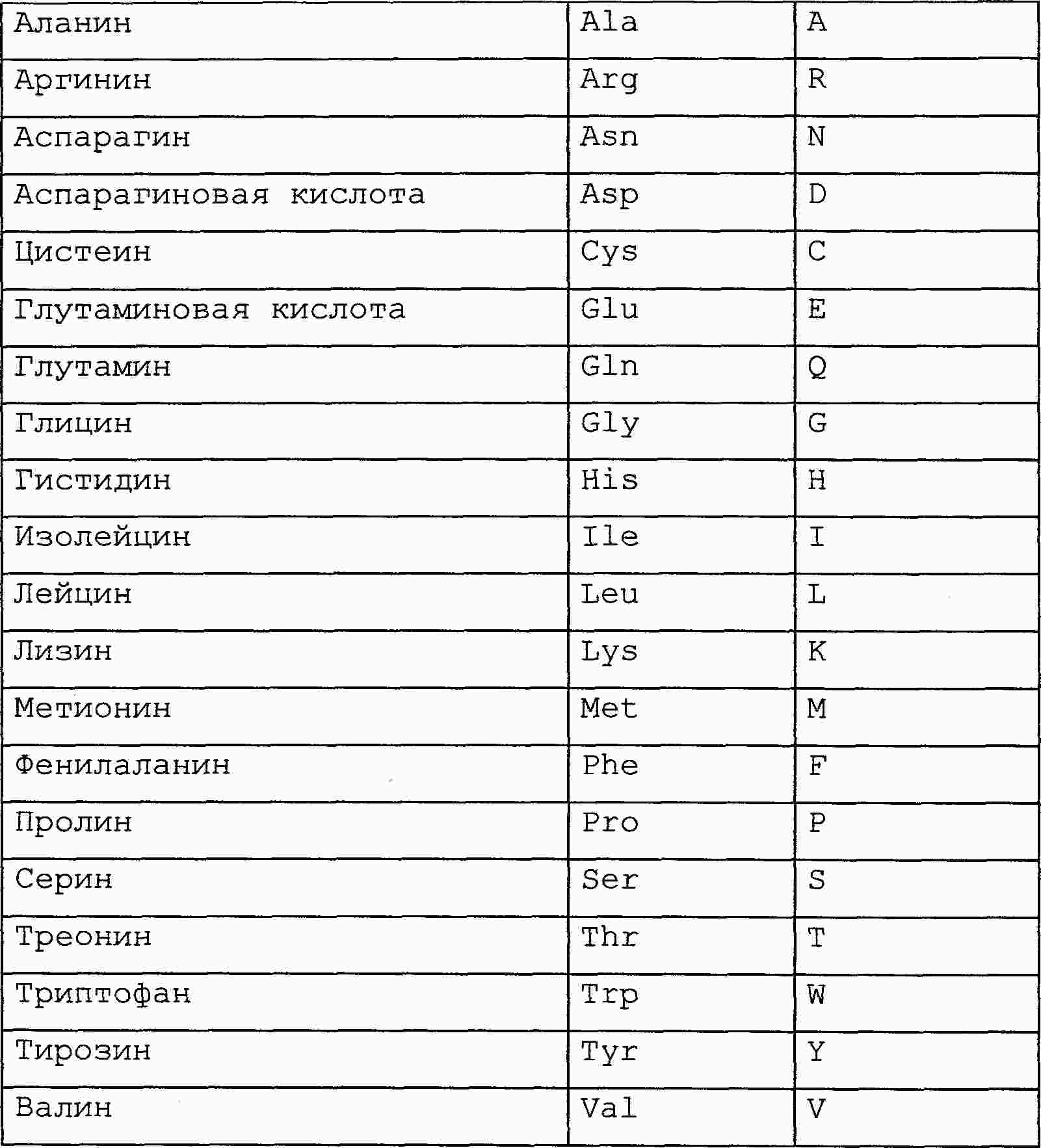
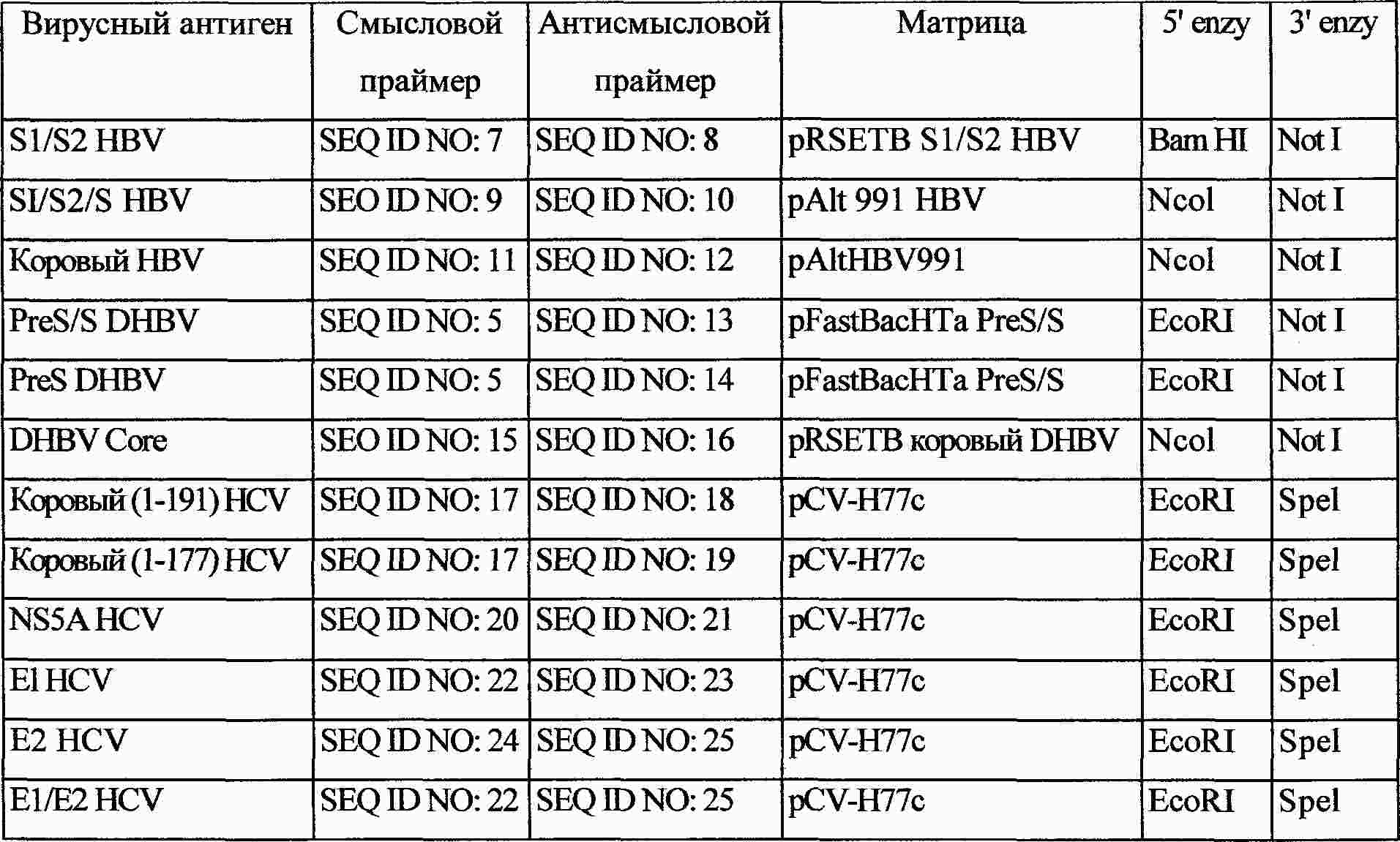
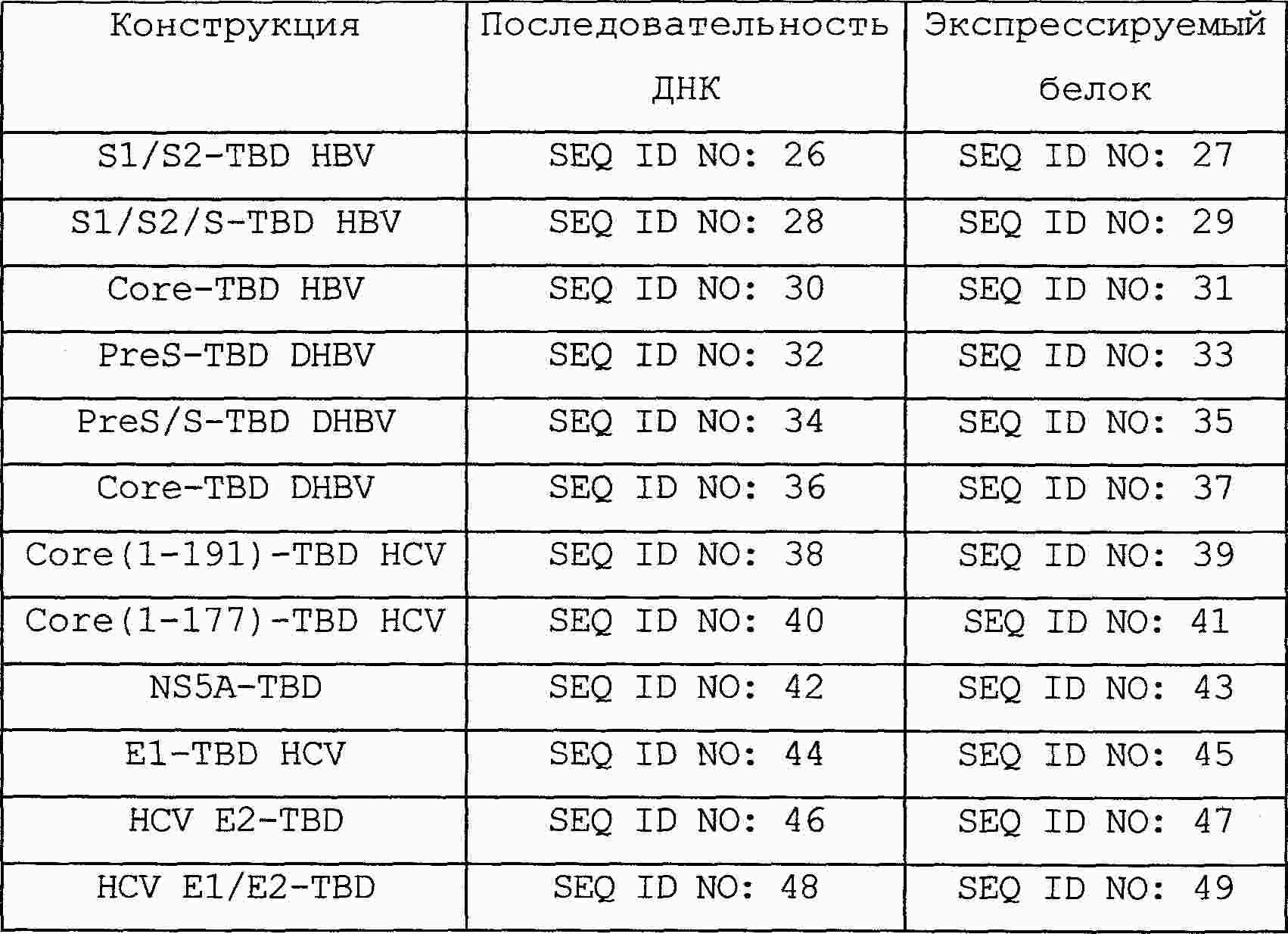
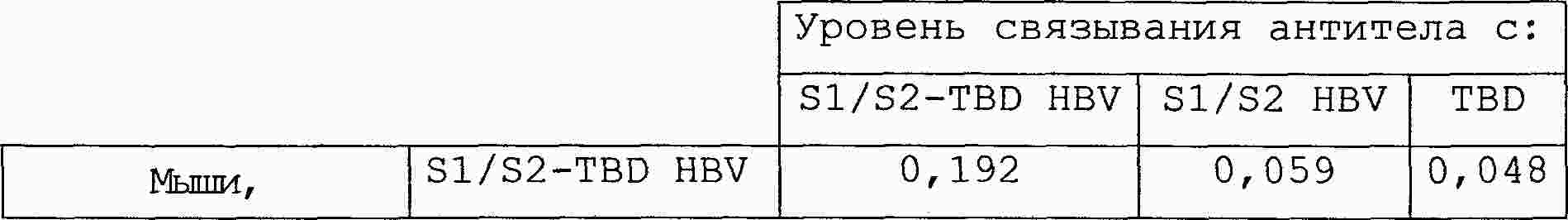
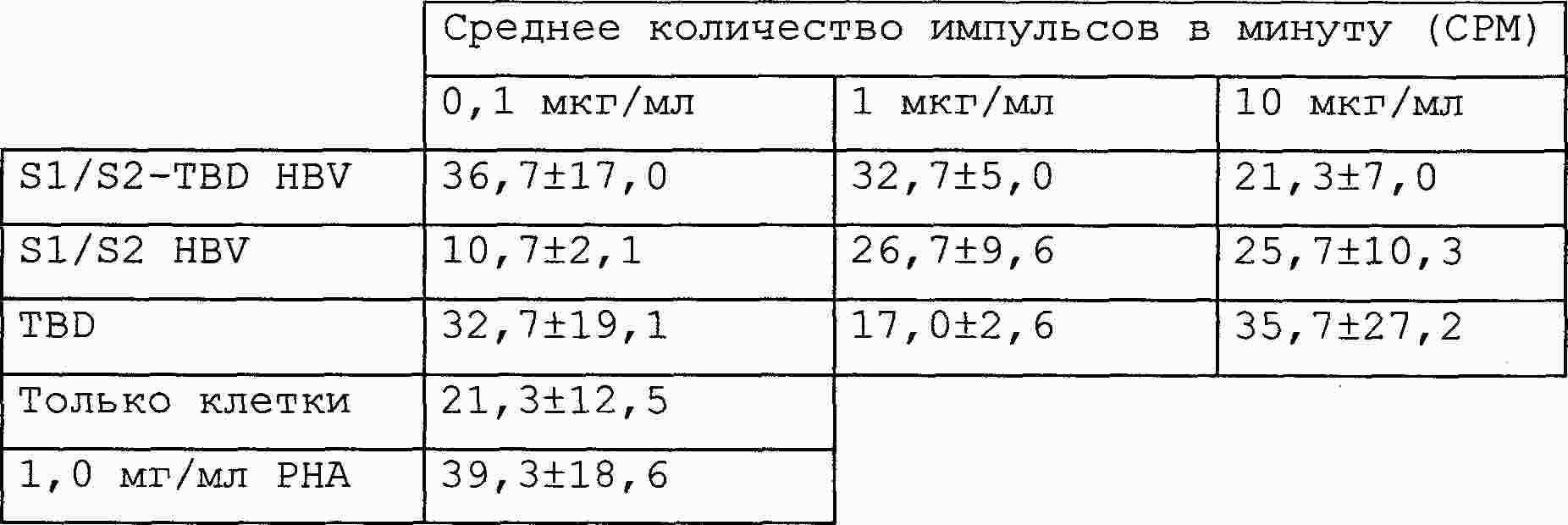
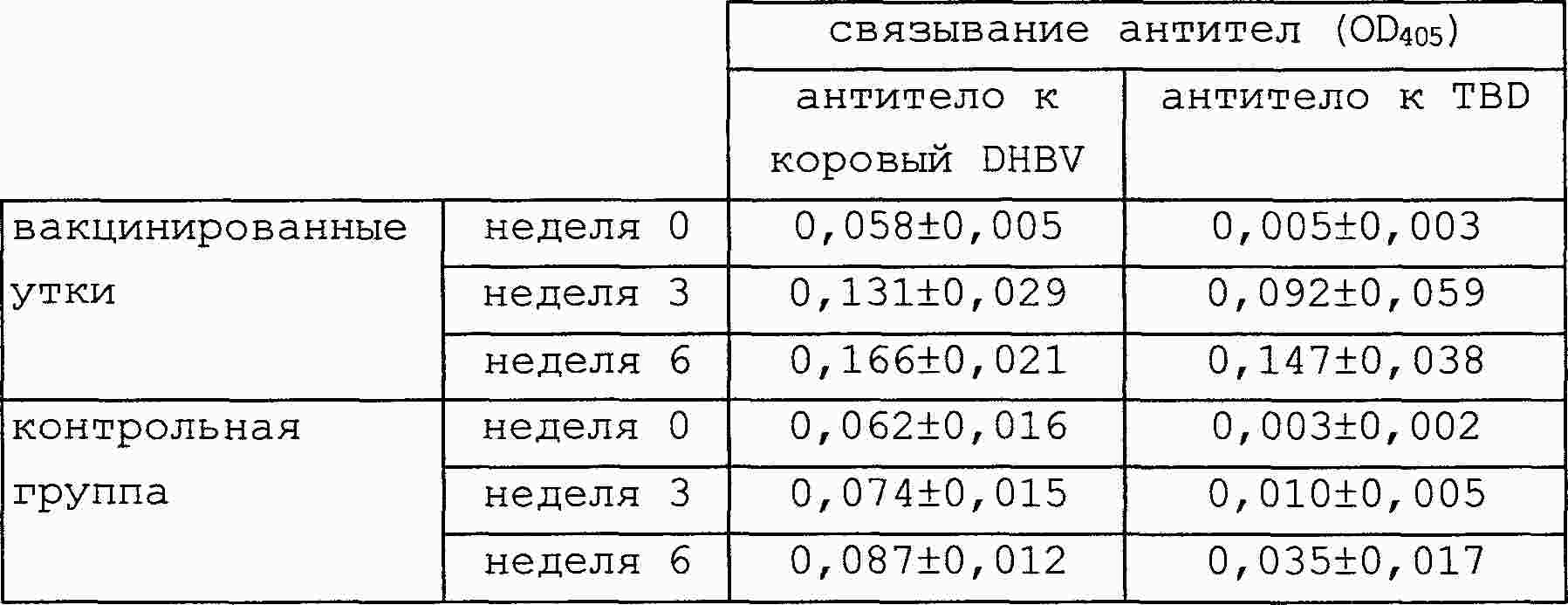
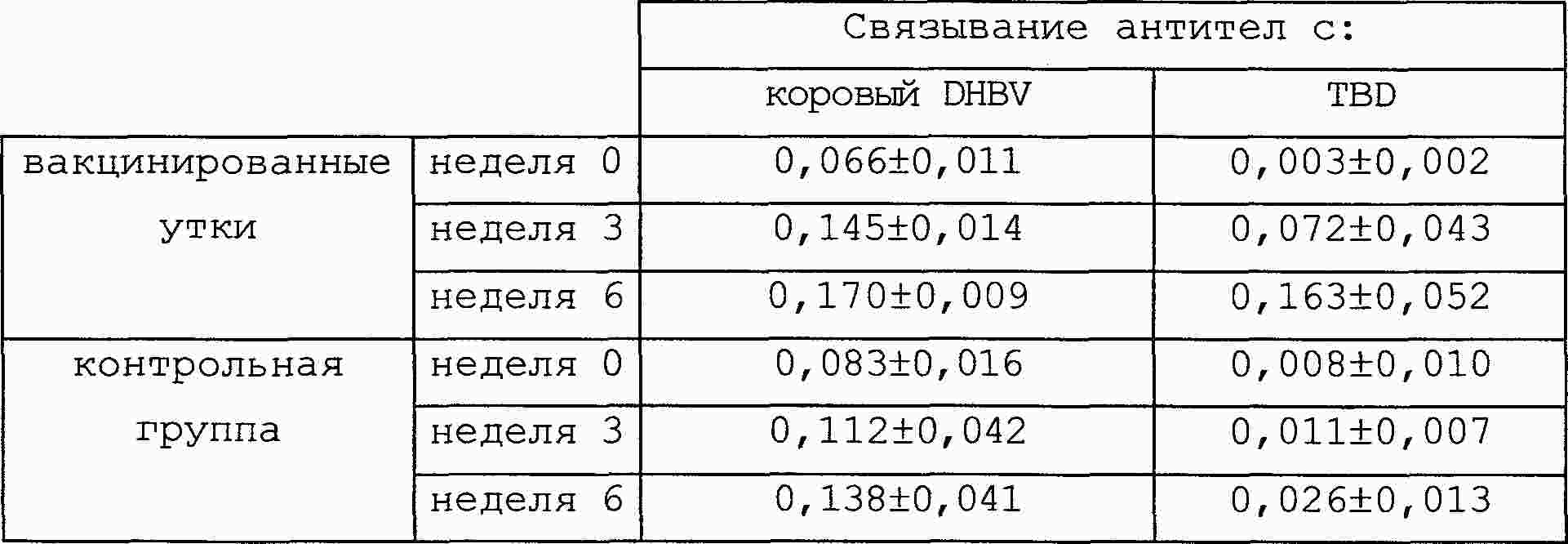
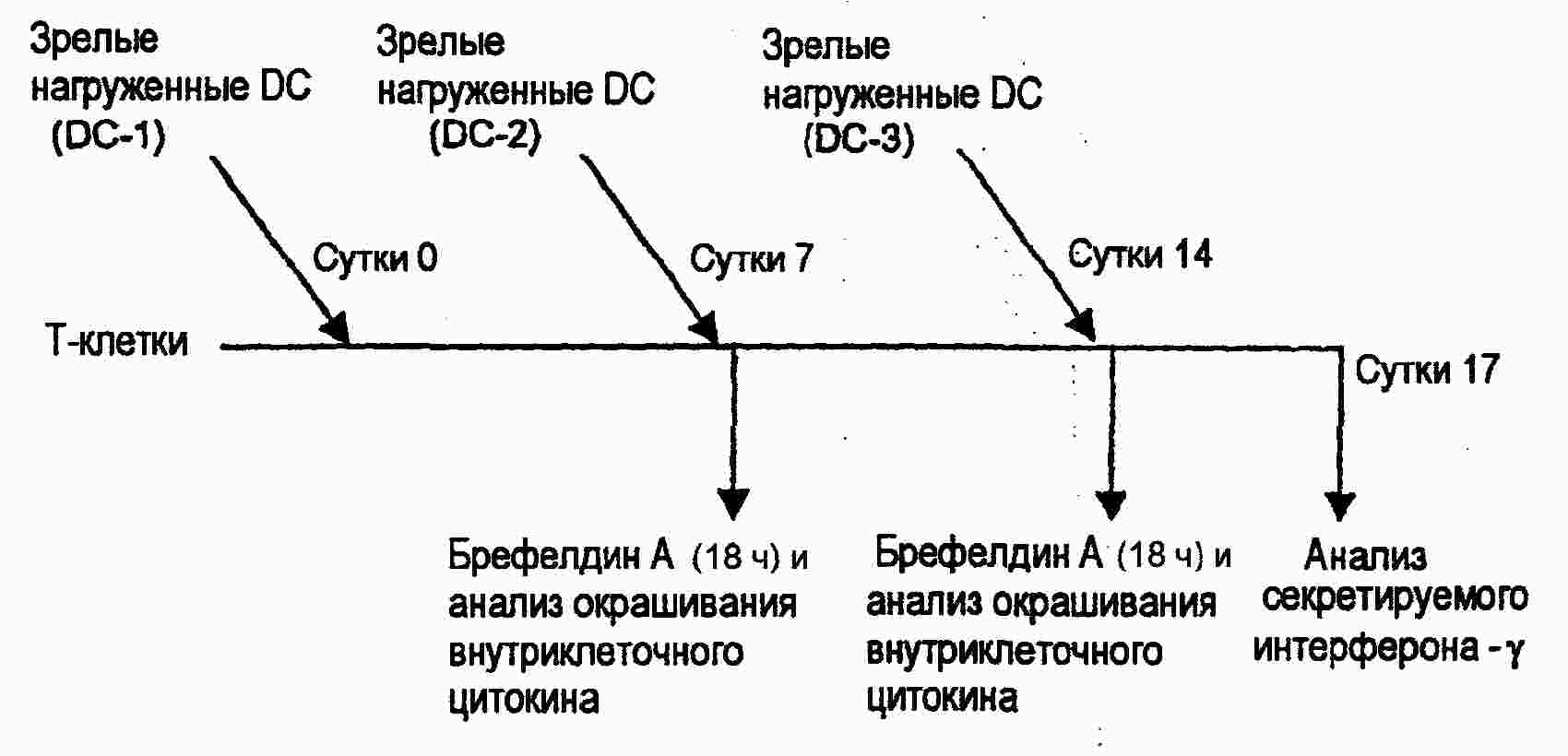
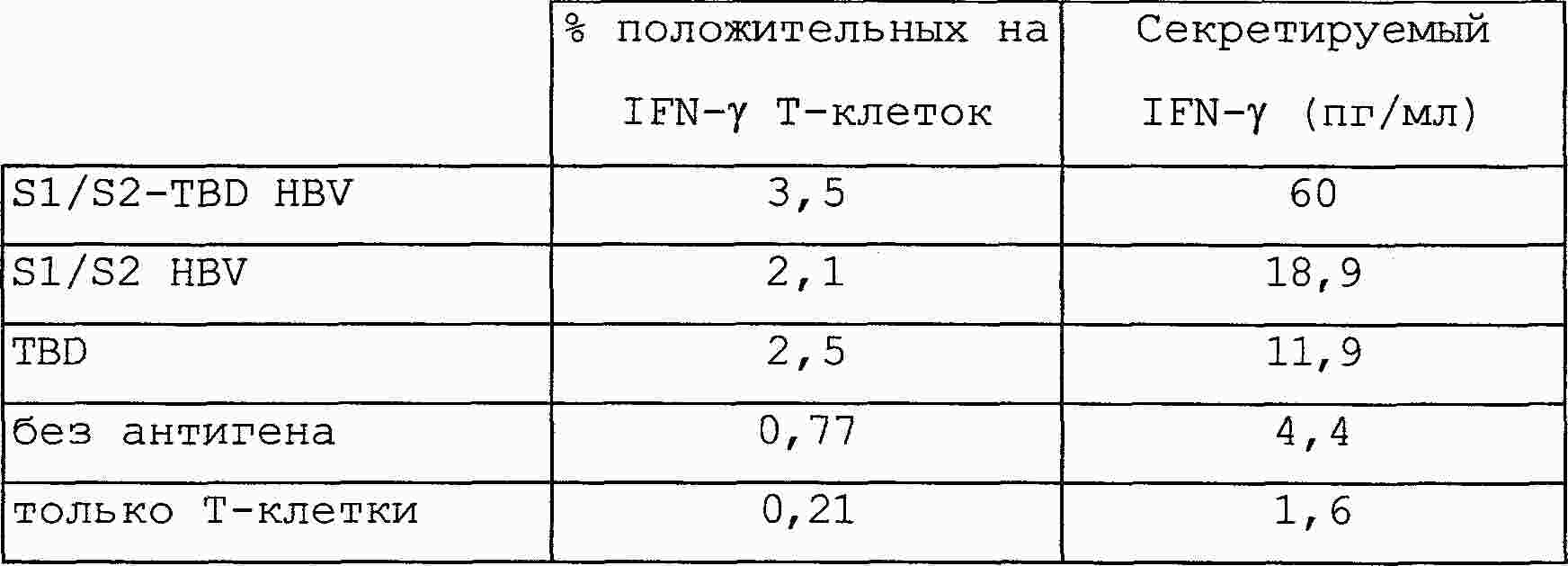
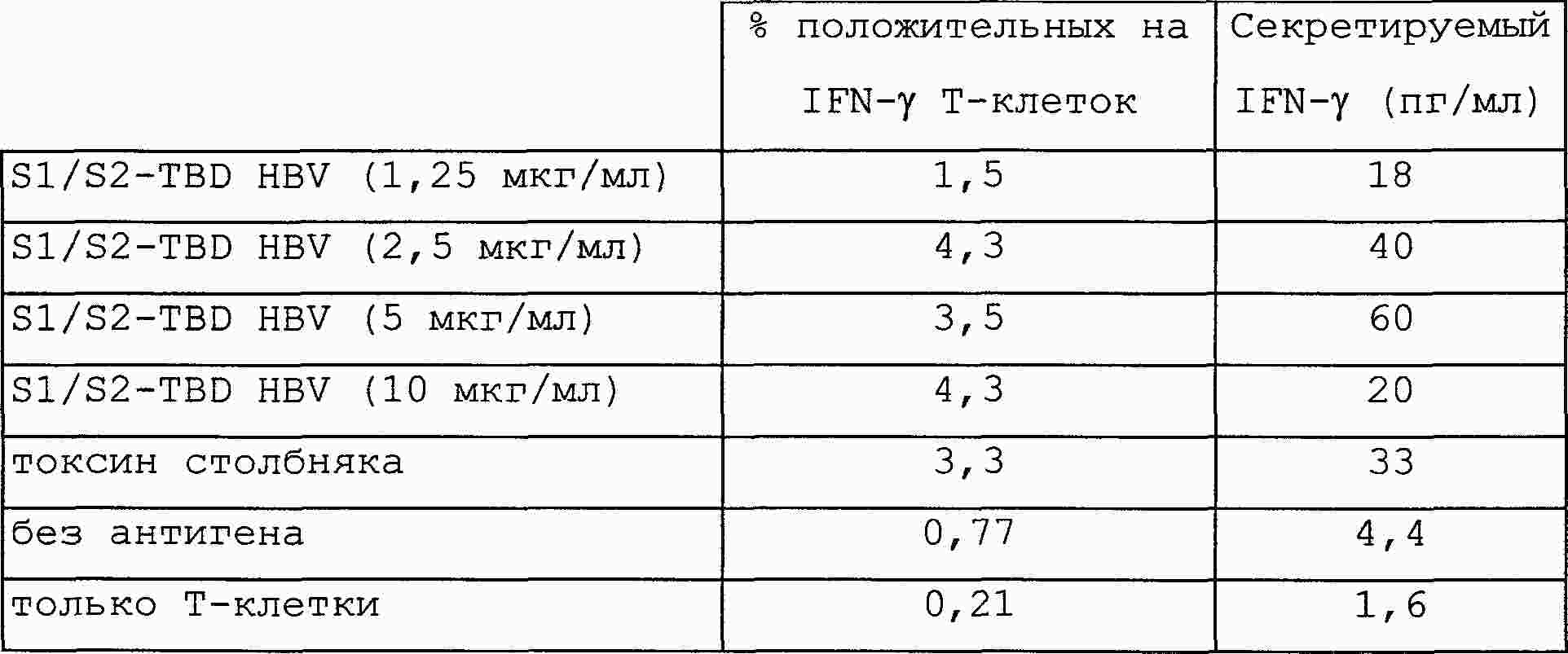
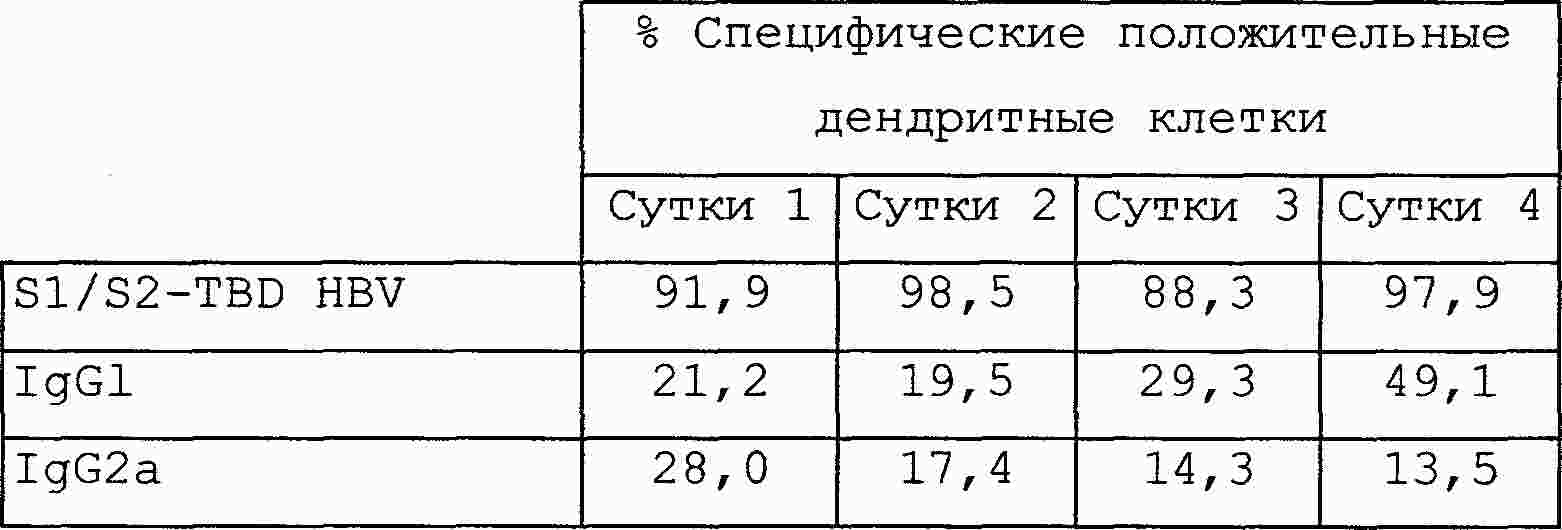
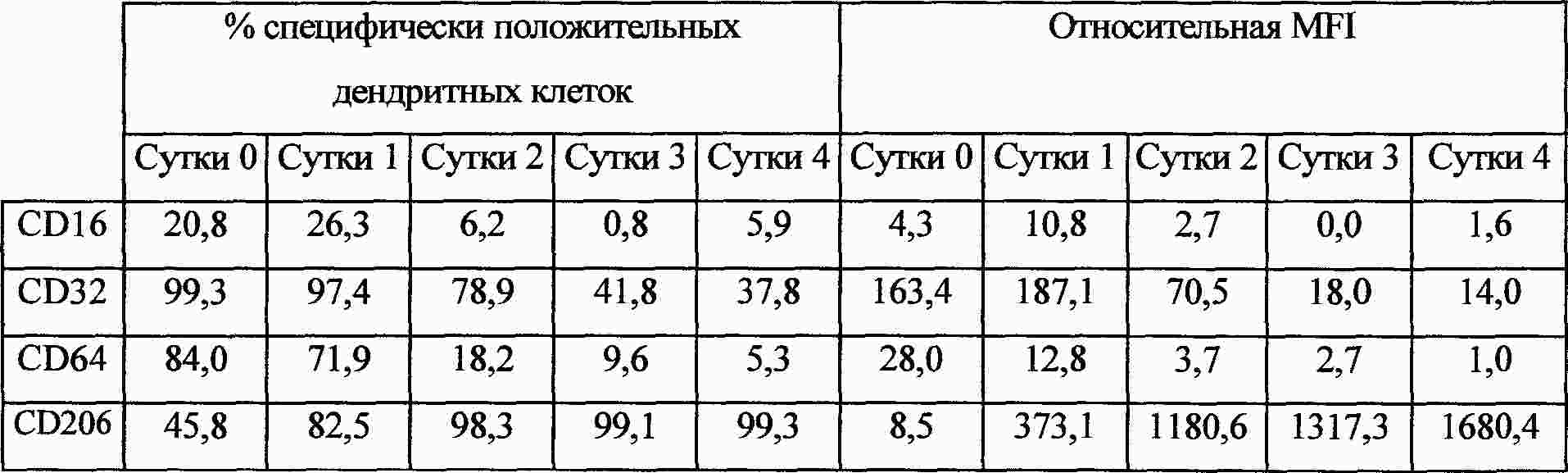
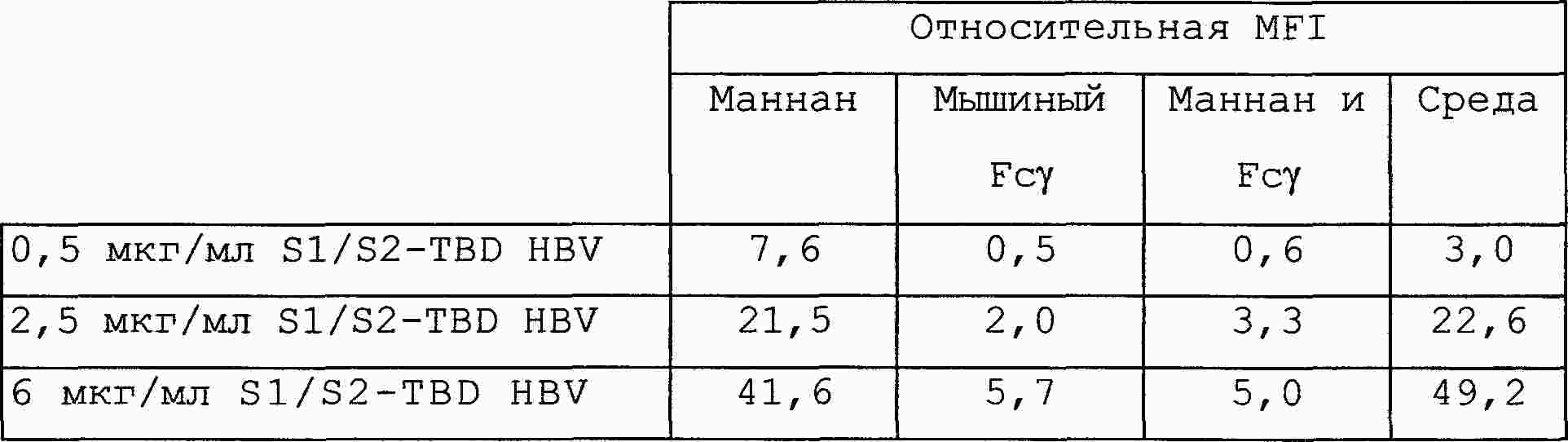
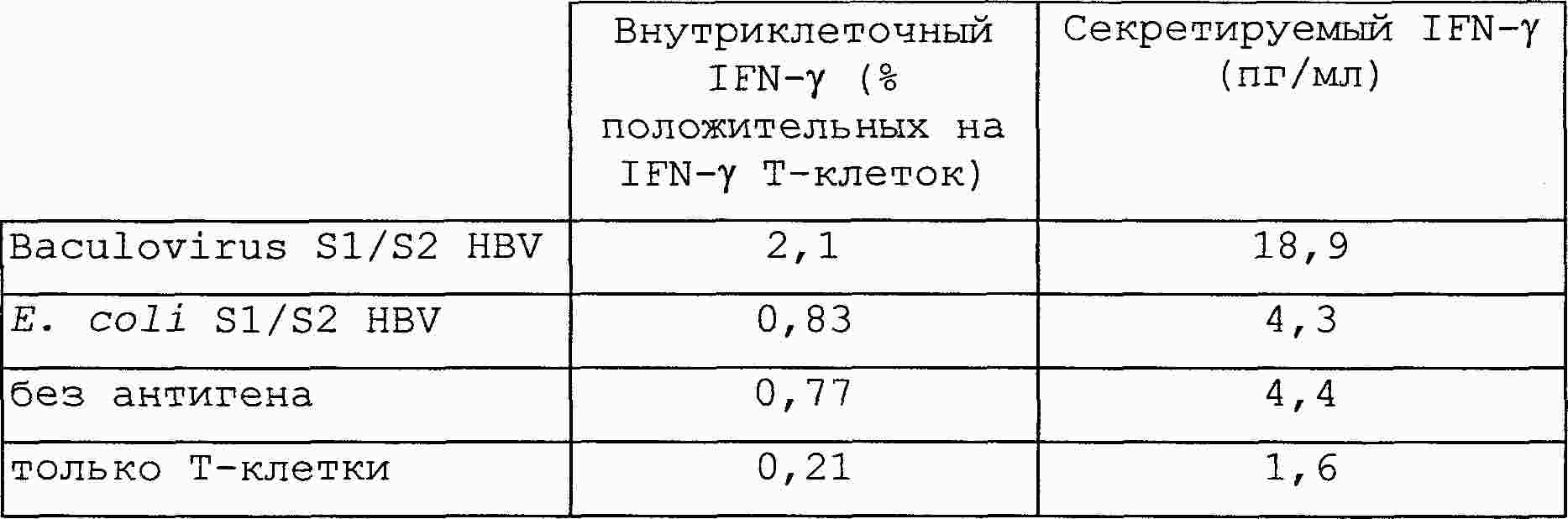
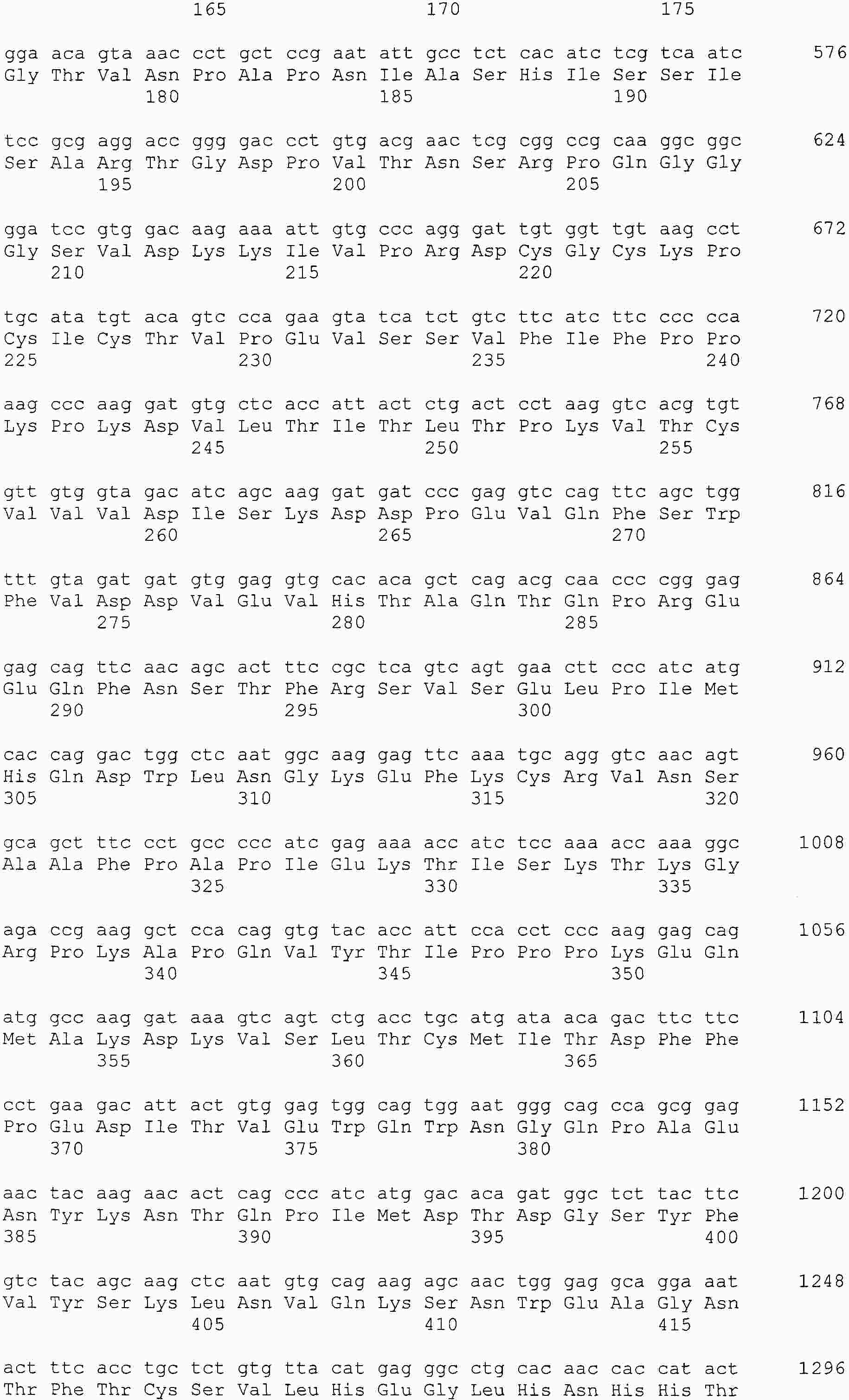
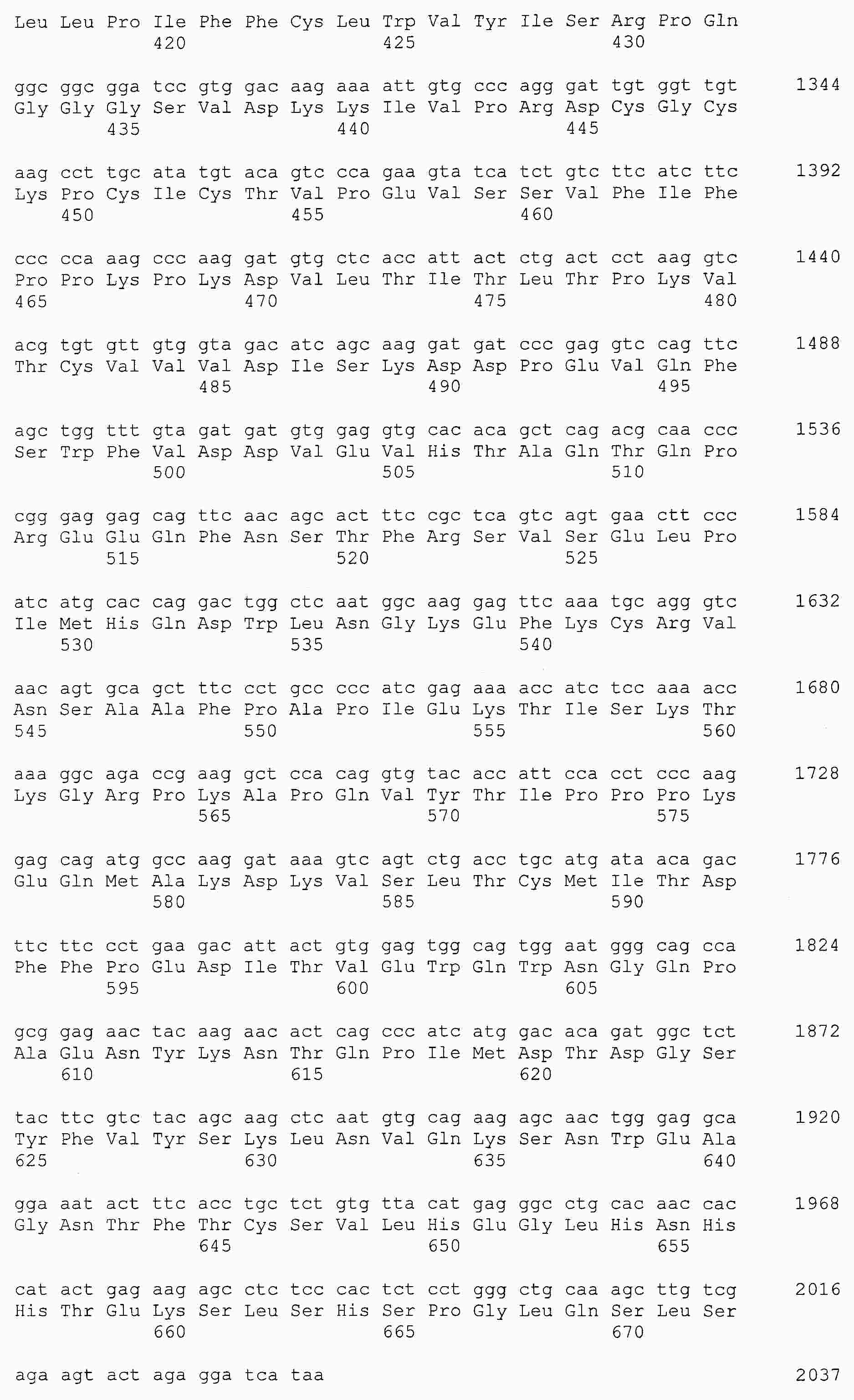
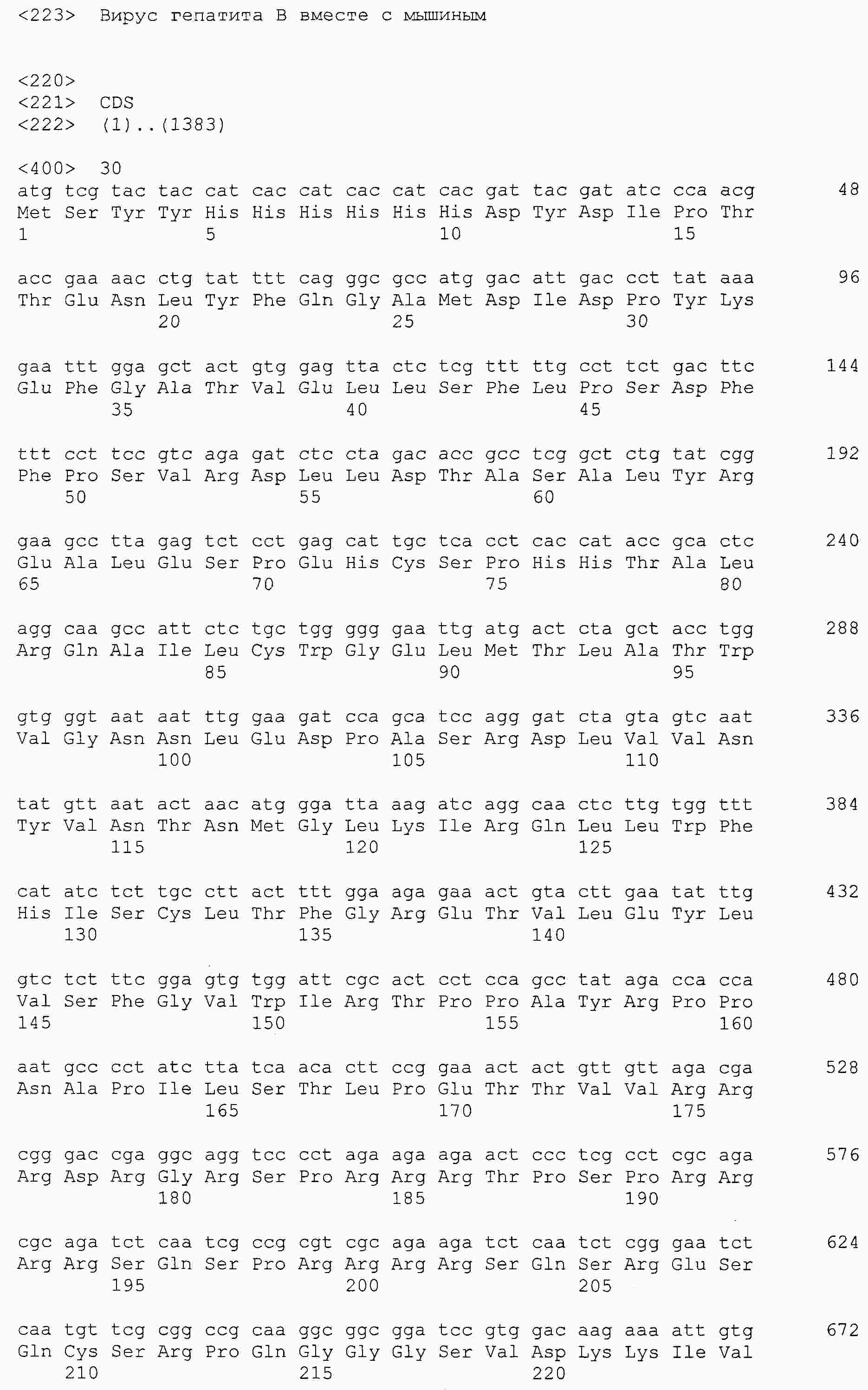
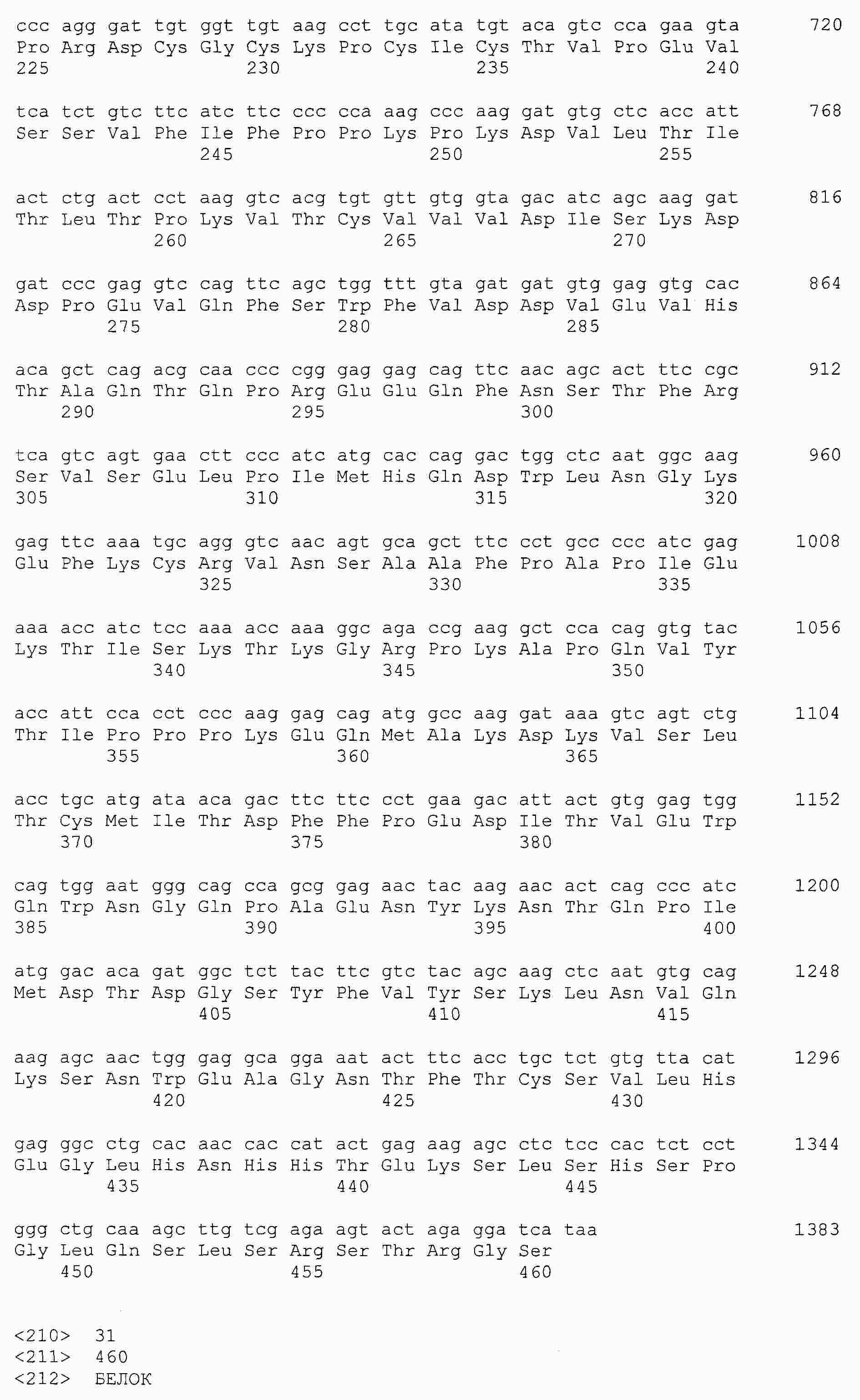
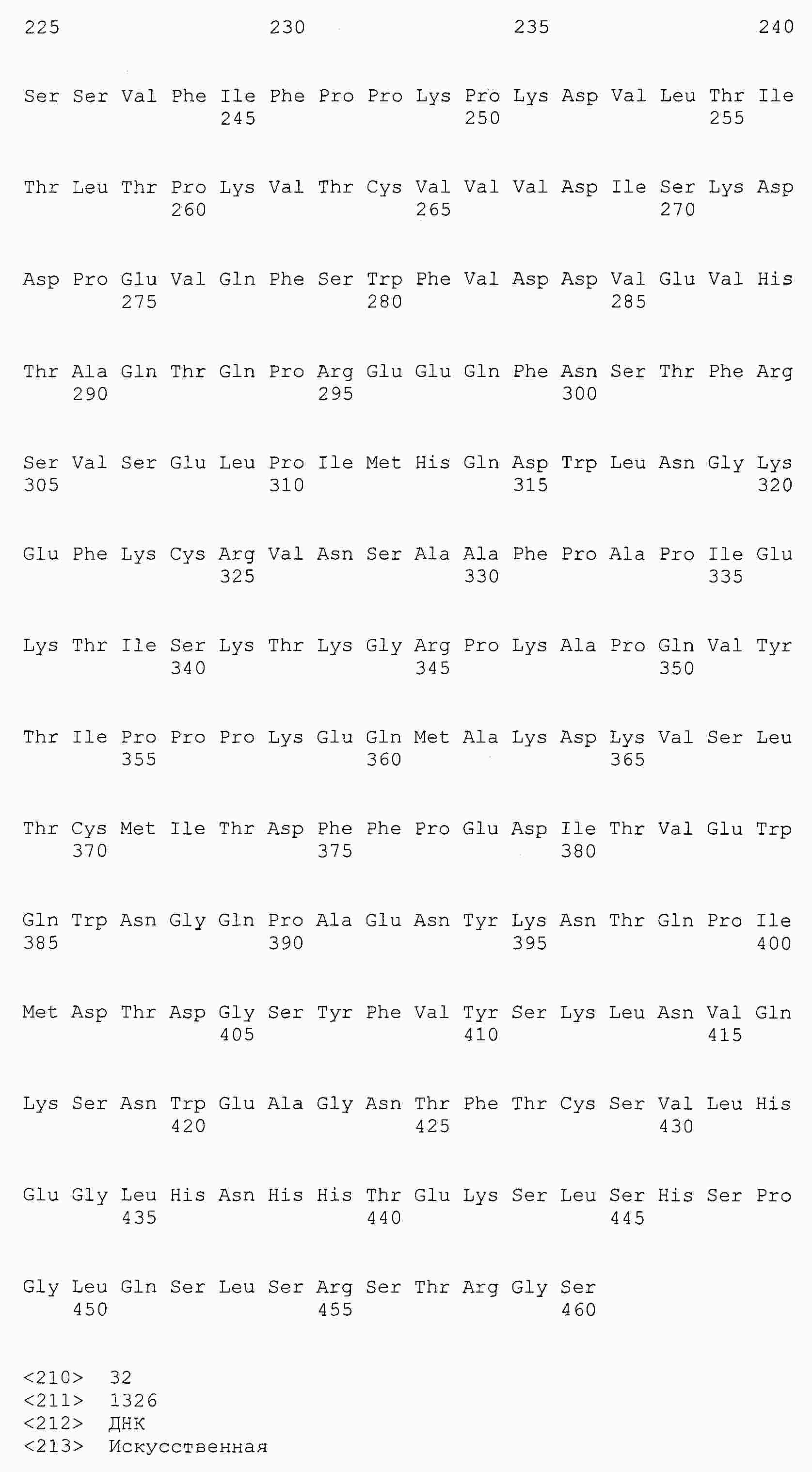
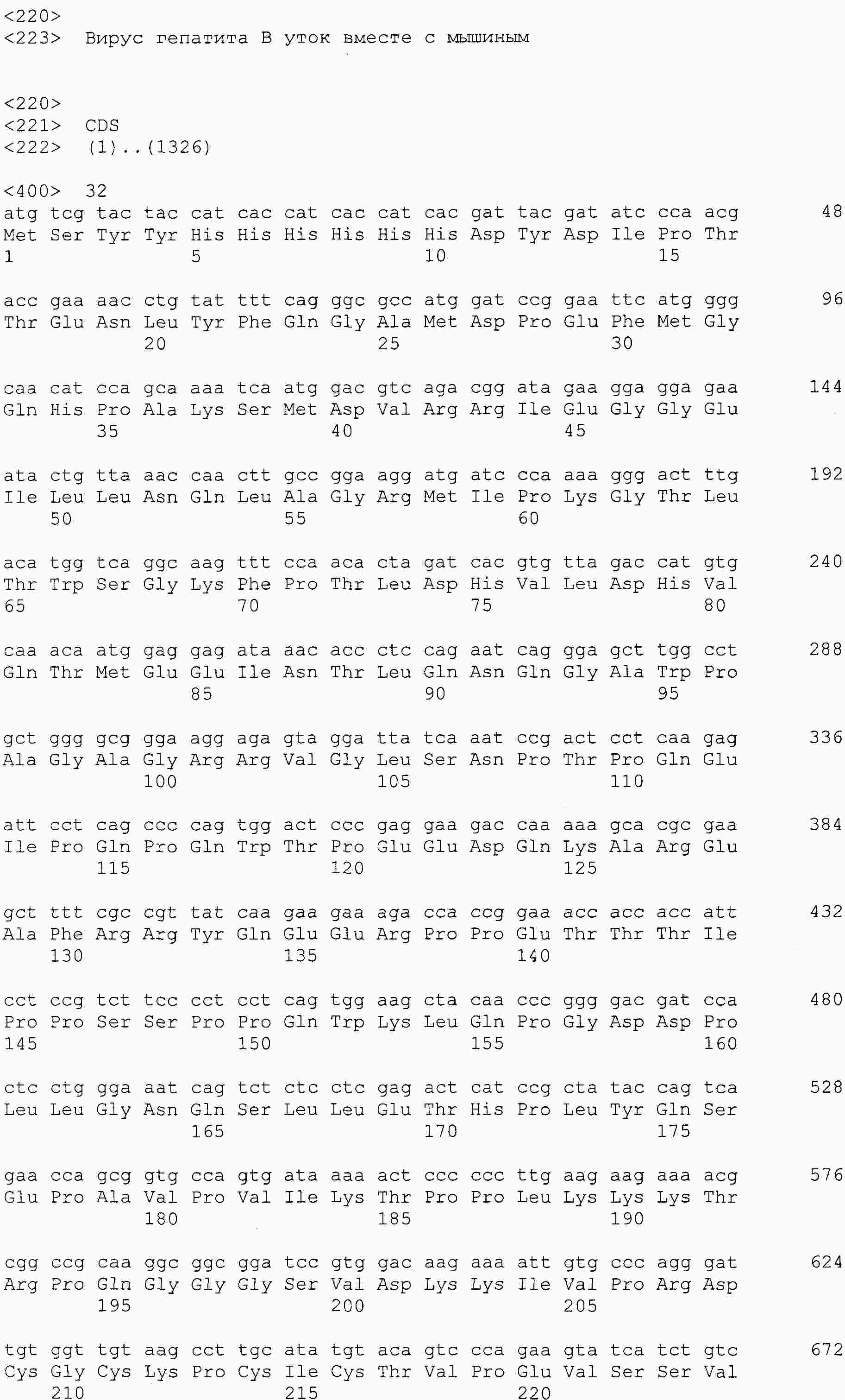
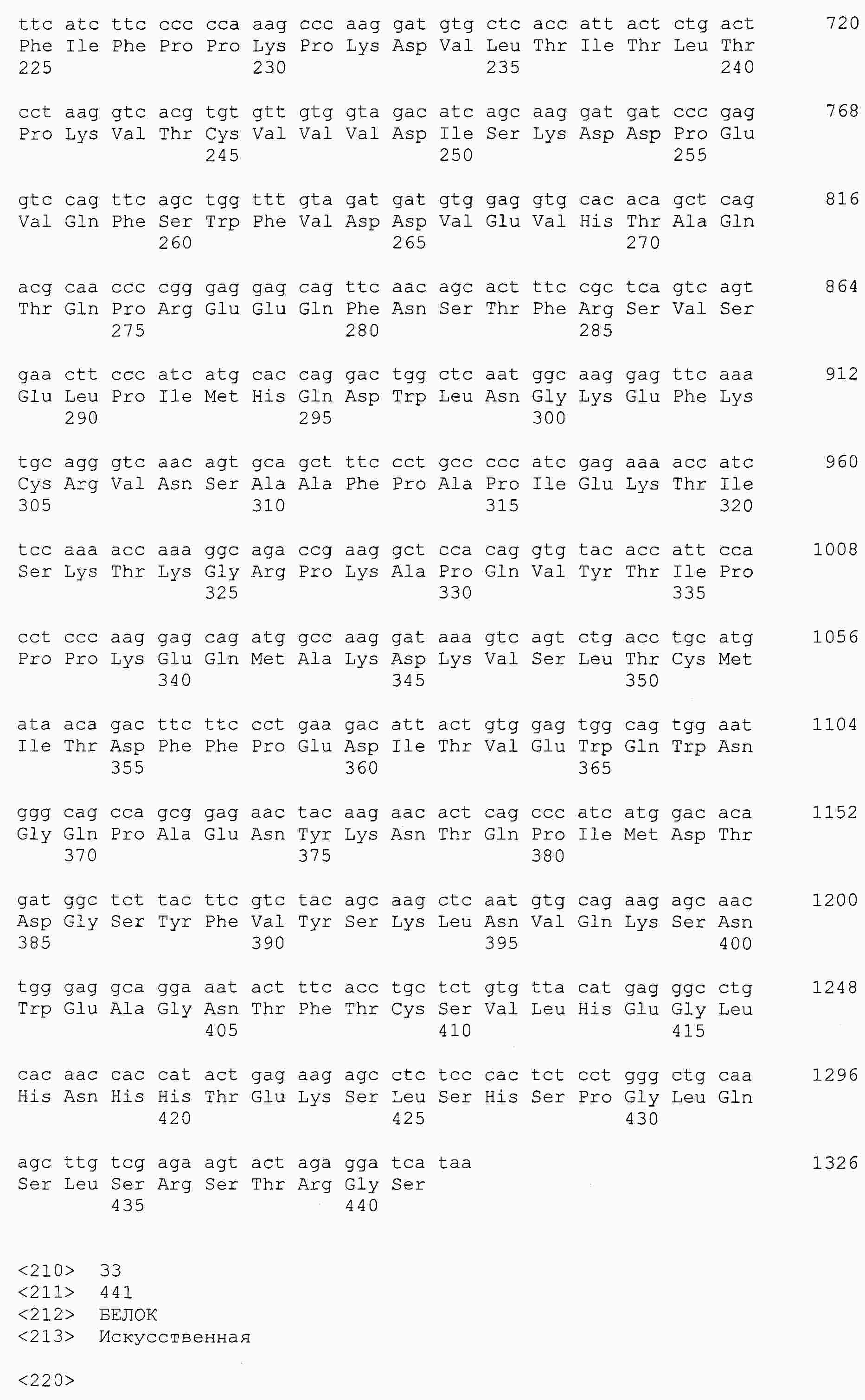
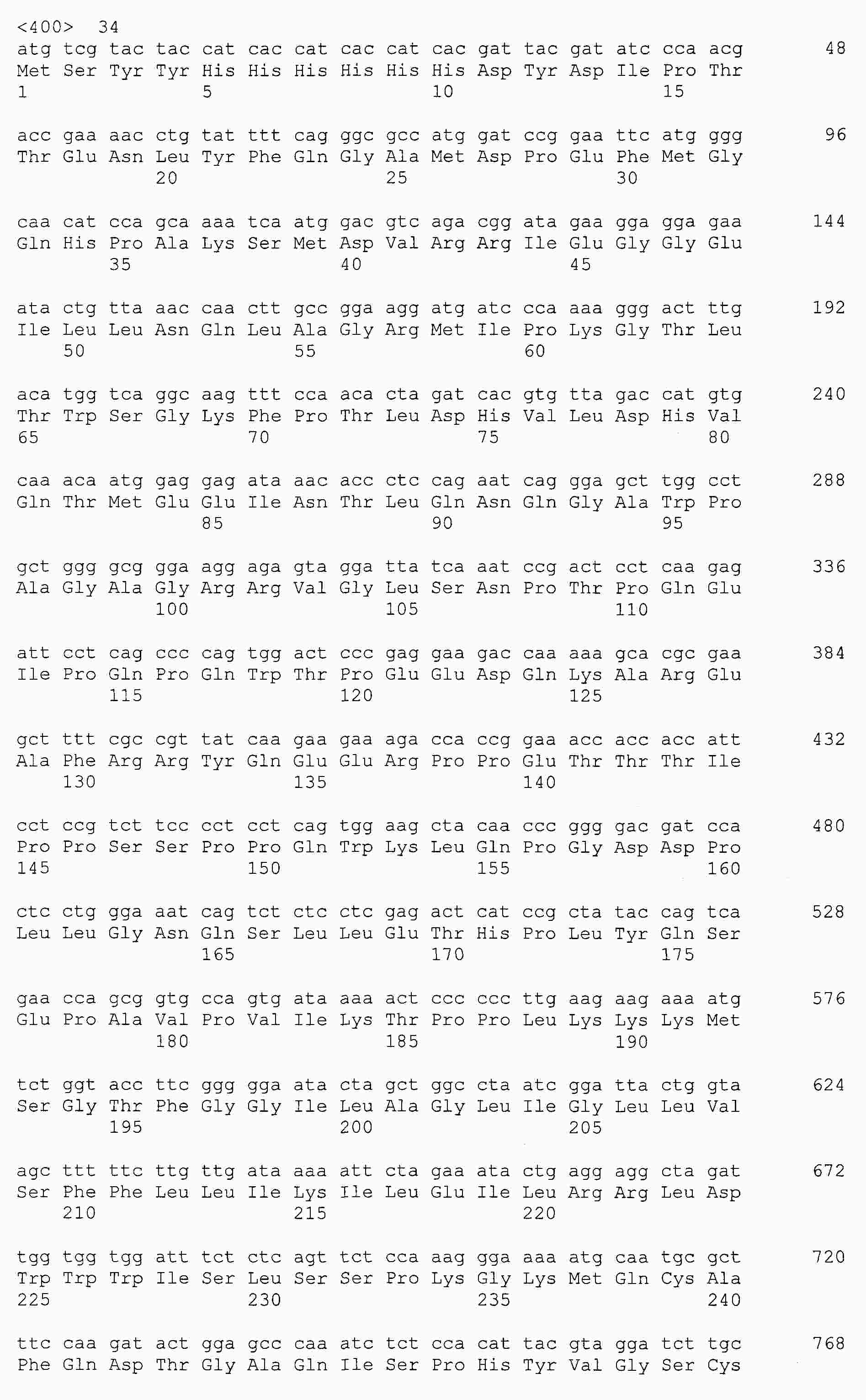
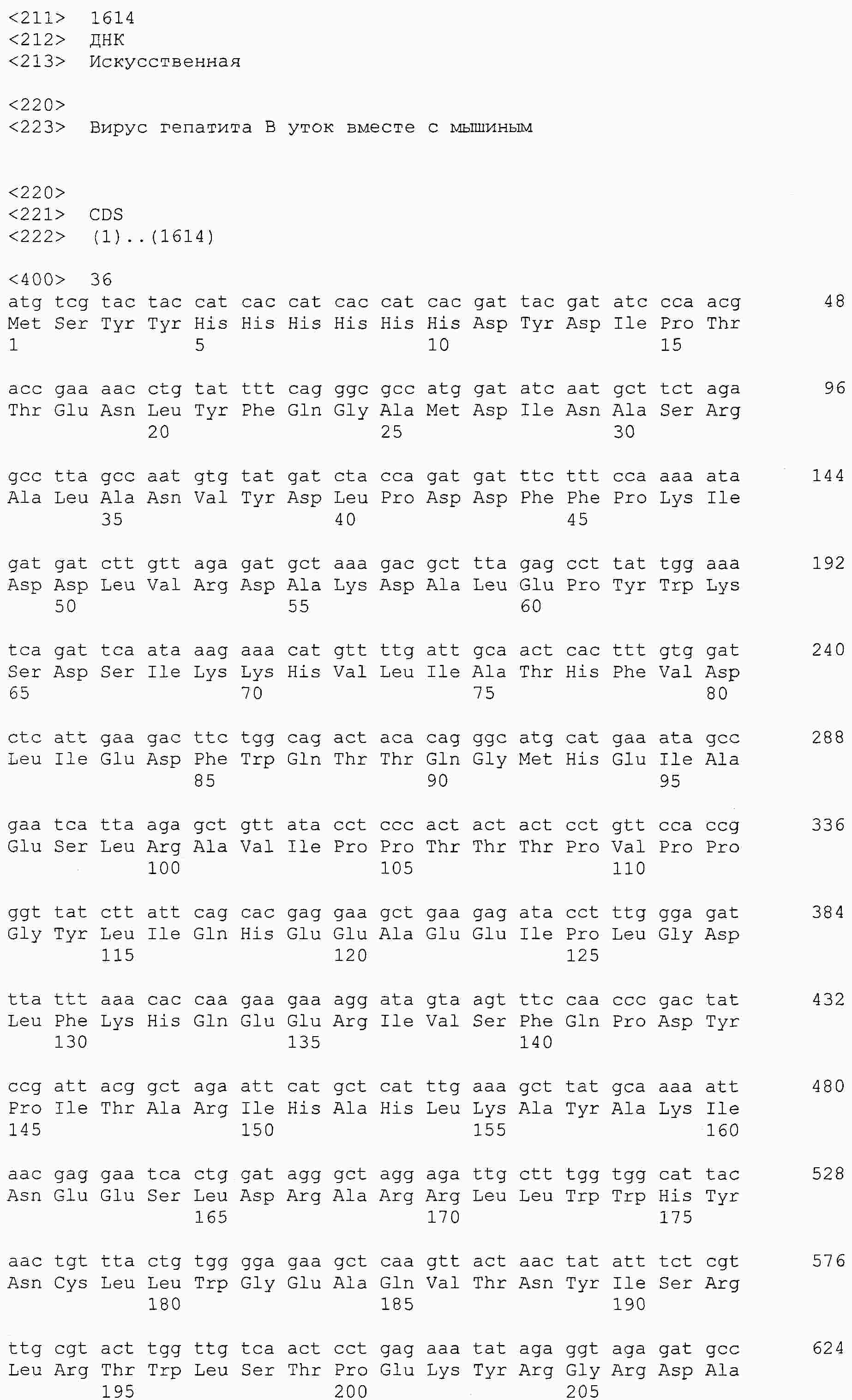
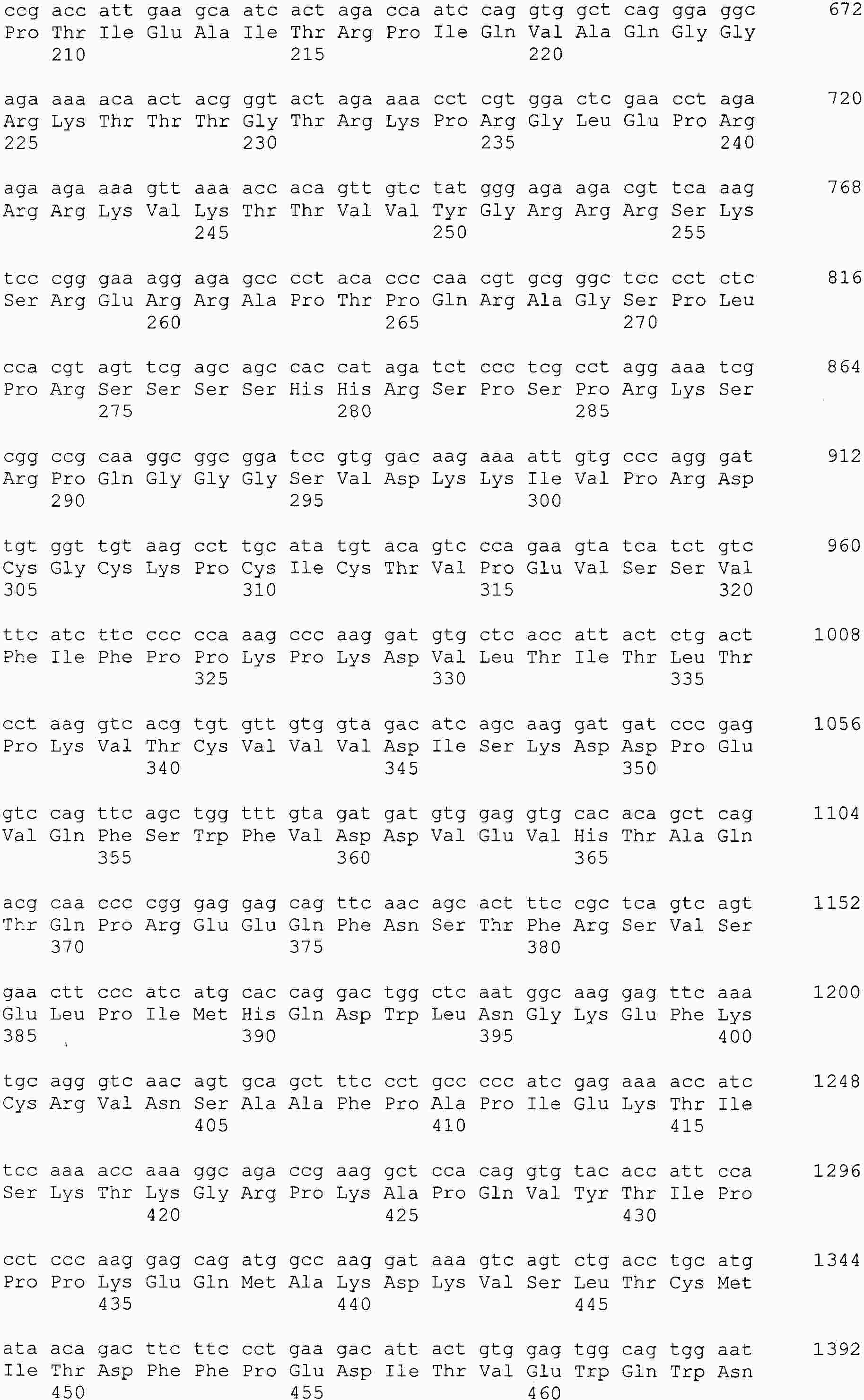
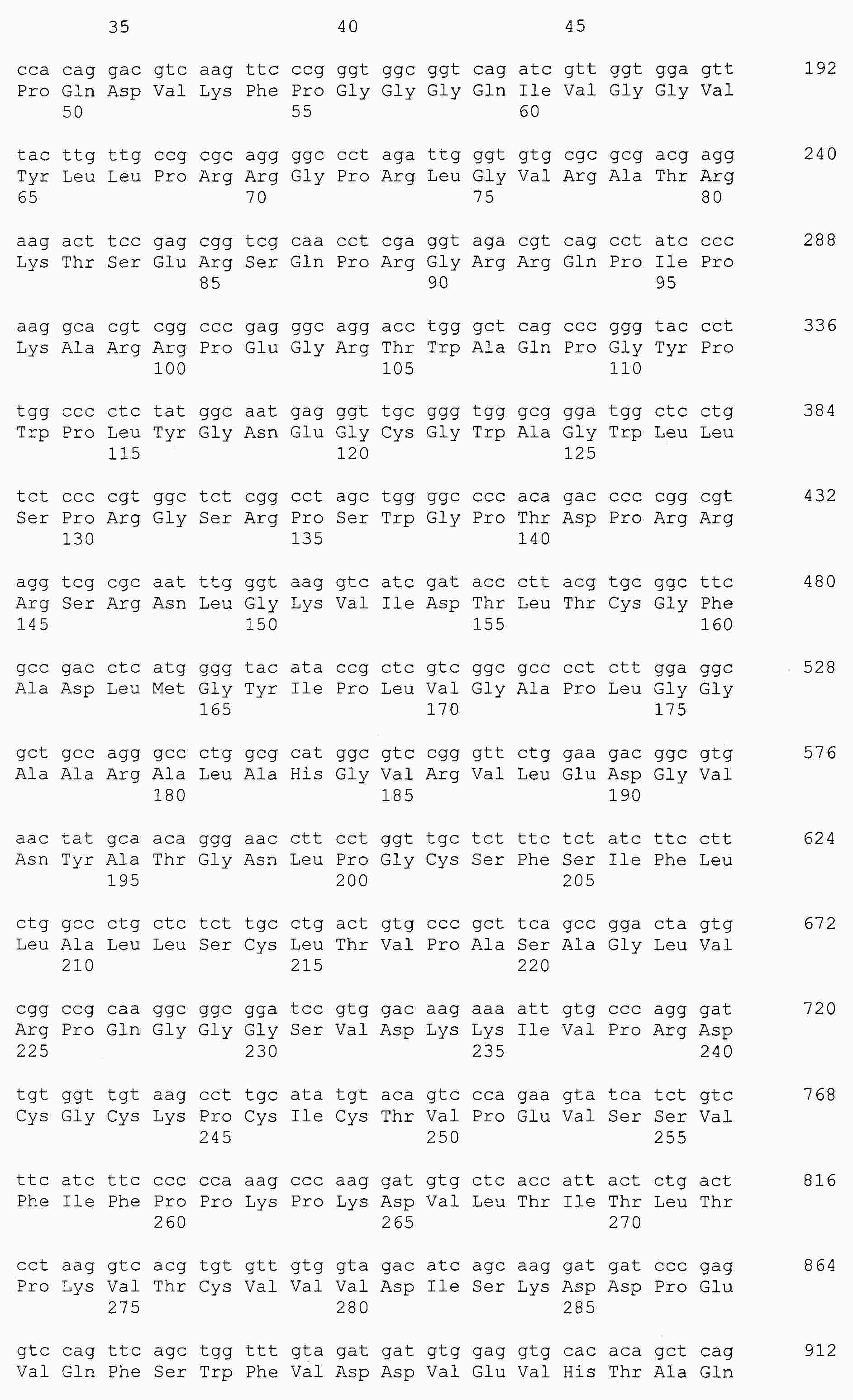
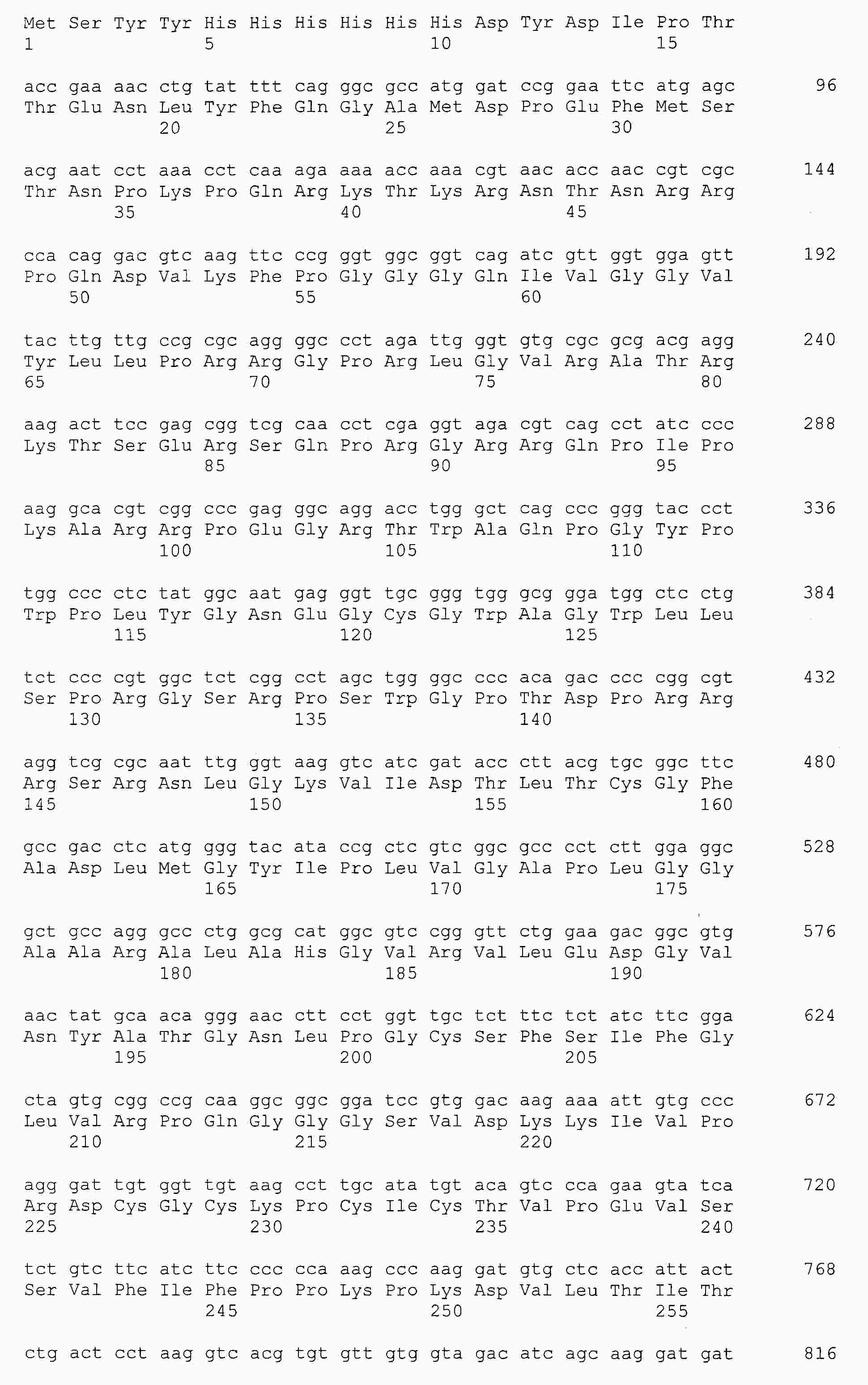
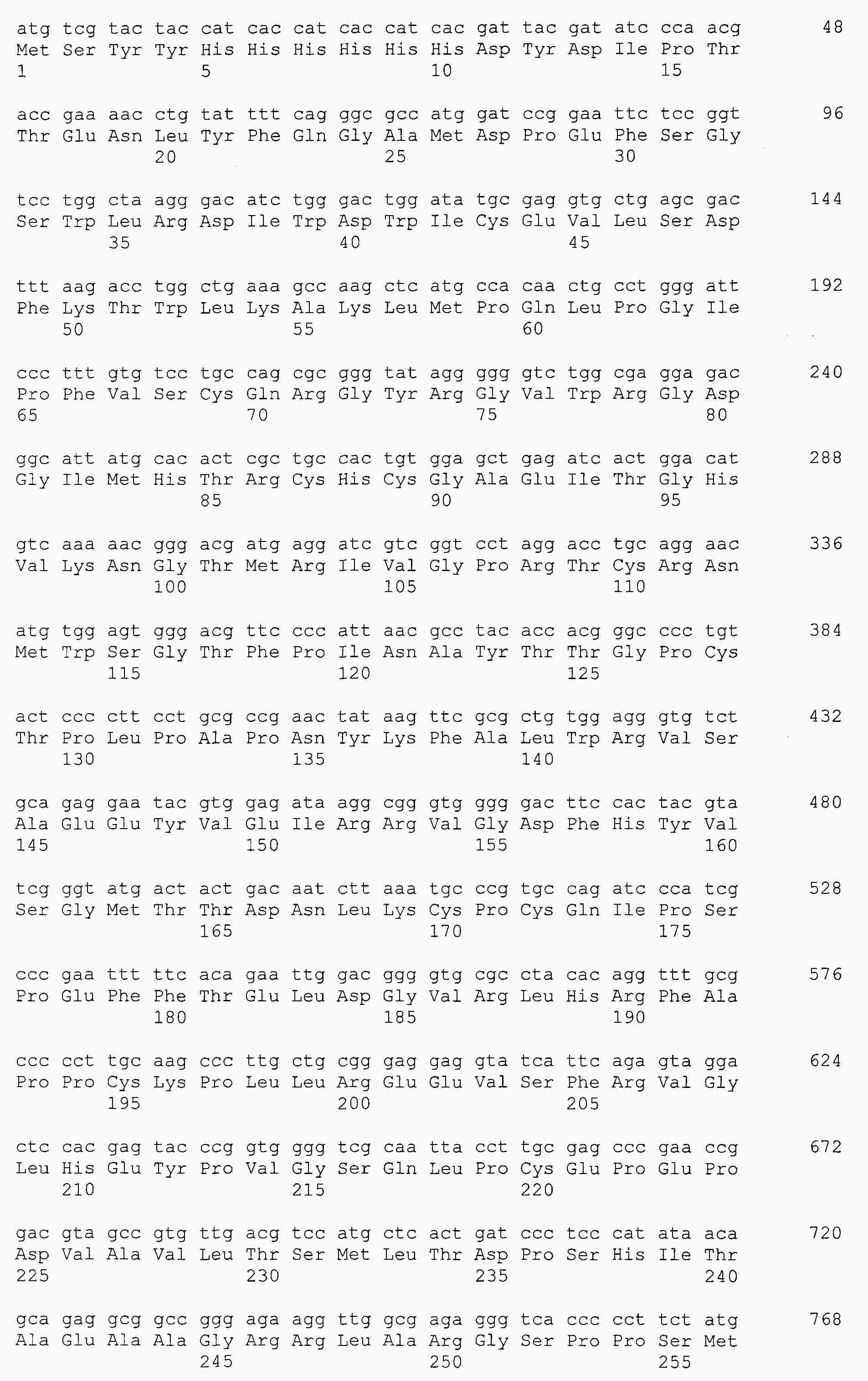
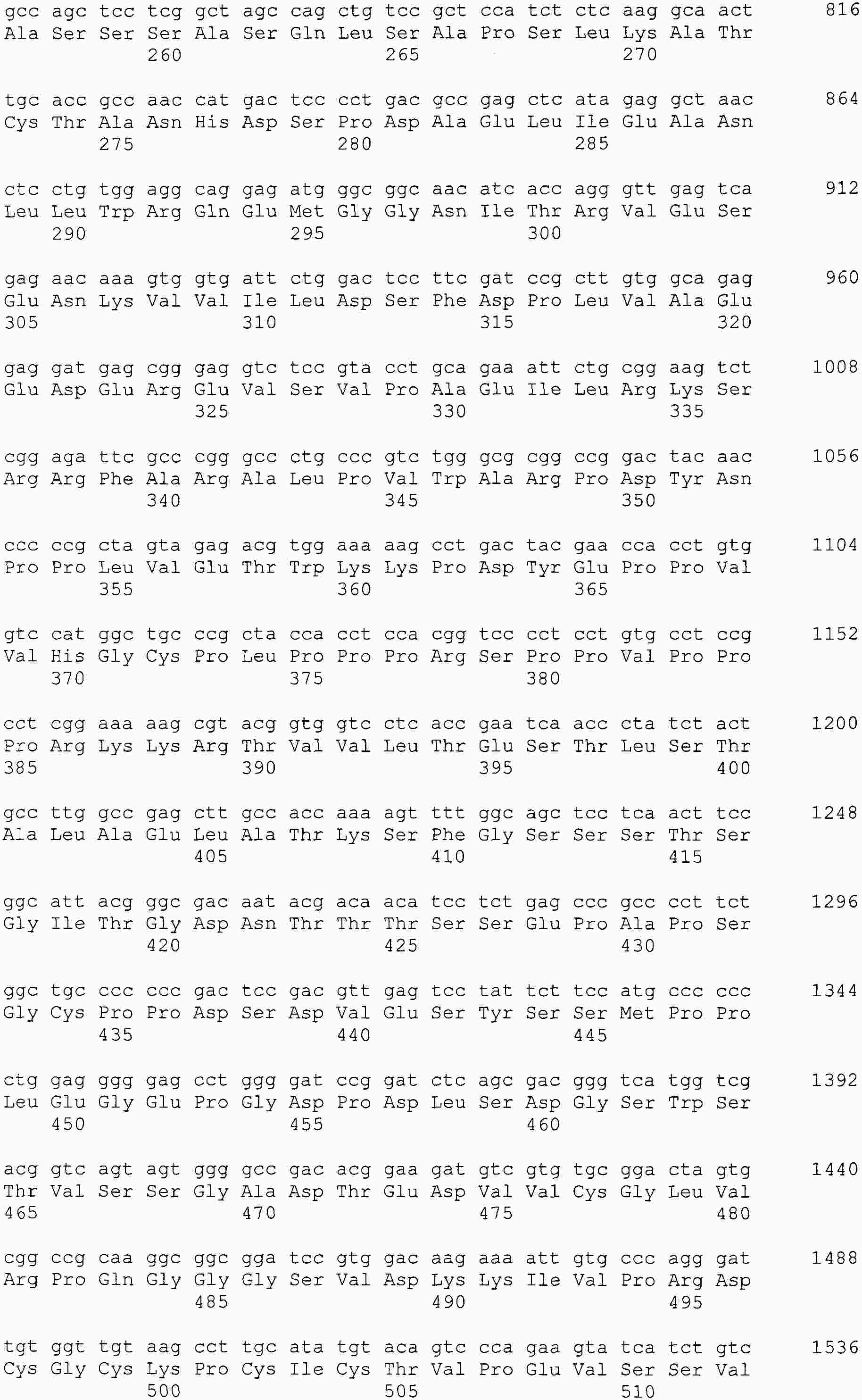
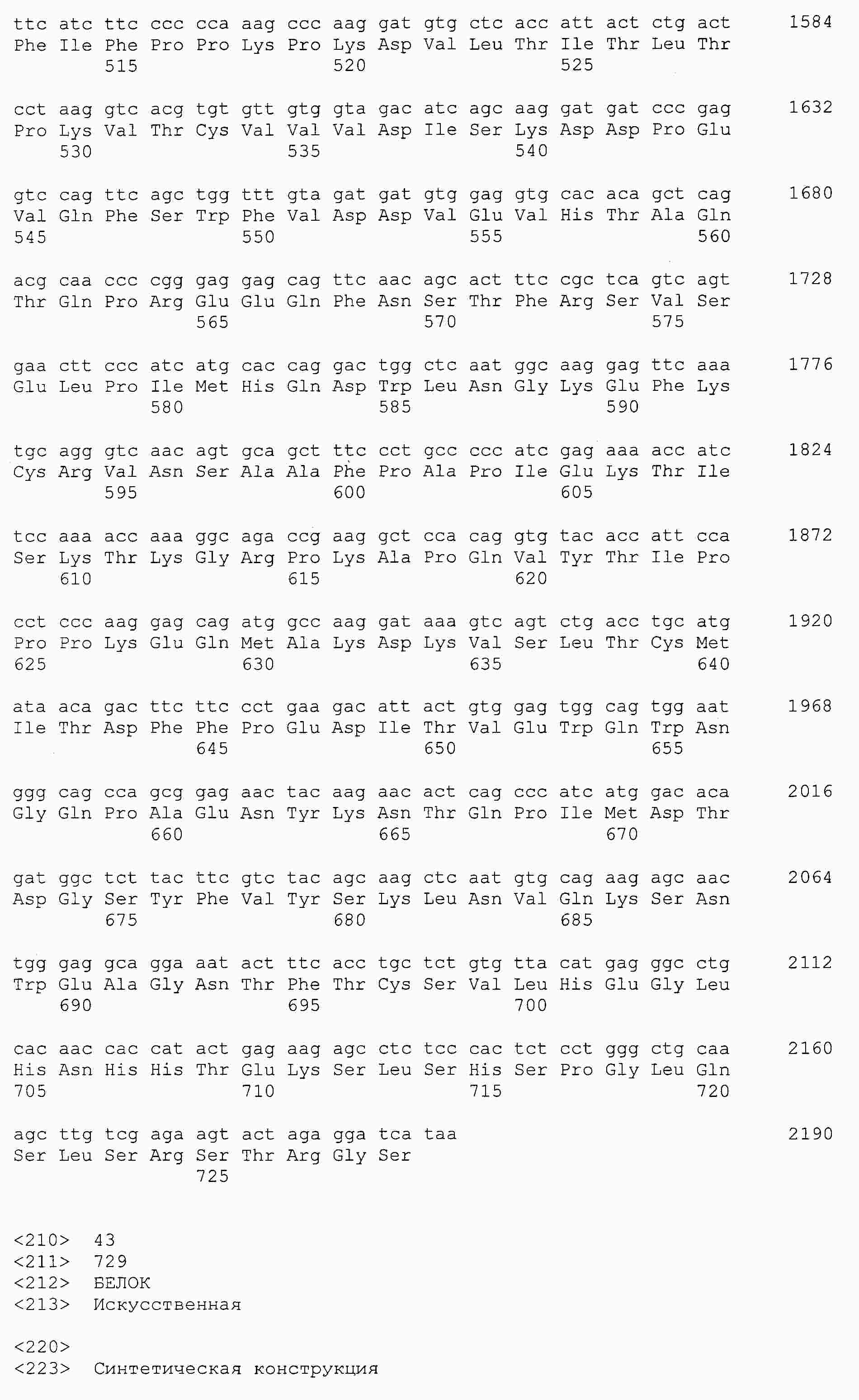
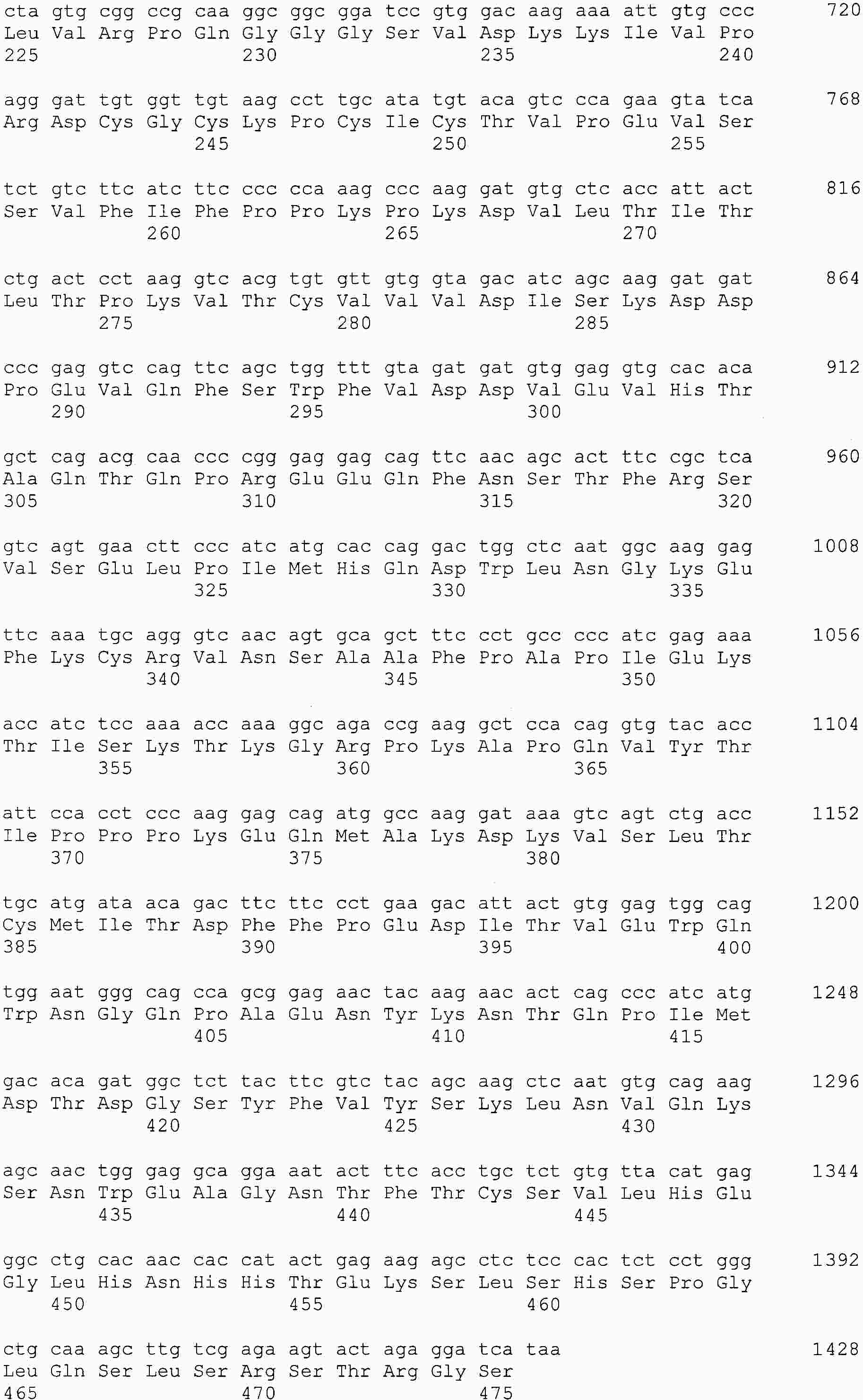
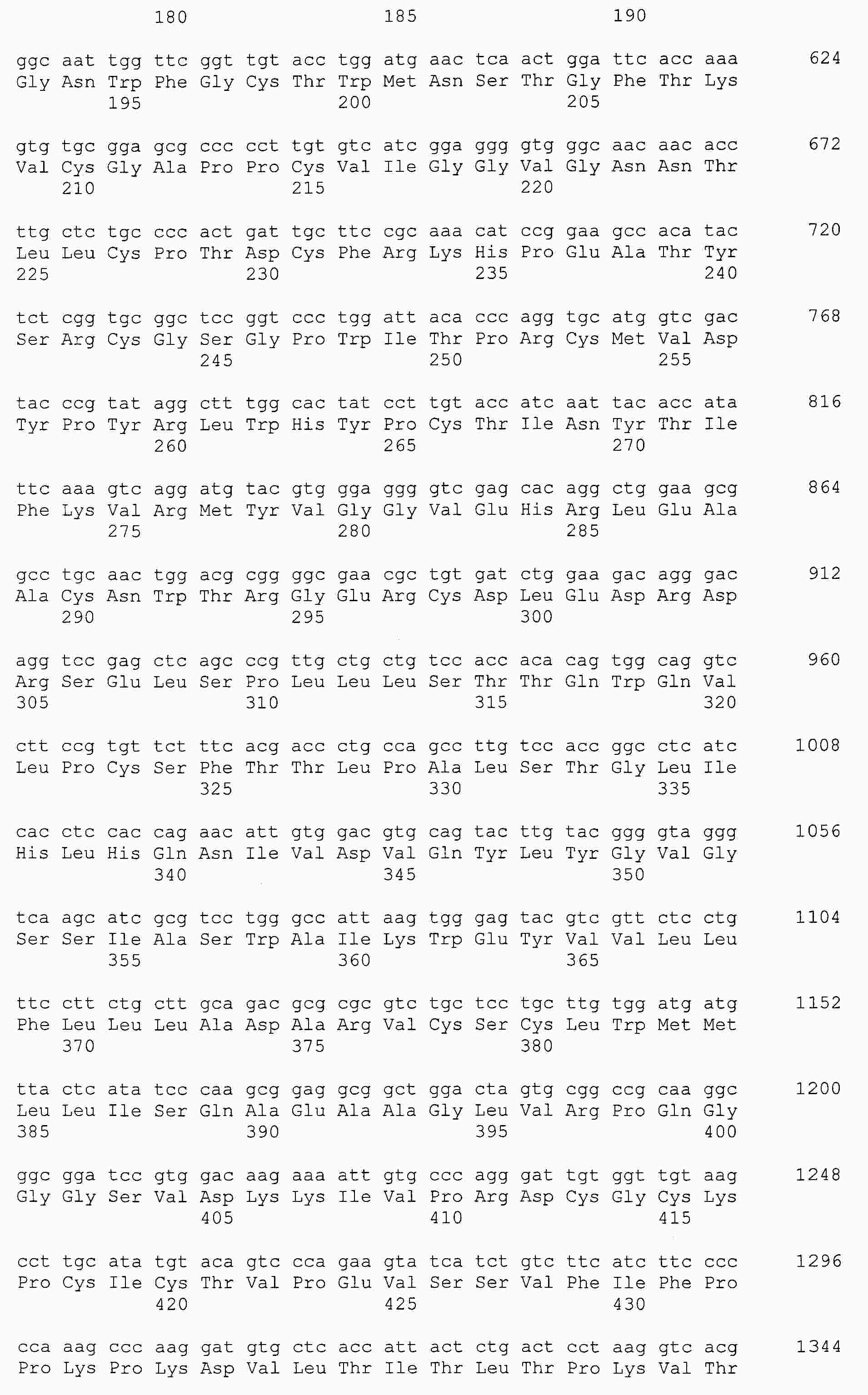
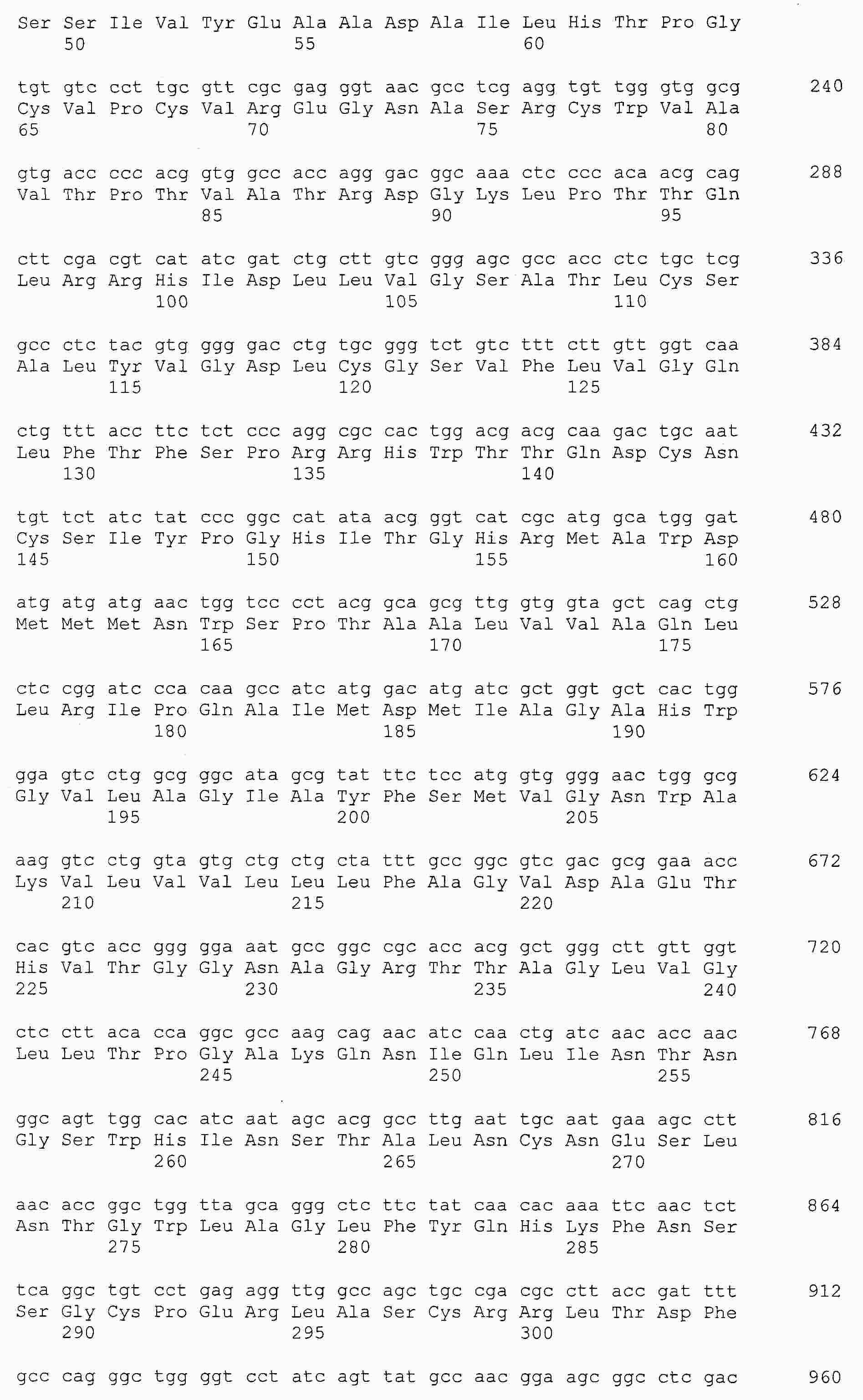
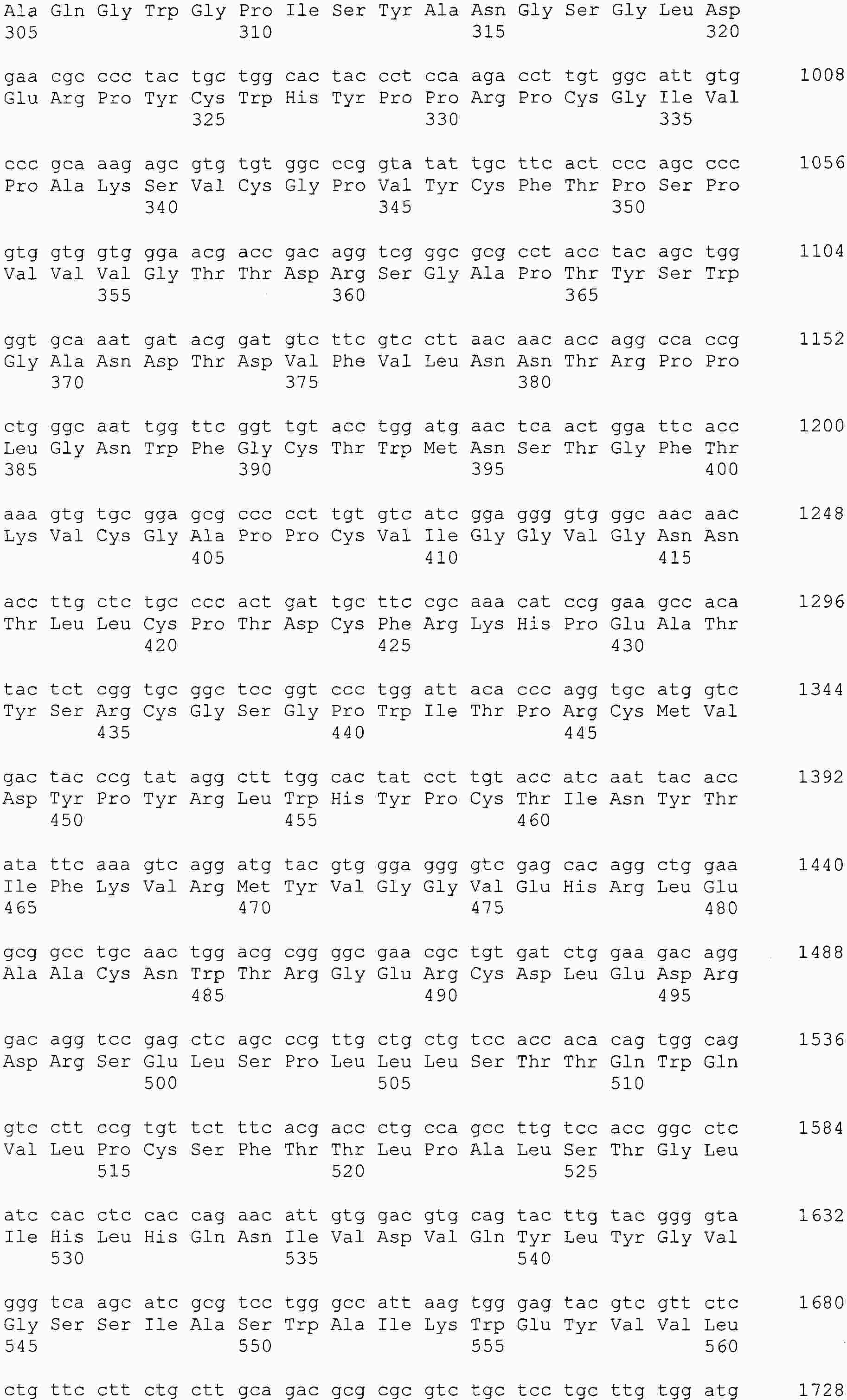
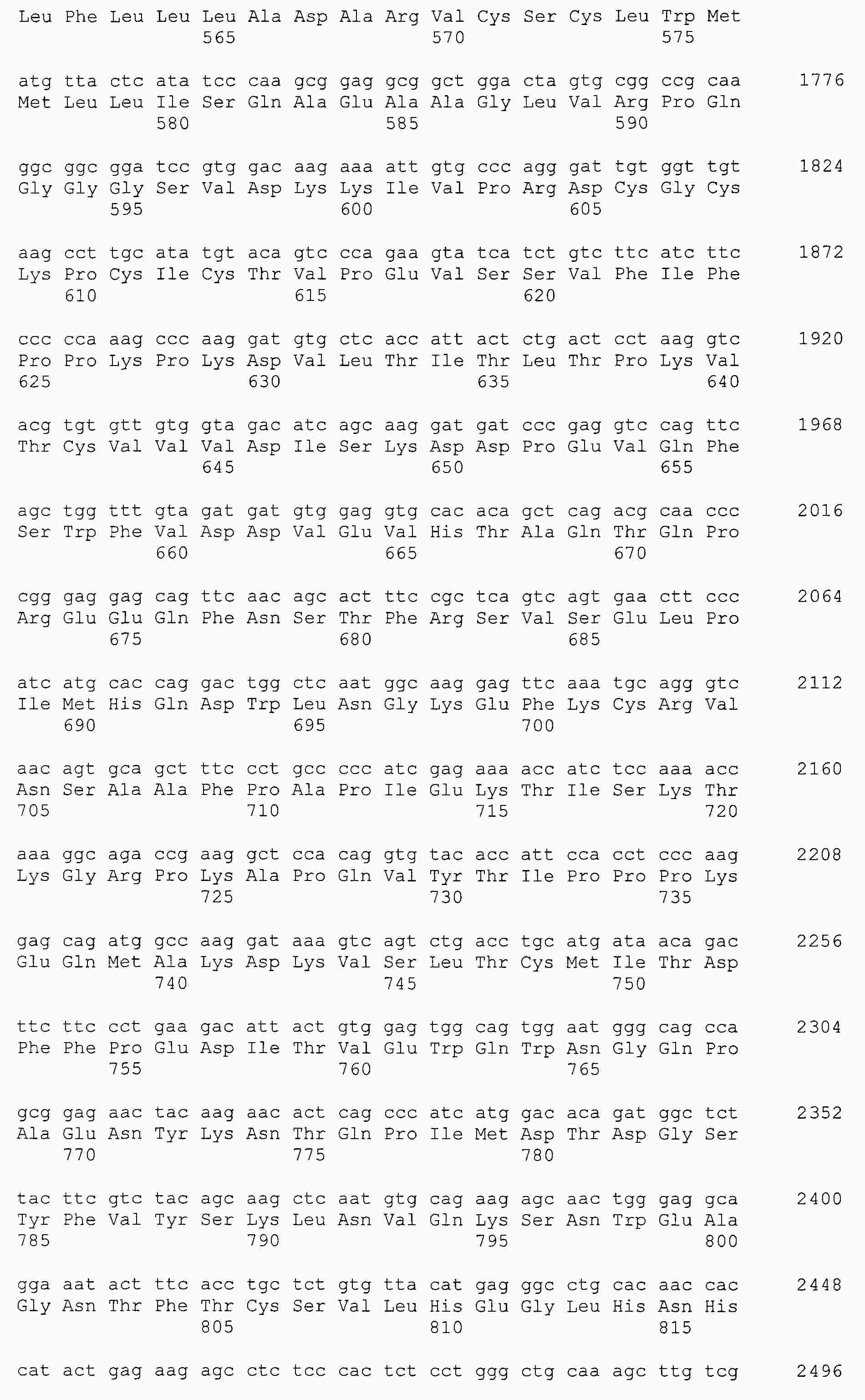
|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000013070b*\id |
|
Термины запроса в документе Реферат Настоящее изобретение относится к композициям и способам генерации иммунного ответа на антигены. Конкретно, раскрыты соединения и способы генерации иммунного ответа на чужеродные антигены, которые в ином случае распознаются хозяином как "собственные" антигены, таким образом, нарушая толерантность хозяина к таким антигенам. Презентация иммунной системе хозяина химерного антигена, содержащего домен, вызывающий иммунный ответ, и домен связывания мишени, где домен связывания мишени содержит фрагмент антитела, усиливает иммунный ответ на чужеродный антиген или антиген, в отношении которого существует толерантность. Антигенпрезентирующие клетки захватывают, подвергают процессингу и презентации химерный антиген, вызывая и гуморальный, и клеточный иммунный ответ на желаемый антиген. Формула [0001] Химерный антиген для генерации иммунного ответа, содержащий по меньшей мере один антигенный домен, где указанный по меньшей мере один антигенный домен содержит по меньшей мере один антигенный участок поверхностного антигена вируса гепатита В (HBV) и домен связывания с антигенпредставляющей клеткой (АРС), где домен связывания с АРС содержит фрагмент антитела, состоящий из фрагмента тяжелой цепи иммуноглобулина (Ig). [0002] Химерный антиген по п.1, в котором фрагмент антитела выбран из группы, состоящей из полноразмерной CH1-области Ig или из части CH1-области Ig и CH2- и CH3-областей Ig. [0003] Химерный антиген по п.1 или 2, содержащий полностью или частично CH2-область Ig, полностью или частично CH3-область Ig или же полностью или частично CH2-область Ig и полностью или частично CH3-область Ig. [0004] Химерный антиген по любому из пп.1-3, в котором фрагмент антитела содержит шарнирную область Ig или ее часть. [0005] Химерный антиген по любому из пп.1-4, в котором фрагмент антитела является фрагментом мышиного антитела. [0006] Химерный антиген по любому из пп.1-5, в котором фрагмент антитела содержит Fc-фрагмент Ig. [0007] Химерный антиген по п.1, в котором антигенный домен связан с меткой и участком расщепления протеазой и домен связывания с АРС содержит одну или несколько аминокислотных последовательностей, выбранных из группы, состоящей из последовательности CH1-домена Ig или его части, CH2-домена Ig или его части и CH3-домена Ig или его части, а антигенный домен связан с доменом связывания с АРС пептидным линкером. [0008] Химерный антиген по любому из пп.1-7, который содержит мономер, содержащий антигенный домен и домен связывания с АРС, или димер, образованный мономерами. [0009] Химерный антиген по любому из пп.2-4 и 7, в котором один или несколько антигенных доменов содержат по меньшей мере одну антигенную часть белка, выбранного из группы, состоящей из белка вируса гепатита В (HBV), белка утиного вируса гепатита В (DHBV) или белка вируса гепатита С (HCV). [0010] Химерный антиген по п.9, в котором белок HBV является поверхностным антигеном HBV. [0011] Химерный антиген по п.10, в котором поверхностный антиген HBV выбран из группы, состоящей из S HBV, S1 HBV, S2 HBV и их комбинаций, корового белка HBV, корового белка HBV ctm и полимеразы HBV. [0012] Химерный антиген по п.10 или 11, в котором поверхностным антигеном HBV является S1/S2 HBV или S1/S2/S HBV. [0013] Химерный антиген по любому из пп.9-12, в котором HBV представляет собой HBV человека. [0014] Химерный антиген по любому из пп.2-4, в котором один или несколько антигенных доменов содержат по меньшей одну антигенную часть белка HCV, выбранного из группы, состоящей из корового белка HCV (1-191), корового белка HCV (1-177), Е1-Е2 HCV, E1 HCV, E2 HCV, NS3 HCV, NS4A HCV и NS5A HCV. [0015] Химерный антиген по любому из пп.1-14, где химерный антиген связан с одной или несколькими метками. [0016] Химерный антиген по п.15, в котором одна или несколько меток содержат по меньшей мере одну метку 6 ×His. [0017] Химерный антиген по п.16, в котором один или несколько антигенных доменов имеют связанную с ними метку 6 ×His. [0018] Химерный антиген по любому из пп.2-5, в котором один или несколько антигенных доменов содержат одну или несколько антигенных частей белка из HPV, HIV, HSV, облигатного внутриклеточного паразита или антигена злокачественной опухоли. [0019] Химерный антиген по п.18, в котором облигатным внутриклеточным паразитом является бактерия. [0020] Химерный антиген по п.19, в котором облигатный внутриклеточный паразит является простейшим. [0021] Химерный антиген по п.1, в котором фрагмент тяжелой цепи Ig содержит полностью или частично CH1-область Ig. [0022] Химерный антиген по любому из пп.1-21, где химерный антиген содержит часть CH1-области Ig, состоящей из аминокислотной последовательности VDKKI (SEQ ID NO:2). [0023] Химерный антиген по любому из пп.1-22, содержащий линкер, который связывает антигенный домен с доменом связывания с АРС. [0024] Химерный антиген по п.23, в котором линкер представляет собой пептид. [0025] Химерный антиген по п.24, в котором пептид содержит последовательность XRPQGGGS (SEQ ID NO:1), где X означает V или S. [0026] Химерный антиген по любому из пп.1-23, в котором и антигенный домен, и домен связывания с АРС дополнительно содержат одну из пары линкерных молекул, которые связывают антигенный домен с доменом связывания с АРС. [0027] Химерный антиген по п.26, в котором пара линкерных молекул выбрана из группы, состоящей из (а) лейциновых молний и (b) биотина и авидина. [0028] Химерный антиген по любому из пп.1-27, где химерный антиген содержит посттрансляционную модификацию. [0029] Химерный антиген по любому из пп.1-28, который содержит гликозилирование. [0030] Химерный антиген по любому из пп.1-29, который содержит гликозилирование маннозой. [0031] Химерный антиген по п.30, в котором один или несколько антигенных доменов содержат гликозилирование маннозой. [0032] Химерный антиген по п.31, в котором домен связывания с АРС содержит гликозилирование маннозой. [0033] Химерный антиген по п.32, который содержит гликозилирование маннозой в низкой степени или гликозилирование маннозой в высокой степени. [0034] Химерный антиген по любому из пп.1-33, содержащий (а) линкерный пептид, состоящий из аминокислотной последовательности XRPQGGGS (SEQ ID NO:1), где X означает S; (b) часть CH1-области Ig, состоящей из аминокислотной последовательности VDKKI (SEQ ID NO: 2); (с) CH2-домен Ig и (d) CH3-домен Ig. [0035] Химерный антиген по любому из пп.1-34, в котором по меньшей мере один антигенный домен содержит множественные эпитопы, которые способны вызывать иммунный ответ. [0036] Химерный антиген по любому из пп.1-35, в котором домен связывания с АРС содержит по меньшей мере один эпитоп, который способен вызывать иммунный ответ. [0037] Химерный антиген по любому из пп.1-36, способный вызывать гуморальный иммунный ответ и/или клеточный иммунный ответ. [0038] Химерный антиген по любому из пп.1-37, способный вызывать (а) иммунный Th1-ответ, (b) иммунный Th2-ответ, (с) CTL-ответ или (d) любое сочетание (а), (b) и (с). [0039] Способ активации АРС, предусматривающий обеспечение контактирования АРС с химерным антигеном по любому из пп.1-38. [0040] Способ по п.39, согласно которому контактирование АРС происходит в условиях ex vivo. [0041] Способ по п.39, согласно которому АРС находится у субъекта in vivo. [0042] Способ по п.39, согласно которому субъектом является человек. [0043] Способ по п.41 или 42, согласно которому контактирование АРС с химерным антигеном вызывает у субъекта (а) иммунный Th1-ответ, (b) иммунный Th2-ответ, (с) иммунный CTL-ответ или (d) любое сочетание (а), (b) и (с). [0044] Способ по пп.41-43, согласно которому один или несколько антигенных доменов содержат антиген, в отношении которого субъект является иммунологически толерантным. [0045] Способ по любому из пп.41-43, согласно которому у субъекта нарушена иммунологическая толерантность. [0046] Способ активации АРС по любому из пп.39-45, согласно которому указанный способ обеспечивает вакцинирование субъекта против HBV посредством введения химерного антигена. [0047] Способ активации АРС по любому из пп.39-46, согласно которому указанный способ обеспечивает терапевтическое или профилактическое вакцинирование субъекта посредством введения химерного антигена. [0048] Способ по любому из пп.41-47, согласно которому введение химерного антигена вызывает более сильный иммунный ответ, чем отдельное введение антигенного домена, где иммунный ответ представляет собой (а) иммунный Th1-ответ, (b) иммунный Th2-ответ, (с) иммунный CTL-ответ или (d) любое сочетание (а), (b) и (с). [0049] Способ по п.48, согласно которому иммунный ответ измеряют по количеству антигенспецифического антитела, присутствующего в организме субъекта. [0050] Способ по п.49, согласно которому количество антигенспецифического антитела измеряют с использованием химерного антигена. [0051] Способ по любому из пп.48-50, согласно которому иммунный ответ измеряют по количеству интерферона- γ, продуцируемого Т-клетками в ответ на их экспозицию с АРС, нагруженными химерным антигеном или только антигенным доменом. [0052] Способ по пп.48-51, согласно которому иммунный ответ измеряют по количеству антигенспецифических CD8+ Т-клеток, полученных в ответ на их экспозицию с АРС, нагруженными химерным антигеном или только антигенным доменом. [0053] Способ по любому из пп.41-52, согласно которому субъект имеет инфекцию или злокачественную опухоль. [0054] Способ по п.53, согласно которому инфекция представляет собой паразитарную инфекцию или вирусную инфекцию. [0055] Способ по п.54, согласно которому вирусная инфекция представляет собой инфекцию HBV. [0056] Способ по любому из пп.53-55, согласно которому инфекция представляет собой хроническую инфекцию. [0057] Способ по п.56, согласно которому хроническая инфекция представляет собой хроническую инфекцию вирусом гепатита С, хроническую инфекцию вирусом папилломы человека, хроническую инфекцию вирусом иммунодефицита человека или хроническую инфекцию вирусом простого герпеса. [0058] Способ по любому из пп.41-57, согласно которому у субъекта развивается иммунный ответ более чем на один эпитоп химерного антигена. [0059] Способ по любому из пп.41-58, согласно которому у субъекта развивается иммунный ответ более чем на один эпитоп одного или нескольких антигенных доменов. [0060] Способ по любому из пп.41-59, согласно которому химерный антиген содержит фрагмент антитела, который является ксенотипическим для субъекта. [0061] Способ по любому из пп.41-60, согласно которому химерный антиген содержит фрагмент антитела, который не является ксенотипическим для субъекта. [0062] Фармацевтическая композиция, содержащая фармацевтически приемлемый наполнитель и химерный антиген по любому из пп.1-61. [0063] Фармацевтическая композиция по п.62 для парентерального введения. [0064] Фармацевтическая композиция по п.62 для трансдермального, интрадермального, внутривенного, подкожного, внутримышечного, назального, легочного, ректального, перорального введения. [0065] Набор, содержащий химерный антиген по любому из пп.1-61, и инструкции для введения химерного антигена нуждающемуся в этом субъекту. [0066] Способ получения химерного антигена, предусматривающий: [0067] Способ по п.66, согласно которому клетка или клеточная линия представляет собой клетку насекомого или линию клеток насекомого. [0068] Способ по п.66, согласно которому микроорганизм или клеточная линия представляет собой эукариотический микроорганизм или клеточную линию. [0069] Способ по п.66, согласно которому микроорганизм или клеточная линия представляет собой дрожжи или линию растительных клеток. [0070] Полинуклеотид, кодирующий химерный антиген по любому из пп.1-60. [0071] Полинуклеотид по п.69, который содержит нуклеотидную последовательность, выбранную из группы, состоящей из нуклеотидов 1-1326 последовательности SEQ ID NO:26, нуклеотидов 1-2004 последовательности SEQ ID NO: 28, нуклеотидов 1-1350 последовательности SEQ ID NO: 30, нуклеотидов 1-1293 последовательности SEQ ID NO: 32, нуклеотидов 1-1794 последовательности SEQ ID NO: 34, нуклеотидов 1-1581 последовательности SEQ ID NO: 36, нуклеотидов 1-1389 последовательности SEQ ID NO: 38, нуклеотидов 1-1347 последовательности SEQ ID NO: 40, нуклеотидов 1-2157 последовательности SEQ ID NO: 42, нуклеотидов 1-1395 последовательности SEQ ID NO: 44, нуклеотидов 1-1905 последовательности SEQ ID NO: 46 или нуклеотидов 1-2484 последовательности SEQ ID NO: 48. [0072] Полинуклеотид по п.69, который кодирует химерный антиген, который по меньшей мере на 90% идентичен полной аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 1-442 последовательности SEQ ID NO:27, аминокислот 1-668 последовательности SEQ ID NO: 29, аминокислот 1-450 последовательности SEQ ID NO: 31, аминокислот 1-431 последовательности SEQ ID NO: 33, аминокислот 1-598 последовательности SEQ ID NO: 35, аминокислот 1-527 последовательности SEQ ID NO: 37, аминокислот 1-463 последовательности SEQ ID NO: 39, аминокислот 1-449 последовательности SEQ ID NO: 41, аминокислот 1-719 последовательности SEQ ID NO: 43, аминокислот 1-465 последовательности SEQ ID NO: 45, аминокислот 1-635 последовательности SEQ ID NO: 47 и аминокислот 1-828 последовательности SEQ ID NO: 49. [0073] Полинуклеотид, который избирательно гибридизуется в жестких условиях с полинуклеотидом, имеющим нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46 и 48. [0074] Микроорганизм, содержащий полинуклеотид по любому из пп.70-72. [0075] Линия клеток, содержащая полинуклеотид по любому из пп.70-72. [0076] Вектор, содержащий полинуклеотид по любому из пп.70-72. Полный текст патента I. Введение A. Связанные заявки. Данная заявка претендует на приоритет предварительной заявки США № 60/493449, поданной 8 августа 2003 г, включенной в настоящее описание в качестве ссылки. B. Техническая область. Настоящее изобретение относится к способам и композициям для получения или усиления иммунного ответа и для нарушения толерантности хозяина к чужеродным антигенам. C. Предпосылки. Когда здоровый хозяин (человек или животное) сталкивается с чужеродным антигеном, таким как происходящий из бактерии, вируса и/или паразита белок, обычно у хозяина возникает иммунный ответ. Данный иммунный ответ может представлять собой гуморальный ответ и/или клеточный ответ. При гуморальном ответе в ответ на антигенный стимул В-клетки продуцируют антитела и секретируют их в кровь и/или лимфу. Затем антитела нейтрализуют антиген, например вирус, специфически связываясь с антигенами на его поверхности, помечая их для разрушения фагоцитирующими клетками и/или опосредованными комплементом механизмами или блокируя связывание или увеличивая выведение свободного антигена из циркуляции. Клеточный ответ характеризуется отбором и экспансией специфических хелперных или цитотоксических Т-лимфоцитов, способных непосредственно или опосредованно уничтожать содержащие антиген клетки. У некоторых индивидов иммунная система не способна к ответу на определенные чужеродные антигены. Когда антиген не стимулирует продукцию специфических антител и/или киллерных Т-клеток, иммунная система не в состоянии предотвратить результирующее заболевание. Таким образом, возбудитель инфекции, например вирус, может вызывать хроническую инфекцию, а иммунная система хозяина приобретает толерантность к антигенам, продуцируемым данным возбудителем инфекции. Хотя механизм, по которому возбудитель инфекции уклоняется от иммунных механизмов хозяина, недостаточно понятен, отсутствие правильной презентации чужеродных антигенов иммунной системе хозяина может являться фактором, способствующим развитию хронических инфекций. Антигенпрезентирующие клетки (АРС) процессируют обнаруженные антигены различным образом в зависимости от локализации антигена. Экзогенные антигены подвергаются эндоцитозу, а затем - процессингу в эндосомах антигенпрезентирующей клетки. Образующиеся из экзогенных антигенов пептидные фрагменты презентируются на поверхности клетки в комплексе с главным комплексом гистосовместимости (МНС) класса II. Презентация данного комплекса CD4 + Т-клеткам стимулирует хелперные CD4 + Т-клетки к секреции цитокинов, стимулирующих В-клетки к продукции антител к экзогенному антигену (гуморальный ответ). С другой стороны, внутриклеточные антигены подвергаются процессингу и презентации в виде комплексов с МНС класса I на поверхности антигенпрезентирующих клеток. Антигенная презентация CD8 + Т-клеткам приводит к иммунному ответу цитотоксическими Т-клетками (CTL) против клеток хозяина, несущих антиген. У субъектов с хроническими вирусными или паразитическими инфекциями (когда организм на некоторой стадии в течение его жизненного цикла постоянно обитает внутри клетки хозяина) антигены продуцируются клеткой хозяина и экспрессируются в ней, а секретируемые антигены присутствуют в циркуляции. В качестве примера, у хронических носителей вируса человеческого гепатита В (HBV) в крови можно выявить вирионы и поверхностные антигены HBV и суррогат сердцевинных антигенов (в виде е-антигена). Для эффективной терапии хронической инфекции необходим сильный CTL-ответ на ассоциированные с возбудителем инфекции антигены. Этого можно достичь или продукцией антигена в клетке хозяина, или посредством доставки антигена в соответствующий клеточный компартмент, так, чтобы он подвергался процессингу и презентации таким образом, чтобы вызывать иммунный ответ. В литературе описано несколько подходов для доставки антигена внутрь клетки. Среди них описаны вирусные векторы (Lorenz et al., Hum. Gen. Ther. 10:623-631 (1999)), применение трансфицированных кДНК клеток (Donnelly et al., Ann. Rev. Immunol. 