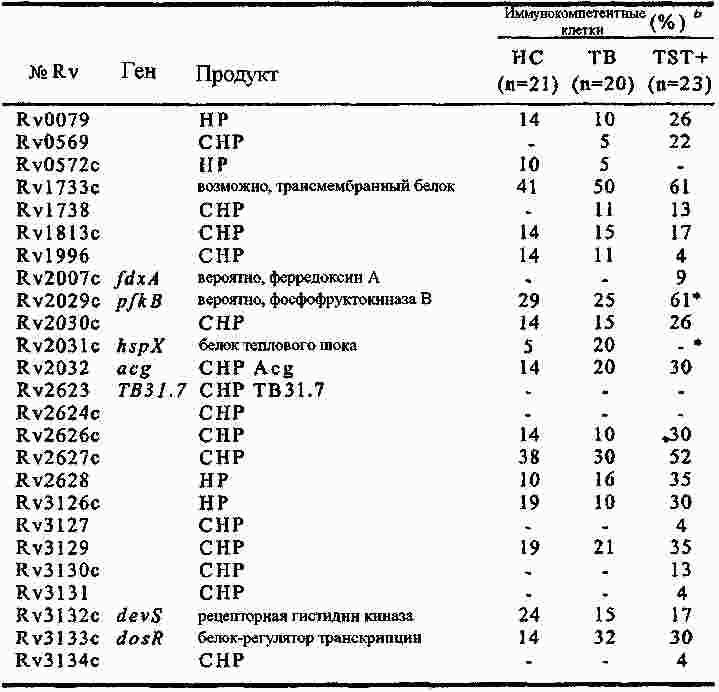
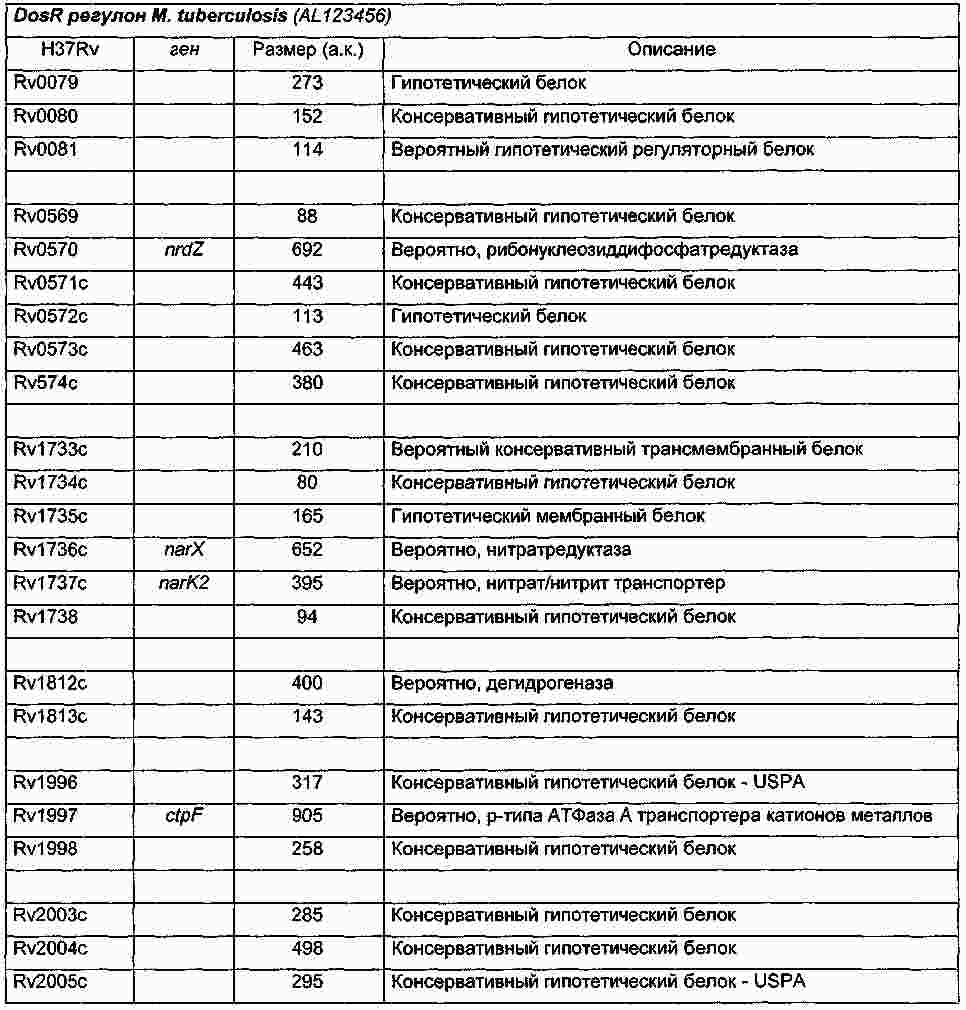
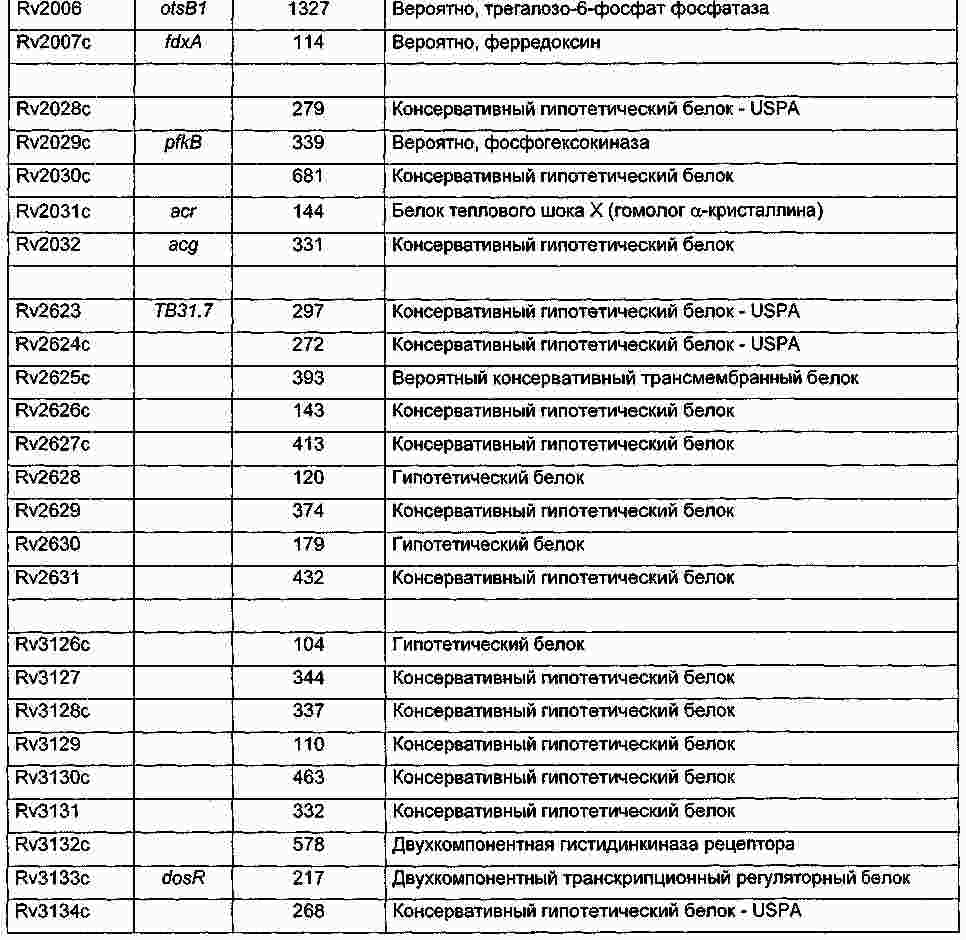
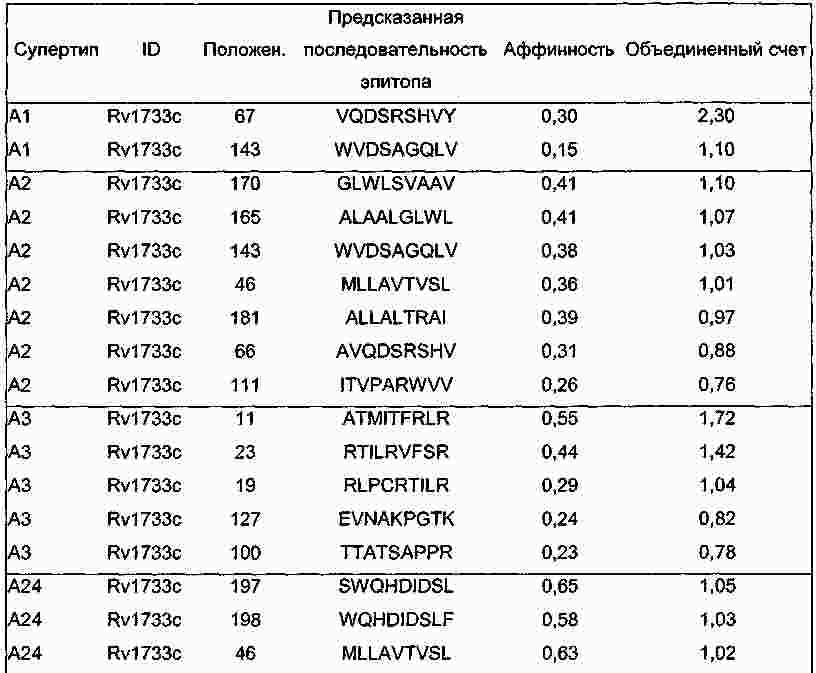
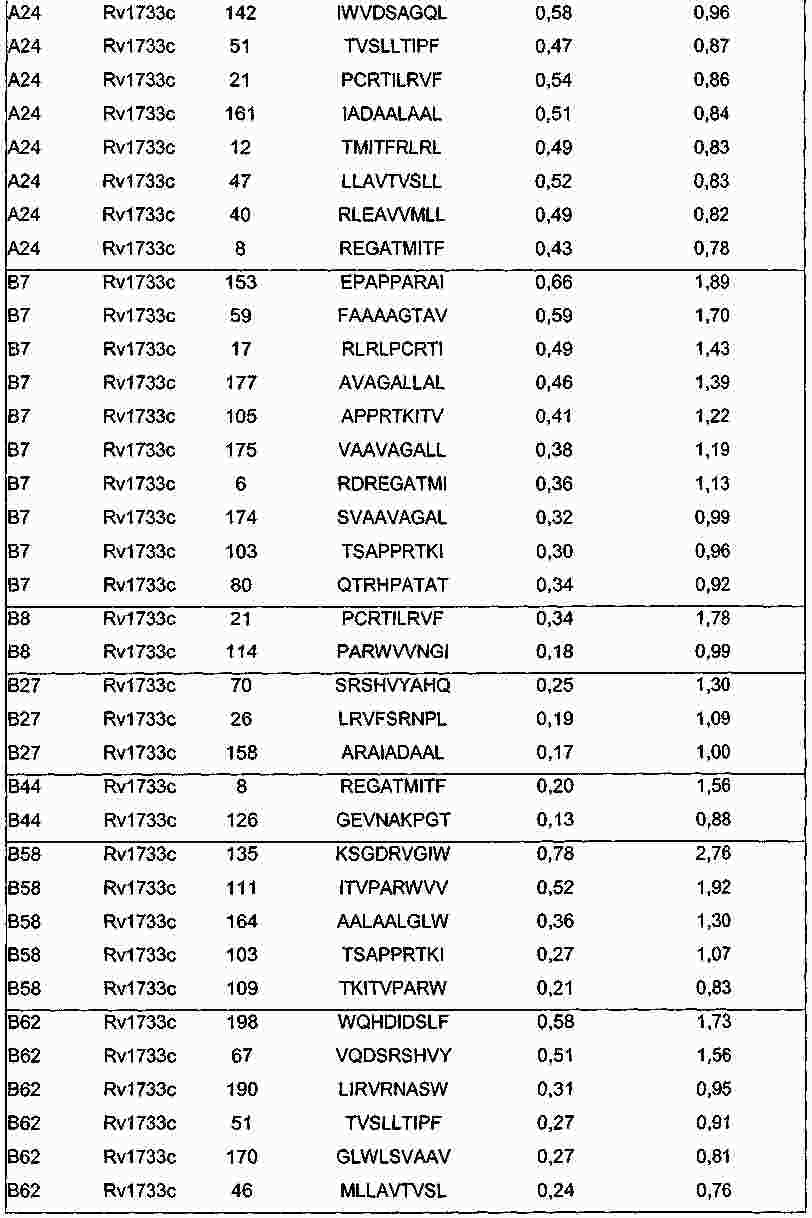
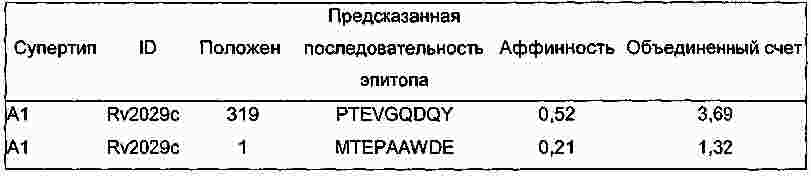
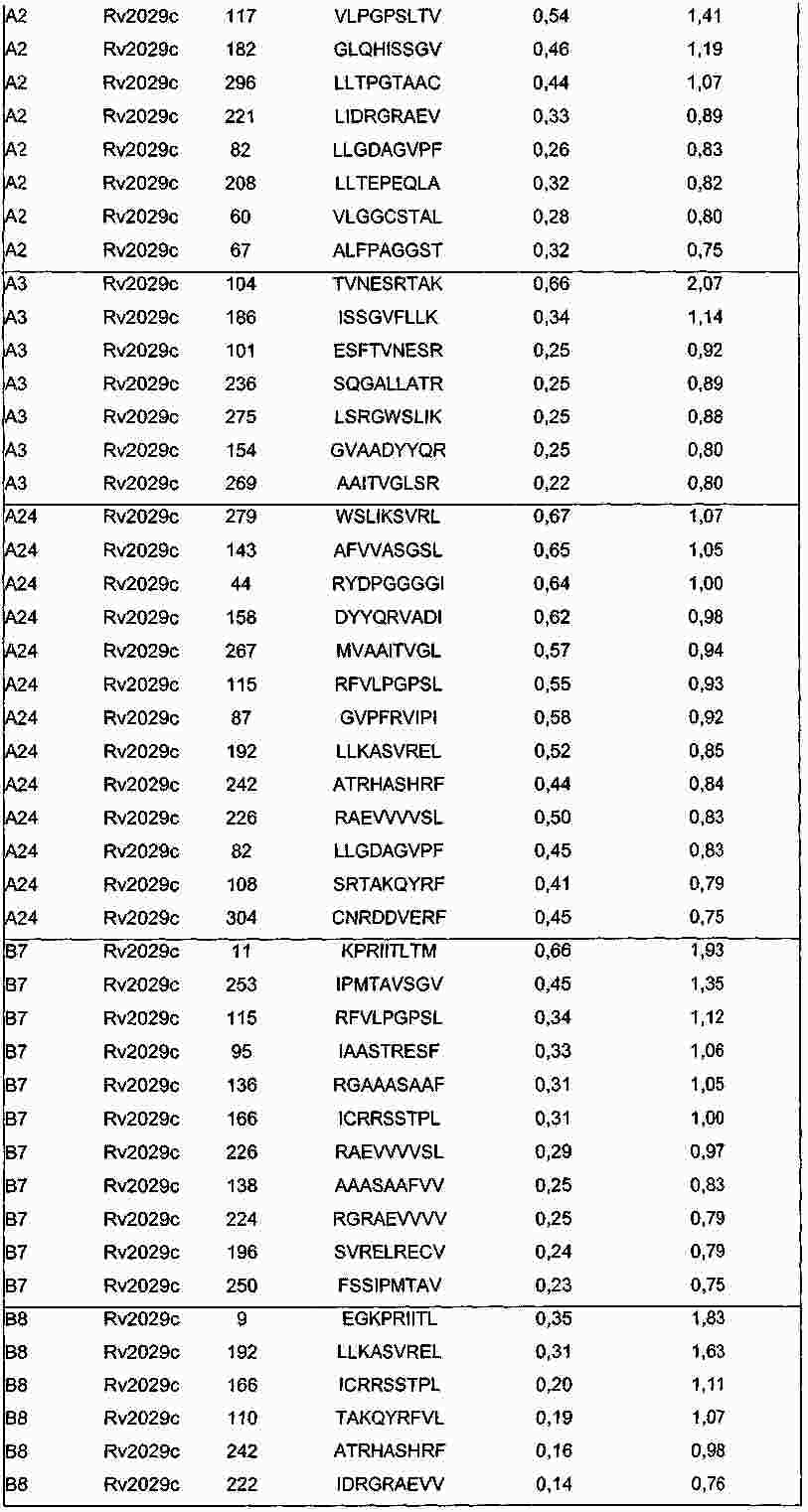
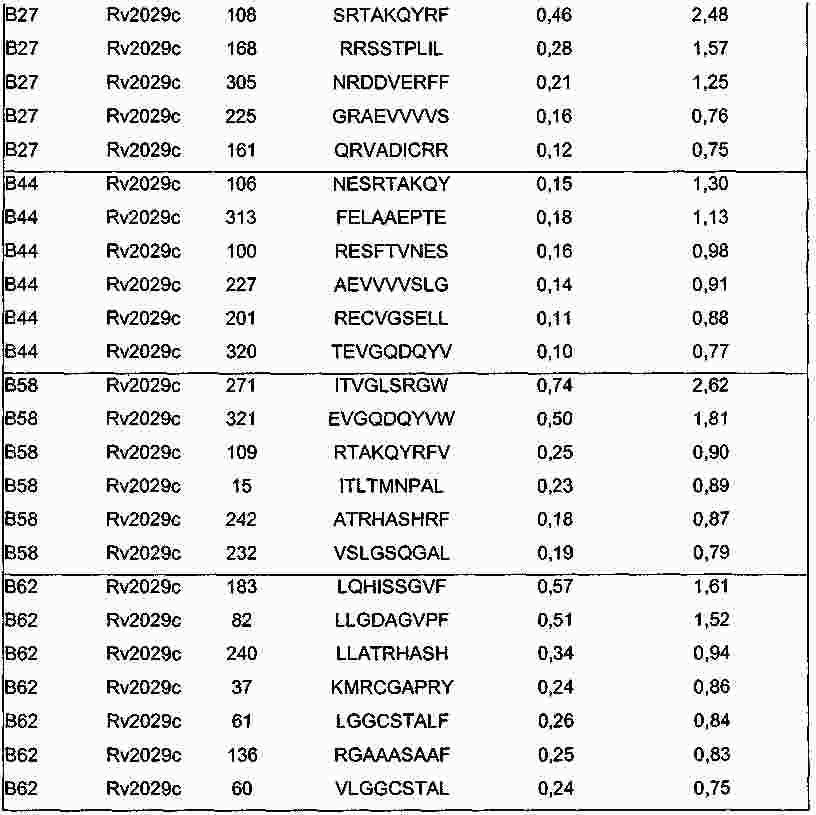
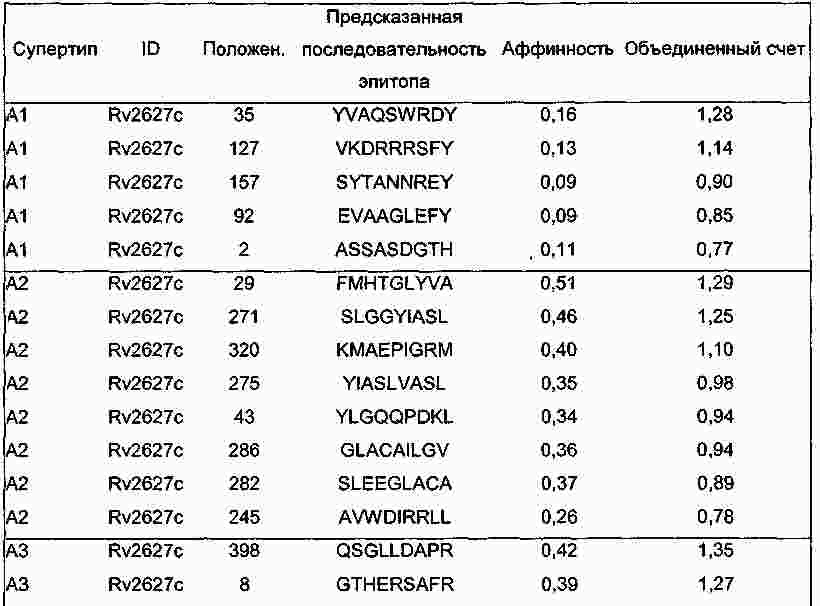
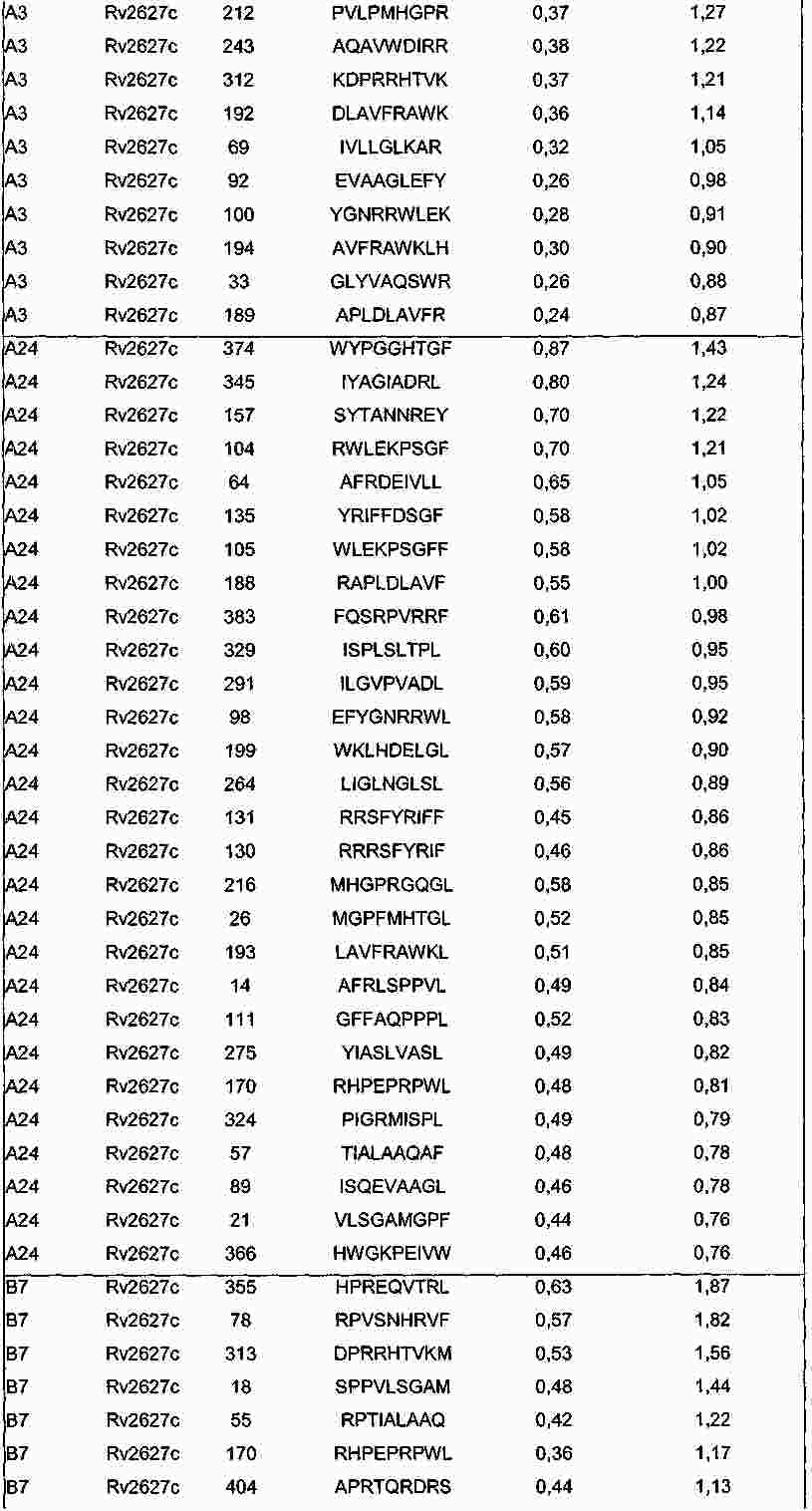
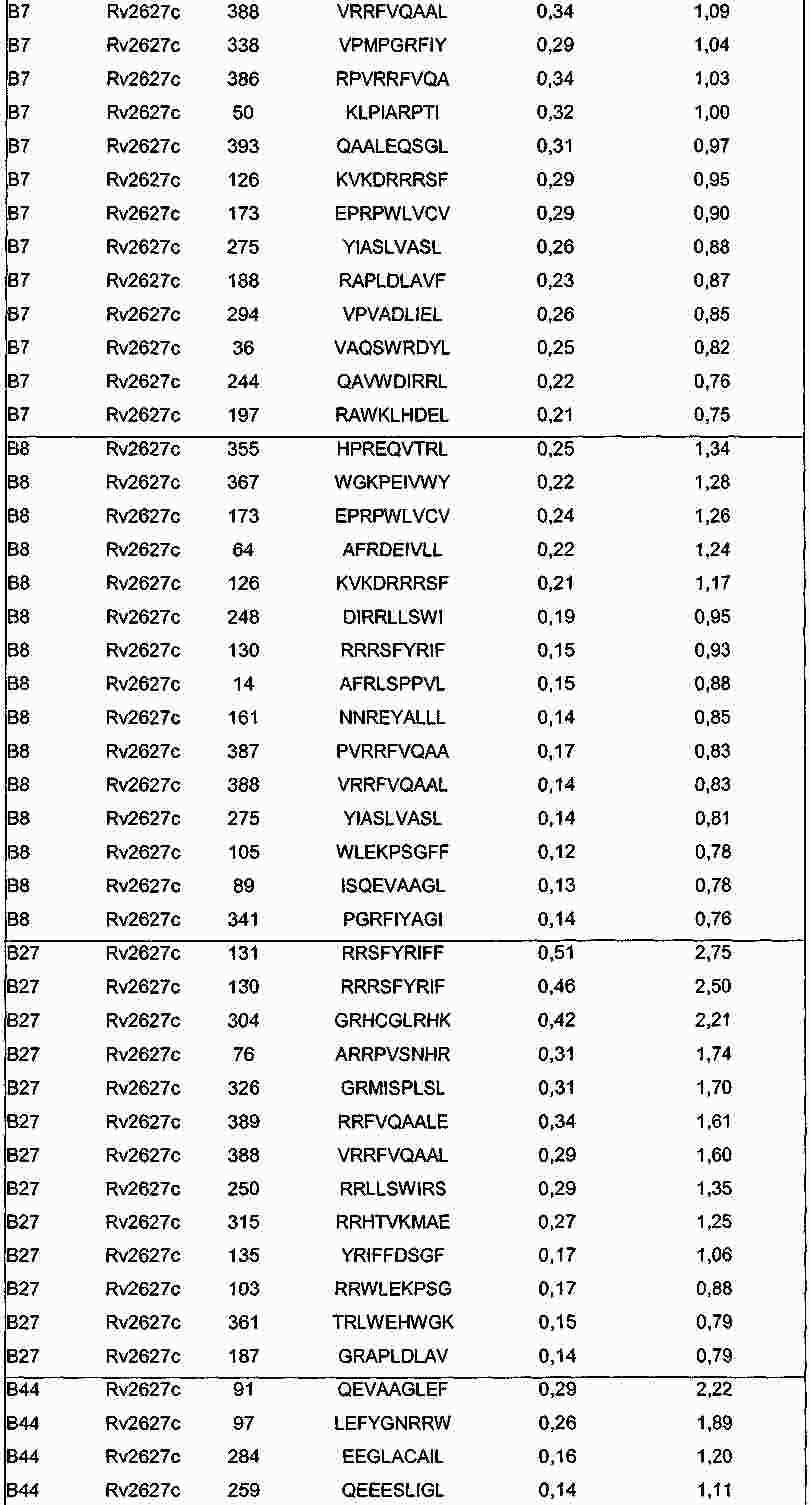
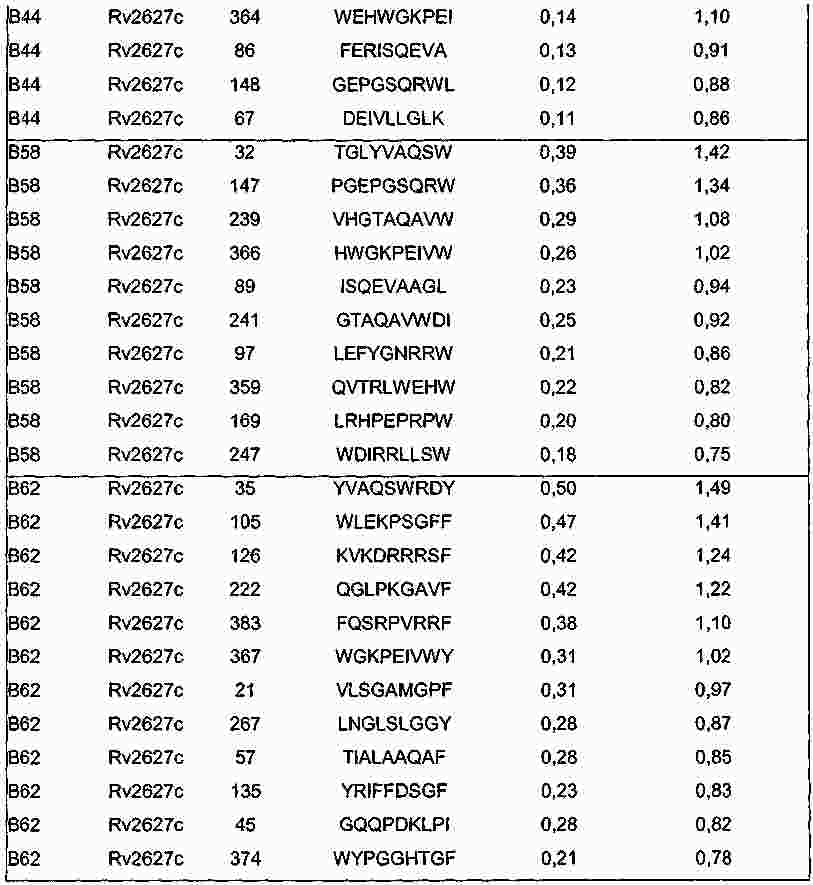
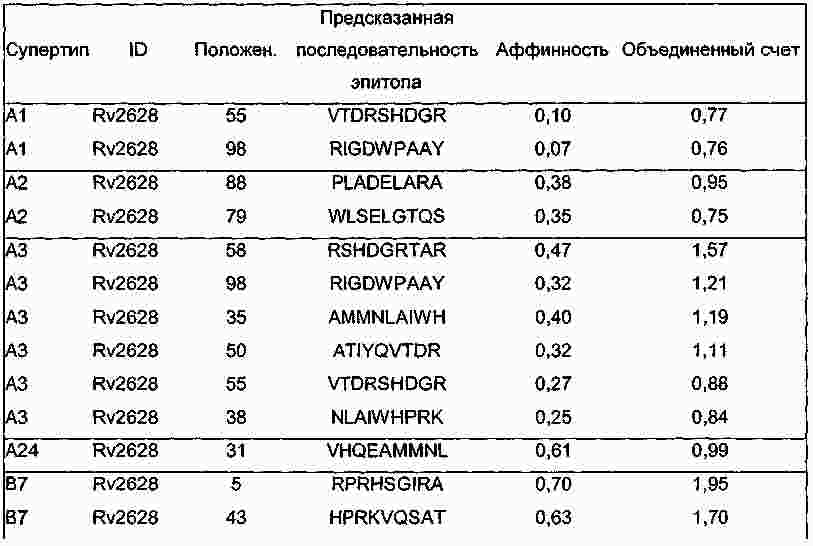
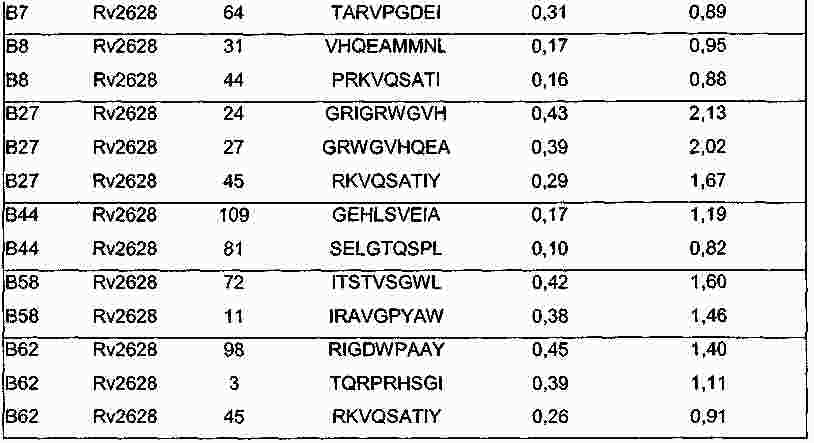
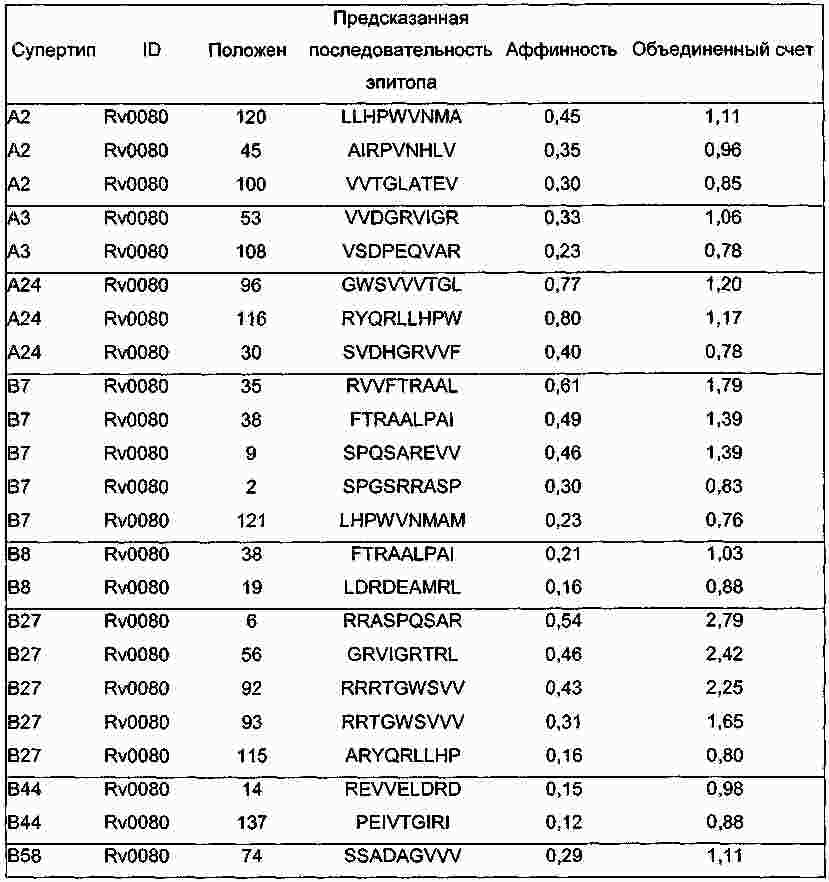
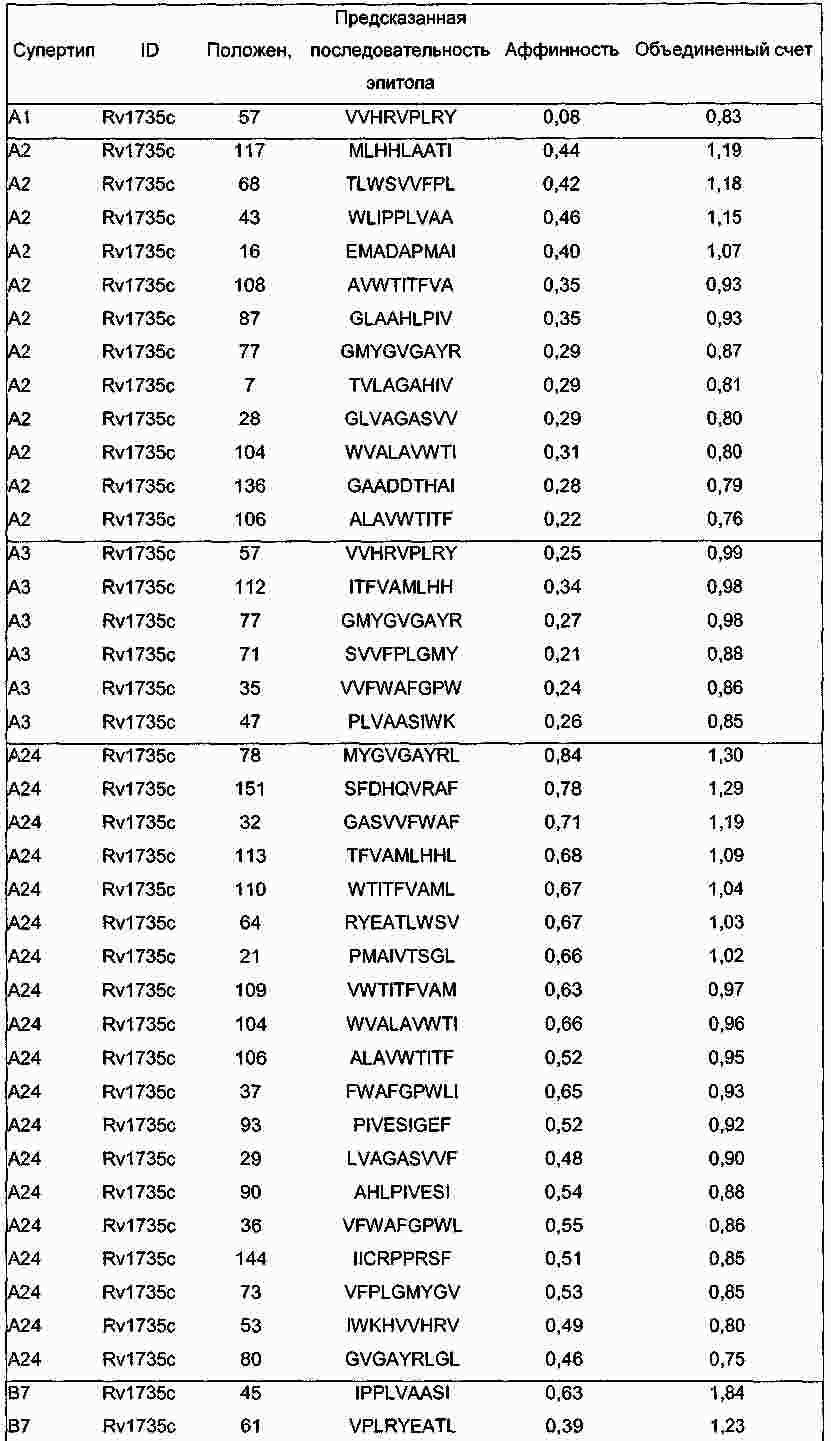
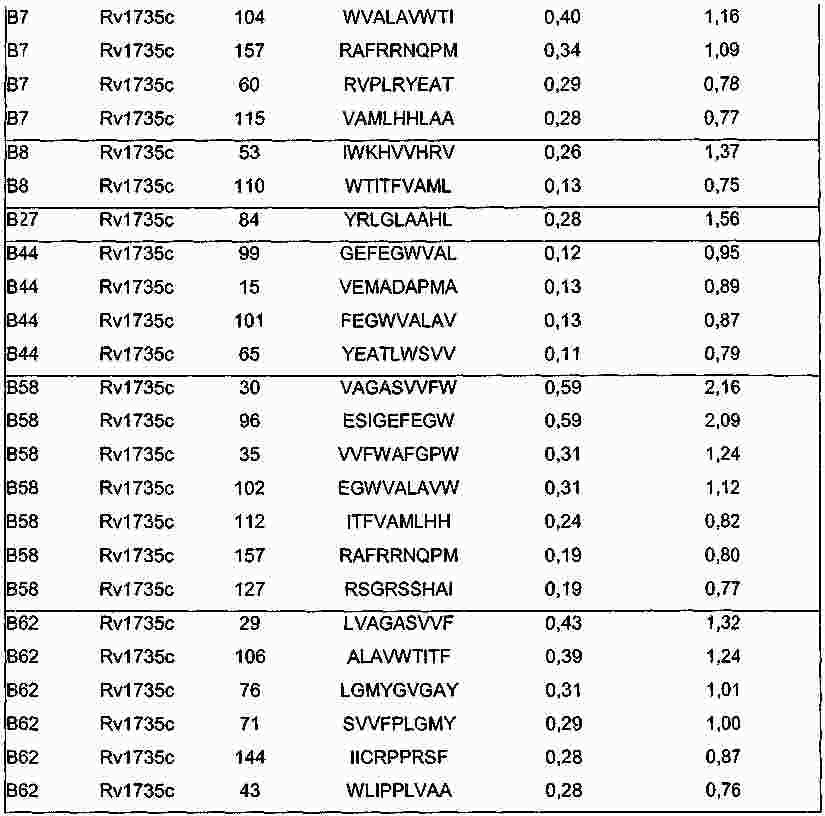
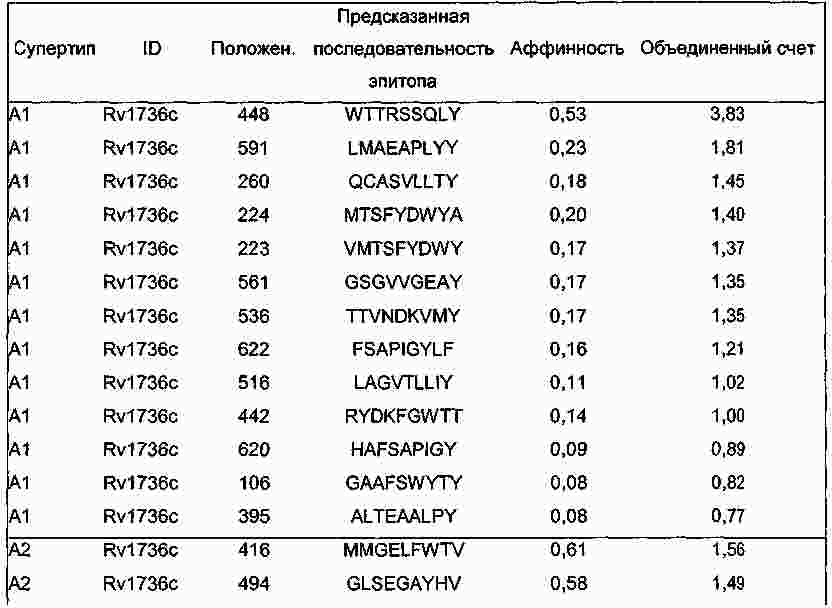
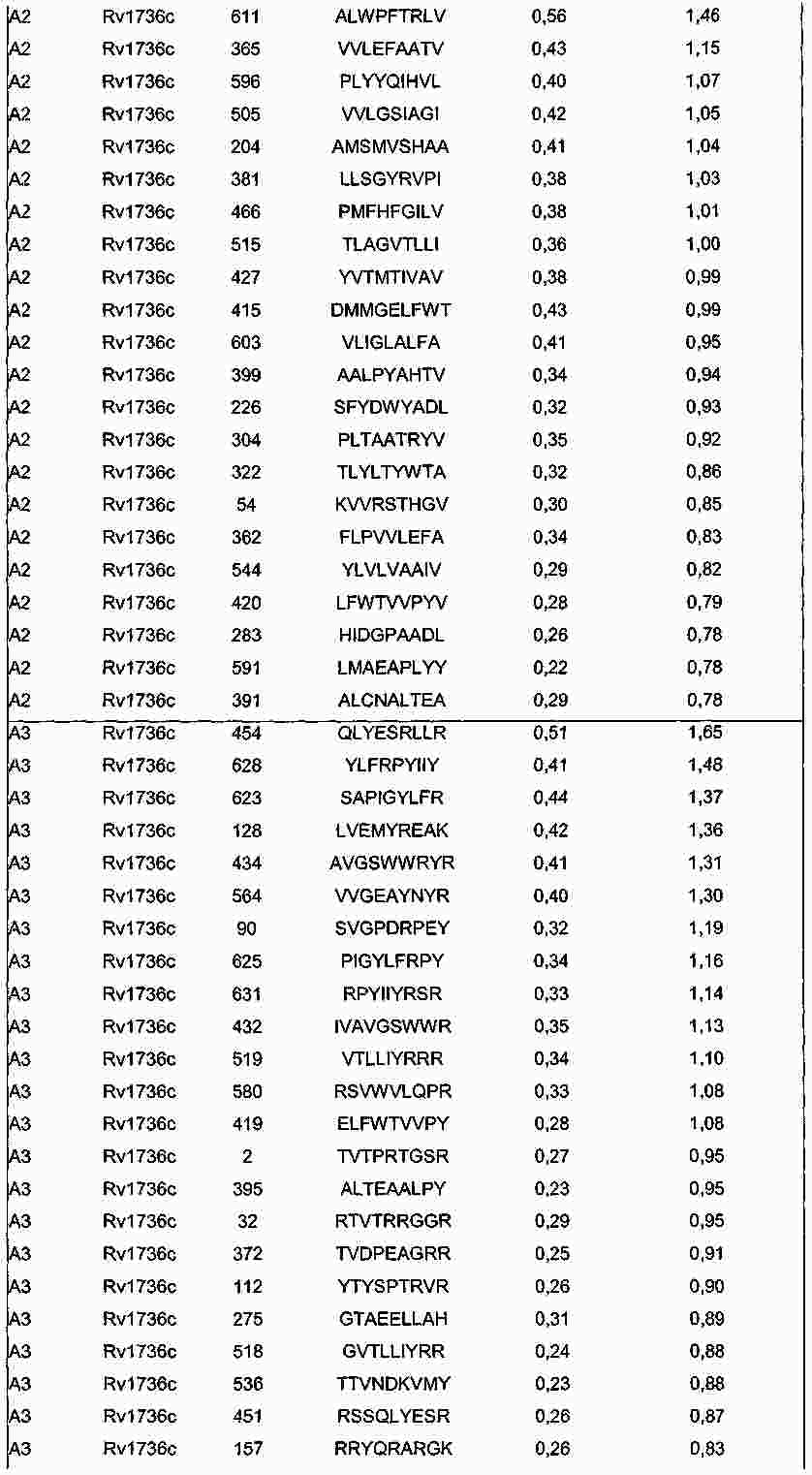
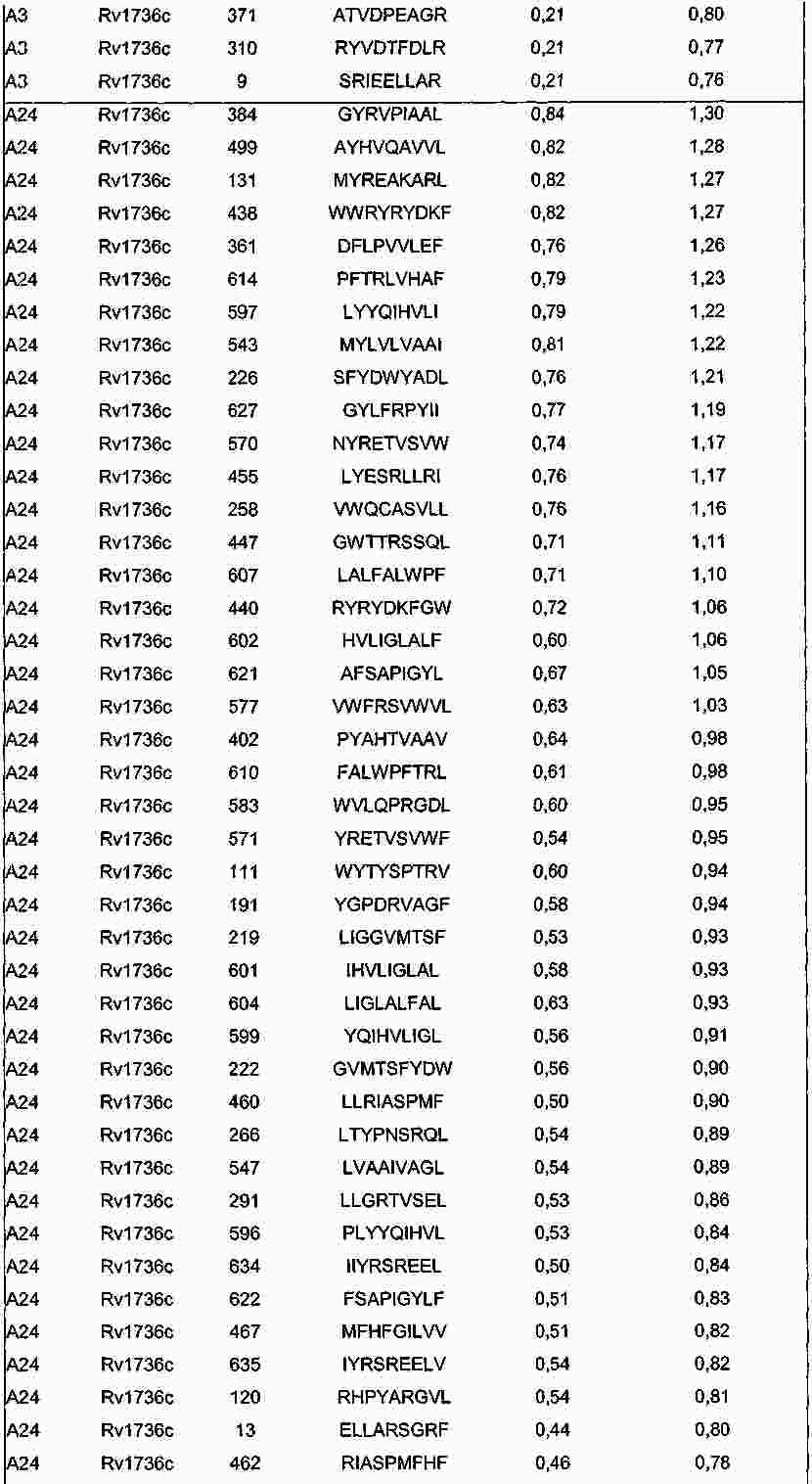
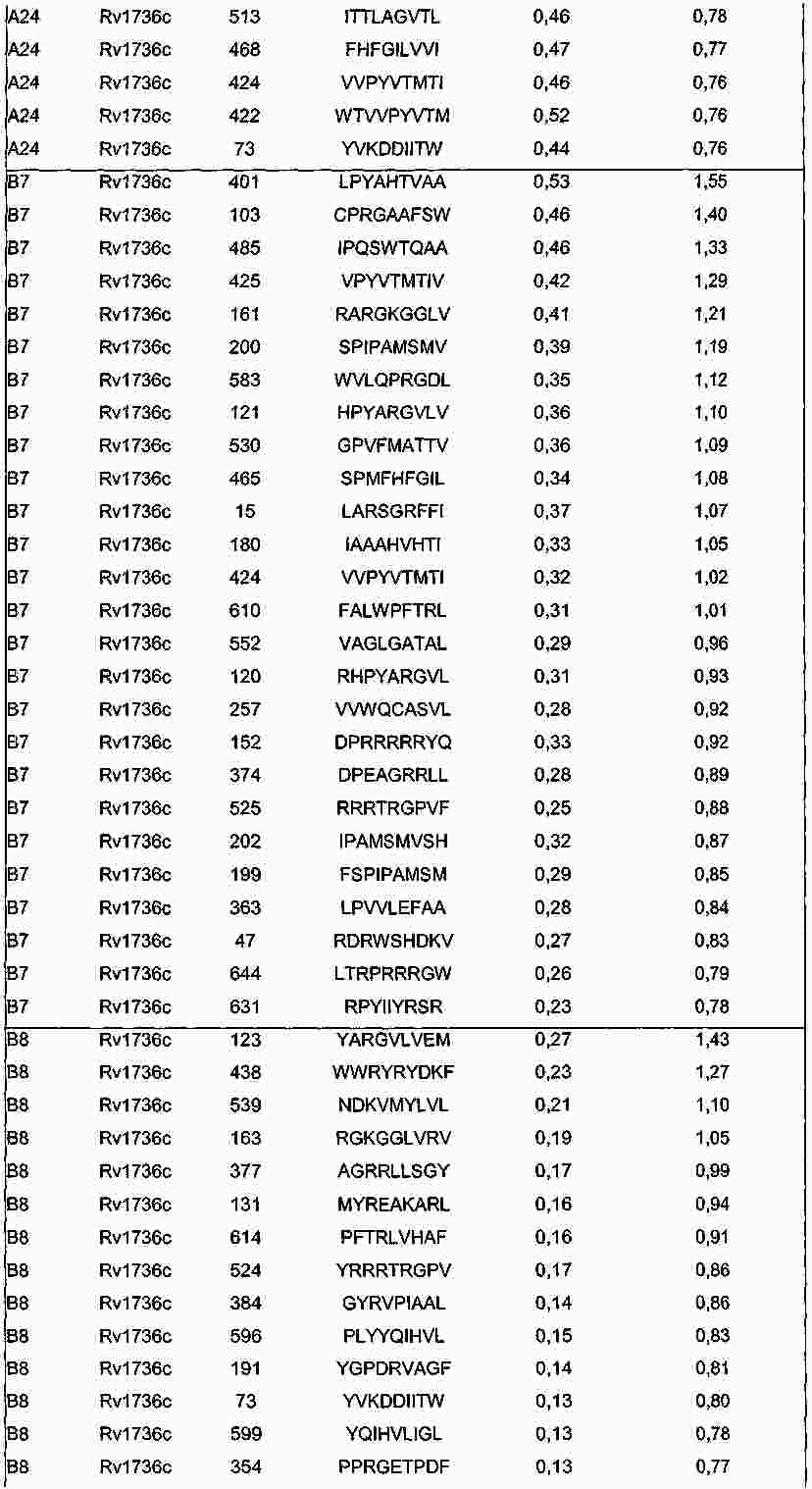
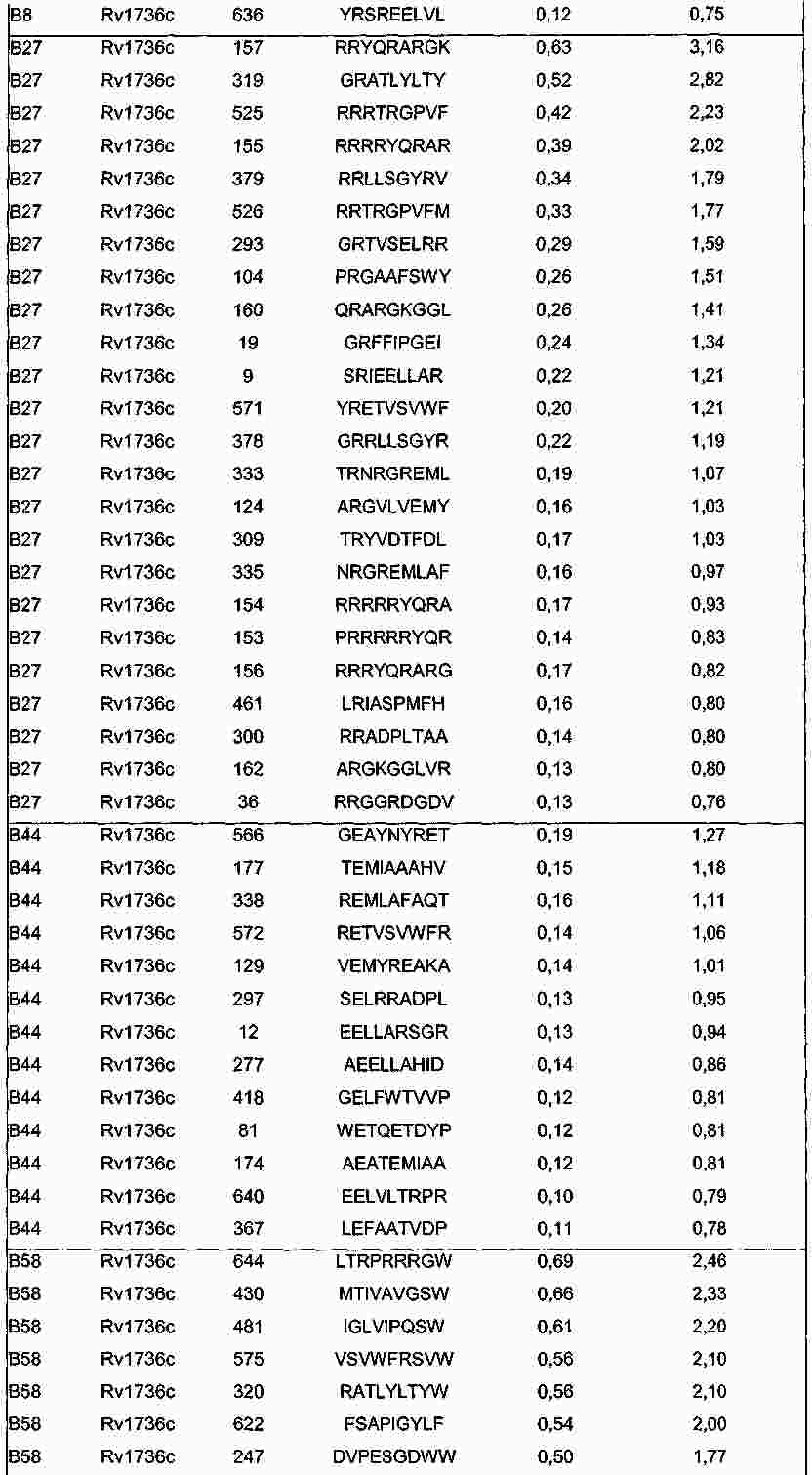
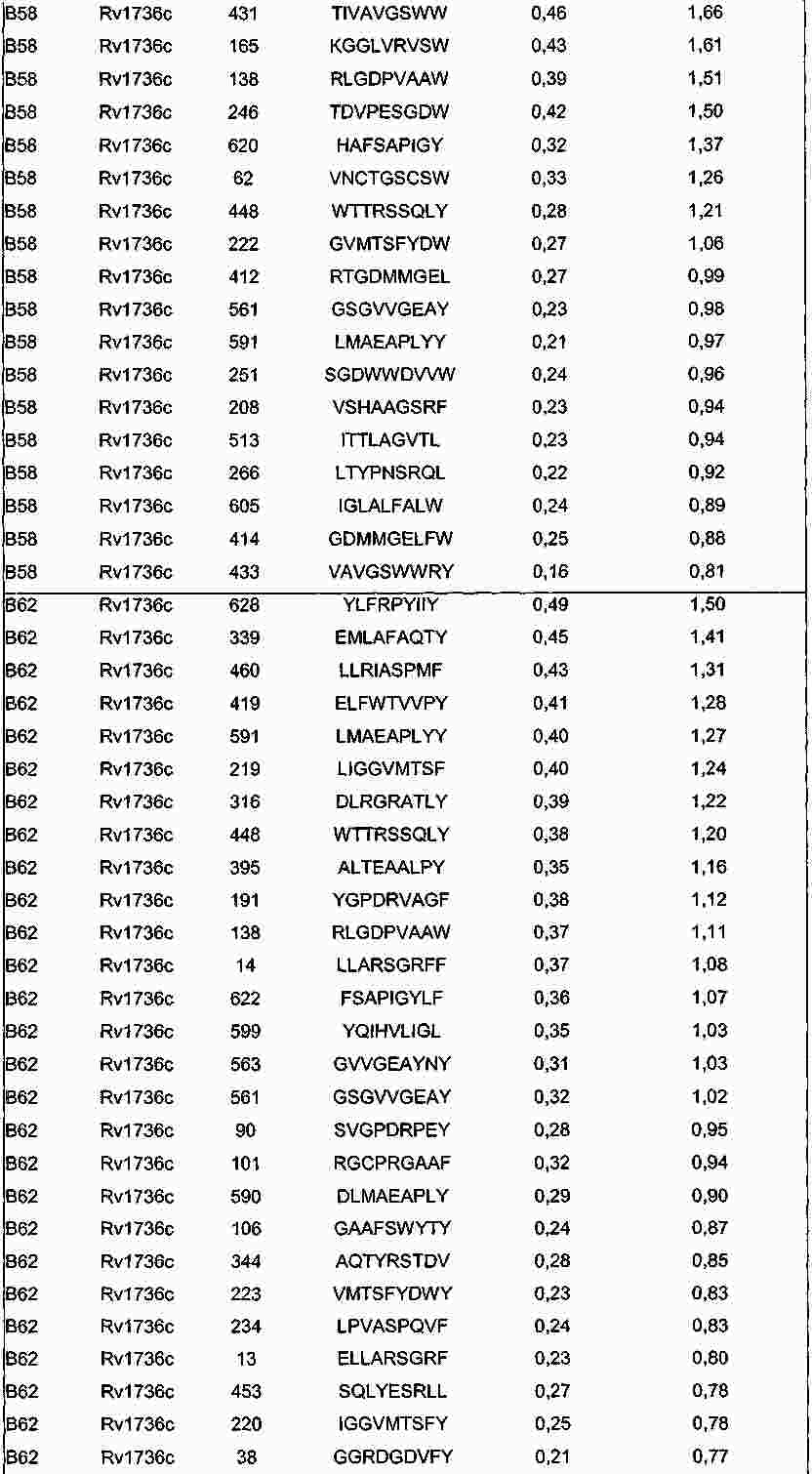
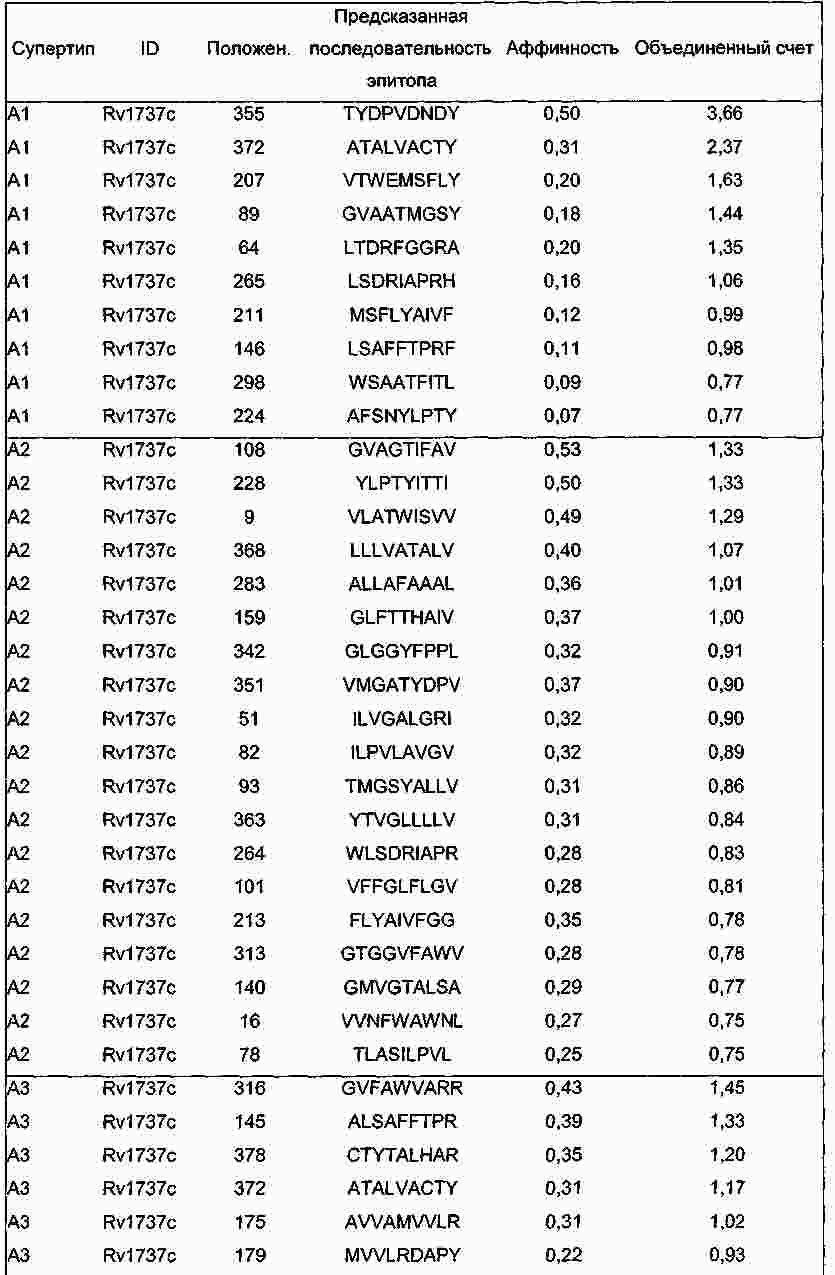
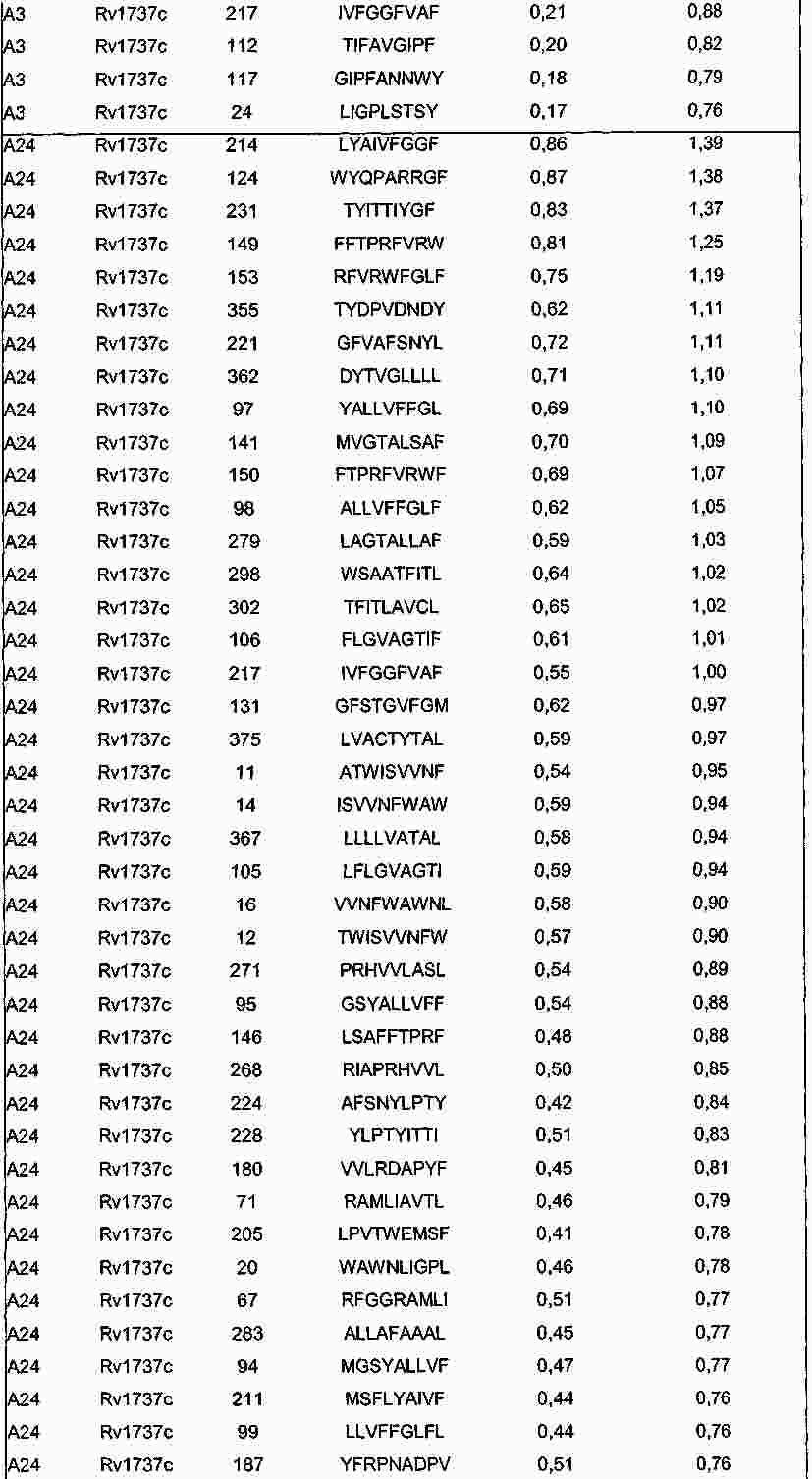
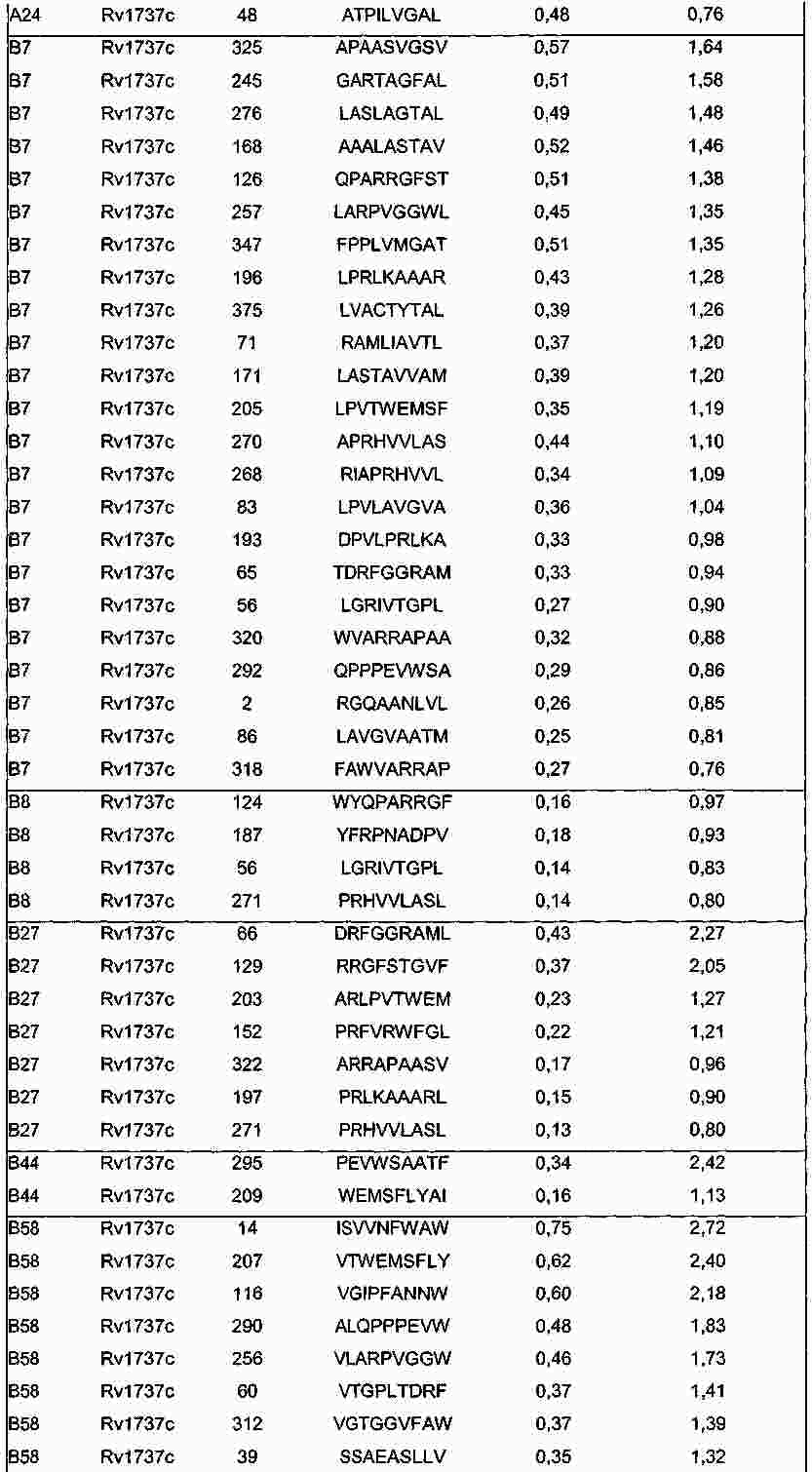
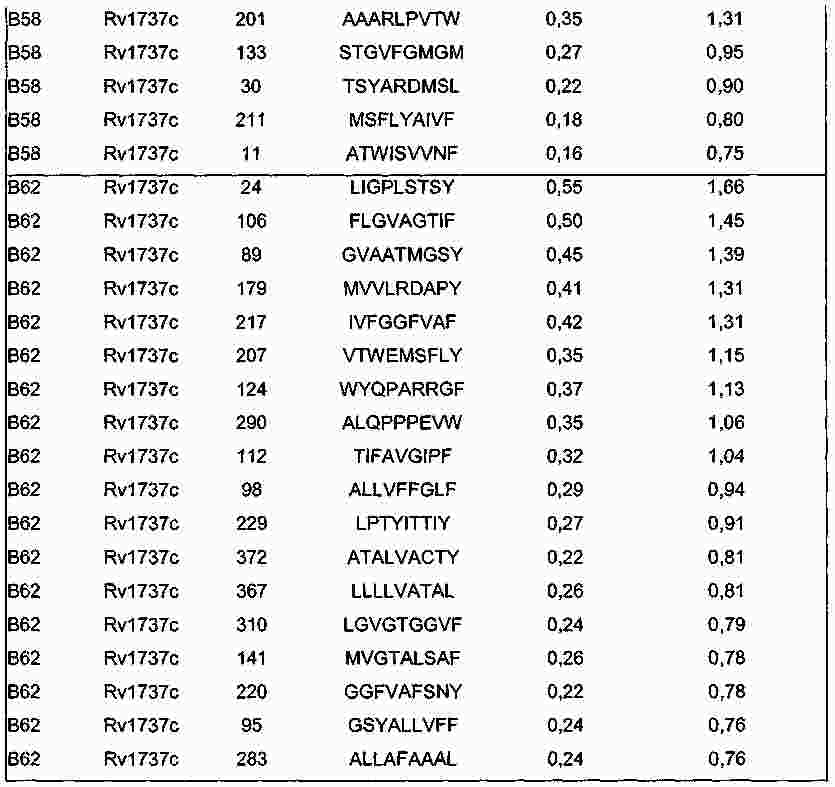
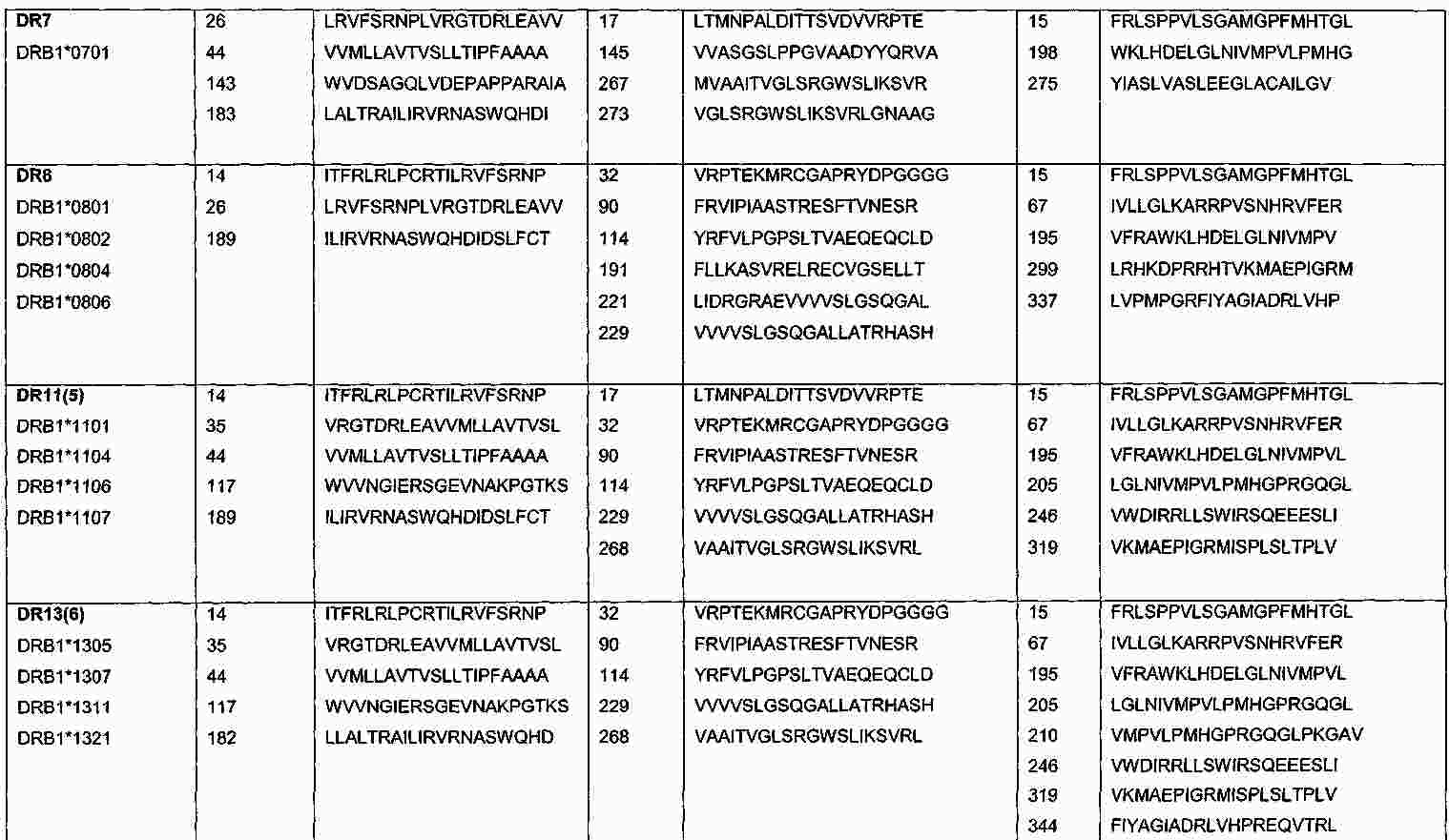
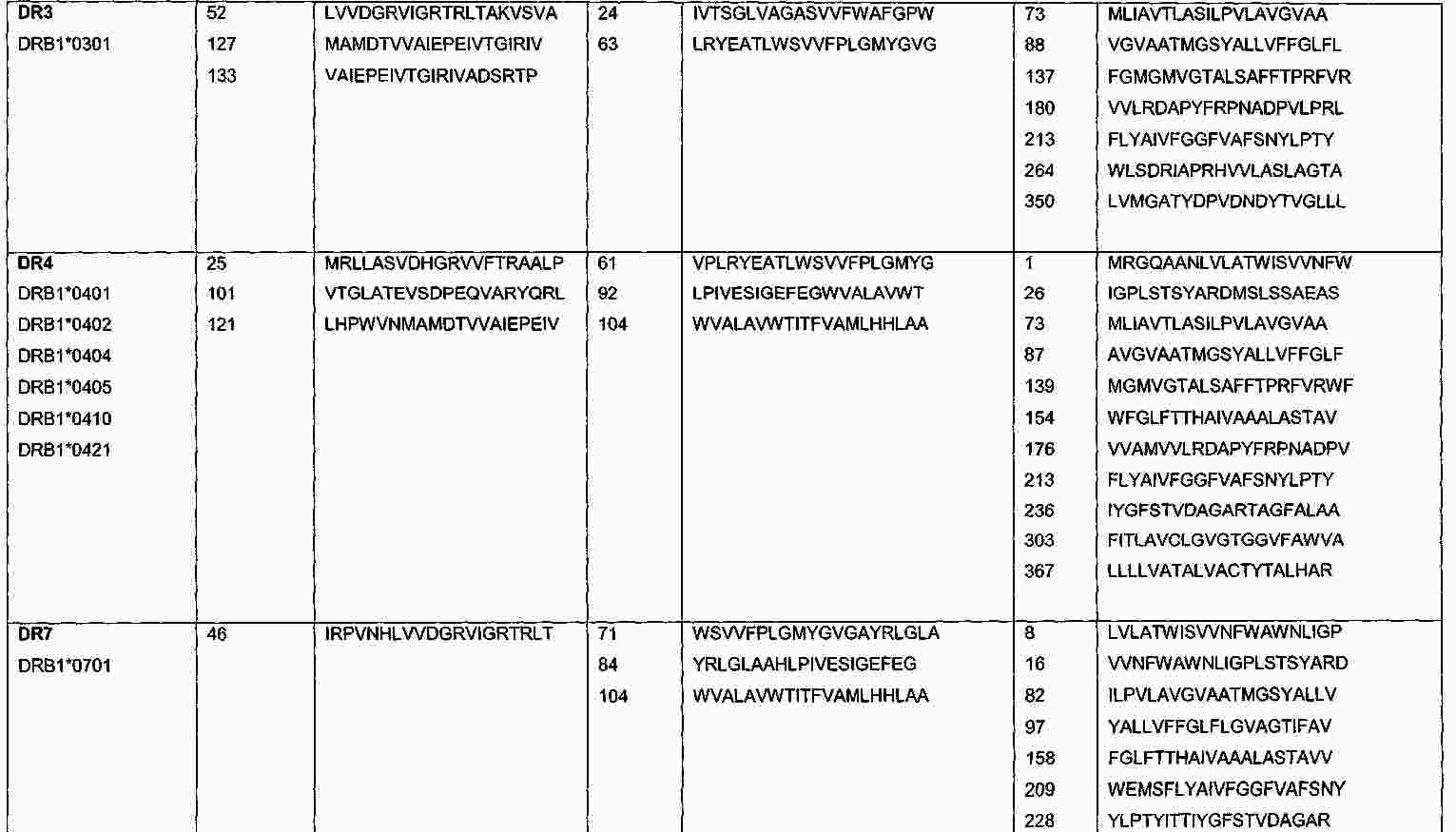
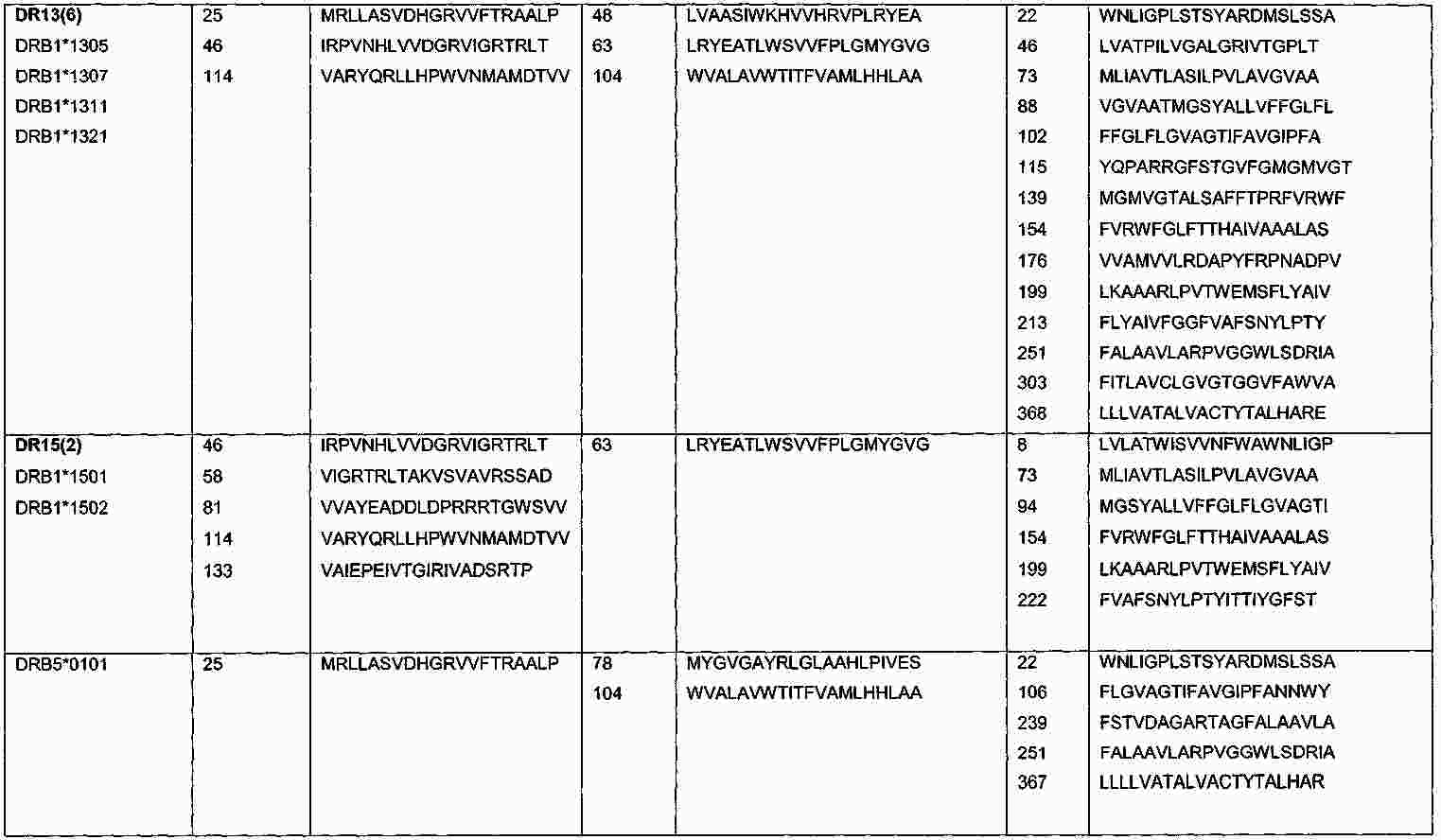
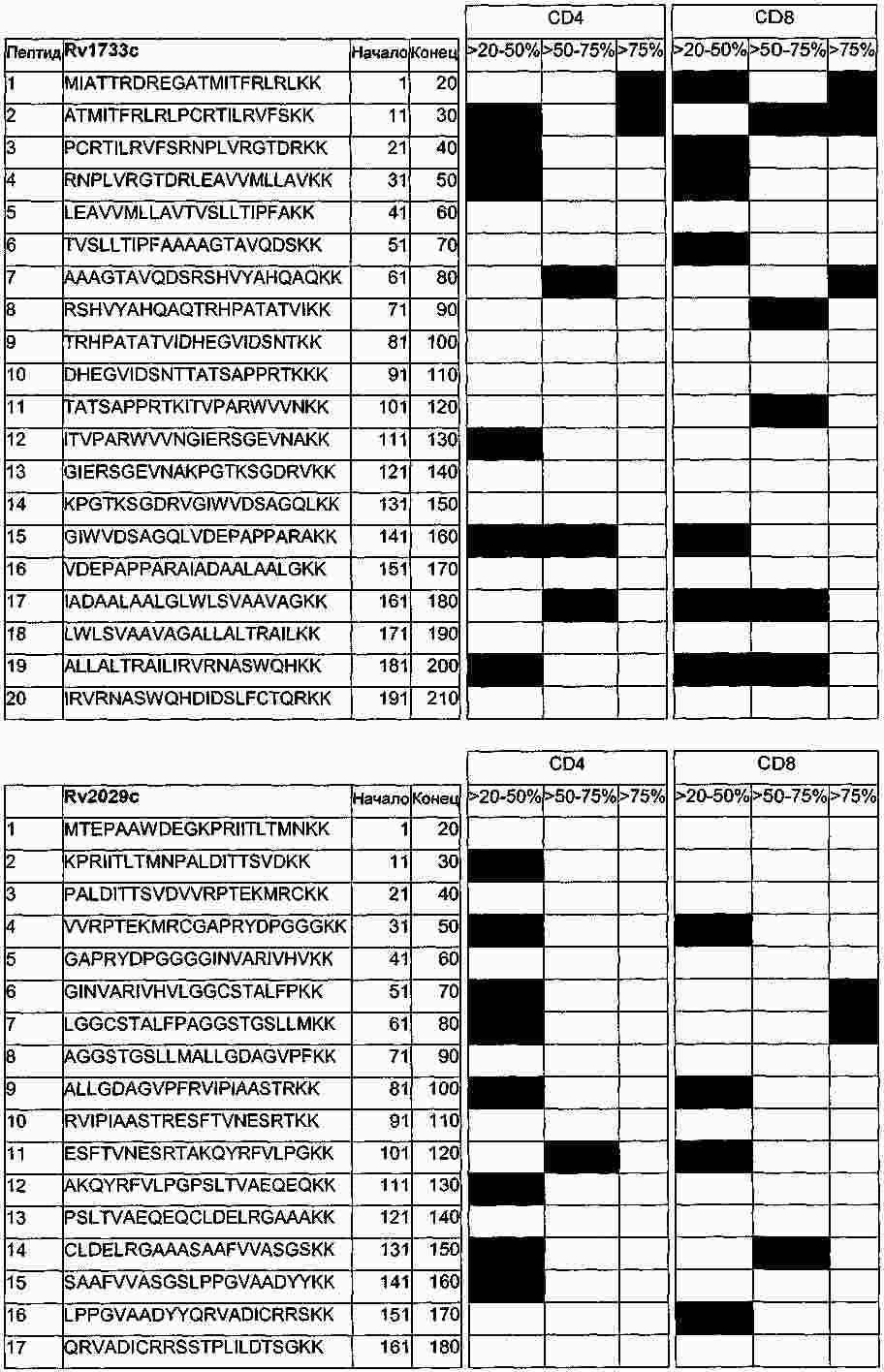
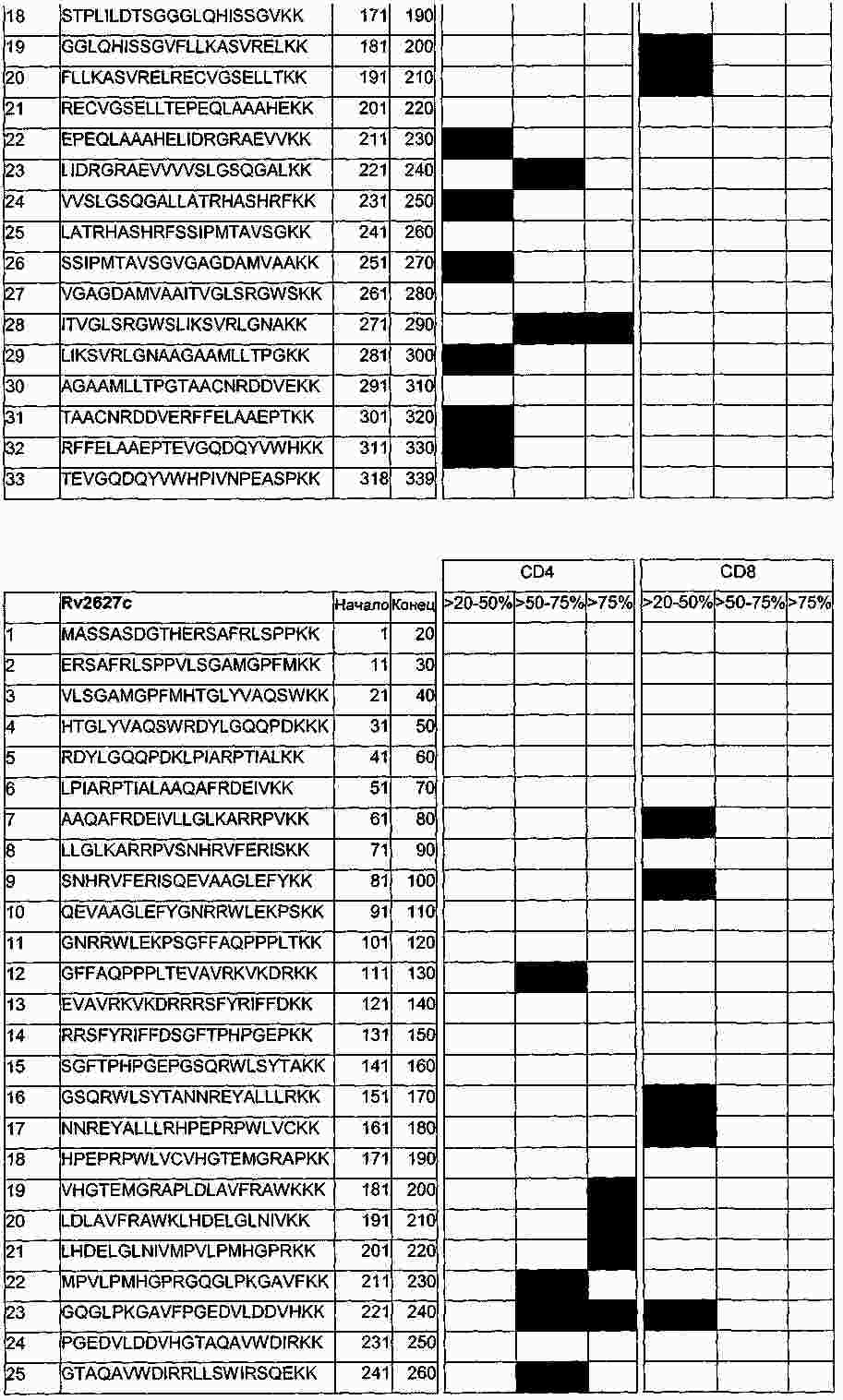
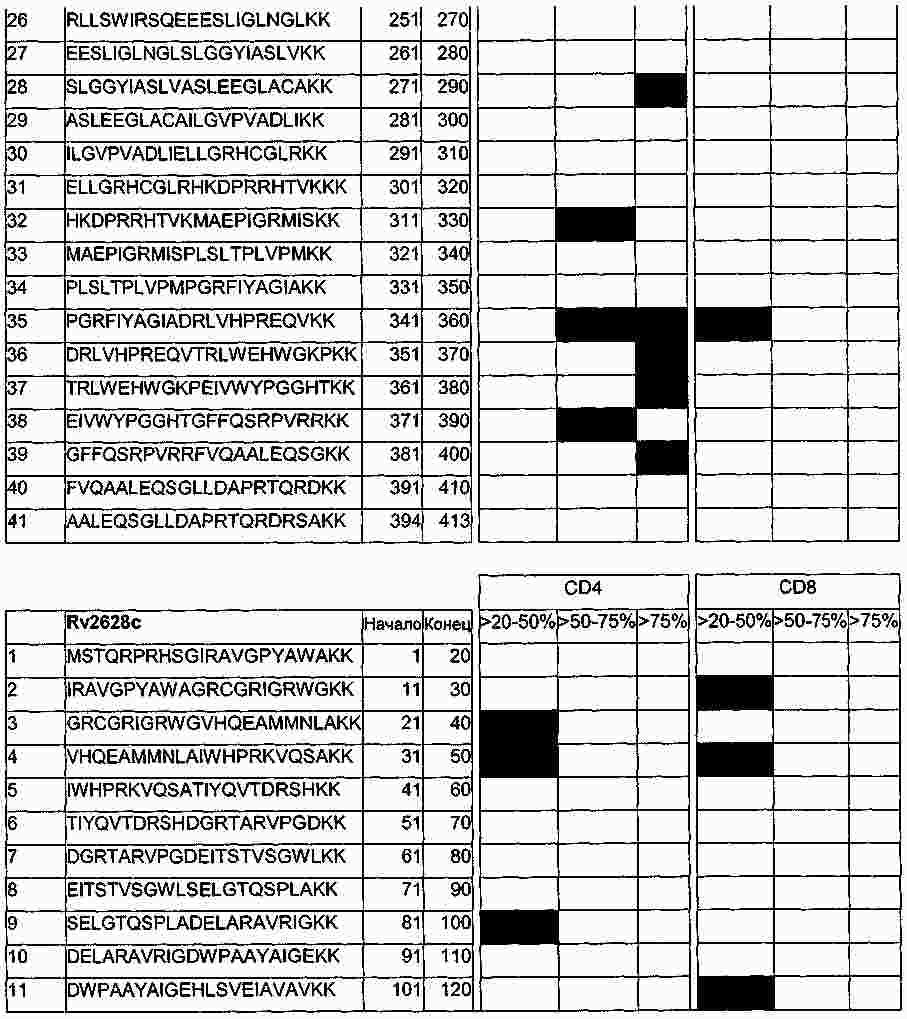
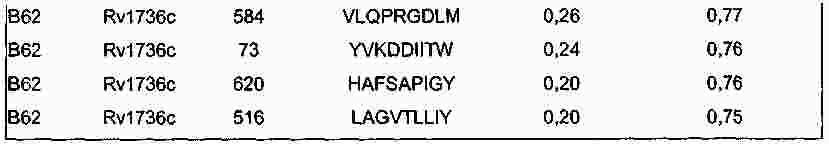|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000013016b*\id |
|
Термины запроса в документе Реферат Изобретение относится к небольшой подгруппе связанных с латентностью антигенов и/или эпитопов Mycobacterium, которые способны вызывать иммунный ответ in vivo и in vitro у млекопитающего. Изобретение относится к способам и композициям для детекции латентной инфекции Mycobacterium и иммунизации против них. Данные композиции содержат те микобактериальные латентные антигены, которые действительно способны вызывать иммунный ответ in vivo у млекопитающих, подвергающихся действию латентной инфекции Mycobacterium. Более предпочтительно композиции содержат те антигены, которые предпочтительно выявляются латентно инфицированными индивидуумами и которые не узнаются или в значительно меньшей степени узнаются у индивидуумов с активной инфекцией Mycobacterium или у индивидуумов с индуцированными Mycobacterium симптомами или заболеваниями, такими как у инфицированных М. tuberculosis больных, страдающих от заболевания туберкулезом (ТВ). Формула [0001] Способ индукции иммунного ответа на инфекцию Mycobacterium у позвоночного, включающий стадии введения позвоночному композиции, содержащей источник одного или более полипептидов, выбранных из группы, состоящей из последовательностей регулона спячки (DosR) M. tuberculosis:Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX), и аналогов, гомологов или их фрагментов. [0002] Способ по п.1, где полипептид выбран из группы, состоящей из Rv1733c, Rv2029c и Rv2627c. [0003] Способ по п.1, где полипептид получен, исходя из последовательностей ТВ регулона спячки (DosR) Rv1733c, Rv2029c (PfkB) и Rv0080. [0004] Композиция для иммунизации против инфекций Mycobacterium, содержащая источник из одного или более полипептидов, выбранных из группы, состоящей из последовательностей ТВ регулона спячки (DosR):Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX), и аналогов, гомологов или их фрагментов; и необязательно включающая адъювант. [0005] Композиция по п.4, где полипептид выбран из группы, состоящей из Rv1733c, Rv2029c и Rv2627c. [0006] Композиция по п.4, где полипептид получен, исходя из последовательностей ТВ регулона спячки (DosR) Rv1733c, Rv2029c (PfkB) и Rv0080. [0007] Композиция для иммунизации по любому из пп.4-6, где адъювант выбран из группы адъювантов, состоящей из polyI:C, CpG, LPS, липида А и их производных, IC31, QS21, липопептида Pam3Cys, бактериальных флагеллинов, DDA/MPL, DDA/TDB и растворимого LAG3. [0008] Композиция для иммунизации по любому из пп.4-7, где полипептид Mycobacterium выбран из ТВ комплекса видов Mycobacterium M. tuberculosis, M. bovis, M. bovis BCG, М. africanum, M. canetti и М. microti. [0009] Композиция для иммунизации по любому из пп.4-8, где источник полипептида представляет собой молекулу рекомбинантной ДНК, кодирующей полипептид, необязательно включенную в вектор и/или в геном. [0010] Композиция для иммунизации по любому из пп.4-9, где источником полипептида является рекомбинантный Mycobacterium, предпочтительно Mycobacterium bovis бацилла Кальмета-Герена (BCG). [0011] Композиция для иммунизации по любому из пп.4-10, где полипептидные фрагменты представляют собой синтетические пептиды, предпочтительно от 18 до 45 аминокислот в длину, необязательно перекрывающиеся или лигированные, необязательно содержащие дополнительные аминокислоты, иммуностимулирующие функциональные группы и/или защитные группы для усиления растворимости и увеличения стабильности in vivo. [0012] Композиция для иммунизации по любому из пп.4-11, дополнительно включающая СD40-связывающую молекулу, выбранную из антител или их фрагмента или лиганда CD40 или его варианта. [0013] Композиция для иммунизации по любому из пп.4-12, дополнительно включающая агонистические анти-4-1ВВ антитела или их фрагмент, способные стимулировать рецептор 4-1ВВ. [0014] Композиция для иммунизации по любому из пп.4-13, где композиция дополнительно включает антиген Mycobacterium, который не является специфичным для латентной стадии. [0015] Способ диагностики инфекции Mycobacterium у индивидуума, включающий стадии: [0016] Способ по п.15, где полипептиды выбраны из группы, включающей Rv1733c, Rv2029c, Rv2627c и Rv0080. [0017] Диагностический набор для осуществления способов диагностики по п.15 или 16, содержащий один или более полипептидов, и реагенты для анализа и количественного связывания антител с указанными полипептидами, где указанный один или более полипептид выбран из группы, состоящей из последовательностей ТВ регулона спячки (DosR):Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX), и аналогов, гомологов или их фрагментов; и необязательно включающая адъювант. Полный текст патента Область изобретения Настоящее изобретение относится к медицине, в частности к диагностике, профилактике и лечению заболеваний, вызываемых Mycobacterium, более конкретно инфекций, вызываемых Mycobacterium tuberculosis. Изобретение также относится к области вакцинации. Туберкулез (ТВ) представляет собой значительную угрозу здоровью человечества, по осторожным оценкам, от ТВ каждую минуту умирает четыре человека, что соответствует двум миллионам ежегодно. По оценкам, одна треть населения мира латентно инфицирована М. tuberculosis. Этот ужасающий по количеству источник скрытого туберкулеза, из которого происходит большинство случаев активного ТВ, заключает в себе основное препятствие для достижения контроля над ТВ. Существует несколько причин, почему латентные инфекции М. tuberculosis осложняют усилия по ликвидации ТВ. Во-первых, отслеживание распространения и излечивание при латентной инфекции достигаются только в условиях, когда большинство индивидуумов дают отрицательный результат в кожных тестах на туберкулин, что имеет место в промышленно развитых странах, где случаи ТВ уже редки. Даже в их условиях эффективность доступных в настоящее время режимов, применяемых для лечения скрытой инфекции М. tuberculosis, является ограниченной. Наряду с очевидными проблемами несоблюдения врачебных инструкций и преобладания штаммов, устойчивых к антибиотикам, это может быть связано с тем, что, как было обнаружено, находящиеся в состоянии покоя организмы М. tuberculosis являются либо умеренно, либо в высокой степени устойчивыми к широко используемым лекарственным средствам, таким как рифампин и изониазид, которые оказывают бактерицидное действие только в отношении реплицирующихся бацилл, что было показано in vitro (1, 2). Идея выживаемости нереплицирующихся М. tuberculosis в скрытом состоянии поддерживается открытием, что генотип М. tuberculosis слабо меняется в течение многих лет латентного состояния, тогда как скорость изменений в структуре ДНК значительно выше при активном заболевании (3, 4). Во-вторых, единственная используемая в настоящее время вакцина против ТВ, бациллы Кальмета-Герена (BCG) M. bovis, не предохраняет от возникновения латентной инфекции М. tuberculosis. BCG обеспечивает сильно вариабельный уровень защиты от реактивации ТВ, который различается в географических регионах и который, возможно, зависит от степени контакта с микобактериями в окружающей среде (5). Недавние попытки по разработке улучшенной вакцины были направлены на профилактические вакцины, предназначенные для введения до того, как произошло инфицирование М. tuberculosis, и которые оценивались в моделях на животных с острой первичной инфекцией. Такие кандидаты в профилактические вакцины были неэффективны или даже вредны, когда применялись в условиях после воздействия в моделях с животными, имитирующими хронические или латентные инфекции (6-8). В отличие от этого, вакцина для применения после воздействия, которую можно без риска вводить инфицированным в скрытой форме индивидуумам и которая препятствует реактивации ТВ, могла бы обладать непосредственным преимуществом, поскольку ее можно применять в высокой степени эндемических областях, где скрытая инфекция М. tuberculosis присутствует в большинстве популяций. Антигены, содержащиеся в таких вакцинах, должны усиливать защитный иммунный ответ, позволяющий распознавать и ликвидировать бациллы М. tuberculosis при латентной инфекции. CD4 Т-клетки играют важную роль в контроле и поддержании инфекции М. tuberculosis, хотя точный механизм этого и антигены-мишени, узнаваемые при скрытом ТВ, в значительной степени неизвестны (9). До недавнего времени было проведено несколько исследований по дифференциальной экспрессии генов и изменениям в метаболизме М. tuberculosis при латентности. Параллельно меньшее количество исследований было посвящено анализу специфических иммунных ответов организма-хозяина у человека, связанных с поддержанием латентности. До сих пор идентифицирован лишь один хорошо описанный белок М. tuberculosis, который по-видимому важен при латентном состоянии (10). Этот белок, HspX (Rv2031c или Acr), обладает существенно повышенным уровнем экспрессии при гипоксии, в условиях in vitro используется как заменитель стресса при воздействии факторов внешней среды, связанных с латентной инфекцией в гранулемах у человека (11). Клеточные иммунные ответы на этот белок теплового шока были обнаружены у латентно инфицированных индивидуумов, то есть связаны с защищенным состоянием, тогда как антитела к этому антигену были обнаружены у лиц с активной формой заболевания ТВ (12, 13). Идентификация дополнительных, связанных с латентностью антигенов является благоприятной для разработки эффективных вакцин для применения после воздействия. Несколько исследований были сфокусированы на устойчивом состоянии инфекции М. tuberculosis с использованием моделей in vitro и in vivo, которые были разработаны для имитации природного состояния латентности. Во-первых, Wayne et al. создали модель латентности in vitro на основе роста М. tuberculosis при постепенном уменьшении парциальных давлений кислорода, которое приводит к обратимой остановке роста бацилл, названной нерепликативным выживанием (NRP) (14). Другие авторы использовали постоянные гипоксические условия культивирования для изучения изменений метаболизма М. tuberculosis (11, 15, 16). Кроме того, Voskuil et al. исследовали профиль экспрессии М. tuberculosis при культивировании в присутствии низких доз оксида азота в качестве еще одного варианта условий in vitro, переживаемых бациллами при латентности и соответствующих появлению Thl иммунитета (19). При помощи микронабора ДНК всего генома Voskuil et al. обнаружили согласованное увеличение экспрессии в группе из 48 генов М. tuberculosis, которое наблюдалось во всех трех in vitro моделях латентности, а именно при NRP, постоянной гипоксии и воздействии малой дозы оксида азота (17). Было также обнаружено, что гены этого так называемого регулона спячки (DosR) обладают увеличенным уровнем экспрессии при выращивании M. tuberculosis в активированных макрофагах мышей in vitro (18). Кроме того, в легких хронически инфицированных мышей и больных ТВ профили транскрипции М. tuberculosis обладали теми же характеристиками, что и при NRP (19, 20). На этом основании было сделано предположение, что в ходе заболевания ТВ субпопуляция бацилл, наиболее вероятно, попадает в условия гипоксии и низкого уровня оксида азота и адаптируется к нерепликативному состоянию. Выяснилось, что белки, связанные с состоянием покоя, индуцируются в процессе перехода к скрытой инфекции и что во время латентности, весьма вероятно, туберкулезные бациллы экспрессируют белки, связанные с состоянием покоя. Функции большинства предполагаемых белков, кодируемых этим регулоном, неизвестны. В этом патентном описании авторы будут упоминать белки, кодируемые генами регулона спячки, как латентные антигены. Исходя из открытия генов (DosR) регулона спячки, использование кодируемых