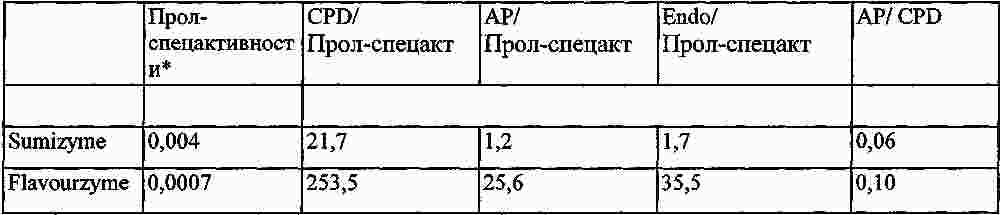
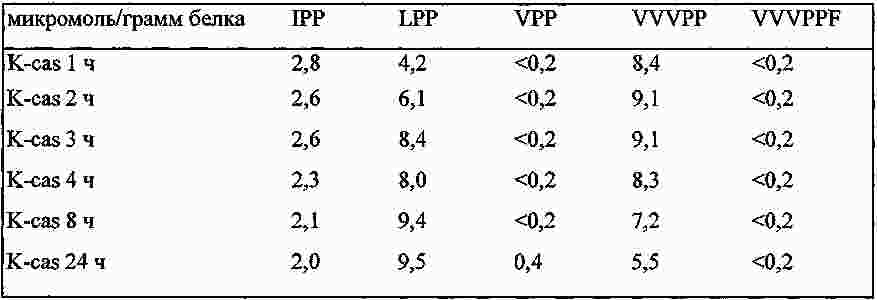
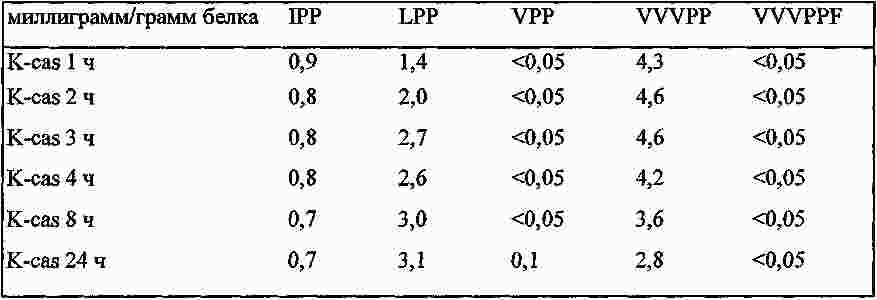
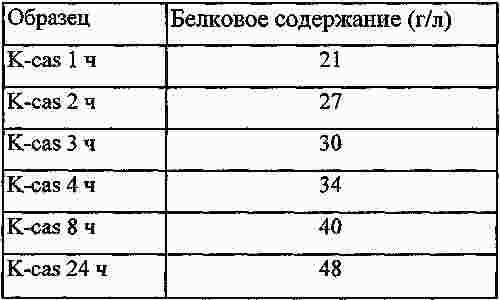
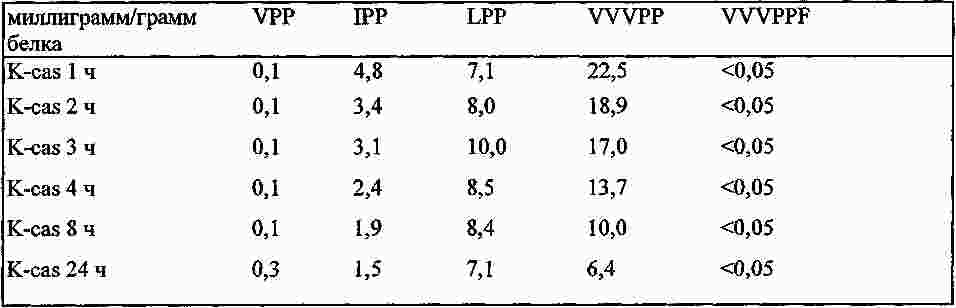
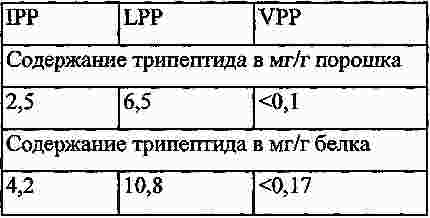
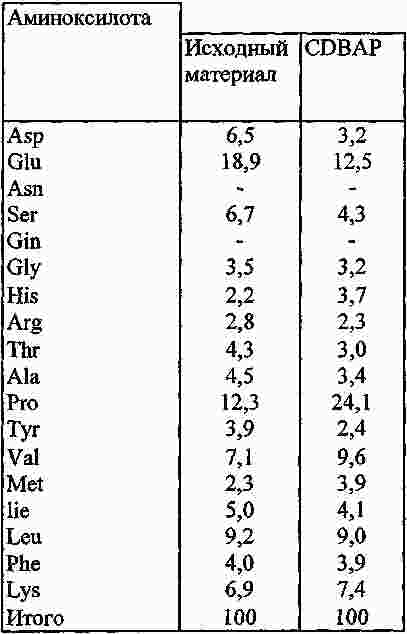
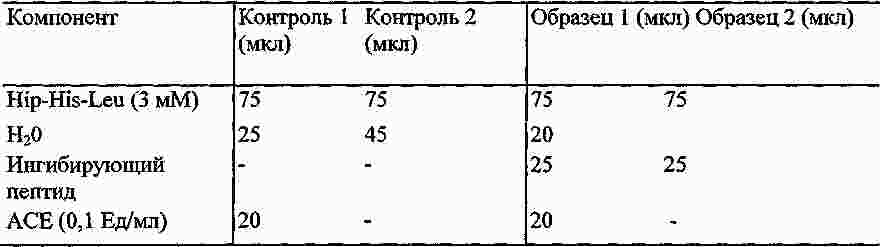
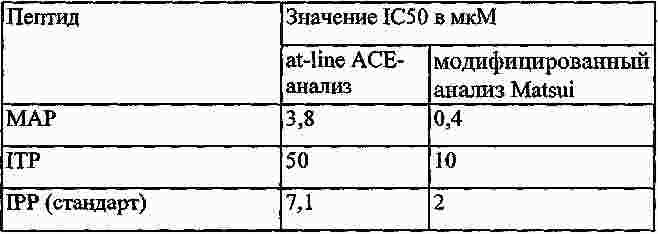
|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000012972b*\id |
|
Термины запроса в документе Реферат Настоящее изобретение раскрывает трипептиды MAP и ITP и их соли, белковые гидролизаты, содержащие указанные трипептиды и их соли, и смеси указанных трипептидов и их солей, используемые для снижения кровяного давления. Изобретение также раскрывает способ получения указанных трипептидов и их солей, который включает ферментативный гидролиз белка пролин-специфичной протеазой и при необходимости преобразование MAP и/или ITP в их соли. Формула [0001] Трипептид MAP или его соль, используемый для снижения кровяного давления. [0002] Трипептид ITP или его соль, используемый для снижения кровяного давления. [0003] Белковый гидролизат, содержащий трипептид MAP по п.1 и трипептид ITP по п.2. [0004] Белковый гидролизат по п.3, который имеет степень гидролиза (DH) от 5 до 50%, предпочтительно от 10 до 40%, более предпочтительно от 20 до 35%. [0005] Смесь трипептидов, содержащая трипептид MAP или его соль по п.1 и трипептид ITP или его соль по п.2. [0006] Смесь трипептидов по п.5, которая содержит по меньшей мере 1 мг МАР/г белка, предпочтительно по меньшей мере 2 мг МАР/г белка, более предпочтительно по меньшей мере 4 мг МАР/г белка. [0007] Смесь трипептидов по п.4 или 5, в которой количество пептидов, имеющих молекулярную массу (MW) менее 500 Да, составляет по меньшей мере 30 мас.% (сухого вещества) смеси пептидов, предпочтительно между 35 и 70 мас.% (сухого вещества) смеси пептидов. [0008] Смесь трипептидов по любому из пп.5-7 или белковый гидролизат по п.3 или 4, который дополнительно содержит IPP или LPP. [0009] Смесь трипептидов по любому из пп.5-7 или белковый гидролизат по п.3 или 4, который содержит от 1 до 90% воды, предпочтительно от 1 до 30% воды, более предпочтительно от 1 до 15% воды. [0010] Способ получения трипептида MAP и/или трипептида ITP и/или их соли по любому из пп.1, 2, который включает ферментативный гидролиз белка пролин-специфичной протеазой и при необходимости преобразование MAP и/или ITP в их соли. Полный текст патента Область техники, к которой относится изобретение Настоящее изобретение относится к получению новых пептидов. Гипертензия представляет собой относительно общее болезненное состояние у людей и является доминирующим фактором риска для развития сердечно-сосудистых заболеваний, почечной недостаточности и нарушений мозгового кровообращения. Возможность использования широкого ряда фармацевтических продуктов, таких как блокаторов кальциевых каналов, бета-блокаторов, диуретических средств, альфа-блокаторов, центральных альфа-антагонистов, антагонистов ангиотензина II и ингибиторов ангиотензин-конвертирующего фермента, демонстрирует, что физиологические механизмы, лежащие в основе генеза гипертензии, являются многосторонними. Физиологическим механизмам гипертензии, в особенности ренин-ангиотензивному механизму, уделяется много научного внимания. В этом механизме ангиотензин секретируется печенью и расщепляется ферментом ренином с образованием биологически неактивного декапептида ангиотензина I. Как только ангиотензин проходит через легочные капилляры, другой фермент, называемый ангиотензин-конвертирующий фермент (именуемый в дальнейшем АСЕ), взаимодействует с ангиотензином I, что приводит к отщеплению последних двух остатков ангиотензина I (His-Leu) и образованию октапептида ангиотензина II. Октапептид ангиотензин II проявляет сильную вазоконстрикторную активность и, вследствие чего, повышает артериальное давление. Ингибирование АСЕ, что влечет за собой низкие уровни ангиотензина II, препятствует вазоконстрикции и, следовательно, высоким кровяным давлениям. Помимо расщепления ангиотензина I АСЕ также может подвергать гидролизу брадикинин, нонапептид, также участвующий в регуляции кровяного давления. В последнем механизме ингибирование АСЕ приводит к повышенным уровням брадикинина, что приводит к расширению кровеносных сосудов и снижению кровяного давления. Ингибирование АСЕ, таким образом, приводит к понижению кровяного давления посредством по меньшей мере двух отдельных механизмов. Также известно, что октапептид ангиотензин II стимулирует высвобождение альдостерона из коры надпочечников. Органом-мишенью для альдостерона являются почки, где альдостерон обеспечивает повышенную реабсорцию натрия из почечных канальцев. Также посредством этого третьего механизма ингибирование АСЕ снижает кровяное давление, но в данном случае - путем снижения реабсорции натрия. Вследствие его многочисленных физиологических действий ингибирование протеолитической активности АСЕ является эффективным способом снижения кровяного давления. Эти наблюдения влекут за собой создание ряда эффективных фармацевтических средств, снижающих кровяное давление, таких как каптоприл и эналаприл (Ondetti, M.A. et al., 1977, Science, Washington DC, 196, 441-444). Поскольку гипертензия является общим болезненным состоянием, то будет предпочтительным уравновесить эти нежелательные результаты современного образа жизни со слабоактивными натуральными ингредиентами, в особенности слабоактивные натуральные ингредиенты, которые могут быть включены в продукты питания или напитки, поскольку такие продукты регулярно потребляются. Альтернативно, такие слабоактивные натуральные ингредиенты могут включаться в диетические добавки. За последние десятилетия было обнаружено, что специфичные пептиды в кисломолочных продуктах обладают способностью ингибирования АСЕ и могут вызывать снижение кровяного давления у субъектов, страдающих гипертензией. На сегодняшний день ряд in vitro и некоторые исследования на животных продемонстрировали АСЕ-ингибирующие действия различных пептидов, полученных из различных источников белка. Несмотря на то что исследования ингибирования АСЕ in vitro обнаружили многие различные пептидные последовательности, следует отметить, что пептиды, ингибирующие АСЕ, нуждаются в циркуляции в крови для проявления своего действия in vivo. Их роль заключается в том, что эффективные пептиды, ингибирующие АСЕ, должны препятствовать расщеплению посредством протеолитической системы пищеварения желудочно-кишечной тракта и должны оставаться интактными в течение дальнейшего переноса через стенку кишечника. Структурно-функциональный анализ различных пептидов, ингибирующих АСЕ, натолкнул на мысль, что они обычно содержат Pro-Pro, Ala-Pro или Ala-Hyp на С-концевой последовательности (Maruyama, S. and Suzuki, H., 1982; Agric Biol Chem., 46(5): 1393-1394). Эта находка частично объясняется тем фактом, что АСЕ является пептидилдипептидазой (ЕС3.4.15.1), не способной расщеплять пептидные связи, включая пролиновые. Таким образом, из трипептидов, имеющих структуру Хаа-Pro-Pro, дипептид Pro-Pro не может быть удален, поскольку связь Хаа-Pro не может быть расщеплена. Вследствие чего можно допустить, что при наличии в высоких концентрациях трипептиды, содержащие структуру Хаа-Pro-Pro, будут ингибировать активность АСЕ. Поскольку не только для АСЕ, но и почти для всех протеолитических ферментов характерны сложности в расщеплении связей Хаа-Pro или Pro-Pro, то убеждение в том, что наличие (многочисленных) пролиновых остатков на карбоксиконцевом конце пептидов приводит к относительной устойчивости молекул к действию протеаз, является почти очевидным. Подобные пептиды, содержащие гидроксипролин (Hyp) вместо пролина, относительно устойчивы к действию протеаз. В связи с чем можно сделать вывод, что пептиды, содержащие один или более (гидрокси)пролиновые остатки на карбоксиконцевом конце, способны избегать протеолитического расщепления в желудочно-кишечном тракте. Эти заключения помогут понять заметное действие по снижению кровяного давление in vivo специфичных пептидов, ингибирующих АСЕ: они не только испытывают трудности структурных требований для ингибирования АСЕ, они также являются стойкими при расщеплении протеолитической системой расщепления желудочно-кишечного тракта и остаются интактными в течение дальнейшего переноса через стенку кишечника. Стойкие активности в отношении ингибирования АСЕ сообщались для трипептидов Leu-Pro-Pro (JP 02036127), Val-Pro-Pro (ЕР 0583074) и Ile-Pro-Pro (J. Dairy Sci., 78:777-7831995)). Исходно, все пептиды, ингибирующие АСЕ, характеризовались на основании их действия in vitro в отношении активности АСЕ и трипептиды Ile-Pro-Pro (именуемый в дальнейшем IPP), Val-Pro-Pro (именуемый в дальнейшем VPP) и Leu-Pro-Pro (именуемый в дальнейшем LPP) выпадали из-за их стойкого действия в отношении ингибирования АСЕ, что приводило к относительно низким значениям IC 50 . В дальнейшем предполагаемые антигипертензивные действия трипептидов VPP, а также IPP могут быть подтверждены на спонтанно-гипертензивных крысах (Nakamura et al., J. Dairy Sci., 78:12531257 (1995)). В этих экспериментах ингибирующие трипептиды были получены из молочно-кислых бактерий сквашенного молока. Во время сквашивания молока желаемые пептиды продуцируются протеазами посредством роста молочно-кислых бактерий. АСЕ ингибирующие пептиды концентрировали из молочных продуктов после электродиализа, диализа с применением мембран из полых волокон или хроматографических способов для предоставления возможности их распространения в виде таблеток, подобных концентрированным диетическим добавкам или пастилкам. Настоящее изобретение относится к новым трипептидам MAP и/или ITP или к соли MAP и/или соли ITP. Настоящее изобретение также относится к белковому гидролизату, содержащему MAP и/или ITP или соль MAP и/или ITP. Предпочтительно белковый гидролизат имеет от 5 до 50% DH (степень гидролиза), более предпочтительно от 10 до 40% и наиболее предпочтительно от 20 до 35% DH. Кроме того, настоящее изобретение относится к смеси пептидов, содержащей MAP и/или ITP или соль MAP и/или ITP. Предпочтительно эта смесь пептидов содержит по меньшей мере 1 мг МАР/г белка, более предпочтительно по меньшей мере 2 мг/г и более предпочтительно по меньшей мере 4 мг/г белка. Предпочтительно смесь пептидов также содержит LPP и/или IPP. Таким образом, смесь пептидов предпочтительно содержит по меньшей мере 1 мг IPP/г белка, более предпочтительно по меньшей мере 2 мг/г и наиболее предпочтительно по меньшей мере 4 мг/г белка и/или смесь пептидов предпочтительно также содержит по меньшей мере 1 мг LPP/г белка, более предпочтительно по меньшей мере 2 мг/г и наиболее предпочтительно по меньшей мере 4 мг/г белка. Эти смеси пептидов предпочтительно содержат пептиды не менее 30 мас.% (сухого вещества), имеющие молекулярную массу (MW) менее чем 500 Да, более предпочтительно от 35 до 70 мас.