15:617 (1997)), а также экспрессия антигена посредством инъецируемых кДНК-векторов (Lai et al., Crit. Rev. Immunol. 18:449-484 (1988) и патент США № 5589466). Кроме того, описаны ДНК-вакцины, экспрессирующие антигены, являющиеся мишенью дендритных клеток (You, et al., Cancer Res 61:3704-3711 (2001)). Также применяли средства доставки, способные доставлять антигены в цитозольный компартмент клетки для пути процессинга МНС класса I. HiIgers, et al. (Vaccine 17:219-228 (1999)) подробно описали применение адъювантов для достижения той же цели. Другой подход представляет собой применение биологически разрушаемых микросфер для доставки антигенов в цитоплазму, иллюстрируемое получением иммунного ответа Th1 на пептид яичного альбумина (Newman, et al., J Control Release 54:49-59 (1998) и Newman, et al., J Biomed Mater Res 50:591-597 (2000)). Кроме того, антигенпрезентирующие клетки, например дендритные клетки, захватывают наносферы PLGA (Newman, et al., J Biomed Mater Res 60:480-486 (2002)). Способность дендритных клеток захватывать, подвергать процессингу и презентации антиген и стимулировать наивные Е-клетки делает их очень важным средством для разработки терапевтических вакцин (Laupeze, et al., Hum Immunol 60:591-597 (1999)). Нацеливание антигена на дендритные клетки представляет собой решающую стадию в антигенной презентации, и для этой цели использовали присутствие на дендритных клетках нескольких рецепторов для Fc-области антител (Regnault, et al., J Exp Med 189:371-380 (1999)). Дополнительные примеры данного подхода включают в себя Mab-В43.13 к раку яичников, антитело к PSA, a также комплексы антигена и антитела к HBV (Wen, et al., Int Rev Immunol 18:251-258 (1999)). Показано, что иммунотерапия злокачественных опухолей с применением дендритных клеток, нагруженных ассоциированными с опухолями антигенами, вызывает опухолеспецифический иммунный ответ и противоопухолевую активность (Fong and Engleman, Ann Rev Immunol 96:1865-1972 (2000) и Campton, et al., J Invest Dermatol 115:57-61 (2000)). Многообещающие результаты получены при клинических испытаниях in vivo с применением сенсибилизированных опухолевыми антигенами дендритных клеток (Tarte and Klein, Leukemia 13:653-663 (1999)). Данные исследования отчетливо демонстрируют эффективность применения дендритных клеток для получения иммунного ответа на опухолевые антигены. Антигенную презентацию также можно проводить посредством маннозных рецепторов, вместо применения рецептора Fc на антигенпрезентирующих клетках или в дополнение к нему. Макрофагальный маннозный рецептор (MMR), также известный как CD206, экспрессируется на таких антигенпрезентирующих клетках, как дендритные клетки (DC). Данная молекула является представителем семейства лектинов С-типа эндоцитируемых рецепторов. CD206 может связывать и интернализировать маннозилированные антигены. Как правило, считают, что экзогенный антиген подвергается процессингу и презентации, главным образом, посредством пути МНС класса II. Однако в случае направления через CD206 существуют доказательства того, что вовлечены оба пути МНС класса I и класса II (Apostolopoulos et al., Eur. J. Immunol. 30:1714 (2000); Apostolopoulos and McKenzie, Curr. Mol. Med. 1:469 (2001); Ramakrishna et al., J. Immunol. 172:2845-2852 (2004)). Инфекционное заболевание и злокачественная опухоль являются главными проблемами государственного здравоохранения. Например, статистика Всемирной Организации Здравоохранения показывает, что более чем 2 миллиарда человек были инфицированы HBV. Среди них 370 миллионов инфицированы хронически и, как результат - повышенная вероятность развития цирроза печени и печеночно-клеточного рака. Приблизительно 170 миллионов человек во всем мире являются хроническими носителями HCV, для которого не существует эффективной профилактической или терапевтической вакцины. По сообщениям Всемирной Организации Здравоохранения, у 10 миллионов человек ежегодно диагностируют злокачественную опухоль. Злокачественная опухоль служит причиной 6 миллионов смертных случаев ежегодно, 12% смертных случаев во всем мире. Таким образом, существует необходимость в новых терапевтически эффективных композициях и способах достижения иммунного ответа против инфекций и злокачественной опухоли, а также в новых способах получения таких композиций. Изобретение относится к химерным антигенам для индукции иммунного ответа, где химерные антигены содержат домен, вызывающий иммунный ответ, и домен связывания мишени, где домен связывания мишени содержит фрагмент антитела. В другом аспекте изобретение относится к способам усиления антигенной презентации у антигенпрезентирующей клети, включающим в себя приведение антигенпрезентирующей клетки в контакт с содержащей химерный антиген композицией согласно изобретению. В еще одном аспекте изобретение относится к способам активации антигенпрезентирующих клеток, включающим в себя приведение антигенпрезентирующих клеток в контакт с химерным антигеном согласно изобретению. Один из аспектов изобретения относится к способам получения иммунного ответа, включающим в себя введение субъекту содержащей химерный антиген композиции согласно изобретению. В другом аспекте изобретение относится к способам нарушения толерантности, включающим в себя введение субъекту химерного антигена согласно изобретению. В предпочтительном варианте осуществления субъект хронически инфицирован вирусом или облигатным внутриклеточным паразитом. Один из аспектов изобретения относится к способам лечения поддающегося иммунотерапии состояния, включающим в себя введение нуждающемуся в этом субъекту терапевтически эффективного количества химерного антигена согласно изобретению. В предпочтительном варианте осуществления поддающееся иммунотерапии состояние представляет собой инфекцию, особенно хроническую инфекцию, или злокачественную опухоль. В еще одном аспекте изобретение относится к способам вакцинации субъекта против инфекции, включающим в себя введение субъекту химерного антигена согласно настоящему изобретению. Субъекта можно вакцинировать профилактически или терапевтически. В предпочтительном варианте осуществления у субъекта развивается иммунный ответ более чем к одному эпитопу химерного антигена, а более предпочтительно более чем к одному эпитопу домена, вызывающего иммунного ответа. Предпочтительно инфекция представляет собой вирусную инфекцию или инфекцию облигатными внутриклеточными паразитами. Другой аспект согласно изобретению относится к фармацевтической композиции, содержащей химерный антиген согласно настоящему изобретению и фармацевтически приемлемый носитель. Один из аспектов изобретения относится к готовым изделиям, содержащим химерный антиген согласно изобретению и инструкции для введения химерного изобретения нуждающемуся в этом субъекту. Другой аспект согласно изобретению относится к кодирующим химерный антиген полинуклеотидам, где указанный полинуклеотид содержит первую полинуклеотидную часть, кодирующую домен, вызывающий иммунный ответ, и вторую полинуклеотидную часть, кодирующую домен связывания мишени, где домен связывания мишени содержит фрагмент антитела. Изобретение также относится к микроорганизмам и клеточным линиям, содержащим такие полинуклеотиды. В еще одном аспекте изобретение относится к способам получения химерного антигена согласно изобретению, включающим в себя предоставление микроорганизма или клеточной линии, содержащих полинуклеотид, кодирующий химерный антиген согласно изобретению, и культивирование микроорганизма или клеточной линии в условиях, когда экспрессируется химерный антиген. Подробности одного или нескольких вариантов осуществления изобретения указаны в сопроводительных чертежах и описании ниже. Другие особенности, цели и преимущества изобретения станут понятны из описания и чертежей, и из формулы изобретения. На фиг. 1А представлена схематическая диаграмма, иллюстрирующая структуру химерного антигена согласно настоящему изобретению в виде мономера, где химерный антиген обладает двумя частями, а именно доменом, вызывающим иммунный ответ, и доменом связывания мишени. На схеме также проиллюстрирован предпочтительный вариант осуществления изобретения, в котором существует шарнирная область. На фиг. 1В представлена схематическая диаграмма, иллюстрирующая структуру химерного антигена согласно настоящему в его обычном состоянии, собранным в виде димера. Данная схема иллюстрирует особенно предпочтительный вариант осуществления изобретения, в котором химерный антиген, в дополнение к домену, вызывающему иммунный ответ, и домену связывания мишени, содержит метку 6xHis и пептидный линкер. На фиг. 2 показано, что стимуляция Т-клеток S1/S2-TBD HBV вызывает ответ цитотоксических Т-клеток (CTL) конкретно на эпитоп из белка S1 HBV. На фиг. 3 показано, что стимуляция Т-клеток S1/S2-TBD HBV вызывает ответ цитотоксических Т-клеток (CTL) конкретно на эпитоп из белка S2 HBV. На фиг. 4 показано сравнение захвата S1/S2-TBD HBV, IgG1 и IgG2 созревающими дендритными клетками в виде функции от концентрации. На фиг. 5 показана корреляция связывания S1/S2-TBD HBV с экспрессией CD32 и CD206 на дендритных клетках. А. Обзор. Здесь описаны композиции и способы получения иммунного ответа на антигены. В частности, соединения и способы вызывают иммунный ответ на чужеродные антигены, которые в ином случае распознаются хозяином как "собственные" антигены, нарушая, таким образом, толерантность к этим антигенам. Презентация иммунной системе хозяина химерного антигена, содержащего домен, вызывающий иммунный ответ, и домен связывания мишени, где домен связывания мишени содержит фрагмент антитела, усиливает иммунный ответ на чужеродный антиген или антиген, в отношении которого существует толерантность. Антигенпрезентирующие клетки захватывают, подвергают процессингу и презентации химерный антиген, вызывая и гуморальный и клеточный иммунный ответ на желаемый антиген. В. Определения. Перед дальнейшим подробным описанием изобретения применяемые в данной заявке термины определяют, как указано ниже, если не указано иначе. "Антитело" относится к продуцируемой В-лимфоцитами молекуле иммуноглобулина со специфической аминокислотной последовательностью, индуцируемой у человека или животных антигеном (иммуногеном). Данные молекулы характеризуются специфическим взаимодействием с антигеном, где каждый определен на основании другого. "Ответ антителами", или "гуморальный ответ", относится к типу иммунного ответа, при котором в ответ на антигенный стимул В-лимфоциты продуцируют антитела и секретируют их в кровь и/или лимфу. При правильно функционирующем иммунном ответе, антитело специфически связывается с антигенами на поверхности клеток (например, патоген), маркируя клетку для разрушения фагоцитирующими клетками и/или опосредованными комплементом механизмами. Антитела также циркулируют системно и могут связываться со свободными вирионами. Данное связывание антител может нейтрализовать вирион и предотвратить инфицирование им клетки, а также маркируя его для удаления из циркуляции фагоцитозом или фильтрацией в почках. "Антиген" относится к любому веществу, которое в результате вступления в контакт с соответствующими клетками индуцирует состояние восприимчивости и/или иммунологической реактивности и которое явным образом реагирует с антителами и/или иммунными клетками сенсибилизированных субъектов in vivo или ex vivo. "Антигенпрезентирующая клетка" относится к вспомогательным клеткам процесса индукции антигеном, главным образом функционирующим процессируя и презентируя антиген лимфоцитам. Взаимодействие антигенпрезентирующих клеток с антигенами представляет собой важнейшую стадию иммуноиндукции, так как оно дает возможность лимфоцитам встретить и распознать антигенные молекулы и активироваться. Примеры антигенпрезентирующих клеток включают в себя макрофаги, дендритные клетки Лангерганса, фолликулярные дендритные клетки и В-клетки. "В-клетка" относится к типу лимфоцитов, продуцирующих иммуноглобулины или антитела, взаимодействующие с антигенами. "Область C H 1" относится к области константного домена тяжелой цепи на антигенсвязывающем фрагменте антитела. "Клеточный ответ" относится к типу иммунного ответа, опосредованному специфическими хелперными или киллерными Т-клетками, способными непосредственно устранять зараженные вирусом или злокачественные клетки. Здесь термин "химерный антиген" относится к полипептиду, содержащему домен, вызывающий иммунный ответ и домен связывания мишени. Домен, вызывающий иммунный ответ и домены связывания мишени могут быть прямо или опосредованно связаны ковалентными или нековалентными связями. "Цитотоксический Т-лимфоцит" представляет собой специализированный тип лимфоцита, способный к разрушению чужеродных клеток и инфицированных возбудителями инфекции клеток хозяина, продуцирующих вирусные антигены. "Эпитоп" относится к простейшей форме антигенной детерминанты, на молекуле сложного антигена; он представляет собой конкретную часть антигена, распознаваемую иммуноглобулином или рецептором Т-клетки. "Гибридный белок" относится к белку, формируемому экспрессией гибридного гена, полученного объединением двух или более генных последовательностей. "Шарнирная область" относится к части антитела, соединяющей Fab-фрагмент с Fc-фрагментом; шарнирная область содержит дисульфидные мостики, ковалентно связывающие две тяжелые цепи. Термин "гомолог" относится к молекуле, проявляющей гомологию с другой молекулой, например, обладая последовательностями химических остатков, являющимися идентичными или сходными в соответствующих положениях. Фраза "гомологичный на столько-то %" или "% гомологии" относится к проценту нуклеотидов или аминокислот в одном и том же положении гомологичных полинуклеотидов или полипептидов, являющихся идентичными или сходными. Например, если 75 из 80 остатков в двух белках идентичны, то два этих белка гомологичны на 93,75%. Процент гомологии можно определить с применением различного программного обеспечения, известного специалисту в данной области. "Хозяин" относится к теплокровному животному, включая человека, страдающему от поддающегося иммунотерапии состояния, такого как инфекция или злокачественная опухоль. Здесь, "хозяин" также относится к теплокровному животному, включая человека, которому вводят химерный антиген. В контексте данного изобретения "гибридизация" означает спаривание комплементарных цепей олигомерных соединений. В настоящем изобретении предпочтительный механизм спаривания включает в себя образование водородных связей, которое может быть по Уотсону-Крику, хугстиновским или обратным хугстиновским образованием водородных связей, между комплементарными нуклеозидными или нуклеотидными основаниями (нуклеиновые основания) цепей олигомерных соединений. Например, аденин и тимин являются комплементарными нуклеиновыми основаниями, спаривающимися посредством образования водородных связей. Гибридизация может происходить в различных условиях. Подразумевают, что термины "гибридизоваться", "гибридизующийся", "гибридизуется" и т.п., применяемые в контексте полинуклеотидов, относятся к общепринятым условиям гибридизации, предпочтительно таким как гибридизация в 50% формамиде/6 ×SSC/0,1% SDS/100 мкг/мл мДНК, где температуры гибридизации составляют выше 37 °С, а температуры отмывки в 0,1 ×SSC/0,1% SDS составляют выше 55 °С. "Иммунитет" или "иммунный ответ" относится к ответу организма на антиген. В конкретных вариантах осуществления он относится к способности антитела противостоять инфекционному заболеванию или защищать себя от него. "Домен, вызывающий иммунный ответ (IRD)" относится к разным образом сконфигурированной антигенной части бифункциональной молекулы химерного антигена. Домен, вызывающий иммунный ответ, содержит один или несколько антигенов и/или один или несколько рекомбинантных антигенов. Здесь фраза "поддающееся иммунотерапии состояние" относится к состоянию или заболеванию, которое можно предотвратить, подавить или ослабить, получая или модулируя иммунный ответ у субъекта. "Лимфоцит" относится к субпопуляции ядерных клеток, находящихся в крови, опосредующих специфический иммунный ответ. "Моноклональное антитело" или "mAb", относится к антителу, продуцируемому клоном или генетически гомогенной популяцией слитых гибридных клеток, т.е. гибридомными клетками. Гибридные клетки клонируют для получения клеточных линий, продуцирующих конкретные моноклональные антитела, являющиеся химически и иммунологически гомогенными, т.е. распознающие только один тип антигена. "Пептидная связь" относится к двум или нескольким аминокислотам, ковалентно связанным замещенной амидной связью между альфа-аминогруппой одной аминокислоты и альфа-карбоксильной группой другой аминокислоты. "Фармацевтически приемлемая" относится к нетоксической композиции, физиологически пригодной для человека или животных. "Фармацевтически приемлемый наполнитель" включает в себя такое вещество, как адъюванты, носители, средства для регуляции рН и буферные средства, регулирующие концентрацию средства, увлажняющие средства, консерванты и т.п., являющиеся нетоксичными и физиологически приемлемыми для человека или животных. Здесь термин "полинуклеотид" относится к полимерной форме нуклеотидов любой длины, как к рибонуклеотидам, так и к дезоксирибонуклеотидам. Данный термин относится только к первичной структуре молекулы. Таким образом, данный термин включает в себя двух- и одноцепочечную ДНК и РНК. Он также включает в себя известные типы модификаций, например известные в данной области метки, метилирование, "кэпы", замещение одного или нескольких встречающихся в природе нуклеотидов аналогом, межнуклеотидные модификации, такие, например, как модификации с незаряженными связями (например, метилфосфонаты, фосфотриэфиры, фосфоамидаты, карбаматы и т.д.) и с заряженными связями (например, фосфоротиоаты, фосфородитиоаты и т.п.), модификации с дополнительными боковыми группами, такими как белки (включающие в себя, например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и т.п.), модификации с интеркаляторами (например, акридином, псораленом и т.д.), модификации, содержащие хелаторы (например, металлы, радиоактивные металлы, бор, окислительные металлы и т.д.), модификации, содержащие алкилирующие средства, модификации с модифицированными связями (например, альфа-аномерные нуклеиновые кислоты и т.