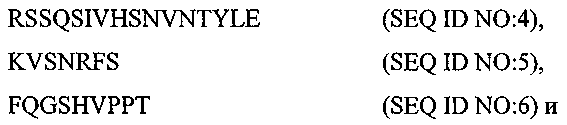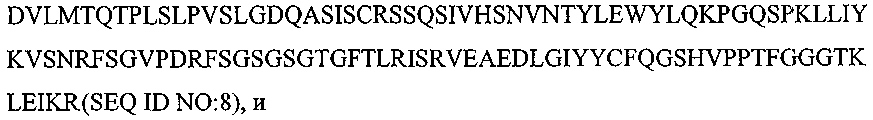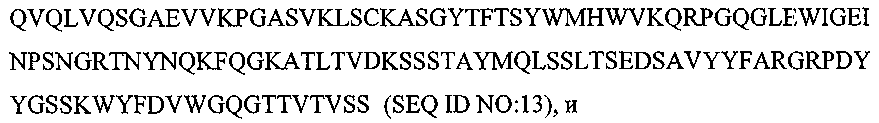|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000009807b*\id |
|
Термины запроса в документе Реферат 1. Композиция, содержащая: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент и где указанное антитело или указанный фрагмент специфически связывается с рецептором инсулин-подобного фактора роста I, выбранный из группы, состоящей из: (i) антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), (ii) антитела с перестроенной поверхностью или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (iii) человеческого или гуманизированного антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (iv) функционального эквивалента антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (v) варианта мышиного антитела ЕМ164 или его эпитопсвязывающего фрагмента, имеющих по меньшей мере одну мутацию, делецию или инсерцию нуклеотида в сравнении с мышиным антителом ЕМ164 и имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, и (vi) мышиного антитела ЕМ164, продуцируемого мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающего фрагмента, и (b) второй терапевтический агент. 2. Композиция по п.1, где указанный второй терапевтический агент выбран из группы, состоящей из таких терапевтических агентов, как доцетаксел, паклитаксел, доксорубицин, эпирубицин, циклофосфамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, анастрозол, фулвестрант, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорелбин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идарубицин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2b, мелфалам, бортезомиб (Velcade), алтретамин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), антитело к EGF-рецептору (Cetuxivmab, Abx-EGF) и эпотилон. 3. Композиция по п.1, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитабина и камптотецина. 4. Композиция по п.1, где указанный первый терапевтический агент вводят пациенту в дозе приблизительно 1 - приблизительно 2000 мг/кв.м и где указанный второй терапевтический агент вводят в дозе приблизительно 10 - приблизительно 2000 мг/кв.м. 5. Композиция по п.1, где указанный первый терапевтический агент вводят пациенту в дозе приблизительно 10 - приблизительно 1000 мг/кв.м и где указанный второй терапевтический агент вводят в дозе приблизительно 50 - приблизительно 1000 мг/кв.м. 6. Фармацевтическая композиция, содержащая композицию по п.1 и фармацевтически приемлемый носитель или разбавитель. 7. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или эпитопсвязывающий фрагмент антитела, содержащий по меньшей мере один определяющий комплементарность район, имеющий аминокислотную последовательность, выбранную из группы, состоящей из (b) второй терапевтический агент, где, когда выбрана SEQ ID NO:5, указанное антитело или фрагмент антитела специфически связывается с рецептором инсулин-подобного фактора роста I (IGF-IR). 8. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или эпитопсвязывающий фрагмент антитела, содержащий по меньшей мере одну вариабельную область тяжелой цепи и по меньшей мере одну вариабельную область легкой цепи, где указанная вариабельная область тяжелой цепи содержит три последовательных определяющих комплементарность района, имеющего аминокислотные последовательности, представленные SEQ ID NO:1-3, соответственно: и где указанная вариабельная область легкой цепи содержит три последовательных определяющих комплементарность района, имеющего аминокислотные последовательности, представленные SEQ ID NO:4-6, соответственно: (b) второй терапевтический агент. 9. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент антитела, причем указанное антитело или указанный фрагмент содержит вариабельную область тяжелой цепи, которая имеет по меньшей мере 90% идентичность последовательности относительно аминокислотной последовательности, представленной SEQ ID NO:7: (b) второй терапевтический агент. 10. Композиция по п.9, где указанная вариабельная область тяжелой цепи имеет по меньшей мере 95% идентичность последовательности относительно указанной аминокислотной последовательности, представленной SEQ ID NO:7. 11. Композиция по п.9, где указанная вариабельная область тяжелой цепи имеет аминокислотную последовательность, которая представлена SEQ ID NO:7. 12. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область легкой цепи, которая имеет по меньшей мере 90% идентичность последовательности относительно аминокислотной последовательности, представленной SEQ ID NO:8: (b) второй терапевтический агент. 13. Композиция по п.12, где указанная вариабельная область легкой цепи имеет по меньшей мере 95% идентичность последовательности относительно указанной аминокислотной последовательности, представленной SEQ ID NO:8. 14. Композиция по п.12, где указанная вариабельная область легкой цепи имеет аминокислотную последовательность, которая представлена SEQ ID NO:8. 15. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область легкой цепи, имеющую последовательность, выбранную из группы, состоящей из: и (b) второй терапевтический агент. 16. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область тяжелой цепи, имеющую последовательность, представленную SEQ ID NO:13: (b) второй терапевтический агент. 17. Композиция по любому из пп.7-16, где указанный второй терапевтический агент выбран из группы, состоящей из доцетаксела, паклитаксела, доксорубицина, эпирубицина, циклофосфамида, трастузумаба (Herceptin), капецитабина, тамоксифена, торемифена, летрозола, анастрозола, фулвестранта, экземестана, госерелина, оксалиплатина, карбоплатина, цисплатина, дексаметазона, антида, бевацизумаба. (Avastin), 5-фторурацила, лейковарина, левамизола, иринотекана, этопозида, топотекана, гемцитабина, винорелбина, эстрамустина, митоксантрона, абареликса, золедроната, стрептозоцина, ритуксимаба (Rituxan), идарубицина, бусульфана, хлорамбуцила, флударабина, иматиниба, цитарабина, ибритумомаба (Zevalin), тозитумомаба. (Bexxar), интерферона альфа-2b, мелфалама, бортезомиба (Velcade), алтретамина, аспарагиназы, гефитиниба (Iressa), эрлонитиба (Tarceva), антитела против EGF-рецептора (Cetuxivmab, Abx-EGF) и эпотилона. 18. Композиция по любому из пп.7-16, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитабина и камптотецина. 19. Способ ингибирования роста раковой клетки, предусматривающий контактирование указанной клетки с композицией по п.1. 20. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества композиции по п.1. 21. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества фармацевтической композиции по п.6. 22. Способ лечения по любому из пп.19-21, где указанный рак является раком, выбранным из группы, состоящей из рака молочной железы, рака ободочной кишки, рака яичника, остеосаркомы, рака шейки матки, рака предстательной железы, рака легкого, синовиального рака, рака поджелудочной железы, меланомы, множественной миеломы, нейробластомы и рабдомиосаркомы. 23. Набор, содержащий: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) второй терапевтический агент и (c) инструкции по применению. 24. Способ ингибирования роста раковой клетки, предусматривающий контактирование указанной клетки с: (a) первым терапевтическим агентом, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) вторым терапевтическим агентом. 25. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества: (a) первого терапевтического агента, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемого мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) второго терапевтического агента. 26. Способ по п.24, где указанную клетку контактируют с указанным первым терапевтическим агентом и указанным вторым терапевтическим агентом одновременно. 27. Способ по п.24, где указанную клетку контактируют с указанным первым терапевтическим агентом и указанным вторым терапевтическим агентом последовательно и в любом порядке. 28. Способ по п.25, где указанный первый терапевтический агент и указанный второй терапевтический агент вводят одновременно. 29. Способ по п.25, где указанный первый терапевтический агент и указанный второй терапевтический агент вводят последовательно и в любом порядке. 30. Способ по п.24 или 25, где указанный второй терапевтический агент выбран из группы состоящей из таких терапевтических агентов, как доцетаксел, паклитаксел, доксорубицин, эпирубицин, циклофосфамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, анастрозол, фулвестрант, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорелбин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идарубицин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2b, мелфолам, бортезомиб (Velcade), алтретамин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), анти-EGF-рецептор-антитело (Cetuxivmab, Abx-EGF) и эпотилон. 31. Способ по п.24 или 25, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитабина и камптотецина.
Полный текст патента (57) Реферат / Формула: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент и где указанное антитело или указанный фрагмент специфически связывается с рецептором инсулин-подобного фактора роста I, выбранный из группы, состоящей из: (i) антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), (ii) антитела с перестроенной поверхностью или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (iii) человеческого или гуманизированного антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (iv) функционального эквивалента антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (v) варианта мышиного антитела ЕМ164 или его эпитопсвязывающего фрагмента, имеющих по меньшей мере одну мутацию, делецию или инсерцию нуклеотида в сравнении с мышиным антителом ЕМ164 и имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, и (vi) мышиного антитела ЕМ164, продуцируемого мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающего фрагмента, и (b) второй терапевтический агент. 2. Композиция по п.1, где указанный второй терапевтический агент выбран из группы, состоящей из таких терапевтических агентов, как доцетаксел, паклитаксел, доксорубицин, эпирубицин, циклофосфамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, анастрозол, фулвестрант, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорелбин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идарубицин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2b, мелфалам, бортезомиб (Velcade), алтретамин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), антитело к EGF-рецептору (Cetuxivmab, Abx-EGF) и эпотилон. 3. Композиция по п.1, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитабина и камптотецина. 4. Композиция по п.1, где указанный первый терапевтический агент вводят пациенту в дозе приблизительно 1 - приблизительно 2000 мг/кв.м и где указанный второй терапевтический агент вводят в дозе приблизительно 10 - приблизительно 2000 мг/кв.м. 5. Композиция по п.1, где указанный первый терапевтический агент вводят пациенту в дозе приблизительно 10 - приблизительно 1000 мг/кв.м и где указанный второй терапевтический агент вводят в дозе приблизительно 50 - приблизительно 1000 мг/кв.м. 6. Фармацевтическая композиция, содержащая композицию по п.1 и фармацевтически приемлемый носитель или разбавитель. 7. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или эпитопсвязывающий фрагмент антитела, содержащий по меньшей мере один определяющий комплементарность район, имеющий аминокислотную последовательность, выбранную из группы, состоящей из (b) второй терапевтический агент, где, когда выбрана SEQ ID NO:5, указанное антитело или фрагмент антитела специфически связывается с рецептором инсулин-подобного фактора роста I (IGF-IR). 8. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или эпитопсвязывающий фрагмент антитела, содержащий по меньшей мере одну вариабельную область тяжелой цепи и по меньшей мере одну вариабельную область легкой цепи, где указанная вариабельная область тяжелой цепи содержит три последовательных определяющих комплементарность района, имеющего аминокислотные последовательности, представленные SEQ ID NO:1-3, соответственно: и где указанная вариабельная область легкой цепи содержит три последовательных определяющих комплементарность района, имеющего аминокислотные последовательности, представленные SEQ ID NO:4-6, соответственно: (b) второй терапевтический агент. 9. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент антитела, причем указанное антитело или указанный фрагмент содержит вариабельную область тяжелой цепи, которая имеет по меньшей мере 90% идентичность последовательности относительно аминокислотной последовательности, представленной SEQ ID NO:7: (b) второй терапевтический агент. 10. Композиция по п.9, где указанная вариабельная область тяжелой цепи имеет по меньшей мере 95% идентичность последовательности относительно указанной аминокислотной последовательности, представленной SEQ ID NO:7. 11. Композиция по п.9, где указанная вариабельная область тяжелой цепи имеет аминокислотную последовательность, которая представлена SEQ ID NO:7. 12. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область легкой цепи, которая имеет по меньшей мере 90% идентичность последовательности относительно аминокислотной последовательности, представленной SEQ ID NO:8: (b) второй терапевтический агент. 13. Композиция по п.12, где указанная вариабельная область легкой цепи имеет по меньшей мере 95% идентичность последовательности относительно указанной аминокислотной последовательности, представленной SEQ ID NO:8. 14. Композиция по п.12, где указанная вариабельная область легкой цепи имеет аминокислотную последовательность, которая представлена SEQ ID NO:8. 15. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область легкой цепи, имеющую последовательность, выбранную из группы, состоящей из: и (b) второй терапевтический агент. 16. Композиция, содержащая: (а) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область тяжелой цепи, имеющую последовательность, представленную SEQ ID NO:13: (b) второй терапевтический агент. 17. Композиция по любому из пп.7-16, где указанный второй терапевтический агент выбран из группы, состоящей из доцетаксела, паклитаксела, доксорубицина, эпирубицина, циклофосфамида, трастузумаба (Herceptin), капецитабина, тамоксифена, торемифена, летрозола, анастрозола, фулвестранта, экземестана, госерелина, оксалиплатина, карбоплатина, цисплатина, дексаметазона, антида, бевацизумаба. (Avastin), 5-фторурацила, лейковарина, левамизола, иринотекана, этопозида, топотекана, гемцитабина, винорелбина, эстрамустина, митоксантрона, абареликса, золедроната, стрептозоцина, ритуксимаба (Rituxan), идарубицина, бусульфана, хлорамбуцила, флударабина, иматиниба, цитарабина, ибритумомаба (Zevalin), тозитумомаба. (Bexxar), интерферона альфа-2b, мелфалама, бортезомиба (Velcade), алтретамина, аспарагиназы, гефитиниба (Iressa), эрлонитиба (Tarceva), антитела против EGF-рецептора (Cetuxivmab, Abx-EGF) и эпотилона. 18. Композиция по любому из пп.7-16, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитабина и камптотецина. 19. Способ ингибирования роста раковой клетки, предусматривающий контактирование указанной клетки с композицией по п.1. 20. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества композиции по п.1. 21. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества фармацевтической композиции по п.6. 22. Способ лечения по любому из пп.19-21, где указанный рак является раком, выбранным из группы, состоящей из рака молочной железы, рака ободочной кишки, рака яичника, остеосаркомы, рака шейки матки, рака предстательной железы, рака легкого, синовиального рака, рака поджелудочной железы, меланомы, множественной миеломы, нейробластомы и рабдомиосаркомы. 23. Набор, содержащий: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) второй терапевтический агент и (c) инструкции по применению. 24. Способ ингибирования роста раковой клетки, предусматривающий контактирование указанной клетки с: (a) первым терапевтическим агентом, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) вторым терапевтическим агентом. 25. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества: (a) первого терапевтического агента, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемого мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) второго терапевтического агента. 26. Способ по п.24, где указанную клетку контактируют с указанным первым терапевтическим агентом и указанным вторым терапевтическим агентом одновременно. 27. Способ по п.24, где указанную клетку контактируют с указанным первым терапевтическим агентом и указанным вторым терапевтическим агентом последовательно и в любом порядке. 28. Способ по п.25, где указанный первый терапевтический агент и указанный второй терапевтический агент вводят одновременно. 29. Способ по п.25, где указанный первый терапевтический агент и указанный второй терапевтический агент вводят последовательно и в любом порядке. 30. Способ по п.24 или 25, где указанный второй терапевтический агент выбран из группы состоящей из таких терапевтических агентов, как доцетаксел, паклитаксел, доксорубицин, эпирубицин, циклофосфамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, анастрозол, фулвестрант, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорелбин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идарубицин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2b, мелфолам, бортезомиб (Velcade), алтретамин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), анти-EGF-рецептор-антитело (Cetuxivmab, Abx-EGF) и эпотилон. 31. Способ по п.24 или 25, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитабина и камптотецина.
009807 Область техники, к которой относится изобретение Данное изобретение относится к антителам, которые связываются с рецептором инсулин-подобного фактора-I (IGF-I-рецептором). Более конкретно, данное изобретение относится к антителам против IGF-I-рецептора, которые ингибируют клеточные функции IGF-I-рецептора. Еще более конкретно, данное изобретение относится к антителам против IGF-I-рецептора, которые являются антагонистами эффектов IGF-I, IGF-II и сыворотки на рост и выживание опухолевых клеток и которые, по существу, лишены аго-нистической активности. Данное изобретение относится также к фрагментам указанных антител, гуманизированным и переделанным версиям указанных антител, конъюгатам указанных антител, производным антител и их применениям в диагностических, исследовательских и терапевтических приложениях. Далее, данное изобретение относится к улучшенным антителам или их фрагментам, которые получены из вышеописанных антител и их фрагментов. В другом аспекте, данное изобретение относится к полинуклеотиду, кодирующему эти антитела или их фрагменты, и к векторам, содержащим эти полинуклеотиды. Уровень техники Рецептор инсулин-подобного фактора-I (IGF-I-рецептор) является трансмембранным гетеротетра-мерным белком, который имеет две внеклеточные альфа-цепи и две простирающиеся через мембрану бета-цепи в дисульфид-связанной конфигурации р-а-а-р. Связывание лигандов, которыми являются инсулин-подобный фактор роста I (IGF-I) и инсулин-подобный фактор роста II (IGF-II), внеклеточным доменом IGF-I-рецептора активирует его внутриклеточный тирозинкиназный домен, приводя к автофосфо-рилированию этого рецептора и фосфорилированию субстрата. IGF-I-рецептор является гомологичным рецептору инсулина, имеющим высокое сходство последовательности 84% в тирозинкиназном домене бета-цепи и низкое сходство последовательности 48% в богатом цистеинами внеклеточном домене (Ul-rich., A. et al., 1986, EMBO, 5, 2503-2512; Fujita-Yamaguchi, Y. et al., 1986, J. BIol. Chem., 261, 1672716731; LeRoith, D. et al., 1995, Endocrine Reviews, 16, 143-163). IGF-I-рецептор и его лиганды (IGF-I и IGF-II) играют важные роли в многочисленных физиологических процессах, в том числе в росте и развитии во время эмбриогенеза, метаболизме, клеточной пролиферации и дифференцировке клеток во взрослых людях (LeRoith, D., 2000, Endocrinology, 141, 1287-1288; LeRoith, D., 1997, New England J. Med., 336, 633-640). IGF-I и IGF-II функционируют как в качестве эндокринных гормонов в крови, где они преимущественно присутствуют в комплексах с IGF-связывающими белками, так и в качестве паракринных и ауток-ринных факторов роста, которые продуцируются локально (Humbel, R.E., 1990, Eur. J. Biochem., 190, 445-462; Cohick, W.S. and Clemmons, D.R. 1993, Annu. Rev. Physiol. 