|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000008252b*\id |
|
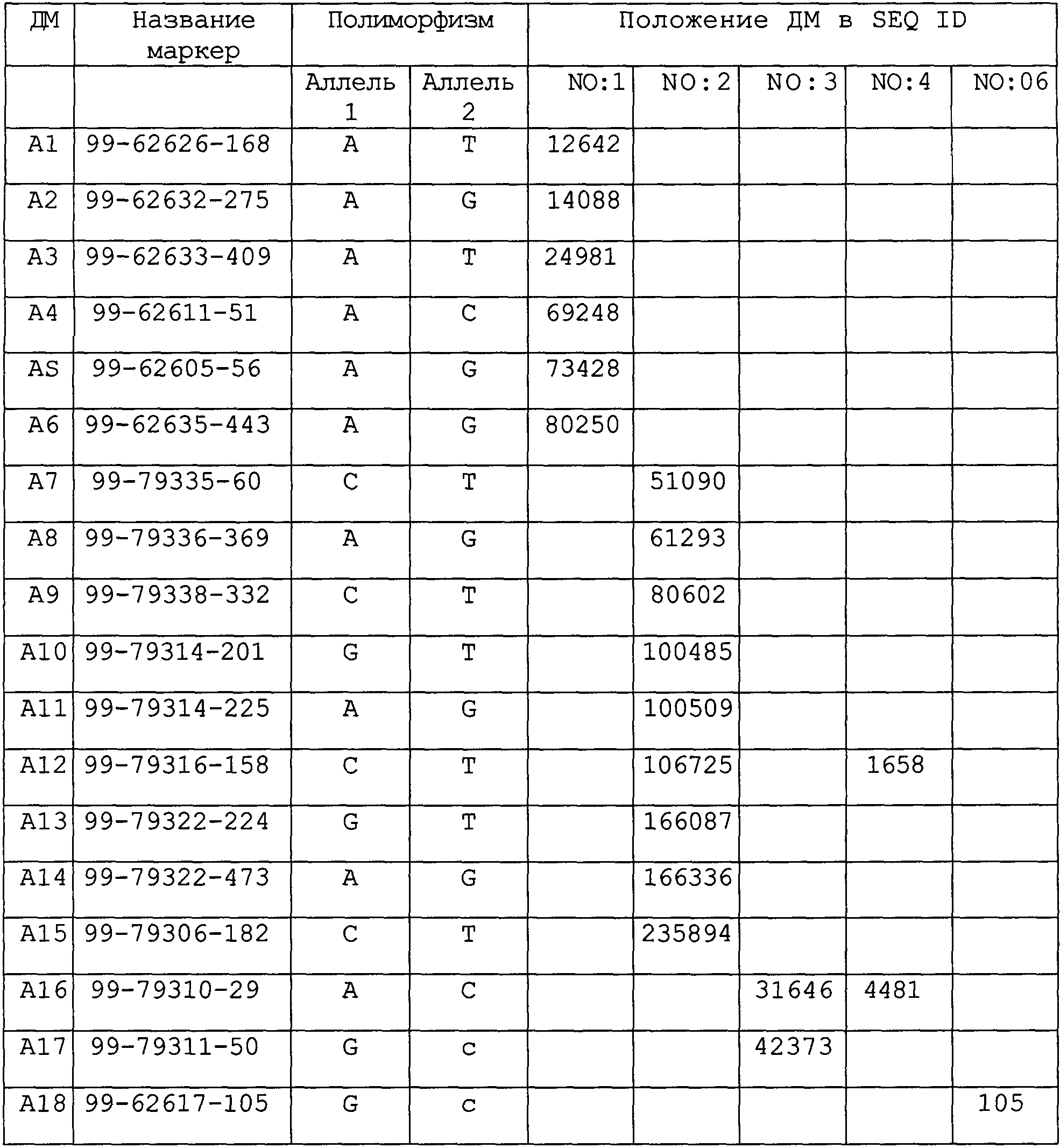
Термины запроса в документе Реферат 1. Антитело или антигенсвязывающий домен, специфически связывающийся с полипептидом с последовательностью SEQ ID NO: 5. 2. Антитело или антигенсвязывающий домен по п.1, который является химерным, гуманизированным антителом, или антигенсвязывающим доменом, или моноклональным или поликлональным антителом, или антигенсвязывающим доменом человека. 3. Антитело или антигенсвязывающий домен по п.1 или 2, которое является антителом или антигенсвязывающим доменом с одной цепью. 4. Антитело или антигенсвязывающий домен по любому из пп.1-3, который является моноспецифичным, биспецифичным, триспецифичным или мультиспецифичным антителом или антигенсвязывающим доменом. 5. Применение антитела или антигенсвязывающего домена по любому из пп.1-3 для диагностики шизофрении или биполярного расстройства. 6. Набор антисмысловых последовательностей, ингибирующих экспрессию полипептида ионного канала, кодируемого полинуклеотидом, показанным в виде SEQ ID NO: 1, 2, 3 или 4, содержащий фрагмент полинуклеотида с последовательностью SEQ ID NO: 1, 2, 3 или 4, для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 7. Набор антисмысловых последовательностей по п.6, которые являются рибозимами. 8. Применение антитела или антигенсвязывающего фрагмента по пп.1-4 для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 9. Применение набора антисмысловых последовательностей по п.6 или 7 для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 10. Фармацевтическая композиция, содержащая антитело или антигенсвязывающий фрагмент по пп.1-4. 11. Фармацевтическая композиция, содержащая набор антисмысловых последовательностей по п.6 или 7. 12. Применение по крайней мере одного двухаллельного маркера (ДМ), связанного с ионным каналом, выбранного из группы двуаллельных маркеров, показанных ниже, для определения подвержен ли индивидуум риску или страдает ли больной шизофренией или биполярным расстройством. 13. Способ генотипирования, включающий стадии: a) выделения нуклеиновой кислоты из биологического образца; b) обнаружения наличия нуклеотида в одном или нескольких зависимых двухаллельных маркерах, связанных с ионным каналом, А1-А18, перечисленных в п.12. 14. Способ по п.13, где способ осуществляют с использованием анализа микросеквенирования. 15. Диагностический набор для диагностики шизофрении или биполярного расстройства, содержащий полинуклеотид, который является фрагментом полинуклеотида с последовательностью SEQ ID NO: 1, 2, 3 или 4, содержащего один или несколько биаллельных маркеров А1-А18, как описано в п.12. 16. Диагностический набор для диагностики шизофрении или биполярного расстройства, содержащий: a) поликлональное или моноклональное антитело, которое специфически связывается с полипептидом ионного канала, содержащим аминокислотную последовательность SEQ ID No 5, или с фрагментом пептида или его вариантом, необязательно меченным; b) реагент, с помощью которого осуществляется определение образовавшегося комплекса антиген-антитела, указанный реагент либо необязательно содержит метку, либо который может быть распознан меченным реагентом.