д.), а также модифицированные формы полинуклеотидов. "Участок расщепления протеазами" относится к участку, в котором протеолитические ферменты гидролизуют (разрывают) полипептидные цепи. В настоящем изобретении фраза "жесткие условия гибридизации" или "жесткие условия" относится к условиям, в которых соединение согласно изобретению гибридизуется с его последовательностью-мишенью, но с минимальным числом других последовательностей. Термин "субъект" относится к любому теплокровному животному, предпочтительно человеку. "Метка" относится к маркеру или маркерной последовательности, применяемой для выделения или очистки молекулы, содержащей метку. Пример метки включает в себя метку 6 ×His. "Т-клетка" относится к типу лимфоцитов, ответственному за антигенспецифические клеточные взаимодействия и опосредующему гуморальный и клеточный иммунный ответ. "Домен связывания мишени (TBD)" относится к белку, содержащему фрагмент антитела, способный к связыванию с рецептором на антигенпрезентирующей клетке, особенно на дендритной клетке, и впоследствии транспортируемый внутрь антигенпрезентирующей клетки путем опосредуемого рецептором захвата. Фраза "терапевтически эффективное количество" относится к количеству химерного антигена или полинуклеотида, кодирующего химерный антиген, достаточному для получения эффективного В-клеточного ответа, ответа цитотоксических Т-лимфоцитов (CTL) и/или ответа хелперных Т-лимфоцитов (Th) на антиген, и для блокирования или для лечения, или, по меньшей мере, для частичной задержки или замедления развития симптомов и/или осложнений заболевания или нарушения. Здесь термины "обработка" и "лечение" относятся к любому лечению состояния, поддающегося у животных, особенно у человека, лечению химерным антигеном, и включают в себя (i) профилактику данного состояния у субъекта, который может быть предрасположен к данному состоянию, но у которого еще не установили его наличие; (ii) ингибирование состояния, например, путем задержки или замедления его развития или (iii) ослабление состояния, например, путем индукции регрессии состояния или его симптомов. Здесь "ксенотипический" относится к происходящему из другого вида, отличного от хозяина. Например, рекомбинантно экспрессированное антитело, клонированное из мышиного генома, будет являться ксенотипическим для человека, но не для мыши, вне зависимости от того, получено ли рекомбинантно экспрессируемое антитело в бактериальной клетке, клетке насекомого или мыши. С. Новые химерные антигены. Изобретение относится к химерным антигенам для получения иммунного ответа, содержащим домен, вызывающий иммунный ответ, и домен связывания мишени, где домен связывания мишени содержит фрагмент антитела. Согласно настоящему изобретению предпочтительно, чтобы химерный антиген был способен к связыванию с Fc-рецептором и/или с макрофагальным маннозным рецептором. Фрагмент антитела может быть ксенотипическим для хозяина или не быть ксенотипическим для хозяина. В предпочтительных вариантах осуществления изобретения химерный антиген способен к индукции гуморального и/или клеточного иммунного ответа. Клеточный иммунный ответ может включать в себя ответ Th1, ответ Th2 и/или ответ цитотоксических Т-лимфоцитов (CTL). В еще одном предпочтительном варианте осуществления изобретения химерный антиген вызывает полиэпитопный иммунный ответ. Полиэпитопный иммунный ответ может включать в себя ответ по меньшей мере на один эпитоп домена, вызывающего иммунный ответ и/или ответ по меньшей мере на один эпитоп домена связывания мишени. Альтернативно, полиэпитопный ответ может быть ограничен ответом более чем к одному эпитопу домена, вызывающего иммунный ответ. Химерный антиген согласно настоящему изобретению содержит две части, а именно домен, вызывающий иммунный ответ, содержащий антигенную последовательность (такую как вирусный антиген) и домен связывания мишени, содержащий фрагмент антитела (фиг. 1). В предпочтительном варианте осуществления домен, вызывающий иммунный ответ, может быть связан с доменом связывания мишени любым известным специалистам в данной области способом. Связующие звенья для связывания домена, вызывающего иммунный ответ, с доменом связывания мишени могут включать в себя в качестве неограничивающих примеров ковалентные пептидные связи, химическую конъюгацию, лейциновые молнии и биотин/авидин. В предпочтительном варианте осуществления домен, вызывающий иммунный ответ, и домен связывания мишени клонированы в виде одного гибридного белка. Ковалентная пептидная связь гибридного белка может включать в себя дополнительные пептидные последовательности, такие как SRPQGGGS или VRPQGGGS (SEQ ID №1). В еще одном предпочтительном варианте осуществления изобретения биотинилируют различные домены, вызывающие иммунный ответ, а домен связывания мишени получают в виде гибридного белка со стрептавидином для облегчения получения широкого ассортимента химерных антигенов. Альтернативно, каждый из доменов, вызывающих иммунный ответ, и домена связывания мишени можно экспрессировать в виде гибрида с мотивом лейциновой молнии, обуславливающим ассоциацию двух частей химерного антигена при смешивании. Наконец, домен, вызывающий иммунный ответ, и домены связывания мишени можно экспрессировать отдельно, а затем химически конъюгировать с применением известных специалисту в данной области способов. Иллюстративные примеры включают в себя применение средств для создания перекрестных сшивок белков, таких как диметилсуберимидат для ковалентного соединения двух доменов. Домен, вызывающий иммунный ответ, главным образом обеспечивает антигенную часть химерного антигена. Домен, вызывающий иммунный ответ, содержит по меньшей мере одну антигенную часть объекта, на который желателен иммунный ответ. Химерный антиген, необязательно, может содержать более чем один домен, вызывающий иммунный ответ. В предпочтительных вариантах осуществления изобретения домен, вызывающий иммунный ответ, содержит по меньшей мере одну антигенную часть возбудителя инфекции, такого как вирус или облигатный внутриклеточный паразит, или антигена злокачественной опухоли. Более предпочтительно, чтобы домен, вызывающий иммунный ответ, содержал по меньшей мере одну антигенную часть инфекционного вируса. Примеры предпочтительных инфекционных вирусов включают в себя: Retroviridae (например, вирусы человеческого иммунодефицита, такие как вирус человеческого иммунодефицита-1 (HIV-1), также обозначаемый как HTLV-III, LAV или HTLV-III/LAV или HIV-III; и другие изоляты, такие как HIV-LP; Picornaviridae (например, вирусы полиомиелита, вирус гепатита А; энтеровирусы, человеческие вирусы Коксаки, риновирусы, ЕСНО-вирусы); Calciviridae (например, вызывающие гастроэнтерит штаммы); Togaviridae (например, вирусы лошадиного энцефалита, вирусы краснухи); Flaviridae (например, вирус гепатита С, вирусы денге, вирусы энцефалита, вирусы желтой лихорадки); Coronaviridae (например, коронавирусы); Rhabdoviridae (например, вирусы везикулярного стоматита, вирусы бешенства); Filoviridae (например, вирусы Эбола); Paramyxoviridae (например, вирусы парагриппа, вирус эпидемического паротита, вирус кори, респираторно-синцитиальный вирус); Orthomyxoviridae (например, вирусы гриппа); Bungaviridae (например, вирусы Хантаан, бунъявирусы, флебовирусы и найровирусы); Arenaviridae (вирус геморрагической лихорадки); Reoviridae (например, реовирусы, орбивирусы и ротавирусы); Birnaviridae; Hepadnaviridae (вирус человеческого гепатита В (HBV), вирус гепатита В уток (DHBV)); Parvoviridae (парвовирусы); Papovaviridae (вирусы папилломы, вирусы полиомы); Adenoviridae (большинство аденовирусов); Herperviridae (вирус простого герпеса (HSV) 1 и 2, вирус ветряной оспы, цитомегаловирус (CMV), вирусы Эпштейна-Барр, вирусы герпеса); Poxviridae (вирусы оспы человека, вирусы коровьей оспы, поксвирусы) и Iridoviridae (например, вирус африканской лихорадки свиней) и неклассифицированные вирусы (например, возбудитель гепатитов дельта, возбудители не-А, не-В гепатитов (класс 1 передающиеся энтерально); класс 2 - передающиеся парентерально; вирусы Норфолк и родственные вирусы и астровирусы). В некоторых вариантах осуществления изобретения домен, вызывающий иммунный ответ, химерного антигена содержит по меньшей мере одну антигенную часть одного или нескольких белков, выбранных из группы, состоящей из белков HBV, белков DHBV и белков HCV. Особенно предпочтительные для применения в настоящем изобретении белки HBV включают в себя в качестве неограничивающих примеров S1/S2 HBV, S1/S2/S HBV, коровый белок HBV, коровый белок HBV ctm (модифицированный по С-концу), е-антиген HBV и полимеразу HBV. Особенно предпочтительные для применения в настоящем изобретении белки DHBV включают в себя в качестве неограничивающих примеров PreS/S DHBV, PreS DHBV, коровый белок DHBV и полимеразу DHBV. Особенно предпочтительные для применения в настоящем изобретении белки HCV включают в себя в качестве неограничивающих примеров коровый белок HCV (1-191), коровый белок HCV (1-177), Е1-Е2 HCV, E1 HCV, E2 HCV, NS3 HCV, NS5A и NS4A HCV. Другие предпочтительные вирусные антигены для применения в настоящем изобретении включают в себя gp120 HIV, щелочную нуклеазу HSV и капсидные белки L1 и L2 вируса человеческой папилломы и белки ранней области E1 HPV, E2 HPV, Е4 HPV, Е5 HPV, Е6 HPV и Е7 HPV. Примеры предпочтительных облигатных внутриклеточных паразитов включают в себя: виды Tetrahymena (например, Т. pyriformis), виды Plasmodium (например, P. falciparum), виды Cryptospiridium, виды Spraguea (например, S. lophii), виды Giardia, виды Toxoplasma (например, Т. gondii, T. cruzi), виды Leishmania, виды Rickettsia (например, R. prowazekii), виды Chlamydia, виды Mycobacterium (например, М. tuberculosis), виды Legionella, виды Listeria, (например, L. monocytogenes), виды Coxiella (например, С. brunette), виды Shigella, виды Erlichia и виды Bartonelia. Предпочтительные антигены злокачественных опухолей включают в себя: специфический антиген простаты (PSA), специфический мембранный антиген простаты (PSMA), MUC1, СА 125, WT1, Her-2/neu, эмбриональный опухолевый антиген (СЕА), MAGE-3, MART-1, gp100, NY-ESS0-1, СА19.9, TAG72, СА15.3, СА 27.9, gp120, простатическую кислую фосфатазу (РАР), белки теплового шока, альфа-фетопротеин (AFP), теломеразу и ras. В еще одном варианте осуществления изобретения домен, вызывающий иммунный ответ, химерного антигена включает в себя метку 6 ×His, слитую с одной или несколькими антигенными частями. Химерный антиген по настоящему изобретению представляет собой белок, который способен связываться с Fc-рецептором и/или CD206 на антигенпрезентирующей клетке, особенно на дендритной клетке, и потом перемещаться внутрь антигенпрезентирующей клетки посредством опосредуемого рецептором захвата. Согласно настоящему изобретению присутствие фрагмента антитела увеличивает захват химерного антигена посредством Fc-рецептора на антигенпрезентирующих клетках, а конкретно - на дендритных клетках. Вследствие этого специфического связывания и интернализации вирусный антиген подвергается процессингу и презентации как чужеродный. Таким образом, можно эффективно вызывать иммунный ответ к антигену, в отношении которого ранее существовала толерантность. Домен связывания мишени содержит фрагмент антитела, который может являться ксенотипическим для хозяина или может являться не ксенотипическим для хозяина. В предпочтительном варианте осуществления изобретения фрагмент антитела содержит мышиный Fc-фрагмент. В более предпочтительном варианте осуществления изобретения домен связывания мишени содержит Fc-фрагмент, шарнирную область и часть области C H 1, а химерный антиген содержит пептидный линкер, пригодный для соединения домена связывания мишени с доменом, вызывающим иммунный ответ. В другом предпочтительном варианте осуществления изобретения домен связывания мишени содержит фрагмент тяжелой цепи иммуноглобулина и, необязательно, дополнительно содержит шарнирную область. В особенно предпочтительном варианте осуществления изобретения фрагмент тяжелой цепи содержит аминокислоты VDKKI (SEQ ID № 2) домена C H 1 и/или часть или все из доменов C H 2 и C H 3. Как обсуждали выше, антигены, связывающиеся и интернализующиеся посредством CD206, могут быть представлены посредством МНС и класса I, и класса II, таким образом вызывая и клеточный, и гуморальный иммунный ответ. Следовательно, в предпочтительном варианте осуществления химерный антиген гликозилирован. Гликозилированным может быть домен, вызывающий иммунный ответ, и/или домен связывания мишени. В особенно предпочтительном варианте осуществления изобретения химерный антиген гликозилирован маннозой или в высокой степени гликозилирования маннозой, или в низкой степени гликозилирования маннозой (Jarvis, Virology 310:1-7 (2003)). D. Новые способы применения химерных антигенов. Изобретение относится к способам вызова иммунного ответа, включающим в себя введение субъекту содержащей химерный антиген композиции согласно изобретению. Для обеспечения эффективной антигенной презентации, авторы изобретения разработали новый химерный антиген, содержащий домен, вызывающий иммунный ответ, и домен связывания мишени, где домен связывания мишени содержит фрагмент антитела. Не являясь ограниченным конкретной теорией согласно изобретению, данная молекула, посредством фрагмента антитела, связывается с специфическими рецепторами на антигенпрезентирующих клетках, а вирусный антиген подвергается процессингу и презентации в комплексе с главным комплексом гистосовместимости (МНС) класса I и класса II. Такие процессинг и презентация антигена посредством МНС класса I вызывает усиленный ответ цитотоксическими Т-лимфоцитами, приводя к устранению любого возбудителя инфекции, ассоциированного с антигеном домена, вызывающего иммунный ответ. Кроме того, антигенная презентация посредством молекул МНС класса II вызывает гуморальный ответ, который также помогает в выведении антигена из инфицированной клетки и/или из циркуляции. Изобретение также относится к способам нарушения толерантности, включающим в себя введение субъекту химерного антигена согласно изобретению. При антигенной презентации для вызова клеточного и/или гуморального иммунного ответа с применением химерных антигенов, антигены, которые при хронической инфекции обрабатываются как "собственные", распознаются как "чужеродные". Таким образом, иммунная система хозяина запускает CTL-ответ для устранения инфицированных клеток. Одновременно образующиеся в ответ на химерный антиген антитела связываются с возбудителем инфекции и удаляют его из циркуляции или блокируют связывание возбудителя инфекции с клетками хозяина. Таким образом, настоящее изобретение предназначено для получения химерных антигенов, способных индуцировать полиспецифический иммунный ответ у субъектов с хроническими инфекциями, в отношении которых в ином случае существует толерантность иммунной системы хозяина. В предпочтительном варианте осуществления химерный антиген нарушает толерантность к антигену у субъекта, хронически инфицированного возбудителем инфекции, таким как вирус или паразит, или у субъекта со злокачественной опухолью. Более предпочтительно, чтобы возбудитель инфекции находился внутри клетки хозяина на некоторой стадии в течение его жизненного цикла. В предпочтительном варианте осуществления иммуногенность предварительно отобранного антигена, не распознающегося иммунной системой хозяина или в отношении которого развивается толерантность иммунной системы хозяина, увеличивается вследствие присутствия фрагмент антитела, а также вследствие наличия гликозилирования, проведенного в эукариотической клеточной экспрессирующей системе, например насекомого. Такой химерный антиген вследствие присутствия компонента антитела и гликозилирования связывается со специфическими рецепторами, находящимися на различных типах иммунных клеток, включающих в себя дендритные клетки, макрофаги, В-клетки и гранулоциты. В еще одном аспекте изобретение относится к способам активации антигенпрезентирующих клеток, включающим в себя приведение антигенпрезентирующих клеток в контакт с химерным антигеном согласно изобретению. Изобретение также относится к способам усиления антигенной презентации в антигенпрезентирующих клетках, включающих в себя приведение антигенпрезентирующей клетки в контакт с содержащей химерный антиген композицией согласно изобретению. Химерный антиген можно приводить в контакт с антигенпрезентирующими клетками, предпочтительно дендритными клетками, in vivo или ex vivo. В предпочтительном варианте осуществления контакт химерного антигена с антигенпрезентирующими клетками активирует антигенпрезентирующие клетки и усиливает антигенную презентацию более чем одного эпитопа. Данный полиэпитопный ответ может включать в себя презентацию одного или нескольких эпитопов домена, вызывающего иммунный ответ, и/или презентацию одного или нескольких эпитопов домена связывания мишени. Изобретение также относится к способам лечения поддающегося иммунотерапии состояния, включающим в себя введение нуждающемуся в этом субъекту терапевтически эффективного количества химерного антигена согласно изобретению. В предпочтительном варианте осуществления поддающееся иммунотерапии состояние представляет собой инфекцию или злокачественную опухоль. Инфекция может быть вирусной инфекцией, паразитарной инфекцией или бактериальной инфекцией. Предпочтительно инфекция протекает со стадией, в течение которой возбудитель инфекции находится в клетке хозяина. Более предпочтительно поддающееся иммунотерапии состояние представляет собой хроническую вирусную инфекцию. Наиболее предпочтительно поддающееся иммунотерапии состояние представляет собой хроническую инфекцию вирусом гепатита В (HBV) или хроническую инфекцию вирусом гепатита С (HCV). Для лечения HBV домен, вызывающего иммунный ответ, предпочтительно содержит по меньшей мере одну антигенную часть белка, выбранного из группы, состоящей из корового белка HBV, белка HBV S, белка HBV S1, белка HBV S2 и их сочетаний. Для лечения HCV домен, вызывающий иммунный ответ, предпочтительно содержит по меньшей мере одну антигенную часть белка, выбранного из группы, состоящей из корового белка HCV (1-191), корового белка HCV (1-177), белка HCV E1, белка HCV Е2, белка HCV E1-E2, белка HCV NS3A, белка HCV NS5A и их сочетаний. В предпочтительном варианте осуществления введение химерного антигена вызывает более сильный иммунный ответ, чем введение одного домена, вызывающего иммунный ответ. Амплитуду иммунного ответа можно измерить, например, (i) посредством количества специфичного к антигену антитела, присутствующего у субъекта; (ii) посредством количества интерферона- γ, секретируемого Т-клетками в ответ на контакт с антигенпрезентирующей клеткой, нагруженной химерным антигеном или одним доменом, вызывающим иммунный ответ, или (iii) посредством количества специфичных к антигену CD8 + Т-клеток, полученных в ответ на контакт с антигенпрезентирующей клеткой, нагруженной химерным антигеном или одним доменом, вызывающим иммунный ответ. Химерный антиген можно оценивать по его эффективности при получении иммунного ответа представлением химерного антигена дендритным клеткам ex vivo или in vivo. Дендритные клетки подвергаются процессингу и