55, 131-153). Предполагалось, что IGF-I-рецептор участвует в стимуляции роста, трансформации и выживания опухолевых клеток (Baserga, R. et al., 1997, Biochem. Biophus. Acta, 1332, F105-F126; Blakesley, V.A. et al., 1997, Journal of Endocrinology, 152, 339-344; Kaleko, M., Rutter, W.J., and Miller, A.D. 1990, Mol. Cell. Biol., 10, 464-473). Так, известно, что несколько типов опухолей экспрессируют более высокие, чем нормальные, уровни IGF-I-рецептора, в том числе рак молочной железы, рак ободочной кишки, рак яичника, синовиальная саркома и рак поджелудочной железы (Khandwala, H.M. et al, 2000, Endocrine Reviews, 21, 215-244; Werner, H. and LeRoith, D., 1995, Adv. Cancer Res., 68, 183-223; Happerfield, L.C. et al., 1997, J. Pathol., 183, 412-417; Frier, S. et al, 1999, Gut, 44, 704-708; van Dam, P.A. et al., 1994, J. Clin. Pathol., 47, 914-919; Xie, Y. et al., 1999, Cancer Res., 59, 3588-3591; Bergmann, U. et al., 1995, Cancer Res., 55, 20072011). In vitro было обнаружено, что IGF-I и IGF-II являются сильными митогенами для нескольких клеточных линий человека, таких как линии клеток рака легкого, рака молочной железы, рака ободочной кишки, остеосаркомы и рака шейки матки (Ankrapp, D.P. and Bevan, D.R., 1993, Cancer Res., 53, 33993404; Cullen, K.J., 1990, Cancer Res., 50, 48-53; Hermanto, U., 2000, Cell Growth & Differentiation, 11, 655664; Guo, Y.S. et al., 1995, J. Am. Coll. Surg., 181, 145-154; Kappel, C.C. et al., 1994, Cancer Res., 54, 28032807; Steller, M.A. et al, 1996, Cancer Res., 56, 1761-1765). Несколько из этих опухолей и линий опухолевых клеток также экспрессируют IGF-I или IGF-II, которые могут стимулировать их рост аутокринным или паракринным образом (Quinn, K.A. et al., 1996, J. Biol. Chem., 271, 11477-11483). Эпидемиологические исследования показали корреляцию повышенного уровня в плазме IGF-I (и более низкого уровня IGF-связывающего белка-3) с увеличенным риском рака предстательной железы, рака ободочной кишки, рака легкого и рака молочной железы (Chan, J.M. et al., 1998, Science, 279, 563566; Wolk, A. et al, 1998, J. Natl. Cancer Inst., 90, 911-915; Ma, J. et al, 1999, J. Natl. Cancer Inst, 91, 620-625; Yu, H. et al., 1999, J. Natl. Cancer Inst., 91, 151-156; Hankinson, S.E. et al., 1998, Lancet, 351, 1393-1396). Стратегии понижения уровня IGF-I в плазме или ингибирования функции IGF-I-рецептора были предложены для предотвращения рака (Wu, Y. et al., 2002, Cancer Res., 62, 1030-1035; Grimberg, A and Cohen P., 2000, J. Cell. Physiol., 183, 1-9). IGF-I-рецептор защищает опухолевые клетки от апоптоза, вызываемого депривацией фактора роста, независимостью сцепления или обработкой цитотоксическими лекарственными средствами (Navarro, M. and Baserga, R., 2001, Endocrinology, 142, 1073-1081; Baserga, R. et al., 1997, Biochem. Biophys. Acta, 1332, F105-F126). Были идентифицированы мутационным анализом домены IGF-I-рецептора, которые являются решающими для его митогенной, трансформирующей и антиапоптотической активностей. - 1 - 009807 Например, остаток тирозина 1251 IGF-I-рецептора был идентифицирован как критический для ан-тиапоптотической и трансформирующей активностей, но не для его митогенной активности (O'Connor, R. et al., 1997, Mol. Cell. Biol., 17, 427-435; Miura, M. et al, 1995, J. Biol. Chem. 270, 22639-22644). Внутриклеточный путь передачи сигнала активируемого лигандом IGF-I-рецептора включает в себя фосфорили-рование остатков тирозина субстратов рецептора инсулина (IRS-1 и IRS-2), которые рекрутируют фосфа-тилинозитол-3-киназу (PI-3-киназу) к мембране. Мембраносвязанные фосфолипидные продукты PI-3-киназы активирует серин/треонинкиназу Akt, субстраты которой включают в себя про-апоптотический белок BAD, который фосфорилируется до неактивного состояния (Datta, S.R., Brunet, A. and Greenberg, M.E., 1999, Genes & Development, 13, 2905-2927; Kulik, G., Klippel, A. and Weber, M.J., 1997, Mol. Cell. Biol. 17, 1595-1606). Митогенная передача сигнала IGF-I-рецептора в клетках MCF-7 рака молочной железы человека требует PI-3-киназы и является независимой от митоген-активируемой протеинкиназы, тогда как передача сигнала выживания в дифференцированных клетках РС12 крысиной феохромоцито-мы требует как PI-3-киназы, так и путей передачи сигнала митоген-активируемой протеинкиназы (Du-fourny, В. et al., 1997, J. Biol. Chem., 272, 31163-31171; Parrizas, M., Saltiel, A.R. and LeRoith, D., 1997, J. Biol. Chem., 272, 154-161). Было показано, что понижающая регуляция уровня IGF-I-рецептора антисмысловыми стратегиями уменьшает онкогенность нескольких линий опухолевых клеток in vivo и in vitro, таких как меланома, рак легкого, рак яичника, глиобластома, нейробластома и рабдомиосаркома (Resnicoff, M. et al., 1994, Cancer Res., 54, 4848-4850; Lee, C.-T. et al, 1996, Cancer Res., 56, 3038-3041; Muller, M. et al., 1998, Int. J. Cancer, 77, 567-571; Trojan, J. et al., 1993, Science, 259, 94-97; Liu, X. et al., 1998, Cancer Res., 58, 5432-5438; Shapiro, D.N. et al, 1994, J. Clin. Invest., 94, 1235-1242). Подобным образом, было сообщено, что доминантный негативный мутант IGF-I-рецептора уменьшает онкогенность in vivo и рост in vitro трансформированных клеток Rat-1, сверхэкспрессирующих IGF-I-рецептор (Prager, D. et al, 1994 Proc. Natl. Acad. Sci. USA, 91, 2181-2185). Опухолевые клетки, экспрессирующие антисмысловую в отношении IGF-I-рецептора мРНК, подвергаются обширному апоптозу при инъецировании в животных в биодиффузионных камерах. Это наблюдение делает IGF-I-рецептор привлекательной терапевтической мишенью на основе гипотезы, что опухолевые клетки являются более чувствительными, чем нормальные клетки, к апоптозу, вызываемому ингибированием IGF-I-рецептора (Resnicoff, M. et al., 1995, Cancer Res., 55, 2463-2469; Baserga, R., 1995, Cancer Res., 55, 249-252). Другой стратегией ингибирования функции IGF-I-рецептора в опухолевых клетках было использование антител против IGF-I-рецептора, которые связываются с внеклеточным доменом IGF-I-рецептора и ингибируют его активацию. Сообщалось о нескольких попытках разработки мышиных моноклональных антител против IGF-I-рецептора, из которых два ингибиторных антитела, IR3 и IH7, являются доступными и об их применении сообщалось в нескольких исследованиях по IGF-I-рецептору. Антитело IR3 было разработано с использованием частично очищенного плацентарного препарата рецептора инсулина для иммунизации мышей, которые образовывали антитело, IR1, которое было селективным в отношении связывания рецептора инсулина, и были получены два антитела, IR2 и IR3, которые показали преимущественную иммунопреципитацию IGF-I-рецептора (рецептора соматометина-С), но только слабую иммунопреципитацию рецептора инсулина (Kull, F.C. et al., 1983, J. Biol. Chem., 258, 6561-6566). Антитело 1 Н7 было получено иммунизацией мышей очищенным плацентарным препаратом IGF-I-рецептора, которая давала ингибиторное антитело 1 Н7, наряду с тремя стимуляторными антителами (Li, S.-L. et al., 1993, Biochem. Biophys. Res. Commun., 196, 92-98; Xiong, L. et al., 1992 Proc. Natl. Acad. Sci. USA, 89, 5356-5360). В другом сообщении панель мышиных моноклональных антител, специфических в отношении IGF-I-рецептора человека, получали иммунизацией мышей трансфицированными клетками 3Т3, экспресси-рующими высокие уровни IGF-I-рецептора, которые были классифицированы на семь групп с использованием исследований конкурентного связывания и по их ингибированию или стимуляции связывания IGF-I с трансфицированными клетками 3Т3 (Soos, M.A. et al., 1992, J. Biol. Chem., 267, 12955-12963). Таким образом, хотя антитело IR3 является наиболее часто используемым ингибиторным антителом для исследований IGF-I-рецептора in vitro, оно имеет недостаток, заключающийся в том, что оно проявляет агонистическую активность в трансфицированных клетках 3T3 и СНО, экспрессирующих IGF-I-рецептор человека (Kato, H. et al., 1993, J. Biol. Chem., 268, 2655-2661; Steele-Perkins, G. and Roth, R.A., 1990, Biochem. Biophys. Res. Commun., 171, 1244-1251). Подобным образом, среди этой панели антител, разработанной Soos et al., большинство ингибиторных антител 24-57 и 24-60 также показали агонистиче-ские активности в трансфицированных клеток 3T3 (Soos, M.A. et al., 1992, J. Biol. Chem., 267, 1295512963). Хотя сообщается, что антитело IR3 ингибирует связывание IGF-I (но не IGF-II) с экспрессируе-мыми рецепторами в интактных клетках и после солюбилизации, было показано, что оно ингибирует способность IGF-I и IGF-II стимулировать синтез ДНК в клетках in vitro (Steele-Perkins, G. and Roth, R.A., 1990, Biochem. Biophys. Res. Commun., 171, 1244-1251). Был сделан вывод на основании химерных конструкций инсулин-IGF-I-рецептора, что связывающим эпитопом антитела IR3 является район 223-274 - 2 - 009807 IGF-I-рецептора (Gustafson, ТА. and Rutter, W.J., 1990, J. Biol. Chem., 265, 18663-18667; Soos, M.A. et al., 1992, J. Biol. Chem., 267, 12955-12963). Линию клеток рака молочной железы человека MCF-7 обычно используют в качестве модельной клеточной линии для демонстрации реакции роста IGF-I и IGF-II in vitro (Dufourny, В. et al., 1997, J. Biol. Chem., 272, 31163-31171). В клетках MCF-7 антитело IR3 полностью блокирует стимулирующее действие экзогенно добавленного IGF-I и IGF-II в бессывороточных условиях на приблизительно 80%. Антитело IR3 также незначительно ингибирует (менее, чем на 25%) рост клеток MCF-7 в 10% сыворотке (Cul-len, K.J. et al., 1990, Cancer Res., 50, 48-53). Это слабое ингибирование стимулируемого сывороткой роста клеток MCF-7 антителом IR3 in vitro может быть связано с результатами исследования in vivo, в которых обработка антителом IR3 не ингибировала значимо рост ксенотрансплантата MCF-7 в голых мышах (Arteaga, C.L. et al., 1989, J. Clin. Invest., 84, 1418-1423). Вследствие слабых агонистических активностей IR3 и других описанных антител и их неспособности значимо ингибировать рост опухолевых клеток, таких как клетки MCF-7, в более физиологических условиях стимуляции сывороткой (вместо стимуляции экзогенно добавленными IGF-I или IGF-II в бессывороточных условиях) существует потребность в новых антителах против IGF-I-рецепторов, которые значимо ингибируют стимулируемый сывороткой рост опухолевых клеток, но которые сами по себе не обнаруживают значимой агонистической активности. Сущность изобретения Таким образом, целью данного изобретения является обеспечение антител, фрагментов антител и производных антител, которые специфически связываются с инсулин-подобным фактором роста I и ин-гибируют клеточную активность этого рецептора антагонистическим действием на этот рецептор, а также, по существу, лишены агонистической активности в отношении этого рецептора. Таким образом, в первом варианте осуществления обеспечено мышиное антитело ЕМ164, которое полностью охарактеризовано здесь в отношении аминокислотных последовательностей вариабельных областей как его легкой цепи, так и его тяжелой цепи, кДНК-последовательностей генов для вариабельных областей легкой и тяжелой цепи, идентификации его CDR (определяющих комплементарность районов), идентификации его поверхностных аминокислот и способов для его экспрессии в рекомбинантной форме. Во втором варианте осуществления обеспечены имеющие перестроенную поверхность или гуманизированные версии антитела ЕМ164, где экспонированные на поверхности остатки этого антитела или его фрагментов заменены как в легкой, так и в тяжелой цепях для более близкого сходства с известными поверхностями антитела человека. Такие гуманизированные антитела могут иметь увеличенную полезность в сравнении с мышиным ЕМ164, в качестве терапевтических или диагностических агентов. Гуманизированные версии антитела ЕМ164 также полностью охарактеризованы здесь в отношении их соответствующих аминокислотных последовательностей вариабельных областей как легкой, так и тяжелой цепи, последовательностей генов для вариабельных областей легкой и тяжелой цепи, идентификации CDR, идентификации их поверхностных аминокислот и раскрытия способов для их экспрессии в реком-бинантной форме. В третьем варианте осуществления обеспечено антитело, которое способно ингибировать рост раковых клеток на более, чем приблизительно 80%, в присутствии стимулятора роста, такого как, например, сыворотка, инсулин-подобный фактор роста I и инсулин-подобный фактор роста II. В четвертом варианте осуществления обеспечено антитело или фрагмент антитела, имеющие тяжелую цепь, включающую в себя CDR, имеющие аминокислотные последовательности, представленные SEQ ID N0:1-3, соответственно: SYWMH (SEQIDNO:l), EINPSNGRTNYNEKFKR (SEQ ID N0:2), GRPDYYGSSKWYFDV (SEQ ID N0:3); и имеющие легкую цепь, которая содержит CDR, имеющие аминокислотные последовательности, представленные SEQ ID N0:4-6: RSSQSIVHSNVNTYLE (SEQ ID NO:4); KVSNRFS (SEQ ID N0:5); FQGSHVPPT (SEQ ID N0:6). В пятом варианте осуществления обеспечены антитела, имеющие тяжелую цепь, которая имеет аминокислотную последовательность, которая имеет по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью, представленной SEQ ID N0:7: QVQLQQSGAELVKPGASVKLSCKASGYTFTSWMHWVKQRPGQGLEWIGEI NPSNGRTNYHEKFKRKATLTVDKSSSTAYMQLSSLTSEDSAVYYFARGRPDY YGSSKWYFDVWGAGTTVTVSS (SEQ ID NO:7) Сходным образом, обеспечены антитела, имеющие легкую цепь, которая имеет аминокислотную - 3 - 009807 последовательность, которая имеет по меньшей мере 90% идентичность последовательности с аминокислотной последовательностью, представленной SEQ ID N0:8: DVLMTQTPLSLPVSLGDQASISCRSSQSIVHSNVNTYLEWYLQKPGQSPKLLIY KVSNRFSGVPDRFSGSGSGTGFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGTK LEIKR(SEQ ID N0:8). В шестом варианте осуществления обеспечены антитела, имеющие гуманизированную или с перестроенную поверхностью вариабельную область легкой цепи, имеющую аминокислотную последовательность, соответствующую одной из SEQ ID N0:9-12: DWMTQTPLSLPVSLGDPASISCRSSQSIVHSNVNTYLEWYLQKPGQSPRLLIY KVSNRFSGVPDRFSGSGAGTDFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGT KLEIKR (SEQ ID N0:9); DVLMTQTPLSLPVSLGDPASISCRSSQSrVHSNVNTYLEWYLQKPGQSPKLLIY KVSNRFSGVPDRFSGSGAGTDFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGT KLEIKR (SEQ ID NO: 10); DVLMTQTPLSLPVSLGDPASISCRSSQSIVHSNVNTYLEWYLQKPGQSPRLLIY KVSNRFSGVPDRFSGSGAGTDFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGT KLEIKR (SEQ ID NO: 11); или DWMTQTPLSLPVSLGDPASISCRSSQSWHSNVNTYLEWYLQKPGQSPKLLIY KVSNRFSGVPDRFSGSGAGTDFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGT KLEIKR (SEQ ED NO: 12). Сходным образом, обеспечены антитела, имеющие гуманизированную или с перестроенной поверхностью вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, соответствующую SEQ ID N0:13: QVQLVQSGAEVVKPGASVKLSCKASGYTFTSYWMHWVKQRPGQGLEWIGEI NPSNGRTNYNQKFQGKATLTVDKSSSTAYMQLSSLTSEDSAVYYFARGRPDY YGSSKWYFDVWGQGTTVTVSS (SEQ ID NO: 13). В седьмом варианте осуществления обеспечены антитела или фрагменты антител данного изобретения, которые имеют улучшенные свойства. Например, антитела или фрагменты антител, имеющие улучшенную аффинность в отношении IGF-I-рецептора, получают созреванием аффинности антитела или фрагмента данного изобретения. Данное изобретение обеспечивает, далее, конъюгаты указанных антител, где цитотоксический агент ковалентно присоединен, непосредственно или через расщепляемый или нерасщепляемый линкер, к антителу или эпитопсвязывающему фрагменту антитела данного изобретения. В предпочтительных вариантах осуществления этим цитотоксическим агентом является таксол, майтансиноид, СС-1065 или аналог СС-1065. Далее данное изобретение обеспечивает антитела или их фрагменты, которые дополнительно помечены для использования в исследовательских или диагностических приложениях. В предпочтительных вариантах осуществления этой меткой является радиоактивная метка, флуорофор, хромофор, визиогра-фический агент или ион металла. Обеспечен также способ диагностики, в котором указанные меченые антитела или фрагменты вводят субъекту, в котором подозревают наличие рака, и измеряют или подвергают мониторингу распределение этой метки в теле этого субъекта. В восьмом варианте осуществления данное изобретение обеспечивает способы лечения субъекта, имеющего рак, введением антитела, фрагмента антитела или конъюгата антитела данного изобретения, отдельно или в комбинации с другими цитотоксическими или терапевтическими агентами. Этот рак может быть, например, одним или несколькими из рака молочной железы, рака ободочной кишки, рака яичника, остеосаркомы, рака шейки матки, рака предстательной железы, рака легкого, синовиального рака, рака поджелудочной железы или другого рака, который еще должен быть определен, в котором уровни IGF-I-рецептора являются повышенными. В девятом варианте осуществления данное изобретение обеспечивает способы лечения субъекта, имеющего рак, введением антитела, фрагмента антитела или конъюгата антитела данного изобретения, отдельно или в комбинации с другими цитотоксическими или терапевтическими агентами. В частности, предпочтительные цитотоксические и терапевтические агенты включают в себя доцетаксел, паклитаксел, доксорубицин, эпирубицин, циклофосфамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торе-мифен, летрозол, анастрозол, фулвестрант, экземестан, госерелин, оксалиплатин, карбоплатин, циспла - 4 - 009807 тин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, это-позид, топотекан, гемцитабин, винорелбин, эстрамустин, митоксантрон, абареликс, золедронат, стрепто-зоцин, ритуксимаб (Rituxan), идарубицин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2^ мелфолам, бортезомиб (Velcade), алтретамин, аспарагиназу, гефитиниб (Iressa), эрлонитиб (Tarceva), анти-EGF-рецептор-антитело (Cetuximab, Abx-EGF) и эпотилон. Более предпочтительно, этим терапевтическим агентом является содержащий платину агент (такой как карбоплатин, оксалиплатин, цисплатин), таксан (такой как паклитак-сел, доцетаксел), гемцитабин или камптотецин. Раком может быть, например, одно или более из раковых заболеваний, таких как рак молочной железы, рак ободочной кишки, рак яичника, остеосаркома, рак шейки матки, рак предстательной железы, рак легкого, синовиальный рак, рак поджелудочной железы, меланома, множественная миелома, нейроб-ластома и рабдомиосаркома или другой рак, который еще должен быть определен, в котором уровни IGF-I-рецептора являются повышенными. В десятом варианте осуществления данное изобретение обеспечивает наборы, содержащие один или несколько описанных здесь элементов и инструкции в отношении использования этих элементов. В предпочтительном варианте осуществления, набор данного изобретения включает в себя антитело, фрагмент или конъюгат антитела данного изобретения и терапевтический агент. Инструкции для этого предпочтительного варианта осуществления включают в себя инструкции в отношении ингибирования роста раковых клеток с использованием антитела, фрагмента или конъюгата антитела данного изобретения и терапевтического агента и/или инструкции в отношении способа лечения пациента, имеющего рак, с использованием антитела, фрагмента или конъюгата антитела данного изобретения и терапевтического агента. Краткое описание фигур Фиг. 1 показывает анализ с использованием клеточного сортинга с возбуждением флуоресценции (FACS) специфического связывания очищенного антитела ЕМ164 с клетками, сверхэкспрессирующими IGF-I-рецептор Y1251F или рецептор инсулина человека; Фиг. 2 - кривую титрования связывания для связывания антитела ЕМ164 с биотинилированным IGF-I-рецептором человека. Фиг. 3 - ингибирование связывания биотинилированного IGF-I-рецептора человека с клетками MCF-7 рака молочной железы человека антителом ЕМ164. Фиг. 4 - ингибирование IGF-I-стимулированного автофосфорилирования IGF-I-рецептора в клетках MCF-7 антителом ЕМ164. Фиг. 5 - ингибирование IGF-I-стимулированного IRS-1-фосфорилирования в клетках MCF-7 антителом ЕМ164. Фиг. 6 - ингибирование IGF-I-рецептор-стимулированной трансдукции сигнала в клетках SaOS-2 антителом ЕМ164. Фиг. 7 - действие антитела ЕМ164 на рост и выживание клеток MCF-7 при различных условиях роста, оцененное при помощи МТТ-анализа. Фиг. 8 - действие антитела ЕМ164 на рост и выживание клеток MCF-7 в присутствии различных концентраций сыворотки. Фиг. 9 - ингибирование стимулированного IGF-I-и сывороткой роста и выживания клеток NCI-H838 антителом ЕМ164. Фиг. 10 - действие обработки антителом ЕМ164, таксолом или комбинацией антитела ЕМ164 и так-сола на рост ксенотрансплантата рака легкого Calu-6 в мышах. Фиг. 11 - конкуренцию между связыванием гуманизированного антитела ЕМ164 (v.1.0) и мышиного антитела ЕМ164. Фиг. 12 - кДНК- (SEQ ID NO:49) и аминокислотную (SEQ ID NO:50) последовательности лидерной области и вариабельной области легкой цепи мышиного антитела против IGF-I-рецептора ЕМ 164. Стрелка показывает начало каркаса 1. Три CDR-последовательности в соответствии с Kabat подчеркнуты. Фиг. 13 - кДНК- (SEQ ID NO:51) и аминокислотную (SEQ ID NO:52) последовательности лидерной области и вариабельной области тяжелой цепи мышиного антитела против IGF-I-рецептора ЕМ 164. Стрелка показывает начало каркаса 1. Три CDR-последовательности в соответствии с Kabat подчеркнуты. Фиг. 14 показывает аминокислотные последовательности CDR легкой и тяжелой цепей антитела ЕМ164, определенные на основании определений канонического класса Chothia. Показаны также определения программы AbM для CDR тяжелой цепи. Легкая цепь: CDR1 является SEQ ID NO:4, CDR2 является SEQ ID NO:5 и CDR 3 является SEQ ID NO:6. Тяжелая цепь: CDR1 является SEQ ID NO:1, CDR2 является SEQ ID NO:2 и CDR3 является SEQ ID NO:3. Тяжелая цепь AbM: CDR1 является SEQ ID NO:53, CDR2 является SEQ ID NO:54 и CDR3 является SEQ ID NO:55. Фиг. 15 показывает аминокислотные последовательности легкой цепи и тяжелой цепи для антитела против IGF-I-рецептора ЕМ164, выровненные с последовательностями зародышевой линии для генов Cr1 - 5 - 009807 (SEQ ID NO:56) и J558.C (SEQ ID NO:57). Пунктир (-) указывает идентичность последовательностей. Фиг. 16 показывает плазмиды, используемые для конструирования и экспрессии рекомбинантных химерных и гуманизированных антител ЕМ 164. А) клонирующая плазмида для легкой цепи, В) клонирующая плазмида для тяжелой цепи, С) экспрессионная плазмида для антитела млекопитающего. Фиг. 17 показывает 10 наиболее гомологичных аминокислотных последовательностей легких цепей, полученных скринингом из 127 антител в наборе файлов структур, используемых для предсказания поверхностных остатков ЕМ164. em164 LC (SEQ ID NO:58), 2jel (SEQ ID NO:59), 2pep (SEQ ID NO:60), 1ngb (SEQ ID NO:61), 1kel (SEQ ID NO:62), 1hyx (SEQ ID NO:63), 1igf (SEQ ID NO:64), 1tet c(SEQ ID NO:65), 1clz (SEQ ID NO:66), 1bln (SEQ ID NO:67, 1cly (SEQ ID NO:68), Консенсус (SEQ ID NO:69). Фиг. 18 показывает 10 наиболее гомологичных аминокислотных последовательностей легких цепей, полученных скринингом из 127 антител в наборе файлов структур, используемых для предсказания поверхностных остатков ЕМ164. em164 НС (SEQ ID NO:70), 1ngb (SEQ ID NO:71), 1ngp (SEQ ID NO:72), 1fbi (SEQ ID NO:73), 1afv (SEQ ID NO:74), 1yuh (SEQ ID NO:75), 1plg (SEQ ID NO:76), 1d5b (SEQ ID NO:77), 1ae6 (SEQ ID NO:78), 1axs (SEQ ID NO:79), 3hf1 (SEQ ID NO:80), Консенсус (SEQ ID NO:81). Фиг. 19 - среднюю доступность для каждого из остатков вариабельной области легкой (А) и тяжелой (В) цепей из 10 наиболее гомологичных структур. Числа представляют номера положений последовательности антитела по Kabat. Фиг. 20 - аминокислотные последовательности вариабельной области легкой цепи для мышиного ЕМ164 (muEM164) и гуманизированного ЕМ164 (huEM164) антител. muEM164 (SEQ ID NO:82), huEM164 V1.0 (SEQ ID NO:83), huEM164 VI.1 (SEQ ID NO:84), huEM164 V1.2 (SEQ ID NO:85), huEM164 V1.3 (SEQ ID NO:86). Фиг. 21 - аминокислотные последовательности вариабельной области тяжелой цепи для мышиного ЕМ164 (muEM164, SEQ ID NO:87) и гуманизированного ЕМ164 (huEM164, SEQ ID NO:88) антител. Фиг. 22 - ДНК-последовательность и аминокислотную последовательность вариабельной области huEM164 v1.0 как для легкой (ДНК, SEQ ID NO:89, аминокислотная SEQ ID NO:90), так и для тяжелой (ДНК, SEQ ID NO:91, аминокислотная SEQ ID NO:92). Фиг. 23 - ДНК-последовательность и аминокислотную последовательность вариабельной области легкой цепи для гуманизированного