ими белков в качестве потенциальных антигенов для детекции и защитной или лечебной иммунизации и/или для вакцинации было рассмотрено в GB 01163856/US 2004/0241826. Более того, в WO 0179274 и US 2004/0057963 описаны способы и композиции для индуцирования иммунного ответа на латентные инфекции Mycobacterium tuberculosis при помощи полипептидов, которые индуцируются именно при латентном состоянии микобактериальных инфекций. Полипептиды выбирали из совокупности, состоящей из 45 генов регулона спячки, которые обладают повышенным уровнем экспрессии при латентности в вышеупомянутых in vitro моделях. Одним из антигенов, выбранным для иммунизации, является HspX (Rv2031c или Acr), кодирующий гомолог α-кристаллина и который может использоваться для диагностики и иммунизации, как описано в US 2004/0146933. В настоящее время неизвестно, действительно ли белки, кодируемые регулоном NRP/спячки (DosR), экспрессируются у М. tuberculosis на достаточно высоком уровне при латентной фазе инфекции у человека для индуцирования значительного иммунного ответа, так как в прототипе для латентности использовались in vitro модели и модели с использованием мышей. Также неизвестно, достаточно ли иммуногенны предполагаемые антигены регулона латентности или спячки, и действительно ли иммунитет к этим гипотетическим антигенам и/или эпитопам латентности имеет отношение к обеспечению защиты от скрытой или вновь приобретенной инфекции Mycobacterium у млекопитающих. В настоящее время, в частности, неизвестно, какие из 48 антигенов, кодируемых регулоном спячки/латентности, могут иметь наибольшее значение во время фактических латентных инфекций Mycobacterium tuberculosis у человека. Невозможно, или нежелательно, или неэффективно комбинировать, например, в фармацевтической композиции или вакцине все 48 предполагаемых латентных антигенов для того, чтобы вызвать иммунный ответ у индивидуумов с латентной инфекцией Mycobacterium. Большинство идентифицированных предполагаемых латентных антигенов не будут эффективны для индукций иммунного ответа, так как они не экспрессируются на достаточном уровне и не содержат адекватные (доминантные) эпитопы для цитотоксических Т-клеток (CTL) или Т-хелперов (Th), которые могут распознаваться иммунной системой латентно инфицированных млекопитающих. Задачей, которая поставлена для решения в настоящем изобретении, и является обеспечение оптимального выбора из 48 известных предполагаемых латентных антигенов и отбор только тех антигенов, которые действительно способны вызывать иммунные ответы in vivo у здоровых индивиуумов с латентной инфекцией Mycobacterium. Настоящее изобретение решает обсуждаемые выше вопросы путем выявления ex vivo доминантных иммунных ответов у людей в отношении связанных с латентным состоянием Mycobacterium антигенов и/или эпитопов in vivo и, тем самым, обеспечивая новые способы и композиции для детекции и иммунизации против латентной инфекции Mycobacterium. Указанные композиции включают только те латентные антигены, которые действительно способны вызывать иммунный ответ in vivo у млекопитающих, подвергнутых скрытой инфекции Mycobacterium, и, более предпочтительно, включают только те антигены, которые предпочтительно узнаются у латентно инфицированных индивидуумов и которые совсем не узнаются или узнаются в значительно меньшей степени у лиц с активной инфекцией Mycobacterium или у лиц с индуцированными Mycobacterium заболеваниями или симптомами, такими же, как у больных, страдающих от туберкулеза (ТВ). Изобретение достигает этой цели за счет выявления небольшой подгруппы доминантных антигенов и/или эпитопов из группы по меньшей мере 48 латентных антигенов М. tuberculosis, известных в данной области. Неожиданно было обнаружено, что наиболее предпочтительные антигены, выявленные в настоящем изобретении, отличаются от тех предполагаемых латентных антигенов, которые были наиболее изучены и применялись до сих пор в прототипе; главным образом Rv2031c (HspX/acr) и Rv0569. Настоящее изобретение отличается от предпочтительных предполагаемых латентных антигенов, отобранных и применяемых в публикациях, патентах и патентных заявках предшествующего уровня техники, что обозначает, что небольшая группа, рассматриваемая в настоящем изобретении, далека от известных примеров. Определенная и небольшая подгруппа латентных антигенов, представленных в описании, представляет собой целенаправленную выборку из известной группы предполагаемых латентных антигенов. По этому изобретению антигены и/или эпитопы были выбраны после обширного анализа группы предполагаемых антигенов. В то время как предполагаемые антигены в предшествующем уровне техники были выбраны на основании моделей in vitro в лабораторных условиях и на моделях с использованием мышей, в настоящем изобретении раскрывается, которые из этих антигенов действительно способны дифференцированно вызывать иммунный ответ in vivo в нормальных условиях при латентной инфекции Mycobacterium и не вызывать или вызывать в значительно меньшей степени у здоровых индивидуумов или больных, страдающих заболеванием ТВ. Настоящее изобретение относится к способам и композициям для индуцирования иммунного ответа на инфекции Mycobacterium у позвоночных, предпочтительно млекопитающих, в частности на латентные инфекции Mycobacterium; способу, включающему стадию введения позвоночному композиции, содержащей источник одного или более полипептидов или их фрагментов, выбранных из группы полипептидов, содержащей белки, кодируемые регулоном NRP/спячки (DosR) Mycobacterium, которые способны вызывать иммунный ответ in vivo у позвоночных, инфицированных Mycobacterium. Используемый в описании термин инфекция Mycobacterium означает включение латентно инфицированных млекопитающих, вновь инфицированных млекопитающих с еще непроявленными симптомами и позвоночных, страдающих от индуцированных Mycobacterium заболеванием и симптомами, такими как активная форма туберкулеза. Предпочтительно инфекция Mycobacterium по изобретению представляет собой латентную инфекцию, на которую следует направить лечение для предотвращения развития туберкулеза. Способ по изобретению также может эффективно применяться в качестве дополнительного лечебного мероприятия во время или после лечения антибиотиками больных ТВ, ТВ с или без (множественной) лекарственной устойчивости; и в отношении здоровых, но подвергавшихся воздействию лиц, предпочтительно, но не исключительно, детей эндемичных по ТВ стран, которые предварительно были вакцинированы BCG. Изобретение относится к способам и композициям, которые могут быть эффективны в отношении латентных инфекций Mycobacterium, но также могут быть легко скомбинированы специалистом с полипептидами или композициями, содержащими эпитопы, предназначенные для индукции иммунного ответа на нелатентные инфекции, такими как профилактические вакцины и/или многостадийные вакцины против микобактерий. К латентной инфекции Mycobacterium, как понимается в описании, относится стадия инфекции, при которой бациллы остаются живыми, однако, медленно реплицируются или выживают в нереплицируемом состоянии и могут быть сосредоточены в локальном очаге внутри органа или ткани, не вызывая активное некротическое заболевание, которое, как правило, наблюдается при ТВ. Латентная стадия может существовать на протяжении всей жизни хозяина, или инфекция может повторно активизироваться во время, например, периода снижения иммунитета хозяина или в ответ на другие стресс-факторы, такие как другие (мико)бактериальные или вирусные инфекции типа ВИЧ или лечение рака и другие иммуносупрессирующие условия или лечения. Изобретение относится к способам и композициям, которые подходят для применения в качестве 1) превентивных (профилактических), 2) применяемых после воздействия инфекционного агента/инфекции или 3) терапевтических/целебных вакцин против латентной инфекции Mycobacterium и родственных заболеваний, таких как, в качестве неограничивающих примеров, инфекции (ослабленных и/или рекомбинантных) Mycobacterium tuberculosis, M. bovis (включая бациллы Кальмета-Герена (BCG), М. africanum, M. smegmatis, М. leprae, М. vaccae, M. intracellulare, М. avium (включая подвид paratuberculosis), М. canettii, М. leprae, M. microti и М. ulcerans. М. tuberculosis, M. bovis (включая штаммы BCG), М. microti, M. africanum и М. canettii (т.е. виды Mycobacterium и штаммы, относящиеся к ТВ комплексу) являются наиболее предпочтительными источниками полипептидов или их фрагментов, индуцируемых в состоянии латентности, для использования по изобретению. Для способов и композиций по изобретению позвоночное, для которого предназначено лечение или диагностика, представляет собой человека, но также включает всех лабораторных и сельскохозяйственных животных, таких как, в качестве неограничивающих примеров, мыши, крысы, морские свинки, кролики, кошки, собаки, овцы, козы, коровы, лошади, верблюды, и домашняя птица, например цыплята, утки, индейка и гусь. Источником полипептида может быть белок, перевар белка и/или его фрагменты, которые могут находиться в очищенном виде или в составе неочищенной композиции, предпочтительно биологического происхождения, такой как бактериальные лизаты, разрушенные ультразвуком или фиксированные клетки. Альтернативно, (поли)пептид может быть химически синтезирован или получен ферментативно in vitro. Источником полипептида или его фрагмента может быть также нуклеиновая кислота, кодирующая полипептид или его фрагмент, с РНК или ДНК в качестве матрицы. Молекулы РНК или ДНК могут быть «голой » ДНК, предпочтительно включенной в везикулы или липосомы, или могут быть в составе вектора. Вектором может быть любой (рекомбинантный) ДНК или РНК вектор, известный в данной области, который предпочтительно представляет собой плазмиду, где гены, кодирующие латентные антигены, функционально связаны с регуляторной последовательностью, направляющей экспрессию и трансляцию кодирующих мессенджеров. Вектором также может быть любой ДНК- или РНК-содержащий вирус, такой как, в качестве неограничивающих примеров, аденовирус, аденоассоциированный вирус (AAV), ретровирус, лентивирус, модифицированная осповакцина (MVA) или вирус оспы птиц, или любой другой вирусный вектор, способный направлять экспрессию полипептидов, содержащих эпитопы латентности, у хозяина. ДНК-содержащие векторы могут быть неинтегрируемыми, такими как эписомные репликативные векторы, или могут быть векторами, интегрируемыми в геном хозяина в результате случайной интеграции или гомологичной рекомбинации. Молекулы ДНК, содержащие гены, кодирующие полипептиды или их фрагменты, по настоящему изобретению, необязательно встроены в векторы, такие как вирусы или плазмиды, могут быть интегрированы в геном хозяина. В предпочтительном варианте осуществления изобретения таким хозяином может быть микроорганизм. Предпочтительно таким рекомбинантным микроорганизмом является Mycobacterium, например виды М. tuberculosis или М. bovis, и наиболее предпочтительно М. bovis бациллы Кальмета-Герена (BCG), способные передавать хозяину полипептиды или их фрагменты по изобретению. Рекомбинантные BCG и способы рекомбинации известны в данной области, например, из WO 2004094469. Такой рекомбинантный микроорганизм может быть получен в виде живой рекомбинантной и/или живой ослабленной вакцины, как описано, например, в Jacobs et al. 1987, Nature, 327(6122):532-5. Вектор может также содержаться в хозяине бактериального происхождения, таком как, в качестве неограничивающих примеров, живые ослабленные и/или рекомбинантные бактерии шигеллы или сальмонеллы. В одном из вариантов осуществления настоящее изобретение относится к способу индуцирования иммунного ответа на инфекции Mycobacterium у млекопитающих, способу, включающему стадию введения млекопитающему источника одного или более полипептидов или их фрагментов, выбранных из группы полипептидов, содержащей белки, кодируемые регулоном NRP/спячки (DosR) Mycobacterium, которые способны вызывать IFN- γ ответ в Т-линии клеток человека, состоящей из Rv079, Rv0569, Rv0572c, Rv1733c, Rv1738, Rv1813c, Rv1996, Rv2007c (FdxA), Rv2029c (PfkB), Rv2030c, Rv2031c (HspX, Acr, 16кДа гомолог α-кристаллина), Rv2032, Rv2623, Rv2624c, Rv2626c, Rv2627c, Rv2628, Rv3126c, Rv3127, Rv3129, Rv3130c, Rv3131, Rv3132c, Rv3133c (DosR), Rv3134c, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX) и их аналоги или гомологи, и необязательно один или более адъювантов. Указанные антигены узнаются кратковременными Т-линиями клеток от инфицированных индивидуумов и приведенных в контакт с разрушенными ультразвуком М. tuberculosis. T-линия клеток показывает интерферон- γ (IFN- γ) ответ предпочтительно не менее > 50 пг IFN- γ/мл в тестах, описанных в примерах 1 и 2. Номенклатура Rv для микобактериальных антигенов и последовательности ДНК и белков регулона NRP/спячки (DosR) хорошо известна в данной области и может быть найдена, например, на http://genolist.pasteur.fr/TubercuList/ или на http://www.ncbi.nlm.nih.gov/entrez (Accession number AL123456). Номенклатура Rv, как используют в описании, может относиться либо к аминокислотной последовательности антигена, либо к нуклеотидной