% (сухого вещества) смеси пептидов пептиды, имеющие MW менее чем 500 Да. Кроме того, настоящее изобретение относится к ферментативному процессу выработки MAP и/или ITP предпочтительно путем ферментативного гидролиза источника белка. Как показано в настоящем изобретении, трипептиды MAP и ITP являются достаточно эффективными в отношении ингибирования АСЕ. Эти ингибирующие эффекты сопоставляются с удивительно низкими значениями IC 50 , соответственно 0,4 для MAP и 10 для ITP (в мкМ), как определено в экспериментальной части настоящего изобретения. Кроме того, обнаружено, что оба трипептида MAP и ITP не поддаются расщеплению протеолитическими ферментами желудочно-кишечного тракта и, следовательно, как ожидается, являются устойчивыми в кишечном тракте человека. Трипептид MAP и/или трипептид ITP и их соли, следовательно, являются чрезвычайно пригодными для эффективного ингибирования АСЕ и могут, например, использоваться в функциональных продуктах питания в качестве нутрицевтиков или лекарств. Понятие нутрицевтик в используемом здесь значении указывает на пригодность как к пищевой, так и к фармацевтической областям применения. Таким образом, новые фармацевтические композиции, содержащие MAP и/или ITP, могут найти свое применение в качестве добавок к продуктам питания или напиткам и в качестве фармацевтических составов или лекарственных средств для энтерального или парентерального применения, которые могут быть в виде твердых составов, таких как капсулы и таблетки, или жидких составов, таких как растворы, суспензии и эмульсии. Трипептиды MAP (Met-Ala-Pro) и ITP (Ile-Thr-Pro) могут приготавливаться с помощью различных способов, включая химический синтез, ферментативный гидролиз и сбраживания растворов, содержащих белки. Идентификация биологически активных пептидов в комплексных смесях, таких как гидролизаты белков, или жидкостях, образованных в результате брожения, является трудной задачей. Кроме основных вопросов, используем ли мы правильный белковый субстрат, используем ли мы правильный фермент, используем ли мы правильную микрокультуру, ожидается, что некоторые биологически активные пептиды могут присутствовать в комплексных образцах, которые содержат тысячи различных пептидов. Традиционные способы идентификации, использующие повторяющиеся циклы фракционирования с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ) и биохимическое определение, обычно являются трудоемкими и подвержены к потерям присутствующих биологически активных пептидов, делая обнаружение соответствующей биологической активности крайне затруднительным. В настоящей работе было использовано довольно сложное оборудование и был проведен скрининг множества различных белковых гидролизатов и ферментационных бульонов, что в конечном итоге привело к идентификации нами двух новых пептидов MAP и ITP, которые обладают свойствами в отношении ингибирования АСЕ. В нашем способе непрерывный биохимический анализ был связан в режиме реального времени с ВЭЖХ системой фракционирования. ВЭЖХ-элюат разделялся между непрерывного АСЕ биоанализа и способа химического анализа (масс-спектрометрия). Неочищенные гидролизаты и ферментационные бульоны разделялись посредством ВЭЖХ и далее наличие биологически активных компонентов определялось с помощью биохимического способа в режиме реального времени. Масс-спектр записывался непрерывно, в результате чего информация о структуре белков являлась легкодоступной, когда пептид демонстрировал положительный сигнал при биохимическом анализе. Трипептиды MAP и ITP, идентифицированные вышеуказанным способом, могут быть получены различными способами, включая экономически эффективные способы получения. Получение посредством химического синтеза является возможным при использовании традиционных способов, которые, к примеру, описываются в книге N. Sewald and H.D. Jakubke "Peptides: Chemistry and Biology" (Eds. Wiley-VCH Verlag GmbH, 2002, Chapter 4). В частности, экономически эффективные способы химического синтеза пептидов, пригодные для массового производства, основаны на применении алкилхлорформиатов или пивалоил хлорида для активации карбоксильной группы и связаны с применением сложных метиловых эфиров для защиты С-конца и бензилоксикарбонила (Z) или трет-бутилоксикарбонильных групп для защиты N-конца. Например, в случае MAP, L-пролина сложный метиловый эфир может связываться с Z-Ala, активированного изобутилхлорформиатом; итоговый дипептид может быть Z-де-защищен посредством гидрогенолиза при использовании водорода и Pd на С и соединен снова с Z-Met, активированного изобутилхлорформиатом; итоговый трипептид метилового эфира подвергается гидролизу при использовании NaOH и после Z-де-защиты посредством гидрогенолиза получается трипептид Met-Ala-Pro. Аналогичным образом может синтезироваться Ile-Thr-Pro, но во время реакций связывания гидрокси-функция Thr нуждается в защите бензилом; затем, на конечной стадии, эта группа синхронно исключается в процессе Z-де-защиты. MAP и/или ITP могут также производиться путем ферментативного гидролиза или посредством ферментационных способов при использовании любого белкового субстрата, содержащего последовательности аминокислот MAP и/или ITP. Преимущественно белковый субстрат содержит оба фрагмента MAP и ITP. Предпочтительными белковыми субстратами для таких ферментативных или ферментационных способов являются коровье молоко или казеиновая фракция коровьего молока. Посредством оптимизации условий ферментации или гидролиза продукция биологически активных молекул MAP и/или ITP может быть максимально повышена. Специалист, пытающийся максимально увеличить продукцию, будет знать, каким образом устанавливать параметры процесса, такие как время гидролиза/ферментации, температура гидролиза/ферментации, тип фермента/микроорганизма и концентрация и др. MAP и/или ITP или композиции, содержащие MAP и/или ITP, являются преимущественно продуктами гидролиза и предпочтительно изготавливаются по способу, включающему следующие стадии: (a) ферментативный гидролиз пригодного белкового субстрата, содержащего MAP или ITP в их аминокислотной последовательности, что приводит к образованию гидролизованного белкового продукта, содержащего трипептиды MAP и/или ITP; (b) отделение от гидролизованного белкового продукта фракции с высоким содержанием трипептида MAP и/или трипептида ITP; и необязательно (c) концентрирование и/или высушивание фракции, полученной по стадии b), для получения концентрированной жидкости или твердого вещества с высоким содержанием трипептида MAP и/или трипептида ITP. Стадия ферментативного гидролиза (а) может представлять собой ферментативную обработку пригодного белкового субстрата, приводящую к гидролизу белка, что приводит к выделению MAP и/или ITP трипептидов. Несмотря на то что для высвобождения желаемых трипептидов из белкового субстрата могут использоваться различные ферментативные комбинации, предпочтительным ферментом, используемым в настоящем процессе, является пролин-специфичная эндопротеаза или пролин-специфичная олигопептидаза. Пригодным белковым субстратом может являться любой субстрат, содержащий аминокислотную последовательность MAP и/или ITP. Известными белковыми субстратами, содержащими MAP, являются, например, казеин, пшеница с высоким содержанием клейковины, белковый изолят подсолнечника, рисовый белок, яичный белок. Пригодные белковые субстраты предпочтительно включают последовательности аминокислот АМАР или РМАР, которые встречаются в бета-казеине коровьего молока, альфа-глиадиновой фракции пшеницы с высоким содержанием клейковины и в 2S-фракции белкового изолята подсолнечника. Казеиновым субстратом может быть любой материал, который содержит значительное количество бета-казеина или альфа-s2-казеина. Примерами пригодных субстратов являются молоко, а также казеин, порошок казеина, казеиновые порошковые концентраты, казеиновые порошковые изоляты, или бета-казеина или альфа-s2-казеина. Предпочтительно субстратом, который имеет высокое содержание казеина, является белковый изолят казеина (CPI). Ферментом может быть любой фермент или комбинация ферментов, которые способны подвергать гидролизу белок, такой как бета-казеин и/или альфа-s2-казеин, что приводит к выделению одного или более трипептидов MAP и/или ITP. Стадия отделения (b) может быть выполнена любым известным для специалиста из уровня техники способом, например путем преципитации, фильтрации, центрифугации, экстракции или посредством хроматографии или их комбинациями. Предпочтительно стадия отделения (b) выполняется при использовании способов микро- или ультрафильтрации. Размер пор мембран, используемый в стадии фильтрации, так же как и заряд мембран могут использоваться для регулирования отделения трипептида MAP и/или трипептида ITP. Фракционирование казеиновых белковых гидрализатов при использовании мембран UF/NF описывается в Y. Poilot et al., Journal of Membrane Science 158 (1999), 105-114. Стадия концентрирования (с) может включать нанофильтрацию или выпаривание фракции, образованной по стадии (b) для получения высоко концентрированной жидкости. Если образованы подходящим образом, например, с низкой активностью воды (Aw), низким значение рН и предпочтительно при наличии консервантов, таких как бензоат или сорбат, такие концентрированные жидкие композиции формируются приемлемым способом при сохранности трипептидов изобретения. Необязательно, за стадией высушивания, например, путем сушки распылением или лиофильной сушки, следует стадия выпаривания для получения твердого вещества, содержащего высокие концентрации MAP и/или ITP. Ферментативный процесс содержит предпочтительно одну стадию инкубации фермента. Ферментативный процесс настоящего изобретения, кроме того, относится к применению пролин-специфичной протеазы, которая предпочтительно не содержит загрязнения ферментативными активностями. Пролин-специфичная протеаза определяется как протеаза, которая подвергает гидролизу пептидную связь на карбоксиконцевом участке пролина. Предпочтительной пролин-специфичной протеазой является протеаза, которая гидролизует пептидную связь на карбоксиконцевом участке пролина и аланиновых остатках. Пролин-специфичная протеаза предпочтительно способна подвергать гидролизу крупные белковые молекулы, такие как полипептиды или сами белки. В целом, время инкубации по способу изобретения составляет менее 24 ч, предпочтительно время инкубации составляет менее 10 ч и более предпочтительно менее 4 ч. Температура инкубации обычно составляет выше 30 °С, предпочтительно выше 40 °С и более предпочтительно выше 50 °С. Другим аспектом настоящего изобретения является процесс очистки и/или отделения трипептидов MAP и ITP от гидролизованного белка. Большинство гидролизованных белков согласно изобретению предпочтительно способны выпадать в осадок при выбранных рН условиях. Этот процесс очистки включает изменение рН до рН, в результате чего большинство гидролизованных и негидролизованных белков осаждаются и отделяются осажденные белков от (биологически активных) трипептидов, которые остаются в растворе. Для получения настоящих трипептидов с пролиновым остатком на карбоксиконцевом конце, применение протеазы, которая может расщепляться на карбоксиконцевом участке пролиновых остатков, отражает предпочтительное мнение. Так называемые пролилолигопептидазы (ЕС 3.4.21.26) обладают уникальной способностью предпочтительного расщепления пептидов с карбоксиконцевой стороны пролиновых остатков. Пролилолигопептидазы также обладают способностью расщеплять пептиды на карбоксильном конце остатков аланина, однако последняя реакция менее эффективна, чем расщепление пептидных связей, включающих пролиновые остатки. Во всех достаточно охарактеризованных пролин-специфичных протеазах, выделенных из млекопитающих, а также из микробных источников, идентифицирован уникальный домен, который не позволяет крупным пептидам приблизиться к активному сайту фермента. Также эти ферменты не способны расщеплять пептиды, содержащие более чем приблизительно 30 аминокислотных остатков, вследствие чего такие ферменты в настоящее время относятся к "пролилолигопептидазам" (Fulop et al. Cell, Vol. 94, 161-170, July 24, 1998). Как следствие, для этих пролилолигопептидаз требуется проведение предварительного гидролиза с другими эндопротеазами перед тем, как они могут проявлять их гидролитическое действие. Однако, как описывается в WO 02/45523, даже комбинация пролилолигопептидазы с другой эндопротеазой приводит к образованию гидролизатов, отличающихся значительно увеличенным содержанием пептидов с карбоксиконцевым пролиновым остатком. Вследствие чего, такие гидролизаты образуют превосходное исходное положение для выделения трипептидов с АСЕ-ингибирующими действиями in vitro, а также повышенную устойчивость к разрушению протеолитическими ферментами в желудочно-кишечном тракте. Термины «пептид » или «олигопептид » в рамках настоящего изобретения определены как цепи, содержащие по меньшей мере два аминокислотных остатков, связанных пептидными связями. Термины «пептид » и «олигопептид » считаются синонимами (что соответствует общепринятому толкованию), и каждый из этих терминов может применяться вместо другого в зависимости от контекста. В рамках настоящего изобретения «полипептид » определен как цепь, содержащая более 30 аминокислотных остатков. Все формулы или последовательности (олиго)пептидов и полипептидов записаны в настоящем патенте слева направо в направлении от аминоконца к карбоксиконцу в соответствии с общепринятой практикой. Буквенные коды аминокислот, используемые в изобретении, общеизвестны в технике и могут быть найдены в Sambrook, et al. (Molecular Cloning: A Laboratory Manual, 2 nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). Смесь пептидов представляет собой композицию, содержащую не менее 30 мас.