ЕМ164 v1.1 (ДНК, SEQ ID NO:93, аминокислотная SEQ ID NO:94), v1.2 (ДНК, SEQ ID NO:95, аминокислотная SEQ ID NO:96) и v1.3 (ДНК, SEQ ID NO:97, аминокислотная SEQ ID NO:98). Фиг. 24 - ингибирование IGF-I-стимулируемого роста и выживания клеток MCF-7 гуманизированным антителом ЕМ164 v1.0 и мышиным антителом: ЕМ164. Фиг. 25 показывает, что ЕМ164 подавляет IGF-I-стимулированное прохождение клеточного цикла клеток MCF-7. Фиг. 26 показывает, что ЕМ 164 подавляет антиапоптотическое действие IGF-I и сыворотки. Обработка ЕМ164 приводит к апоптотической смерти клеток, как показано увеличенными уровнями расщепленного белка СК18. Фиг. 27 показывает действие обработки антителом ЕМ164, гемцитабином или комбинацией антитела ЕМ164 и гемцитабина на рост ксенотрансплантатов рака поджелудочной железы человека ВхРС-3 в иммунодефицитных мышах. Сведения, подтверждающие возможность осуществления изобретения Данное изобретение обнаружило и улучшило новые антитела, которые специфически связываются с рецептором инсулин-подобного фактора роста I (IGF-IR) человека на поверхности клеток. Эти антитела и фрагменты имеют уникальную способность ингибировать клеточные функции этого рецептора без способности активации самого рецептора. Таким образом, в то время как известные ранее антитела, которые специфически связывают и ингибируют IGF-IR, также активируют рецептор даже в отсутствие лигандов IGF-IR, антитела или фрагменты данного изобретения являются антагонистами IGF-IR, но, по существу, лишены агонистической активности. Кроме того, антитела и фрагменты антител данного изобретения ингибируют рост опухолевых клеток человека, таких как клетки MCF-7, в присутствии сыворотки более, чем на 80%, что является более высокой степенью ингибирования, чем степень ингибирова-ния, получаемая с использованием ранее известных антител против IGF-I-рецептора. Данное изобретение начинается от мышиного антитела против IGF-I-рецептора, здесь ЕМ164, которое полностью охарактеризовано в отношении аминокислотных последовательностей как легкой, так и тяжелой цепей, идентификации CDR, идентификации поверхностных аминокислот и способа для его экспрессии в рекомбинантной форме. Последовательности зародышевой линии показаны на фиг. 15, выровненные с последовательностью ЕМ164. Это сравнение идентифицирует возможные соматические мутации в EM164, в том числе одну из каждых мутаций в CDR 1 в легкой цепи и в CDR 2 в тяжелой цепи. Здесь описаны первичные аминокислотная последовательность и ДНК-последовательность легкой и тяжелой цепей антитела ЕМ164 и гуманизированных версий. Однако объем данного изобретения не ограничивается антителами и фрагментами, содержащими эти последовательности. Вместо этого, все антитела и фрагменты, которые специфически связываются с рецептором инсулин-подобного фактора - 6 - 009807 роста I и противодействуют биологической активности этого рецептора, но которые, по существу, лишены агонистической активности, попадают в объем данного изобретения. Таким образом, антитела и фрагменты антител могут отличаться от антитела ЕМ164 или гуманизированных производных по аминокислотной последовательности их каркаса, CDR, легкой цепи и тяжелой цепи, но все еще попадать в объем данного изобретения. CDR антитела ЕМ164 идентифицированы моделированием и их молекулярные структуры были предсказаны. Опять, хотя эти CDR являются важными для узнавания эпитопов, они не являются незаменимыми для антител и фрагментов данного изобретения. Таким образом, обеспечены антитела и фрагменты, которые имеют улучшенные свойства, образованные, например, созреванием аффинности антитела данного изобретения. Разнообразные антитела и фрагменты антител, а также миметики антител могут быть легко получены посредством мутации, делеции и/или инсерции в последовательностях вариабельной и константной областей, которые фланкируют конкретный набор CDR. Так, например, возможны различные классы Ab для конкретного набора CDR, посредством замещения различных тяжелых цепей, посредством чего могут быть получены, например, типы и изотипы антител IgG1-4, IgM, IgA1-2, IgD, IgE. Подобным образом, могут быть получены искусственные антитела в объеме данного изобретения посредством помещения конкретного набора CDR в полностью синтетический каркас. Термин "вариабельные" используется здесь для описания определенных частей вариабельных доменов, которые различаются по их последовательности среди антител и используются в связывании и специфичности каждого конкретного антитела в отношении его антигена. Однако эта вариабельность не распределяется обычно равномерно по всем вариабельным доменам этих антител. Обычно она сконцентрирована в трех сегментах, называемых определяющими комплементраность районами (CDR) или гипервариабельными районами в вариабельных доменах как легкой, так и тяжелой цепи. Более высококонсервативные части вариабельных доменов называют каркасом (FR). Вариабельные домены тяжелой и легкой цепей, каждый, содержат четыре каркасных района, принимающих в значительной степени конфигурацию бета-складок, соединенных тремя CDR, которые образуют петли, соединяющие бета-складчатую структуру и в некоторых случаях образующие часть бета-складчатой структуры. CDR в каждой цепи удерживаются вместе в тесной близости посредством FR-районов и, с CDR из другой цепи, способствуют образованию антигенсвязывающего сайта антител (Е.А. Kabat et al., Sequences of Proteins of Immunological Interest, fifth edition, 1991, NIH). Константные домены не участвуют непосредственно в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие этого антитела в антителозависимой клеточной токсичности. Гуманизированные антитела или антитела, адаптированные в отношении неотторжения другими млекопитающими, могут быть получены с использованием нескольких технологий, таких как перестройка поверхности антител и трансплантация CDR. В технологии перестройки поверхности, молекулярное моделирование, статистический анализ и мутагенез объединяют для доведения не-CDR-поверхностей вариабельных областей до сходства с поверхностями известных антител хозяина-мишени. Стратегии и способы для переделывания поверхностей антител и другие способы для уменьшения иммуногенности антител в отличающемся хозяине описаны в Патенте США 5639641, который включен здесь в его полном виде в качестве ссылки. В технологии трансплантации CDR, CDR мышиных тяжелой и легкой цепей трансплантируют в полностью человеческую каркасную последовательность. Данное изобретение включает в себя также функциональные эквиваленты антител, описанных в данном описании. Функциональные эквиваленты имеют характеристики связывания, которые сравнимы с характеристиками связывания этих антител, и включают в себя, например, химерные, гуманизированные и одноцепочечные антитела, а также их фрагменты. Способы получения таких функциональных эквивалентов описаны в Заявке РСТ WO 93/21319, Заявке на Европейский патент № 239400; Заявке РСТ WO 89/09622; Заявке на Европейский патент 338745 и Заявке на Европейский патент 332424, которые включены в их полном объеме путем отсылки. Функциональные эквиваленты включают в себя полипептиды с аминокислотными последовательностями, по существу такими же, что и аминокислотная последовательность вариабельных или гипервариабельных районов антител данного изобретения. "По существу такая же" в применении к аминокислотной последовательности определяется здесь как последовательность по меньшей мере с приблизительно 90% и более предпочтительно по меньшей мере приблизительно 95% идентичностью последовательности относительно другой аминокислотной последовательности, определяемой при помощи способа поиска FASTA в соответствии с Pearson and Lipman, Proc. Natl. Acad. Sci. USA 85, 2444-2448 (1988). Химерные антитела предпочтительно имеют константные области, произведенные, по существу, или исключительно из константных областей антитела человека, а вариабельные области, произведенные по существу или исключительно из последовательности вариабельной области из млекопитающего, другого, чем человек. Гуманизированные формы этих антител получают замещением определяющих ком-плементарность районов, например, мышиного антитела, в каркасный домен человека, например, см. публикацию РСТ № WO92/22653. Гуманизированные антитела предпочтительно имеют константные области и вариабельные области, другие, чем определяющие комплементарность районы (CDR), полу - 7 - 009807 ченные, по существу, или исключительно из соответствующих районов антител человека, a CDR, полученные, по существу, или исключительно из млекопитающего, другого, чем человек. Функциональные эквиваленты включают в себя также одноцепочечные фрагменты антител, также известные как одноцепочечные антитела (scFv). Эти фрагменты содержат по меньшей мере один фрагмент вариабельной аминокислотной последовательности тяжелой цепи антитела (VH), присоединенный по меньшей мере к одному фрагменту вариабельной последовательности легкой цепи антитела (VL) с одним или несколькими соединяющими их линкерами. Такой линкер может быть коротким, гибким пептидом, выбранным для гарантии того, что образуется правильная трехмерная укладка доменов (VL) и (VH) при их связывании, так чтобы сохранялась специфичность связывания молекулы-мишени целого антитела, из которого получен этот одноцепочечный фрагмент антитела. Обычно карбоксильный конец последовательности (VL) или (VH) может быть ковалентно связан таким пептидным линкером с аминоконцом комплементарной последовательности (VL) и (VH). Одноцепочечные фрагменты антител могут быть генерированы молекулярным клонированием с использованием библиотеки фагового дисплея антител или сходными способами. Эти белки могут быть продуцированы либо в эукариотических клетках, либо в прокариотических клетках, в том числе бактериях. Одноцепочечные фрагменты антител содержат аминокислотные последовательности, имеющие по меньшей мере один из вариабельных или определяющих комплементарность районов (CDR) целых антител, описанных в этом описании, но лишены некоторых или всех константных областей этих антител. Эти константные области не являются обязательными для связывания антигена, но составляют основную часть структуры целых антител. Одноцепочечные фрагменты антител могут, следовательно, преодолевать некоторые из проблем, связанных с применением антител, содержащих часть константной области или всю константную область. Например, одноцепочечные фрагменты антител имеют тенденцию быть свободными от нежелательных взаимодействий между биологическими молекулами и константной областью тяжелой цепи или от другой нежелательной биологической активности. Кроме того, одноцепочеч-ные фрагменты антител являются значительно меньшими, чем целые антитела и могут, следовательно, иметь более высокую капиллярную проникаемость, чем целые антитела, позволяющую одноцепочечным фрагментам более эффективно определять местонахождение антигенсвязывающих сайтов-мишеней и связываться с ними. Фрагменты антител могут в относительно большом масштабе продуцироваться в прокариотических клетках, что облегчает их получение. Кроме того, относительно малый размер одно-цепочечных фрагментов приводит к тому, что они с меньшей вероятностью индуцируют иммунную реакцию в реципиенте, чем целые антитела. Функциональные эквиваленты дополнительно включают в себя фрагменты антител, которые имеют такие же или сравнимые характеристики связывания, что и характеристики связывания целого антитела. Такие фрагменты могут содержать один или оба Fab-фрагмента или F(ab')2-фрагмент. Предпочтительно, эти фрагменты антител содержат все шесть определяющих комплементарность районов целого антитела, хотя фрагменты, содержащие меньше, чем все такие районы, например, три, четыре или пять CDR, также являются функциональными. Далее функциональные эквиваленты могут быть членами или могут быть объединены с членами следующих классов иммуноглобулинов: IgG, IgM, IgA, IgD или IgE и их подклассов. Знание аминокислотных последовательностей и последовательностей нуклеиновых кислот для антител против IGF-I-рецептора ЕМ 164 и его гуманизированных вариантов, которые описаны здесь, может быть использовано для развития других антител, которые также связываются с IGF-I-рецептором человека и ингибируют клеточные функции этого IGF-I-рецептора. Некоторые исследования изучали действие введения одной или нескольких аминокислотных замен в различных положениях в последовательности антитела, на основе знания первичной последовательности антитела, на такие его свойства, как связывание и уровень экспрессии (Yang, W.P. et al., 1995, J. Mol. Biol., 254, 392-403; Rader, С. et al., Proc. Natl. Acad. Sci. USA, 95, 8910-8915; Vaughan, T.J. et al., 1998, Nature Biotechnology, 16, 535-539). В этих исследованиях варианты первичного антитела генерировали изменением последовательностей генов тяжелой и легкой цепей в районах CDR1, CDR2, CDR3 или каркасных районах с использованием таких способов, как олигонуклеотид-опосредованный сайт-направленный мутагенез, кассетный мутагенез, склонная к ошибкам ПЦР, перетасовка ДНК или мутаторные штаммы Е. coli (Vaughan, T.J. et al., 1998, Nature Biotechnology, 16, 535-539; Adey, N.B. et al., 1996, Chapter 16, pp. 277-291, in "Phage Display of Peptides and Proteins", Eds. Kay, B.K. et al., Academic Press). Эти способы изменения последовательности первичного антитела привели к улучшенным аффинностям вторичных антител (Gram, H. et al., 1992, Proc. Natl. Acad. Sci. USA, 89, 3576-3580; Boder, E.T. et al., 2000, Proc. Natl. Acad. Sci. USA, 97, 10701-10705; Davies, J. and Riechmann, L., 1996, Immunology, 2, 169-179; Thompson, J. et al., 1996, J. Mol. Biol., 256, 77-88; Short, M.K. et al., 2002, J. Biol. Chem., 277, 16365-16370; Furukawa, K. et al, 2001, J. Biol. Chem., 276, 27622-27628). С использованием подобной направленной стратегии изменения одного или нескольких аминокислотных остатков антитела, последовательности антител, описанные в данном изобретении, могут быть использованы для развития антител против IGF-I-рецептора с улучшенными функциями. Конъюгаты данного изобретения содержат антитело, фрагменты и их аналоги, описанные здесь, - 8 - 009807 связанные с цитотоксическим агентом. Предпочтительными цитотоксическими агентами являются май-тансиноиды, таксаны и аналоги СС-1065. Эти конъюгаты могут быть получены способами in vitro. Для связывания цитотоксического агента с антителом используют связывающую группу. Подходящие связывающие группы хорошо известны в данной области и включают в себя дисульфидные группы, простые тиоэфирные группы, неустойчивые к кислоте группы, фотолабильные группы, пептидаза-лабильные группы и эстераза-лабильные группы. Предпочтительными связывающими группами являются дисуль-фидные группы и тиоэфирные группы. Например, могут быть сконструированы конъюгаты с использованием реакции дисульфидного обмена или посредством образования простой тиоэфирной связи между антителом и цитотоксическим агентом. Майтансиноиды и аналоги майтансиноидов находятся среди предпочтительных цитотоксических агентов. Примеры подходящих майтансиноидов включают в себя майтансинол и аналоги майтансинола. Подходящие майтансиноиды описаны в патентах США с номерами 4424219; 4256746; 4294767; 4307016; 4313946; 4315929; 4331598; 4361650; 4362663; 4364866; 4450254; 4322348; 4371533; 6333410; 5475092; 5585499 и 5846545. Таксаны также являются предпочтительными цитотоксическими агентами. Таксаны, подходящие для использования в данном изобретении, описаны в Патентах США с номерами 6372738 и 6340701. СС-1065 и его аналоги являются также предпочтительными цитотоксическими лекарственными средствами для использования в данном изобретении. СС-1065 и его аналоги описаны в Патентах США с номерами 6372738; 6340701; 5846545 и 5585499. Привлекательным кандидатом на получение таких цитотоксических конъюгатов является СС-1065, который является сильным противоопухолевым антибиотиком, выделенным из культурального бульона Streptomyces zelensis. СС-1065 является приблизительно в 1000 раз более сильным, чем обычно используемые антираковые лекарственные средства, такие как доксорубицин, метотрексат и винкристин (ВХ. Bhuyan et al., Cancer Res., 42, 3532-3537 (1982)). Цитотоксические лекарственные средства, такие как метотрексат, даунорубицин, доксорубицин, винкристин, винбластин, мелфалан, митомицин С, хлорамбуцил и калихеамицин, также являются подходящими для приготовления конъюгатов данного изобретения, и эти молекулы лекарственных средств могут быть также связаны с молекулами антител через промежуточную молекулу-носитель, такую как сывороточный альбумин. Для диагностических приложений антитела данного изобретения обычно метят детектируемой молекулой. Детектируемой молекулой может быть любая молекула, которая способна продуцировать, прямо или опосредованно, детектируемый сигнал. Например, детектируемой молекулой может быть радио- 3 14 32 35 131 изотоп, такой как 3Н, 14С, 32Р, 35S или 131I; флуоресцентное или хемилюминесцентное соединение, такое как изотиоцианат флуоресцеина, родамин или люциферин; или фермент, такой как щелочная фосфатаза, бета-галактозидаза или пероксидаза хрена. Любой способ, известный в данной области, может быть использован для конъюгирования антитела с детектируемой молекулой, в том числе способы, описанные Hunter, et al., Nature 144:945 (1962); David, et al., Biochemistry 13:1014 (1974); Pain, et al, J. Immunol. Meth. 40:219 (1981) и Nygren, J. Histochem. and Cytochem. 30:407 (1982). Антитела данного изобретения могут быть использованы в любом известном способе анализа, таком как конкурентные анализы связывания, прямые или опосредованные сэндвич-анализы и иммунопре-ципитационные анализы (Zola, Monoclonal Antidodies: A Manual of Techniques, pp. 47-158 (CRC Press, Inc., 1987)). Антитела данного изобретения применимы также для визиографии in vivo, где антитело, меченое детектируемой молекулой, такой как непрозрачный для излучения агент или радиоактивный изотоп, вводят субъекту, предпочтительно в кровоток, и анализируют присутствие или местонахождение этого меченого антитела в хозяине. Этот способ визиографии применим в определении стадии и лечении злока-чественностей. Антитело может быть помечено любой молекулой, которая является детектируемой в хозяине, при помощи ядерного магнитного резонанса, радиологии или других средств детектирования, известных в данной области. Антитела данного изобретения применимы также в качестве агентов аффинной очистки. В этом способе антитела иммобилизуют на подходящем носителе, таком как смола Сефадекс или фильтровальная бумага, с использованием способов, хорошо известных в данной области. Антитела данного изобретения применимы также в качестве реагентов в биологических исследованиях, основанных на их ингибировании функции IGF-I-рецептора в клетках. Для терапевтических применений антитела и конъюгаты данного изобретения вводят субъекту в фармацевтически приемлемой дозированной форме. Они могут быть введены внутривенно в виде болюса или непрерывной инфузией на протяжении некоторого периода времени, внутримышечным, подкожным, внутрисуставным, интрасиновиальным, интратекальным, пероральным, местным или ингаляционным способами. Антитело может также вводиться внутриопухолевым, околоопухолевым способами, способом введения в повреждение или около повреждения, для проявления локальных, а также системных терапевтических действий. Подходящие фармацевтически приемлемые носители, разбавители и - 9 - 009807 эксципиенты хорошо известны и могут быть определены специалистами в данной области, как того может потребовать клиническая ситуация. Примеры подходящих носителей, разбавителей и/или эксципи-ентов включают в себя: (1) забуференный фосфатом солевой раствор Дульбекко, рН приблизительно 7,4, содержащий около 1 мг/мл - 25 мг/мл сывороточного альбумина человека, (2) 0,9% солевой раствор (0,9% мас./об. NaCl) и (3) 5% (мас./об.) декстроза. Способ данного изобретения может практиковаться in vitro, in vivo или ex vivo. В других терапевтических обработках антитела, фрагменты или конъюгаты антител данного изобретения вводят совместно или вводят последовательно с одним или несколькими дополнительными терапевтическими агентами. Подходящие терапевтические агенты включают в себя, но не ограничиваются ими, цитотоксические или цитостатические агенты. Таксол является предпочтительным терапевтическим агентом, который является также цитотоксическим агентом. Противораковыми терапевтическими агентами являются такие агенты, которые стремятся убить или ограничить рост раковых клеток, вызывая лишь минимальное повреждение хозяина. Таким образом, такие агенты могут использовать любое отличие в свойствах раковых клеток (например, метаболизм, васкуляризацию или презентацию антигенов клеточной поверхности) от свойств здоровых клеток хозяина. Различия в морфологии опухоли являются потенциальными сайтами для вмешательства: например, вторым терапевтическим веществом может быть такое антитело, как антитело против VEGF, которое применимо в замедлении васкуляризации внутренней части солидной опухоли, которое замедляет скорость роста опухоли. Другие терапевтические агенты включают в себя, но не ограничиваются ими, дополнительные средства, такие как гранистерон, HCl, ингибиторы андрогена, такие как ацетат лейпроли-да, антибиотики, такие как доксорубицин, антиэстрогены, такие как тамоксифен, антиметаболиты, такие как интерферон альфа-2а, цитотоксические агенты, такие как таксол, ингибиторы ферментов, такие как ингибитор фарнезилтрансферазы, иммуномодуляторы, такие как алдеслейкин и производные азотных аналогов горчичного газа (иприта), такие как мелфалан, HCl и т.