последовательности, кодирующей антиген. В предпочтительном варианте осуществления изобретения способ индукции иммунного ответа на инфекцию Mycobacterium у позвоночного включает введение источника полипептидов или их фрагментов, которые выбраны из группы последовательностей регулона NRP/спячки (DosR) Mycobacterium, который содержит латентные антигены, способные вызывать иммунный ответ у позвоночных, обладающих латентной микобактериальной инфекцией, состоящей из Rv0079, Rv0569, Rv1733c, Rv1738, Rv1813c, Rv1996, Rv2007c (FdxA), Rv2029c (PfkB), Rv2030c, Rv2031c (HspX, Acr, 16кДа гомолог α-кристаллина), Rv2032, Rv2626c, Rv2627c, Rv2628, RV3126C, Rv3129, Rv3130c, Rv3132c, Rv3133c (DosR), Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX) и их аналоги или гомологи. Эта специфическая подгруппа латентных антигенов способна индуцировать интерферон- γ (IFN- γ) ответ у моноцитов периферической крови (РВМС), полученных от индивидуумов с латентной микобактериальной инфекцией, с более чем 100 пг IFN- γ/мл и у не менее 5, 10, 20, 30, 40 или 50% всех инфицированных Mycobacterium индивидуумов. В наиболее предпочтительном варианте осуществления изобретение относится к способу индукции иммунного ответа у позвоночного, предпочтительно у индивидуума, страдающего от или подверженного риску приобретения латентной инфекции Mycobacterium, включающему введение источника полипептидов или их фрагментов, которые выбраны из группы последовательностей регулона NRP/спячки (DosR) Mycobacterium, которые способны предпочтительно вызывать иммунный ответ у индивидуумов, зараженных латентной инфекцией Mycobacterium, состоящей из антигенов Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX). Указанные восемь антигенов включают доминантные эпитопы, которые предпочтительно узнаются у латентно инфицированных индивидуумов и которые не способны или в значительно меньшей степени способны индуцировать IFN- γ ответ у неинфицированных индивидуумов или у индивидуумов с симптомами заболевания активной формой инфекции Mycobacterium. Указанные антигены индуцировали наиболее высокий уровень IFN- γ у мононуклеаров периферической крови (РВМС) среди 48 исследованных латентных антигенов. Кроме того, эти восемь антигенов способны индуцировать значительную продукцию IL-10 в РВМС от латентно инфицированных индивидуумов, но не в РВМС от больных с симптомами, связанными с активной формой инфекции Mycobacterium tuberculosis. Три наиболее предпочтительных полипептида для использования в способе по изобретению представляют собой последовательности регулона NRP/спячки (DosR) Mycobacterium Rv1733c и Rv2029c (PfkB) и Rv2627c. Из всех 48 исследованных полипептидов Rv1733c и Rv2029c (PfkB) и Rv0080 наиболее часто отмечались как вызывающие иммунные ответы у индивидуумов с латентной инфекцией Mycobacterium, что определялось по индукции IFN- γ и/или IL-10 в РВМС от этих индивидуумов. В другом аспекте изобретение относится к композициям, содержащим источник полипептидов или их фрагментов последовательностей регулона NRP/спячки (DosR) Mycobacterium, которые способны вызывать иммунный ответ у индивидуумов с латентной микобактериальной инфекцией. Предпочтительно композиция для иммунизации против инфекции Mycobacterium и индуцированных заболеваний по изобретению включает источник одного или более полипептидов или их фрагментов, выбранных из группы полипептидов, содержащей белки, кодируемые регулоном NRP/(DosR) Mycobacterium, способные вызывать IFN- γ ответ у Т-линии клеток человека, состоящей из Rv0079, Rv0569, Rv0572c, Rv1733c, Rv1738, Rv1813c, Rv1996, Rv2007c (FdxA), Rv2029c (PfkB), Rv2030c, Rv2031c (HspX, Acr, 16кДа гомолог α-кристаллина), Rv2032, Rv2623, Rv2624c, Rv2626c, Rv2627c, Rv2628, Rv3126c, Rv3127, Rv3129, Rv3130c, Rv3131, Rv3132c, Rv3133c (DosR), Rv3134c, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX) и их аналоги или гомологи, и необязательно содержит по меньшей мере один адъювант. Гомолог или аналог, как используется в описании, включает пептид, который по меньшей мере на 70, 80, 90, 95, 98 или 99% идентичен аминокислотной последовательности с отмеченными выше полипептидами, кодируемыми нативным регулоном NRP/спячки (DosR) М. Tuberculosis, и сохраняет способность индуцировать иммунный ответ, который не слабее иммунного ответа, вызываемого полипептидом М. tuberculosis. Гомолог или аналог может содержать замены, инсерции, делеции и дополнительные N- или С-концевые аминокислоты или химические части для повышения стабильности, растворимости и иммуногенности. Под фрагментом полипептидных антигенов по изобретению понимают фрагмент, содержащий, по меньшей мере, эпитоп. Фрагмент, таким образом, включает не менее 4, 5, 6, 7 или 8 соседних аминокислот последовательности полипептидного антигена. Более предпочтительно фрагмент включает, по меньшей мере, эпитоп Т-клеток, т.е. не менее 8, 9, 10, 11, 12, 13 или 14 соседних аминокислот последовательности полипептидного антигена. Еще более предпочтительно фрагмент содержит как CTL эпитоп, так и эпитоп для Т-хелперов. Однако наиболее предпочтительно фрагмент представляет собой пептид, для которого необходим процессинг антиген-представляющей клеткой, т.е. фрагмент имеет длину не менее 18 аминокислот, и эти 18 аминокислот необязательно являются смежной последовательностью из полипептидного антигена. Более предпочтительно композиция по изобретению содержит источник полипептидов или их фрагментов, которые выбраны из группы последовательностей регулона NRP/спячки (DosR) Mycobacterium, содержащего латентные антигены, способные к иммунному ответу у латентно инфицированных Mycobacterium индивидуумов, состоящей из Rv0079, Rv0569, Rv1733c, Rv1738, Rv1813c, Rv1996, Rv2007c (FdxA), Rv2029c (PfkB), Rv2030c, Rv2031c (HspX, Acr, 16кДа гомолог α-кристаллина), Rv2032, RV2626C, Rv2627c, Rv2628, Rv3126c, Rv3129, Rv3130c, Rv3132c, Rv3133c (DosR), Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX). Указанные антигены и композиции способны индуцировать IFN- γ ответ у мононуклеаров периферической крови (РВМС) от индивидуумов, инфицированных Mycobacterium. В более предпочтительном варианте осуществления композиция по изобретению содержит источник последовательностей регулона NRP/спячки (DosR) Mycobacterium, которые способны предпочтительно на индукции иммунного ответа у индивидуумов, латентно инфицированных Mycobacterium, при этом антигены выбираются из одного или более из Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarK). Полипептиды Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX) способны индуцировать наиболее сильный ответ у индивидуумов с латентной инфекцией в терминах - продукции IFN- γ в РВМС по сравнению с другими 48 проверенными полипептидами регулона NRP/спячки (DosR) Mycobacterium. Указанные антигены также способны стимулировать продукцию IL-10 в РВМС от латентно инфицированных больных, в то время как индукция IL-10 не наблюдалась или наблюдалась в значительно меньшей степени в РВМС от больных, имеющих активную форму инфекции Mycobacterium и/или симптомы заболевания ТВ. В другом варианте осуществления композиция по изобретению включает, по меньшей мере, источник полипептида или его фрагментов, полученных из последовательности регулона NRP/спячки (DosR) Mycobacterium Rv1733c, и/или Rv2029c, и/или Rv2627c, которые являются наиболее часто распознаваемыми антигенами у латентно инфицированных индивидуумов из всех исследованных полипептидов, кодируемых регулоном NRP/спячки (DosR). Композиция для иммунизации по изобретению, содержащая кодируемые регулоном NRP/спячки (DosR) полипептиды или их фрагменты, предпочтительно содержит по меньшей мере один наполнитель. Наполнители хорошо известны в данной области фармации и приведены в руководствах, таких как Remmington's pharmaceutical sciences, Mack Publishing, 1995. Композиция для иммунизации по изобретению может предпочтительно содержать по меньшей мере один адъювант. Адъюванты могут включать любой адъювант, известный в области вакцинации, и могут быть выбраны из руководств, например Current Protocols in Immunology, Wiley Interscience, 2004. Наиболее предпочтительно адъюванты выбраны из следующего перечня адъювантов: катионные (противомикробные) пептиды и лиганды Toll-подобного рецептора (TLR), такие как, в качестве неограничивающих примеров, поли (I:С), CpG мотивы, LPS, липид А, липопептид Pam3Cys и бактериальные флагеллины или их части, и их производные, имеющие химические модификации. Другими предпочтительными адъювантами для использования в способе и в композиции по изобретению являются смеси из живых или убитых BCG, иммуноглобулиновые комплексы с указанными антигенами латентности или их частями, IC31 (с www.intercell.com; в WO03047602), QS21/MPL (US2003095974), DDA/MPL (WO2005004911), DA/TDB (WO2005004911; Holten-Andersen et al., 2004 Infect Immun. 2004 Mar; 72(3): 1608-17.) и растворимый LAG3 (CD223) (с www.Immunotep.com; US2002192195). Способ и композиция для иммунизации по настоящему изобретению могут также включать использование и/или добавление связывающей CD40 молекулы для усиления CTL ответа и тем самым усиливать терапевтические эффекты способов и композиций по изобретению. Использование связывающих CD40 молекул описано в WO 99/61065, который приводится в настоящем документе в качестве ссылки. CD40 связывающая молекула предпочтительно представляет собой антитела или их фрагмент или CD40 лиганд или их вариант и может добавляться отдельно или может быть включен в композицию по настоящему изобретению. Способ и композиция для иммунизации по настоящему изобретению может также включать использование и/или добавление агонистических анти-4-1ВВ антител или их фрагмента, или другой молекулы, способной к взаимодействию с рецептором 4-1ВВ. Использование 4-1ВВ-рецептора агонистических антител и молекул описано в WO 03/084999, который приводится в описании в качестве ссылки. 4-1ВВ агонистические антитела могут использоваться с или без добавления CD40 связывающих молекул, для усиления CTL иммунитета посредством переключения/стимулирования рецепторов 4-1ВВ и/или CD40. 4-1ВВ связывающая молекула или антитела могут добавляться отдельно или могут быть включены в композицию по настоящему изобретению. Полипептиды по изобретению в целях иммунизации могут быть слиты с белками, такими как, в качестве неограничивающих примеров, столбнячный токсин/токсоид, дифтерийный токсин/токсоид или другими молекулами-носителями. Полипептиды по изобретению можно также успешно слить с белками теплового шока, такими как рекомбинантный эндогенный gp96 (GRP94) (мыши) в качестве носителя иммунодоминантных пептидов, как описано в (ссылки Rapp UK and Kaufmann SH, Int. Immunol. 20 04 Apr.; 16(4): 597-605; Zugel U., Infect Immun. 2001 Jun.; 69(6): 4164-7), или слитых белков с Hsp70 (Triebel et al.; WO 9954464). Способ и композиция для иммунизации по изобретению предпочтительно включают использование полипептидных фрагментов, полученных из полипептидов по изобретению, содержащих доминантные CTL или Th эпитопы, и такие пептиды имеют длину от 18 до 45 аминокислот. Наличие CTL или Th эпитопов в последовательности может быть обнаружено специалистом с использованием общеизвестных биоинформационных программ, таких как HLA_BIND, SYFPEITHI, NetMHC и TEPITOPE 2000 (ссылки 43, 44, 45, 46, 47 и 48), или опытным путем при помощи стандартных экспериментов (Current Protocols in Immunology, Wiley Interscience 2004). Было отмечено, что пептиды по изобретению с длиной от 18 до 45 аминокислот обеспечивают превосходные иммуногенные свойства, как описано в WO 02/070006. Пептиды могут быть легко синтезированы химически и могут необязательно (частично) перекрываться и/или также могут быть связаны с другими молекулами, такими как TLR лиганды, пептиды или белки. Пептиды могут быть слиты с образованием синтетических белков, как описано в PCT/NL 03/00929 и в Welters et al. (Vaccine. 