презентации химерный антиген Т-лимфоцитам, которые оценивают на пролиферацию Т-клеток и на продукцию интерферона- γ в качестве маркеров Т-клеточного ответа. Конкретно, в случае ex vivo из крови выделяют наивные дендритные клетки. Активацию Т-клеток дендритными клетками оценивают известными способами посредством измеряющих маркеров, например уровней интерферона- γ. См., например, Berlyn, et al., Clin. Immunol 101 (3):276-283 (2001). Увеличение процента Т-клеток, секретирующих интерферон- γ, по меньшей мере на 50% предсказывает эффективность in vivo. В случае in vivo химерный антиген непосредственно парентерально вводят хозяину, где доступные дендритные и другие антигенпроцессирующие клетки имеют возможность взаимодействовать с антигенами для соответствующего их процессинга. Кроме того, изобретение относится к способам вакцинации субъекта от инфекции, включающим в себя введение субъекту химерного антигена согласно настоящему изобретению. Субъекта можно вакцинировать профилактически или терапевтически. Предпочтительно инфекция представляет собой вирусную инфекцию. Бифункциональная природа молекулы помогает направить антиген к антигенпрезентирующим клеткам, например дендритным клеткам, делая ее уникальным подходом в терапии хронических инфекционных заболеваний посредством специфического направления к антигенпрезентирующим клеткам с наиболее эффективным стехиометрическим отношением антигена к антителу. Это пригодно для разработки терапевтических вакцин для хронических вирусных инфекций, таких как гепатит В, гепатит С, вирус человеческого иммунодефицита, вирус человеческой папилломы и простой вирус герпеса, облигатные внутриклеточные паразиты, и можно применять для всех аутологических антигенов при таких заболеваниях, как злокачественная опухоль и аутоиммунные нарушения. Введение данных гибридный белков может вызвать у хозяина полиспецифический иммунный ответ, включая и клеточный, и гуморальный ответ. Таким образом, кроме их пригодности в качестве профилактических вакцин для иммунизации субъектов с риском развития конкретной инфекции, их можно применять в качестве терапевтических вакцин для лечения субъектов, иммунологически толерантных к существующей инфекции. Е. Способы получения химерных антигенов. Один из аспектов изобретения относится к способам получения химерного антигена, включающим в себя (а) получение микроорганизма или клеточной линии, предпочтительно эукариотической, более предпочтительно микроорганизма или клеточной линии, не относящихся к млекопитающим, содержащих полинуклеотид, кодирующий химерный антиген; и (b) культивирование указанных микроорганизма или клеточной линии в условиях, когда экспрессируется химерный антиген. Предпочтительно микроорганизм или клеточная линия представляют собой дрожжи, клеточную линию растений или клеточную линию насекомых. Более предпочтительно клеточная линия представляет собой клеточную линию насекомых, выбранную из группы, состоящей из Sf9, Sf21, Drosophila S2 и High Five ™. В одном из вариантов осуществления настоящего изобретения для получения гибридных белков выбранного антигена(ов) и домена связывания мишени, которые необходимы для осуществления изобретения на практике, применяют общепризнанную технологию рекомбинантных ДНК. Конструкции гибридных белков получают на уровне ДНК, вводя участки распознавания специфических рестрикционных ферментов, используемые при введении фрагмента желаемой ДНК в экспрессирующие векторы и применяемые для экспрессии желаемых гибридных белков в гетерологичной экспрессирующей системе. Как применяют здесь, термин "вектор" означает способные нести ДНК плазмиды, кодирующие желательный белок(ки). Предпочтительные плазмидные векторы для применения по настоящему изобретению включают в себя в качестве неограничивающих примеров pFastBac HTa и соответствующие рекомбинантные "бакмиды", полученные в DH10Bac ™ E. coli (Invitrogen). Кодирующий домен связывания мишени ген можно получить из любой продуцирующей антитела клетки, например гибридомы, продуцирующей моноклональное антитело, посредством полимеразной цепной реакции (ПЦР). Для облегчения более поздних этапов клонирования предпочтительно конструировать праймеры с добавлением уникальных участков распознавания рестрикционных ферментов. Аналогично, антигенные части домена, вызывающего иммунный ответ, можно получать из любой клеточной или вирусной РНК или ДНК, содержащих ген, кодирующий антигенную часть желаемой мишени. Предпочтительно ПЦР применяют для получения ДНК, кодирующей антигенные части домена, вызывающего иммунный ответ, а праймеры для ПЦР конструируют с добавлением уникальных участков распознавания рестрикционными ферментами для облегчения клонирования. Однако для получения ДНК, кодирующей антигенные части домена, вызывающего иммунный ответ, можно применять любой способ рекомбинантных ДНК. Затем кодирующие домен связывания мишени и домен, вызывающий иммунный ответ, полинуклеотиды можно объединять в одну конструкцию с применением стандартных способов клонирования. Альтернативно, отдельные домены можно клонировать с ДНК, кодирующей линкеры, такие как лейциновые молнии, стрептавидин или сигналы биотинилирования. Предпочтительно для экспрессии химерного антигена согласно изобретению применяют бакуловирусную систему, не только вследствие продукции больших количеств гетерологичных белков, но также вследствие посттрансляционных модификаций эукариотических белков, происходящих в инфицированной клетке насекомого, таких как фосфорилирование и гликозилирование. Так как прямое клонирование в клетки насекомых может быть сложным, предпочтительно получить кодирующий химерный антиген полинуклеотид в бактериальной системе и перенести полученную в результате конструкцию в экспрессирующую систему бакуловирус/клетка насекомого. Системы переноса, например система Вас-То-Вас ™ (Invitrogen), известны специалистам в данной области. В системе Bac-to-Bac ™ для переноса представляющего интерес гена в челночный вектор Е.coli-клетка насекомого (бакмиду) применяют сайт-специфическую транспозицию с бактериальным транспозоном Tn7. Полученные рекомбинантные бакмиды трансфицируют в клетки насекомых для получения экспрессирующих рекомбинантные белки бакуловирусов. Для получения бакуловирусов бакмиду трансфицируют в клетки насекомых, такие как клетки Sf9. После трансфекции клетки инкубируют в течение периода времени, достаточного для увеличения популяции бакуловирусов. Содержащую бакуловирусы среду собирают и хранят при 4 °С в темноте. Трансфекцию можно подтвердить посредством проверки на продукцию бакуловирусной ДНК, подвергая вирусную культуру ПЦР с применением праймеров, специфичных для желаемой вставки ДНК. Экспрессию гетерологического белка в клетках можно подтвердить любым известным в данной области способом, например электрофорезом в полиакриламидном геле с SDS (SDS-PAGE) или "вестерн-блоттингом". Для инфицирования клеток насекомых применяют рекомбинантные бакмиды стандартизированной множественности заражения (MOI). Клетки высевают при плотности приблизительно 3 ×10 5 клеток/мл и инкубируют в суспензионной культуре при 27,5 °С при перемешивании, пока клеточная плотность не достигает приблизительно 2-3 ×10 6 клеток/мл. Затем к клеткам добавляют стандартизированные количества соответствующего рекомбинантного бакуловируса. Температура инкубации составляет 27,5 °С, а соответствующий период заражения стандартизуют для экспрессии отдельных белков. Клетки собирают центрифугированием и используют для очистки рекомбинантных белков. Неиспользуемые порции клеток можно быстро замораживать в жидком азоте и хранить при -70 °С. Предпочтительно химерные антигены очищают в денатурирующих условиях. Экспрессирующие химерные антигены клетки лизируют в денатурирующем буфере, например буфере, содержащем 6М гуанидиний-HCl. Лизис можно усилить механическими средствами, такими как разрушение ультразвуком. Затем лизат центрифугируют для удаления неразрушенных клеток и клеточного дебриса. Затем супернатант наносят на колонку с бусинами Ni-NTA Super Flow (Qiagen), предварительно уравновешенную буфером для лизиса. После нанесения колонку отмывают буферизованным денатурирующим раствором, предпочтительно содержащим 6М гуанидиний-HCl, приблизительно при рН 8. На данном этапе денатурирующее средством можно заменить, например, на 8М мочевину в буферизованном растворе. Буферы для лизиса, нанесения и отмывки предпочтительно содержат низкую концентрацию, например 1-40 мМ имидазола. После замены буфера колонку следует отмывать буфером до тех пор, пока OD 280 не снизится, например, до <0,1. Связанный белок можно элюировать буфером, содержащим 8М мочевину и 250 мМ имидазол, рН 8 (элюирующий буфер). Содержащие белок фракции объединяют и подвергают диализу при 4 °С против многократных смен низкоконцентрированного (например, 100 мМ) солевого денатурирующего буфера для диализа, предпочтительно содержащего 8М мочевину. Диализированный белок затем наносят на ионообменную колонку, такую как колонку с DEAE (диэтиламиноэтил). В предпочтительном варианте осуществления до нанесения на ионообменную колонку к белку добавляют дитиотреитол (DTT) или другой восстановитель. Химерный антиген пропускают через колонку с DEAE. Следовательно, проходящий поток DEAE собирают и поэтапно подвергают диализу против буферов, содержащих уменьшающиеся концентрации денатурирующего средства. В иллюстративном способе белок затем подвергают диализу против буферизованной 4М мочевины в течение по меньшей мере 12 ч, затем против буферизованной 2М мочевины в течение по меньшей мере 12 ч, затем против буферизованной 1М мочевины в течение по меньшей мере 12 ч, затем против буферизованной 0,5М мочевины в течение по меньшей мере 12 ч и, наконец, подвергают диализу против буфера, не содержащего денатурирующего средства в течение по меньшей мере 12 ч, предпочтительно с последующими двумя дополнительными периодами диализа по 12 ч против свежего буфера, не содержащего денатурирующего средства. Очищенные подвергнутые рефолдингу белки можно концентрировать и охарактеризовывать с применением стандартных биохимических способов, включающих в себя, например, электрофорез в геле с SDS, изоэлектрофокусирование и анализ "вестерн-блот" с применением антител к различным доменам экспрессируемого белка. При практическом применении настоящего изобретения, если не указано иначе, применяют традиционные способы молекулярной биологии, микробиологии, рекомбинантных ДНК и иммунологии, известные специалистам в данной области. Такие способы полностью объяснены в литературе. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, 2 nd Ed., New York: Cold Spring Harbor Press, 1989; и Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, ( ©1995, as Supplemented April 2004, Supplement 66). F. Новые полинуклеотиды. Другой аспект согласно изобретению относится к кодирующим химерный антиген полинуклеотидам, содержащим первую полинуклеотидную часть, кодирующую домен, вызывающий иммунный ответ, и вторую полинуклеотидную часть, кодирующую домен связывания мишени. Первая и вторая полинуклеотидные части могут быть расположены на одной и той же или различных нуклеотидных цепях. Изобретение относится к полинуклеотидам, соответствующим или комплементарным генам, кодирующим химерные антигены, мРНК и/или кодирующим последовательностям, предпочтительно в изолированной форме, включая кодирующие варианты белков химерных антигенов полинуклеотиды; ДНК, РНК, гибриды ДНК/РНК и родственные молекулы, полинуклеотиды или олигонуклеотиды, комплементарные или обладающие по меньшей мере 90% гомологии с генами, кодирующими химерный антиген или последовательностями мРНК или их части, и полинуклеотиды или олигонуклеотиды, гибридизующиеся с генами, кодирующими химерный антиген, мРНК или с полинуклеотидами, кодирующими химерный антиген. Дополнительно изобретение включает в себя аналоги кодирующих химерный антиген генов, конкретно описанные здесь. Аналоги включают в себя, например, мутанты, сохраняющие способность вызывать иммунный ответ и предпочтительно обладающие гомологией по меньшей мере 80%, более предпочтительно - 90% и наиболее предпочтительно - 95% с любым из кодирующих химерный антиген полинуклеотидов, как конкретно приведено в SEQ ID № № 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46 и 48. Как правило, такие аналоги отличаются изменениями кодонов в количестве только от 1 до 15. Примеры включают в себя полипептиды с незначительными отличиями аминокислот от природной аминокислотной последовательности вирусного антигена или фрагмента антитела, в частности консервативные замены аминокислот. Консервативные замены представляют собой такие замены, которые происходят в пределах семейства аминокислот, родственных по их боковым цепям. Кодируемые генами аминокислоты, как правило, делят на четыре семейства: (1) кислые=аспартат, глутамат; (2) основные=лизин, аргинин, гистидин; (3) неполярные=аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан и (4) незаряженные полярные=глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин. Фенилаланин, триптофан и тирозин иногда группируют вместе как ароматические аминокислоты. Например, разумно ожидать, что отдельная замена лейцина на изолейцин или валин, аспартата на глутамат, треонина на серии или подобная консервативная замена аминокислоты структурно родственной аминокислотой не будет оказывать большое влияние на биологическую активность. Полипептидные молекулы, обладающие по существу идентичной аминокислотной последовательностью как любой из полипептидов, описанных в любой из SEQ ID № № 27, 2 9, 31, 33, 35, 37, 39, 41, 43, 45, 47 и 49, но несущие незначительные аминокислотные замены, не оказывающие значительного воздействия на способность химерных антигенов вызывать иммунный ответ, находятся в пределах определения химерного антигена, обладающего последовательностью, как указано в SEQ ID № № 27, 29, 31, 33, 35, 37, 39, 41, 43, 45, 47 и 49, соответственно. Производные включают в себя агрегационные конъюгаты с другими молекулами химерных антигенов и ковалентные конъюгаты с неродственными химическими группами. Ковалентные производные получают посредством связывания функциональностей с группами, находящимися на аминокислотных цепях химерных антигенов или на N- или С-концевых остатках посредством известных в данной области способов. Сокращения аминокислот предоставлены в табл. 1. Консервативные замены аминокислот в белке можно проводить без изменения конформации или функции белка. Белки могут содержать от 1 до 15 консервативных замен. Такие изменения включают в себя замену любого изолейцина (I), валина (V) и лейцина (L) на любую другую из данных гидрофобных аминокислот; аспарагиновой кислоты (D) на глутаминовую кислоту (Е) и наоборот; глутамина (Q) на аспарагин (N) и наоборот; и серина (S) на треонин (Т) и наоборот. Другие замены также можно рассматривать как консервативные, в зависимости от окружения конкретной аминокислоты и ее роли в трехмерной структуре белка. Например, глицин (G) и аланин (А) часто являются взаимозаменяемыми, как и аланин (А) и валин (V). Метионин (М), являющийся относительно гидрофобным, часто можно обменивать с лейцином или изолейцином, а иногда с валином. Лизин (K) и аргинин (R) часто являются взаимозаменяемыми в тех положениях, в которых важным свойством аминокислотного остатка является его заряд, а различия рК данных двух аминокислотных остатков не важны. В конкретных видах окружения можно рассматривать как "консервативные" еще некоторые изменения (см., например, Biochemistry 4 th Ed., Lubert Stryer ed. (W.H. Freeman and Co.), страницы 18-23; Henikoff and Henikoff, Proc Nat'l Acad Sci USA 89:10915-10919 (1992); Lei et al., J Biol Chem 270 (20): 11882-6 (1995)). Изобретение также относится к полинуклеотидам, которые гибридизуются предпочтительно в строгих условиях с кодирующим химерный антиген полинуклеотидом, как конкретно приведено в SEQ ID № № 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46 и 48. Специалист в данной области легко определит строгость реакций гибридизации, и, как правило, она представляет собой эмпирический расчет, зависящий от длины зонда, температуры отмывки и концентрации солей. Как правило, более длинные зонды требуют более высоких температур для правильного отжига, тогда как более короткие зонды требуют более низких температур. Как правило, гибридизация зависит от способности денатурированных последовательностей нуклеиновых кислот к повторному отжигу, когда в окружающей среде с температурой ниже температуры их плавления присутствуют комплементарные цепи. Чем выше степень желаемой гомологии между зондом и гибридизуемой последовательностью, тем выше относительная температура, которую можно применять. Как результат отсюда следует, что более высокие относительные температуры приводят к получению более строгих реакционных условий, тогда как более низкие температуры снижают их. Для дополнительных подробностей и объяснения строгости реакций гибридизации см., например, Ausubel et al., выше, на страницах 2.9.1-2.10.8 и 4.9.1-4.9.13. Как определено здесь, "жесткие условия" или "высокожесткие условия" в качестве неограничивающих примеров определяют как условия, в которых (1) для отмывки используют низкую ионную силу и высокую температуру, например 0,015М хлорид натрия/0,0015 М цитрат натрия/0,1% додецилсульфат натрия при 50 °С; (2) в течение гибридизации используют денатурирующее средство, такое как формамид, например 50% (об./об.) формамида с 0,1% бычьего сывороточного альбумина/0,1% фиколла/0,1% поливинилпирролидона/50 мМ натрий-фосфатного буфера при рН 6,5 с 7 50 мМ хлорида натрия, 75 мМ цитратом натрия при 42 °С или (3) используют 50% формамид, 5 ×SSC (0,75M NaCl, 0,075M цитрат натрия), 50 мМ фосфат натрия (рН 6,8), 0,1% пирофосфат натрия, 5х раствор Дэнхарда, разрушенную ультразвуком ДНК спермы лосося (50 мкг/мл), 0,1% SDS и 10% сульфат декстрана при 42 °С, с отмывкой при 42 °С в 0,2 ×SSC (хлорид натрия/цитрат натрия) и 50% формамид при 55 °С, с последующей высокострогой отмывкой, состоящей из 0,1 ×SSC, содержащего ЭДТА при 55 °С. "Умеренно жесткие условия" в качестве неограничивающих примеров описывают как условия в Sambrook et al., выше, и они включают в себя применение отмывочного раствора и условий гибридизации (например, температуры, ионной силы и % SDS) менее строгих, чем описанные выше условия. Пример умеренно строгих условий представляет собой инкубацию в течение ночи при 37 °С в растворе, содержащем: 20% формамида, 5 ×SSC (150 мМ NaCl, 15 мМ трехосновный цитрата натрия), 50 мМ фосфат натрия (рН 7,6), 5 × раствор Дэнхарда, 10% сульфат декстрана и 20 мг/мл денатурированной разрезанной ДНК спермы лосося, с последующей отмывкой фильтров в 1 ×SSC приблизительно при 37-50 °С. Специалисты в данной области знают, как отрегулировать температуру, ионную силу и т.д., как необходимо, чтобы удовлетворять таким факторам, как длина зонда и т.