п. Терапевтические агенты, которые могут быть комбинированы с ЕМ164 для улучшенной противораковой эффективности, включают в себя различные агенты, используемые в практике онкологии (Ссылка: Cancer, Principles & Practice of Oncology, DeVita, V.T., Helman, S., Rosenberg, S.A., 6th edition, Lippencott-Raven, Philadelphia, 2001), такие как доцетаксел, паклитаксел, доксорубицин, эпирубицин, циклофосфа-мид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, аиастрозол, фулвестрант, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорел-бин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идаруби-цин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2^ мелфолам, бортезомиб (Velcade), алтретамин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), анти-EGF-рецептор-антитело (Cetuxivmab, Abx-EGF) и эпотилоны и конъю-гаты цитотоксических лекарственных средств и антител против рецепторов клеточной поверхности. Предпочтительными терапевтическими агентами являются содержащие платину агенты (такие как кар-боплатин, оксалиплатин, цисплатин), таксаны (такие как паклитаксел, доцетаксел), гемцитабин или кам-птотецин. Один или несколько дополнительных терапевтических агентов могут вводиться до, одновременно или после антитела, фрагмента или конъюгата антитела, данного изобретения. Специалисту с квалификацией в данной области будет понятно, что для каждого терапевтического агента могут быть преимущества в отношении конкретного порядка введения. Подобным образом, специалисту с квалификацией в данной области будет понятно, что для каждого терапевтического агента продолжительность интервалов, с которыми вводят каждый агент и антитело, фрагмент или конъюгат антитела данного изобретения, будет варьировать. Хотя специалисту с квалификацией в данной области будет понятно, что доза каждого терапевтического агента будет зависеть от конкретного агента, предпочтительные дозы могут находиться в диапазоне от приблизительно 10 до приблизительно 2000 мг/кв. м, более предпочтительно от приблизительно 50 до приблизительно 1000 мг/кв. м. Для предпочтительных агентов, таких как содержащие платину агенты (карбоплатин, оксалиплатин, цисплатин), предпочтительная доза равна приблизительно 10 - приблизительно 400 мг/кв. м, для таксанов (паклитаксела, доцетаксела) предпочтительно доза равна приблизительно 20 - приблизительно 150 мг/кв.м, для гемцитабина предпочтительная доза равна приблизительно 100 -приблизительно 2000 мг/кв.м и для камптотецина предпочтительная доза равна приблизительно 60 мг/кв. м - приблизительно 350 мг/кв.м. Доза этих и других терапевтических агентов может зависеть от того, вводят ли антитело, фрагмент или конъюгат антитела данного изобретения одновременно или последовательно с терапевтическим агентом. Введение антитела, фрагмента или конъюгата антитела данного изобретения и одного или нескольких дополнительных терапевтических агентов, независимо от того вводят ли их совместно или вводят последовательно, может выполняться, как описано выше для терапевтических применений. Подходящие фармацевтически приемлемые носители, разбавители и эксципиенты для совместного введения будут известны специалисту с квалификацией в данной области и будут зависеть от идентичности конкретного - 10 - 009807 терапевтического агента, который вводят совместно. Когда антитело присутствует в водной дозированной форме, а не в лиофилизированной форме, это антитело готовят обычно в концентрации приблизительно 0,1-100 мг/мл, хотя допускается широкое варьирование за пределами этих диапазонов. Для лечения заболевания подходящая доза антитела или конъюгата будет зависеть от типа подлежащего лечению заболевания, как определено выше, тяжести и протекания этого заболевания, от того, вводят ли эти антитела для профилактических или терапевтических целей, хода предыдущей терапии, клинической истории пациента и реакции на это антитело и от усмотрения лечащего врача. Предпочтительно, это антитело вводят пациенту один раз или на протяжении ряда введений. В зависимости от типа и тяжести заболевания, предпочтительно начальной дозой для введения пациенту является доза от приблизительно 1 до приблизительно 2000 мг/кв.м антитела, более предпочтительно от приблизительно 10 до приблизительно 1000 мг/кв.м антитела, например, посредством одного или нескольких отдельных введений или непрерывной инфузией. Для повторяемых введений на протяжении нескольких дней или более продолжительно, в зависимости от состояния, введение повторяют, пока не наблюдается желаемая супрессия симптомов заболевания. Однако могут быть использованы и не исключены и другие схемы введения доз. Данное изобретение включает в себя также наборы, содержащие один или несколько описанных здесь элементов и инструкции в отношении использования этих элементов. В предпочтительном варианте осуществления, набор данного изобретения включает в себя антитело, фрагмент или конъюгат антитела данного изобретения и терапевтический агент. Инструкции для этого предпочтительного варианта осуществления включают в себя инструкции в отношении ингибирования роста раковых клеток с использованием антитела, фрагмента или конъюгата антитела данного изобретения и терапевтического агента и/или инструкции в отношении способа лечения пациента, имеющего рак, с использованием антитела, фрагмента или конъюгата антитела данного изобретения, и терапевтического агента. Предпочтительно антитело, используемое в этом наборе, имеет ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (АТСС accession number PTA-4457), или это антитело является его эпитопсодержащим фрагментом, причем как антитело, так и этот фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I. Антителом и фрагментом антитела, используемыми в этом наборе, могут быть также переделанная в отношении поверхности антитела версия антитела ЕМ164, имеющая по меньшей мере одну нуклеотидную мутацию, делецию или инсерцию. Антитела и фрагменты антител каждой из этих трех версий сохраняют ту же самую специфичность связывания, которую имеет антитело ЕМ164. Предпочтительно, терапевтический агент, используемый в этом наборе, выбран из группы, состоящей из таких терапевтических агентов, как доцетаксел, паклитаксел, доксорубицин, эпирубицин, цикло-фосфамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, анастрозол, фулве-странт, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорел-бин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идаруби-цин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2^ мелфолам, бортезомиб (Velcade), алтретамин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), анти-EGF-рецептор-антитело (Cetuxivmab, Abx-EGF) и эпотилон. Более предпочтительно, этим терапевтическим агентом является содержащий платину агент (такой как карбоп-латин, оксалиплатин, цисплатин), таксан (такой как паклитаксел, доцетаксел), гемцитабин или камптоте-цин. Элементы наборов данного изобретения находятся в подходящей для набора форме, например, в форме раствора или лиофилизированного порошка. Квалифицированному специалисту будет понятно, что концентрация или количество этих элементов наборов будут варьировать в зависимости от идентичности и предполагаемого использования каждого элемента этого набора. Типы рака и клеток из этих типов рака, на которые делается ссылка в инструкциях наборов, включают в себя рак молочной железы, рак ободочной кишки, рак яичника, остеосаркому, рак шейки матки, рак предстательной железы, рак легкого, синовиальный рак, рак поджелудочной железы, меланому, множественную миелому, нейробластому и рабдомиосаркому. Примеры Теперь данное изобретение описывается со ссылкой на следующие примеры, которые являются только иллюстративными и не предназначены для ограничения данного изобретения. Пример 1. Мышиное антитело ЕМ164. В этом первом примере описана полная первичная аминокислотная структура и кДНК-последовательность мышиного антитела данного изобретения, вместе с его связывающими свойствами и способом его экспрессии в рекомбинантной форме. Таким образом, обеспечено полное и законченное описание антитела данного изобретения и его получение, так чтобы средний специалист в области иммунологии был способен получить указанное антитело без чрезмерного экспериментирования. - 11 - 009807 А. Генерирование гибридомы моноклонального антитела против IGF-I-рецептора. Клеточную линию, экспрессирующую IGF-I-рецептор человека с мутацией Y1251F, использовали для иммунизации, так как она экспрессировала высокое количество IGF-I-рецепторов (~107 на клетку). Мутация Y1251F в цитоплазматическом домене IGF-I-рецептора приводила к потере трансформации (превращения клеток в опухолевые клетки) и антиапоптотической передачи сигнала, но не влияла на связывание IGF-I и IGF-I-стимулируемую митогенную передачу сигнала (O'Connor, R. et al., 1997, Mol. Cell. Biol., 17, 427-435; Miura, M. et al, 1995, J. Biol. Chem., 270, 22639-22644). Эта мутация не влияла иным образом на генерирование антител, так как антитело этого примера связывалось с внеклеточным доменом IGF-I-рецептора, который был идентичным как для мутанта Y125IF, так и для рецептора дикого типа. Клеточную линию, экспрессирующую IGF-I-рецептор с мутацией Y1251F, генерировали из 3T3-подобных клеток дефицитной по IGF-I-рецептору мыши трансфекцией геном IGF-I-рецептора человека вместе с геном устойчивости к пуромицину и отбирали с использованием пуромицина (2,5 мкг/мл) и FACS-сортингом на высокую экспрессию IGF-I-рецептора (Miura, M. et al., 1995, J. Biol. Chem., 270, 22639-22644). Клеточную линию, имеющую высокий уровень экспрессии IGF-I-рецептора, дополнительно отбирали с использованием высокой концентрации пуромицина, например, 25 мкг/мл, которая была токсичной для большинства этих клеток. Выжившие колонии отбирали, и колонии, обнаруживающие высокий уровень экспрессии IGF-I-рецептор, отбирали. Самок мышей CAF1/J, в возрасте 6 месяцев, иммунизировали внутрибрюшинно в день 0 клетками, сверхэкспрессирующими Y1251F-мутантный IGF-I-рецептор человека (5х106 клеток, суспендированных в 0,2 мл ЗФР). Животным повторно вводили 0,2 мл клеточной суспензии следующим образом: день 2, 5,2х106 клеток; день 5, 2х106 клеток; дни 7, 9, 12 и 23, 1х107 клеток. В день 26 мышь умерщвляли и извлекали ее селезенку. Селезенку измельчали между двумя матированными стеклянными предметными стеклами с получением суспензии отдельных клеток, которую промывали бессывороточной средой RPMI, содержащей пенициллин и стрептомицин (SFM). Осадок клеток селезенки ресуспендировали в 10 мл 0,83% (масса/объем) раствора хлорида аммония в воде в течение 10 мин на льду для лизиса эритроцитов и затем промывали бессывороточной средой (SFM). Клетки селезенки (1,2х108) объединяли с миеломными клетками (4х107) из несекретирующей линии клеток мышиной миеломы P3X63Ag8.653 (АТСС, Rockville, MD; Cat. # CRL1580) в пробирке и промывали бессывороточной средой RPMI 1640 (SFM). Супернатант удаляли и осадок клеток ресуспендировали в остальной среде. Пробирку помещали в химический стакан с водой при 37°С и медленно добавляли при скорости капель 0,5 мл/мин 1,5 мл раствора полиэтиленгли-коля (50% ПЭГ (мас./об.), средняя молекулярная масса 1500 в 75 мМ HEPES, рН 8) при осторожном встряхивании пробирки. После выдерживания в течение одной минуты добавляли 10 мл SFM следующим образом: 1 мл на протяжении первой минуты, 2 мл на протяжении второй минуты и 7 мл на протяжении третьей минуты. Затем другие 10 мл добавляли медленно на протяжении одной минуты. Клетки осаждали центрифугированием, промывали в SFM и ресуспендировали в среде для выращивания RPMI-1640, дополненной 5% фетальной телячьей сывороткой (ФТС), гипоксантином/аминоптери-ном/тимидином (HAT), пенициллином, стрептомицином и 10% клонирующей гибридому добавкой (HCS). Клетки высевали в 96-луночные плоскодонные планшеты для культуры ткани при 2х105 клеток селезенки в 200 мкл на лунку. Спустя 5-7 дней 100 мкл на лунку удаляли и заменяли средой для выращивания, дополненной гипоксантином/тимидином (НТ) и 5% ФТС. Общие условия, используемые для иммунизации и получения гибридомы, были такими как описанные J. Langone and H. Vunakis (Eds., Methods in Enzymology, Vol. 121, "Immunological Techniques, Part I", 1986; Academic Press, Florida) и E. Harlow and D. Lane ("Antibodies: A Laboratory Manual", 1988; Cold Spring Harbor Laboratory Press, New York). Другие способы иммунизации и получения гибридом могут быть также использованы, как это хорошо известно специалистам с квалификацией в данной области. Культуральные супернатанты из гибридомных клонов подвергали скринингу на связывание с очищенным IGF-I-рецептором человека и на отсутствие специфического связывания с клетками, сверхэкспрессирующими рецептор инсулина человека, при помощи ELISA и FACS-скрининга, как описано ниже. Клоны, проявляющие более высокую аффинность связывания с клетками, сверхэкспрессирующими IGF-I-рецептор человека, чем с клетками, сверхэкспрессирующими рецептор инсулина человека, размножали и субклонировали. Культуральные супернатанты этих субклонов дополнительно подвергали скринингу посредством вышеуказанных анализов связывания. При помощи этой процедуры, был отобран субклон 3F1-C8-D7 (ЕМ164), и гены тяжелой и легкой цепей клонировали и секвенировали, как описано ниже. IGF-I-рецептор человека выделяли для использования в скрининге супернатантов из гибридомных клонов на их связывание с IGF-I-рецептором при помощи описанного ниже способа. Биотинилирован-ный IGF-I получали модификацией рекомбинантного IGF-I с использованием реагентов биотинилирова-ния, таких как сульфо-NHS-LC-биотин, сульфо-NHS-SS-биотин или NHS-PEOzj-биотин. Биотинилиро-ванный IGF-I абсорбировали на стрептавидин-агарозных гранулах и инкубировали с лизатом из клеток, которые сверхэкспрессировали IGFR дикого типа человека или Y1251 Р-мутантный IGFR. Эти гранулы - 12 - 009807 промывали и элюировали буфером, содержащим 2-4 М мочевину и детергент, такой как Тритон Х-100 или октил-р-глюкозид. Элюированный IGF-I-рецептор диализовали против ЗФР и анализировали на чистоту электрофорезом в ДСН-ПААГ при восстанавливающих условиях, который показал полосы альфа- и бета-цепей IGF-I-рецептора с молекулярными массами приблизительно 135 кДа и 95 кДа, соответственно. Для проверки на связывание супернатантов гибридом с очищенным IGF-I-рецептором планшет Immulon-4HB ELISA покрывали пробой очищенного IGF-I-рецептора человека (полученного диализом из элюции смесью мочевина/октил-р-глюкозид аффино очищенной пробы), разбавленного в 50 мМ CHES-буфере при рН 9,5 (100 мкл; 4°С, в течение ночи). Лунки блокировали 200 мкл блокирующего буфера (10 мг/мл БСА в TBS-T-буфере, содержащем 50 мМ Трис, 150 мМ NaCl, рН 7,5, и 0,1% Твин-20) и инкубировали с супернатантами из гибридомных клонов (100 мкл; разведенные в блокирующем буфере) в течение приблизительно 1-12 ч, промывали TBS-T-буфером и инкубировали с конъюгатом козьи антитела против мышиного IgG-Fc -пероксидаза хрена (HRP) (100 мкл; 0,8 мкг/мл в блокирующем буфере; Jackson ImmunoResearch Laboratories), с последующими промывками и детектированием с использованием субстрата ABTS/H2O2 при 405 нм (0,5 мг/мл ABTS, 0,03% Н2О2 в 0,1 М цитратном буфере, рН 4,2). Обычно, супернатант из гибридомного субклона 3F1 давал сигнал приблизительно 1,2 единицы оптической плотности в пределах 3 мин проявления, в противоположность величинам 0,0, полученным для су-пернатантов из некоторых других гибридомных клонов. Общие условия для этого ELISA были сходными со стандартными условиями ELISA для связывания антител и детектирования, описанными Е. Harlow and D. Lane ("Antibodies: A Laboratory Manual", 1988; Cold Spring Harbor Laboratory Press, New York), которые также могут быть использованы. Скрининг гибридомных супернатантов на специфическое связывание с IGF-I-рецептором человека, но не с рецептором инсулина человека выполняли с использованием ELISA на клеточных линиях, которые сверхэкспрессировали Y1251F-IGF-I-рецептор человека, и на клеточных линиях, которые сверхэкс-прессировали рецептор инсулина человека. Обе клеточные линии получали из 3T3-подобных клеток IGF-I-рецептор-недостаточньгх мышей. Сверхэкспрессирующие IGF-I-рецептор клетки и сверхэкспресси-рующие рецептор инсулина клетки отдельно собирали из колб для культуры ткани быстрой обработкой трипсином/ЭДТА, суспендировали в среде для выращивания, содержащей 10% ФТС, осаждали центрифугированием и промывали ЗФР. Промытые клетки (100 мкл приблизительно 1-3 х 106 клеток/мл) добавляли в лунки Immulon-2НВ-планшета, покрытого фитогемагглютинином (100 мкл 20 мкг/мл РНА), центрифугировали и давали прикрепляться к РНА-покрытым лункам в течение 10 мин. Планшет с клетками переворачивали с постукиванием для удаления ЗФР и затем сушили в течение ночи при 37°С. Лунки блокировали 5 мг/мл-раствором БМА в ЗФР в течение 1 ч при 37°С и затем промывали осторожно ЗФР. Затем аликвоты супернатантов из гибридомных клонов (100 мкл; разбавленные в блокирующем буфере) добавляли в лунки, содержащие сверхэкспрессирующие IGF-I-рецептор клетки, и в лунки, содержащие сверхэкспрессирующие рецептор инсулина клетки, и инкубировали при температуре окружающей среды в течение 1 часа. Лунки промывали ЗФР, инкубировали с конъюгатом козьи антитела против мышиного IgG-Fc - пероксидаза хрена (100 мкл; 0,8 мкг/мл в блокирующем буфере) в течение 1 ч с последующими промывками и затем связывание детектировали с использованием субстрата ABTS/H2O2. Типичный супериатант из гибридомного субклона 3F1 после инкубирования с клетками, сверхэкспрессирующими IGF-I-рецептор, давал сигнал 0,88 единиц оптической плотности в пределах 12 минут проявления, в противоположность величине 0,22 единицы оптической плотности, полученной с клетками, сверхэкспрессирующими рецептор инсулина человека. Эту гибридому выращивали в колбах Integra CL 350 (Integra Biosciences, Maryland) в соответствии с описаниями изготовителя, для получения очищенного антитела ЕМ164. Выход приблизительно 0,5-1 мг/мл антитела получали в собранных супернатантах из колб Integra, на основе количественного определения ELISA и на основе электрофореза в ДСН-ПААГ/окрашивания Кумасси синим с использованием стандартов антител. Это антитело очищали аффинной хроматографией на колонке с гранулами Белок А-агарозы при стандартных условиях очистки в отношении нанесения и промывания в 100 мМ Трис-буфере, рН 8,9, содержащем 3 М NaCl, с последующей элюцией в 100 мМ растворе уксусной кислоты, содержащем 150 мм NaCl. Элюированные фракции, содержащие антитело, нейтрализовали холодным 2 М раствором K2HPO4 и диализовали в ЗФР при 4°С. Концентрацию антитела определяли измерением оптической плотности при 280 нм (коэффициент экстинкции =1,4 мг-1мл см-1). Пробу очищенного антитела анализировали электрофорезом в ДСН-ПААГ при восстанавливающих условиях и окрашивании Кумасси синим, которое показало только полосы тяжелой и легкой цепей антитела при приблизительно 55 кДа и 25 кДа, соответственно. Изотип этого очищенного антитела был IgG1 с легкой цепью каппа. В. Характеристика связывания антитела БМ164. Специфическое связывание очищенного антитела ЕМ164 демонстрировали при помощи клеточного сортинга с возбуждением флуоресценции (FACS) с использованием клеток, сверхэкспрессирующих IGF-I-рецептор человека, и с использованием клеток, сверхэкспрессирующих рецептор инсулина человека (фиг. 1). Инкубирование антитела ЕМ164 (50-100 нМ) в 100 мкл холодного FACS-буфера (1 мг/мл БСА в - 13 - 009807 среде MEM в модификации Дульбекко) выполняли с использованием клеток, сверхэкспрессирующих IGF-I-рецептор человека, и с использованием клеток, сверхэкспрессирующих рецептор инсулина человека (2 х 105 клеток/мл) в круглодонном 96-луночном планшете в течение 1 ч. Клетки осаждали центрифугированием и промывали холодным FACS-буфером осторожным стряхиванием с последующим инкубированием с конъюгатом козьи антитела против мышиного IgG-Fc - пероксидаза хрена (100 мкл; 10 мкг/мл в FACS-буфере) на льду в течение 1 ч. Клетки осаждали, промывали и ресуспендировали в 120 мкл 1% раствора формальдегида в ЗФР. Планшет анализировали с использованием FACSCalibur-ридера (BD Biosciences). Сильное смещение флуоресценции получали после инкубирования сверхэкспрессирующих IGF-I-рецептор клеток с антителом ЕМ164, в противоположность незначительному смещению после инкубирования сверхэкспрессирующих рецептор инсулина клеток с антителом ЕМ164 (фиг. 1), что показало, что антитело ЕМ 164 было селективным в его связывании с IGF-I-рецептором и не связывалось с рецептором инсулина. Контрольные антитела, антитело против IGF-I-рецептора 1Н7 (Santa Cruz Biotechnology) и анти-рецептор инсулина-антитело (BD Pharminogen Laboratories), давали смещения флуоресценции после инкубации с клетками, сверхэкспрессирующими IGF-I-рецептор и рецептор инсулина, соответственно (фиг. 1). Сильное смещение флуоресценции наблюдали также при помощи FACS-анализа с использованием антитела ЕМ164 и клеток MCF-7 рака молочной железы человека, которые экспрессирова-ли IGF-I-рецептор (Dufourny, В. et al., 1997, J. Biol. Chem., 272, 31171), которое показало, что антитело ЕМ164 связывалось с IGF-I-рецептором человека на поверхности опухолевых клеток человека. Константу диссоциации (KD) для связывания антитела ЕМ164 с IGF-I-рецептором человека определяли при помощи титрования ELISA связывания антитела при нескольких концентрациях либо с непосредственно сенсибилизацией IGF-I-рецептором (аффинно очищенным с использованием биотинилиро-ванного IGF-I, как описано выше), либо с опосредованно захваченным биотинилированным IGF-I-рецептором. Биотинилированный IGF-I-рецептор получали биотинилированием солюбилизированного детергентом лизата из сверхэкспрессирующих IGF-I-рецептор клеток с использованием реагента РЕО-малеимид-биотин (Pierce, Molecular Biosxiences), который аффинно очищали с использованием антитела против бета-цепи IGF-I-рецептора, иммобилизованного на NHS-агарозных гранулах, и элюировали 2-4 М мочевиной в буфере, содержащем детергент NP-40, и диализовали в ЗФР. Определение KD для связывания антитела ЕМ164 с биотинилированным IGF-I-рецептором проводили сенсибилизацией Immulon-2НВ-планшета 100 мкл 1 мкг/мл стрептавидином в карбонатном буфере (150 мМ карбонат натрия, 350 мМ бикарбонат натрия) при 4°С в течение ночи. Сенсибилизированные стрептавидином лунки блокировали 200 мкл блокирующего буфера (10 мг/мл БСА в TBS-T-буфере), промывали TBS-T-буфером и инкубировали с биотинилированным IGF-I-рецептором (10-100 нг) в течение 4 часов при температуре окружающей среды. Затем лунки, содержащие захваченный биотинилиро-ванный IGF-I-рецептор, промывали и инкубировали с антителом ЕМ164 в блокирующем буфере при нескольких концентрациях (5,1 х 10-13 М - 200 нМ) в течение 2 ч при температуре окружающей среды и затем инкубировали в течение ночи при 4°С. Затем лунки промывали TBS-T-буфером и инкубировали с конъюгатом козьи антитела против мышиного IgGH+L - пероксидаза хрена (100 мкл; 0,5 мкг/мл в блокирующем буфере) с последующими промывками и детектированием с использованием субстрата ABTS/H2O2 при 405 нм. Величину KD определяли нелинейной регрессией для односайтного связывания. Подобное