2004 Dec. 2; 23(3): 305-11). Также можно добавлять на N- или С-конец пептида химические функциональные группы или дополнительные (модифицированные или D-) аминокислоты для повышения стабильности и/или уменьшения биодеградации пептида. Для увеличения иммуногенности/иммуностимуляции могут присоединяться функциональные группы, например липидация. Для увеличения растворимости пептида и увеличения стабильности можно in vivo использовать присоединение заряженных или полярных аминокислот. В еще одном варианте осуществления композиция для индукции иммунного ответа или иммунизации по изобретению дополнительно содержит антигены Mycobacterium, неспецифичные для латентного состояния. Такие антигены с успехом могут быть высокоспецифичными для других стадий инфекционного процесса. Для целей иммунизации может быть эффективно предоставление композиций, которые не только нацеливают иммунный ответ исключительно на латентные инфекции Mycobacterium, но которые способны на иммунный ответ, направленный на симптомы заболевания инфицированного млекопитающего, вызванные активными инфекциями Mycobacterium. В таких способах и для таких композиций эффективно комбинировать защитный иммунитет в отношении Mycobacteria при различных фазах инфекционного процесса и тем самым обеспечивать лучшую защиту в целом. Композиции по изобретению, содержащие специфические латентные антигены, которые способны вызывать иммунный ответ, таким образом, можно комбинировать с микобактериальными антигенами, для которых известно, что они вызывают иммунный ответ на активные инфекции, такими как, в качестве неограничивающих примеров, антигены М. tuberculosis ESAT-6 (Rv3875), Ag85A (FbpA/MPT59, Rv3804c), Ag85B (Rv1886c), Ag85C (Rv3803c), CFP10 (Rv3874), TB10.3 (Rv3019c), TB10.4 (Rv0288), MPT64 (Rv1980c), MPT32 (RV1860) и МРТ57 (Rv3418c). В другом варианте осуществления специфические латентные антигены и композиции по изобретению применяются в способах и реагентах для диагностики латентных инфекций Mycobacterium. Способ диагностики латентной или устойчивой инфекции Mycobacterium, в частности латентной инфекции у индивидуума по изобретению, включает стадии: a) контактирования образца жидкости организма и/или клеток (в частности, лейкоцитов) индивидуума, необязательно выделенных, с одним или более полипептидов или их фрагментов, выбранных из группы последовательностей регулона NRP/спячки (DosR) Mycobacterium, состоящей из Rv0079, Rv0569, Rv1733c, Rv1738, Rv1813c, Rv1996, Rv2007c (FdxA), Rv2029c (PfkB), Rv2030c, Rv2031c (HspX, Acr, 16 кДа гомолог α-кристаллина), Rv2032, Rv2626c, Rv2627c, Rv2628, Rv3126c, Rv3129, Rv3130c, Rv3132c, Rv3133c (DosR), Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX); и b) детектирования иммунного ответа на указанные полипептиды, определяемого по пролиферации или продукции цитокинов, что является индикатором инфекции Mycobacterium. Способ диагностики наиболее предпочтительно включает не менее одного или более полипептидов Rv1733c, Rv2029c (PfkB), Rv2627c, RV2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX). В другом предпочтительном варианте осуществления способ диагностики включает детекцию иммунного ответа в отношении полипептидов Rv1733c, Rv2029c (PfkB), Rv2627c и Rv0080 или их фрагментов, которые представляют собой наиболее часто обнаруживаемые антигены при латентной инфекции Mycobacterium из всех проверенных здесь латентных антигенов. Жидкость организма в данном случае включает мочу, слюну, сперму, слезы, лимфатические жидкости и наиболее предпочтительно кровь, включая эритроциты. Из образцов крови можно получить клетки РВМС и выращивать их с использованием общепринятых методик. Для увеличения чувствительности и специфичности способа диагностики в значительной степени предпочтительно совместное детектирование некоторых специфических латентных антигенов, и в конкретном предпочтительном варианте осуществления способ включает обнаружение иммунного ответа в отношении антигенов Rv1733c, Rv2029c (PfkB), Rv2627c и Rv0080. Способ также может быть объединен с детектированием других известных в данной области антигенов, которые не являются специфичными для латентной фазы инфекции Mycobacterium, например специфические для активной фазы инфекционного процесса. В случае М. tuberculosis такие антигены могут включать, в качестве неограничивающих примеров, ESAT-6 (Rv3875), Ag85A (FbpA/MPT59, Rv3804c), Ag85B (Rv1886c), Ад85С (Rv3803c), CFP10 (Rv3874), TB10.3 (Rv3019c), TB10.4 (Rv0288), MPT64 (Rv1980c), MPT32 (Rv1860) и МРТ57 (Rv3418c). Изобретение также относится к диагностическому набору для проведения описанного выше способа диагностики, набор включает один или более полипептидов или их фрагментов по изобретению и необязательно включает реагенты для анализа и количественного связывания антител с указанными полипептидами или для определения клеточных иммунных ответов. Такие реагенты могут предпочтительно содержать реагенты, необходимые для определения связывания антигена, такие как, в качестве неограничивающих примеров, ELISA или объединенные анализы СВА, или реагенты для определения IFN- γ и/или IL-10 ответа, например те, которые использовались в предоставленных здесь примерах. Полипептиды или их фрагменты для определения микобактериальных инфекций можно присоединить к твердофазному носителю, такому как плашки для белков/пептидных (микро)анализов или для микротитрования в лунках. Изобретение также охватывает предпочтительные фрагменты специфического антигена латентности и/или эпитопы, включая полипептиды Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX). Эти пептиды предпочтительно используются в способах и композициях для индукций иммунного ответа и для диагностических целей по изобретению и предпочтительно имеют длину от 18 до 45 аминокислот и включают или состоят из следующих последовательностей: VDEPAPPARAIADAALAALG (SEQ ID NO: 1) или одного из В- и Т-клеточного эпитопов, идентифицированных и описанных в описании, на фиг. 13, 14, 15 и 16. Определения. Идентичность аминокислотной последовательности означает, что две (поли)пептидные последовательности при оптимальном наложении, например, при помощи программ GAP или BESTFIT с используемыми по умолчанию параметрами, имеют не менее определенного процента одинаковой последовательности, что определено в описании в другом месте. GAP использует универсальный алгоритм совмещения Needleman и Wunsch для совмещения двух последовательностей по всей их длине, максимально увеличивая число пар и минимизируя количество пропусков. Как правило, GAP по умолчанию использует параметры со штрафом за создание пропуска = 8 и штрафом за продолжение пропуска = 2. Для белков заданной по умолчанию матрицей счета является Blosum62 (Henikoff & Henikoff, 1992, PNAS 89, 915-919). Совмещение последовательностей и оценка процента идентичности последовательностей могут быть определены при помоши компьютерной программы, такой как GCG Wisconsin Package, Version 10.3, available from Accelrys Inc., 9685 Scranton Road, San Diego, CA 92121-3752, USA. Альтернативно процент сходства или идентичности можно определить по поиску в базах данных, таких как FASTA, BLAST и др. Регулоны у эукариотов представляют собой генетические единицы, состоящие из несмежных групп генов, контролируемых одним регуляторным геном. У бактерий регулоны представляют собой универсальные регуляторные системы, вовлеченные во взаимодействие регуляторных доменов, и могут состоять из одного или нескольких оперонов. Регулон NRP/спячки (DosR) M. tuberculosis находится под контролем транскрипционного регулятора DosR - (Rv3133c) и включает по меньшей мере 48 последовательностей, описанных в Voskuil et al. (J. Exp. Med. 2003, 198(5): 705-13), перечисленных в табл. 2. Термин «индивидуум », как применяют в описании, относится к живым многоклеточным позвоночным организмам, понятие включает как людей, так и не относящихся к человеку млекопитающих. Термин «индивидуум » включает как людей, так и ветеринарных и лабораторных индивидуумов. Антиген в данном случае представляет собой свойство молекулы или ее фрагмента, означающее способность индуцировать иммунный ответ у млекопитающих. Термин включает иммуногены и участки, ответственные за антигенность или антигенные детерминанты или эпитопы. Антиген представляет собой химическую или биохимическую структуру, детерминант, антиген или их часть, способные индуцировать формирование клеточного (Т-cell) или гуморального (антитела) иммунных ответов. Иммунный ответ in vivo или in vitro может быть определен и/или проконтролирован при помощи одного из способов, предоставленных в данной спецификации, но также при помощи других способов, известных и очевидных специалисту и с которыми можно ознакомиться в Current Protocols in Immunology, Wiley Interscience 2004. Клеточный иммунный ответ может быть определен по индукции высвобождения соответствующих цитокинов, таких как IFN- γ или IL-10, из (или по индукции пролиферации) лимфоцитов, взятых у млекопитающего, вновь или предварительно инфицированного (опасными) микобактериями или иммунизированного в качестве неограничивающего примера полипептидом(ами). Для контроля клеточной пролиферации клетки могут быть импульсно мечены радиоактивным тимидином или подсчитаны в (проточном) цитометре или под микроскопом. Индукцию цитокинов можно контролировать различными иммунохимическими способами, такими как, в качестве неограничивающих примеров, тесты ELISA или Elispot. Клеточный ответ in vitro также можно определить с использованием линий Т-клеток, полученных от здорового индивидуума или инфицированного Mycobacterium млекопитающего, где линия Т-клеток отобрана при помощи живых и/или убитых, ослабленных или рекомбинантных Mycobacteria, латентных Mycobacteria или выделенных или полученных от них выбранных антигенов. Термины «вакцина » или «иммуногенная композиция » используются в настоящем документе для описания композиции, пригодной для индукций специфического иммунного ответа у млекопитающего, необязательно включающей адъюванты и другие активные компоненты для усиления или управления определенным типом иммунного ответа, предпочтительно CTL или Th ответа. Специфический для латентного состояния полипептид или антиген экспрессируется на высоких уровнях (или исключительно) у Mycobacterium в скрытой или стационарной, но не в активной или логарифмической фазе роста, и для М. tuberculosis может кодироваться регулоном NRP/спячки (DosR) (Voskuil et al., 2003). Под TST положительным индивидуумом понимается включение млекопитающего с положительным тестом Манту (отвердение > 5 мм) или индивидуума, у которого очищенное белковое производное (PPD) (=туберкулин) индуцирует положительный ответ отзыва in vitro, определенный по высвобождению IFN- γ и, следовательно, обладающего латентной микобактериальной инфекцией, т.е. индивидуума, который был инфицирован (вирулентными) Mycobacterium, например М. tuberculosis, но не проявляет никаких признаков (активного) заболевания, такого как туберкулез (ТВ). Под больным ТВ понимается индивидуум, инфицированный (вирулентными) микобактериями, что подтверждено высевами и микроскопически, и/или индивидуум с клиническим диагнозом ТВ и который быстро реагирует на противотуберкулезную химиотерапию. Посевы, микроскопия и клинический диагноз ТВ хорошо известны любому практикующему врачу-специалисту. Последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, когда последовательность первой нуклеиновой кислоты устанавливает функциональное взаимоотношение со второй последовательностью нуклеиновой кислоты. Например, промотор функционально связан с кодирующей последовательностью, если промотор оказывает влияние на транскрипцию или экспрессию кодирующей последовательности. Как правило, функционально связанные последовательности ДНК являются смежными и, где необходимо соединить две белок-кодирующие области, находятся в одной рамке считывания. Вектор, как используют в описании, относится к молекуле нуклеиновой кислоты, которая при введении в клетку-хозяин осуществляет ее трансформацию. Вектор может включать последовательность нуклеиновой кислоты, которая позволяет ему реплицироваться в клетке-хозяин, такую как точка начала репликации. Вектором может быть, например, плазмида, фагмида, фаг, космида, вирус, ретровирус, эписома или мобильный генетический элемент. Вектор может также включать один или несколько селективных (устойчивость к антибиотику) или визуальных (например, GFP, иммунная метка) маркерных генов и других генетических элементов, известных в данной области. Белки, пептиды и полипептиды представляют собой линейные полимерные цепочки аминокислот (как правило, L-аминокислот), чьи α-углеродные атомы связаны посредством пептидных связей, образованных в результате реакции конденсации между карбоксильной группой α-углеродного атома одной аминокислоты и аминогруппой α-углеродного атома другой аминокислоты. Концевая аминокислота на одном конце цепи (т.е. аминотерминальная) содержит свободную аминогруппу, тогда как концевая аминокислота на другом конце цепи (т.