% (сухого вещества) пептидов, определенных способом определения азота Kjeldahl в сочетании с определением пептида, имеющего MW менее 3500 Да. Эндопротеаза определяется здесь как фермент, который гидролизует пептидные связи в полипептиде в эндофазе и принадлежит к группе ЕС 3.4. Эндопреотеазы подразделяются на подподклассы на основании каталитического механизма. Существуют подподклассы серинэндопротеаз (ЕС 3.4.21), цистеинэндопротеаз (ЕС 3.4.22), аспарагиновых эндопротеаз (ЕС 3.4.23), металлоэндопротеаз (ЕС 3.4.24) и треонинэндопротеаз (ЕС 3.4.25). Экзопротеазы определены здесь как ферменты, которые гидролизуют пептидные связи, расположенные рядом с концевой α-аминогруппой ( «аминопептидазы »), или пептидные связи между концевой карбоксильной группой и предпоследней аминокислотой ( «карбоксипептидазы »). В WO 02/45524 описаны пролин-специфичные протеазы, полученные из Aspergillus niger. Фермент, полученный из A. niger, расщепляет предпочтительно на карбоксиконце пролина, но также может расщеплять на карбоксиконце гидроксипролина и, с более низкой эффективностью, на карбоксиконце аланина. В WO 02/45524 также указано, что не существует гомологии между этим ферментом, полученным из A. niger, и известной пролилолигопептидазой из другого микробного источника или млекопитающего. В отличие от известных пролилолигопептидаз, фермент из A. niger имеет кислый оптимум рН. Несмотря на то что известные пролилолигопептидазы, а также фермент, полученный из A. niger, являются так называемыми сериновыми протеазами, мы демонстрируем в настоящем изобретении (пример 1), что фермент из A. niger принадлежит к belongs to a совершенно другому подсемейству. Секретируемый фермент из A. niger, по-видимому, является членом семейства S28 сериновых пептидаз, а не семейства S9, в котором сгруппированы большинство цитозольных пролилолигопептидаз (Rawling N.D. and Barrett, A.J.; Biochim. Biophys. Acta 1298 (1996) 1-3). В примере 2 показывается рН и температурный оптимумы пролин-специфичной протеазы, полученной из A. niger. В примере 3 проиллюстрировано высокое преимущество полученной из A. niger пролин-специфичной эндопротеазы в отношении расщепления карбоксиконца пролиновых и аланиновых остатков. Кроме того, нами продемонстрировано в этом примере, что ферментативный препарат, полученный из A. niger, как используется в настоящем изобретении, является предпочтительно более чистым, означая, что не присутствует другая значимая эндопротеолитическая активность, чем эндопротеолитическая активность, присущая чисто пролин-специфичной эндопротеазе. Нами также продемонстрировано, что ферментативный препарат, полученный из A. niger, предпочтительно используемый согласно настоящему изобретению, не включает какие-либо экзопротеолитические, в особенности аминопептидолитические побочные активности. Предпочтительно экзопротеолитическая активность отсутствует в ферментативном препарате, полученном из A. niger, используемом в способе изобретения. Экспериментальное подтверждение того, что пролин-специфичная эндопротеолитическая активность фактически отсутствует в нерекомбинантных штаммах Aspergillus может быть найдено в WO 02/45524 и также проиллюстрировано в примере 3 настоящего патента. Поскольку осуществление способа настоящего изобретения возможно посредством инкубации казеинового субстрата только с пролин-специфичной эндопротеазой, условия для оптимальной инкубации, такие как температура, рН и др., могут быть без труда подобраны и не должны фиксироваться на недостаточно оптимальных условиях, как это может быть в случае, если используются два или более фермента. Более того, предотвращается образование нежелательных побочных продуктов, как, например, дополнительных, не биологически активных пептидов или свободных аминокислот, приводящих к постороннему вкусу бульона. Обладание большими степенями свобод в выборе условий реакции делает более легким выбор других возможных критериев. Например, более легким является выбор условий, которые менее чувствительны к микробным инфекциям и к оптимизированным рН условиям относительно дальнейших стадий осаждения белков. В примере 4 мы демонстрируем, что фермент из Aspergillus является не олигопептидазой, а настоящей эндопептидазой, способной гидролизовать интактные белки, крупные пептиды, а также более мелкие пептидные молекулы, без необходимости в присутствии дополнительно эндопротеазы. Это новое и неожиданное открытие открывает возможность применения фермента из A. niger для приготовления гидролизатов с беспрецедентно высокими содержаниями пептидов с карбоксиконцевым пролиновым остатком, поскольку не требуется дополнительной эндопротеазы. Такие новые гидролизаты могут быть приготовлены из различных белковоподобных исходных материалов, происходящих из овощей и животных. Примерами таких исходных материалов являются казеины, желатин, рыбьи или яичные белки, пшеница с высоким содержанием клейковины, соевый и гороховый белки, а также рисовый белок и белок подсолнечника. Поскольку известно, что натрий играет важную роль в гипертензии, предпочтительные субстраты для продукции пептидов, обладающих способностью ингибирования АСЕ, представляют собой кальций и калий, а не соли натрия этих белков. Оптимум рН пролилэндопротеаз, полученных из A. niger, составляет около 4,3 (см. фиг. 1). Из-за такого низкого оптимума рН инкубирование казеина коровьего молока с пролилэндопротеазой из A. niger не является очевидным. Казеин коровьего молока будет выпадать в осадок, если рН опустится ниже 6,0, но при рН, равном 6,0, фермент из A. niger обладает только ограниченной активностью. Однако в примере 5 показано, что даже при таком неблагоприятном условии при инкубировании с полученной из A. niger пролилэндопротеазой можно привести к образованию некоторых известных пептидов, обладающих способностью ингибировать АСЕ, таких как IPP и LPP. Достаточно неожиданно, что при таких условиях не синтезировались VPP. Казеин коровьего молока объединяет ряд различных белков, включая бета-казеин и каппа-казеин. Согласно известным аминокислотным последовательностям бета-казеин включает трипептиды IPP, VPP и LPP, ингибирующие АСЕ. Каппа-казеин включает только IPP. Тот факт, что фермент, полученный из A. niger, не обладает какой-либо измеряемой аминопептидазной активностью, убедительно указывает, что сформированный IPP высвобождается из -А107-I08-Р109-Р110-последовательности, присутствующей в каппа-казеине. По-видимому, пептидная связь карбоксиконца IPP расщепляется посредством основной активности полученной из A. niger пролилэндопротеазы, тогда как расщепление предыдущей Ala-Ile связи осуществляется Ala-специфичной концевой активностью. Аналогично, отсутствие VPP может объясняться, исходя из отсутствия аминопептидазной концевой активности. VPP содержится в бета-казеине в последовательности -P 81 -V 82 -V 83 -V 84 -P 85 -P 86 -. Таким образом, пролин-специфичная эндопротеаза удаляет последовательность VVVPP, но не способна отщеплять VPP. Эти результаты были получены путем инкубирования казеина с эндопротеазой, полученной из A. niger, в простом одностадийном derived ферментативном способе. Водные растворы, содержащие белок, высоко чувствительны к микробным инфекциям, особенно если хранятся в течение многих часов при значениях рН выше 5,0 и при температуре 50 °С или ниже. В особенности микробные токсины, которые могут образовываться в течение таких длительных инкубационных стадий и которые способны сохраняться при последующих стадиях нагревания и образовывать потенциальную угрозу пищевым процессам. По настоящему изобретению предпочтительно применяется температура инкубации выше 50 °С. В сочетании с одностадийным ферментативным способом, в котором инкубация фермента осуществляется в течение периода менее чем 24 ч, предпочтительно менее 8 ч, более предпочтительно менее 4 ч, способ изобретения дает преимущество повышенной к воздействию микроорганизмов. Используя настоящее соотношение фермент-субстрат в сочетании с высокими температурными условиями, удаление IPP и LPP завершается в 3-часовой инкубационный период. Поскольку АСЕ-ингибирующие пептиды IPP и LPP могут быть удалены из казеина при использовании одной практически очищенной эндопротеазы, осуществление способа настоящего изобретения приводит к образованию малых количеств растворимых в воде пептидов, чем при осуществлении способов, известных из уровня техники. Среди таких водорастворимых пептидов IPP и LPP присутствуют в больших количествах. Это является особенно значимым в случае высоких концентраций трипептидов, ингибирующих АСЕ, которые требуются без многих других, часто менее активных соединений. Согласно настоящему способу предпочтительно не менее 20%, более предпочтительно не менее 30%, наиболее предпочтительно не менее 40% -I-P-P- или -L-Р-Р-последовательности, присутствующей в белке, превращено в трипептид IPP или LPP соответственно. В примере 6 показана пятикратная степень очистки биоактивных пептидов путем новой и удивительной стадии очистки. Основа такого способа очистки формируется на основании уникальных свойств пролин-специфичной эндопротеазы из А. niger. Инкубация с этим ферментом высвобождает наиболее биоактивные части субстратной молекулы в виде водорастворимых трипептидов. Не- или наименее биоактивные части субстратной молекулы сохраняются во многом в нерасщепленном и, следовательно, наиболее крупных пептидных или полипептидных частях субстратных молекул. Вследствие ограниченной водной растворимости этих более крупных пептидных или полипептидных частей при выбранных условиях рН, эти не- или наименее биоактивные части субстратной молекулы без труда отделяются от намного более растворимых биоактивных трипептидов. В этом процессе исходный гидролизат формируется во время короткого ферментативного инкубационного периода при 55 °С, рН 6,0 и затем, необязательно, нагревается до температуры выше 80 °С для уничтожения всех заражающих микроорганизмов и для инактивации полученной из A. niger пролилэндопептидазы. Затем гидролизат подкислялся для достижения снижения рН до 4,5 или по меньшей мере ниже 5,0. При таком значении рН, которое не может использоваться для инактивации полученной из A. niger пролин-эндопептидазы, поскольку является оптимальным условием для фермента, все крупные пептиды из казеинового осадка, таким образом, только более мелкие пептиды остаются в растворе. Как осажденные казеинаты могут быть без труда удалены путем декантации или стадии фильтрации или центрифугирования при низкой скорости (т.е. ниже 5000 rpm), водная фаза содержит высокие пропорции биоактивных пептидов относительно количества присутствующего белка. Согласно данным Kjeldahl от 80 до 70% казеината белка удаляется посредством стадии низкоскоростного центрифугирования, которая предполагает 4-5-кратную очистку АСЕ ингибирующих пептидов. Нами обнаружено, что такой принцип очистки может успешно применяться для получения биологически активных пептидов, полученных из белковоподобного материала, кроме казеина. Также не только ферментативно синтезированные гидролизаты, но и белки, которые сбраживаются пригодными микроорганизмами, могут быть отделены и очищены в соответствии со способом настоящего изобретения. Инкубация фермента и субстрата при значении рН, близком к таковому, при котором субстрат будет осаждаться и фермент будет сохранять активность, будет обеспечивать данную стадию очистки. Благодаря низкому оптимуму рН пролилэндопротеазы, могут рассматриваться субстратные преципитаты в диапазоне значений рН от 1,5 до 6,5. Учитывая специфическую реакцию осаждения, глютен осаждается при рН выше 3,5, белок подсолнечника осаждается при рН выше 4,0 и ниже 6,0, яичный белок осаждается при рН выше 3,5 и ниже рН 5,0, при этом образуются примеры условий, с помощью которых гидролизованные белковые преципитаты и осажденные белки могут отделяться от гидролизованных белков или пептидов. После декантации, фильтрации или низкоскоростного центрифугирования супернатанты, содержащие биологически активные пептиды, могут быть удалены в очищенном состоянии. Последующее испарение и стадия сушки распылением будут образовывать экономичный способ для получения пищевого состава или порошка с высокой биоактивностью. При расщеплении казеинатов в соответствии со способом, который описывается, образуется белый и без запаха порошок с высоким содержанием пептидов, ингибирующих АСЕ. Альтернативно могут применяться выпаривание и нанофильтрация для дальнейшего концентрирования биоактивных пептидов. Правильный состав концентрируется путем увеличения активности воды (Aw) в сочетании с регулированием рН и добавление пищевых консервантов, таких как бензоат или сорбат, приведет к образованию устойчивых к действию микроорганизмов, пищевых жидких концентрированных пептидов, снижающих кровяное давление. Если должным образом разбавить до правильной трипептидной концентрации, универсальный исходный материал получается пригодным для наделения всех видов продуктов питания и напитков свойствами ингибирования АСЕ. При необходимости, супернатант, полученный после декантации, фильтрации или низкоскоростного центрифугирования, может в дальнейшем подвергаться обработке для улучшения вкусовых качеств итогового продукта. Например, супернатант может вступать в реакцию с порошкообразным активированным углем, с последующей стадией фильтрации для удаления угля. Для уменьшения горечи итогового продукта супернатант, полученный после декантации, фильтрации или низкоскоростного центрифугирования, также может использоваться для инкубации с другой протеазой, такой как субтилизин, трипсин, нейтральная протеаза или глутамат-специфичная эндопротеаза. При необходимости, концентрация биоактивных ингредиентов MAP и/или ITP может повышаться в дальнейшем путем последовательных стадий очистки, в которых целью является приготовление специфичного гидрофильного/гидрофобного свойства трипептидов MAP и ITP. Предпочтительные способы очистки включают нанофильтрацию (разделение по размеру), экстракцию, например, гексаном или бутанолом, за которой следует выпаривание/осаждение или взаимодействие подкисленного гидролизата, который получен с хроматографическими смолами из класса Amberlite XAD (Roehm). Также могут применяться бутилсефарозные смолы, которые обеспечиваются Pharmacia. В примере 7 описывается идентификация новых пептидов MAP и ITP, ингибирующих АСЕ, в казеиновом гидролизате, полученных при использовании пролин-специфичной эндопротеазы, полученной из A. niger, в сочетании с новым способом очистки пептидов. Только применение такой одной (фактически чистой) эндопротеазы в сочетании с большим процентом небиоактивных пептидов и высокосложное оборудование для отделения и идентификации позволили нам выявить и идентифицировать эти новые трипептиды, обладающие способностью ингибировать АСЕ. В полученных из казеина биоактивных пептидах (CDBAP), приготовленных по примеру 7 (после осаждения), были идентифицированы трипептиды MAP и ITP в количествах, равняющихся 2,9 мг МАР/г CDBAP (4,8 мг МАР/г белка в CDBAP) и 0,9 мг ITP/г CDBAP (1,4 мг ITP/грамм белка в CDBAP). Кроме того, характеристикой для CDBAP является их чрезвычайно высокое пролиновое содержание в 24% по молярному значению. Тесты, описанные в примере 7, иллюстрируют очень низкие значения IC 50 для двух новых трипептидов в модифицированном тесте Matsui, т.