титрование связывания проводили с использованием Fab-фрагмента антитела ЕМ164, полученного расщеплением папаином этого антитела, как описано Е. Harlow and D. Lane ("Using Antibodies: A Laboratory Manual", 1988; Cold Spring Harbor Laboratory Press, New York). Кривая титрования связывания для связывания антитела ЕМ164 с биотинилированным IGF-I-рецептором человека давала величину KD 0,1 нМ (фиг. 2). Fab-фрагмент антитела ЕМ164 также связывал IGF-I-рецептор человека очень прочно с величиной KD 0,3 нМ, что указывало на то, что мономерное связывание антитела Е164 с IGF-I-рецептором было также очень сильным. Эта крайне низкая величина константы диссоциации для связывания IGF-I-рецептора антителом ЕМ164 была отчасти обусловлена очень низкой скоростью koff, подтверждаемой сильными сигналами связывания, наблюдаемыми после пролонгированных промывок в течение 1-2 дней антитела, связанного с иммобилизованным IGF-I-рецептором. Антитело ЕМ 164 может быть использовано для иммунопреципитации IGF-I-рецептора, как показано инкубированием солюбилизированного детергентом лизата клеток MCF-7 молочной железы человека с антителом ЕМ164, иммобилизованным на белок G-агарозных гранулах (Pierce Chemical Company). Вес-терн-блот иммунопреципитата антитела ЕМ164 детектировали с использованием кроличьего поликло-нального антитела против бета-цепи (С-конца) IGF-I-рецептора (Santa Cruz Biotechnology) и конъюгата козьи антитела против мышиного IgG - пероксидаза хрена с последующими промывками и детектированием усиленной хемилюминесценции (ECL). Вестерн-блот иммунопреципитата ЕМ164 из клеток MCF-7 обнаруживал полосы, соответствующие бета-цепи IGF-I-рецептора при приблизительно 95 кДа и про-IGF-I-рецептор при приблизительно 220 кДа. Сходные иммунопреципитации проводили для других типов клеток для проверки видовой специфичности связывания антитела ЕМ164, которое также связыва - 14 - 009807 лось с IGF-I-рецептором из клеток COS-7 (африканская зеленая мартышка), но не связывалось с IGF-I-рецептором клеток 3T3 (мышь), клеток СНО (китайский хомячок) или фибробластных клеток козы (коза). Антитело EM164 не детектировало денатурированный ДСН IGF-I-рецептор человека в Вестерн-блоте лизатов из клеток MCF-7, что указывало на то, что оно связывалось с конформационным эпитопом на-тивного, неденатурированного IGF-I-рецептора человека. Связывающий домен антитела ЕМ163 дополнительно характеризовали с использованием укороченной конструкции альфа-цепи, которая содержала богатый цистеинами домен, фланкированный доменами L1 и L2 (остатки 1-468), слитый с 16-мерным участком С-конца (остатки 704-719) и которая заканчивалась С-концевой эпитопной меткой. Сообщалось, что этот IGF-I-рецептор меньшего размера, который был лишен остатков 469-703, связывается с IGF-I, хотя и менее прочно в сравнении с нативным полноразмерным IGF-I-рецептором (Molina, L. et al., 2000, FEBS Letters, 467, 226-230; Kristensen, С. et al., 1999, J. Biol. Chem., 274, 37251-37356). Таким образом, получали укороченную конструкцию альфа-цепи IGF-I-рецептора, содержащую остатки 1-468, слитые с С-концевым участком, который содержит остатки 704719 и фланкирован С-концевой эпитопной меткой myc. Конструировали стабильную клеточную линию, которая экспрессировала эту конструкцию и которая также экспрессирует эту конструкцию транзиторно в клетках 3T3 эмбриональной почки человека. Наблюдали сильное связывание антитела ЕМ164 с этой конструкцией укороченной альфа-цепи IGF-I-рецептора. Из двух испытанных антител, IR3 (Calbiochem) также связывалось с этой укороченной альфа-цепью, но антитело 1 Н7 (Santa Cruz Biotechnology) не связывалось, что указывало на то, что эпитоп антитела ЕМ164 был явно отличающимся от эпитопа антитела 1 Н7. С. Ингибирование связывания IGF-I с клетками MCF-7 антителом ЕМ164. Связывание IGF-I с клетками рака молочной железы человека ингибировали антителом ЕМ164 (фиг. 3). Клетки MCF-7 инкубировали в присутствии или в отсутствие 5 мкг/мл антитела ЕМ 164 в течение 2 ч в бессывороточной среде с последующим инкубированием с 50 нг/мл биотинилированного IGF-I в течение 20 мин при 37°С. Затем эти клетки промывали дважды бессывороточной средой для удаления несвязанного биотин-IGF-I и затем лизировали в 50 мМ HEPES, рН 7,4, содержащем 1% NP-40 и ингибиторы протеаз. ^ПШЮП^НВ -планшет сенсибилизировали мышиным моноклональным антителом против бета-цепи IGF-I-рецептора и использовали для захвата IGF-I-рецептора и связанного биотин-IGF-I из лизата. Связывание сенсибилизирующего антитела с цитоплазматическим С-концевым доменом бета-цепи IGF-I-рецептора не мешало связыванию биотин-IGF-I с внеклеточным доменом IGF-I-рецептора. Эти лунки промывали, инкубировали с конъюгатом стрептавидин-пероксидаза хрена, опять промывали и затем детектировали с использованием субстрата ABTS/H2O2. Ингибирование IGF-I-связывания с клетками MCF-7 антителом ЕМ164 (5 мкг/мл) было по существу количественным и было почти эквивалентным связыванию фона ELISA, полученного с использованием контроля, не содержащего биотин-IGF-I. Кроме описанного выше анализа на ингибирование связывания IGF-I с клетками MCF-7 антителом ЕМ164, следующий анализ показал, что антитело ЕМ164 было высокоэффективным в вытеснении связанного IGF-I из клеток MCF-7, как это и желательно при физиологических условиях, чтобы антагонистическое антитело против IGF-I-рецептора вытесняло связанный эндогенный физиологический лиганд (такой как IGF-I или IGF-II). В этом анализе вытеснения IGF-I клетки MCF-7, выращенные в 12-луночном планшете, подвергали голоданию в бессывороточной среде и затем инкубировали с биотини-лированным IGF-I (20-50 нг/мл) в бессывороточной среде при 37°С (или при 4°С) в течение 1-2 ч. Затем эти клетки со связанным биотинилированным IGF-I обрабатывали антителом ЕМ164 или контрольным антителом (10/100 мкг/мл) при 37°С (или при 0°С) в течение 30 мин - 1 ч. Затем клетки промывали ЗФР и лизировали в лизисном буфере, содержащем 1% NP-40, при 4°С. ELISA проводили, как описано выше, для захвата IGF-I-рецептора из лизата и затем детектирования биотинилированного IGF-I, связанного с рецептором, с использованием конъюгата стрептавидин - пероксидаза хрена. Этот ELISA показал, что антитело ЕМ164 было способно вытеснять предварительно связанный биотинилированный IGF-I из клеток почти полностью (90% в пределах 30 мин и ~ 100% в пределах 4 ч) при 37°С и приблизительно на 50% за 2 ч при 4°С. В другом эксперименте, клетки рака легкого NCI-H838 инкубировали с биотин-IGF-I, затем промывали и инкубировали с антителом ЕМ164 при 4°С в течение 2 ч, что приводило к 80% уменьшению предварительно связанного биотин-IGF-I. Таким образом, антитело ЕМ164 было высокоэффективным в вытеснении предварительно связанного биотин-IGF-I из раковых клеток, что было бы важным терапевтически для антагонизма в отношении IGF-I-рецептора вытеснением связанного эндогенного физиологического лиганда. Инкубирование клеток MCF-7 с антителом ЕМ164 при 4°С в течение 2 ч (или при 37°С в течение 30 мин) не приводило к значимой понижающей регуляции IGF-I-рецептора на основе Вестерн-блот-анализа с использованием антитела против бета-цепи IGF-I-рецептора (Santa Cruz Biotechnology; sc-713), хотя более продолжительное инкубирование с антителом ЕМ164 при 37°С в течение 2 ч приводило к 25% понижающей регуляции IGF-I-рецептора. Таким образом, ингибирование связывания IGF-I и вытеснение связанного IGF-I антителом ЕМ164 как при 4°С, так и при 37°С в этих экспериментах с коротким периодом времени не могут быть объяснены понижающей регуляцией рецептора вследствие связывания антитела ЕМ164. Механизм сильного ингибирования связывания IGF-I с IGF-I-рецептором и вытеснения - 15 - 009807 предварительно связанного IGF-I антителом ЕМ164 является, по-видимому, конкуренцией за связывание, либо через наличие общего сайта связывания, либо через стерическую окклюзию, либо через алло-стерические эффекты. D. Ингибирование опосредованной IGF-I-рецептором клеточной передачи сигнала антителом ЕМ164. Обработка клеток рака MCF-7 и клеток остеосаркомы SaOS-2 антителом ЕМ164 почти полностью ингибировала внутриклеточную передачу сигнала IGF-I-рецептора, как показано ингибированием авто-фосфорилирования IGF-I-рецептора и ингибированием фосфорилирования его расположенных далее по ходу трансдукции сигнала эффекторов, таких как субстрат-1 рецептора инсулина (IRS-1), Akt и Erk1/2 (фиг. 4-6). На фиг. 4, клетки MCF-7 выращивали в 12-луночном планшете в нормальной среде в течение 3 дней и затем их обрабатывали 20 мкг/мл антитела ЕМ164 (или контрольного анти-В4-антитела) в бессывороточной среде в течение 3 ч с последующей стимуляцией 50 нг/мл IGF-I в течение 20 мин при 37°С. Затем эти клетки лизировали в охлажденном на льду лизисном буфере, содержащем ингибиторы протеаз и фосфатаз (50 мМ HEPES-буфер, рН 7,4, 1% NP-40, 1 мМ ортованадат натрия, 100 мМ фторид натрия, 10 мМ пирофосфат натрия, 2,5 мМ ЭДТА, 10 мкМ лейпептин, 5 мкМ пепстатин, 1 мМ PMCF, 5 мМ бен-замид и 5 мкг/мл апротинин). Планшет ELISA предварительно сенсибилизировали моноклональным антителом ТС123 против С-конца бета-цепи ингредиента и инкубировали с пробами лизата в течение 5 ч при температуре окружающей среды для захвата IGF-I-рецептора. Затем лунки, содержащие захваченный IGF-I-рецептор, промывали и инкубировали с биотинилированным антифосфотирозин-антителом (PY20; 0,25 мкг/мл; BD Transduction Laboratories) в течение 30 мин с последующими промывками и инкубированием с конъюгатом стрептавидин-пероксидаза хрена (0,8 мкг/мл) в течение 30 мин. Лунки промывали и детектировали с использованием субстрата ABTS/H2O2. Использование контрольного анти-В4-антитела не обнаруживало ингибирования IGF-I-стимулируемого автофосфорилирования IGF-I-рецептора. В противоположность этому, полное ингибирование IGF-I-стимулируемого автофосфорилирования IGF-I-рецептора получали после обработки антителом ЕМ 164 (фиг. 4). Для демонстрации ингибирования фосфорилирования субстрата-1 рецептора инсулина (IRS-1) использовали ELISA с применением иммобилизованного антитела против IRS-1 для захвата IRS-1 из лиза-тов с последующим измерением ассоциированной субъединицы р85 фосфатидилинозитол-3-киназы (PI-3-киназы), которая связывается с фосфорилированным IRS-1 (Jackson, J.G. et al., 1998, J. Biol. Chem., 273, 9994-10003). Ha фиг. 5 клетки MCF-7 обрабатывали 5 мкг/мл антитела (ЕМ164 или IR3) в бессывороточной среде в течение 2 ч с последующей стимуляцией 50 нг/мл IGF-I в течение 10 мин при 37°С. Анти-IRS-1-антитело (кроличье поликлональное; Upstate Biotechnology) непосредственно захватывалось посредством инкубирования нанесенным в виде покрытия козьим антителом против кроличьего IgG на ELISA-планшете, который затем использовали для захвата IRS-1 из проб клеточных лизатов инкубированием в течение ночи при 4°С. Затем эти лунки инкубировали с мышиным моноклональным антителом против р85-PI-3-киназы (Upstate Biotechnology) в течение 4 ч с последующей обработкой конъюгатом козьи антитела против мышиного IgG - HRP в течение 30 мин. Затем лунки промывали и детектировали с использованием субстрата ABTS/H2O2 (фиг. 5). Как показано на фиг. 5, антитело ЕМ164 было более эффективным в ингибировании IGF-I-стимулируемого фосфорилирования IRS-1 при инкубировании с клетками в отсутствие IGF-I. Активация других эффекторов, находящихся ниже по ходу передачи сигнала, таких как Akt и Erk1/2, также ингибировалась зависимым от дозы образом антителом ЕМ164 в клетках SaOS-2 (фиг. 6) и в клетках MCF-7, как было показано Вестерн-блотами лизатов и специфическими в отношении фосфо-рилирования антителами (кроличьими поликлональными антителами против фосфо-Ser473 Akt и поли-клональными антителами против фосфо-ERK1/2; Cell Signaling Technology). Антитело против тотального ERK показало одинаковые нагрузки белка во всех дорожках (фиг. 6). Обработка клеток SaOS-2 антителом ЕМ164 не ингибировала EGF-стимулируемое фосфорилирование Erk1/2 демонстрируя специфичность ингибирования пути передачи сигнала IGF-I-рецептора антителом ЕМ 164. E. Ингибирование стимулируемого IGF-I, IGF-II и сывороткой роста и выживания опухолевых клеток человека антителом ЕМ164. Несколько линий опухолевых клеток человека испытывали в бессывороточных условиях на их реакцию роста и выживания на IGF-I. Эти клеточные линии обрабатывали антителом ЕМ164 в присутствии IGF-I, IGF-II или сыворотки и их реакции роста и выживания измеряли с использованием МТТ-анализа спустя 2-4 дня. Приблизительно 1500 клеток высевали в 96-луночном планшете в нормальной среде с сывороткой, которую заменяли бессывороточной средой на следующий день (либо бессывороточной средой RPMI, дополненной трансферрином и БСА, либо не содержащей фенолового красного средой, описанной Dufourny, В. et al., 1997, J. Biol. Chem., 272, 31163-31171). После одного для роста в бессывороточной среде эти клетки инкубировали с приблизительно 75 мкл 10 мкг/мл антитела в течение 30 мин -3 ч с последующим добавлением 25 мкл раствора IGF-I (или IGF-II или сыворотки) с получением конечной концентрации 10 нг/мл IGF-I или 20 нг/мл IGF-II или 0,04-10% сыворотки. В некоторых экспериментах эти клетки стимулировали сначала IGF-I в течение 15 мин перед добавлением антитела ЕМ164, или - 16 - 009807 добавляли вместе как IGF-I, так и антитело ЕМ164. Затем клеткам давали расти в течение следующих 2-3 дней. Затем добавляли раствор МТТ (бромида 3-(4,5)-диметилтиазол-2-ил)-2,5-дифенилтетразолия; 25 мкл 5 мг/мл раствора в ЗФР) и эти клетки возвращали в термостат на 2-3 ч. Затем среду удаляли и заменяли 100 мкл ДМСО, смешивали и измеряли оптическую плотность этого планшета при 545 нм. Несколько линий опухолевых клеток человека обнаружили реакцию роста и выживания после добавления IGF-I или IGF-II или сыворотки, которая значимо ингибировалась антителом ЕМ164, независимо от того, добавляли антитело до IGF-I или IGF-I добавляли до антитела или как IGF-I, так и антитело добавляли вместе (табл. 1). Таблица 1. Ингибирование IGF-I-стимулируемого роста и выживания опухолевых клеток антителом ЕМ 164 Тип опухолевых клеток Реакция роста на IGF-I (МТТ-анализ: отношение IGF-I-обработанных клеток к необработанным клеткам в бессывороточной среде (в указанное число раз)а % ингибирования антителом ЕМ 164 1GF-I-сти мул ируемого роста в бессывороточной среде Ингибирование антителом ЕМ 164 роста/выживания (слеток в 1,25-10% сыворотке1* MCF-7 (молочная железа) 1.7-2.8 100% 85% НТ-3 (шейка матки 70-90% Coto 205 (ободочная кишка)) 2.3 50% НТ-29 1.5 60% NCI-H838 (легкое) 100% 85-90 % Calu-6 1.6-1.8 85% SK-LU-1 1.4 100% нет NCI-H596 1.4 100% Weakly А 549 1.2 80% 1MD А 375 (меланома) 1.6 90% нет SK-Mel-37 1.4 85% RD (рабдомиосаркома) 1.7 85-100 % SaOS-2 (остеосаркома) 2.5 100% А 431 (эпидермоид) 2.2 85% SK-N-SH (нейробластома) 85% 30-50 % а МТТ-анализ 3-4-дневного роста/выживания клеток в реакции на 10 нг/мл IGF-I в бессывороточной среде, содержащей 5-10 мкг/мл антитела ЕМ 164 b Ингибирование роста клеток в 1,25-10% сыворотке в присутствии 5-10 мкг/мл антитела ЕМ164 в соответствии с МТТ-анализом или анализом образования колоний на основании сравнения с контролем (с сывороткой, но без антитела); степень ингибирования количественно измеряли для клеток MCF-7, NCI-H838 и SK-N-SH в расчете на контроли (без сыворотки, но с антителом и с сывороткой, но без антитела) для объяснения аутокрин-ной/паракринной IGF-стимуляции клетками. ND указывает отсутствие данных или плохие данные вследствие трудностей окрашивания. Антитело ЕМ164 сильно ингибировало стимулируемые IGF-I или сывороткой рост и выживание клеток MCF-7 рака молочной железы (фиг. 7 и 8). В отдельном эксперименте антитело ЕМ164 сильно ингибировало стимулируемые IGF-II рост и выживание клеток MCF-7. Предыдущие сообщения с использованием коммерчески доступных антител, таких как антитело IR3, показали только слабое ингиби-рование стимулируемых сывороткой роста и выживания клеток MCF-7, как подтверждено на фиг. 7 для антител IR3 и 1Н7 (Cullen, K.J. et al., 1990, Cancer Res., 50, 48-53). В противоположность этому, антитело ЕМ164 было сильным ингибитором стимулируемого сывороткой и IGF-I роста клеток MCF-7. Как показано на фиг. 8, антитело ЕМ164 было одинаково эффективным в ингибировании роста и выживания клеток MCF-7 в широком диапазоне концентраций сыворотки (0,04-10% сыворотка). Ингибирование роста клеток MCF-7 антителом ЕМ164 измеряли подсчетом клеток. Так, в 12 - 17 - 009807 луночном планшете приблизительно 7500 клеток высевали в среде RPMI с 10% ФТС, в присутствии и в отсутствие 10 мкг/мл антитела ЕМ164. После 5 дней роста количество клеток для необработанной контрольной пробы было равно 20,5 х 104 клеток, в противоположность количеству клеток только 1,7 х 104 клеток для обработанной антителом ЕМ 164 пробы. Обработка антителом ЕМ 164 ингибировала рост клеток MCF-7 приблизительно в 12 раз за 5 дней. Это ингибирование антителом ЕМ164 было значимо большим, чем сообщенное ингибирование в 2,5 раза с использованием антитела IR3 в 6-дневном анализе для клеток MCF-7 (Rohlik, Q.T. et al., 1987, Biochem. Biophys. Res. Commun., 149, 276-281). Стимулируемые IGF-I и сывороткой рост и выживание линии клеток немелкоклеточного рака легкого NCI-H838 также сильно ингибировались антителом ЕМ 164 в сравнении с контрольным антителом В4 (фиг. 9). Обработка антителом ЕМ164 в бессывороточной среде давала меньший сигнал, чем необработанная проба как для клеток HCI-H838, так и для клеток MCF-7, предположительно вследствие того, что антитело ЕМ164 ингибировало также аутокринную и паракринную стимуляцию IGF-I и IGF-II этих клеток (фиг. 7 и 9). Размер колоний клеток рака ободочной кишки НТ29 был также значительно уменьшен после обработки антителом ЕМ164. Таким образом, в антитело ЕМ164 является уникальным среди всех известных антител против IGF-I-рецептора по его эффективности в ингибировании стимулируемого сывороткой роста опухолевых клеток, таких как клетки MCF-7 и NCI-H838, на более, чем 80%. Антитело ЕМ164 вызывало задержку роста в фазе G0/G1 клеточного цикла и устраняло митогенное действие IGF-I. Для анализа клеточного цикла клетки MCF-7 обрабатывали IGF-I (20 нг/мл) в присутствии или в отсутствие ЕМ164 (20 мкг/мл) в течение 1 дня и затем анализировали окрашивание иодидом пропидия и проточной цитометрией. Как показано на фиг. 25, прохождение клеточного цикла клетками в ответ на стимуляцию IGF-I в отсутствие ЕМ 164 (с 41% этих клеток в S-фазе и 77% этих клеток с G0/G1-фазе) подавлялось в ЕМ164-обработанных клетках (с только 9% в S-фазе и 77% этих клеток в G0/G1фазе). Кроме ингибирования пролиферации клеток обработка антителом ЕМ164 приводила к апоптозу клеток. Для измерения апоптоза расщепление белка цитокератина СК18 каспазой измеряли в клетках рака легкого NCI-H838, инкубированных с: IGF-I или сывороткой в присутствии или в отсутствие ЕМ164, для дня 1 (фиг. 26). В отсутствие ЕМ164 добавление IGF-I или сыворотки приводило к более низкому сигналу расщепленного каспазой СК18 в сравнении с контролем без IGF-I, что указывало на то, что IGF-I и сыворотка предотвращают активацию каспазы. Обработка антителом ЕМ164 подавляла анти-апоптотические действия IGF-I и сыворотки, как показано более высокими уровнями расщепленного СК18, полученными в присутствии ЕМ164, чем в отсутствие ЕМ164 (фиг. 26). F. Синергическое ингибирование антителом ЕМ164 роста и выживания опухолевых клеток человека в комбинациях с другими цитотоксическими и цитостатическими агентами. Комбинированное введение антитела ЕМ164 с таксолом было значимо более ингибиторным в отношении роста и выживания клеток немелкоклеточного рака легкого Calu6, чем один таксол. Подобным образом, комбинация антитела ЕМ164 с камптотецином была значительно более ингибирующей, чем один камптотекцин, в отношении роста и выживания клеток рака ободочной кишки НТ29. Поскольку не ожидалось, что одно антитело ЕМ164 является таким же токсичным в отношении клеток, как органические хемотоксичные лекарственные средства, синергизм между преимущественно цитостатическим действием антитела ЕМ164 и цитотоксическим действием хемотоксичного лекарственного средства мог быть высокоэффективным в комбинации раковых терапевтических веществ в клинических условиях. Комбинированное действие антитела ЕМ164 с антителом против IGF-I-рецептора (KS77) было значимо более ингибиторным, чем одного антитела ЕМ164 или одного антитела KS77, на рост и выживание нескольких линий опухолевых клеток, таких как клетки НТ-3, клетки RD, клетки MCF-7 и клетки А431. Таким образом, синергическое действие комбинирования нейтрализующих антител для двух рецепторов фактора роста, таких как IGF-I-рецептор и EGF-рецептор, может быть также применимым в клиническом лечении рака. На основе эффективности антитела ЕМ164 в виде единственного агента в ингибировании пролиферации и выживания различных линий раковых клеток человека, показанной в табл. 1, проводили дополнительные исследования эффективности с использованием комбинаций антитела ЕМ164 с другими противораковыми терапевтическими агентами. В этих исследованиях на различных линиях раковых клеток комбинированная обработка антителом ЕМ164 и другими противораковыми терапевтическими агентами приводила к даже более высокой противораковой эффективности, чем с использованием либо ЕМ164, либо другого терапевтического агента по отдельности. Таким образом, комбинации ЕМ164 с другими терапевтическими агентами являются высокоэффективными в ингибировании пролиферации и выживания раковых клеток. Терапевтическими агентами, которые могут комбинироваться с ЕМ164 для улучшенной противораковой эффективности, включают в себя разнообразные агенты, используемые в онкологической практике (Ссылка: Cancer, Principles & Practice of Oncology, DeVita, V.T., Helman, S., Rosenberg, S.A., 6th edition, Lippencott-Raven, Philadelphia, 2001), такие как доцетаксел, паклитаксел, док-сорубицин, эпирубицин, циклофосфамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, анастрозол, фулвестрант, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дек - 18 - 009807 саметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорелбин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идарубицин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибриту-момаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2^ мелфолам, бортезомиб (Velcade), алтрета-мин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), антитело