п. Варианты осуществления кодирующего химерный антиген полинуклеотида включают в себя: кодирующий химерный антиген полинуклеотид с последовательностью, выбранной из любой из SEQ ID № № 27, 29, 31, 33, 35, 37, 39, 41, 43, 45, 47 и 49, или нуклеотидную последовательность химерного антигена, выбранную из любой из SEQ ID № № 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46 и 48, где Т, необязательно, может представлять собой U. Например, варианты осуществления нуклеотидов химерного антигена включают в себя в качестве неограничивающих примеров: (a) полинуклеотид, содержащий последовательность, как описано в SEQ ID № №: нуклеотиды 1-1326 последовательности SEQ ID № 26, нуклеотиды 1-2004 последовательности SEQ ID № 28, нуклеотиды 1-1350 последовательности SEQ ID № 30, нуклеотиды 1-1293 последовательности SEQ ID № 32, нуклеотиды 1-1794 последовательности SEQ ID № 34, нуклеотиды 1-1581 последовательности SEQ ID № 36, нуклеотиды 1-1389 последовательности SEQ ID № 38, нуклеотиды 1-1347 последовательности SEQ ID № 40, нуклеотиды 1-2157 последовательности SEQ ID № 42, нуклеотиды 1-1395 последовательности SEQ ID № 44, нуклеотиды 1-1905 последовательности SEQ ID № 46 или нуклеотиды 1-2484 последовательности SEQ ID № 48, где Т также может представлять собой U, или состоящий из них; (b) полинуклеотид, последовательность которого по меньшей мере на 80% гомологична последовательности, описанной в SEQ ID № № 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46 или 48; (c) полинуклеотид, кодирующий химерный антиген, последовательность которого закодирована ДНК, содержащейся в одной из сконструированных плазмид: pFastBacHTa S1/S2 HBV-TBD, pFastBacHTa HBV core-TBD, pFastBacHTa HCV core(1-177)-TBD, pFastBacHTa HCV NS5A-TBD и pFastBacHTa HCV E2-TBD, хранящихся в International Depository Authority Канады (Bureau of Microbiology at Health Canada) под инвентарными № № 080504-03, 080504-04, 080504-05, 080504-02 и 080504-01, соответственно; (d) полинуклеотид, кодирующий химерный антиген, последовательность которого составляют аминокислоты 1-442 последовательности SEQ ID № 27, аминокислоты 1-668 последовательности SEQ ID № 29, аминокислоты 1-450 последовательности SEQ ID № 31, аминокислоты 1-431 последовательности SEQ ID № 33, аминокислоты 1-598 последовательности SEQ ID № 35, аминокислоты 1-527 последовательности SEQ ID № 37, аминокислоты 1-463 последовательности SEQ ID № 39, аминокислоты 1-449 последовательности SEQ ID № 41, аминокислоты 1-719 последовательности SEQ ID № 43, аминокислоты 1-465 последовательности SEQ ID № 45, аминокислоты 1-635 последовательности SEQ ID № 47 или аминокислоты 1-828 последовательности SEQ ID № 49; (e) полинуклеотид, кодирующий родственный с антигеном белок, который по меньшей мере на 90% идентичен полной аминокислотной последовательности, описанной аминокислотами 1-442 последовательности SEQ ID № 27, аминокислотами 1-668 последовательности SEQ ID № 29, аминокислотами 1-450 последовательности SEQ ID № 31, аминокислотами 1-431 последовательности SEQ ID № 33, аминокислотами 1-598 последовательности SEQ ID № 35, аминокислотами 1-527 последовательности SEQ ID № 37, аминокислотами 1-463 последовательности SEQ ID № 39, аминокислотами 1-449 последовательности SEQ ID № 41, аминокислотами 1-719 последовательности SEQ ID № 43, аминокислотами 1-465 последовательности SEQ ID № 45, аминокислотами 1-635 последовательности SEQ ID № 47 или аминокислотами 1-828 последовательности SEQ ID № 49; (f) полинуклеотид, полностью комплементарный полинуклеотиду любого из (a)-(d); и (g) полинуклеотид, селективно гибридизующийся с полинуклеотидом из (a)-(f) при строгих условиях. Изобретение также относится к рекомбинантным молекулам ДНК или РНК, содержащим полинуклеотид химерного антигена, его аналог или гомолог, включающим в себя в качестве неограничивающих примеров фаги, плазмиды, фагмиды, космиды, YAC (искусственные хромосомы дрожжей), ВАС (искусственные хромосомы бактерий), а также различные вирусные и не вирусные векторы, хорошо известные в данной области, и клетки, трансформированные или трансфицированные такими рекомбинантными молекулами ДНК или РНК. Способы получения таких молекул хорошо известны (см., например, Sambrook et al., 1989, выше). Изобретение дополнительно относится к системе вектор-хозяин, содержащей рекомбинантную молекулу ДНК, содержащую полинуклеотид химерного антигена, его аналог или гомолог, в пригодной прокариотической или эукариотической клетке-хозяине. Примеры пригодных эукариотических клеток-хозяев включают в себя дрожжевую клетку, растительную клетку или животную клетку, такую как клетка млекопитающего или клетка насекомого (например, зараженная бакуловирусом клетка, такая как Sf9, Sf21, Drosophila S2 или клетка High Five ™). Примеры пригодных клеток млекопитающих включают в себя различные клеточные линии рака простаты, такие как DU145 и TsuPr1, другие трансфицированные или трансдуцированные клеточные линии рака простаты, первичные клетки (PrEC), а также ряд клеток млекопитающих, регулярно применяемых для экспрессии рекомбинантных белков (например, клетки COS, СНО, 293, 293Т). Более подробно, полинуклеотид, содержащий кодирующую последовательность химерного антигена или его фрагмент, аналог или гомолог, можно использовать для получения кодируемого ею химерного антигена с применением любого ряда традиционно применяемых и широко известных в данной области систем хозяин-вектор. Для экспрессии в них химерных антигенов доступен широкий диапазон систем хозяин-вектор, см., например, Sambrook et al., 1989, выше; Ausubel, выше, на страницах 1.0.1-1.16.16, 9.01-9.17.3 и 13.4.1-13.6.5). Предпочтительные векторы для экспрессии в клетках насекомых включают в себя в качестве неограничивающих примеров pFastBac HTa (Invitrogen). С применением таких экспрессирующих векторов химерные антигены можно экспрессировать в нескольких клеточных линиях, включающих в себя, например, Sf9, Sf21, Drosophila S2 и High Five. Альтернативно, предпочтительные дрожжевые экспрессирующие системы включают в себя Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichia pastoris и Pichia august. Системы хозяин-вектор согласно изобретению пригодны для получения химерного антигена. Химерный антиген или его аналог или гомолог можно получать в клетках, трансфицированных конструкцией, кодирующей химерный антиген. Например, клетки Sf9 можно трансфицировать экспрессирующей плазмидой, кодирующей химерный антиген, или его аналог, или гомолог, химерный антиген экспрессируется в клетках Sf9, и химерный антиген выделяют с применением стандартных способов очистки. Также можно применять различные другие экспрессирующие системы, хорошо известные в данной области. Экспрессирующие конструкции, кодирующие лидерный пептид, объединенный в одну рамку считывания с кодирующей химерный антиген последовательностью, можно применять для получения секретируемой формы химерного антигена. Как обсуждают здесь, вырожденность генетического кода допускает вариацию в генетических последовательностях химерных антигенов. В частности, в данной области известно, что конкретные виды хозяев обладают конкретными кодонными предпочтениями, и, таким образом, можно адаптировать описываемую последовательность, как предпочтительно для желаемого хозяина. Например, предпочтительные кодонные последовательности аналога, как правило, включают в себя редкие кодоны (т.е. кодоны с частотой использования менее чем приблизительно 20% от известных последовательностей желательного хозяина), заменяемые кодонами с большей частотой. Кодонные предпочтения для конкретных видов рассчитывают, например, применяя таблицы использования кодонов, доступные в Интернете, такие как на URL всемирной паутины www.kazusa.or.jp/codon. Для увеличения экспрессии белка в клеточном хозяине известны дополнительные модификации последовательности. Они включают в себя элиминацию последовательностей, кодирующих ложные сигналы полиаденилирования, сигналы сплайсинга экзонов/интронов, подобные транспозонам повторы и/или другие такие хорошо охарактеризованные последовательности, вредные для экспрессии генов. Содержание GC в последовательности корректируют до средних уровней для данного клеточного хозяина, рассчитанного исходя из известных генов, экспрессирующихся в клетке-хозяине. Когда возможно, последовательность модифицируют так, чтобы избежать предсказанных вторичных структур мРНК в виде шпилек. Другие пригодные модификации включают в себя добавление на старте открытой рамки считывания консенсусной последовательности инициации трансляции, как описано у Kozak, Mol. Cell Biol., 9:5073-5080(1989). Специалисты в данной области понимают, что основное правило о том, что эукариотические рибосомы инициируют трансляцию исключительно на 5' проксимальном кодоне, AUG нарушается только в редких условиях (см., например, Kozak Proc Nat'l Acad Sci USA 92 (7):2662-2666 (1995) и Kozak Nucl Acids Res 15 (20):8125-8148 (1987)). G. Фармацевтические композиции согласно изобретению. Один из аспектов изобретения относится к фармацевтическим композициям, содержащим фармацевтически приемлемый наполнитель и химерный антиген, содержащий домен, вызывающий иммунный ответ, и домен связывания мишени, где домен связывания мишени содержит фрагмент антитела. В терапевтических применениях фармацевтические композиции можно вводить субъекту в количестве, достаточном для вызова эффективного В-клеточного ответа, ответа цитотоксических Т-лимфоцитов (CTL) и/или хелперных Т-лимфоцитов (Th) на антиген и для блокирования инфекции, или для лечения или, по меньшей мере, для частичной задержки или замедления развития симптомов и/или осложнений, или заболевания, или нарушения. Количества, эффективные для данного применения, будут зависеть, например, от конкретной вводимой композиции, способа введения, стадии и тяжести заболевания, подлежащего лечению, веса и общего состояния здоровья субъекта и решения лечащего врача. Дозировка для начальной терапевтической иммунизации (химерным антигеном), как правило, находится в диапазоне единиц дозирования, где нижнее значение составляет приблизительно 1, 5, 50, 500 или 1000 нг, а верхнее значение составляет приблизительно 10000, 20000, 30000 или 50000 мкг. Размер доз для человека, как правило, находится в диапазоне приблизительно от 500 нг до приблизительно 50000 мкг в расчете на индивидуума массой 70 кг. Поддерживающие дозы приблизительно от 1,0 нг до приблизительно 50000 мкг химерного антигена, в соответствии с поддерживающим режимом в течение от недель до месяцев, можно вводить в зависимости от ответа и состояния субъекта. Введение следует продолжать до тех пор, пока, по меньшей мере, клинические симптомы или лабораторные тесты не укажут, что состояние предотвращено, задержано, замедлено или устранено, и в течение некоторого периода после этого. Дозировки, способы введения и протоколы дозирования корректируют в соответствии с известными в данной области методологиями. Стандартная лекарственная форма химерного антигена, как правило, включает в себя фармацевтическую композицию, содержащую человеческую единицу дозирования приемлемого носителя, в одном из вариантов осуществления - водный носитель, и ее вводят в таком объеме/количестве, которое известно специалистам в данной области как подходящее для введения таких полипептидов людям (см., например, Remington: The Science and Practice of Pharmacy, 20 th Edition, A. Gennaro, Editor, Lippincott Williams & Wilkins, Baltimore, Md., 2000). Как понятно специалистам в данной области, на идеальную дозу в каждом конкретном случае могут влиять различные факторы. Такие факторы включают в себя, например, период полувыведения химерного антигена, аффинность связывания химерного антигена, иммуногенность композиции, желаемый стационарный уровень, способ введения, частота лечения и влияние других средств, применяемых в сочетании со способом лечения согласно изобретению, а также состояние здоровья конкретного субъекта. В определенных вариантах осуществления композиции по настоящему изобретению применяют при тяжелых болезненных состояниях, т.е. опасных для жизни или потенциально опасных для жизни ситуациях. В таких случаях, в результате относительно нетоксической природы химерного антигена в предпочтительных композициях, возможно и может быть очень желательно лечащими врачами вводить существенный избыток данных химерных антигенов относительно и указанных количеств в дозах. Концентрация химерного антигена согласно изобретению в фармацевтических препаратах может широко варьировать, т.е. от менее чем приблизительно 0,1%, как правило от или по меньшей мере приблизительно 2%, до количества от 20 до 50% или более по массе, и ее следует выбирать главным образом по объемам жидкости, вязкости и т.д. в соответствии с конкретным выбранным способом введения. Фармацевтические композиции можно доставлять посредством любого способа, известного в данной области, такого как парентеральный, интратекальный, внутрисосудистый, внутривенный, внутримышечный, трансдермальный, интрадермальный, подкожный, интраназальный, топический, пероральный, ректальный, вагинальный, пульмональный, или интраперитонеальный. Предпочтительно композицию доставляют парентеральными способами, такими как подкожное или интрадермальное введение. Фармацевтические композиции можно получать смешиванием желательных химерных антигенов с соответствующим носителем, подходящем для планируемого пути введения. При изготовлении фармацевтических композиций по данному изобретению химерный антиген, как правило, смешивают с наполнителем, разбавляют наполнителем или заключают внутри носителя, который может быть в форме капсулы, саше, бумаги или другого носителя. Когда фармацевтически приемлемый наполнитель служит в качестве разбавителя, он может быть твердым, полутвердым или жидким веществом, которое действует в качестве наполнителя, носителя или среды для терапевтического средства. Таким образом, композиции могут находиться в форме таблеток, пилюль, порошков, пастилок, саше, облаток, эликсиров, суспензий, эмульсий, растворов, сиропов, аэрозолей (как твердых, так и в жидкой среде), мазей, содержащих, например, до 10% по массе химерного антигена, мягких и твердых желатиновых капсул, суппозиториев, стерильных растворов для инъекций и стерильных расфасованных порошков. Некоторые примеры подходящих наполнителей включают в себя в качестве неограничивающих примеров декстрозу, сахарозу, сорбит, маннит, крахмалы, гуммиарабик, фосфат кальция, альгинаты, трагакант, желатин, силикат кальция, микрокристаллическую целлюлозу, поливинилпирролидон, целлюлозу, стерильную воду, патоку и метилцеллюлозу. Препараты могут дополнительно содержать: смазывающие средства, такие как тальк, стеарат магния и минеральное масло; увлажнители; эмульгаторы и суспендирующие средства; консерванты, такие как метил- и пропилгидроксибензоаты; подсластители и средства для придания вкуса. Композиции согласно изобретению можно формулировать так, чтобы обеспечить быстрое, длительное или замедленное высвобождение химерного антигена после введения субъекту посредством применения известных в данной области процедур. См., например, Remington, выше, на страницах 903-92 и страницах 1015-1050. Для получения твердой композиции, такой как таблетки, химерный антиген смешивают с фармацевтическим наполнителем с формированием твердой предпрепаратной композиции, содержащей гомогенную смесь химерного антигена по настоящему изобретению. Когда такие предпрепаратные композиции обозначают как гомогенные, это означает, что химерный антиген равномерно распределен во всем объеме композиции так, что композицию можно легко разделить на равноэффективные стандартные лекарственные формы, такие как таблетки, пилюли и капсулы. Таблетки или пилюли по настоящему изобретению можно покрывать или составлять иным образом для обеспечения формы дозирования, предоставляющей преимущество пролонгированного действия. Например, таблетка или пилюля может содержать компонент внутренней дозы и компонент внешней дозы, где последний находится в форме оболочки над формообразователем. Два компонента можно разделить растворимым в кишечнике слоем, служащим для препятствия разрушению в желудке и дающим возможность внутреннему компоненту пройти интактным в двенадцатиперстную кишку или высвобождаться с задержкой. Для таких растворимых в кишечнике слоев или покрытий можно применять множество веществ, такие вещества включают в себя ряд полимерных кислот и смесей полимерных кислот с такими веществами, как шеллак, цетиловый спирт и ацетат целлюлозы. Жидкие формы, в которые для перорального введения или инъекции можно помещать новые композиции по настоящему изобретению, включают в себя водные растворы, ароматизированные сиропы, жидкие или масляные суспензии и ароматизированные эмульсии с пищевыми маслами, такими как кукурузное масло, хлопковое масло, кунжутное масло, кокосовое масло или арахисовое масло, а также эликсиры и подобные фармацевтические носители. При получении композиции для парентерального введения определенное внимание необходимо уделять регуляции тоничности для уменьшения болезненной чувствительности. Восстанавливаемая композиция представляет собой стерильное твердое вещество, расфасованное в сухой форме. Восстанавливаемая композиция является предпочтительной, так как она является более стабильной при хранении в качестве сухого твердого вещества, чем в растворе, готового к непосредственному введению. Сухое твердое вещество, как правило, расфасовывают в стерильный контейнер с бутилкаучуковой пробкой для обеспечения того, что твердое вещество останется при оптимальном диапазоне влажности. Восстанавливаемое сухое твердое вещество формируют посредством способов сухого заполнения, распылительной сушки или лиофилизации. Описания данных способов можно найти, например, у Remington, выше, на страницах 681-685 и 802-803. Композиции для парентеральной инъекции, как правило, разводят, а компонент, присутствующий в наибольшей пропорции, представляет собой носитель. Носитель, как правило, не обладает терапевтической активностью и является нетоксичным, но представляет химерный антиген тканям организма в форме, подходящей для всасывания. Всасывание, как правило, происходит наиболее быстро и полностью, когда химерный антиген представлен в виде водного раствора. Однако модификация носителя водорастворимыми жидкостями или замещение водонерастворимыми жидкостями может влиять на скорость всасывания. Предпочтительно для данной композиции носителем с наибольшей значимостью является изотонический солевой раствор. При получении композиции, пригодной для инъекции, можно применять водные носители, водорастворимые носители и неводные носители. Для улучшения или сохранения качества композиции в композиции согласно изобретению для инъекции можно добавлять дополнительные вещества. Таким образом, добавляемое вещество может влиять на растворимость, обеспечивая субъекту комфорт, увеличивая химическую стабильность или защищая препарат от роста микроорганизмов. Таким образом, композиция может включать в себя подходящий растворитель, вещества, действующие в качестве антиоксидантов, и вещества, действующие в качестве консервантов для профилактики роста микроорганизмов. Данные вещества должны присутствовать в количестве, подходящем для их функционирования, но не должны неблагоприятно влиять на действие композиции. Примеры подходящих противомикробных средств включают в себя тимеросал, хлорид бензэтония, хлорид бензалкония, фенол, метил-п-гидроксибензоат и пропил-п-гидроксибензоат. Подходящие антиоксиданты можно найти в Remington, выше, на стр. 1015-1017. В определенных вариантах осуществления для введения химерных антигенов по настоящему изобретению применяют липосомы, нанокапсулы, микрочастицы, липидные частицы, везикулы и т.