е. 0,5 мкмоль/л для MAP и 10 мкмоль/л для ITP. Эти данные являются еще более удивительными, если мы осознаем, что IPP, один из наиболее эффективных известных натуральных пептидов, ингибирующих АСЕ, имеет значение IC 50 , полученное в модифицированном тесте Matsui, равное 2,0 мкмоль/л. В соответствии с настоящим способом предпочтительно не менее 20%, более предпочтительно не менее 30%, наиболее предпочтительно не менее 40% последовательности -М-А-Р- или -I-T-P-, присутствующей в белке, преобразовывается в трипептид MAP или ITP соответственно. Применимость недавно идентифицированных пептидов MAP и ITP, ингибирующих АСЕ, в дальнейшем иллюстрируется в примере 8. В последнем примере показывается, что оба пептида успешно сохраняются в инкубационных условиях, искусственно воспроизводящих условия пищеварения, типично созданные в желудочно-кишечном тракте. На основании этого мы сделали заключение, что новые трипептиды способны сохраняться в желудочно-кишечном тракте млекопитающего (например, человека), что подразумевает значительный экономический потенциал при применении для лечения гипертензии. В примере 9 продемонстрировано, что лучший АСЕ-ингибирующий пептид MAP может быть синтезирован не только в экспериментах ферментативного гидролиза, но также выявляется в молочных препаратах, подвергнутых ферментации соответствующими пищевыми микроорганизмами. Однако мы не смогли продемонстрировать присутствие пептида ITP в таком продукте, подвергнувшемся ферментации. Пептиды MAP и/или ITP, полученные либо перед, либо после дополнительных, например, стадий хроматографической очистки, могут использоваться для включения в продукты питания, которые широко потребляются регулярно. Примерами таких продуктов являются маргарины, жиры, различные молочные продукты, такие как масло или йогурты либо молоко или напитки, содержащие молочную сыворотку. Несмотря на то что такие композиции обычным образом поступают в организм человека, они также могут применяться для животных, предпочтительно млекопитающих, для облегчения гипертензии. Кроме того, содержание в продуктах ингибиторов АСЕ в высоких концентрациях делает эти продукты пригодными для включения их в диетические добавки в виде пилюль, таблеток, или высококонцентрированных растворов, или паст, или порошков. Медленно высвобождающиеся диетические добавки, которые обеспечат непрерывное высвобождение пептидов, ингибирующих АСЕ, представляют особый интерес. Пептиды MAP и/или ITP согласно изобретению могут иметь форму сухого порошка, например пилюли, таблетки, гранулы, саше или капсулы. Альтернативно, ферменты согласно изобретению могут иметь форму жидкости, например в сиропе или капсуле. Композиции, использующие различные формы и содержания ферментов, согласно изобретению также включают по меньшей мере одно соединение группы, состоящей из физиологически пригодного носителя, адъюванта, наполнителя, стабилизатора, буфера и растворителя, где термины используются в их обычных смыслах для определения субстанций, которые участвуют в сборке, доставке, поглощении, стабилизации или, в случае адъюванта, для повышения физиологического эффекта ферментов. Соответствующие фоновые значения различных соединений, которые могут использоваться в сочетании с ферментами согласно изобретению в порошковых формах, могут быть найдены в "Pharmaceutical Dosage Forms", второе издание, Vol. 1-3, ISBN 0-8247-8044-2 Marcel Dekker, Inc. Несмотря на то что пептиды, ингибирующие АСЕ, согласно изобретению имеющие форму сухого порошка, могут храниться в течение достаточно долгого времени, контакт с увлажнителем или влажным воздухом должен избегаться путем выбора пригодной упаковки, такой как, например, алюминиевый блистер. Относительно новыми формами для перорального применения являются применение различных типов желатиновых капсул или таблеток на желатиновой основе. Принимая во внимание значимость природных пептидов, ингибирующих АСЕ, в борьбе с гипертензией, настоящий новый и экономически эффективный способ обеспечивает привлекательную отправную точку для пищевых и даже ветеринарных продуктов, обладающих умеренно гипотензивным действием. Поскольку настоящий способ также включает удивительно простую стадию очистки, то возможности концентрированных диетических добавок снижать кровяное давление увеличиваются. Под пролин-специфичной эндопротеазой настоящего изобретения или примененной по настоящему изобретению подразумевается полипептид, упомянутый в пп.1-5, 11 и 13 формулы изобретения заявки WO 02/45524. Следовательно, эта пролин-специфичная эндопротеаза представляет собой полипептид, который обладает пролин-специфичной эндопротеолитической активностью, выбранный из группы, состоящей из: (a) полипептида, обладающего аминокислотной последовательностью, которая по крайней мере на 40% идентична аминокислотной последовательности, включающей аминокислоты с 1 по 526 из SEQ ID NO:2 или ее фрагменту; (b) полипептида, кодируемого полинуклеотидом, который гибридизуется в условиях пониженной жесткости с (i) последовательностью нуклеиновых кислот SEQ ID NO:1 или ее фрагментом, который идентичен не менее чем на 80 или 90% на участке более 60, предпочтительно более 100 нуклеотидов, более предпочтительно идентичен не менее чем на 90% на участке более 200 нуклеотидов, или (ii) последовательностью нуклеиновых кислот, комплементарной последовательности нуклеиновых кислот SEQ ID NO:1. Последовательности SEQ ID NO:1 и SEQ ID NO:2 соответствуют приведенным в WO 02/45524. Полипептид предпочтительно присутствует в изолированной форме. Предпочтительный полипептид, используемый по настоящему изобретению, имеет аминокислотную последовательность, которая не менее чем на 50%, предпочтительно не менее чем на 60%, предпочтительно не менее чем на 65%, предпочтительно не менее чем на 70%, более предпочтительно не менее чем на 80%, еще более предпочтительно не менее чем на 90%, наиболее предпочтительно не менее чем на 95% и даже еще более предпочтительно не менее чем приблизительно на 97% идентична аминокислотам с 1 по 526 из SEQ ID NO:2 или содержащей аминокислотную последовательность SEQ ID NO:2. Полипептид предпочтительно кодируется полинуклеотидом, который гибридизуется в условиях пониженной жесткости, более предпочтительно в условиях средней жесткости и наиболее предпочтительно в жестких условиях с (i) последовательностью нуклеиновых кислот SEQ ID NO:1 или ее фрагментом или (ii) последовательностью нуклеиновых кислот, которая комплементарна последовательности нуклеиновых кислот SEQ ID NO:1. Термин «способный гибридизоваться » означает, что целевой полинуклеотид по настоящему изобретению может гибридизоваться с нуклеиновой кислотой, используемой в качестве зонда (например, с последовательностью нуклеотидов, приведенной в SEQ ID NO:1, или ее фрагментом или комплементом SEQ ID NO:1) на уровне, значительно превосходящем фоновый. Изобретение также включает полинуклеотиды, которые кодируют пролин-специфичную эндопротеазу по настоящему изобретению, а также нуклеотидную последовательность, ей комплементарную. Нуклеотидная последовательность может представлять собой РНК или ДНК, включая геномную ДНК, синтетическую ДНК или кДНК. Предпочтительно нуклеотидная последовательность представляет собой ДНК и наиболее предпочтительно последовательность геномной ДНК. Как правило, полинуклеотид по настоящему изобретению содержит непрерывную последовательность нуклеотидов, которая способна селективно гибридизоваться с кодирующей последовательностью или комплементом кодирующей последовательности SEQ ID NO:1. Такие нуклеотиды могут быть синтезированы хорошо известными из уровня техники способами. Полинуклеотид по настоящему изобретению может гибридизоваться с кодирующей последовательностью или комплементом кодирующей последовательности SEQ ID NO:1 на уровне значительно выше фонового. Фоновая гибридизация может иметь место, например, из-за того, что другая кДНК представлена в библиотеке кДНК. Уровень сигнала, генерируемого взаимодействием между полинуклеотидом по настоящему изобретению и кодирующей последовательностью или комплементом кодирующей последовательности SEQ ID NO:1, как правило, не менее чем в 10 раз, предпочтительно не менее чем в 20 раз, более предпочтительно не менее чем в 50 раз и еще более предпочтительно не менее чем в 100 раз превосходит по интенсивности уровень взаимодействия между другими полинуклеотидами и кодирующей последовательностью SEQ ID NO:1. Интенсивность взаимодействия может быть измерена, например, внесением в зонд радиоактивной метки, например 32Р. Селективная гибридизация, как правило, может быть достигнута при использовании условий пониженной жесткости (0,3 М хлорид натрия и 0,03 М цитрат натрия при температуре около 40 °С), условий средней жесткости (например, 0,3 М хлорид натрия и 0,03 М цитрат натрия при температуре около 50 °С) и жестких условий (например, 0,3 М хлорид натрия и 0,03 М цитрат натрия при температуре около 60 °С). Пакет UWGCG предоставляет программу BESTFIT, которая может применяться для вычисления идентичности (например, используя ее установки по умолчанию). Алгоритмы PILEUP и BLAST N также могут быть использованы для вычисления идентичности последовательностей или для сравнения последовательностей (как, например, для идентификации эквивалентных или аналогичных последовательностей, например при установках по умолчанию). Программное обеспечение для осуществления анализа BLAST доступно всем желающим через Национальный центр биотехнологической информации (http://www.ncbi.nlm.nih.gov/). Этот алгоритм включает, во-первых, идентификацию пар последовательностей с высокой степенью соответствия (HSPs) путем идентификации коротких наборов символов длины W в запрашиваемой последовательности, которые либо совпадают, либо удовлетворяют определенному положительному пороговому значению Т при сравнении с набором символов той же длины в базе данных последовательностей. Т называют пороговым значением близости набора символов. Эти первоначальные совпадения близких наборов символов действуют в качестве отправной точки для инициирования поиска с целью обнаружения содержащих их HSPs. Совпадения наборов символов расширяются в обоих направлениях вдоль каждой последовательности до тех пор, пока может быть увеличено общее количество соответствий. Расширение в каждом из направлений для поиска совпадения набора символов прекращается в случае, если: общее количество соответствий уменьшается на число X по сравнению с максимальным достигнутым значением; общий счет соответствий доходит до нуля или ниже за счет накопления одной или нескольких групп остатков, вносящих отрицательный счет; или если достигнут конец любой из последовательностей. Параметры алгоритма BLAST W, T и X определяют чувствительность и скорость сверки. В программе BLAST по умолчанию используется длина набора символов (W), равная 11, матрица подсчета соответствий BLOSUM62 (В), равная 50, ожидание (Е), равное 10, М=5, N=4 и сравнение обеих цепей. Алгоритм BLAST осуществляет статистический анализ подобия между двумя последовательностями. Одной из мер подобия, предоставляемых алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), показывающая вероятность, с которой совпадение между двумя