против EGF-рецептора (Cetuxivmab, Abx-EGF), эпотилоны и конъюгаты цитотоксических лекарственных средств и антител против рецепторов клеточной поверхности. Для этих комбинированных способов терапии ЕМ164 комбинируют с одним или несколькими противораковыми агентами различных механизмов действия, такими как алкилирующие агенты, содержащие платину агенты, гормональные терапевтические агенты, антиметаболиты, ингибиторы топоизомера-зы, антимикробные агенты, агенты дифференцировки, антиангиогенные агенты или агенты, действующие против васкуляризации, лучевая терапия, агонисты и антагонисты рилизинг-фактора лютеинизи-рующего гормона (LHRH) или гонадотропин-рилизинг-гормона (GnRH) и другие химиотерапевтические агенты (Ссылка: Cancer, Principles & Practice of Oncology, DeVita, V.T., Helman, S., Rosenberg, S.A., 6th edition, Lippencott-Raven, Philadelphia, 2001). В одном варианте осуществления, комбинация антагониста LHRH антида (0,1-10 микромолярного) и антитела ЕМ164 (0,1-10 наномолярного) ингибировала пролиферацию раковых клеток молочной железы MCF-7 значительно в большей степени, чем в случае либо одного ЕМ164, либо одного антида. В примере комбинированной терапии с содержащим платину агентом, эта комбинированная обработка антителом ЕМ164 (10 мкг/мл) и цисплатином (0,1-60 мкг/мл) приводила к большему ингибированию пролиферации и выживания раковых клеток молочной железы MCF-7 в сравнении с ингибированием либо одним антителом ЕМ164, либо одним цисплатином. Эти комбинации антитела ЕМ164 с другими терапевтическими агентами являются эффективными против нескольких типов раков, в том числе рака молочной железы, легкого, ободочной кишки, поджелудочной железы, шейки матки, яичника, меланомы, множественной миеломы, нейробластомы, рабдо-миосаркомы и остеосаркомы. Антитело ЕМ164 и этот терапевтический агент могут вводиться для раковой терапии либо одновременно, либо последовательно. Конъюгаты антитела ЕМ164 с цитотоксическими лекарственными средствами также являются ценными в нацеленной доставке цитотоксических лекарственных средств к опухолям, сверхэкспрессирую-щим IGF-I-рецептор. Конъюгаты антитела ЕМ 164 с радиоактивными метками могут быть использованы в лечении и визиографии опухолей, которые сверхэкспрессируют IGF-I-рецептор. G. Действие обработки ЕМ164, в качестве единственного агента или в комбинации с противораковыми агентами, в раковых ксенотрансплантатах человека в иммунодефицитных мышах. Ксенотрансплантаты немелкоклеточного рака человека Calu-6 прививали в иммуно дефицитным мышам подкожными инъекциями 1 х 107 клеток Calu-6. Как показано на фигуре 10, мышей, содержащих привитые ксенотрансплантаты Calu-6 100 мм3, обрабатывали одним антителом ЕМ164 (6 инъекций 0,8 мг/мышь, i.v., два раза в неделю) или одним таксолом (пять инъекций таксола, i.p., каждые два дня; 15 мг/кг) или комбинацией обработок таксолом и антителом ЕМ164 или одним ЗФР (200 мкг/мышь, 6 инъекций, два раза в неделю, i.v.) с использованием пяти мышей на группу обработки. Рост опухолей значимо замедлялся обработкой антителом ЕМ 164 на основании измерений массы этих мышей. Хотя обработка одним таксолом была эффективной до дня 14, затем эта опухоль снова начинала расти. Однако рост опухоли значимо задерживался в группе, которую обрабатывали комбинацией таксола и антитела ЕМ164, в сравнении с группой, которую обрабатывали одним таксолом. Ксенотрансплантаты поджелудочной железы человека прививали 5-недельным самкам мышей SCID/ICR (Talconic) подкожными инъекциями 107 клеток ВхРС-3 в ЗФР (день 0). Затем мышей, несущих привитые опухоли 80 мм3, обрабатывали одним ЕМ164 (13 инъекций 0,8 мг/мышь, i.v., в латеральную хвостовую вену, в дни 12, 16, 19, 23, 26, 29, 36, 43, 50, 54, 58, 61 и 64), одним гемцитабином (две инъекции 150 мг/кг/мышь, i.p. в дни 12 и 19), комбинацией гемцитабина и ЕМ164 в соответствии с вышеуказанными схемами введения, одним ЗФР и одним контрольным антителом (в соответствии со схемами, приведенными для ЕМ164) с использованием пяти мышей в каждой из пяти групп обработки. Как показано на фиг. 27, обработка одним ЕМ164 или в комбинации с гемцитабином приводила к общему регрессу опухолевых ксенотрансплантатов в 4 из 5 животных в группе обработки ЕМ164 и во всех 5 животных в группе обработки комбинацией. Измеримое возобновление роста опухоли наблюдали только в более, чем одном животном, в день 43 в группе ЕМ164 и в день 68 в группе обработки комбинацией, приводящее к значимо меньшим средним объемам опухолей в день 74 в сравнении с контрольными обработками (Р = 0,029 и 0,002, соответственно; двухсторонний Т-критерий; фиг. 27). В другом исследовании, обработка антителом ЕМ164 (одним или в комбинации с антитело против IGF-I-рецепторам; внутрибрюшин-ные инъекции) ингибировала рост привитых ксенотрансплантатов ВхРС-3 в мышах. Мышиные ЕМ164 и гуманизированные антитела ЕМ164 показали эквивалентное ингибирование роста привитых ксенотрансплантатов ВхРС-3 в мышах, демонстрируя, следовательно, что эффективность гуманизированного ЕМ164 является эквивалентной эффективности мышиного EMI64 in vivo. В сравнении с различными способами введения антитела ЕМ164, как внутрибрюшинное, так и внутривенное введения антитела ЕМ164 показали эквивалентное ингибирование роста привитых ксенотрансплан - 19 - 009807 татов ВхРС-3 в мышах. В другом исследовании с ксенотрансплантатами обработка антителом ЕМ164 обнаруживала значимую задержку роста привитых ксенотрансплантатов рабдомиосаркомы человека А-673/саркомы Эвинга в мышах. Н. Клонирование и секвенирование легкой и тяжелой цепей антитела ЕМ164. Тотальную РНК очищали из гибридомных клеток ЕМ164. Реакции обратной транскриптазы выполняли с использованием 4-5 мкг тотальной РНК и либо олиго-dT, либо случайных гексамерных прайме-ров. 1ТЦР-реакции выполняли с использованием способа RACE (быстрой амплификации концов кДНК), описанного в Со et al. (J. Immunol., 148, 1149-1154 (1992)), и с использованием вырожденных праймеров, описанных в Wang et al. (J. Immunol. Methods, 233, 167-177 (2000). Способ ТЦР RACE требовал промежуточной стадии добавления поли G-хвоста на 3'-концах кДНК первых цепей. ОТ-реакционные смеси очищали с использованием колонок Qianeasy (Qiagen) и элюировали в 50 мкл 1XNEB-буфера 4. Реакция присоединения dG-хвоста выполняли на элюате с использованием 0,25 мМ CoCl2, 2 мМ dGTP и 5 единиц терминальной трансферазы (NEB) в 1 X NEB-буфере 4. Эту смесь инкубировали при 37°С в течение 30 мин и затем 1/5 этой реакционной смеси (10 мкл) добавляли непосредственно в ТЦР-реакцию, где она служила в качестве ДНК-матрицы. RACE- и вырожденные ТЦР-реакции были идентичными, за исключением различий в праймерах и матрицах. Реакцию терминальной трансферазы использовали непосредственно для матрицы RACE-PCR, тогда как реакционную смесь ОТ-реакции использовали непосредственно для вырожденных ТЦР-реакций. Как в RACE-, так и в вырожденных ТЦР-реакциях использовали один и тот же 3'-праймер легкой цепи: HindKL - tatagagctcaagcttggatggtgggaagatggatacagttggtgc (SEQ ID NO:14) и 3'-праймер тяжелой цепи: Bgl2IgG1 - ggaagatctatagacagatgggggtgtcgttttggc (SEQ ID NO:15). В RACE-ТЦР один поли С-5'-праймер использовали как для тяжелой, так и для легкой цепи: EcoPolyC - TATATCTAGAATTCCCCCCCCCCCCCCCCC (SEQ ID NO:16), тогда как вырожденными 5'-концевыми праймерами были: Sac1MK - GGGAGCTCGAYATTGTGMTSACMCARWCTMCA (SEQ ID NO:17) для легкой цепи и равная смесь из: EcoRIMH1 - CTTCCGGAATTCSARGTNMAGCTGSAGSAGTC (SEQ ID NO:18) и EcoRIMH2 - CTTCCGGAATTCSARGTNMAGCTGSAGSAGTCWGG (SEQ ID NO:19) для тяжелой цепи. В приведенных выше последовательностях праймеров смешанные основания определены следующим образом: Н=А+Т+С, S=G+C, Y=C+T, K=G+T, М=А+С, R=A+G, W=A+T, V=A+C+G. ТЦР-реакции вьшолняли с использованием следующей программы: 1) 94°С 3 мин, 2) 94°С 15 с, 3) 45°С 1 мин, 4) 72°С 2 мин, 5) цикл опять до стадии №2 29 раз, 6) окончание с конечной стадией удлинения при 72°С в течение 10 мин. ТЦР-продукты клонировали в pBlueScript II SK+ (Stratagene) с использованием рестрикционных ферментов (рестриктаз), созданных этими ТЦР-праймерами. Несколько отдельных клонов легкой и тяжелой цепи секвенировали общепринятыми способами для идентификации и с целью избежать возможных ошибок генерированной полимеразой последовательности (фиг. 12 и 13). С использованием определений канонической классификации Chothia были идентифицированы CDR трех легких цепей и тяжелых цепей (фиг. 12-14). Тоиск с использованием базы данных NCBI IgBlast показал, что вариабельная область легкой цепи антитела против IGF-I-рецептора, возможно, происходила из гена мышиной зародышевой линии IgVk Cr1, тогда как вариабельная область тяжелой цепи, возможно, происходила из гена зародышевой линии IgVh J558.c (фиг. 15). Секвенирование белка мышиного антитела ЕМ164 выполняли для подтверждения последовательностей, показанных на фиг. 12 и 13. Полосы белков тяжелой и легкой цепей очищенного антитела ЕМ164 переносили на мембрану PVDF из геля (ДСН-ТААГ, восстанавливающие условия), вырезали из мембраны PVDF и анализировали путем секвенирования белковой последовательности. Было определено секве-нированием по Эдману, что N-концевая последовательность легкой цепи представляет собой: DVLMTQTPLS (SEQ ID NO:20), которая соответствует N-концевой последовательности клонированного гена легкой цепи из гибридомы ЕМ164. Было обнаружено, что N-конец тяжелой цепи является блокированным для секвенирования белка по Эдману. Тептидный фрагмент расщепления трипсином тяжелой цепи с массой 1129,5 (М+Н+, моноизотопный) фрагментировали при помощи post-source decay (PSD), и было определено, что его последовательность представляет собой GRPDYYGSSK (SEQ ID NO:21). Другой пептидный фрагмент триптиче-ского расщепления тяжелой цепи с массой 2664,2 (М+Н+, моноизотопный) был также фрагментирован при помощи post-source decay (PSD), и было определено, что его последовательность представляет собой SSSTAYMQLSSLTSEDSAVYYFAR (SEQ ID NO:22. Обе эти последовательности точно соответствуют последовательностям CDR3 и каркаса 3 (FR3) клонированного гена тяжелой цепи, полученного из гиб - 20 - 009807 ридомы ЕМ 164. I. Рекомбинантная экспрессия антитела ЕМ164. Парные последовательности легкой и тяжелой цепей клонировали в единый экспрессионный вектор млекопитающего (фиг. 16). ПЦР-праймеры для вариабельных последовательностей человека создавали сайты рестрикции, которые позволяли присоединяться сигнальной последовательности человека в клонирующем векторе pBlueScriptll, и эти вариабельные последовательности клонировали в экспрессион-ную плазмиду млекопитающего с использованием сайтов EcoRI и BsiWI или HindIII и Apal для легкой цепи или тяжелой цепи, соответственно (фиг. 16). Вариабельные последовательности легкой цепи клонировали в рамке считывания в константную область IgK человека, а вариабельные последовательности тяжелой цепи клонировали в последовательность константной области Igy1 человека. В конечных экс-прессионных плазмидах, промоторы CMV человека запускали экспрессию кДНК-последовательностей как легкой, так и тяжелой цепей. Экспрессию и очистку рекомбинантного мышиного антитела ЕМ 164 выполняли в соответствии со способами, которые хорошо известны в данной области. Пример 2. Гуманизированные версии антитела ЕМ 164. Перестраивание поверхности антитела ЕМ164 для обеспечения гуманизированных версий в качестве терапевтических или диагностических агентов обычно выполняли в соответствии с принципами и способами, описанными в Патенте США 5639641, следующим образом. А. Предсказание поверхности. Доступность для растворителя остатков вариабельной области для ряда антител с расшифрованными структурами использовали для предсказания поверхностных остатков для вариабельной области мышиного антитела против IGF-I-рецептора (ЕМ164). Доступность аминокислот для растворителя для набора из 127 файлов уникальных структур антител (табл. 2) рассчитывали с использованием пакета программ МС (Pedersen et al., 1994, J. Biol. Chem., 235, 959-973). Десять наиболее сходных аминокислотных последовательностей легкой цепи и тяжелой цепи из этого набора из 127 структур определяли при помощи выравнивания последовательностей. Среднюю доступность для растворителя для каждого остатка вариабельной области рассчитывали и считали, что положения с большей, чем 30% средней доступностью, являются поверхностными остатками. Положения со средними доступностями между 25 и 35% дополнительно исследовали расчетом доступности индивидуального остатка только для структур с двумя идентичными фланкирующими остатками. Таблица 2. 127 структур антител из базы данных Brookhaven, используемые для предсказания поверхности антитела против IGF-I-рецептора (ЕМ 164) Файлы структур Brookhaven, использованные для предсказаний _последовательности_ 2rcs 3hfl 3hfin laif la3r lbbj 43c9 4fab 6fab 7fab 2gfb 2hlp 2hfl la6t laxt Ibog 2hrp 2jel 2mcp 2pcp lyuh 2bfv 2cgr 8fab 1ае6 lbvl 2dbl 2fl9 2fb4 2fbj lsm3 ltet lvfa glb2 Ia4j lcly lvge lyec lyed lyee lnsn lopg losp laj7 layl lclz Iplg lpsk lrmf lsbs lncd lnfd lngp lacy lafv lcbv lnld lnma Inmb lnqb lmcp lmfb lmim 15c8 la5f laxs Imlb lmpa lnbv lncb ljrh lkb5 lkel lap2 lb2w ladq lkip lkir live Imam ligi ligm ligt ladO lbaf lcfv ligy likf ljel Ijhl lgpo Ihil Ihyx laOq lbjm lclo liai libg ligc ligf Ifpt lfrg lfVc laqk lbln ld5b lgaf Iggi lghf lgig lfai lfbi lfdl lad9 lbbd lf58 lfgv lfig lflr lfor ldbl Idfb la31 lbfo leap ldsf ldvf B. Молекулярное моделирование. Молекулярную модель мышиного ЕМ164 генерировали с использованием пакета программ Oxford Molecular soft package AbM. Каркас антитела строили из файлов структур для антител с наиболее сходными аминокислотными последовательностями, которыми были 2jel для легкой цепи и 1nqb для тяжелой цепи. Неканонические CDR встраивали с использованием поиска в базе данных С-а-структур, содержащей неизбыточные расшифрованные структуры. Определяли остатки, которые лежат в 5А от CDR. C. Отбор Ab человека. Поверхностные положения мышиного ЕМ164 сравнивали с соответствующими положениями последовательностей антител человека в базе данных Kabat (Johnsob, G. and Wu, T.T. (2001) Nucleic Acids - 21 - 009807 Research, 29: 205-206). Программу SR управления базой данных антител (Searle 1998) использовали для извлечения и сопоставления поверхностных остатков антител из природных пар тяжелых и легких цепей антител человека. Поверхность антитела человека с наиболее идентичными поверхностными остатками с особым обращением внимания на положения, которые находятся в пределах 5А от CDR, были выбраны для замены поверхностных остатков мышиного антитела против IGF-I-рецептора. D. ПЦР-мутагенез. ПЦР-мутагенез выполняли на кДНК-клоне мышиного ЕМ164 (см. выше) для построения переделанного на поверхности антитела ЕМ 164 человека (здесь huEM164). Наборы праймеров конструировали для получения 8 аминокислотных замен, требуемых для всех испытанных версий huEM164, и конструировали дополнительные праймеры для альтернативного создания двух замен остатков в пределах 5А (табл. 3). ПЦР-реакции выполняли с использованием следующей программы: 1) 94°С 1 мин, 2) 94°С 15 сек, 3) 55°С 1 мин, 4) 72°С 1 мин, 5) цикл опять до стадии №229 раз, 6) окончание с конечной стадией удлинения при 72°С в течение 4 мин. ПЦР-продукты расщепляли их соответствующими рестриктазами и клонировали в клонирующие векторы pBlueScript, как описано выше. Клоны секвенировали для подтверждения желаемых аминокислотных замен. Таблица 3. ПЦР-праймеры, используемые для конструирования 4 гуманизированных антител ЕМ164 Праймер Последовательность SEQ ГО NO: Eml64hcvv CAGGTGTACACTCCCAGGTCCAACTGGTGCAGTCTGGGG CTGAAGTGGTG AAGCCTG Eml64hcqqgoll CAATCAGAAGTTCCAGGGGAAGGCCACAC Em]64hcqqgol2 CdTCCCCTGG A ACrTCTG ATTGTAG TT AGTACG Eml641cv3 CAGGTGTACACTCCGATGTTGTGATGACCCAAACrCC Eml641cl3 CAGGTGTACACTCCGATGTnTGATGACCCAAACTCC Eml641cpl8 G ACrAG ATCTGC AAG AG ATGG AGGCrGG ATCTCCA A GA С Eml641cbgl2 TTG СAGATCTAGTC AGAGCATAGTAC ATAGTAATG Eml64r45 GAATGGTACCrGCAGAAACCAGGCCAGTCTCCAAGGCrC CTGATCTAC Eml64a67oll GTGGCAGTGGAGCAGGGACAGATTTCAC Eml64a67ol2 GAAATCTGTCCCTGCTCCACTGCCACTG Е. Остатки поверхности вариабельной области. Способы перестройки поверхности антител, описанные Pedersen et al. (J. Biol. Chem., 235, 959-973, 1994) и Roguska et al. (Protein Eng., 9, 895-904, 1996), начинаются предсказанием остатков поверхности вариабельных последовательностей мышиного антитела. Остатком поверхности считают аминокислоту, которая имеет по меньшей мере 30% ее общей площади поверхности, доступной для молекулы воды. Были идентифицированы 10 наиболее гомологичных антител в наборе из 127 файлов структур антител (фиг. 17 и 18). Доступность для растворителя каждого положения Kabat усредняли для этих выровненных последовательностей, и распределение относительных доступностей для каждого остатка было таким, какое показано на фиг. 19. Как легкая, так и тяжелая цепи имеют 26 остатков со средними относительными доступностями по меньшей мере 30% (фиг. 19); таким образом, эти остатки являются предсказанными остатками поверхности для ЕМ164. Несколько остатков имели средние доступности между 25 и 35%, и их дополнительно исследовали усреднением только антител с двумя идентичными остатками, фланкирующими каждую сторону данного остатка (табл. 4 и 5). После этого дополнительного анализа исходный набор поверхностных остатков, который был идентифицирован выше, оставался неизменным. - 22 - 009807 Таблица 4. Поверхностные остатки и средняя доступность (ср. дост.) для вариабельных последовательностей легкой и тяжелой цепей антитела ЕМ 164 Остатки поверхности ЕМ 164 Легкая цепь Тяжелая цепь ЕМ164 № Kabat Ср. досг. ЕМ 164 № Kabat Ср. дост. 45.89 58.19 41.53 34.08 31.40 34.36 50.08 38.01 35.45 47.72 39.79 46.51 34.36 31.49 32.63 31.42 34.35 34.41 36.38 31.23 43.05 36.24 46.56 44.01 34.92 42.62 30.58 46.85 30.40 46.68 41.46 44.87 42.41 38.92 45.96 40.06 38.20 35.92 42.61 48.91 38.46 82В 32.72 95Е 34.83 35.21 103 31.10 39.62 107 36.94 36.00 108 60.13 106 37.65 109 53.65 113 43.42 Таблица 5 Остатки границы поверхности Легкая цепь Тяжелая цепь ЕМ164 № Kabat Ср. дост. ЕМ164 № Rabat Ср. дост. 28.68 31.62 30.24 36.07 26.59 29.88 25.20 30.87 25.73 25.64 25.37 35.06 36.73 31.48 31.00 30.53 32.29 27.12 27А 29.78 27С 29.05 27.71 24.65 34.92 18.80 32.24 82В 32.87 30.02 29.50 26.03 103 111 25.95 26.64 29.61 95Е 42.12 100 29.82 103 31.10 205 25.78 Остатки, которые имеют средние доступности между 25% и 35%, дополнительно анализировали усреднением субпопуляции антител, которые имеют два идентичных остатка, фланкирующих каждую сторону рассматриваемого остатка. NA обозначает остатки, не имеющие идентичных фланкирующих - 23 - 009807 остатков в 10 наиболее сходных антителах. F. Молекулярное моделирование для определения, какие остатки попадают в пределы 5А от CDR. Описанную выше молекулярную модель, генерированную с использованием пакета программ AbM, анализировали для определения, какие остатки поверхности ЕМ164 были в пределах 5А от CDR. Для переделывания поверхности мышиного антитела ЕМ 164 все остатки поверхности, находящиеся вне CDR, должны быть заменены копиями человека, но остатки в пределах 5А от CDR обрабатывали с особой осторожностью, так как они могут также вносить вклад в антигенную специфичность. Таким образом, эти последние остатки должны быть идентифицированы, и к ним следует осторожно относиться в ходе всего процесса гуманизации. Определения CDR, используемые для переделывания поверхности, объединяют AbM-определение для CDR 2 тяжелой цепи и определения Kabat для остальных 5 CDR (фиг. 14). Табл. 6 показывает остатки, которые находятся в пределах 5 А от любого остатка CDR в последовательности либо легкой, либо тяжелой цепи модели ЕМ164. Таблица 6. Остатки поверхности каркаса антитела ЕМ 164 в пределах 5А от CDR Легкая цепь Тяжелая цепь Т28 К73 S74 Р40 Q42 К45 G57 D60 Е81 G. Идентификация наиболее гомологичных поверхностей антител человека. Кандидатные поверхности антител человека для перестройки поверхности ЕМ164 идентифицировали в пределах базы данных антител Kabat, с использованием программного обеспечения SR, которое обеспечило поиск только указанных положений остатков против этой базы данных. Для сохранения природных пар, сравнивали вместе поверхностные остатки как легкой, так и тяжелой цепей. Наиболее гомологичные человеческие остатки из базы данных Kabat сопоставляли в ранжированном порядке идентичности последовательности. Верхние 5 поверхностей приведены в табл. 7. Затем эти последовательности сравнивали для идентификации, какие из них, могли бы требовать наименьших замен в 5 А от CDR. Антитело против лейкозных В-клеток, CLL 1.69, требовало наименьшего количества замен поверхностных остатков (всего 10), и только два из этих остатков находились в пределах 5 А от CDR. Полноразмерную последовательность вариабельной области для ЕМ 164 также сопоставляли против базы данных Kabat, и CLL 1.69 опять идентифицировали как наиболее сходную последовательность вариабельной области человека. Вместе, эти сравнения последовательностей идентифицировали антитело лейкозных В-клеток CLL 1.69 как предпочтительный выбор в качестве человеческой поверхности для ЕМ164. Таблица 7. Верхние 5 человеческих последовательностей, извлеченных из базы данных Kabat 5 наиболее гомологичных поверхностей антитела человека Антитело Легкая цепь SEQ ID N0: МиЕМ164 DLTLLQPGQKGDS REKKRA 33 34 35 36 37 38 CLL1.69 DVTLLP PGQRGDAREKKR-DQSLI Р PG QKGDS RDKKRA DMS S VR PGQKGS S S DKKR-Е VS GPRPGQRGDS REKKR-ЕVS GPRPG QRGDS REKKR- MSL5 CDP57I ЬСЗаРВ SSbPB Антитело Тяжелая цепь SEQIDNO: МиЕМ164 Q QQALKPGKKTPGQEKKRKSSSEAS CLL1.69 Q QVAVKPGKKT Р GQQKQGKS S S EQS MSL5 Q QQP LKPGKKT РGKDDKGTSNNEQS CDP571 Q QVAVKPGKKT Р GQQKKGKS S S EQS ЬСЗаРВ - QVAVKPGKKTРGQQKOGKSSSEQS SSbPB - QVAVKPGKKT Р GQQKQGE S SEQS Сопоставления были получены с использованием SR (Pedersen 1993). Остатки поверхности ЕМ164, которые находятся в пределах 5 А CDR, подчеркнуты. Н. Конструирование гуманизированных генов ЕМ164. Десять замен поверхностных остатков для ЕМ164 (табл. 7) производили с использованием способов ПЦР-мутагенеза, как описано выше. Поскольку восемь из поверхностных остатков для CLL 1.69 не находились в пределах 5 А от CDR, эти остатки заменяли с мышиных на человеческие во всех версиях гуманизированного ЕМ 164 (табл. 8 и 9). Два остатка поверхности легкой цепи, которые находились в пределах 5 А от CDR, (положения