Полный текст патента (57) Реферат / Формула: 2. Антитело или антигенсвязывающий домен по п.1, который является химерным, гуманизированным антителом, или антигенсвязывающим доменом, или моноклональным или поликлональным антителом, или антигенсвязывающим доменом человека. 3. Антитело или антигенсвязывающий домен по п.1 или 2, которое является антителом или антигенсвязывающим доменом с одной цепью. 4. Антитело или антигенсвязывающий домен по любому из пп.1-3, который является моноспецифичным, биспецифичным, триспецифичным или мультиспецифичным антителом или антигенсвязывающим доменом. 5. Применение антитела или антигенсвязывающего домена по любому из пп.1-3 для диагностики шизофрении или биполярного расстройства. 6. Набор антисмысловых последовательностей, ингибирующих экспрессию полипептида ионного канала, кодируемого полинуклеотидом, показанным в виде SEQ ID NO: 1, 2, 3 или 4, содержащий фрагмент полинуклеотида с последовательностью SEQ ID NO: 1, 2, 3 или 4, для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 7. Набор антисмысловых последовательностей по п.6, которые являются рибозимами. 8. Применение антитела или антигенсвязывающего фрагмента по пп.1-4 для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 9. Применение набора антисмысловых последовательностей по п.6 или 7 для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 10. Фармацевтическая композиция, содержащая антитело или антигенсвязывающий фрагмент по пп.1-4. 11. Фармацевтическая композиция, содержащая набор антисмысловых последовательностей по п.6 или 7. 12. Применение по крайней мере одного двухаллельного маркера (ДМ), связанного с ионным каналом, выбранного из группы двуаллельных маркеров, показанных ниже, для определения подвержен ли индивидуум риску или страдает ли больной шизофренией или биполярным расстройством. 13. Способ генотипирования, включающий стадии: a) выделения нуклеиновой кислоты из биологического образца; b) обнаружения наличия нуклеотида в одном или нескольких зависимых двухаллельных маркерах, связанных с ионным каналом, А1-А18, перечисленных в п.12. 14. Способ по п.13, где способ осуществляют с использованием анализа микросеквенирования. 15. Диагностический набор для диагностики шизофрении или биполярного расстройства, содержащий полинуклеотид, который является фрагментом полинуклеотида с последовательностью SEQ ID NO: 1, 2, 3 или 4, содержащего один или несколько биаллельных маркеров А1-А18, как описано в п.12. 16. Диагностический набор для диагностики шизофрении или биполярного расстройства, содержащий: a) поликлональное или моноклональное антитело, которое специфически связывается с полипептидом ионного канала, содержащим аминокислотную последовательность SEQ ID No 5, или с фрагментом пептида или его вариантом, необязательно меченным; b) реагент, с помощью которого осуществляется определение образовавшегося комплекса антиген-антитела, указанный реагент либо необязательно содержит метку, либо который может быть распознан меченным реагентом.