последовательностями нуклеотидов или аминокислот может появиться случайно. Например, считается, что последовательность подобна другой последовательности, если наименьшая суммарная вероятность при сравнении первой последовательности со второй последовательностью менее чем приблизительно 1, предпочтительно менее чем приблизительно 0,1, более предпочтительно менее чем приблизительно 0,01 и наиболее предпочтительно менее чем приблизительно 0,001. Штаммы рода Aspergillus имеют статус пригодных для употребления в пищу, причем известно, что ферменты, выделенные из этих микроорганизмов, получены из источника, несомненно пригодного для употребления в пищу. В соответствии с другим предпочтительным вариантом осуществления изобретения фермент секретируется продуцирующей его клеткой, а не является не секретируемым, так называемым цитозольным или периплазматическим ферментом. Таким образом, ферменты могут быть повторно получены из клеточного бульона в основном в чистом состоянии, без дорогостоящих стадий очистки. Фермент предпочтительно имеет высокое сродство к своему субстрату при широко распространенных значениях рН и температурных условиях. Фиг. 1 - графическое представление оптимального значения рН пролин-специфичной эндопротеазы, выделенной из A. niger. Фиг. 2 - профильная специфичность пролин-специфичной эндопротеазы, выделенной из A. niger. Фиг. 3 - SDS-PAGE интактного овальбумина и синтетического 27-мерного пептида после инкубации с очищенной хроматографией пролин-специфичной эндопротеазой, выделенной из A. niger. Материалы. Пригодный к употреблению в пищу раствор казеината калия (88%) был получен от DMV International, The Netherlands. Синтетические хромогенные пептиды были получены либо от Pepscan Systems B.V. The Netherlands, либо от Bachem, Switzerland. Пролин-специфичная эндопротеаза из A. Niger. Сверхсинтез пролин-специфичной эндопротеазы из Aspergillus niger проводили, как описано в WO 02/45524. Активность фермента испытывали на синтетическом пептиде Z-Glu-Pro-pNA при 37 °С в буфере цитрат/динатрийфосфат рН 4,6. За продуктом реакции наблюдали спектрофотометрически при 405 нм. Единица определена как количество фермента, которое способствует выделению 1 мкмоль п-нитроанилида в минуту при указанных тестовых условиях. Хроматографическая очистка эндопротеазы, полученной из A. Niger. Культуральный бульон, полученный из сверхсинтезирующего штамма A. niger, использовался для хроматографической очистки протеазы для удаления любых контаминирующих эндо- и экзопротеолитических активностей. Для этого ферментационный бульон был сначала центрифугирован для удаления основной грибковой массы и супернатант был в дальнейшем пропущен через ряд фильтров с уменьшающимися размерами пор для удаления всех клеточных фрагментов. В итоге, полученный ультрафильтрат разбавлялся десятикратно в 20 ммоль/л ацетате натрия с рН 5,1 и вносился в колонку Q-Sepharose FF. Белки извлекались в градиенте от 0 до 0,4 моль/л NaCl в 20 ммоль/л ацетате натрия с рН 5,1. Пиковые фракции, проявляющие активность по отношению к расщеплению Z-Gly-Pro-pNA, были собраны и объединены, в соответствии с протоколом, описанным в World Journal of Microbiology & Biotechnology 11, 209-212 (1995), но при незначительно модифицированных условиях анализа. Учитывая кислотный оптимум рН пролин-специфичной эндопротеазы, полученной из А. niger, ферментативный анализ осуществлялся при рН 4,6 в цитрат/дифосфатном буфере при 37 °С. Объединяя активные фракции, с последующим концентрированием, полученный в итоге препарат показывал только простую полосу на SDS-PAGE и один пик на HP-SEC. Дальнейший анализ путем гидрофобной хроматографии подтвердил чистоту полученного ферментного препарата. Анализ ЖХ/МС/МС, применяемый для определения IPP, LPP и VPP. Для количественного определения в белковом ферментативном гидролизате представляющих интерес пептидов, в числе которых IPP, LPP и VPP, полученных при действии смеси ферментов по настоящему изобретению, применяли ВЭЖХ, использующую масс-спектрометр с ионной ловушкой (Thermoquest ®, Breda, the Netherlands), соединенный с насосом Р4000 (Thermoquest ®, Breda, the Netherlands). Полученные пептиды разделяли при использовании колонки lnertsil 3 ODS 3,3 мкм, 150 ×2,1 мм (Varian Belgium, Belgium) в сочетании с градиентом 0,1% муравьиной кислоты в Milli Q water (Millipore, Bedford, MA, USA; раствор А) и 0,1% муравьиной кислоты в ацетонитриле (раствор В) для элюирования. Градиент начинали с 100% раствора А, сохраняя в течение 5 мин, увеличивая линейно до 5% раствора В в течение 10 мин, с последующим линейным увеличением до 45% раствора В в течение 30 мин и немедленно возвращаясь к начальным условиям, и сохраняя в течение 15 мин для стабилизации. Вводимый объем составлял 50 мкл, скорость потока равнялась 200 мкл/мин и температуру колонки поддерживали на уровне 55 °С. Концентрация белка во вводимом образце составляла приблизительно 50 мкг/мл. Подробную информацию об отдельных пептидах получали, применяя специальный МС/МС алгоритм для пептидов, представляющих интерес, при использовании оптимальной энергии столкновения около 30%. Количественное определение отдельных пептидов осуществлялось, используя внешнюю калибровку, путем применения наиболее многочисленных заряженных фрагментов, наблюдаемых в МС/МС методе. Трипептид LPP (М=325,2) использовался для регулировки оптимальной чувствительности алгоритма МС и оптимальной фрагментации в способе МС/МС, выполняя постоянное введение 5 мкг/мл, что приводит к образованию протонированной молекулы в МС алгоритме и оптимальной энергии столкновения около 30% в МС/МС алгоритме, образуя В- и Y-ионные серии. До проведения ЖХ/МС/МС ферментные белковые гидролизаты центрифугировались при комнатной температуре и 13000 rpm в течение 10 мин, фильтровались через 0,22 мкм фильтр и супернатант разбавлялся 1:100 MilliQ водой. Определение азота по Kjeldahl. Содержание общего азота способом Kjeldahl измерялось путем анализа методом впрыскивания в поток. Используя систему Tecator FIASTAR 5000, оснащенную кассетным методом TKN 5000-040, компьютером Pentium 4 с программным обеспечением SOFIA и автодозатором Tecator 5027, аммоний, высвобождающийся из раствора, содержащего белки, определялся при длине волны 590 нм. Количество образца, соответствующего динамическому диапазону способа (0,5-20 мг N/л), помещалось в пищеварительную трубку вместе с 95-97% серной кислотой и таблетки Kjeltab подвергались действию пищеварительной программы в течение 30 мин при 200 °С, с последующим воздействием в течение 90 мин при 360 °С. После введения в систему FIASTAR 5000 измерялся пик азота, по которому можно судить о количестве измеренного белка. Аминокислотный анализ. Точно взвешенный образец белковоподобного материала растворялся в разбавленной кислоте и осадок удалялся путем центрифугирования в центрифуге Eppendorf. Аминокислотный анализ выполнялся на чистом супернатанте в соответствии со способом PicoTag, как определено в руководстве по эксплуатации системы для аминокислотного анализа Waters (Milford MA, USA). Для этого пригодный образец был получен из жидкости, затем высушен и подвергнут паровой фазе кислотного гидролиза и дериватизирован, используя фениловый эфир изотиоциановой кислоты. Содержание различных дериватизированных аминокислот определялось, используя способы ВЭЖХ, и суммировалось для определения общего уровня свободных аминокислот во взвешенном образце. Аминокислоты Cys и Tip не включались в результаты, полученные при этом анализе. Степень гидролиза (DH) белкового гидролизата измерялась при использовании теста ОРА и рассчитывалась, как описано Nielsen et al. (JFS, Vol. 66, № 5, 642-646, 2001). Расчет молекулярной массы пептидов и белков, присутствующих в гидролизатах. Анализ распределения размера частиц протеазы, обработанной белковыми образцами, осуществлялось на автоматизированной системе ВЭЖХ, оснащенной насосом высокого давления, устройством введения, способным вводить 10-100 мкл образца, и УФ-детектором, способным контролировать элюент при 214 нм. Колонкой, используемой для этого анализа, была колонка Superdex Peptide HR 10/300 GL (Amersham), уравновешенная 20 мМ фосфатом натрия/250 мМ хлоридом натрия буфером с рН 7,0. После введения образца (обычно 50 мкл) различные компоненты извлекались из колонки буфером в течение 90 мин при скорости потока 0,5 мл/мин. Система калибровалась, используя смесь цитохрома С (молекулярная масса 13500 Да), апротинина (молекулярная масса 6510 Да) и тетраглицина (молекулярная масса 246 Да) в качестве маркеров молекулярной массы. Пример 1. Фермент, полученный из A. niger, представляет собой новый класс пролин-специфичных ферментов. Из всей кодирующей последовательности пролин-специфичной эндопротеазы, получаемой из A. niger, которая описана в WO 02/45524, может быть определена белковая последовательность из 526 аминокислот. Новизна фермента подтверждена с помощью баз данных для поиска BLAST, таких как SwissProt, PIR и trEMBL. К удивлению авторов не удалось обнаружить явной гомологии между ферментом A. niger и известными пролилолигопептидазами. Более тщательное исследование аминокислотной последовательности выявило, однако, низкую, но значимую, степень гомологии с карбоксипептидазами Pro-Х (ЕС3.4.16.2), дипептидиламинопептидазами I (EC3.4.14.2) и специфичной для тимуса серинпротеазой. Все эти ферменты отнесены к семейству серинпептидаз S28. Также, у всех этих ферментов и эндопротеазы A. niger сохранена конфигурация GxSYxG вокруг активного сайта серина. Кроме того, члены семейства S28 имеют кислотный рН-оптимум, обладают специфичностью в отношении расщепления с карбоксиконцевой стороны пролиновых остатков и синтезируются с сигнальной последовательностью и пропептидом, точно так же как пролин-специфичная эндопротеаза, вырабатываемая A. niger. Также размер фермента A. niger сходен с размером членов семейства S28. Следовательно, пролин-специфичная эндопротеаза А. niger, по-видимому, является членом семейства серинпротеаз S28, а не семейства S9, в котором сгруппировано большинство цитозольных пролилолигопептидаз, включая фермент, выделенный из Flavobacterium meningosepticum. На основе этих структурных и физиологических особенностей мы пришли к выводу, что фермент A. niger принадлежит к семейству S28, а не к семейству серинпротеаз S9. Еще одной особенностью, которая отличает фермент A. niger от пролилолигопептидаз, принадлежащих к семейству S9, является то, что в отличие от цитозольных пролилэндопротеаз, принадлежащих к последнему семейству, недавно идентифицированный фермент A. niger секретируется в ростовую среду. Настоящая заявка является первым сообщением о выделении члена семейства S28 из низших эукариотов и описанием его характеристик. Пример 2. Оптимальные значения рН и температуры пролин-специфичной эндопротеазы, полученной из A. niger. Для установления оптимального значения рН пролин-специфичной эндопротеазы, вырабатываемой A. niger, приготавливали буферные растворы с различными значениями рН. Буферные растворы с рН 4,0-4,5-4,8-5,0-5,5 и 6,0 получали, используя 0,05 моль/л ацетата Na и 0,02М CaCl 2 ; буферные растворы с рН 7,0 и 8,0 получали, используя 0,05 М буферы Tris/HCl, содержащие 0,02 М CaCl 2 . Значения рН регулировали, используя уксусную кислоту и HCl соответственно. В качестве субстрата использовали хромогенный синтетический пептид Z-Glu-Pro-pNA. Буферный раствор, раствор субстрата и предварительно разбавленную эндопротеазу (с активностью 0,1 Ед/мл) нагревали точно до температуры 37,0 °С на водяной бане. После перемешивания за протеканием реакции наблюдали спектрофотометрически при длине волны 405 нм и температуре 37,0 °С в течение 3,5 мин, осуществляя измерения каждые 0,5 мин. Из результатов, показанных на фиг. 1, ясно, что пролин-специфичная эндопротеаза, вырабатываемая A. niger, имеет оптимальное значение рН около 4. Кроме этого, устанавливали температурный оптимум пролилэндопротеазы. С этой целью очищенные препараты фермента инкубировали в 0,1 моль/л ацетате Na, содержащем 0,02 моль/л CaCl 2 при рН 5,0 в течение 2 ч при различных температурах, используя в качестве субстрата Казеин Резоруфин (Roche version 3), и количественно определяли ферментную активность путем измерения при длине волны 574 нм. Согласно полученным результатам, пролин-специфичная эндопротеаза из A. niger имеет температурный оптимум около 50 °С. Пример 3. Специфичность и чистота пролин-специфичной протеазы, полученной из A. niger. Исходные, так же как и хроматографически очищенные ферментные образцы, полученные из штамма A. niger, содержащие множественные копии полигенного экспрессирующего кластера (WO 02/45524), были протестированы против набора хромогенных пептидных субстратов для определения специфичности кодированной эндопротеазы. Используя различные наборы хромогенных пептидов, определялось наличие возможного загрязнения ферментативной активности. Эндопротеолитическая специфичность кодированной эндопротеазы тестировалась на различных Ala-Ala-XpNA (AAXpNA) субстратах. В данном контексте X относится к различным аминокислотным остаткам и pNA - к р-нитроанилиду. Расщепление пептидной связи между X остатком и pNA части молекулы вызывает изменение цвета, что может контролироваться путем усиления светового поглощения при длине волны = 405 или 410 нм AAXpNA субстрат. Для этого исходные растворы AAX-pNA субстратов (150 ммоль/л) разбавлялись в 100 раз в 0,1 М ацетатном буфере с рН 4,0, содержащем 20 CaCl 2 . 