п. Конкретно, композиции по настоящему изобретению можно формулировать для доставки инкапсулированным в любое из липидной частицы, липосомы, везикулы, наносферы или наночастицы или т.п. Альтернативно, композиции по настоящему изобретению можно или ковалентно, или нековалентно связывать с поверхностью таких носителей. Композиции, вводимые посредством липосом, также могут служить: 1) для направления химерного антигена в конкретную ткань, такую как лимфоидная ткань; 2) для селективного направления к антигенпрезентирующим клеткам или 3) для увеличения времени полувыведения пептидной композиции. Липосомы включают в себя эмульсии, пены, мицеллы, нерастворимые монослои, жидкие кристаллы, фосфолипидные дисперсии, ламинарные слои и т.п. В данных препаратах химерный антиген для доставки введен как часть липосомы, отдельно или в сочетании с молекулой, связывающейся с рецептором, преобладающим на лимфоидных клетках, такой как моноклональные антитела, связывающие антиген CD45, или с другими терапевтическими или иммуногенными композициями. Таким образом, липосомы, или наполненные, или декорированные желательным химерным антигеном согласно изобретению, можно направлять к участку лимфоидных клеток, куда липосомы затем доставляют химерные антигены. Липосомы для применения согласно изобретению формируют из стандартных формирующих везикулы липидов, которые, как правило, включают в себя нейтральные или отрицательно заряженные фосфолипиды и стерол, такой как холестерин. Выбор липидов, как правило, проводят при учете, например, размера липосомы, устойчивости к кислотам и стабильности липосом в кровотоке. Для получения липосом доступны множество способов, как описано, например, у Szoka, et al., Ann. Rev. Biophys. Bioeng. 9:467-508(1980) и в патентах США № № 4235871, 4501728, 4837028 и 5019369. Суспензию липосом, содержащую химерный антиген, можно вводить внутривенно, локально, топически и т.д. в дозе, в числе прочего, варьирующей в зависимости от способа введения, вводимого химерного антигена и стадии заболевания, подвергаемого лечению. Композиции для ингаляции или инсуффляции включают в себя растворы и суспензии в фармацевтически приемлемых водных или органических растворителях или их смесях и порошки. Жидкие и твердые композиции могут содержать подходящие фармацевтически приемлемые наполнители, как описано здесь. Композиции можно вводить или пероральным, или респираторным путем для локального или системного воздействия. Композиции в фармацевтически приемлемых растворителях можно распылять с применением инертных газов. Распыляемые растворы можно ингалировать непосредственно из распыляющего устройства или распыляющее устройство можно присоединять к куполу лицевой маски или дыхательному устройству с пульсирующим избыточным давлением. Раствор, суспензию или порошок композиции можно вводить предпочтительно перорально или назально, из устройств, доставляющих препарат подходящим способом. В другом применяемом в способах по настоящему изобретению препарате используют устройства трансдермальной доставки ("пластыри"). Такие трансдермальные пластыри можно применять для обеспечения непрерывной или прерывающейся инфузии химерного антигена по настоящему изобретению в регулируемых количествах. Конструкция и применение трансдермальных пластырей для доставки фармацевтических средств хорошо известны в данной области. См., например, патент США № 5023252, включенный сюда в качестве ссылки. Такие пластыри можно сконструировать для непрерывной, пульсирующей доставки фармацевтических средств или доставки их по потребности. Кроме того, в дополнение к химерному антигену и фармацевтическим наполнителям может являться полезным включение в состав, как правило, одного противовирусного терапевтического или химиотерапевтического средства. Противовирусные терапевтические средства включают в себя в качестве неограничивающих примеров пептидомиметики (такие как ампренавир, индинавир, лопинавир, нелфинавир, ритонавир и саквинавир), полинуклеотиды (такие как амплиген и фомивирсен), пурин/пиримидиноны (такие как абакавир, ацикловир, адефовир, цидофовир, цитарабин, диданозин, дидезоксиаденозин, дипивоксил, эдоксудин, эмтрицитамин, энтековир, фамцикловир, ганцикловир, идоксуридин, инозин пранобекс, ламивудин, MADU, пенцикловир, соривудин, ставудин, тенофовир, трифлуридин, валацикловир, валганцикловир, видарабин, залцитабин и зидовудин), аналоги сиаловых кислот (такие как озельтамивир и занамивир), ацеманан, моноэтаноламин ацетиллейцина, амантадин, амидиномицин, атевиридин, каправирин, делавирдин, н-докозанол, эфавиренз, фоскарнет натрий, интерферон- α, интерферон- β, интерферон- γ, кетоксал, лизоцим, метизазон, мороксидин, невирапин, пентафузид, плеконарил, подофиллотоксин, рибавирин, римантидин, сталлимицин, статолон, термакамра и траомантадин. Другие подходящие противовирусные средства обсуждают в Remington, выше, в главе 87: Anti-Infectives, стр. 1507-1561, конкретно стр. 1555-1560. Предпочтительные противовирусные терапевтические средства для включения в фармацевтические композиции по настоящему изобретению включают в себя адефовир, дипивоксил, энтековир, ламивудин и рибавирин. В некоторых вариантах осуществления может быть желательным включить в фармацевтические композиции согласно изобретению по меньшей мере один компонент, примирующий В-лимфоциты или Т-лимфоциты. В качестве средств, способных к примированию CTL in vivo, идентифицированы липиды. Например, остатки пальмитиновой кислоты можно присоединить к ε- и α-аминогруппам лизинового остатка, а затем связать, например, посредством одного или нескольких связывающих остатков, таких как Gly, Gly-Gly-, Ser, Ser-Ser или т.п. с иммуногенным пептидом. Липидированный пептид затем можно вводить или непосредственно в мицелле или частице, внедренным в липосому или эмульгированным в адъюванте, например неполном адъюванте Фрейнда. В предпочтительном варианте осуществления, особенно эффективная иммуногенная композиция содержит пальмитиновую кислоту, присоединенную к ε- и α-аминогруппам Lys, который присоединен посредством связывающих звеньев, например Ser-Ser, к аминоконцу иммуногенного пептида. Для примирования специфичных к вирусам CTL можно применять другой пример липида, примирующего CTL-ответ, липопротеины Е. coli, такие как трипальмитоил-S-глицерилцистеинилсерилсерин (P 3 CSS) , когда они ковалентно присоединены к соответствующему пептиду (см., например, Deres, et al., Nature 342:561 (1989)). Химерные антигены согласно изобретению можно соединять, например, с P 3 CSS и вводить липопептид индивидууму для специфического примирования иммунного ответа на антиген-мишень. Хотя композиции по настоящему изобретению не требуют применения адъювантов, адъювант применять можно. Для увеличения иммунного ответа можно применять различные адъюванты, в зависимости от вида хозяина и включающие в качестве неограничивающих примеров адъювант Фрейнда (полный и неполный), минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, детергенты, плюрониловые полиолы, полианионы, пептиды, масляные эмульсии, гемоцианины морского блюдца, динитрофенол, иммуностимулирующие полинуклеотидные последовательности и потенциально применимые человеческие адъюванты, такие как BCG (бацилла Кальмета-Гирена) и Corynebacterium parvum. Дополнительные адъюванты также хорошо известны в данной области. Н. Готовое изделие. Другой аспект данного изобретения относится к готовому изделию, включающему в себя контейнер, хранящий композицию, содержащую химерный антиген, пригодную для инъекции или восстановления для инъекции в сочетании с напечатанными прикрепленными инструкциями, обеспечивающими рассмотрение того, как парентерально вводить композицию, например подкожно, внутримышечно, интрадермально, назально или внутрисосудистым способом. Композиция должна содержаться в любом подходящем контейнере, который не будет значимо взаимодействовать с композицией и должен быть помечен соответствующей меткой, указывающей, что она предназначена для парентерального введения. К контейнеру должны быть прикреплены инструкции, в соответствии со способом лечения, как описано выше. Контейнер, содержащий композицию по данному изобретению, может представлять собой контейнер с жидкой композицией, пригодной для инъекции, с подходящей иглой для инъекции и шприцом, так чтобы пациент, доктор, медицинская сестра или другой исполнитель мог ввести химерный антиген. Альтернативно, композиция может представлять собой сухую или концентрированную композицию, содержащую растворимую версию химерного антигена, для объединения или разбавления водным или неводным носителем для растворения или суспендирования композиции. Альтернативно, может содержать суспензию в жидкости или может содержать нерастворимый вариант соли для объединения с носителем, в котором нерастворимый вариант должен суспендироваться. Подходящие контейнеры обсуждают в Remington, выше, страницы 788-789, 805, 850-851 и 1005-1014. Набор согласно изобретению, как правило, содержит контейнер, описанный выше, и один или несколько других контейнеров, содержащих вещества, желательные с коммерческой или пользовательской позиции, включая буферы, разбавители, фильтры, иглы, шприцы и листовки-вкладыши с инструкциями по применению. На контейнере может присутствовать этикетка для указания того, что композицию применяют для конкретной терапии или не терапевтического приложения, и она также может указывать указания по применению in vivo или ex vivo, так как описано выше. Указания или другую информацию также можно включать во вкладыш, содержащийся в наборе. Следующие ниже неограничивающие примеры предоставляют дополнительную иллюстрацию изобретения. А. Пример 1. Конструирование экспрессирующего TBD вектора. Последовательности ДНК мышиных IgG1, кодирующие аминокислоты части области C H 1-шарнир-C H 2-C H 3, получали с мРНК, выделенной из гибридомы (2С12), продуцирующей mAb к поверхностному антигену HBV (sAg). Тотальную мРНК выделяли с применением реагента тризол (Trizol ®) (Gibco BRL, каталожный № 15596-026), а кДНК домена связывания мишени (TBD; фрагмент мышиных иммуноглобулинов) получали посредством ОТ-ПЦР с применением синтеза Superscript First-strand Synthesis (Invitrogen, каталожный № 11904-018). Праймеры для ПЦР содержали на 5'-конце линкерные последовательности, кодирующие линкерный пептид - SRPQGGGS - (SEQ ID № 1), уникальный участок Not I, а на 3'-конце уникальный рестрикционный участок Hind III. Полученная кДНК содержит (5' Not I)-линкерную последовательность-C H 1(VDKKI) (SEQ ID № 2). Шарнирную область C H 2-C H 3-(3' Hind III). После расщепления соответствующими ферментами фрагмент лигируют в экспрессирующую векторную плазмиду pFastBac HTa (Invitrogen), с применением тех же участков рестрикционных ферментов. Применяемый для амплификации ПЦР 5'-праймер представлял собой 5' TGTCATTCTGCGGCCGCAAGGCGGCGGATCCGTGGACAAGATTGTG CCCAGG (SEQ ID № 3) (смысловой), а 3'-праймер представлял собой 5' ACGAATCAAGCTTTGCA GCCCAGGAGAGTGGGAGAG (SEQ ID № 4) (антисмысловой), и они содержат участки Not I и Hind III, соответственно. Далее следует протокол для направленного клонирования. Полученные фрагменты расщепляли соответствующими ферментами, очищали на агарозном геле и клонировали в векторную плазмиду. Последовательность ДНК и корректность ORF подтверждали стандартными способами определения последовательностей. После клонирования ДНК кодирующей домен связывания мишени в донорную плазмиду pFastBac HTa, рекомбинантные белки экспрессировали с применением бакуловирусной экспрессирующей системы Bac-to-Bac ™ (Invitrogen). Клонированный ген переносили в бакуловирусный челночный вектор посредством сайт-специфической транспозиции в штамме Е. coli, DH10Bac. Клетки DH10Bac содержат челночный вектор, придающий устойчивость к канамицину и плазмиду-помощника, кодирующую транспозазу и придающую устойчивость к тетрациклину. Аликвоту 100 мкл компетентных клеток DH10Bac помещали на оттаивали на льду, добавляли основанные на pFastBac HTa плазмиды и смесь инкубировали на льду в течение 30 мин. Затем смесь подвергали тепловому шоку в течение 45 с при 42 °С, а затем охлаждали на льду в течение 2 мин. Затем смесь добавляли к 900 мкл среды LB и инкубировали в течение 4 ч при 37 °С. Трансформированные клетки последовательно разводили LB до 10 -1 и 10 и 100 мкл каждого из разведений помещали на планшеты с LB-агаром, дополненным 50 мкг/мл канамицина, 7 мкг/мл гентамицина, 10 мкг/мл тетрациклина, 100 мкг/мл X-gal и 40 мкг/мл IPTG, и инкубировали по меньшей мере в течение 36 ч при 37 °С. Устойчивость к гентамицину придавала pFastBac HTa, a X-gal и IPTG (изопропилтио- β-D-галактозид) применяли для различения белых колоний (рекомбинантные плазмиды) и голубых колоний (не рекомбинантные плазмиды). Отбирали белые колонии и инокулировали в 2 мл LB, дополненной 50 мкг/мл канамицина, 7 мкг/мл гентамицина, 10 мкг/мл тетрациклина и инкубировали в течение ночи при 37 °С при перемешивании. Для отбора небольшого количества ночной культуры применяли стерильную петлю и образец штрихами наносили на свежий планшет с LB-агаром, дополненным 50 мкг/мл канамицина, 7 мкг/мл гентамицина, 10 мкг/мл тетрациклина, 100 мкг/мл X-gal и 40 мкг/мл IPTG, и инкубировали в течение по меньшей мере 36 ч при 37 °С для подтверждения белого фенотипа. Рекомбинантные бакмиды выделяли посредством стандартных протоколов (Sambrook, выше), образец ДНК растворяли в 40 мкл ТЕ (10 мМ Tris-HCl pH 8, 1 мМ ЭДТА) и применяли для трансфекций. Для получения бакуловирусов, бакмиду трансфицировали в клетки насекомых Sf9. Клетки Sf9 (9 ×10 5 ) высевали в каждую лунку 6-луночной чашки для культивирования клеток (35 мм лунки) в 2 мл ESF 921 (Expression Systems) и позволяли прикрепляться в течение по меньшей мере 1 ч при 27 °С. Трансфекций проводили с применением реагента Cellfection ® Reagent (Invitrogen, каталожный № 10362-010) по протоколам, предоставленным поставщиком клеток Sf9. После трансфекций клетки инкубировали при 27 °С в течение 72 ч. Содержащую бакуловирус среду собирали и хранили при 4 °С в темноте. Эффективность трансфекций подтверждали проверкой на продукцию бакуловирусной ДНК. Выделенную бакуловирусную ДНК подвергали ПЦР для скринирования вставленного гена, кодирующего TBD. Применяемые праймеры представляли собой 5' TATTCCGGATTATTCATACCG (SEQ ID № 5) (смысловой) и 3' 5' CTCTACAAATGTGGTATGGC (SEQ ID № 6) (антисмысловой). Амплифицированные продукты пропускали через агарозный гель (0,8%). Экспрессию гетерологичного белка в клетках подтверждали электрофорезом в полиакриламидном геле с SDS (SDS-PAGE) и "вестерн-блотами" с применением моноклонального антитела с 6 ×His меткой (Clonetech) в качестве зонда. После подтверждения продукции бакуловируса и экспрессии белка увеличивали продукцию вируса для получения концентрированного маточного раствора бакуловируса, несущего ген, кодирующий домен связывания мишени. Стандартной практикой в данной области является амплификация бакуловируса по меньшей мере два раза и во всех описанных здесь протоколах придерживалась данной стандартной практике. После второго раунда амплификации подсчитывали концентрацию полученного бакуловируса с применением анализа бляшкообразования по протоколам, описанным производителем набора (Invitrogen). Также рассчитывали наиболее подходящую концентрацию вируса для заражения клеток High Five ™ и оптимальный период времени для продукции желаемого белка. Как правило, для экспрессии TBD применяли MOI 1 и период времени длительностью 48 ч. В. Пример 2. Конструирование экспрессирующих химерные антигены векторов. ДНК, кодирующую желательный вирусный антиген, получали с матрицы с применением методологии ПЦР с применением 5'-смыслового и 3'-антисмыслового праймеров, указанных в таблице 2. 5'-конец полученного амплифицированного фрагмента содержал уникальный участок рестрикции "5'enzy", a 3'-конец содержал уникальный участок рестрикции "3'enzy", каждый из которых применяли для лигирований. Амплифицированную ДНК расщепляли соответствующими 5' и 3' рестрикционными ферментами и лигировали с экспрессирующим вектором pFastBac HTa для получения экспрессирующей плазмиды только для вирусного антигена. Тот же фрагмент ДНК также лигировали с плазмидой pFastBac HTa-TBD, описанной в примере 1, после расщепления соответствующими ферментами, для получения экспрессирующей плазмиды для вирусного антигена, слитого с доменом связывания мишени. Полученные плазмиды применяли для получения рекомбинантного бакуловируса, как описано в примере 1, для последующего применения в экспрессии химерного антигена. Последовательности ДНК и аминокислотные последовательности химерных антигенов представлены в табл. 3. С. Пример 3. Экспрессия и очистка TBD, вирусных антигенов и химерных антигенов. Рекомбинантные бакмиды стандартизированной множественности заражения (MOI) применяли для заражения клеток насекомых High Five ™. Для суспензионных культур клетки высевали при плотности 3 ×10 5 клеток/мл и инкубировали при 27,5 °С при перемешивании при 138 об./мин до того момента, пока клеточная плотность не достигала 2-3 ×10 6 клеток/мл. К клеткам добавляли стандартизированные количества соответствующего рекомбинантного бакуловируса. Температура инкубации составляла 27,5 °С, а соответствующий период заражения стандартизировали для экспрессии индивидуального белка. Клетки собирали посредством центрифугирования при 2500 об./мин в течение 10 мин при 42 °С и применяли для очистки рекомбинантных белков. Неиспользуемые порции клеток подвергали быстрой заморозке в жидком азоте и хранили при -70 °С. Рекомбинантные белки очищали в денатурирующих условиях. Клетки лизировали в буфере, содержащем 6М гуанидиний-HCl в 100 мМ NaH 2 PO 4 , 10 мМ Tris, 300 мМ NaCl, 10 мМ имидазол, рН 8,0 (буфер для лизиса). Суспензию подвергали разрушению ультразвуком на льду с применением 5 импульсов по 1 мин на импульс при установках мощности в размере 60 Ватт и перемешивали при комнатной температуре в течение 1 ч. Лизат центрифугировали при 10000 ×g в течение 10 мин для удаления неразрушенных клеток и клеточного дебриса. Супернатант наносили на колонку с гранулами Ni-NTA агарозы (Qiagen) (1 ×5 см/100 мл клеточного лизата), предварительно уравновешенную лизирующим буфером. После нанесения колонку отмывали 20 объемами колонки, содержащими 6М гуанидиний-HCl в 100 мМ NaH 2 PO 4 , 10 мМ Tris, 300 мМ NaCl, 40 мМ имидазол, рН 8,0 (отмывочный буфер 1), с последующими отмывками 20 объемами колонки, содержащими 8М мочевину в 100 мМ NaH 2 PO 4 , 10 мМ Tris, 300 мМ NaCl, 40 мМ имидазол, рН 8,0 (отмывочный буфер 2). Связанный белок отмывали буфером, содержащим 8 М мочевину, 100 мМ NaH 2 PO 4 , 10 мМ Tris, 300 мМ NaCl, 250 мМ имидазол, рН 8 (буфер для элюции). Содержащие белок фракции объединяли и подвергали диализу при 4 °С против многократных замен буфера для диализа (10 мМ NaH 2 PO 4 , 300 мМ NaCl). Очищенные белки охарактеризовывали с применением стандартных биохимических способов, включающих в себя электрофорез в геле с SDS, изоэлектрофокусирование и анализ "вестерн-блот" с применением антител к различным доменам экспрессируемого белка. D. Пример 4. Нарушение толерантности к "собственному" белку с применением гибридного белка химерного антигена. Для оценки иммунного ответа на гибридные белки химерных антигенов мышей иммунизировали очищенными белками S1/S2 HBV, TBD или S1/S2-TBD и количественно определяли антитела, продуцирующиеся к отдельным белкам. Пролиферацию полученных от иммунизированных мышей селезеночных Т-клеток оценивали после стимуляции соответствующими белками. Для иммунизаций использовали мышей BALB/c в возрасте 15 недель. Мышам четыре раза с двухнедельными интервалами подкожно инъецировали S1/S2-TBD (4,15 мкг), S1/S2 (4,15 мкг) или TBD (4,15 мкг). Перед началом иммунизаций и через неделю после каждой из иммунизаций собирали образцы крови. Из свернувшихся образцов крови получали сыворотку и применяли для оценки уровней антител, продуцируемых животным-хозяином к соответствующим инъецированным антигенам. 