008252 Область техники, к которой относится изобретение Настоящее изобретение относится к антителу или антигенсвязывающему домену, специфически связывающимся с полипептидом потенциалзависимого воротного ионного канала, к их применению для диагностики шизофрении или биполярного расстройства, к набору антисмысловых последовательностей, ингибирующих экспрессию ионного канала, к соединению, которое активирует или ингибирует экспрессию или активность ионного канала указанного полипептида, к фармацевтической композиции, содержащей антитело или антигенсвязывающий фрагмент, набор антисмысловых последовательностей или такое соединение и к диагностическому набору для диагностики шизофрении или биполярного расстройства. Уровень техники Достижения в техническом оснащении лабораторий, занятых фундаментальными и клиническими исследованиями, в огромной мере способствовали более глубокому изучению влияния головного мозга и нервной системы на здоровье и болезни. Выдвинут ряд нейробиологических и фармакологических гипотез в отношении нейрохимических и генетических механизмов, имеющих место при нарушениях центральной нервной системы (ЦНС), включающих психические нарушения и нейродегенеративные заболевания. Однако нарушения ЦНС характеризуются сложным и недостаточным пониманием этиологии, а также симптомы, которые перекрываются, плохо описаны, и трудны для измерения. В итоге, будущие схемы лечения и усилия по созданию лекарств нуждаются в усовершенствовании и сосредоточении внимания на мультигенных причинах, а это требует проведения новых демографических исследований заболеваний и получения более точной диагностической и прогностической информации о пациентах, страдающих нарушениями ЦНС. Нарушения ЦНС охватывают широкий ряд расстройств и соответствующую широкую группу генетических факторов. Примеры расстройств ЦНС включают нейродегенеративные нарушения, психотические расстройства, расстройства в эмоциональном состоянии, аутизм, зависимость от химических веществ и алкоголизм, нарушения болевых ощущений, эпилепсию, задержку психического развития и другие психические заболевания, включая когнитивную функцию, страх, нарушение функции приема пищи, нарушение функции мотивации и аномалии характера. Нарушения можно определить, используя классификацию Diagnosis and Statistical Manual of Mental Disorders fourth edition (Диагностический и Статистический Справочник Психических Нарушений) (DSM-IV). Даже при рассмотрении всего лишь небольшой подгруппы нарушений ЦНС очевидно, что из-за недостатка адекватного лечения и для понимания молекулярной основы нарушений при шизофрении и биполярном расстройстве, нужны новые цели для создания лекарств и усовершенствования способов лечения. Что касается шизофрении и биполярного расстройства, все ныне известные молекулы, используемые для их лечения, обладают побочными эффектами и действуют только против симптомов данного заболевания. Существует, таким образом, очень большая потребность в новых молекулах, не вызывающих побочных эффектов и направленных против мишеней, которые вовлечены в причинные механизмы шизофрении и биполярного расстройства. Поэтому, инструментальные средства, облегчающие открытие и характеристику данных целей, необходимы и полезны. Потенциалзависимые воротные ионные каналы Потенциалзависимые воротные ионные каналы являются частью большого семейства макромолекул, функции которых включают контроль и поддержание электрического потенциала клеточных мембран, выделение и трансдукцию сигнала. Эти белки-каналы участвуют в контроле высвобождения ней-ромедиаторов из нейронов и играют существенную роль в регуляции разнообразных клеточных функций, включая мембранную возбудимость, мышечное сокращение и синаптическую передачу. Основные альфа-субъединицы №+-каналов и альфа-1-субъединицы Са+-каналов состоят, приблизительно, из 2000 аминокислот и составляют путь протекания ионов. Биохимический анализ показал, что физиологически активный ионный канал составлен из нескольких разных субъединиц. Существует две вспомогательные субъединицы, бета-1- и бета-2-субъединица, которые очищаются совместно с альфа-субъединицей N+-каналов. Что касается Са+-каналов, для них идентифицированы дополнительные субъединицы (альфа-2, бета, гамма и сигма) с модуляторным действием. Субъединицы альфа-2 и бета, похоже, усиливают функциональную активность альфа-1-субъединицы Са+-каналов. Альфа-субъединицы К+-каналов ассоциируются с бета-субъединицами в соотношении 1:1, что приводит к усложнению К+-каналов, проявляющих (альфа)4(бета)4 стехиометрию (Terlau и соавт., Naturwissenschaften 85:437-444 (1998)). Базовая структура и примеры кальциевых и натриевых ионных каналов рассматриваются, кроме того, например, у Williams, и соавт., Science 257:389-395 (1992); Mori, и соавт., Nature 350:398-402 (1991); и Koch, и соавт., J. Biol. Chem. 265 (29):17786-17791 (1990). Нуклеотидная последовательность Са+- и №+-ионного канала крысы обладает высоким уровнем гомологии с последовательностью CanIon-канала, впоследствии описанного у Lee и соавт., FEBS Lett. 445:231-236 (1999) . Альфа-субъединица обладает последовательностью, характерной для всех потенциалзависимых ка-тионных каналов, и использует один и тот же структурный мотив, включающий пучок из 6 спиралей в доменах, предположительно пронизывающих мембрану. И в натриевых, и в кальциевых каналах данный мотив повторяется в указанной последовательности 4 раза, образуя пучок из 24 спиралей. Данные аминокислотные последовательности высококонсервативны у видов (например, человека и Drosophila), a также у разных ионных каналов. - 1 - 008252 Существует несколько тканеспецифичных фармакологически и электрофизиологически различимых изоформ кальциевых каналов, кодируемых отдельными генами мультигенного семейства. В скелетной мышце каждая группа из плотно связанных альфа-, бета- и гамма-субъединиц ассоциирована с 4 другими с образованием пятичленной макромолекулы. Например, альфа-1-субъединицы кальциевого канала нейрона являются продуктом, по меньшей мере, семи разных генов, именуемых от А до Н. В иммуноци-тохимических исследованиях показано разное распределение альфа-1-субъединиц кальциевого канала. Альфа-1А и альфа-1В экспрессируются главным образом в дендритах и пресинаптических окончаниях, и альфа-1А обычно концентрируется в большем числе нервных окончаний, в альфа-1В. В нервно-мышечном соединении крысы и человека альфа-1А локализуется пресинаптически, тогда как альфа-1В, так и альфа-1А представлены в ассоциированных с аксоном швановских клетках. Альфа-1Е локализуется главным образом в клеточных телах, в некоторых случаях - в проксимальных дендритах, а также в дис-тальных ответвлениях клеток Пуркинье. Альфа-1С и альфа-Ш локализуются в клеточных телах и в проксимальных дендритах центральных нейронов. Нативные кальциевые каналы классифицируются на основе их фармакологических и/или электрофизиологических свойств. Классификация потенциалзависимых кальциевых каналов разделяет их на каналы, активируемые высоким потенциалом (HVA), включающие типы L, N, Р и О; промежуточным (^АД-тип); и активируемые низким потенциалом (LVA, Т-тип). (Могеш и соавт., Annals NY Acad. Sci. 102-117 (1999)). Основные субъединицы (альфа-1) относятся к генному семейству, члены которого могут сами образовывать функциональные каналы при экспрессии в гетерологичных экспрессионных системах. (Zhang и соавт., Neuropharmacology 32 (11):1075-1088 (1993), включенный в данное описание путем ссылки). В нативных клетках альфа-1-субъединицы экспрессируются в виде мультисубъединичных комплексов вместе с вспомогательными субъединицами, которые модифицируют функциональные характеристики альфа-1-субъединицы. Во многих случаях коэкспрессия вспомогательных субъединиц влияет на биофизические свойства данных каналов. Бета-субъединицы, в частности, имеют тенденцию к существенному влиянию на альфа-1-субъединицы; было показано, что бета-субъединицы изменяют активационные свойства, стационарную инактивацию, кинетику инактивации и максимальное протекание тока. Большое молекулярное разнообразие каналов получают благодаря существованию множественных форм альфа-1-субъединиц. Например, было показано, что по-разному сплайсированные формы кальциевых каналов и экспрессируются по-разному и обладают разной чувствительностью к фосфорилированию серин-треониновыми киназами. (Hell и соавт., Annals NY Acad. Sci. 747:282-293 (1994)). Установлено, что мутации генов ионных каналов вовлечены