10-минутные измерения кинетики на TECAN Genios MTP Reader (Salzburg, Vienna) при длине волны 405 нм записывали увеличения оптической плотности, что посредством данных, обработанных в Excel, получили диаграмму, показанную на фиг. 2. Из результата наглядно видно, что эндопротеаза из A. niger является высоко специфичной к пролиловым пептидным связям с концевой активностью по отношению к аланиловым связям. Исходные и хроматографически очищенные препараты демонстрировали аналогичные профили активности. Возможное загрязнение эндопротеазы, полученной из A. niger, чужеродными ферментативными активностями приводит к ряду нежелательных побочных реакций. Например, наличие экзопротеаз, таких как карбоксипептидаз или аминопептидаз, приводит к образованию пептидных препаратов, содержащих повышенный уровень свободных аминокислот. Эти экстрасвободные аминокислоты разбавляют относительные концентрации содержащихся биоактивных пептидов и, более того, придают хлебные вкусы в результате усиленных реакций Maillard. Представлены вопросы, должны ли значительные уровни загрязненных эндопротеаз присутствовать в препарате сверхэкспрессированной пролин-специфичной эндопротеазы, сайты расщепления в других положениях, чем карбоксиконцевые пролиновые или аланиновые остатки. Такие экстрасайты расщепления генерируют экстрапептиды, при этом также разбавляя содержание биоактивных пептидов, имеющих карбоксиконцевые пролиновые остатки. Для снижения до минимума всех этих нежелательных побочных реакций предпочтительным является применение практически чистых пролин-специфичных протеаз. Под понятием практически чистые понимается, что активности загрязненных эндопротеаз, так же как и загрязненных экзопептидаз, при инкубационных условиях используются в минимальных количествах или предпочтительно отсутствуют. Следующая тестовая процедура была разработана для количественного определения таких загрязненных экзо- и эндопептидазных активностей. Основа для тестовой процедуры формировалась путем сбора различных избирательных хромогенных пептидов. Поскольку только пролин-специфичные олиго- и эндопротеазы могут удалить pNA из пептида Z-AAAP-pNA, то этот специфический пептид применялся для количественного определения желаемой пролин-специфичной эндопротеолитической активности. Поскольку многие эндопротеазы могут удалить pNA из пептидов Z-AAAF-pNA и Z-AAAR-pNA, эти два пептида применялись для определения загрязненной, не-пролин-специфичной эндопротеолитической активности. Так как многие аминопептидазы могут эффективно удалить Phe и Gln из пептидных субстратов, хромогенные пептиды Q-pNA и V-pNA применялись для определения загрязненных аминопептидазных активностей. Поскольку многие карбоксипептидазы могут удалить Phe- и Arg-остатки из пептидных субстратов, пептиды, содержащие эти остатки, были выбраны для определения загрязненных карбоксипептидазных активностей. Однако не существует пригодных хромогенных групп, пригодных для количественного определения карбоксипептидазных активностей, в связи с чем был разработан альтернативный способ при использовании синтетических пептидов Z-AF и Z-AR. Этот альтернативный способ приводится ниже. Во всех синтетических пептидах используемый Z представляет бензилоксикарбонил и pNA - хромофорный паранитроанилид. Все хромогенные пептиды были получены от Pepscan (Lelystad, The Netherlands). Пептиды Z-AF и Z-AR были куплены у Bachem (Switserland). Все инкубации производились при 40 °С. Разбавленные ферментные препараты были пересчитаны в соответствии с концентрацией коммерческого продукта. Для демонстрации уровней различных ферментативных активностей, которые обычно присутствуют в промышленно доступных ферментных препаратах, в данном примере нами описывается тестирование трех коммерческих ферментных препаратов на их различные протеолитические активности, а именно Flavourzyme 1000L Batch HPN00218 (Novozymes), Sumizyme FP (Shin Nihon, Japan) и Corolase LAP Ch.: 4123 (AB Enzymes, UK). Известно, что оба Flavourzyme и Sumizyme FP являются комплексными ферментными препаратами, которые содержат несколько аминопептидолитических ферментных активностей кроме неспецифичных эндопротеолитических и карбоксипептидолитических активностей. Corolase LAP представляет относительно чистую, клональную и сверхэкспрессированную лейцин-аминопептидазную активность из Aspergillus. Измерение аминопептидазных активностей. Исходные растворы 150 ммоль/л F-pNA и Q-pNA в 100% DMSO были разбавлены в 80 раз в 0,1 М BisTris буфере с рН 6 для приготовления 3,75 ммоль/л F-pNA+ Q-pNA-субстратного раствора, содержащего F-pNA и Q-pNA в соотношении 1:1. 200 мкл аликвоты из этого аминопептидазного субстратного раствора были распипетировано в отдельные лунки микротитровой планшеты (МТР). МТР предварительно инкубировали при 40 °С в Tecan Genios МТР (Salzburg, Vienna) под управлением программного обеспечения Magellan4. Реакция начиналась путем добавления 50 мкл подходящего ферментного раствора, таким образом, инкубации проводились при концентрации субстрата 3 мМ. Обычно, применялось разбавление 1:50 жидких ферментных образцов Flavourzyme, Corolase LAP и пролин-специфичной эндопротеазы. Был использован 1% раствор сухого продукта Sumizyme FP. Развитие желтой окраски, измеренной при длине волны 405 нм с помощью Tecan Genios МТР, как результат расщепления связи аминокислота-pNA, следовало по меньшей мере через 20 кинетических циклов (около 10 мин). Программное обеспечение генерировало полученные данные как OD 405 /мин (OD - оптическая плотность). Измерение пролин-специфичной эндопротеазной активности. Это измерение проводилось практически аналогично измерению аминопептидазной активности, но в данном случае Z-AAAP-pNA применялся только в качестве субстрата в итоговой концентрации 3 ммоль/л. Этот субстрат был растворен путем нагревания суспензии в буфере с рН 6 до 50-55 °С, что приводило к образованию прозрачного раствора при комнатной температуре. Измерения проводились при 40 °С. Обычно применялось разбавление 1:50 жидких ферментных образцов Flavourzyme и Corolase LAP. Sumizyme FP применялся как 1% раствор. Пролин-специфичная эндопротеаза обычно использовалась в разведении 1:5000. Программное обеспечение генерировало данные как OD 405 /мин. Измерение загрязнения не-пролин-специфичных эндопротеазных активностей. Также эти измерения проводились практически аналогичным образом, как описано для анализа аминопептидазной активности, но в данном тесте в качестве субстрата использовались Z-AAAF-pNA и Z-AAAR-pNA в соотношении 1:1 и в конечной концентрации 3 ммоль/л. Субстрат Z-AAAF-pNA оказался плохорастворимым при тестовых условиях при рН 6,0, но тестовая инкубация с субтилизином привела к быстрому растворению субстрата одновременно с высвобождением pNA. Измерения проводились при 40 °С. Однако для компенсации такой плохой растворимости устройство для чтения МТР было запрограммировано на встряхивание между кинетическими циклами. Аналогично, программное обеспечение генерировало данные как OD 405 /мин. Измерение загрязнения карбоксипептидазных активностей. Поскольку не существует чувствительных хромогенных пептидов, пригодных для измерения карбоксипептидазных активностей, используемый способ основан на протоколе Boehringer для измерения карбоксипептидазы С. Два исходных раствора Z-A-F и Z-A-R в этаноле по 150 ммоль/л были разбавлены в 80 раз в 0,1 моль/л BisTris буфере с рН 6 для приготовления 3,75 ммоль/л Z-A-F+Z-A-R субстратного раствора, содержащего Z-A-F и Z-A-R в соотношении 1:1. Затем 200 мкл субстратного раствора было раскапано в эппендорфную пробирку и предварительно инкубировано при 40 °С. Реакция начиналась путем добавления 50 мкл подходящего ферментного раствора. Обычно применялось разбавление 1:50 образцов Flavourzyme и Corolase LAP и пролин-специфичной эндопротеазы. 1% раствор использовался для Sumizym FP. Через 5 мин реакция останавливалась добавлением 250 мкл нингидринового реагента. Нингидриновый реагент был приготовлен из 400 мг нингидрина (Merck) и 60 мг гидриндантина, растворенных в 15 мл DMSO, к которым добавляли 5 мл 4,0 моль/л буфера ацетата лития с рН 5,2. 4,0 моль/л буфера ацетата лития приготавливали путем растворения LiOH (Sigma), после чего рН раствора доводили до рН 5,2, используя безводную уксусную кислоту (Merck). После остановки реакции каждый образец нагревался в течение 15 мин при 95 °С для обеспечения формирования окраски, и его последовательно разбавляли в 10 раз чистым этанолом. Образованную окраску измеряли при 578 нм в спектрофотометре Uvikon. Контрольные пробы приготавливались таким же образом, как образцы для измерения активности, но нингидриновый реагент и фермент не добавлялись. Для определения количества свободных аминокислот, образованных карбоксипептидазной активностью, использовалась аминокислота L-фенилаланин для создания калибровочной кривой. Растворы в буфере с рН 6, содержащие 0,1875, 0,375, 0,75, 1,5 и 3,0 ммоль/л L-фенилаланин (Sigma), обрабатывались аналогично образцам, т.е. по 250 мкл на пробирку. Из полученных значений OD578 была построена кривая в Excel. Концентрации аминокислот, находящихся в образцах, содержащих субстраты Z-A-F и Z-A-R, были рассчитаны при использовании кривой. Из полученных значений была рассчитана карбоксипептидазная активность в микромолях на минуту на количество тестируемого фермента. Вычисление индексов активности. Для установления пригодности различных ферментных препаратов для способа изобретения индексы значимых ферментативных активностей были вычислены. Анализ, основанный на показаниях устройства для чтения МТР, показал, что ферментативные активности характеризуются высвобождением pNA в динамике по времени, а именно как ΔOD 405 /мин. Индексы ферментативных активностей, полученных при использовании устройства для чтения МТР, рассчитывались путем простого деления значений ΔOD/мин, полученных аналогичных индексов фермента. Однако в случае карбоксипептидазного анализа OD генерируется, при этом не может сравниваться непосредственно с ΔOD/мин, полученным анализом MTP-pNA. В данном случае измеренную OD сначала конвертировали в мкмоль аминокислоты, высвобожденной за минуту (мкмоль/мин). Затем ΔOD/мин высвобожденного pNA конвертировалось в мкмоль/мин. Для этого калибровочная кривая была получена по показаниям устройства для чтения МТР, где измерялись разбавления чистого pNA (Sigma) 0,25, 0,125, 0,625, 0,0312 и 0,015 ммоль/л и 250 мкл на лунку. Из полученных данных была построена калибровочная кривая в Excel. Из данной калибровочной кривой ΔOD/мин переводился мкмоль/мин, таким образом, измерения, основанные на pNA, могли сравниваться с измерениями, основанными на нингидрине. На основании полученных данных вышеуказанных тестов различные используемые ферментные препараты были охарактеризованы в виде желаемых пролин- и аланин-специфичных активностей и загрязненных эндопротеазных, аминопептидазных и карбоксипептидазных активностей. Данные на пролин-специфичные олиго- или эндопротеолитические активности, представленные для каждого ферментного препарата, показаны в табл. 1 в графе "Прол-спецактивности". Данные на загрязненные аминопептидазные активности (АР/Прол-спецакт), загрязненные карбоксипептидазные аминопептидазные активности (CPD/Прол-спецакт) и загрязненные эндопротеолитические активности (Endo/Прол-спецакт) показаны в соотношении с настоящими пролин-специфичными активностями. Уровень загрязненной аминопептидазной активности по отношению к загрязненной карбоксипептидазной активности, присутствующей в каждом препарате, показан как (AP/CPD). Очевидно, что ни один из коммерческих ферментных препаратов не содержит какие-либо значительные пролин-специфичные олиго- или эндопротеолитические активности. Более того, все тестируемые коммерческие ферментные препараты содержат значительные загрязненные эндо- и экзопротеолитические активности. Таблица 1 * Sumizyme измерялся в 1% растворе, Flavourzyme и Corolase как разведение 1:50. Прол-специфичная активность, полученная из A. niger, измерялась как разведение 1:5000. Затем данные рассчитывались для определения присутствующей активности в представленном продукте. Пример 4. Пролин-специфичная эндопротеаза, полученная из