1. ELISA для детекции антител к S1/S2 HBV, TBD или S1/S2-TBD. 96-луночный планшет покрывали антигенами S1/S2 HBV, TBD или S1/S2-TBD при концентрации 1,0 мкг/мл в течение ночи при 4 °С. Планшет отмывали PBS, содержащим 2% БСА. К каждой из лунок при различных разведениях (1:10-1:500) добавляли сыворотку от соответствующего животного и инкубировали при 37 °С в течение 1 часа. Планшет отмывали PBS, содержащим 0,05% Tween 20 (отмывочный буфет). К лункам добавляли разведение (1:5000) козьих IgG с Fab с пероксидазой хрена (HRP) к мышиным антителам и инкубировали при 37 °С в течение 1 ч. Планшет отмывали отмывочным буфером и получали окраску с применением 2-2'азиноди-(3-этилбензилтиазолин-6-сульфоната) (KPL, Guildford, UK). Оптическую плотность полученной в образцах окраски измеряли с применением планшетного сканера ELISA (Molecular Devices, USA) при длине волны 405 нм. Отрицательным контролем для эксперимента служила преимунная сыворотка от того же животного, значение которой вычитали из всех экспериментальных значений. Результаты для мышей, иммунизированных S1/S2-TBD HBV, представлены в табл. 4. Химерный антиген вызывает сильный ответ антителами против химерного антигена (S1/S2-TBD). Здесь ответ антителами является поливалентной (или полиэпитопной) природы. Представленные в таблице 4 результаты показывают, что антитела, продуцируемые иммунизированными S1/S2 HBV-TBD мышами, связываются с химерным антигеном и с белком-мишенью S1/S2, нанесенным на планшет. Таким образом, антитела продуцируются к компоненту химерного антигена S1/S2. Аналогично, антитела, продуцируемые иммунизированными S1/S2 HBV-TBD мышами, связываются с белком домена связывания мишени (таблица 4). Химерный антиген, содержащий белок мышиного происхождения, может вызывать гуморальный иммунный ответ у мыши, доказывая то, что химерный антиген может преобразовывать "собственный" антиген в "чужеродный". Таким образом, возможно нарушение толерантности к белку, в ином случае воспринимаемому как "собственный" белок. 2. Анализ Т-клеточной пролиферации. Через неделю после четвертой иммунизации животных умерщвляли, удаляли селезенку и получали суспензию отдельных клеток. Клетки в трех экземплярах высевали при клеточной плотности 4 ×10 5 клеток/лунку в 96-луночный планшет. Их нагружали соответствующими антигенами, S1/S2 HBV, TBD или S1/S2-TBD, при концентрациях 0,1 мкг/мл, 1,0 мкг/мл и 10 мкг/мл. К клеткам с отрицательным контролем добавляли только среду, а положительным контролем для Т-клеточной пролиферации служил фитогемагглютинин (РНА) при 1,0-5,0 мкг/мл. Клеточные культуры инкубировали в течение 4 суток при 37 °С в атмосфере 7% СО 2 . Каждую из лунок с клетками импульсно метили 1,0 мКи 3 [Н]-тимидина и инкубировали дополнительные 18 ч. Клетки собирали с применением клеточного харвестра ТОМТЕС МАСН 3 (Hamden, CT, USA) и связавшуюся с фильтром из стекловолокна (Wallac Oy, Turku, Finland) радиоактивность количественно определяли с применением жидкостного сцинтиллятора Wallac Trilux 1450 Microbeta и счетчика люминесценции (Wallac, USA) . Результаты показаны в табл. 5. Т-клеточную пролиферацию наблюдали при стимуляции S1/S2 HBV-TBD, S1/S2 или TBD. Иммунизация химерным антигеном индуцировала поливалентный Т-клеточный ответ, т.е., ответ к различным частям одного и того же белка. Химерный антиген, содержащий белок мышиного происхождения, может вызывать клеточный иммунный ответ у мыши, доказывая то, что химерный антиген может преобразовывать "собственный" антиген в "чужеродный". Таким образом, возможно нарушение толерантности к белку, в ином случае воспринимаемому как "собственный" белок. E. Пример 5. Анализы антигенной презентации. Способность S1/S2 HBV-TBD вызывать иммунный ответ измеряли с применением анализа антигенной презентации ex vivo. Получение эффективного Т-клеточного ответа после множественной стимуляции наивных Т-клеток нагруженными антигенами антигенпрезентирующими клетками (АРС), такими как дендритные клетки (DC), оценивали количественным определением увеличения количества специфичных к антигену Т-клеток, а также способностью Т-клеток продуцировать Th1-цитокин IFN- γ. 1. Отбор моноцитов посредством адгезии. Мононуклеарные клетки периферической крови (РВМС) оттаивали посредством добавления AIM-V (в отношении 9 мл AIM-V добавляемых к 1 мл замороженных клеток). Затем клетки центрифугировали при 200 ×g в течение 5 мин, супернатант удаляли, а клетки ресуспендировали в AIM-V/1% соответствующей сыворотке и добавляли в 100 мм культуральную чашку или в культуральный флакон Т-25. РВМС инкубировали в течение 1 ч при 37 °С во влажном инкубаторе в 7% СО 2 . Для удаления неприкрепившихся клеток культуру несколько раз растирали, супернатант удаляли, а клетки однократно отмывали средой AIM-V. Моноциты собирали клеточным скребком и центрифугировали при 300 ×g в течение 5 мин. Клеточный осадок ресуспендировали в AIM-V/2,5% соответствующей сыворотке при 2 ×10 6 клеток/мл и высевали в 24-луночную чашку. Для запуска дифференцировки моноцитов в незрелые DC добавляли цитокины IL-4 и GM-CSF (1000 МЕ/мл каждого). 2. Быстрый или медленный анализ антигенной презентации. Для быстрого анализа антигенной презентации (АРА) антиген добавляли к незрелым DC в период от 4 до 24 ч после выделения. После дополнительных 24 ч нагруженных антигеном незрелых моноцитов стимулировали к созреванию посредством культивирования с PGE 2 (1 мкМ) , IL-1 β (10 нг/мл) и TNF α (10 нг/мл) в течение 24 ч. Затем зрелые DC совместно культивировали (первая стимуляция) с аутологичными Т-клетками. Т-клетки получали из тех же РВМС, что и DC посредством отрицательной селекции с применением набора с магнитами для выделения Т-клеток (Dynal) по инструкциям производителя. Затем Т-клетки через 7 суток повторно стимулировали нагруженными антигенами зрелыми DC в присутствии IL-2 (20 МЕ/мл), IL-7 (10 нг/мл) и IL-15 (5 нг/мл). Через 7 суток инкубации Т-клетки третий раз стимулировали нагруженными антигенами зрелыми DC. Третья стимуляции длилась в течение 6 ч, после чего Т-клетки собирали и подвергали иммуноокрашиванию на экспрессию CD3, CD8 и IFN- γ и анализировали проточной цитометрией. Для медленного АРА, моноцитам перед добавлением антигена позволяли дифференцироваться до незрелых DC в присутствии GM-CSF и IL-4 в течение от 5 до 6 суток. Через два часа после добавления антигенов незрелые DC превращали в зрелые с применением TNFa (10 нг/мл) и IFN- α (50 МЕ/мл). Через семь суток после выделения зрелые DC совместно культивировали (первая стимуляция) с аутологичными Т-клетками (описано выше). Затем Т-клетки через 7 суток повторно стимулировали нагруженными антигенами зрелыми DC в присутствии IL-2, IL-7 и IL-15. Через 7 суток инкубации клетки третий раз стимулировали нагруженными антигенами зрелыми DC. Через 18 ч инкубации Т-клетки собирали и подвергали иммуноокрашиванию на экспрессию CD3, CD8 и IFN- γ и анализировали проточной цитометрией. 3. Анализ антигенной презентации РВМС. В данном анализе исходная культура состоит из тотальных РВМС (например, лимфоцитов и моноцитов), которые инкубируют с антигеном и IL-2 в предположении, что система будет иметь сходство с иммунным ответом in vivo, так как для участия представлены все типы клеток (Maini, M. K. et. al J. Exp. Med. 191:1269-1280, 2000). РВМС оттаивали, отмывали и сразу же инкубировали с антигеном. Через 4 суток культивирования для обеспечения захвата и презентации антигенов добавляли IL-2 (20 МЕ/мл) и оставляли в течение дополнительных 8 суток (т.е. сутки 12 эксперимента). За сутки до второй стимуляции (т.е. сутки 10 эксперимента), DC выделяли посредством адгезии, как описано выше, и непосредственно инкубировали с GM-CSF, IL-4 и антигеном в течение 24 ч. Как и при быстром АРА, незрелым клеткам позволяли дифференцироваться в течение 24 ч после добавления PGE2, IL-1 β и TNF α. Затем к культуре РВМС добавляли нагруженные зрелые DC (вторая стимуляция, сутки 12 эксперимента) в присутствии IL-2, IL-7 и IL-15. Третью стимуляцию проводили на 21 сутки эксперимента нагруженными антигенами зрелыми DC, полученными за 2 суток до этого, как описано. После 6 ч инкубации Т-клетки собирали и подвергали иммуноокрашиванию на экспрессию CD3, CD8 и IFN- γ и анализировали проточной цитометрией. Для всех обсуждаемых выше анализов антигенной презентации порцию Т-клеток в конце анализа инкубировали в течение дополнительных 3-5 суток и проверяли на специфические Т-клетки посредством анализа тетрамеров (см. ниже). 4. S1/S2 HBV вызывает Т-клеточный ответ к S1 и S2 HBV. Пептиды. Для получения Т-клеток, которые затем оценивали на их антигенную специфичность, применяли АРА РВМС. Таким образом, РВМС от здоровых индивидуумов HLA-A2 культивировали в AIM-V, содержащей 2,5% соответствующих сывороток, в 96-луночных планшетах при 5 ×10 5 клеток/мл. Добавляли антиген (т.е. 10 мкг/мл S1/S2-TBD) и клетки культивировали в течение 4 суток при 37 °С. Затем добавляли IL-2 при 20 МЕ/мл и клетки культивировали в течение дополнительных 8 суток с заменой сред (AIM-V/2,5% соответствующей сыворотки и 20 МЕ/мл IL-2) каждые 2-3 суток. Большинство клеток, оставшихся к концу 12 суточного культивирования, представляли собой Т-клетки и данные Т-клетки повторно стимулировали аутоолгичными нагруженными антигеном зрелыми DC в присутствии IL-2 (20 МЕ/мл), IL-7 (10 нг/мл) и IL-15 (5 нг/мл). Нагруженные антигеном зрелые DC для второй и третьей стимуляции Т-клеток в АРА получали в течение периода 48 ч с применением описанной ниже процедуры. Моноциты выделяли из тотальных РВМС посредством адгезии на пластиковые чашки для тканевых культур. Клетки, приблизительно 85% которых представляли собой моноциты, как определяли посредством анализа FACS (CD11c + , CD14 + , CD19 - и CD3 - ), культивировали в 96-луночном планшете при 1 ×10 5 клеток/лунку содержащей 100 мкл AIM-V/2,5% соответствующих сывороток с цитокинами IL-4 и GM-CSF при 1000 МЕ/мл, и через 4 ч добавляли антиген, такой как S1/S2-TBD. После 20 ч инкубации полученные незрелые DC дифференцировались в зрелые посредством культивирования в течение дополнительных 24 ч в присутствии PGE 2 (1 ×10 -6 М), IL-1 β (10 нг/мл) и TNF α (10 нг/мл). Т-клетки культивировали в течение 7 суток после второй стимуляции с заменами сред (AIM-V с 2,5% соответствующей сыворотки и 20 МЕ/мл IL-2) каждые 1-2 суток. Затем Т-клетки (сутки 19 культивирования) стимулировали третий раз нагруженными антигеном зрелыми DC (полученными в течение 2 суточной процедуры, как описано выше) в присутствии IL-2, IL-7 и IL-15 (как выше), и или оценивали на продукцию IFN- γ после 6 часов культивирования или культивировали в течение 5 суток (с заменой сред каждые 1-2 суток на AIM-V/2,5% соответствующей сыворотки и 20 МЕ/мл IL-2), а затем оценивали на специфичность Т-клеток к антигену preS HBV с применением тетрамеров preS HBV (сутки 24 культивирования). Анализ тетрамеров проводили с применением самостоятельно синтезированных по протоколам производителей тетрамеров МНС класса I iTag (Beckman Coulter). Таким образом клетки собирали, отмывали и переносили в 96-луночный планшет с v-образным дном при ~2 ×10 5 клеток/лунку в 20 мл. Клетки метили при 20 °С в течение 30 мин mAb, специфичными к CD3 (антитела к CD3 с FITC) и CD8 (антитела к CD8 с Су-Chrome), совместно с 2 мкл конъюгированного с РЕ тетрамера HLA-A*0201 preS1 (GMLTPVSTI, SEQ ID № 50) или тетрамера preS2 (NIASHISSI, SEQ ID № 51) . Затем клетки отмывали, фиксировали 2% параформальдегидом в PBS и переносили в 5 мл пробирки для FACS. Клетки выявляли на FACSCalibur (BD Biosciences) с 80000-100000 событиями на образец. Анализ проводили с применением программного обеспечения CellQuest (BD Biosciences) с воротами, определенными на живой (основываясь на профиле FSC/SSC) популяции CD3 + и доле клеток CD8 + , меченых тетрамером. Когда РВМС культивировали с S1/S2-TBD HBV при 10 мкг/мл и дважды повторно стимулировали нагруженными S1/S2 HBV-TBD зрелыми DC, заметная доля клеток положительно метилась тетрамером S1 (фиг. 2) и тетрамером S2 (фиг. 3). Это отличается от Т-клеток, культивированных со зрелыми DC, не нагруженными антигеном, где количество положительных на тетрамер клеток являлось незначимым. Таким образом, нагруженные S1/S2-TBD зрелые DC были способны индуцировать образование значительного количества Т-клеток со специфическими детерминантами к антигенам S1 HBV и S2 HBV. F. Пример 6. Нарушение толерантности к антигенам DHBV и DHBV с применением гибридного белка химерного антигена. DHBV служил эффективной животной моделью в разработке противовирусной терапии для HBV. Для изучения механизма репликации вируса и для скрининга противовирусных соединений применяли пекинских уток, врожденно зараженных DHBV. В настоящем изобретении применяли два типа моделей на утках. Первая представляет собой врожденно зараженных DHBV уток. Она похожа на вертикальный перенос HBV у человека. Вторая модель представляет собой персистирующую модель инфекции, когда только что вылупившихся утят заражают DHBV, и они переносят инфекцию. Данная вторая модель похожа на горизонтальный перенос инфекции HBV у человека. 1. Врожденно зараженные DHBV утки. Врожденно зараженных DHBV уток в возрасте четырех недель разделяли на две группы. Для получения точки отсчета уровней антитела до иммунизации собирали образец крови (1,0 мл) и образцы крови собирали каждую неделю до вакцинаций. Экспериментальной группе, получавшей гибридный белок химерного антигена Core-TBD DHBV в количестве 19,95 мг/дозу, проводили подкожную инъекцию каждую неделю в одни и те же сутки до недели 5. В течение недели 6 дозу удваивали и инъецировали однократно каждые четыре недели до прерывания вакцинации на неделе 26. Группа плацебо получала равный объем буфера (20 мМ фосфат натрия рН 8,0, 300 мМ NaCl). 96-луночный планшет покрывали антигенами Core DHBV, TBD или Core DHBV-TBD, при концентрации 1,0 мкг/мл в течение ночи при 42 °С. Планшет отмывали фосфатно-солевым буфером (PBS), содержащим 2% BSA. К каждой из лунок при различных разведениях (1:10-1:500) добавляли разведенную сыворотку от соответствующего животного и инкубировали при 37 °С в течение 1 ч. Планшет отмывали PBS, содержащим 0,05% Tween 20 (отмывочный буфер). К лункам добавляли разведение (1:5000) козьих IgG-HRP к утиным антителам и инкубировали при 37 °С в течение 1 ч. Планшет отмывали отмывочным буфером и получали окраску с применением 2-2'азиноди-(3-этилбензилтиазолин-6-сульфоната) (KPL, Guildford, UK). Оптическую плотность полученной в образцах окраски измеряли с применением планшетного сканера ELISA (Molecular Devices, USA). Титры антител подсчитывали относительно преимунной сыворотки от того же животного. Уровни антител к коровому белку в сыворотке от врожденно инфицированных DHBV уток в контрольной и экспериментальной группа уток на неделях 0, 3 и 6 показаны в табл. 6. Хотя у уток была хроническая инфекция DHBV, уровни антител были низкими, вследствие хронической природы инфекции и так как иммунная система не распознавала антиген как чужеродную молекулу. При иммунизации химерным антигеном Core DHBV-TBD иммунная система хозяина распознавала вирусный антиген и запускала гуморальный ответ на сердцевинный антиген, который уже присутствовал у хозяина, таким образом, нарушая толерантность хозяина к вирусному антигену. Подобным образом иммунная система утки распознает компонент TBD химерного антигена в качестве чужеродного антигена и также запускает иммунный ответ к данной части гибридного белка. Планшеты покрывали TBD и сыворотку от отдельных уток оценивали на уровни антител посредством ELISA. Результат данного исследования представлен в табл. 6. 