в целый ряд заболеваний, включая некоторые заболевания центральной нервной системы. Примеры мутаций ионных каналов, вызывающих ряд эпизодических нарушений, включая периодический паралич, эпизодическую атаксию, гемикранию, синдром удлинения от интервала QT, пароксизмальную дискинезию, рассматриваются в обзоре Bulman и соавт., Hum. Mol. Gen. 6(10):1679-1685 (1997). Некоторые нарушения, связанные с мутацией Са+-канала, представлены, например, в табл. А (из Moreno, выше). Таблица А 1 Заболевание Субъединица кальциевого канала Модель Семейная гемиплегическая мигрень Альфа-1А Человек Эпизодическая атаксия, тип 2 Альфа-1А Человек Спинально-церебеллярная атаксия, тип 6 Альфа-1А Человек Фенотипы пьяного и беспомощного Альфа-1А Мышь Синдром Ламберт-Итона Альфа-1А, ex-IB Человек Гипокалиемический периодический паралич Альфа-lS Человек Мышиная дисплазия Альфа-lS Мышь Диабетические жировые фенотипы Цукер (Х-1С; а-Ю Крыса Летаргический фенотип Бета-4 Мышь Злокачественна гипотермия Альфа-2/5 Человек Сзерцание Гамма Мышь Модуляторы кальциевых и натриевых каналов обычно используются для лечения различных заболеваний и состояний. Например, было показано, что блокаторы кальциевого и/или натриевого канала могут использоваться для лечения или профилактики одного или нескольких симптомов, связанных с различными заболеваниями или состояниями, такими как различные заболевания и состояния сердца (например, стенокардия, аритмия), гипертензия, мигрени, неврологические эффекты нарушений мозгово - 2 - 008252 го кровообращения, мания, нейролептически индуцированная поздняя дискинезия стенок миокарда, биполярное расстройство, боль, эпилепсия и другие. Было показано, что между разными ионными каналами нервной системы имеются существенные функциональные различия. Кроме того, существенные различия имеются между разными мутациями в одном и том же гене ионного канала, а также между вариантами сплайсинга одного и того же ионного канала. Таким образом, несмотря на причастность ионных каналов к заболеваниям ЦНС, то, что касается конкретного заболевания, трудно определить какой ионный канал может представлять собой эффективную мишень для терапевтического вмешательства. Первоочередная задача состоит в том, чтобы предоставить ген ионного канала, причастный к шизофрении, биполярному расстройству или заболеваниям, связанными с ними. Настоящее изобретение направлено на решение этих и других задач. Краткое изложение существа изобретения Настоящее изобретение относится к антителу или антигенсвязывающему домену, специфически связывающемуся с полипептидом с последовательностью SEQ ID No 5. Антитело или антигенсвязываю-щий домен может быть химерным, гуманизированным антителом или антигенсвязывающим доменом, или моноклональным или поликлональным антителом или антигенсвязывающим доменом человека. В одном из вариантов осуществления антитело или антигенсвязывающий домен имеет одну цепь. В другом варианте осущетвления антитело или антигенсвязывающий домен является моноспецифичным, биспе-цифичным, триспецифичным или мультиспецифичным антителом или антигенсвязывающим доменом. Настоящее изобретение также относится к применению антитела или антигенсвязывающего домена для диагностики шизофрении или биполярного расстройства. Настоящее изобретение, кроме того, относится к набору антисмысловых последовательностей, ин-гибирующих экспрессию ионного канала, кодируемого полинуклеотидом с SEQ ID No 1, 2, 3 или 4, содержащим фрагмент поликулеотида с последовательностью SEQ ID No 1, 2, 3 или 4, для получения лекарственного средства для лечения шизофрении или биполярного расстройства. В одном из вариантов осуществления такой набор антисмысловых последовательностей представляет собой набор рибозимов. Настоящее изобретение относится к применению соединения, которое активирует или ингибирует экспрессию или активность ионного канала полипептида, содержащего полипептид с последовательностью SEQ ID No 5 для получения лекарственного средства для лечения шизофрении или биполярного расстройства. Кроме того, настоящее изобретение относится к фармацевтической композиции, содержащей вышеуказанное антитело или антигенсвязывающий фрагмент, вышеуказанный набор антисмысловых последовательностей или