A. niger, способна гидролизовать крупные белки, также как и небольшие пептиды и, таким образом, является настоящей эндопротеазой. Благодаря характерной особенности структуры, пролилолигопептидазы, принадлежащие к семейству S9, не могут разлагать пептиды размером свыше 30 аминокислотных остатков. Это ограничение является очевидным недостатком для фермента, который, как подразумевается, должен гидролизовать различные белки так быстро и эффективно, как только возможно. Для того чтобы выяснить, характерны ли эти ограничения, относящиеся к молекуле субстрата, для пролин-специфичной эндопротеазы, полученной из A. niger, авторы инкубировали хроматографически очищенную пролилэндопротеазу из A. niger с небольшим синтетическим пептидом, а также с крупной молекулой овальбумина и подвергли образовавшиеся продукты гидролиза анализу с помощью SDC-PAGE. Используемый синтетический пептид представлял собой 27-мерный пептид, включающий последовательность NH 2 -FRASDNDRVIDPGKVETLTIRRLHIPR-COOH, и был подарен компанией Perscan (Lelystad, The Netherlands). Как видно из его аминокислотной последовательности, этот пептид содержит 2 пролиновых остатка, один в середине и один возле самого конца пептида. Интактная молекула овальбумина (Pierce Imject, пузырьки, содержащие 20 мг замороженного высушенного вещества) состоит из 385 аминокислот и имеет молекулярную массу 42750 Да. Эта молекула содержит 14 пролиновых остатков, один из которых расположен в самой С-концевой части молекулы и его нельзя расщепить пролин-специфичной эндопротеазой. Овальбумин и олигопептид по отдельности инкубировали при 50 °С с очищенной пролин-специфичной эндопротеазой, полученной из A. niger. Через определенные отрезки времени отбирали несколько образцов, которые анализировали, применяя SDS-PAGE. Хроматографически очищенную пролин-специфичную эндопротеазу, полученную из A. niger, с активностью 4,5 Ед/мл разбавляли в 100 раз 0,1М ацетатным буфером с рН 4, содержащим 20 мМ CaCg 2 . Овальбумин растворяли в ацетатном буфере с рН 4 до концентрации 1 мг/мл (22 мкМ). 27-мерный пептид растворяли в таком же буфере до достижения концентрации 0,48 мг/мл (152 мкМ). Молярности растворов овальбумина и 27-мера выбирали таким образом, чтобы оба раствора имели одинаковую молярность по отношению к расщепляемым пролиновым остаткам. Овальбумин содержит 13 потенциальных сайта расщепления пролина, тогда как 27-мерный пептид имеет только два. 0,5 мл растворов обоих субстратов инкубировали с 10 мкл (0,45 мЕд) раствора фермента в термомиксере Эппендорфа при 50 °С. Через определенные интервалы времени отбирали 10 мкл образцов инкубируемых смесей и до проведения SDS-PAGE хранили при 20 °С. Все материалы, использованные для SDS-PAGE и окрашивания, были приобретены у Invitrogen. Образцы получали, используя буфер LDS в соответствии с инструкциями производителя, и разделяли на 12% бис-трис-гелях, используя буферную систему MES-SDS согласно инструкциям производителя. Окрашивание выполняли, используя краситель Simply Blue Safe Stain (Collodial Coomassie G250). Как можно видеть на фиг. 3, овальбумин расщепляется ферментом, полученным из Aspergillus, на дискретную линию, приблизительно 35-36 КДа, в первые 4,75 ч инкубирования (полоса 3). Увеличение продолжительности инкубирования приводит к дальнейшему распаду на более мелкие продукты различной молекулярной массы (полоса 7). 27-мерный пептид также расщепляется, как можно судить по более слабым линиям в полосах 4, 6 и 8 по сравнению с полосой 2. Сдвиг, соответствующий очень малой молекулярной массе продукта (ср. полосы 9 и 8), вероятнее всего, вызван отщеплением аргининового остатка от карбоксильного конца пептида. Разница составляет приблизительно 200 Да (измерена с использованием программного обеспечения Alphalmager 3.3d на системе Alphalmager 2000), и аргинин обладает молекулярной массой 174. Этот сдвиг, соответствующий малой молекулярной массе, вероятно, соответствует первой стадии в расщеплении пептида. Дальнейший распад продукта может быть виден за счет уменьшения интенсивности линии на геле SDS. Продукты дальнейшего распада невидимы, так как окрашивание в геле красителем кумасси бриллиантовый голубой компонентов с молекулярной массой около 1000 невозможно. Из этого эксперимента можно сделать вывод, что в отличие от известных пролилолигопептидаз, принадлежащих к семейству S9, пролин-специфичная эндопротеаза, выделенная из A. niger, не имеет конкретного предпочтения в отношении расщепления пептидов небольшого размера по сравнению со значительно более крупными белками. В силу этого выделенный из A. niger фермент представляет собой настоящую эндопротеазу и является предпочтительным ферментом для гидролиза различных видов белков. Это наблюдение привело к неожиданному применению фермента, что показано в следующем примере. Пример 5. При инкубации казеината калия с пролин-специфичной эндопротеазой из A. niger быстро выделяются IPP и LPP, но не VPP. В этом эксперименте сверхпродуцирующая и практически чистая пролин-специфичная эндопротеаза из A. niger инкубировалась с казеинатом калия для тестирования выделения пептидов IPP и VPP, ингибирующих АСЕ, а также LPP. Используемая эндопротеаза была практически чистой, это означает, что не наблюдалось значимой эндопротеолитической активности, за исключением эндопротеолитической активности, свойственной чистой пролин-специфичной эндопротеазе (т.е. карбоксиконцевое расщепление пролиновых и аланиновых остатков). Кроме того, ферментный препарат лишен значимых экзопротеолитических активностей, как описывается в примере 3. Для ограничения поступления натрия в результате применения пептидов, ингибирующих АСЕ, насколько возможно, казеинат калия использовался в качестве субстрата при данной инкубации. Казеинат суспендировался в воде при 65 °С в концентрации 10% (м/м) белка, после чего рН доводилось до 6,0 при использовании фосфорной кислоты. Затем суспензия охлаждалась до 55 °С и добавлялась пролин-специфичная эндопротеаза, полученная из А. niger, в концентрации 4 ед./г белка (см. раздел «Материалы и методы для определения единиц »). При непрерывном перемешивании эта смесь инкубировалась в течение 24 ч. Образцы отбирались через 1, 2, 3, 4, 8 и 24 ч инкубации. Для каждого образца ферментативная активность завершалась путем нагревания образца до 90 °С в течение 5 мин. После охлаждения рН каждого образца быстро снижалось до 4,5, используя для этого фосфорную кислоту, после чего суспензия центрифугировалась в течение 5 мин при 3000 rpm в центрифуге настольного типа Hereaus. Абсолютно чистый супернатант использовался для анализа ЖХ/МС/МС для определения содержания пептидов VPP, IPP, LPP, VVVPP и VVVPPF в супернатанте (см. раздел «Материалы и методы »). Казеин коровьего молока включает ряд различных белков, включая бета- и каппа-казеин. В соответствии с известными амино-последовательностями бета-казеин охватывает трипептиды IPP, VPP и LPP, ингибирующие АСЕ. В бета-казеине IPP находится в последовательности -P 71 -Q 72 -N 73 -I 74 -P 75 -P 76 -, VPP содержится в последовательности -P 81 -V 82 -V 83 -V 84 -P 85 -Р 86 - и LPP содержится в последовательности -P 150 -L 151 -P 152 -P 153 -. Каппа-казеин, который присутствует в препаратах казеина, полученных путем кислотного осаждения, в молярной концентрации почти 50% от концентрации бета-казеина, включающего только IPP. В каппа-казеине IPP содержится в последовательности -A 107 -I 108 -P 109 -P 110 -. Другие белковые компоненты казеина не содержат ни IPP, VPP, ни LPP. Табл. 2 и 3 показывают концентрации пептидов, присутствующих в подкисленных и процентрифугированных супернатантах, вычисленные на грамм казеинат калия, добавленного к инкубационной смеси. Как показано в табл. 2, концентрация IPP увеличивается через 1 ч инкубации. Помимо этого концентрация IPP не увеличивается в дальнейшем. Образование пентапептида VVVPP показывает такие же кинетики, что и образование IPP. Как теоретически предполагается, молярный выход VVVPP аналогичен молярному выходу пептида LPP. Выход обоих LPP и VVVPP увеличивается почти на 60% от того, что мы теоретически допустили. То, что максимум концентрации LPP достигается только через 3 ч инкубации, предполагает, что расщепление этой определенной части молекулы бета-казеина, возможно, является довольно более сложным. По сравнению с VVVPP, гексапептид VVVPPF не формируется вообще. Это наблюдение предполагает, что пролин-специфичная эндопротеаза эффективно расщепляет связь -P-F-, вследствие чего образуется VVVPP. Трипептид IPP образуется сразу же, но его молярный выход составляет не более трети максимального выхода VVVPP или LPP. Поскольку трипептид IPP содержатся как в бета-казеине, так и в каппа-казеине, такой результат является неожиданным. Вероятным объяснением этого результата является то, что пролин-специфичная протеаза может генерировать IPP, но только из каппа-казеиновой части казеинатов. Учитывая соответствующую аминокислотную последовательность каппа-казеина, предполагается, что пептидная связь -A 107 -I 108 - расщепляется аланин-специфичной активностью фермента. Если это верно, то количество отделенного IPP увеличится приблизительно на 40% от количества, которое присутствует в каппа-казеине, однако не более чем приблизительно на 10% от IPP, которое теоретически присутствует в бета- плюс каппа-казеине. Этот механизм расщепления для высвобождения IPP также объясняет, почему VPP не может образовываться из его молекулы предшественника VVVPP: просто необходимые эндопротеолитические активности не присутствуют в используемых ферментных препаратах, полученных из A. niger. Таблица 2 Таблица 3 Пример 6. Введение стадии осаждения кислого казеина приводит к пятикратному увеличению концентрации пептидов, ингибирующих АСЕ. Как описывается в примере 5, казеинат калия в концентрации 10% (мас./мас.) белка инкубировался с пролин-специфичной эндопротеазой, полученной из A. niger, при рН 6,0. Через различные инкубационные периоды образцы нагревались для остановки дальнейшей ферментативной активности, после чего рН опускали до 4,5 для уменьшения растворимости казеина. Не растворенные молекулы казеина удалялись с помощью низкоскоростного центрифугирования. В табл. 2 и 3 мы представляем концентрации пептидов, ингибирующих АСЕ, рассчитанные из расчета исходной концентрации 10% белка. Однако в результате подкисления и последующей стадии центрифугирования, большой процент добавленного белка удалялся. Для учета этого удаленного белкового содержания из подкисленных супернатантов выполнялись анализы содержания общего азота (Kjeldahl). В соответствие с последними данными, было обнаружено, что различные супернатанты содержали уровни белка, приведенные в табл. 4. Таблица 4 Учитывая эти данные, мы пересчитали концентрацию пептидов, ингибирующих АСЕ, находящихся в каждом супернатанте, но на этот раз используя их истинные белковые содержания. Эти пересчитанные данные приведены в табл. 5. Таблица 5 Сравнение данных, представленных в табл. 3 и 5, четко показывают, что стадия простого подкисления с последующим осуществлением промышленно возможной декантации, фильтрации или низкоскоростного центрифугирования приводит к пятикратному увеличению концентрации специфичных пептидов и ингибирующих АСЕ. Пример 7. Идентификация новых и активных трипептидов MAP и ITP, ингибирующих АСЕ, в концентрированных казеиновых гидролизатах. Для обеспечения более тщательного анализа присутствующих биоактивных пептидов по препаративному синтезу был приготовлен казеиновый гидролизат, полученный путем ферментации чистой пролин-специфичной эндопротеазы из A. niger и очищенный путем осаждения кислотой. Для этого 3000 г казеинат калия суспендировали в 25 л воды при 75 °С. После тщательной гомогенизации рН медленно доводили до 6,0, используя разбавленную фосфорную кислоту. После охлаждения до 55 °С получены из A. niger пролин-специфичные в концентрации 4 ферментативные единицы/грамм казеината (см. раздел «Материалы и методы для определения единиц »). После инкубации (при помешивании) в течение 3 ч при 55 °С, рН уменьшали до 4,5 путем медленного добавления концентрированной фосфорной кислоты. В этом препаративном синтезе стадия термической обработки для инактивации пролин-специфичной эндопротеазы в этой части способа упускалась. Затем суспензия быстро охлаждалась до 4 °С и сохранялась в течение ночи (без перемешивания) при этой температуре. На следующее утро чистый верхний слой сливался и выпаривался до достижения уровня 40% сухого вещества. Последняя концентрированная жидкость подвергалась обработке UHT (ультравысокой температуры) в течение 4 с при 140 °С и затем ультрафильтрации при 50 °С. После бактериальной фильтрации жидкость высушивалась распылением. Этот материал именуется в дальнейшем как биоактивные пептиды, полученные из казеина (CDBAP (Casein Derived Bio-Active