2. Утки, инфицированные DHBV после вылупления. Нормальных утят инфицировали содержащей DHBV сывороткой утки через сутки после вылупления утят. Это является стандартной практикой в области исследования DHBV. Наличие постоянной виремии подтверждали с применением общеизвестных способов на неделе четыре перед началом иммунизации. Инфицированных DHBV уток разделили на две групп. Для получения точки отсчета уровней антитела до иммунизации собирали образец крови (1,0 мл) и образцы крови собирали каждую неделю до вакцинаций. Экспериментальной группе, получавшей гибридный белок химерного антигена Core-TBD DHBV в количестве 19,95 мкг/дозу проводили подкожную инъекцию каждую неделю в одни и те же сутки до недели 5. В течение недели 6 дозу удваивали и инъецировали однократно каждые четыре недели до прерывания вакцинации на неделе 30. Образцы крови собирали от группы плацебо, получавшей равный объем буфера (20 мМ фосфат натрия рН 8,0, 300 мМ NaCl). Представлены уровни антител в сыворотках, собранных у уток на неделях 0, 3 и 6. Уровни антител к коровому белку в сыворотке от уток, инфицированных после вылупления DHBV, в контрольной и экспериментальной группа уток показаны в табл. 7. Хотя у уток была хроническая инфекция DHBV, уровни антител были низкими, так как иммунная система не распознавала антиген как чужеродную молекулу. При иммунизации химерным антигеном Core DHBV-TBD, иммунная система хозяина распознавала вирусный антиген и запускала гуморальный ответ на сердцевинный антиген, который уже присутствовал у хозяина, таким образом, нарушая толерантность хозяина к вирусному антигену. Уровни антител к TBD также возрастали (табл. 7). Следовательно, появляется поливалентный (или полиэпитопный) иммунный ответ на различные части одного и того же химерного антигена. G. Пример 7. Перекрестносвязанные химическим способом sAg HBV-Fc (мышиный). Растворы 100 мкг sAg (US Biologicals; каталожный № Н 1910-27) и 100 мкг Fc-фрагмента мышиных поликлональных IgG (Harlan Sera-Lab Ltd., каталожный № PP-19-01) подвергали диализу против 100 мМ HEPES рН 8,7 в течение ночи при 4 °С. Растворы белков смешивали вместе, сразу же добавляли диметилсуберимидат (DMS; Pierce Cat № 20700) до конечной концентрации 10 мМ и смесь инкубировали при комнатной температуре в течение 1 ч. Реакцию останавливали добавлением 0,1М Tris HC1 рН 7,8. Реакционную смесь наносили на колонку Sephadex G 75 column (0,7 ×12 см), а фракции элюировали с применением фосфатно-солевого буфера. Собирали 0,5 мл фракции и фракции, содержащие sAg/Fc в молярном соотношении 1:1, как оценивали посредством ELISA с применением соответствующих антител, объединяли. Объединенные фракции применяли для анализов антигенной презентации (Berlyn, et al., Clin. Immunol. 101:276-283, (2001)). Незрелые дендритные клетки культивировали в течение четырех суток с GM-CSF/IL4, инкубировали с конъюгатом sAg-Fc и давали созреть в присутствии TNF α и интерферона- α. К зрелым дендритным клеткам добавляли аутологичные CD3 + Т-клетки. После трех раундов контакта со зрелыми дендритными клетками количественно подсчитывали стимуляцию Т-клеток измерением продукции внутриклеточного интерферона- γ с применением проточной цитометрии. Уровни внутриклеточного интерферона- γ, продуцируемого в Т-клетках в присутствии конъюгата, были существенно выше, чем в присутствии только sAg или только Fc-фрагмента (табл. 8). Н. Пример 8. Анализы антигенной презентации. Анализы антигенной презентации проводили с применением дендритных клеток, полученных из человеческих РВМС по разработанным протоколам (Berlyn, et al., выше (2001)). Краткое изложение протокола для анализа стимуляции Т-клеток представлено в схематической форме. 1. Получение зрелых, нагруженных дендритных клеток. Моноциты получали из образцов лейкафереза от здоровых доноров и уменьшали количество лимфоцитов и гранулоцитов посредством инкубации с антителами к CD2, CD7, CD16, CD19 и CD56. За этим следовала инкубация с конъюгированными с магнитными бусами IgG к антителам мыши и разделение на магните (Dynal). Отрицательно отобранные клетки являлись более чем на 95% чистыми моноцитами, как охарактеризовано посредством проточной цитометрии с применением широкой панели маркеров CD (CD14 + , CD11c + , CD19 - , CD3 - , CD4 - , CD64 + , CD32 + , CD86 + , CD16 - ) . Далее моноциты инкубировали с IL-4 и GM-CSF (R &D Systems) в течение 4 суток в AIM-V с 2,5% соответствующей человеческой сыворотки для получения незрелых дендритных клеток. Снова Аликвоту клеток окрашивали широкой панелью маркеров CD для гарантии чистоты и идентичности клеток. Затем клетки нагружали S1/S2 HBV-TBD (5,0 мкг/мл) , S1/S2 HBV (2,5 мкг/мл) или TBD (2,5 мкг/мл) в течение 2-4 ч при 37 °С и позволяли созревать в присутствии интерферона- α и TNF α в течение 3 суток. Дендритные клетки снова контролировали с применением проточной цитометрии на матрице маркеров CD для гарантии того, что клетки претерпели правильное созревание. Полученные в результате зрелые, нагруженные дендритные клетки применяли для анализа стимуляции Т-клеток. 2. Анализ стимуляции Т-клеток: анализ цитокинов. Т-клетки получали из того же образца РМВС, что и дендритные клетки посредством негативной селекции с применением набора выделения Т-клеток с применением магнита (Dynal) по инструкциям производителя. Зрелые, нагруженные дендритные клетки (DC-1) тщательно отмывали и добавляли к Т-клеткам (сутки 0). Т-клетки и дендритные клетки инкубировали в течение 7 суток. На сутки 7 Т-клетки повторно стимулировали зрелыми, нагруженными дендритными клетками (DC-2). Через 2 ч отбирали аликвоту клеток. Аликвоту клеток инкубировали с брефелдином A (GoIgiPlug ™, R &D Systems) в течение 18 ч, а затем анализировали на окрашивание внутриклеточного цитокина, как описано ниже. Оставшиеся клетки инкубировали в течение дополнительных 7 суток. На сутки 14 оставшиеся клетки стимулировали третьей порцией зрелых, нагруженных дендритных клеток (DC-3). Через 2 ч отбирали аликвоту клеток. Аликвоту клеток инкубировали с брефелдином A (GoIgiPlug ™, R &D Systems) в течение 18 ч, а затем анализировали на окрашивание внутриклеточного цитокина, как описано ниже. Для внутриклеточного окрашивания цитокина клетки окрашивали антителом к CD3 с FITC и антителом к CD8 с Cy-Chrome в течение 30 мин, отмывали, фиксировали, нарушали проницаемость мембраны, а затем окрашивали с применением антител к интерферону- γ с РЕ в течение 30 мин на льду. Клетки отмывали и анализировали проточной цитометрией (FACScan, BD Biosciences). Результаты показаны в табл. 9. После удаления аликвоты на сутки 14 оставшиеся Т-клетки инкубировали в течение дополнительных трех суток, а затем супернатант применяли для измерения уровня секретируемого интерферона- γ посредством ELISA (набор Opt E1A ELISA, BD Biosciences). Стимуляцию Т-клеток оценивали посредством измерения внутриклеточных и секретируемых уровней интерферона- γ. Результаты представлены в табл. 10. Химерный антиген S1/S2-TBD индуцировал продукцию повышенных уровней интерферона- γ по сравнению или с доменом, вызывающим иммунный ответ или с доменом TBD молекулы при тестировании по отдельности при равных концентрациях. Необходимо отметить, что 5 мкг дозы S1/S2-TBD приблизительно содержали 2,5 мкг каждого из компонентов. На Т-клеточный ответ тестировали различные концентрации S1/S2-TBD. Эффект S1/S2-TBD был большим, чем обработка токсином столбняка при тех же концентрациях. При концентрациях меньших, чем 5 мкг/мл, химерный антиген вызывал зависимое от концентрации увеличение продукции и секреции интерферона- γ. Как показано в табл. 11, продукция и секреция интерферона- γ CD3 + T-клетками после презентации антигена S1/S2-TBD дендритными клетками увеличивалась зависимым от концентрации образом. Положительный ответ при низких концентрациях мог бы являться выгодным в отношении дозы, необходимой для вакцинации, и стоимости производства вакцины. I. Пример 9. Связывание и захват химерных антигенов. 1. Получение зрелых, нагруженных дендритных клеток. Мононуклеарные клетки периферической крови (РВМС) получали при обработке фиколл/гистопаком (Sigma) препаратов лейкофереза клеток (Berlyn, et al., выше (2001)). Моноциты отделяли от популяции РВМС отрицательной селекцией с применением набора для выделения моноцитов (Dynal) по инструкциям производителя. Моноциты были более чем 95% чистоты, как оценивали посредством анализа антителами и проточной цитометрией (CD14 + , CD11c + , CD19 - , CD3 - , CD4 - , CD64 + , CD32 + , CD86 + , CD16 - ) . Моноциты дважды отмывали средой AIM-V (Invitrogen), содержащей L-глутамин, стрептомицина сульфат (50 мкг/мл) и гентамицина сульфат (10 мкг/мл) с 1% донорскими соответствующими сыворотками (выделенными, как описано у Berlyn, et al., выше (2001)). Далее моноциты культивировали в среде AIM-V, содержащей 2,5% донорские соответствующие сыворотки и цитокины GM-CSF и IL-4 для дифференцировки клеток в направлении линии дендритных клеток (DC). Клетки инкубировали в 12-луночных планшетах для тканевых культур при 37 °С в атмосфере 7% СО 2 . Полученные из моноцитов дендритные клетки собирали в течение суток от 1 до 4. Потом клетки однократно отмывали средой AIM-V с 0,1% BSA (Sigma) и дважды фосфатно-солевым буфером Дульбеко (Invitrogen) с 0,1% (масс./об.) BSA (PBSB). Полученные из моноцитов дендритные клетки применяли в анализах мечения и связывания при 4 °С или в анализах связывания/захвата при 37 °С. 2. Связывание химерных антигенов с созревающими дендритными клетками. Сравнивали избыточность связывания S1/S2-TBD с созревающими дендритными клетками по отношению к мышиным IgG1 и IgG2a. Дендритные клетки выделяли на различные сутки культуры ex vivo (от суток 0 до суток 4) и обрабатывали S1/S2-TBD (10 мкг/мл) или мышиными IgG1 (2C12, родительское mAb, из которого получен TBD) или IgG2a (G155-178, 90 мкг/мл) в течение 1 часа при 4 °С. Клетки обрабатывали F(ab') 2 козьими антителами к антителам мыши Alexa-488 (10 мкг/мл) в PBSB в течение 20 минут. Клетки дважды отмывали PBSB и ресуспендировали в PBSB с 2% параформальдегидом (PF) и выявляли посредством FACScan Becton Dickinson (BD) оборудованного системой обнаружения CellQuest и программным обеспечением для анализа (BD). Ворота строили на живой клеточной популяции как определяли посредством профиля рассеивания FSC и SSC и выявили ≥10000 событий. Для определения доли положительных клеток ворота установили на основе клеток, обработанных отрицательным контролем (меченых контролем изотипа или клеток, меченых только F(ab') 2 козьих антител против мышиных антител Alexa-488). Процент специфических положительных клеток рассчитывали как: Относительную среднюю интенсивность флуоресценции (MFI) определяли как MFI тестового образца минус MFI контрольного образца. Связывание S1/S2-TBD по отношению к IgG1 и IgG2a на DC после периода от 1 до 4 суток культивирования показано в табл. 12. Таблица 12. Связывание химерного антигена или антитела с созревающими дендритными клетками Связывание S1/S2-TBD было отчетливо более сильным, чем связывание или IgGl, или IgG2a с очевидно большим S1/S2-TBD связыванием на сутки 1, чем на сутки 4. Данные эксперименты демонстрируют, что S1/S2-TBD связывается с созревающими дендритными клетками. 3. Захват химерных антигенов созревающими дендритными клетками. Для определения степени захвата химерных антигенов (например, S1/S2 HBV-TBD) по сравнению с IgG1 и IgG2a клетки инкубировали с различными концентрациями антигена, IgG1 (2C12, родительское антитело mAb, из которого получен TBD) или IgG2a (G155-178) в течение 1 ч при 37 °С в среде AIM-V с 0,1% BSA. Клетки дважды отмывали в PBSB и фиксировали PBS с 2% PF в течение ночи при 4 °С. Далее, клетки дважды отмывали в PBSB и нарушали проницаемость клеточной мембраны PBS, содержащим 0,1% (мас./об.) сапонин (Sigma), в течение 40 мин при 20 °С. Клетки дважды отмывали PBSB и инкубировали с F(ab') 2 козьих антител к мышиным антителам Alexa-488 (10 мкг/мл) в PBSB с 0,1% (мас. /об.) сапонина в течение 20 мин при 4 °С. После двукратной отмывки в PBSB клетки ресуспендировали в PBSB. Вариант данного анализа включает в себя обработку клеток как выше химерным антигеном, IgG1или IgG2a в течение 10 мин с последующим добавлением F(ab') 2 козьих антител к мышиным антителам Alexa-488 (10 мкг/мл) на 50 мин. Затем клетки отмывали и ресуспендировали в PBS с 2% PF. Клетки выявляли посредством FACScan Becton Dickinson (BD) оборудованного системой обнаружения CellQuest и программным обеспечением для анализа (BD). Ворота строили на живой клеточной популяции, как определяли посредством профиля рассеивания FSC и SSC и выявили ≥10000 событий. Для определения доли положительных клеток ворота установили на основе клеток, обработанных отрицательным контролем (меченых контролем изотипа или клеток, меченых только F(ab') 2 козьих антител против мышиных антител Alexa-488). Процент специфических положительных клеток рассчитывали как: Относительную среднюю интенсивность флуоресценции (MFI) определяли как MFI тестового образца минус MFI контрольного образца. Захват S1/S2-TBD в сравнении с мышиными IgG1 и IgG2a оценивали как функцию концентрации на 4 сутки созревания дендритных клеток. Захват количественно подсчитывали при 37 °С в течение 1 ч, а результаты показаны на фиг. 4. Существовало линейное увеличение захвата S1/S2-TBD в зависимости от концентрации. IgG1 захватывался на значительно более низком уровне, и существовал очень слабый захват IgG2a. Следовательно, химерный антиген S1/S2-TBD захватывается дендритными клетками более эффективно, чем иммуноглобулины. J. Пример 10. Экспрессия рецепторов Fc- γ и CD206 на созревающих DC. На антигенпрезентирующих клетках находятся несколько рецепторов, связывающих и захватывающих антигены. Относительное содержание этих рецепторов на созревающих дендритных клетках оценивали с применением флуоресцентно меченых специфичных к рецепторам антител. Для расчета доли положительных на специфические рецепторы клеток в общей популяции дендритных клеток применяли анализ FACS. Степень экспрессии рецепторов оценивали посредством определения относительной средней интенсивности флуоресценции и как функцию от относительной интенсивности флуоресценции (табл. 13). Экспрессия CD64 (рецептор Fc γ I) уменьшалась в течение периода культивирования и на сутки 4 была почти ничтожной. Напротив, CD32 (рецептор Fc γ II) и в меньшей степени CD16 (рецептор Fc γ III) продолжали экспрессироваться после 4 суток культивирования DC. На сутки 0 культивирования по существу не было экспрессии CD206 (макрофагальный маннозный рецептор). Но экспрессия индуцировалась при культивировании с IL-4 и GM-CSF, и на сутки 4 CD206 экспрессировался на очень высоких уровнях. Таким образом, на сутки 4, когда в анализах антигенной презентации нагружали антигеном, дендритные клетки несли по меньшей мере два потенциальных рецептора для связывания химерных антигенов: CD32 и CD206. Кроме того, они обладают полным комплектом костимулирующих молекул (данные не показаны). Экспрессия HLA-DR (класс II) и HLA-ABC (класс I) также увеличивалась с течением времени культивирования. Костимулирующие молекулы CD86 (В7.2) и CD80 (В7.1) экспрессировались на протяжении всего периода анализа. Данные результаты указывают на то, что полученные из моноцитов дендритные клетки дифференцировались в зрелые дендритные клетки и были способны к процессингу и презентации антигенов Т-клеткам. K. Пример 11. Корреляция экспрессии CD32/CD206 и S1/S2-TBD. Связывание с созревающими DC. Существует прямая связь между экспрессией рецепторов CD32/CD206 и связыванием S1/S2-TBD с созревающими дендритными клетками. Так как было известно, что мышиные IgG1 связываются с человеческим CD32, ожидали, что S1/S2-TBD, содержащий мышиный компонент Fc IgGl, также может связываться с CD32. Кроме того, можно ожидать, что S1/S2-TBD вследствие высокой степени его гликозилирования маннозой, также будет связываться с дендритными клетками через рецептор CD206. На точечных диаграммах на фиг. 5 показано связывание S1/S2-TBD (10 мкг/мл) и экспрессия CD32, а также связывание S1/S2-TBD и экспрессия CD206. Существовала прямая корреляция между степенью связывания S1/S2-TBD и степени экспрессии CD32, которая являлась относительно гетерогенной, т.е., существовала широкая степень экспрессии. Данные результаты демонстрируют, что S1/S2-TBD связывается с CD32 и что чем больше экспрессия CD32, тем больше степень связывания химерного антигена S1/S2-TBD. Точечная диаграмма связывания S1/S2-TBD и экспрессии CD206 показывает, что абсолютное большинство клеток, экспрессирующих CD206, также связывали S1/S2-TBD. Небольшая доля клеточной популяции являлась негативной по CD206 и, следовательно, являлась негативной по связыванию S1/S2-TBD. Следовательно, и рецептор CD32, и рецептор CD206 коррелируют со связыванием S1/S2-TBD. L. Пример 12. Связывание и захват S1/S2-TBD первично происходит посредством CD32 при меньшей степени вовлеченности CD206. Захват S1/S2-TBD по сравнению с мышиными IgG1 и IgG2a оценивали как функцию концентрации на сутки 4 созревания DC. Захват количественно оценивали при 37 °С в течение 1 ч в присутствии среды, маннана (2 мг/мл, Sigma) и/или мышиного Fc γ (2 мг/мл, Jackson ImmunoResearch Laboratories). Маннан представляет собой конкурентный ингибитор связывания CD206 и, следовательно, захвата антигенов через CD206 на дендритных клетках. Fc γ представляет собой конкурентный ингибитор связывания CD32 и, следовательно, опосредованного CD32 захвата антигенов. Результаты показаны в табл. 14. Таблица 14. Ингибирование связывания химерного антигена посредством Fc или маннана Существовало прогрессирующее увеличение связывания химерного антигена при росте его концентрации. Инкубация клеток с высокой концентрацией фрагмента мышиного Fc γ устраняла данное связывание, тога как маннан, ингибитор связывания рецептора CD206, оказывал только незначительное действие. Следовательно, CD32 может являться первичным рецептором, вовлеченным в связывание и захват химерного антигена. М. Пример 13. Гликозилирование антигена S1/S2 HBV придает иммуногенность. Метаболический путь гликозилирования белка в клетке насекомого отличается от метаболического пути клетки млекопитающего тем, что белки, синтезируемые в клетках насекомых, претерпевают гликозилирование, приводящее к высокому содержанию маннозы и отсутствию концевых остатков сиаловой кислоты в секретируемом белке (Altman, et al., Glycoconjug 16:109-123(1999)). S1/S, и в клетках насекомых High Five ™ (гликозилирование маннозой). 1. Воздействие гликозилирования на связывание антигена. Данные антигены сравнивали по их связыванию с дендритными клетками, как описано в примере 9. Созревающие дендритные клетки нагружали 10 мкг/мл S1/S2 HBV экспрессированного в клетках насекомых или в E.coli. Гликозилированный белок продемонстрировал лучшее связывание с дендритными клетками (табл. 15). 2. Воздействие гликозилирования на вызов иммунного ответа. Гликозилирование S1/S2 HBV вызывает увеличенную иммуногенность и Т-клеточный ответ. S1/S2 HBV, экспрессированный и в Е. Coli, и в клетках насекомых High Five ™, сравнивали на Т-клеточный ответ при презентации дендритными клетками. Измеряли уровни и клеточного, и секретируемого интерферона- γ, как описано в примере 8 (с применением 2,5 г/мл белка S1/S2 HBV), а результаты представлены в табл. 16. S1/S2 HBV, экспрессируемый в клетках насекомых, вызывал более высокий уровень внутриклеточного и секретируемого интерферона по сравнению с негликозилированным белком, экспрессируемым в Е. coli. Все публикации и патенты, указанные в приведенном выше описании, включены сюда в качестве ссылки. Специалистам в данной области должны быть очевидны различные модификации и вариации описанного способа и системы согласно изобретению без отклонения от объема и сущности изобретения. Хотя изобретение описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что изобретение, как заявлено, не следует неправильно ограничивать такими конкретными вариантами осуществления. Фактически подразумеваются, что в объеме следующей ниже формулы изобретения находятся различные модификации описанных способов выполнения изобретения, которые очевидны специалистам в данной области. |