Kabat 3 и 45), были либо заменены на человеческие, либо оставлены как мышиные. Вместе эти опции генерируют четыре гуманизированные версии ЕМ164, которые были скон - 24 - 009807 струированы (фиг. 22 и 23). Из этих четырех гуманизированных версий, версия 1.0 имеет все 10 человеческих поверхностных остатков. Наиболее консервативной версией в отношении замен вблизи от CDR является версия 1.1, которая сохранила оба из мышиных поверхностных остатков, которые находились в пределах 5 А от CDR. Все четыре гуманизированных антитела ЕМ 164 клонировали в экспрессирующую антитело плазмиду (фиг. 16) для использования в транзиторной и стабильной трансфекциях. Таблица 8. Замены остатков для версий 1.0-1.3 гуманизированного антитела ЕМ 164 Изменения во всех версиях Легкая цепь: muQ18 в huP18; muS67BhuA67 Тяжелая цепь muQ5 в huV5; muR65 ehuG65: muLl 1 в huV 11; muE61 to huQ61; muK64 to huQ64; muA106BhuQ106__ Изменения huEM 164 Аминокислота 45 легкой цепи Аминокислота 3 легкой цепи Всего Мышиные остатки vl.O vl.l vl.2 vl.3 I. Сравнение аффинностей гуманизированных версий антитела EM164 с аффинностями мышиного антитела ЕМ 164 в отношении связывания с полноразмерным IGF-I-рецептором и укороченной альфа-цепью IGF-I-рецептора. Афинности гуманизированных версий 1.0-1.3 антитела ЕМ164 сравнивали с аффинностями мышиного антитела ЕМ164 при помощи конкурентных анализов связывания с использованием биотинилиро-ванного полноразмерного IGF-I-рецептора человека или меченой myc-эпитопной меткой укороченной альфа-цепи IGF-I-рецептора, описанной выше. Пробы гуманизированных антител ЕМ164 получали транзиторной трансфекцией подходящих экспрессирующих векторов в клетках 293Т эмбриональной почки человека, и концентрации антител определяли при помощи ELISA с использованием стандартов очищенного гуманизированного антитела. Для измерений конкурентного связывания при помощи ELISA смеси проб гуманизированных антитела и различных концентраций мышиного антитела ЕМ164 инкубировали с захваченными опосредованным способом битинилированного полноразмерного IGF-I-рецептора или меченой myc-эпитопной меткой укороченной альфа-цепи IGF-I-рецептора. После уравновешивания связанное гуманизированное антитело детектировали с использованием конъюгата козье антитело против Fab'2 человека-пероксидаза хрена. Графики ([связанное мышиное Ab]/[связанное гуманизированное Ab]) против ([мышиное Ab]/[гуманизированное Ab]), которые дают теоретически прямую линию с наклоном = (K D гуманизированного Ab/KD мышиного Ab, b), использовали для определения относительных аффинностей гуманизированных и мышиных антител. Пример конкурентного анализа показан на фиг. 11. Immulon-2НВ-планшет ELISA покрывали 100 мкл 5 мкг/мл стрептавидина на лунку в карбонатном буфере при температуре окружающей среды в течение 7 ч. Покрытые стрептавидином лунки блокировали 200 мкл блокирующего буфера (10 мг/мл БСА в TBS-T-буфере) в течение 1 ч, промывали TBS-T-буфером и инкубировали с биотинилированным IGF-I-рецептором (5 нг на лунку) в течение ночи при 4°С. Затем лунки, содержащие опосредованно захваченный биотинилированный IGF-I-рецептор, промывали и инкубировали со смесями гуманизированного антитела ЕМ164 (15,5 нг) и мышиного антитела (0 нг или 16,35 нг или 32,7 нг или 65,4 нг или 163,5 нг) в 100 мкл блокирующего буфера в течение 2 ч при температуре окружающей среды и затем инкубировали в течение ночи при 4°С. Затем лунки промывали TBS-T-буфером и инкубировали с конъюгатом козье антитело против Fab'2 человека-пероксидаза хрена в течение 1 ч (100 мкл; 1 мкг/мл в блокирующем буфере) с последующими промывками и детектированием с использованием субстрата ABTS/H2O2 при 405 нм. График ([связанное мышиное Ab]/[связанное гуманизированное Ab]) против ([мышиное Ab]/[гуманизированное Ab]) давал прямую линию (r2 = 0,996) с наклоном (=KD гуманизированного Ab/KD мышиного Ab) 0,52. Таким образом, версия 1.0 гуманизированного антитела связывалась с IGF-I-рецептором более прочно, чем мышиное антитело ЕМ164. Сходные величины для градиента в диапазоне от приблизительно 0,5 до 0,8 получали для конкуренций версий 1.0, 1.1, 1.2 и 1.3 гуманизированных антител ЕМ164 с мышиным антителом ЕМ 164 за связывание с полноразмерным IGF-I-рецептором или с укороченной альфа-цепью IGF-I-рецептора, что указывало на то, что все из этих гуманизированных версий антитела ЕМ164 имели сходные аффинности, которые все были лучшими, чем аффинность исходного мышиного антитела ЕМ164. Химерная версия антитела ЕМ164 с мутацией 92F-C в тяжелой цепи показала наклон приблизительно 3 в подобной конкуренции связывания с мышиным антителом ЕМ164, что указывало на - 25 - 009807 то, что мутант 92F- C EM164 имел в 3 раза более низкую аффинность, чем мышиное антитело ЕМ164 в отношении связывания IGF-I-рецептора. Гуманизированное антитело ЕМ164 v1.0 показало сходное ингибирование IGF-I-стимулируемого роста и выживания клеток MCF-7 в сравнении с мышиным антителом ЕМ164 (фиг. 24). Ингибирование стимулируемого сывороткой роста и выживания клеток MCF-7 гуманизированным антителом ЕМ164 v1.0 было сходным с ингибированием мышиным антителом ЕМ164. Таблица 9 Сегмент Легкая цепь Тяжелая цепь FR1 1 -23 (со случайным остатком при 0 и делецией при 10 в цепях Vi} 1-30 (со случайным остатком при 0) CDR1 24-34 (с возможными инсерция- 31-35 (с возможными инсерциями, ями, пронумерованными как 27А, В, С, D, Е, F) пронумерованными как 35А, В) FR2 35-49 36-49 CDR2 50-56 50-65 (с возможными инсерциями, пронумерованными как 52А, В, С) FR3 57-88 66-94 (с возможными инсерциями, пронумерованными как 82А, В, С) CDR3 89-97 (с возможными инсерция- 95-102 (с возможными инсерциями, ми, пронумерованными как 95А, В, С, D, Е, F) пронумерованными как 100А, В, С, D, Е, F, G, Н, I, J, К) FR4 98-107 (с возможной инсерцией, пронумерованной как 106А) 103-113 Использовали систему нумерации Kabat для полипептидов вариабельных областей легкой и тяжелой цепей различных версий ЕМ164 Ab. Аминокислотные остатки сгруппированы в каркасные (FR) и определяющие комплементарность районы (CDR) в соответствии с положением в полипептидной цепи. Взято из Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, 1991, NIH Publication No. 91-3242. J. Способ обеспечения улучшенных антител против IGF-I-рецептора на основе последовательностей мышиного и гуманизированных антител, описанных здесь. Аминокислотные последовательности и последовательности нуклеиновых кислот антитела против IGF-I-рецептора ЕМ 164 и его гуманизированных версий использовали для развития других антител, которые имели улучшенные свойства и которые также находятся в объеме данного изобретения. Такие улучшенные свойства включают в себя увеличенную аффинность в отношении IGF-I-рецептора. Несколько исследований изучили действия введения одной или нескольких аминокислотных замен в различных положениях в последовательности антитела, на основании знания последовательности первичного антитела, на их свойства, такие как связывание и уровень экспрессии (Yang, W.P. et al., 1995, J. Mol. Biol, 254, 392-403; Rader, С et al., 1998, Proc. Natl. Acad. Sci. USA, 95, 8910-8915; Vaughan, T.J. et al., 1998, Nature Biotechnology, 16, 535-539). В этих исследованиях варианты первичного антитела получали изменением последовательностей генов тяжелой и легкой цепей в районах CDR1, CDR2, CDR3 или каркасных районах с использованием таких способов, как опосредованный олигонуклеотидом сайт-направленный мутагенез, кассетный мутагенез, склонная к ошибкам ПЦР, перетасовка ДНК или мутаторные штаммы Е. coli (Vaughan, T.J. et al., 1998, Nature Biotechnology, 16, 535-539; Adey, N.B. et al., 1996, Chapter 16, pp. 277-291, in "Phage Display of Peptides and Proteins", Eds. Kay, B.K. et al., Academic Press). Эти способы изменения последовательности первичного антитела привели, посредством применения стандартных способов скрининга, к улучшенным аффинностям таких вторичных антител (Gram, H. et al, 1992, Proc. Natl. Acad. Sci. USA, 89, 35763580; Boder, E.T. et al., 2000, Proc. Natl. Acad. Sci. USA, 97, 10701-10705; Davies, J. and Riechmann, L., 1996, Immunology, 2, 169-179; Thompson, J. et al., 1996, J. Mol. Biol., 256, 77-88; Short, M.K. et al., 2002, J. Biol. Chem., 277, 16365-16370; Furukawa, K. et al., 2001, J. Biol. Chem., 276, 27622-27628). С использованием подобной направленной стратегии изменения одного или нескольких аминокислотных остатков антитела, последовательности антител, описанные в данном изобретении, могут быть использованы для развития антител против IGF-I-рецептора с улучшенными функциями, например, антител, имеющих подходящие группы, такие как свободные аминогруппы или тиолы, в удобных точках присоединения для ковалентной модификации, например, для использования в присоединении терапевтических агентов. К. Альтернативная система экспрессии для мышиного, химерного и других антител против IGF-I-рецептора. Мышиное антитело против IGF-I-рецептора экспрессировали также при помощи экспрессирующих плазмид млекопитающих, сходных с плазмидами, используемыми для экспрессии гуманизированного антитела (см. выше). Известно, что экспрессирующие плазмиды имеют мышиные константные области, включающие в себя последовательности легкой цепи каппа и тяжелой цепи гамма-1 (McLean et al., 2000, - 26 - 009807 Mol Immunol., 37, 837-845). Эти плазмиды сконструированы для принятия любой вариабельной области антитела, например, мышиного антитела против IGF-I-рецептора, посредством простого расщепления рестриктазами и клонирования. Обычно требуется дополнительная ПЦР этого антитела против IGF-I-рецептора для создания рестрикции, совместимой с рестрикцией в этой экспрессирующей плазмиде. Альтернативным подходом для экспрессии полностью мышиного антитела против IGF-I-рецептора была замена константных областей человека в плазмиде, экспрессирующей химерное антитело против IGF-I-рецептора. Эту плазмиду химерной экспрессии (фиг. 16) конструировали с использованием кассет для вариабельных областей и для константных областей как легкой, так и тяжелой цепей. Как раз, когда вариабельные последовательности антитела клонировали в эту экспрессионную плазмиду посредством расщеплений рестриктазами, отдельные продукты расщепления рестриктазами использовали для клонирования в любых последовательностях константной области. Например, кДНК легкой цепи каппа и кДНК тяжелой цепи гамма-1 клонировали из РНК мышиной гибридомы, такой как РНК, описанная выше для клонирования вариабельных областей антитело против IGF-I-рецептора. Подобным образом, конструировали подходящие праймеры из последовательностей, доступных в базе данных Kabat (см. табл. 10). Например, использовали ОТ-ПЦР для клонирования последовательностей константной области и для создания сайтов рестрикции, необходимых для клонирования этих фрагментов в плазмиду, экспресси-рующую химерное антитело против IGF-I-рецептора. Затем эту плазмиду использовали для экспрессии полностью мышиного антитела против IGF-I-рецептора в стандартных экспрессионных системах млекопитающего, таких как клеточная линия СНО. Таблица 10. Праймеры, сконструированные для клонирования мышиной константной области гамма-1 и мышиной константной области каппа, соответственно Праймеры мышиной константной области Название праймера Последовательность праймера SEQ ID NO: MulgGI C3endX TTTTGAGCTClTATTTACCAGGAGAGTGGGAGA GGCTCTT MulgGI C5endH TTTTAAGCTTGCCAAAACGACACCCCCATCTGTCTAT MulgKap C3endB TTTTGGATCCTAACACTCATTCCTGTTGAAGC MulgKap C5endE TTTTGAATTCGGGCTGATGCTGCACCAACTG Эти праймеры были сконструированы на основании последовательностей, доступных в базе данных Kabat (Johnson, G. and Wu, T.T. (2001) Nucleic Acids Research, 29: 205-206). Заявление о депонировании Гибридома, продуцирующая мышиное антитело ЕМ 164, была депонирована в Американской Коллекции Типовых Культур, РО Box 1549, Manassas, VA 20108, 14 июня 2002 г. согласно Будапештскому Договору и получила номер доступа АТСС (АТСС accession number) PTA-4457. В данном описании делались ссылки на некоторые патенты и печатные публикации, содержание которых, в каждом случае, включены здесь путем отсылки в их соответствующем полном виде. Хотя данное изобретение было описано подробно и со ссылкой на его конкретные варианты осуществления, специалисту с квалификацией в данной области будет понятно, что различные изменения и модификации могут быть произведены в нем без отклонения от его идеи и объема. ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Композиция, содержащая: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент и где указанное антитело или указанный фрагмент специфически связывается с рецептором инсулин-подобного фактора роста I, выбранный из группы, состоящей из: (i) антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), (ii) антитела с перестроенной поверхностью или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (iii) человеческого или гуманизированного антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (iv) функционального эквивалента антитела или его эпитопсвязывающего фрагмента, имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, (v) варианта мышиного антитела ЕМ164 или его эпитопсвязывающего фрагмента, имеющих по меньшей мере одну мутацию, делецию или инсерцию нуклеотида в сравнении с мышиным антителом ЕМ164 и имеющих ту же самую специфичность связывания, что и мышиное антитело ЕМ164, и (vi) мышиного антитела ЕМ164, продуцируемого мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязывающего фрагмента, и (b) второй терапевтический агент. - 27 - 009807 2. Композиция по п.1, где указанный второй терапевтический агент выбран из группы, состоящей из таких терапевтических агентов, как доцетаксел, паклитаксел, доксорубицин, эпирубицин, циклофос-фамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, анастрозол, фулвест-рант, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорел-бин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идаруби-цин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2^ мелфалам, бортезомиб (Velcade), алтретамин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), антитело к EGF-рецептору (Cetuxivmab, Abx-EGF) и эпотилон. 3. Композиция по п.1, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитабина и камптотецина. 4. Композиция по п.1, где указанный первый терапевтический агент вводят пациенту в дозе приблизительно 1 - приблизительно 2000 мг/кв.м и где указанный второй терапевтический агент вводят в дозе приблизительно 10 - приблизительно 2000 мг/кв. м. 5. Композиция по п.1, где указанный первый терапевтический агент вводят пациенту в дозе приблизительно 10 - приблизительно 1000 мг/кв. м и где указанный второй терапевтический агент вводят в дозе приблизительно 50 - приблизительно 1000 мг/кв.м. 6. Фармацевтическая композиция, содержащая композицию по п.1 и фармацевтически приемлемый носитель или разбавитель. 7. Композиция, содержащая: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или эпитопсвязывающий фрагмент антитела, содержащий по меньшей мере один определяющий комплементарность район, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SYWMH {SEQIDNO:l), EINPSNGRTNYNEKFKR (SEQ ID N0:2), GRPDYYGSSKWYFDV (SEQ ID N0:3), RSSQSIVHSNVNTYLE (SEQ ID N0:4), KVSNRFS (SEQ ID NO:5) и FQGSHVPPT (SEQ ID N0:6) и (b) второй терапевтический агент, где, когда выбрана SEQ ID NO:5, указанное антитело или фрагмент антитела специфически связывается с рецептором инсулин-подобного фактора роста I (IGF-IR). 8. Композиция, содержащая: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или эпитопсвязывающий фрагмент антитела, содержащий по меньшей мере одну вариабельную область тяжелой цепи и по меньшей мере одну вариабельную область легкой цепи, где указанная вариабельная область тяжелой цепи содержит три последовательных определяющих комплементарность района, имеющего аминокислотные последовательности, представленные SEQ ГО N0:1-3, соответственно: SYWMH (SEQIDNO:l), EINPSNGRTNYNEKFKR (SEQ ГО N0:2), GRPDYYGSSKWYFDV (SEQ ID NO:3), и где указанная вариабельная область легкой цепи содержит три последовательных определяющих ком-плементарность района, имеющего аминокислотные последовательности, представленные SEQ ID NO:4-6, соответственно: RSSQSIVHSNVNTYLE (SEQ ID N0:4), KVSNRFS (SEQ ID N0:5), FQGSHVPPT (SEQ ID N0:6) и (b) второй терапевтический агент. 9. Композиция, содержащая: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент антитела, причем указанное антитело или указанный фрагмент содержит вариабельную область тяжелой цепи, которая имеет по меньшей мере 90% идентичность последовательности относительно аминокислотной последовательности, представленной SEQ ID N0:7: QVQLQQSGAELVKPGASVKLSCKASGYTFTSYWMHWVKQRPGQGLEWIGEI NPSNGRraYHEKFKRKATLTVDKSSSTAYMQLSSLTSEDSAVYYFARGRPDY YGSSKWYFDVWGAGTTVTVSS (SEQ ID NO:7), и (b) второй терапевтический агент. 10. Композиция по п.9, где указанная вариабельная область тяжелой цепи имеет по меньшей мере - 28 - 009807 95% идентичность последовательности относительно указанной аминокислотной последовательности, представленной SEQ ID NO:7. 11. Композиция по п.9, где указанная вариабельная область тяжелой цепи имеет аминокислотную последовательность, которая представлена SEQ ID NO:7. 12. Композиция, содержащая: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область легкой цепи, которая имеет по меньшей мере 90% идентичность последовательности относительно аминокислотной последовательности, представленной SEQ ID N0:8: DVLMTQTPLSLPVSLGDQASISCRSSQSIVHSNVNTYLEWYLQKPGQSPKLLIY KVSNRFSGVPDRFSGSGSGTGFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGTK LEIKR(SEQIDNO:8),H (b) второй терапевтический агент. 13. Композиция по п.12, где указанная вариабельная область легкой цепи имеет по меньшей мере 95% идентичность последовательности относительно указанной аминокислотной последовательности, представленной SEQ ID NO:8. 14. Композиция по п.12, где указанная вариабельная область легкой цепи имеет аминокислотную последовательность, которая представлена SEQ ID NO:8. 15. Композиция, содержащая: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область легкой цепи, имеющую последовательность, выбранную из группы, состоящей из: DVVMTQTPLSLPVSLGDPASISCRSSQSIVHSNVNTLEWYLQKPGQSPRLLIY KVSNRFSGVPDRFSGSGAGTDFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGTKLEIK R (SEQ ID N0:9); DVLMTQTPLSLPVSLGDPASISCRSSQSrVHSNVNTYLEWYLQKPGQSPKLLIY KVSNRFSGVPDRPSGSGAGTDFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGTKLEIK R (SEQ ID N0:10); DVLMTQTPLSLPVSLGDPASISCRSSQSRVHSNVNTYLEWYLQKPGQSPRLLIY KVSNRFSGVPDRFSGSGAGTDFTLRISRVEAEDLGIYYCFQGSHVPPTFGGGTKLEIK R(SEQ ID NO: 11); DVVMTQTPLSLPVSLGDPASISCRSSQSRVHSNVNTYLEWYLQKPGQSPKLLIY KVSNRFSGVPDRFSGSGAGTDFTLRISRVEAEDLGIYYGFQGSHVPPTFGGGTI":LEIK R(SEQ ID N0:12), (b) второй терапевтический агент. 16. Композиция, содержащая: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело или его эпитопсвязывающий фрагмент, причем указанное антитело или указанный фрагмент содержит вариабельную область тяжелой цепи, имеющую последовательность, представленную SEQ ID N0:13: QVQLVQSGAEVVKPGASVKLSCKASGYTFTSYWMHWVKQRPGQGLEWIGEI NPSNGRTNYNQKFQGKATLTVDKSSSTAYMQLSSLTSEDSAVYYFARGRPDY YGSSKWYFDVWGQGTTVTVSS (SEQ ID NO:13), И (b) второй терапевтический агент. 17. Композиция по любому из пп.7-16, где указанный второй терапевтический агент выбран из группы, состоящей из доцетаксела, паклитаксела, доксорубицина, эпирубицина, циклофосфамида, тра-стузумаба (Herceptin), капецитабина, тамоксифена, торемифена, летрозола, анастрозола, фулвестранта, экземестана, госерелина, оксалиплатина, карбоплатина, цисплатина, дексаметазона, антида, бевацизума-ба. (Avastin), 5-фторурацила, лейковарина, левамизола, иринотекана, этопозида, топотекана, гемцитаби-на, винорелбина, эстрамустина, митоксантрона, абареликса, золедроната, стрептозоцина, ритуксимаба (Rituxan), идарубицина, бусульфана, хлорамбуцила, флударабина, иматиниба, цитарабина, ибритумомаба (Zevalin), тозитумомаба. (Bexxar), интерферона альфа-2b, мелфалама, бортезомиба (Velcade), алтретами-на, аспарагиназы, гефитиниба (Iressa), эрлонитиба (Tarceva), антитела против EGF-рецептора (Cetux-ivmab, Abx-EGF) и эпотилона. 18. Композиция по любому из пп.7-16, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитаби - 29 - 009807 на и камптотецина. 19. Способ ингибирования роста раковой клетки, предусматривающий контактирование указанной клетки с композицией по п.1. 20. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества композиции по п.1. 21. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества фармацевтической композиции по п.6. 22. Способ лечения по любому из пп.19-21, где указанный рак является раком, выбранным из группы, состоящей из рака молочной железы, рака ободочной кишки, рака яичника, остеосаркомы, рака шейки матки, рака предстательной железы, рака легкого, синовиального рака, рака поджелудочной железы, меланомы, множественной миеломы, нейробластомы и рабдомиосаркомы. 23. Набор, содержащий: (a) первый терапевтический агент, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязываю-щий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) второй терапевтический агент и (c) инструкции по применению. 24. Способ ингибирования роста раковой клетки, предусматривающий контактирование указанной клетки с: (a) первым терапевтическим агентом, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемое мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязы-вающий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) вторым терапевтическим агентом. 25. Способ лечения пациента, имеющего рак, предусматривающий введение указанному пациенту эффективного количества: (a) первого терапевтического агента, где указанным первым терапевтическим агентом является антитело, имеющее ту же самую аминокислотную последовательность, что и мышиное антитело ЕМ164, продуцируемого мышиной гибридомой ЕМ164 (номер доступа АТСС РТА-4457), или его эпитопсвязы-вающий фрагмент, где указанное антитело или указанный фрагмент специфически связываются с рецептором инсулин-подобного фактора роста I, (b) второго терапевтического агента. 26. Способ по п.24, где указанную клетку контактируют с указанным первым терапевтическим агентом и указанным вторым терапевтическим агентом одновременно. 27. Способ по п.24, где указанную клетку контактируют с указанным первым терапевтическим агентом и указанным вторым терапевтическим агентом последовательно и в любом порядке. 