е. карбокситерминальная) содержит свободную карбоксильную группу. По существу, термин «аминоконец (N-конец) » относится к свободной α-аминогруппе аминокислоты на аминотерминальном конце пептида или к α-иминогруппе (иминогруппа, когда принимает участие в пептидной связи) аминокислоты в любом другом положении в пептиде. Термин «карбоксиконец (С-конец) » относится к свободной карбоксильной группе аминокислоты на карбокситерминальном конце пептида или к карбоксильной группе аминокислоты в любом другом положении в пептиде. Синтетический полипептид относится к полипептиду, образованному in vitro, путем присоединения аминокислот в определенном порядке, с использованием средств органической химии для образования пептидных связей. Как правило, составляющие пептид аминокислоты пронумерованы в порядке, начиная от N-конца и увеличиваясь в направлении к С-концу пептида. Пояснения к чертежам. Фиг. 1. Число долговременно культивируемых линий Т-клеток (n=12), реагирующих на латентные антигены М. tuberculosis, гипоксические лизат М. tuberculosis и фильтрат культуры (CF). IFN- γ ответ в > 50 пг/мл рассматривался как положительный. Линии были образованы путем стимулирования РВМС, полученных от TST+ индивидуумов (n=2) или больных ТВ (n=4), лизатами (n=4) □ или CF (n=4) | культур М. tuberculosis, выращенных при гипоксических условиях, или с лизатами культур М. tuberculosis, выращенных при стандартных аэрированных условиях (n=4) ■. Столбцы показывают число отвечающих линий на каждый антиген латентности, и число на вершине каждого столбца показывает усредненные значения IFN- γ продукции этих отвечающих линий. Фиг. 2. Число узнаваемых латентных антигенов у TST положительных индивидуумов (TST+) и больных ТВ. РВМС от 23 TST+ индивидуумов и 20 больных ТВ были стимулированы 25 антигенами латентности М. tuberculosis. Антиген латентности считался узнаваемым, если он индуцировал IFN- γ ответ в ≥50 пг/мл. Для каждого индивидуума было подсчитано число узнаваемых латентных антигенов. Столбцы показывают число индивидуумов, узнающих определенное число латентных антигенов. (a) TST+ индивидуумы. Белые столбцы показывают число TST индивидуумов с недавней конверсией (общее n=12) и черные столбцы - число TST индивидуумов с отдаленной конверсией (общее n=11). (b) Больные ТВ. Белые столбцы показывают число больных с активной формой ТВ (общее n=11) и черные столбцы - число излеченных больных ТВ (общее n=9). Фиг. 3. Профили ответа для четырех наиболее узнаваемых латентных антигенов М. tuberculosis. Продукция IFN- γ клетками РВМС от здоровых контрольных индивидуумов (НС), больных ТВ (ТВ) и TST положительных индивидуумов (TST+) в ответ на четыре антигена латентности М. tuberculosis, а именно Rv1733c (a), Rv2029c (b), Rv2627c (с), Rv2628 (d). РВМС также оценивались по чувствительности к лизату (е) или фильтрату культуры (f) M. tuberculosis, выращенной при гипоксических условиях. Среднее значение для групп индивидуумов показано горизонтальной линией. * Р <0,05; ** Р <0,01; *** Р <0,001. Фиг. 4. Ответ здоровых контролей (НС) на латентные антигены М. tuberculosis. (а) Здоровые контрольные индивидуумы, не инфицированные М. tuberculosis, были разделены на две группы на основании их ответа на лизат М. tuberculosis, выращенных в гипоксических условиях. НС с IFN- γ ответом <100 пг/мл были классифицированы как HC low (n=11) Фиг. 5. Пролиферация CFSE-меченных CD4 лимфоцитов, следующая за стимуляцией пулами пептидов Rv1733c M. tuberculosis. РВМС от TST+ индивидуумов, для которых известно, что они реагируют на рекомбинантный белок Rv1733c, были мечены сукцинимидиловым сложным эфиром карбоксифлуоресцеиндиацетата (CFSE) и стимулированы средой (а), PPD (b), рекомбинантным белком Rv1733c (с) или пулами пептидов Rvl733c (d-f). Клетки окрашивались на CD4, затем пролиферация CD4-положительных клеток оценивалась путем измерения разбавления CFSE при помощи проточного цитометра. Фиг. 6. Профиль цитокинов в ответ на латентные антигены. РВМС от больных ТВ (n=10) и TST положительных индивидуумов (n=10) стимулировали указанными антигенами латентности и лизатами гипоксических М. tuberculosis. После 6 дней супернатант собирали и профиль цитокинов определяли при помощи Cytometric Bead Array (BD) с использованием проточного цитометра FACSCalibur. Количественно оценивались следующие цитокины: TNF α (красный), IL-10 (оранжевый), IL-5 (желтый), IL-4 (зеленый) и IL-2 (голубой); средние уровни цитокинов на антиген указаны для каждой группы. Значительные IL-10 ответы наблюдались для TST позитивов по сравнению с больными ТВ, в частности, для Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628 и RV3129. Фиг. 7. Проверка иммуногенности всех 48 DosR латентных антигенов. Фиг. 8. Частота узнавания предпочтительных ТВ латентных антигенов. Фиг. 9. Диаграмма Эйлера, показывающая распределенные ответы на ТВ латентные антигены. Фиг. 10. Фиг. D. Иммунные ответы у людей, последовавшие за BCG вакцинацией. Индивидуальные (i) и средние (ii) IFN- γ ответы вакцинированных BCG индивидуумов на ТВ латентные антигены. Вакцинированные BCG индивидуумы, не подвергавшиеся какому-либо воздействию М. tuberculosis, показывают низкую продукцию IFN- γ на ТВ латентные антигены (слева), в то время как вакцинированные BCG индивидуумы с признаком воздействия ТВ (т.е. положительный in vitro ответ на ТВ-специфические антигены ESAT6 или CFP10) обладали значительной продукцией IFN- γ на ТВ латентные антигены (справа). Фиг. 11. (Верхняя панель I). Вакцинация HLA-DR3 трансгенных мышей BCG индуцирует слабые иммунные ответы на ТВ антиген латентности HspX и его HLA-DR3 рестриктированный Т-клеточный эпитоп, в то время как значительный ответ наблюдался в отношении рекомбинантных белков Hsp65 и Ag85 и их HLA-DR3 рестриктированные пептиды (как описано в Geluk et al. PNAS 95:10797-802). (Нижняя панель ii). IFN- γ ответы на ТВ латентные антигены у BALB/c мышей. Спленоциты от вакцинированных BCG мышей, стимулированные in vitro ТВ антигенами латентности, продуцировали низкий уровень IFN- γ. В отличие от этого клетки, стимулированные секретированными антигенами Ag85А, продуцируют значительные количества IFN- γ (A). Однако мыши способны на генерирование иммунных ответов на тестируемые ТВ латентные антигены: после трех иммунизаций плазмидной ДНК, кодирующей индивидуальные ТВ латентные антигены, спленоциты продуцируют значительные количества IFN- γ (B). Фиг. 12. ТМНММ (трансмембранный) анализ апостероидных вероятностей ТВ антигенов. Исследование индивидуумов. Исследование включало 20 больных ТВ, 23 здоровых, положительных по кожному туберкулиновому тесту (TST) индивидуумов и 21 здоровый неинфицированный контрольный имдивидуум. Так как нашей изначальной целью был скрининг Т-клеточных ответов на новые латентные антигены, не было сделано предположения в отношении фазы инфекции М. tuberculosis, в которой с наибольшей вероятностью происходит узнавание. Таким образом, гетерогенная группа больных ТВ и TST индивидуумов с конверсией была выбрана в качестве исследуемых индивидуумов. Из 20 больных ТВ 11 имели активное заболевание ТВ, и они лечились от 2 недель до 6 месяцев (в среднем 2,5 месяца), а 9 - с излеченным ТВ, которые лечились от 4 до 63 лет до взятия образца крови (средний интервал 29 лет). Одиннадцать пациентов имели легочный и 9 - внелегочный ТВ. Средний возраст при взятии образца крови составлял 46 лет (от 17 до 75), 14 были мужчинами. Девять больных имели голландское происхождение, 3 были из Северной Африки, 6 были африканцами и 3 были выходцами из Азии. Ни один из больных не имел факторов риска на СПИД. 23 TST-положительных индивидуума все были здоровыми, не вакцинированными BCG индивидуумами с документируемыми TST результатом - затвердением с размером ≥10 мм, главным образом, после контакта с больным, мазок-положительным на легочный ТВ (n=14). Средний возраст составлял 37 лет (диапазон от 21 до 63), 14 были мужчинами. Все были голландцами. От 12 индивидуумов кровь бралась в течение 6 месяцев после TST конверсии, из которых только 5 лечились изониазидом от латентной ТВ инфекции. У оставшихся 11 TST-положительных индивидуумов средний интервал между конверсией и взятием крови составлял 6 лет (диапазон от 2 до 12 лет). Только 2 из этих TST индивидуумов с отдаленной конверсией получили изониазид. Вплоть до времени написания этого документа ни у одного из TST-положительных индивидуумов не развилось активное заболевание ТВ после периода в среднем 4,9 года после TST конверсии. Большинство индивидуумов в этой группе может расцениваться как латентно инфицированные индивидуумы, которые показывают определенный уровень естественной защиты против развития активного заболевания ТВ. В качестве контрольной группы был исследован 21 здоровый, не вакцинированный BCG индивидуум. Ни один из здоровых представителей контрольной группы не обладал каким-либо известным контактом с ТВ. Они были либо TST отрицательными (n=18; у других оставшихся TST не делался), или были тестированы как негативные в отношении специфических белков М. tuberculosis ESAT-6 и CFP-10 при помощи теста ELISPOT на IFN- γ (21). Все представители контрольной группы были голландцами, средний возраст которых составлял 30 лет (диапазон 22-44 года), 7 были мужчинами. Образцы крови были получены от всех исследованных индивидуумов при помощи стандартного отбора из вены с использованием покрытых гепарином патрубков после того, как было получено письменное согласие. Затем с использованием градиента плотности фиколла выделяли РВМС и хранили в жидком азоте, как описано ранее (22). Протокол исследования (Р207/99) был одобрен ревизионным советом Leiden University Medical Center. Антигены и пептиды М. tuberculosis. Для настоящего исследования клетки штамма M. tuberculosis H37Rv выращивали в течение 24 ч в пробирках с плотно закрученными крышками, собирали и лизировали, как описано ранее (16). Эти лизаты, которые в дальнейшем будут называться гипоксическими лизатами, преципитировали ацетоном и диализировали против PBS. Культуральный фильтрат этой культуры, выращенной при низком содержании кислорода, концентрировали при помощи центрипреп-концентратора. Концентрацию белка в конечных препаратах определяли при помощи теста ВСА (Pierce, Rockford, Ilinois). Гипоксический лизат и фильтрат гипоксической культуры были любезно предоставлены Statens Serum Institute (Copenhagen, Denmark). Лизат М. tuberculosis, выращенных в стандартных аэрируемых лабораторных условиях, был предоставлен National Institute of Public Health и Environment (Bilthoven, Нидерланды). Были получены рекомбинантные белки 25 генов регулона спячки М. tuberculosis с наибольшими уровнями экспрессии при регуляции (табл. 1). Гены были амплифицированы при помощи PCR и клонированы при помощи Gateway Technology (Invitrogen, San Diego, CA) в бактериальном экспрессирующем векторе, содержащем N-концевой гистидиновый хвост. Белки сверхэкспрессировали в штамме BL21 (DE3) Escherichia coli В и очищали, как описано ранее (23). Для подтверждения того, что экспрессируются правильные последовательности, у всех вставок была определена нуклеотидная последовательность. Размер и степень очистки проверяли гель-электрофорезом и Western-блоттингом с антителами против нескольких гистидинов. Остаточные уровни экзотоксина составляли менее 50 I.U./мг белка, что определяли в тесте Limulus Amebocyte Lysate (BioWhittaker, Walkersville, MD). Были получены двадцать синтетических пептидов длиной 20 аминокислот (а.к.) каждый, соответствующих антигенам латентности Rv1733c, Rv2029c, Rv2627c и Rv2628, с перекрыванием в 10 а.к. и охватывающих всю аминокислотную последовательность указанных латентных антигенов (22). Для увеличения растворимости последовательность всех пептидов удлиняли на два остатка лизина с С-конца. Для оптимального использования РВМС было решено образовать 9 пулов пептидов с 4 или 5 пептидами в каждом, причем каждый конкретный пептид был представлен в двух различных пулах. Этот способ давал возможность идентификации конкретного пептида в пуле, который узнавался антиген-специфическими Т-клетками. Линии Т-клеток. Были образованы восемь долговременно культивируемых линий Т-клеток в отношении либо лизатов (n=4), либо фильтратов культур (n=4) M. tuberculosis, растущих в обедненных кислородом условиях, с использованием РВМС, полученных от двух больных ТВ и двух TST-положительных индивидуумов, для которых был зафиксирован ответ на HspX. Для сравнения, 4 дополнительных М. tuberculosis-специфических линий Т-клеток были получены стимулированием РВМС, взятыми от трех больных ТВ и от одного TST-положительного индивидуума, лизатами М. tuberculosis, выращенными в стандартных аэрируемых лабораторных условиях. Линии Т-клеток получали, как описано ранее (24). Вкратце, РВМС инкубировались в концентрации 1 ×10 6 клеток/лунка в 24-луночных плашках (Nunc, Roskilde, Denmark) в присутствии 5 мкг/мл антигена, как отмечено выше. После 6 дней добавляли 25 U/мл интерлейкина-2 (Cetus, Amsterdam, Нидерланды), продолжали культивирование еще от 2 до 3 недель. Т-клетки замораживали и хранили в жидком азоте до использования. Анализ пролиферации Т-клеток. Т-клетки (5 ×10 4 /лунка) культивировали с автологическими или совпадающими по HLA-DR облученными РВМС в качестве антиген-представляющих клеток (1,5 ×10 4 /лунка) в трех повторностях, в 96-луночных плашках для микротитрования с лунками с плоским дном (NUNC) в присутствии или при отсутствии антигена. Среду DMEM, модифицированную Iscoves (Gibco, Paisley, Шотландия), дополненную 10% полной плазмой человека, 40 U/мл пенициллина и 40 мкг/мл стрептомицина, использовали в качестве стандартной культуральной среды. Все 25 рекомбинантных латентных антигенов, как показано в табл. 1, исследовали при конечной концентрации 0,33 мкМ, а стандартный лизат М. tuberculosis, гипоксические лизат и фильтрат культуры - при концентрации 1 мкг/мл. Митоген РНА (2 мкг/мл) использовали в качестве положительного контроля. После трехдневного культивирования при 37 °С и 5% СО 2 супернатанты (50 мкл/лунка, объединенные в трех повторностях) собирали для определения IFN- γ, и пролиферацию Т-клеток измеряли по включению [ 3 Н]-тимидина, как описано ранее (22). Пролиферацию выражали индексом стимуляции, вычисленным как отношение количества импульсов в минуту в стимулированных лунках к количеству импульсов в минуту в нестимулированных лунках. Индекс стимуляции ≥4 был предварительно определен как положительный ответ. Анализ стимуляции лимфоцитов. РВМС (1,5 ×10 5 /лунка) культивировали в стандартной культуральной среде в трех повторностях в 96-луночных плашках для микротитрования с округлым дном лунок при 37 °С, 5% СО 2 , в присутствии или в отсутствие антигенов латентности. Те же антигены и в тех же количествах использовали на протяжении всех экспериментов. Из-за нехватки количества Rv1733c число исследованных индивидуумов для этого антигена было уменьшено до 17 здоровых человек контрольной группы, 18 TST+ лиц и 16 больных ТВ. На 6 день супернатанты были собраны (75 мкл/лунка, объединенные в трех повторностях) для определения IFN- γ, и пролиферация Т-клеток измерялась, как описано в другой работе (22). Определение IFN- γ. Концентрацию IFN- γ в супернатантах измеряли методом ELISA (U-CyTech, Utrecht, Нидерланды). Предел чувствительности теста составлял 20 пг IFN- γ/мл. Образцы для ELISA тестировали в двух повторностях. Среднее значение для нестимулированных культур вычитали из среднего значения для стимулированных культур. Положительный ответ был предварительно определен как уровень IFN- γ ≥50 пг/мл в супернатантах от стимулированных линий Т-клеток и в > 100 пг/мл от РВМС культур. Одновременное определение цитокинов. Уровни IFN- γ, TNF α, IL-10, IL-5, IL-4 и IL-2 в супернатантах культур определяли при помощи набора Cytometric Bead Array (CBA) для Th1/Th2 цитокинов человека (BD Biosciences), который позволяет одновременно детектировать несколько цитокинов в одном образце. Анализы проводились по инструкциям производителя. Пролиферация CFSE-меченных РВМС. РВМС оттаивали и ресуспендировали в PBS/0,5% BSA (37 °С) в концентрации 10 ×10 6 клеток/мл. CFSE добавляли до конечной концентрации 5 мкМ и инкубировали 10 мин при 37 °С в темноте. После инкубации добавляли FCS (10%) и клетки дважды промывали в PBS/0,5% BSA. Меченые РВМС (1 ×10 6 клеток/лунка) культивировали в 24-луночных плашках в стандартной культуральной среде в присутствии либо PPD (5 мкг/мл), рекомбинантного белка Rv1733 (20 мкг/мл), пула пептидов Rv1733c (10 мкг/мл каждого пептида), PHA (2 мкг/мл) или стандартной среды. После 6 дней клетки промывали PBS/0,1% BSA и окрашивали на CD4 и затем оценивали пролиферацию CD4 положительных клеток при помощи разведений CFSE с использованием проточного цитометра. Статистический анализ. Для сравнения пропорций иммунокомпетентных клеток в каждой исследованной группе использовали критерий хи-квадрат. Средние IFN- γ ответы оценивали непараметрически при помощи теста Крускал-Валлиса для сравнения всех трех исследованных групп, и U-тест Манн-Уитни использовали в качестве последующего теста, когда сравнивались две группы. Поскольку основной целью исследования был предварительный скрининг потенциальных иммуногенных латентных антигенов, коррекция Бонферрони не применялась. Непараметрический тест Фридмана (отклонение по рангам) применяли для проверки гипотезы, что у TST-положительных индивидуумов 25 латентных антигенов в большинстве случаев должны узнаваться лучше, чем у больных ТВ. Значение Р <0,05 полагали статистически значимым. Для статистического анализа использовали SPSS 10,0 для Windows. Пример 1. Отбор иммуногенных латентных антигенов. Антигены были выбраны из недавно идентифицированного регулона спячки М. tuberculosis, состоящего из 48 генов (табл. 2), которые, как выяснилось, индуцируются во время NRP, при недостатке кислорода и при воздействии малых доз оксида азота (17). Поскольку большинство из этих генов содержит гипотетические открытые рамки считывания с неизвестными функциями, отбор генов для этого постгеномного проекта по обнаружению антигенов не мог основываться на функции белка. Поэтому было решено отбирать гены по их уровню индукции. Для этих целей была рассчитана средняя степень индукции для каждого индивидуального гена, на основе кратности индукций, как наблюдали Voskuil et al. в трех различных in vitro моделях латентности (17). Исходя из данных для 48 генов-кандидатов, 25 наиболее сильно индуцированных генов были выбраны для клонирования и экспрессии рекомбинантных белков (табл. 1; фиг. 7i и 8i). Эти гипотетические белки впоследствии тестировали в эквимолярных концентрациях для того, чтобы иметь возможность прямого сравнения между антигенами латентности, которые значительно отличаются по размеру (от 9 до 74 кДа). Все 25 антигенов узнаются кратковременными линиями Т-клеток, полученных при помощи разрушенных ультразвуком М. tuberculosis. Для первоначальной оценки иммуногенности этих гипотетических латентных антигенов были образованы долговременно культивируемые линии Т-клеток к лизату (n=4) или фильтрату культуры (n=4) M. tuberculosis, выращенной в условиях недостатка кислорода или в стандартных аэрируемых условиях (n=4), и специфичность подтверждали тестами по пролиферации Т-клеток. Важно, что все 25 латентных антигенов узнавались (IFN- γ > 50 пг/мл) по меньшей мере 1 из 12 проверенных линий Т-клеток и 20 антигенов не менее 4 линиями Т-клеток (фиг. 1). Латентные антигены HspX (Rv2031c) и Rv2032 узнавались наиболее часто, 75% проверенных линий Т-клеток со средними IFN- γ уровнями 507 и 129 пг/мл, соответственно, среди отвечающих линий. Большинство латентных антигенов узнавалось линиями Т-клеток, выращенных с гипоксическими лизатами, а также линиями, образованными против фильтрата гипоксической культуры, что свидетельствует о том, что латентные антигены могут также присутствовать вне клеток, в фильтрате культуры. Это подтверждает предыдущее исследование, показывающее, что белки Rv0569, Rv2623 и Rv2626c представлены в фильтрате культуры М. tuberculosis, выращенной в гипоксических условиях (16). Специфические для стандартных лизатов линии Т-клеток узнавали латентные антигены почти так же хорошо, как и линии клеток, специфические для гипоксических лизатов (фиг. 1). Эти данные согласуются с недавним наблюдением, что латентные антигены также экспрессируются в М. tuberculosis в стандартных аэрируемых культурах, когда бактерии собираются во время стационарной фазы роста, хотя и в меньшей степени, чем при культивировании в определенных гипоксических условиях (25). Western-блоттинг-анализ лизатов стандартно-аэрируемых М. tuberculosis, которые использовались для образования линий Т-клеток, подтвердили присутствие в этих препаратах антигена латентности HspX (данные не представлены). Сходные результаты из данных по пролиферации (данные не представлены) подтверждают отвечаемость линий Т-клеток на латентные антигены. Этот первый пробный скрининг продемонстрировал, что все 25 новых микобактериальных латентных антигенов потенциально способны вызывать клеточный иммунный ответ. Пример 2. Продукция интерферона- γ клетками РВМС в ответ на латентные антигены М. tuberculosis. Затем 25 латентных антигенов исследовали на индукцию продукции IFN- γ клетками РВМС от 20 больных ТВ, 23 TST-положительных здоровых индивидуумов и 21 неинфицированного контрольного индивидуума. Для каждого индивидуального антигена латентности была рассчитана пропорция исследования ответов (IFN- γ ≥100 пг/мл) индивидуумов в группе (табл. 1). Авторы также рассчитали пропорцию иммунокомпетентных клеток среди всех 43 инфицированных М. tuberculosis индивидуумов, объединяющих вместе 20 больных ТВ и 23 TST-положительных индивидуума. Недавние анализы показали, что 19 латентных антигенов узнавались по меньшей мере 5% инфицированных М. tuberculosis индивидуумов, с Rv1733c, который узнавался большинством (56%) инфицированных индивидуумов. Остальные 6 проверенных антигенов, Rv0572c, Rv2623, Rv2624c, Rv3127, Rv3131, Rv3134c совсем не узнавались или очень слабо узнавались инфицированными М. tuberculosis индивидуумами. Сходные профили узнавания были получены после анализа данных по пролиферации (не приводятся). а Сокращения: НС, здоровые представители; ТВ, больные ТВ; TST+, положительные по туберкулиновой кожной пробе индивидуумы; HP, гипотетический белок; СНР, консервативный гипотетический белок. Примечание с http://genolist.pasteur.fr/TubercuList/ b IFN- γ ответ в ≥100 пг/мл считался положительным, нет ни одного реагирующего индивидуума. * Р <0,05, X 2 критерий сравнения больных ТВ и TST+ индивидуумов. В дополнение к 25 наиболее сильно индуцируемым DosR генам, вся группа из 48 DosR генов снова была проверена на продукцию INF- γ, включая остальные 23 еще не проверенных ТВ антигена латентности. Все DosR гены проверялись сходным образом, как описано выше, результаты приведены на фиг. 7ii и 8ii. Из нижней панели выявляются 4 дополнительных антигена, которые можно выбрать в качестве потенциальных кандидатов на вакцину: Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX), дающие сильную индукцию INF- γ, в дополнение к Rv1733c, Rv2029c, Rv2627c и RV2628. Пример 3. Больные ТВ и TST-положительные индивидуумы по-разному реагируют на латентные антигены. Средние INF- γ ответы в группах больных ТВ и TST индивидуумов с конверсией были определены для каждого латентного антигена. Рассматривая 25 латентных антигенов как группу, средние INF- γ ответы были сообразно и значительно выше у TST-положительных индивидуумов, которых рассматривали как латентно инфицированных М. tuberculosis, чем у больных ТВ (Р <0,01; тест Фридмана). Чтобы в дальнейшем проанализировать разницу в узнавании антигена, авторы подсчитали отношение между INF- γ ответом для каждого антигена латентности и общим ответом на гипоксический лизат М. tuberculosis у того же индивидуума. Этот анализ корректирует возможные вариации между индивидуумами в общей чувствительности Т-клеток. Когда вышеупомянутый анализ Фридмана был повторен, на этот раз сравнивали средние этих отношений, то было подтверждено, что латентные антигены предпочтительно узнавались у TST-положительных индивидуумов (Р <0,01). При сравнении пропорций иммунокомпетентных клеток в группе TST-положительных индивидуумов и группе больных ТВ для каждого отдельного антигена латентности было обнаружено, что почти все латентные антигены узнавались у большей части TST-положительных индивидуумов по сравнению с больными ТВ (табл. 1). Однако эта тенденция была значительной (Р=0,02) только для Rv2029c, который узнавался у 61% TST-положительных индивидуумов и у 25% больных ТВ. Профили ответов различаются между индивидуумами как в отношении числа латентных антигенов, так и плане специфичности антигенов, которые узнавались (фиг. 2). TST-положительные индивидуумы узнавали в среднем 4 из 25 тестированных латентных антигенов, тогда как больные ТВ, в отличие от первых, узнавали в среднем только один антиген латентности. На фиг. 8 приводится обзор частот узнавания ТВ латентных антигенов Манту-положительными индивидуумами. Для первых серий 25 ТВ латентных антигенов оказалось, что не все антигены узнавались одинаково (Q тест Кочрена, Р <0,001). На основании этого представления авторы выбирали латентные антигены по меньшей мере с 50% узнаванием в первых сериях: Rv1733c, Rv2029c (PfkB), Rv2627c и Rv2628, и из второй серии из 23 ТВ латентных антигенов с теми же характеристиками были выбраны Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX). Эти 8 антигенов предоставляют специфическую подгруппу ТВ латентных антигенов, которая наиболее соответствует целям диагностики и вакцинации, в качестве индивидуальных антигенов или их фрагментов, но наиболее предпочтительно использование в комбинациях из 1, 2, 3, 4, 5, 6, 7 или всех 8 продуктов DosR генов, выбранных из группы, состоящей из Rv1733c, Rv2029c, Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX). Пример 4. Интерферон- γ ответы на часто узнаваемые латентные антигены. Четыре антигена латентности, а именно Rv1733c, Rv2029c, Rv2627c и Rv2628, широко распознавались преимущественно латентно инфицированными индивидуумами и индуцировали сильнейший Th1 ответ, что определялось по продукции IFN- γ. Для каждой изучаемой группы IFN- γ ответы на эти 4 антигена представлены на фиг. 3. Средняя продукция IFN- γ в группе TST-положительных индивидуумов в ответ на Rv1733c, Rv2029c, Rv2627c и Rv2628 составляла 213, 281, 107 и 51 пг/мл соответственно. В отличие от этого, значительно меньший уровень IFN- γ ответов на эти четыре антигена наблюдался у больных ТВ, в среднем 98, 16, <10 и <10 пг/мл соответственно. Это различие в IFN- γ ответе было статистически значимым (Р=0,03) для антигена Rv2029c (фиг. 3). Удивительно, один антиген латентности, Rv2031c, узнавался (IFN- γ > 100 пг/мл) значительно чаще у ТВ, чем у TST индивидуумов с конверсией (Р=0,02) (табл. 1). Однако когда был проведен количественный анализ, при котором непосредственно сравнивалась продукция IFN- γ в двух группах, никакого статистически значимого различия обнаружено не было. Интересно, что TST индивидуумы с конверсией, которые демонстрируют по меньшей мере некоторый ответ на Rv2031c (уровни IFN- γ от 20 до 100 пг/мл), все были с недавней TST конверсией ( <6 месяцев) и поэтому только в последнее время подверглись действию М. tuberculosis. Другой интересный антиген латентности, который авторы изучали, представлял собой Rv3133c/dosR, для которого было показано, что он действует как транскрипционный фактор, опосредующий гипоксический ответ М. tuberculosis (15, 26, 27). Недавно был изучен мутантный штамм Rv3133c/dosR; он характеризовался уменьшенными патологическими изменениями и бактериальной нагрузки у морских свинок, однако, не менял проникновение, выживаемость и размножение М. tuberculosis в моноцитах человека in vitro (28). В авторском исследовании Rv3133c узнавался приблизительно у одной трети TST-положительных индивидуумов и больных ТВ, с усредненными IFN- γ ответами иммунокомпетентных клеток 227 и 145 пг/мл соответственно. На фиг. 9 (диаграмма Эйлера) перекрывание IFN- γ ответов представлено для лучших 4 ТВ латентных антигенов первых серий для индивидуумов, у которых оценивались все индивидуальные антигены: 82,4% Манту-положительных индивидуумов реагировали на один или более из следующих латентных антигенов: Rv1733c, Rv2029c (PfkB), Rv2627c и Rv2628. Сочетание Rv1733c и Rv2627c распознается наиболее часто и является исключительно важным для этого явления; без добавления величины Rv2029c (PfkB) и/или Rv2628. Rv2029c (PfkB) узнается наиболее часто (70,6%) ТВ латентных антигенов у Манту-позитивов. Комбинация этих антигенов, предпочтительно в сочетании с Rv0080, Rv1735c, Rv1736c и/или Rv1737c, могла бы быть весьма подходящей и предпочтительной для проведения диагностических испытаний и для композиции латентной и/или многоэтапной вакцины. На фиг. 12 показана предполагаемая трансмембранная спираль для ТВ латентных антигенов Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c, и Rv1736c (NarX). Предсказание было сделано с использованием ТМНММ 2,0 Server. Krogh A, Larsson В, von Heijne G, Sonnhammer EL. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. 2001. J. Mol. Biol. 305(3): 567-80 (http://www.cbs.dtu.dk/services/TMHMM). В сочетании с данными предсказания эпитопов МНС класса I и II в табл. 3 и 4 и данных in vitro в табл. 5 это исследование демонстрирует, что CD8 Т-клеточные ответы на ТВ антигены могут детектироваться независимо от того, являются ли эти антигены секретируемыми или связанными с мембраной (Klein M.R. et al., HLA-B*35-restricted CD8 T cell epitope in Mycobacterium tuberculosis Rv2903c. 2002 Infect. Immun. 70(2): 981-4). Пример 5. Узнавание латентных антигенов здоровыми контролями. Что неожиданно, 16 из 25 латентных антигенов узнавались Т-клетками меньшинства здоровых индивидуумов (табл. 1), хотя сила иммунного ответа была в целом ниже у инфицированных М. tuberculosis индивидуумов. Поскольку все здоровые не вакцинированные BCG контроли были TST отрицательными и in vitro ответы на М. tuberculosis-специфические иммунодоминантные антигены ESAT6 и CFP10 были также отрицательными в этой группе (данные не представлены), авторы сделали заключение, что наблюдаемые ответы на латентные антигены не были вызваны инфицированием видами М. tuberculosis complex. Тем не менее, 10 из 21 (48%) здоровых контролей дали ответ в значительной степени к гипоксическим лизатам М. tuberculosis, с усредненным IFN- γ ответом среди иммунокомпетентных клеток в 563 пг/мл (фиг. 3 и 4). Это заключение согласуется с данными Т-клеточной перекрестной реактивности к микобактериальным антигенам, полученными в результате предшествующего воздействия микобактерий из окружающей среды. Поскольку ответы на латентные антигены преимущественно наблюдались в той группе здоровых контролей, которые сильно реагировали на гипоксические лизаты М. tuberculosis (фиг. 4), эти ответы, весьма вероятно, также отражают предшествующее воздействие перекрестно-реагирующих микобактерий из окружающей среды. Только Rv1733c также иногда узнавался здоровыми контролями, которые не сильно узнавали гипоксические лизаты М. tuberculosis, со средним IFN- γ в 41 пг/мл, что позволяет сделать предположение о перекрестной реактивности с антигенами, отличными от микобактериальных антигенов. Пример 6. Пролиферация CFSE-меченных РВМС в ответ на пептиды Rv1733c. Для подтверждения белок-специфической активации антигенами латентности авторы определили пептид-специфическую пролиферацию Rv1733c как наиболее часто узнаваемого антигена. РВМС от TST положительных индивидуумов и здоровых контролей, для которых известно реагирование на Rv1733c, были мечены CFSE и стимулированы рекомбинантным белком или пулом пептидов Rv1733c. Через 6 дней клетки были окрашены на CD4, и пролиферация CD4 Т-клеток оценивалась при помощи проточного цитометра. Стимуляция PPD и рекомбинантным белком Rv1733c в обоих случаях индуцировала сильную пролиферацию CD4+ Т-клеток. Кроме того, некоторые пулы пептидов Rv1733c, в частности пул, содержащий пептид 16, обладали способностью индуцировать пролиферацию CD4+ Т-клеток. Это показано на фиг. 5 для РВМС от TST-положительных индивидуумов. Предсказание эпитопов Rv1733c in silico для HLA типа этого донора (DRB 1*15) привело к обнаружению нескольких возможных эпитопов, из которых два расположены в пептиде 16 (VDEPAPPARAIADAALAALG и VDEPAPPARAIADAALAALG), что согласуется с наблюдаемой пролиферацией CD4 Т-клеток в ответ на пептид 16. Пептиды Rv1733c также индуцируют пролиферацию CD4 клеток с использованием РВМС от здоровых контролей, которые реагировали на рекомбинантный белок Rv1733c, что говорит о том, что этот ответ является антигенспецифичным. В дополнение к пептиду 16 из Rv1733c, который был упомянут в первоначальной аппликации как последовательность, содержащая Т-клеточные эпитопы, сейчас определены следующие дополнительные Т- и В-клеточные эпитопы и пептиды в выбранных ТВ антигенах латентности: В табл. 3 и табл. 4 все предсказанные HLA класса-I и -II рестриктированные Т-клеточные эпитопы представлены для лучших 4 ТВ латентных антигенов первой серии (т.е. Rv1733c, Rv2029c (PfkB), Rv2627c и Rv2628) и второй серии (т.е. Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX)) соответственно. Девятичленные аминокислотные последовательности перечислены в табл. 4, которая представляет предсказанные CD8 Т-клеточные эпитопы, рестриктируемые всеми известными HLA класса-I супертипами: А1, А2, A3, А24, В7, В8, В27, В44, В58 и В62. Предсказания делались при помощи NetCTL 1.0 Server, который предсказывает CD8 Т-эпитопы в белковой последовательности. Способ объединяет предсказание пептида МНС связывания, протеасомного С-концевого расщепления и эффективность ТАР транспорта. Сервер позволяет предсказывать CTL-эпитопы, рестриктируемые 10 HLA супертипами. МНС связывание и протеасомное расщепление осуществляется при помощи искусственных нейронных сетей. Эффективность ТАР транспорта предсказывается при помощи матрицы весов. Ссылка: Larsen MV et al., 2005. Eur. J. Immunol. 35(8): 2295-303 (www.cbs.dtu.dk/services/NetCTL). В табл. 4 перечислены двадцатичленные аминокислотные последовательности, которые представляют предсказанные CD4 Т-клеточные эпитопы, рестриктированные 25 HLA класса-II аллелями: DRl (DRBl*0101, *0102), DR3 (DRBl*0301), DR4, (DRBl*0401, *0402, *0404, *0405, *0410, *0421), DR7 (DRBl*0701), DR8 (DRBl*0801, *0802, *0804, *0806), DR11(5) (DRBl*1101, *1104, *1106, *1107), DR13(6) (DRBl*1305, *1307, *1307, *1321), DR15(2) (DRBl* 1501, *1502) и DRB5*0101. Предсказания делались с использованием TEPITOPE 2000 (Vaccinome). TEPITOPE является моделью для предсказания Т-клеточных эпитопов, основанной на HLA класса-II пептидном связывании, и позволяет быстро идентифицировать разнородные HLA класса-II лиганды и зпитопы в множестве белковых последовательностей. Ссылка: Bian H, Hammer J. Discovery of promiscuous HLA-II-restricted T cell epitopes with TEPITOPE. 2004. Methods. 34(4): 468-75. Все перечисленные в табл. 5 20-членные пептиды получены из RVL733C, Rv2029c (PfkB), Rv2627c и Rv2628, которые были проверены на узнавание у двадцати PPD положительных доноров. Клетки были мечены CFSE, стимулированы пептидом, рекомбинантным белком или контрольными антигенами. Пролиферация CD4 и CD8 Т-клеток измерялась при помощи проточного цитометра, и супернатанты собирались и анализировались при помощи одновременных тестов на цитокины. Выделенное красным представляет собой пептиды, которые дали сильную пролиферацию (> 75%) CD4 или CD8 Т-клеток у нескольких доноров, зеленым - пептиды с > 50-75% пролиферацией и светло-зеленым - пептиды с > 20-50% пролиферацией (табл. 5). Сходные результаты ожидаются для других ТВ латентных антигенов (т.е. Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX)). В табл. 6 приведены аминокислотные последовательности Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, RV0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX), а выделенные жирным и подчеркнутые представляют собой линейные и конформационные предсказанные В-клеточные эпитопы соответственно. Предсказания делались с использованием BepiPred 1,0 Server, который предсказывает положение линейных В-клеточных эпитопов при помощи сочетания скрытой модели Маркова и масштабного способа предрасположенности. Конформационные В-клеточные эпитопы были предсказаны только для ТВ латентных антигенов с гомологией последовательности другим известным белкам с подходящими структурными и функциональными параметрами (т.е. Rv2029c (PfkB), Rv1736c (NarX) и Rv1737c (NarK2). (Ref: Larsen, JEP, Lund O, Nielsen M. 2006, Improved method for predicting linear B-cell epitopes (http://www.cbs.dtu.dk/services/BepiPred). Пример 7. Продукция других цитокинов в ответ на латентные антигены. В дополнение к данным для IFN- γ, полученным при помощи ELISA (примеры 1-6), в 6-дневных супернатантах от РВМС, стимулированных рекомбинантными антигенами, были измерены серии других цитокинов. РВМС от больных ТВ (n=10) и от Манту-положительных индивидуумов (n=10) стимулировались рекомбинантными белками Rv1733c, Rv2029c (PfkB), HspX (Rv2031c), Rv2032, Rv2626c, Rv2627c, Rv2628, RV3129, ESAT-6 и CFP10 и дополнительно в качестве положительного контроля гипоксическим лизатом М. tuberculosis (фиг. 6). Данные СВА для IFN- γ подтверждают результаты наблюдений, сделанных при помощи ELISA. TNF α и IL-5 ответы были обнаружены у обеих групп без видимых отклонений. Очень слабые ответы были зафиксированы для IL-2 и IL-4; и если были какие-либо детектируемые ответы, то они наблюдались у больных ТВ. Интересно, что для ряда латентных антигенов авторы наблюдали значительные IL-10 ответы у Манту-позитивов и не у больных ТВ (фиг. 6). Для ТВ вакцин, применяемых до воздействия (т.е. профилактических) в связанной с ТВ областью, обычно принято, что доклинические испытания ТВ вакцин должны включать постепенный отбор и исследования на соответствующих моделях с использованием мышей, морских свинок и не относящихся к человеку приматов (Brandt et al. Infect. Immun. 2000; 68(2): 791-795; Olsen et al. Infect. Immun. 2004; 72(10): 6148-50; Langermans et al. Vaccine 2005; 23(21): 2740-50), включая модели заражения аэрозольной инфекцией в малой дозе (Williams et al., 2005, Tuberculosis (Edinb). 2005; 85(1-2): 29-38). Для ТВ вакцин, применяемых после воздействия (т.е. лечебных), существующие животные модели имитируют заболевание ТВ у человека только отчасти (McMurray, Clin. Infect. Dis. 2000; 30 Suppl. 3: S210-2) и экстраполяция на людей остается неясной. Существующее мнение в области разработки ТВ вакцин состоит в усилении иммунного ответа, индуцированного BCG, новой ТВ вакциной (например, McShane et al. Nat Med. 2004; 10(11): 1240-4; Orrae Tuberculosis (Edinb). 2005; 85(1-2): 13-7). Тем не менее, усиление иммунных ответов может быть достигнуто, только если сначала BCG инициирует ответы. ТВ вакциной, используемой в настоящее время, является Mycobacterium bovis BCG. BCG защищает детей младшего возраста от распространения и тяжелых форм заболевания ТВ, однако, не предохраняет от наиболее распространенных и контагиозных форм пульмонального ТВ у взрослых. Было выдвинуто множество гипотез для объяснения неуспеха BCG, в частности, что иммунные ответы на BCG зависят от воздействия микобактерий окружающей среды (Fine РЕ, 1995 Lancet. 346: 1339-45). Настоящее изобретение предоставляет лучший вариант. Условия, в которые BCG попадают при вакцинации на коже, отличаются от условий, с которыми туберкулезные бациллы сталкиваются при сопротивлении иммунокомпетентных хозяев, главным образом, в иммунных гранулемах в легких. Поэтому профили экспрессии антигенов должны быть различными, включая соответствующие профили иммунного узнавания. Авторы обнаружили, что вакцинация BCG не индуцирует иммунные ответы на ТВ латентные антигены: когда образцы крови (РВМС) вакцинированных BCG здоровых индивидуумов тестируются in vitro, наблюдаются очень слабые иммунные ответы на ТВ латентные антигены по сравнению с индивидуумами, которые подверглись действию ТВ (in vitro положительные на ESAT-6 и CFP10) (фиг. 10). Результат наблюдений этих перекрестно-групповых сравнений был подтвержден на (HLA-A2 и -DR3 трансгенных) мышах для HspX, равно как и для других ТВ латентных антигенов на обыкновенных инбредных мышах при вакцинации BCG (фиг. 11). Поскольку иммунодоминантные ТВ латентные антигены опознаются как часть естественного иммунитета в отношении ТВ у человека и не выявляются при имеющей место в настоящее время процедуре BCG вакцинации, можно представить наиболее обещающих кандидатов для терапевтических ТВ вакцин. (Известно, что среди RD антигенов ни один из ТВ латентных антигенов не входил в число RD антигенов, которые отсутствуют в BCG (Behr et al., 1999). In vitro анализ профиля экспрессии BCG обнаруживает, что экспрессироваться способны все ТВ латентные антигены, за исключением NarX/K2. Таким образом, хотя BCG, в принципе, способны экспрессировать ТВ латентные антигены, что определяется in vitro, условия для экспрессии антигенов, кодируемых регулоном DosR, по-видимому, не являются подходящими. Настоящее изобретение предоставляет DosR латентные антигены, которые наиболее эффективны in vivo, выбранные из группы, состоящей из Rv1733c, Rv2029c (PfkB), Rv2627c, Rv2628, Rv0080, Rv1737c (NarK2), Rv1735c и Rv1736c (NarX), наиболее предпочтительно выбранные из Rv1733c, Rv2029c, Rv2627c и Rv0080, для использования в композициях и/или вакцинах, или альтернативно для экспрессии и/или проявления в рекомбинантных (BCG) микобактериях. Изобретение также предоставляет эпитопы для Т- и В-клеток внутри этих DosR антигенов. Примечание: Результаты получены при помощи TEPITOPE 2000, Vaccinome (Bian H., Hammer J. 2004, Discovery of promiscuous HLA-II-restricted T cell epitopes with TEPITOPE. Methods. 