Peptides)). Используя процедуры ЖХ/МС, изложенные в разделе «Материалы и методы », определялось содержание IPP, LPP и VPP порошкового продукта. В зависимости от содержания азота, в порошковом продукте содержание белка составляло около 60% (используя коэффициент пересчёта 6,38). Содержания IPP, LPP и VPP порошка представлено в табл. 6. Аминокислотная композиция продукта CDBAP представлена в табл. 7. Довольно примечательным являлось увеличение молярного содержания пролина в материале, высушенном распылением, полученного после осаждения кислотой: от исходного 12 до приблизительно 24%. Таблица 6 Таблица 7 Наличие новых пептидов, ингибирующих АСЕ, в CDBAP изучалось путем использования двумерного хроматографического разделения в сочетании с at-line анализа ингибирования АСЕ и масс-спектрометрии для идентификации. В первом анализе смесь пептидов отделялась на колонке для жидкостной хроматографии (ЖХ) ODS3 и профили ингибирования АСЕ были получены из различных образованных фракций. Во втором анализе фракции из первой колонки, показывающие высокое ингибирование АСЕ, затем отделялись на колонке для ЖХ Biosuite при использовании различного профиля градиента. Фракции, собранные из второй колонки, разделялись на две части: одна часть использовалась для измерения ингибирования АСЕ at-line, в то время как другая часть подвергалась анализам МС и МС-МС для идентификации присутствия пептидов. Все анализы выполнялись при использовании системы ВЭЖХ Alliance 2795 (Waters, Etten-Leur, the Netherlands), оснащенной двулучевым УФ-детектором. Для идентификации пептидов ВЭЖХ-система соединялась с масс-спектрометром Q-TOF того же производителя. В тестах 20 мкл 10% (мас./об.) раствора CDBAP в Milli-Q воде вводилось в колонку ODS3 150 ×2,1 Inertsil 5 с размером частиц 5 мкм (Varian, Middelburg, the Netherlands). Подвижная фаза А состояла из 0,1% раствора трифторуксусной кислоты (TFA) в Milli-Q воде. Подвижная фаза В состояла из 0,1% раствора TFA в ацетонитриле. Исходная элюентная композиция составляла 100% раствора А. Элюент сохранялся при 100% А в течение 5 мин. Затем линейный градиент начинали в течение 10 мин с 5% В, с последующим линейным градиентом в течение 10 мин до 30% В. Колонка промывалась путем увеличения концентрации раствора В до 70% в течение 5 мин и сохранялась при 70% раствора В в течение последующих 5 мин. После чего элюент менялся на 100% раствор А в течение 1 мин и уравновешивался в течение 9 мин. Общее время пробега составило 50 мин. Скорость потока элюента составляла 0,2 мл ∙мин -1 и температура колонки поддерживалась при 60 °С. УФ-хроматограмма записывалась при длине волны 215 нм. Фракции элюента собирались в 96-луночную планшету, используя одноминутный интервал времени, в результате чего получились объемы фракций в 200 мкл. Элюент в лунках нейтрализовался путем добавления 80 мкл 0,05% водного раствора гидроксида аммония (25%). Растворитель испарялся до сухости в азотной среде при 50 °С. После чего остаток разбавлялся в 40 мкл Milli-Q воды и перемешивался в течение 1 мин. Для at-line анализа ингибирования АСЕ добавлялось 27 мкл 33,4 мЕд мл -1 АСЕ (фермент получен от Sigma Chemicals) в фосфатно-буферном солевом растворе (PBS) с рН 7,4 с концентрацией хлорида 260 мМ, и смесь инкубировалась в течение 5 мин в смесителе для 96-луночных планшет при 700 RPM. После инкубационного периода добавляли 13 мкл 0,35 мМ раствора гиппуровая кислота-гистидин-лейцин (HHL) в буфере PBS и перемешивали в течение 1 мин при 700 RPM. Смесь вступала в реакцию в течение 60 мин при 50 °С в ГХ-термостате. После завершения реакции планшеты охлаждали в тающем льде. Затем 96-луночная планшета анализировалась на flash-ВЭЖХ-колонке. Из реакционной смеси каждой лунки 30 мкл вводилось в ВЭЖХ-колонку Chromlith Flash RP18e 25 ×4,6 мм (Merck, Darmstadt, Germany), оснащенной защитной колонкой RP18e 10 ×4,6 мм того же производителя. Изократическая подвижная фаза состояла из 0,1% раствора TFA в воде/ацетонитриле 79/21. Поток элюента составлял 2 мл ∙мин -1 и температура колонки составляла 25 °С. Введения осуществлялись с интервалом времени в 1 мин. Гиппуровая кислота Н и HHL контролировались при 280 нм. Пиковые значения Н и HHL измерялись и ингибирование АСЕ (ACEI) каждой фракции рассчитывалось по формуле где ACEI α - процент ингибирования аналита; DC w - степень расщепления HHL до Н и HL в воде посредством АСЕ; DC α - степень расщепления HHL до Н и HL для аналита. Степень расщепления рассчитывали путем выражения пикового значения Н как доли от суммы пиковых значений Н и HHL. Наибольшее ингибирование АСЕ определялось во фракциях, извлеченных между 18 и 26 мин. Эта часть собиралась и повторно вводилась в колонку Biosuite 150 ×2,1 мм с размером частиц 3 мкм (Waters, Etten-Leur, the Netherlands). Подвижная фаза состояла из 0,1% раствора муравьиной кислоты (FA) в Milli-Q воде. Подвижная фаза В состояла из 0,1% раствора FA в метаноле. Исходная элюентная композиция являлась 100% А. Элюент сохранялся при 100% А в течение 5 мин. После этого линейный градиент начинался в течение 15 мин с 5% В с последующим линейным градиентом в течение 30 мин с 60% В. Элюент сохранялся при 60% В в течение последующих 5 мин. Окончательно элюент снижался до 100% подвижной фазы А в течение 1 мин и уравновешивался в течение 10 мин. Общее время пробега равнялось 65 мин. Поток элюента составлял 0,2 мл ∙мин -1 и температура колонки поддерживалась при 60 °С. УФ-излучение записывалось при 215 нм. Фракции собирались из колонки Biosuite при 10-секундном интервале времени. Фракции опять разделялись на две части, одна часть использовалась для измерения активности, используя at-line-способ определения ингибирования АСЕ, описанный ранее, тогда как другая часть использовалась для идентификации активных пептидов, используя анализ МС и МС-МС. Два хроматографических пика с молекулярными ионами массой 326,2080 Да и два других пика с молекулярными ионами 330,2029 и 318,1488 Да соответствовали повышенному ингибированию АСЕ, измеренные в период между 18 и 26 мин. Используя анализ МС-МС, эти пептиды были определены как структурные изомеры IPP и LPP (-0,6 ppm), ITP (-4,8 ppm) и MAP (+2,8 ppm) соответственно. Белковыми источниками пептидов являлись каппа-казеин f108-110 (IPP), β-казеин f151-153 (LPP), α-s2-казеин f119-121 (ITP) и β-казеин f102-104 (MAP). IPP и LPP были представлены ранее как пептиды, ингибирующие АСЕ, со значением IC 50 в 5 и 9,6 мкМ соответственно (Y. Nakamura, M. Yamamoto, K. Sakai, A. Okubo, S. Yamazaki, Т. Takano, J. Dairy Sci. 78 (1995) 777-783; Y. Aryoshi, Trends in Food Science and Technol. 4 (1993) 139-144). Однако трипептиды ITP и MAP не были представлены, насколько нам известно, ранее как эффективные пептиды, ингибирующие АСЕ. MAP, ITP и IPP были химически синтезированы и активность каждого пептида измерялась при использовании модифицированного анализа Matsui, описанного ниже. Количественное определение содержания MAP и ITP в различных образцах осуществлялось с помощью устройства Micromass Quattro II MS, работающего в положительном многореакционном режиме мониторинга электрораспыления. Используемый способ ВЭЖХ был подобен способам, описанным выше. МС-настройки (ESI+) были следующими: напряжение при распылении 37 В, напряжение в капиллярах 4 кВ, осушение азота при 300 л/ч. Температура источника и распылителя 100 и 250 °С соответственно. Синтетические пептиды использовались для приготовления калибровочной кривой, используя ион-предшественник 318,1 и суммированные ион-продукты 227,2 и 347,2 для MAP и используя ион-предшественника 320,2 и суммированные ион-продукты 282,2 и 501,2 для ITP. В соответствии с этим анализом новые трипептиды MAP и ITP, ингибирующие АСЕ, присутствовали в продукте CDBAP в количествах, соответствующих 2,9 мг МАР/г CDBAP или 4,8 мг МАР/г белка в CDBAP и 0,9 мг ITP/г CDBAP и 1,4 мг ITP/г белка в CDBAP. Для определения ингибирующей АСЕ-активности MAP и ITP химически синтезированные трипептиды анализировались способом, описанным Matsui et al. (Matsui, Т. et al. (1992) Biosci. Biotech. Biochem. 56: 517-518) с некоторыми незначительными модификациями. Различные инкубации показаны в табл. 8. Таблица 8 Каждый из четырех образцов содержал 75 мкл 3 мМ гиппурил-гистидин-лейцин (Hip-His-Leu, Sigma Chemicals), растворенного в 250 мМ раствора бората, содержащего 200 мМ NaCl, pH 8,3. АСЕ был получен от Sigma Chemicals. Смеси инкубировались при 37 °С и останавливались через 30 мин путем добавления 125 мкл 0,5 М HCI. Далее, было добавлено 225 мкл раствора бицин/NaOH (1 M NaOH: 0,25 М бицина(4:6)), с последующим добавлением 25 мкл 0,1 М TNBS (2,4,6-Тринитробензолсульфоновая кислота, Fluka, Switzerland; в 0,1 М Na 2 HPO 4 ). После инкубации в течение 20 мин при 37 °С добавлялось 4 мл 4 мМ Na 2 SO 3 в 0,2 М NaH 2 PO 4 и измерялось поглощение света при длине волны 416 нм на спектрофотометре UV/Vis (Shimadzu UV-1601 с CPS контроллером, Netherlands). Величина ингибирующей АСЕ-(ACEI)-активности рассчитывалась как процент ингибирования по сравнению с коэффициентом пересчёта АСЕ при отсутствии ингибирования по следующей формуле: где Контроль 1 - поглощение без АСЕ ингибирующего компонента (=макс. АСЕ активности) [AU]; Контроль 2 - поглощение без АСЕ ингибирующего компонента и без АСЕ (фон) [AU]; Образец 1 - поглощение при наличии АСЕ и АСЕ ингибирующего компонента [AU]; Образец 2 - поглощение при наличии АСЕ ингибирующего компонента, но без АСЕ [AU]. Полученное значение IC 50 химически синтезированных трипептидов MAP и ITP представлено в табл. 9 вместе со значениями IC 50 , полученными при at-line-измерениях, используемых в фазе скрининга эксперимента. Измерение химически синтезированного IPP включалось как внутренний стандарт для различных измерений. Таблица 9 Пример 8. Новые пептиды MAP и ITP, ингибирующие АСЕ, способны сохраняться в желудочно-кишечном тракте человека. После употребления диетические белки и пептиды подвергаются различным пищеварительным ферментативным процессам в желудочно-кишечном тракте. Для определения стабильности вновь идентифицированных биоактивных пептидов в желудочно-кишечном тракте человека препарат CDBAP (приготовленный, как описывалось в примере 7) подвергался желудочно-кишечной обработке (GIT), воспроизводящей пищеварительные условия, обычно найденные в организме человека. Образцы, полученные после различных инкубационных периодов в GIT-модельной системе, были проанализированы, используя on-line-систему ВЭЖХ-Bioassay-MS или систему HRS-MS для количественного определения любых остаточных пептидов MAP и ITP. Процедура GIT осуществлялась в стандартном смесителе, содержащем 100 мл флакон (как снабжено Vankel, US). Температура водяной бани поддерживалась при 37,5 °С и лопастная скорость - таким образом, чтобы образец сохранялся в суспензии (100 rpm). Около 3,4 г CDBAP (уровень белков приблизительно 60%) было растворено/суспендировано в 100 мл Milli-Q воды. Во время желудочной стимуляции использовалось 5 М HCl для уменьшения рН. В конце желудочной стимуляции и во время дуоденальной фазы использовалось 5 М NaOH для увеличения рН. Суспензия CDBAP предварительно нагревалась до 37,5 °С и 5 мл суспензии извлекалось для растворения 0,31 г пепсина (Fluka порядковый номер 77161). При t=0 мин 5 мл с вновь растворенным пепсином добавлялось обратно в суспензию. Затем значение рН суспензии CDBAP регулировалось медленно вручную при использовании раздельного рН-метра по следующей схеме: При t=90 мин 0,139 г восьмикратного USP панкреатина (Sigma порядковый номер Р7545) был аккуратно смешан с другими 5 мл суспензии CDBAP и немедленно добавлен обратно. Инкубация продолжалась по следующей схеме: Эксперимент останавливался при t=125 мин и значение рН сверялось (рН составляло 7). Затем образцы переносились в сосуд и помещались в микроволновую печь. Затем образцы перемещались в стеклянные пробирки и инкубировались при температуре 95 °С в течение 60 мин для инактивации всей протеазной активности. После охлаждения образцы помещались в пробирки Falcon и центрифугировались в течение 10 мин при 3000 ×g. Супернатант подвергался лиофилизации. Содержание общего азота полученного порошка определялось и конвертировалось с уровнем белка, используя Kjeldahl-фактор казеина (6,38). В соответствии с этими данными уровень белка препарата CDBAP после процедуры GIT составлял 48,4%. Уровни MAP и ITP, сохранившиеся при протеолитической обработке по процедуре GIT, определялись, как описано в примере 7, и полученные данные показаны в табл. 10. В соответствии с результатами экспериментов оба MAP и ITP проявляют высокую устойчивость против расщепления GIT. В сочетании с низкими значениями IC 50 для этих трипептидов (как показано в примере 7) данные указывают на значительный потенциал двух новых пептидов, ингибирующих АСЕ, как пептидов, снижающих кровяное давление. |