28. Способ по п.25, где указанный первый терапевтический агент и указанный второй терапевтический агент вводят одновременно. 29. Способ по п.25, где указанный первый терапевтический агент и указанный второй терапевтический агент вводят последовательно и в любом порядке. 30. Способ по п.24 или 25, где указанный второй терапевтический агент выбран из группы состоящей из таких терапевтических агентов, как доцетаксел, паклитаксел, доксорубицин, эпирубицин, цикло-фосфамид, трастузумаб (Herceptin), капецитабин, тамоксифен, торемифен, летрозол, анастрозол, фулве-странт, экземестан, госерелин, оксалиплатин, карбоплатин, цисплатин, дексаметазон, антид, бевацизумаб (Avastin), 5-фторурацил, лейковарин, левамизол, иринотекан, этопозид, топотекан, гемцитабин, винорел-бин, эстрамустин, митоксантрон, абареликс, золедронат, стрептозоцин, ритуксимаб (Rituxan), идаруби-цин, бусульфан, хлорамбуцил, флударабин, иматиниб, цитарабин, ибритумомаб (Zevalin), тозитумомаб (Bexxar), интерферон альфа-2^ мелфолам, бортезомиб (Velcade), алтретамин, аспарагиназа, гефитиниб (Iressa), эрлонитиб (Tarceva), анти-EGF-рецептор-антитело (Cetuxivmab, Abx-EGF) и эпотилон. 31. Способ по п.24 или 25, где указанный второй терапевтический агент выбран из группы, состоящей из карбоплатина, оксалиплатина, цисплатина, паклитаксела, доцетаксела, гемцитабина и камптоте-цина. - 30 - 009807 Список последовательностей <110> ImmunoGen, inc. <120> ANTI-IGF-I RECEPTOR ANTIBODY <130> F190322 <140> 10/729,441 <141> 2003-12-08 <150> 10/170,390 <151> 2002-06-14 <Ш> 98 <170> Patentln version 3.3 <210> 1 <211> S <212> PRT <213> Artificial sequence <220;> <223> antibody heavy chain complementarity determining region <400> 1 ser туг Тгр Met His <210> 2 <211> 17 <212> PRT <213> Artificial sequence <220> <223> antibody heavy chain complementarity determining region <400> 2 Glu lie Asn Pro Ser Asn Gly Arg Thr Asn Туг Asn Glu Lys Phe Lys 15 10 15 Arg <210> 3 <211> 15 <212> PRT <213> Artificial Sequence <220> <223> antibody heavy chain complementarity determining region <400> 3 Gly Arg Pro Asp туг туг Gly ser Ser Lys тгр туг Phe Asp val 15 10 15 <210> 4 <211> 16 <212> PRT <213> Artificial sequence <220> <223> antibody light chain complementarity determining region <400> 4 Arg Ser Ser Gin ser lie val His Ser Asn Val Asn Thr Tyr Leu Glu 15 10 15 <210> 5 <211> 7 <212> PRT <213> Artificial sequence <220> <223> antibody light chain complementarity determining region - 31 - 009807 - 32 - 009807 - 33 - 009807 - 34 - 009807 <2XQ> 16 <211> 30 <212> DNA <213> Artificial <220> Sequence - 35 - 009807 - 36 - 009807 - 37 - 009807 - 38 - 009807 - 39 - 009807 - 40 - 009807 - 41 - 009807 - 42 - 009807 - 43 - 009807 - 44 - 009807 - 45 - 009807 - 46 - 009807 - 47 - 009807 - 48 - 009807 ¦ 49 ¦ 009807 Gin val Gin Leu Gin Gin ser Gly Ala Glu Leu val Lys Pro Gly Ala 15 10 IS Ser Val Lys Leu ser cys Lys Ala ser Gly Tyr Thr Phe Thr Ser туг 20 25 30 тгр Met His Trp Val Lys Gin Arg pro Gly Arg Gly Leu Glu Тгр Пе 35 40 45 Gly Arg lie Asp Pro Asn ser Gly Gly Thr Lys Tyr Asn Glu Lys Phe 50 55 60 Lys Ser Lys Ala Thr Leu Thr val Asp Lys pro Ser Ser Thr Ala Tyr Met Gin Leu ser ser Leu Thr Ser Glu Asp Ser Ala Val туг туг cys 85 90 95 Ala Arg туг Asp туг Tyr Gly Ser Ser Tyr Phe Asp Tyr Trp Gly Gin 100 105 110 Gly Thr Thr Val Thr Val Ser Ser 115 120 <210> 72 <211> 120 <212> PRT <213> Artificial sequence <220> <22J> synthetic antibody structure <400> 72 Gin val Gin Leu Gin Gin pro Gly Ala Glu Leu val Lys Pro Gly Ala 15 10 15 ser val Lys Leu ser cys Lys Ala Ser Gly туг Thr Phe Thr ser туг 20 25 30 тгр Met His Trp val Lys Gin Arg pro Gly Arg Gly Leu Glu тгр lie 35 40 45 Gly Arg lie Asp Pro Asn ser Gly Gly Thr Lys Tyr Asn Glu Lys Phe 50 55 60 Lys ser Lys Ala Thr Leu Thr Val Asp Lys Pro Ser ser Thr Ala Tyr 65 70 75 80 Met Gin Leu ser Ser Leu Thr Ser Glu Asp ser Ala val туг туг Cys 85 90 95 Ala Arg Tyr Asp Tyr Tyr Gly Ser Ser Tyr Phe Asp Tyr Trp Gly Gin 100 105 110 Gly Thr Thr Leu Thr val Ser Ser 11S 120 <210> 7 J <211> 122 <212> PRT <213> Artificial sequence <220> <223> synthetic antibody structure <400> 73 Gin Val Gin Leu Gin Gin Pro Gly Ala Glu Leu Val Lys Pro Gly Ala ¦ 50 ¦ 009807 10 15 Ser Val Lys Leu ser cys Lys Ala ser Gly Tyr Thr Phe Thr ser туг 20 25 30 Trp Met His Trp val Lys Gin Gly Pro Gly Gin Gly Leu Glu Trp lie 35 40 45 Gly Glu lie Asp pro ser Asp ser туг Pro Asn Tyr Asn Glu Lys Phe 50 55 60 Lys Gly Lys Ala Thr Leu Thr val Asp Lys Ser ser Ser Thr Ala туг 65 70 75 80 Met Gin Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala val туг ту^г cys 85 90 Ala Ser Leu Tyr Tyr Tyr Gly Thr ser Tyr Gly val Leu Asp туг тгр 100 105 110 Gly Gin Gly Thr ser Val Thr Val ser Ser 115 120 <210> 74 <211> 120 <212> PRT <213> Artificial Sequence <220> <223> synthetic antibody structure <400> 74 Gin Val Gin Leu Gin Gin Pro Gly ser val Leu val Arg pro Gly Ala 15 10 15 Ser Val Lys Leu Ser Cys Lys Ala ser Gly Tyr Thr Phe Thr ser Ser 20 25 30 тгр He His Trp Ala Lys Gin Arg Pro Gly Gin Gly Leu Glu тгр lie 35 40 45 Gly Glu He His Pro Asn ser Gly Asn Thr Asn туг Asn Glu Lys Phe 50 55 60 Lys Gly Lys Ala Thr Leu Thr val Asp Thr Ser ser ser Thr Ala Tyr 65 70 75 80 val Asp Leu Ser ser Leu Thr ser Glu ASP Ser Ala val туг туг cys 85 90 95 Ala Arg Trp Arg туг Gly ser Pro туг Туг Phe Asp туг тгр Gly Gin 100 105 110 Gly Thr Thr Leu Thr val Ser Ser 115 120 <210> 75 <211> 118 <212> PRT <213> Artificial sequence <220> <22i> synthetic antibody structure <400> 75 Gin val Gin Phe Gin Gin Ser Gly Ala Glu Leu val Lys Pro Gly Ala IS 10 15 - 51 - 009807 - 52 - 009807 - 53 - 009807 - 54 - 009807 - 55 - 009807 - 56 - 009807 - 57 - 009807 - 58 - 009807 Ser Val Lys Leu ser Cys Lys Ala ser Gly туг Thr Phe Thr Ser туг 20 25 30 тгр Met His Trp val Lys Gin Arg pro Gly Gin Gly Leu Glu Trp lie 35 40 45 Gly Glu lie Asn Pro Ser Asn Gly Arg Thr Asn Ty^r Asn Gin Lys Phe Gin Gly Lys Ala Thr Leu Thr val Asp Lys Ser ser Ser Thr Ala Tyr 65 70 75 80 Met Gin Leu ser Ser Leu Thr ser Glu Asp ser Ala val туг Tyr Phe 85 90 95 Ala Arg Gly Arg Pro Asp туг Tyr Gly ser Ser Lys Trp Tyr Phe Asp 100 105 110 val тгр Gly Gin Gly Thr Thr val Thr val ser 115 120 <210> 89 <211> 339 <212> DNA <213> Artificial Sequence <220> <223> variable region of humanized EM164 antibody - light chain <220> <221> CDS <222> (D..C339) <400> 89 gat gtt gtg atg acc caa act cca etc tec ctg cct gtc agt ctt gga Asp Val Val Met Thr Gin Thr pro Leu ser Leu pro Val Ser Leu Gly gat cca gec tec ate tct tgc aga tct agt cag age ata gta cat agt Asp Pro Ala ser lie Ser cys Arg ser ser Gin ser lie val His Ser 20 25 30 aat gta aac acc tat tta gaa tgg tac ctg cag aaa cca ggc cag tct Asn val Asn Thr туг Leu Glu Тгр Tyr Leu Gin Lys Pro Gly Gin Ser 35 40 45 cca agg etc ctg ate tac aaa gtt tec aac cga ttt tct ggg gtc cca pro Arg Leu Leu Пе туг Lys val ser Asn Arg phe ser Gly val Pro 50 55 60 gac agg ttc agt ggc agt gga gca ggg аса gat ttc аса etc agg ate Asp Arg Phe ser Gly ser Gly Ala Gly Thr Asp Phe Thr Leu Arg lie age aga gtg gag get gag gat ctg gga att tat tac tgc ttt caa gat Ser Arg val Glu Ala Glu Asp Leu Gly He туг туг cys Phe Gin Gly tea cat gtt cct ccg aCg ttc ggt gga ggc acc aaa ctg gaa ate aaa ser His val Pro pro Thr phe Gly Gly Gly Thr Lys Leu Glu He Lys 100 105 110 cgt Arg <210> 90 <211> 113 <212> PRT <213> Artificial sequence <220> <223> Synthetic construct <400> 90 Asp val val Met Thr Gin Thr pro Leu ser Leu pro val Ser Leu Gly - 59 - 009807 - 60 - 009807 - 61 - 009807 - 62 - 009807 - 63 - 009807 <210> 98 <211> 113 <212> PRT <213> Artificial Sequence <220> <223> synthetic construct <4Q0> 98 Asp Val Val Met Thr Gin Thr Pro Leu Ser Leu Pro val Ser Leu Gly 15 10 15 Asp Pro Ala Ser lie Ser cys Arg ser ser Gin ser He val His ser 20 25 30 Asn val Asn Thr туг Leu Glu Trp Tyr Leu Gin Lys Pro Gly Gin ser 35 40 45 pro Lys Leu Leu Пе Tyr Lys val Ser Asn Arg Phe ser Gly val Pro 50 55 60 Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu Arg lie 65 70 75 80 ser Arg val Glu Ala Glu Asp Leu Gly lie Tyr Tyr cys Phe Gin Gly 85 90 95 Ser His val pro pro Thr Phe Gly Gly Gly Thr Lys Leu Glu lie Lys 100 105 110 Arg Клетки, экспрессирующие IGF-I-рецептор Y1251F Клетки, экспрессирующие рецептор инсулина Только козье антитело против мышиного IgG-rTTC Только козье антитело против мышиного IgG-FTTC t0° "Г flT 1 <Г 10 Я.1-Н О J 10° То1 Те^ То3 "г 100 нМ оЕМ164 5 J |_J llWIll'il |1ИВ|' I 1JWI|-I HJM 10° "Г 10* 1(Г 10 FL14H 100 нМ ОЕМ164 101 10* 10Г 10ч FL1-H 100 нМ аШ7 FL1-H 100 нМ aLR IIID 101 10* 103 ID4 FL1-H Фиг. 1 - 64 - 009807 Кривая титрования для связывания антитела ЕМ 164 с биотинилированным IGF-1-рецептором Е X ё 1 Ё О 10"' т 10 10'11 Ю-10 10" Концентрация, М Фиг. 2 Ингибирование связывания биотин-IGF-I с клетками MCF-7 антителом ЕМ 164 + биотин-IGF-I Фиг. 3 - 65 - 009807 Ингибирование IGF-I-стимулированното автоёюсфорилирования IGF-I-рецептора в клетках MCF-7 антителом ЕМ164 + IGF-I Фиг. 4 Ингибирование IGF-1-стимулированного фосфорилирования IRS-1 в клетках MCF-7 антителом ЕМ 164 0.7 -г 0.6 - 0.5 - 0.4 - 0.3 - 0.2 - 0.1 - 0.0 - IGF-I Фиг. 5 + IGF-I - 66 - 009807 Ингибирование IGF-I-стимулированной трансдукции сигнала в клетках SaOS-2 антителом ЕМ164 -IGF-I +IGF-I (50 нг/мл ) ЕМ164 (мкг/мл) Фиг. 6 Блот фосфо-Akt (S473) фосфо-Erk Весь Erk (повторный блот после блота на (Jxxxbo-Erk) Действие антител на рост и выживание клеток MCF-7 з бессывороточных условиях, +IGF-I, или в условиях +1,25% сыворотки 1.2 IIой ОТ 10 0.6" II 0.4 0.2 0.0 ЕИЗ без антител ШШ антитело ЕМ164 антитело 1Н7 антитело IR3 анти-В4-антитело -IGF-I +IGF-1 +1.25% сыворотка Фиг. 7 Действие антитела ЕМ 164 на рост и выживание клеток MCF-7 в 0,04 -10% сыворотке Ё О • без антител ¦ + 10 мкг/мл антитела ЕМ 164 Фиг. 8 - 67 - 009807 ie О Ингибирование стимулированного IGF-и сывороткой роста и выживания клеток NCI-H838 IGF-I + IGF-I Фиг. 9 + сыворотка Действие обработки антителом ЕМ 164 или таксолом или комбинацией антитела ЕМ164 и таксола на рост ксенотрансплантата рака легкого Calu-б в мышах ЗФР ЕМ16 Таксол ЕМ164 + 10 15 Таксол tftt t ft ft t *фуппа, умерщвленная вследствие быстрого роста опухоли 1 дни Фиг. 10 - 68 - 009807 Конкуренция связывания гуманизированного антитела ЕМ164 (версии 1.0) и мышиного антитела ЕМ164 (диапазон 1,06-10,6 молярной концентрации) с иммобилизованным биотинилированным IGF-1-рецептором 0 2 4 6 8 10 12 Отношение ([мышиное АЬ]/[гуманизированное Ab]) Фиг. 11 Легкая цепь мышиного ЕМ 164 1 atgaagttgcctgttaggctcjttggtgctgatgttctggattcct 1MKLPVRLLVLMFWIP 4G gcttccagtagtgatgttttgatgacccaaactccactctGcctg 16 AS SSDVbMTQ T B LSIi J-VK 91 cctgtcagtcttggagatcaagcctccatctcfctgcagatcfcagt 31PVSLGDQASISC R S_S CDR1 136 cagagcattgtacatagt aatgtaaacacctat 11 agaatggt ac 46 QS IVHSNVtTTYbE W Y CDR1 181 ctgcagaaaccaggccagtctccaaagctcctgatctaeaaagtfc 61LQKPGQS P К L L I Y К_V CDR2 22 6 tecaaccgattttctggggtcccagacaggttcagtggcagtgga 76 S N R F S GVPDRFSGSG CDR2 271 fccagggacagatttcacactcaggatcagcagagtggaggctgag 91SGTDFTLRISRVEAK 316 gatctgggaatttattactgctttcaaggttcacatgttcctccg 106 D L G I Y Y С F Q G 5 H V P_P CDR3 361 acgttcggtggaggcaccaagctggaaatcaaacgg 121 TFGGGTKLEI KR Фиг. 12 - 69 - 009807 Тяжелая цепь мышиного ЕМ 164 1 atgggatggagctatatcatcctctttttggtagcaacagctaca 1 MGWS Y I 1 L F Ь V А Т ~А Т 46 gaagtccactcccaggtccaactgcagcagtctggggctgaactg 16EVHSQVQLQQSGA"EL J-VH 91 gtgaagcctggggcttcagtgaagctgtcctgtaaggcttctggc 31VKPGASVKLSCKASG 136 tacaccttcaccagctactggatgcactgggtgaagcagaggcct 46 Y T F T SYWMH W V К Q R P CDR1 181 ggacaaggccfctgagtggattggagagattaatcctagcaacggt 61GQGLEWIG В I N P S N G CDR2 226 cgtactaactacaatgagaagttcaagaggaaggccacactgact 76 RTNYNBKFKR К A T h T CDR2 271 gtagacaaatcctccagcacagcctacatgcaactcagcagcctg 91.VDKS SSTAYMQLSSL 316 acatctgaggactctgcggtctattactttgcaagaggaagacca 106 TSEDSAVYYFAR G_Jt_p CDR3 361 gattactacggtagtagcaagtggtacttcgatgtctggggcgca 121 DYYGSSKWYFDV W G A CDR3 406 gggaccacggtcaccgtctcctca 136 GTTVTVSS Фиг. 13 - 70 - 009807 CDR мышиного EM164 Легкая цепь CDR1: RSSQSIVHSNVNTYLE CDR2: KVSNRFS CDR3: FQGSHVPPT Тяжелая цепь CDR1: SYWMH CDR2: EINPSNGRTNYNEKFKR CDR3: GRPDYYGSSKWYFDV Тяжелая цепь АЬМ CDR1: GYTFTSYWMH CDR2: EINPSNGRTN CDR3: GRPDYYGSSKWYFDV Фиг. 14 Сравнения с последовательностями зародышевой линии Легкая цепь | 50~ muEM164 .......-............--------------v---------------- 100 Crl - bblYK^SmtFSGVTORFSasaSQTDFTI^ISRVK^I^VryCFOGaire шиЕМ164 - ---------------......-------R----------X......---- Тяжелая цепь| 50 J55 8.C " QVQbQQPeAELVKPGS^VKLRCKASGTTPTSTOMHWVKQTLPGCGbTJWIGE muKMl64-------s-........-............-......-------------- J558 . с muEM164 njPSHGRTHYHEKFKS К ATbTVDKS SSTAYHQLS S PTS ЕП S AVY У CAE ---------------R-------------------L---------F-- Фиг. 15 - 71 - 009807 Клонирование и карты экспрессионных плазмид млекопитающих Фиг. 16 - 72 - 009807 Выравнивание 10 наиболее гомологичных последовательностей легкой цепи 1 so ЕМ164 DVIJtTQTPLS LPVSLGDQAS TSCRSSQSIV HSHVNTYLEW YLQKPGQSPK 2jel DVLMTQTPLS LPVSLGDQAS ISCESSQSIV HSHGHTYLEW YLQKPGQSPK 2pcp DVTJMTQTPLS LPVSLGBQAS ISCRSSQTIV HSHGHTYLEW YbQKPGQSPK Inqb DIELTQTPLS LPVSLODQAS ISCRSSQSTV HSHGHTYLEW YLQKPGQSPK lkel DVLMTQTPLS LPVSLGDQAS ISCRF3QSIV HSUGNTYLEW YLQKSGQSPK lfayx ELVMTQTPLS LPVSLGDQAS ISCRSSQTIV HSBODTYLDW FLQKPGQiJPK ligf DVLMTQTPLS LPVSLGDQAS ISCRSNQTIL LSDGDTYLEW YLQKPGQSPK ltet DVLMTQTPLS LPVSLGDQAS ISCKSSQSXV HSSGBTYFEW YLQKPGQSPK lclz DVLMTQIPVS LPVSLGDQAS ISCRSSQIIV HRWGNTYLEW YLQKPGQSPCJ lbln DVLMTQTPVS LSVSLGDQA5 XSCRSSQSXV HSTGNTYLKW YLQKPGQSPK lely DVLMTQIPVS LPVSLGDQAS ISCRSSQIIV HNWGNTYLHW YLQKPGQ3PQ Консенсус dVlraTQtPIS LpVSLGDQAS ISCrsaQxIv hengnTYleW yLQKpGQSPk EK164 LLIYKVSHRF SGVPDRFSGS 2jel LLIYSISSRF SGVPDRFSGS 2pcp LLIYKVTKRF SGVPDRFSGS Inqb LLIYKVSHRF SGVPDRFSGS lkel LLIYKVSHRF SGVPDRFSGS lhyx LLIYKVSBRF SGVPDRFSGS Ugf LLIYKVSHRF SGVPDRFSGS ltet LLIYKVSHRF SGVPDRFSGS lola LLIYKVSHRF SGVPDRFSGS lbln LLIYKISHRF SGVPDRFSGS lcly LLIYKVSHRF SGVPDRFSGS KOHCeHCyC LLIYkvsnRF SGVPDRFSGS 100 GSGTDFTLRI SRVEAEDLGI YYCFQGSHVP GSGTDFTLKI SRVQAEDLGV YYCFQQSHVP GSGTDFTLKI SRVEAEDLGV YYCFQGTHAP GSGTDFTLKI SRVEAEDLGV YYCFQQSHVP GSGTDFTLKI SRVEAEDLGV YYCFQQSHVP GSGTDFTLKI SRVEAEDLGV YYCFQQSHVP GSGTDFTLKI SRVEAEDLGV YYCFQGSHVP GSGTDFTLKI SRVEAEDLGV YYCFQSSHIP GSGTDFTLKI SRVEAEDLGV YYCFQGSHVP GSGTDFTLKI SRVEAEDLGV YYCFQASHAP GSGTDFTLKI SRVEAEDLGV YYCFQGSHVP GSGTDFTLkl SRVeAEDLGv YYCPQgeHvp 101 113 EMI 64 PTFGGGTKLE IKR 2jel YTFGGGTKLE IKR 2pcp YTFGGGTKLE IKR lnqb YTFGGGTKLE IKR lkel RTFGGGTKLE IKR lhyx PTFGGGTKLE IKR ligf PTFGGGTKLE IKR ltet FTFGSGTKUE IKR lclz FTFGSOTKLE IKR lbln RTFGGGTKLE IKR lely FTFGSGTKLS IKK Консенсус xTFGgGTKLE IKR Фиг. 17 - 73 - 009807 Выравнивание 10 наиболее гомологичных последовательностей тяжелой цепи 1 50 EM1S4 QVQLQQSGAE LVKPGASVKL SCKASGYTFT SYWMHWVKQR PGQGLEWIGE lnqb QVQLQQSGAE LVKPGASVKL SCKASGYTFT SYWMHWVKQR PGRGLEWIGR lngp QVQLQQPGAE LVKPGASVKL SCKASGYTFT SYWMHWVKQR PGRGLEWIGR Ifbi QVQbQQPGAE LVKPGASVKL SCKASGYTFT SYWMHWVKOQ PGQGLEWIGE lafv QVQLQQPGSV LVKPGASVKL SCKASGYTFT SSHIHWAKQR PGQQLEW1G5 lyuh QVQFQQSGAB LVKPGASVKL SCKASGYTFT SYLMHWIKQR PGRGLEWIGR lplg QIQLQQSGPE LVRPGASVKT SCKASGYTFT DYYIHWVKQR PGEGLBWIGW IdSb QVQLQQSGAE LMKPGASVKT SCKATGYTFS SFWIEHVKQR PGBGLEWIGB lae6 QIQLQQSGPB LVKPGASVKI SCKASGYTFT DYYXBXMKQK PGQGLEWIGW laxs QVQLLESGAE LHKPGASVKI SCKATGYTFS SFWIEWVKQR PGHGLBWJGE 3UC1 .VQLQQSGJVE LHKPGASVKI SCKASGYTFS DYWIEWVKQR PGHGLEW1GE Консенсус qvQlqqsGae LvkPGASVKx SCKAsGYTFt sywxhWvKQr PGxGLEWlGx 51 100 EM1S4 INPSNGRTNY NEKPKRKATL TVDKSSSTAY MQLSSLTSED SAvYYFARQR lnqb IDPNSGGTKY HEKPKSKATL TVDKPSSTAY MQLSSLTSED SAVYYCAR. - lngp IDPNSGGTKY HEKFKSKATL TVDKPSSTAY MQLSSLTSED SAVYYCAR. , lfbi IDPSDSYPHY NEKFKGKATL TVDKSSSTAY MQLSSLTSED SAVYYCAS.. lafv IHPKSGSTHY NEKFKGKATL TVDTSSSTAY VDbSSLTSHD SAVYYCAR. . lyuh IDPKiTWTKF HEKFKSKATL TVDKPSSTAY MBLSSLTSED SAVYYCAR.. lplg IYPGSGSTKY HEKFKGKATL TVDTSSSTAY MQLSSLTSED SAVYFCAH. . IdSb IIiPGSGGTHY NBKFKGKATF TADKSSNTAY MQLSSLTSED SAVYYCARGH 1авб IDPGSGNTKY HEKFKGKATL TVDTSSSTAY MQLSSLTSED TAVYFCAa.. laxs IIiPGSGGTHY HBKFKGXATF TADKSSHTAY MQLSSLTSED SAVYYCARGH 3hfl ILPGSGSTHY HERFKGKATF TADTSSSTAY MQLHSLTSED SGVYYCLHGN Консенсус TxPxsgxtxy nEkFKgKATl TvDksSsTAY mqLsSLTSED saVYycar-- 101 124 EM164 PDYYGSSKHY FDVWGAGTTV TVSS lnqb YTJYYGSS. . Y FDYW3QGTTV TVSS Ungp YDYYGSS. . Y FDYWGQGTTL TVSS Ifbi LYYYGTSYGV LDYWGQGTSV TVSS lafv .WRYGSP.YY FDYWGQGTTL TVSS lyuh YAYCKP----MDYWGQGTTV TVSS lplg . .GGK. . .FA KDYWGOGTSV TVSS ld5b S.YYF.. .YD GDYWGQGT5V TVSS lae6 . .EKTTYYYA MDYWGQGTSV TVSA laxs S.YYF. .-YD GDYWGOGTSV TVSS 3bfl ........YD FTJGWGQGTTL TVSS Консенсус xxyyxxx-xx xOyWGqGTxv TVSe Фиг. 18 - 74 - 009807 Доступность легкой цепи ЕМ 164 т- <Ч И Р> Номер по нумерации Kabat Доступность тяжелой цепи ЕМ164 Номер по нумерации Kabat Фиг. 19 Аминокислотные последовательности вариабельной области леской цепи для мышиного и гуманизированного антител ЕМ164 Kabat # muEM164 huEM164 VI, huEM164 vl. huEM164 vl, huEM164 vl, замены 1 10 20 27C 35 45 DVbMTQTPLS LPVSLGDQAS ISCRSSQSIV HSKVHTXLEW YLQKPGQSPK 0 DWMTQTPLS LPVSLGDPAS ISCRSSQSIV HSNVNTYLEW YLQKPGQSPR 1 DVLMTQTPLS LPVSLGDPAS ISCRSSQSIV HSNVBTYLEW YLQKPQQSPK 2 DVLMTQTPLS LPVSLGDPAS ISCRSSQSIV HSKVHTYLEW YLQKPGQSPR 3 DWMTQTPLS LPVSLGDPAS ISCRSSQSIV HSNVNTYLEW YLQKPGQSPK Kabat # muEM164 huEM164 vl.O huEM164 vl.l huEM164 vl-2 huEM164 vl.3 замены 46 55 LLIYKVSHRF LLFHCVSNRF LLIYKVSHRF LLIYKVSHRF LLIYKVSHRF 65 SGVPDRFSGS SGVPDRFSGS SGVPDRFSGS SGVPDRFSGS SGVPDRFSGS 75 3SGTDFTLRI GAGTDFTLRI GAGTDFTLRI QAGTDFTLRI GAGTDFTLRI 85 SRVEAEDLGI SRVEAEDLGI SRVEAEDLGI SRVEAEDLGI SRVEAEDLGI 95 f YYCFQGSHVP YYCFQGSHVP YYCFQGSHVP YYCFQGSHVP YYCFQGSHVP Kabat # ЯШЕМ164 huEM164 vl.O huEM164 Vl.l huEM164 vl-2 huEM164 Vl.3 замены 96 105 108 PTFGGGTKLE IKR PTFGGGTKLE IKR PTFGGGTKLE IKR PTFGGGTKLE IKR PTFGGGTKLE IKR Фиг. 20 - 75 - 009807 Аминокислотные последовательности вариабельной области тяжелой цепи для мышиного и гуманизированного антител ЕМ164 Kabat # 1 10 20 30 40 50 ВШИ4164 QVQLQQSGAE LVKPGASVKL SCKASGYTFT SYWMHWVKQR PGQGLEHISB huEMlG4 QVQLVQSGAE WKPGASVKL SCKASGYTFT SYWMHWVKQR PGQGLEWIGB замены * * Kabat # 51 59 69 muEM164 IHPSHGRTKY HEKFKRKATL huHO.64 rNPSNGRTHY HQKFQGKATL замены * ** 79 86 96 TVDKSSSTAY MQLSSLTSED SAVYYFAKGIt TVDKSSSTAY MQLSSLTSED SAVYYFARGK Kabat # 97 lOOf 109 113 muEM164 PDYYGSSKWY FDVWGAGTTV TVSS huB"164 PDYYGSSKWY FDVWGKJQTTV TVSS замены 4 Фиг. 21 ДНК-последовательность и аминокислотная последовательность вариабельной области huEM164 vl.O Легкая цепь 1 gatgttgtgatgacccaaactccactctcccfcgcctgtcagtctt 1DVVMTQTPLSLPVSL 4S ggagatccagcctccatctcttgcagatctagtcagagcatagta 16GDPASIS CRSSQSIV 91 catagtaatgtaaacacctatttagaatggtaectgcagaaacca 31 HSMVNTY LEWYLQKP 136 ggccagtctccaaggctcctgatctacaaagtttccaaccgattt 46GQSPRLL I YKVSN RF 181 tctggggtcccagacaggttcagtggcagtggagcagggacagat 61SGVPDRFSGSGAGTD 226 ttcacactcaggatcagcagagtggaggctgaggafcctgggaatt 76FTLRISRVEAEDLGI 271 eat tactgct tt caaggt tcacatgtt cctccgacgt fccggtgga 91YYCFQGSHVPPTFGG 316 ggcaccaaactggaaatcaaacgt 106 GTKLEIKR Тяжелая цепь 1 caggtccaactggtgcagtctggggctgaagtggtgaagcctggg 1QVQLVQSGABVVKPG 46 gcttcagtgaagctgtcctgtaaggcttctggctacaccttcacc 16ASVKLSCKASGYTFT 91 agctactggatgcactgggtgaagcagaggcctggacaaggcctt 31 SYWMHWVKQRPGQGL 136 gagtggattggagagattaatcctagcaacggtcgtactsactac 46BWIGEINPSKGRTNY 181 aatcagaagttccaggggaaggccacactgactgtagacaaatcc 61NQKFQGKATLTVDKS 226 tccagcacagcctacatgcaactcageagectgacatctgaggae 76SSTAYMQLSSLTSED 271 tctgcggtctattactttgcaagaggaagaccagattactacggt 91 SAVYYFARGRPDYYG 316 agtagcaagtggtaefctcgatgtctggggccaagggaccacggtc 106 SSKWYFDVWGQGTTV 361 accgtctcctca 121 TVSS Фиг. 22 - 76 - 009807 ДНК-последовательность и аминокислотная последовательность вариабельной области легкой цепи huEM164 vl.l, 1.2,1.3 vl.l 1 gatgttttgatgacccaaactccactctccctgcctgtcagtctt 1DVLMTQ.TP1iSE.PVSL 4 6 ggagatceagectccatetct tgcagatctagtcagagcatag ta 16GDPASISCRSSQSIV 91 catagtaafcgtaaacacctatttagaatggtacctgcagaaacca 31HSJSrVHTYLEWYIiQKP 13 6 ggccagtetecaaagctccfcgatcfcacaaagtttccaaccgattt 46GQSPKLL IYKVSNRF 181 tetggggtcccagacaggttcagtggcagtggagcagggacagat 61SGVPDRFSGSGAGTD 226 ttcacactcaggat cagcagagtggaggctgaggatctgggaatt 76FTLRISRVEAEDLGI 271 tattactgcfcttcaaggttcacatgttcotccgacgtteggtgga 91YYCFQGSHVPPTFGG 316 ggcaccaaactggaaatcaaacgt 106 GTKLE1KR Vl.2 1 gatgttttgatgacccaaactccactctccctgcctgtcagtctt IDVItMTQTPbSIiP-VSL 46 ggagat ce agcct с cat сtctt gcagatctagtcagagcatagta 16GDPASISCRSSQSIV 91 catagtaatgtaaacacctatttagaatggtacctgcagaaacca 31HSHVNTYLBWYLQKJP 136 ggccagtetceaaggetcctgatetacaaagtttccaaccgattt 46GQSPRLLIYKVSNKF 181 tctggggtcecagacaggttcagtggcagtggagcagggacagat 61SGVPDRFSGSGAGTD 226 ttcacactcaggatcagcagagtggaggctgaggatetgggaatt 76FTbR2SRVBAEDLGI 271 tattactgctttcaaggttcacatgttcctccgacgttcggtgga 91YYCFQGSHVPFTFGQ 316 ggcaccaaactggaaatcaaacgt 106. GTKLEIKR - 77 - 009807 vl.3 1 gatgttgtgatgacccaaactccactctccctgcctgfccagtetfc IDVVMTQTPLSLPVSb 46 ggagatccagcctceatctcttgcagatctagtcagagcatagfca 16GDPA3 ISCRSS QSI V 91 catagtaatgtaaacacctatttagaatggtacctgcagaaacca 31HSMVHTYI> EWYLQKP 13e ggccagtctccaaagctectgatctacaaagtttccaaccgattt 46GQSPKLL IYKVSNRF 181 tctggggtcccagacaggttcagtggcagtggagcagggacagat 61SGVPDRFSGSGAGTD 226 ttcacactcaggatcagcagagtggaggctgaggatctgggaatt 76FTI.RISRVEAEDLG I 271 tattactgctctcaaggttcaeatgctcctccgacgttcggtgga 91YYCFQGS HVPPT FGO 316 ggcaccaaactggaaatcaaacgt 106 GTKLEIKR Фиг. 23 Сравнение ингибирования IGF-I-стимулированного роста и выживания клеток MCF-7 гуманизированным антителом ЕМ164 vl.O (625 мкг/мл) в сравнении с мышиным антителом ЕМ164 (5-10 мкг/мл) - IGF-i + IGF-! Фиг. 24 - 78 - 009807 Задержка клеточного цикла клеток MCF-7 антителом ЕМ 164 Клетки MCF-7 | G2/M 100 2 о и. О r-t ¦ S ? GQ/GI 1-1 Фиг. 25 Индукция апоптоза в клетках рака легкого NCI-H838 антителом ЕМ164 (измеренная на основании расщепления белка СК18 цитокератина каспазой) NCI-H838 Cells без антитела Антитело ЕМ164 Фиг. 26 - 79 - 009807 Подавление роста ксенотрансплантатов рака поджелудочной железы человека ВхРС-3 в мышах обработкой антителом ЕМ164 в качестве единственного агента или в комбинации с гемцитабином ВхРС-3 -о- Гемцитабин -°- Контрольное антитело -о-ЗФР -•- Антитело ЕМ164 -*~ ЕМ164 + Гемцитабин 10 20 30 40 50 60 70 Время (дни) t t Гемцитабин ttt ttt t t ttttt Антитело EM164 Фиг. 27 Евразийская патентная организация, ЕАПВ Россия, 109012, Москва, Малый Черкасский пер., 2/6 - 80 - |