34(4): 468-75). Примечание: Результаты получены при помощи TEPITOPE 2000, Vaccinome (Bian H., Hammer J. 2004, Discovery of promiscuous HLA-II-restricted T cell epitopes with TEPITOPE. Methods. 34(4): 468-75). 1. Wayne, L.G. and H.A. Sramek. 1994. Metronidazole is bactericidal to dormant cells of Mycobacterium tuberculosis. Antiraicrob. Agents Chemother. 38: 2054-2058. 2. Wayne, L.G. and K.Y. Lin. 1982. Glyoxylate metabolism and adaptation of Mycobacterium tuberculosis to survival under anaerobic conditions. Infect. Immun. 37: 1042-1049. 3. Lillebaek, Т., Dirksen, A., Vynnycky, E., Baess, I., Thomsen, V.O. and Andersen, A.B. 2003. Stability of DNA patterns and evidence of Mycobacterium tuberculosis reactivation occurring decades after the initial infection. J. Infect. Dis. 188: 1032-1039. 4. Warren, R.M., van der Spuy, G.D., Richardson, M., Beyers, N., Borgdorff, M.W., Behr, M.A., and van Helden P.D. 2002. Calculation of the stability of the IS6110 banding pattern in patients with persistent Mycobacterium tuberculosis disease. J. Clin. Microbiol. 40: 1705-1708. 5. Fine, P.E. 1995. Variation in protection by BCG: implications of and for heterologous immunity. Lancet 346: 1339-1345. 6. Turner, J., Rhoades, E.R., Keen, M., Belisle, J.T., Frank, A.A., and Orme I.M. 2000. Effective preexposure tuberculosis vaccines fail to protect when they are given in an immunotherapeutic mode. Infect. Immun. 68: 1706-1709. 7. Moreira, A.L., Tsenova, L., Aman, M.H., Bekker, L.G., Freeman, S., Mangaliso, B., Schroder, U., Jagirdar, J., Rom, W.N., Tovey, M.G., Freedman, V.H., and Kaplan G. 2002. Mycobacterial antigens exacerbate disease manifestations in Mycobacterium tuberculosis-infected mice. Infect. Immun. 70: 2100-2107. 8. Repique, C.J., Li, A., Collins, F.M., and Morris S.L. 2002. DNA immunization in a mouse model of latent tuberculosis: effect of DNA vaccination on reactivation of disease and on reinfection with a secondary challenge. Infect. Immun. 70: 3318-3323. 9. Chan, J. and Flynn J. 2004. The immunological aspects of latency in tuberculosis. Clin. Immunol. 110: 2-12. 10. Cunningham, A.F. and Spreadbury C.L. 1998. Mycobacterial stationary phase induced by low oxygen tension: cell wall thickening and localization of the 16-kilodalton alpha-crystallin homolog. J. Bacteriol. 180: 801-808. 11. Yuan, Y., Crane, D.D., and Barry III, C.E. 1996. Stationary phase-associated protein expression in Mycobacterium tuberculosis: function of the mycobacterial alpha-crystallin homolog. J. Bacteriol. 178: 4484-4492. 12. Wilkinson, R.J., Wilkinson, K.A., De Smet, K.A., Haslov, K., Pasvol, G., Singh, M., Svarcova, I. and Ivanyi, J. 1998. Human T- and B-cell reactivity to the 16kDa alpha-crystallin protein of Mycobacterium tuberculosis. Scand. J. Immunol. 48: 403-409. 13. Oftung, F., Borka, E. and Mustafa, A.S. 1998. Mycobacterium tuberculosis reactive T cell clones from naturally converted PPD-positive healthy subjects: recognition of the M. tuberculosis 16-kDa antigen. FEMS Immunol. Med. Microbiol. 20: 319-325. 14. Wayne, L.G. and Hayes L.G. 1996. An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence. Infect. Immun. 64: 2062-2069. 15. Sherman, D.R., Voskuil, M., Schnappinger, D., Liao, R., Harrell, M.I. and Schoolnik, G.K. 2001. Regulation of the Mycobacterium tuberculosis hypoxic response gene encoding alpha-crystallin. Proc. Natl. Acad. Sci. U.S.A. 98: 7534-7539. 16. Rosenkrands, I., Slayden, R.A., Crawford, J., Aagaard, С., Barry, III, C.E. and Andersen, P.. 2002. Hypoxic response of Mycobacterium tuberculosis studied by metabolic labeling and proteome analysis of cellular and extracellular proteins. J. Bacteriol. 184: 3485-3491. 17. Voskuil, M.I., Schnappinger, D., Visconti, K.C., Harrell, M.I., Dolganov, G.M., Sherman, D.R. and Schoolnik, G.K. 2003. Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program. J. Exp. Med. 198: 705-713. 18. Schnappinger, D., Ehrt, S., Voskuil, M.I., Liu, Y., Mangan, J.A., Monahan, I.M., Dolganov, G., Efron, B., Butcher, P.D., Nathan, C. and Schoolnik, G.K. 2003. Transcriptional Adaptation of Mycobacterium tuberculosis within Macrophages: Insights into the Phagosomal Environment. J. Exp. Med. 198: 693-704. 19. Shi, L., Jung, Y.J., Tyagi, S., Gennaro, M.L. and North, R. J. 2003. Expression of Th1-mediated immunity in mouse lungs induces a Mycobacterium tuberculosis transcription pattern characteristic of nonreplicating persistence. Proc. Natl. Acad. Sci. U.S.A. 100: 241-246. 20. Timm, J., Post, F.A., Bekker, L.G., Walther, G.B., Wainwright, H.C., Manganelli, R., Chan, W.T., Tsenova, L., Gold, B., Smith, I., Kaplan, G. and McKinney, J.D. 2003. Differential expression of iron-, carbon-, and oxygen-responsive mycobacterial genes in the lungs of chronically infected mice and tuberculosis patients. Proc. Natl. Acad. Sci. U.S.A 100: 14321-14326. 21. Arend, S.M., Andersen, P., van Meijgaarden, K.E., Skjot, R.L., Subronto, Y.W., van Dissel, J.T. and Ottenhoff, Т.Н. 2000. Detection of active tuberculosis infection by T cell responses to early-secreted antigenic target 6-kDa protein and culture filtrate protein 10. J. Infect. Dis. 181: 1850-1854. 22. Arend, S.M., Geluk, A., van Meijgaarden, K.E., van Dissel, J.T., Theisen, M., Andersen, P. and Ottenhoff, Т.Н. 2000. Antigenic equivalence of human T-cell responses to Mycobacterium tuberculosis-specific RD1-encoded protein antigens ESAT-6 and culture filtrate protein 10 and to mixtures of synthetic peptides. Infect.mmun. 68: 3314-3321. 23. Franken, K.L., Hiemstra, H.S., van Meijgaarden, K.E., Subronto, Y., den Hartigh, J., Ottenhoff, Т.Н. and Drijfhout, J.W. 2000. Purification of his-tagged proteins by immobilized chelate affinity chromatography: the benefits from the use of organic solvent. Protein Expr. Purif. 18: 95-99. 24. Ottenhoff, Т.Н., Elferink, D.G., Hermans, J. and de Vries, R.R. 1985. HLA class II restriction repertoire of antigen-specific T cells. I. The main restriction determinants for antigen presentation are associated with HLA-D/DR and not with DP and DQ. Hum. Immunol. 13: 105-116. 25. Voskuil, M.I., Visconti, K.C. and Schoolnik, G.K. 2004. Mycobacterium tuberculosis gene expression during adaptation to stationary phase and low-oxygen dormancy. Tuberculosis.(Edinb.) 84: 218-227. 26. Florczyk, M.A., McCue, L.A., Purkayastha, A., Currenti, E., Wolin, M.J. and McDonough, K.A. 2003. A family of acr-coregulated Mycobacterium tuberculosis genes shares a common DNA motif and requires Rv3133c (dosR or devR) for expression. Infect. Immun. 71: 5332-5343. 27. Park, H.D., Guinn, K.M., Harrell, M.I., Liao, R., Voskuil, M.I., Tompa, M., Schoolnik, G.K. and Sherman, D.R. 2003. Rv3133c/dosR is a transcription factor that mediates the hypoxic response of Mycobacterium tuberculosis. Mol. Microbiol. 48: 833-843. 28. Malhotra, V., Sharma, D., Ramanathan, V.D., Shakila, H., Saini, D.K., Chakravorty, S., Das, Т.К., Li, Q., Silver, R.F., Narayanan, P.R. and Tyagi, J.S. 2004. Disruption of response regulator gene, devR, leads to attenuation in virulence of Mycobacterium tuberculosis. FEMS Microbiol. Lett. 231: 237-245. 29. Yuan, Y., Crane, D.D., Simpson, R.M., Zhu, Y.Q., Hickey, M.J., Sherman, D.R. and Barry, III, C.E. 1998. The 16-kDa alpha-crystallin (Acr) protein of Mycobacterium tuberculosis is required for growth in macrophages. Proc. Natl. Acad. Sci. U.S.A 95: 9578-9583. 30. Monahan, I.M., Betts, J., Banerjee, D.K. and Butcher, P.D. 2001. Differential expression of mycobacterial proteins following phagocytosis by macrophages. Microbiology 147: 459-471. 31. Florczyk, M.A., McCue, L.A., Stack, R.F., Hauer, C.R. and McDonough, K.A. 2001. Identification and characterization of mycobacterial proteins differentially expressed under standing and shaking culture conditions, including Rv2623 from a novel class of putative ATP-binding proteins. Infect. Immun. 69: 5777-5785. 32. Boon, С., Li, R., Qi, R. and Dick, T. 2001. Proteins of Mycobacteriura bovis BCG induced in the Wayne dormancy model. J. Bacteriol. 183: 2672-2676. 33. Boon, C. and Dick, T. 2002. Mycobacterium bovis BCG response regulator essential for hypoxic dormancy. J. Bacteriol. 184: 6760-6767. 34. Ottenhoff, Т.Н., Verreck, F.A., Lichtenauer-Kaligis, E.G., Hoeve, M.A., Sanal, O. and van Dissel, J.T. 2002. Genetics, cytokines and human infectious disease: lessons from weakly pathogenic mycobacteria and salmonellae. Nat. Genet. 32: 97-105. 35. Flynn, J.L., Chan, J., Triebold, K.J., Dalton, D.K., Stewart, T.A. and Bloom, B.R. 1993. An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J. Exp. Med. 178: 2249-2254. 36. Mayuri, G. Bagchi, Das, Т.K. and Tyagi, J.S. 2002. Molecular analysis of the dormancy response in Mycobacterium smegmatis: expression analysis of genes encoding the DevR-DevS two-component system, Rv3134c and chaperone alpha-crystallin homologues. FEMS Microbiol. Lett. 211: 231-237. 37. Behr, M.A., Wilson, M.A., Gill, W.P., Salamon, H., Schoolnik, G.K., Rane, S. and Small, P.M. 1999. Comparative genomics of BCG vaccines by whole-genome DNA microarray. Science 284: 1520-1523. 38. Colditz, G.A., Brewer, T.F., Berkey, C.S., Wilson, M.E., Burdick, E., Fineberg, H.V. and Mosteller, F. 1994. Efficacy of BCG vaccine in the prevention of tuberculosis. Meta-analysis of the published literature. JAMA 271: 698-702. 39. Brandt, L., Feino, C.J., Weinreich, O.A., Chilima, B., Hirsch, P., Appelberg, R. and Andersen, P. 2002. Failure of the Mycobacterium bovis BCG vaccine: some species of environmental mycobacteria block multiplication of BCG and induction of protective immunity to tuberculosis. Infect. Immun. 70: 672-678. 40. Vekemans, J., Ota, M.O., Sillah, J., Fielding, K., Alderson, M.R., Skeiky, Y.A., Dalemans, W., McAdam, K.P., Lienhardt, C. and Marchant, A. 2004. Immune responses to mycobacterial antigens in the Gambian population: implications for vaccines and immunodiagnostic test design. Infect. Immun. 72: 381-3 88. 41. Ravn, P., Boesen, H., Pedersen, B.K. and Andersen, P. 1997. Human T cell responses induced by vaccination with Mycobacterium bovis bacillus Calmette-Guerin. J. Immunol. 158: 1949-1955. 42. Lim, A., Eleuterio, M., Hutter, B., Murugasu-Oei, B. and Dick, T. 1999. Oxygen depletion-induced dormancy in Mycobacterium bovis BCG. J. Bacteriol. 181: 2252-2256. 43. Parker, K.C., Bednarek, M.A. and Coligan, J.E. 1994. Scheme for ranking potential HLA-A2 binding peptides based on independent binding of individual peptide side-chains. J. Immunol. 152:163. HLA_BIND (http://bimas.cit.nih.gov/molbio/hla_bind/). 44. Rammensee, Friede, Stevanovic, MHC ligands and peptide motifs: 1st listing, Immunogenetics 41, 178-228, 1995; SYFPEITHI (http://www.syfpeithi.de/) and Rammensee, Bachmann, Stevanovic:MHC ligands and peptide motifs. Landes Bioscience 1997 (International distributor - except North America: Springer Verlag GmbH & Co. KG, Tiergartenstr. 17, D-69121 Heidelberg. 45. Buus S., Lauemoller S.L., Worning P., Kesmir C., Frimurer Т., Corbet S., Fomsgaard A., Hilden J., Holm A., Brunak S. Sensitive quantitative predictions of peptide-MHC binding by a 'Query by Committee 1 artificial neural network approach, in Tissue Antigens., 62: 378-84, 2003; NetMHC (http://www.cbs.dtu.dk/services/NetMHC/). 46. Nielsen M., Lundegaard C., Worning P., Lauemoller S.L., Lamberth K., Buus S., Brunak S., Lund O., Reliable prediction of T-cell epitopes using neural networks with novel sequence representations. Protein Sci., 12: 1007-17, 2003. 47. Improved prediction of MHC class I and class II epitopes using a novel Gibbs sampling approach, Nielsen M., Lundegaard C, Worning P., Hvid CS, Lamberth K., Buus S., Brunak S., Lund O., Bioinformatics, 20(9): 1388-97, 2004. 48. Sturniolo, T. et al., Nature Biotechnology 17, 555-562, 1999, Generation of tissue-specific and promiscuous HLA ligand databases using DNA chips and virtual HLA class II matrices; TEPITOPE (http://www.vaccinome.com/pages/597444/). |