вышеупомянутое соединение, которое активирует или ингибирует экспрессию или активность ионного канала полипептида, содержащего полипептид с последовательностью SEQ ID No 5, и подходящий носитель или эксципиент. Настоящее изобретение также относится к применению по крайней мере одного двуаллельного маркера (ДМ), связанного с ионным каналом, выбранного из группы двуаллельных маркеров, показанных в табл. 1' ниже, для определения подвержен ли индивидуум риску или страдает ли больной шизофренией или биполярным расстройством. Таблица Г Название маркер Полиморфизм Положение ДМ в SEQ ID Аллель Аллель N0:1 N0 : 2 N0:3 N0:4 N0:06 99-62626-168 12642 99-62632-275 14088 99-62633-409 24981 99-62611-51 69248 99-62605-56 73428 99-62635-443 80250 99-79335-60 51090 99-79336-369 61293 99-79338-332 80602 А10 99-79314-201 100485 99-79314-225 100509 А12 99-79316-158 106725 1658 А13 99-79322-224 166087 А14 99-79322-473 166336 А15 99-79306-182 235894 А16 99-79310-29 31646 4481 А17 99-79311-50 42373 А18 99-62617-105 105 - 3 - 008252 Настоящее изобретение относится к способу генотипирования, включающему стадии: a) выделения нуклеиновой кислоты из биологического образца; b) обнаружения наличия нуклеотида в одном или нескольких зависимых двуаллельных маркерах, связанных с ионным каналом, А1-А18, перечисленных в п.13. В одном из вариантов осуществления такой способ генотипирования осуществляют с использованием анализа микросеквенирования. Настоящее изобретение относится к диагностическому набору для диагностики шизофрении или биполярного расстройства, содержащему полинуклеотид, который является фрагментом полинуклеотида с последовательностью SEQ ID NO: 1, 2, 3 или 4, содержащего один или несколько биаллельных маркеров А1-А18. В одном из конкретных вариантов осуществления настоящего изобретения диагностический набор для диагностики шизофрении или биполярного расстройства, содержит a) поликлональное или моноклональное антитело, которое специфически связывается с полипептидом ионного канала, содержащим аминокислотную последовательность SEQ ID No 5, или с фрагментом пептида или его вариантом, необязательно меченным; b) реагент, с помощью которого осуществляется определение образовавшегося комплекса антиген-антитела, указанный реагент либо необязательно содержит метку, либо который может быть распознан меченным реагентом. Краткое описание чертежей На фиг. 1 изображена диаграмма, представляющая ВАС-карту области хромосомы 13q, содержащей CanIon-ген. На фиг. 2 изображен блок диаграмм типовой компьютерной системы. На фиг. 3 изображена структурная схема, иллюстрирующая один из вариантов осуществления процесса 200 по сравнению новой нуклеотидной или белковой последовательности с базой данных последовательностей с целью установления уровней гомологии между новой последовательностью и соответствующей последовательностью базы данных. На фиг. 4 изображена схема последовательности операций, иллюстрирующая один из вариантов осуществления процесса 250 в компьютере, чтобы определить действительно ли две последовательности гомологичны. На фиг. 5 изображена схема последовательности операций, иллюстрирующая один из вариантов процесса-идентификатора 300 для выявления в последовательности наличия характерной особенности. Краткое описание последовательностей, представленных в списке последовательностей SEQ ID No 1 содержит геномную последовательность Canlon, включающую 5'-регуляторную область ("левая" нетранскрибируемая область) и экзоны 1-7. SEQ ID No 2 содержит геномную последовательность CanIon, включающую экзоны 8-27. SEQ ID No 3 содержит геномную последовательность CanIon, включающую экзоны 28-44, а также 3'-регуляторную область ("правая" нетранскрибируемая область). SEQ ID No 4 содержит кДНК-последовательность CanIon. SEQ ID No 5 содержит аминокислотную последовательность, кодируемую кДНК SEQ ID No 4. SEQ ID No 6 содержит нуклеотидную последовательность ампликона, который включает двуал-лельный маркер А18. SEQ ID No 7 содержит праймер, включающий дополнительную 5'-последовательность PU, ниже описанную в примере 2. SEQ ID No 8 содержит праймер, содержащий дополнительную 5'-последовательность RP, ниже описанную в примере 2. В соответствии с правилами, относящимися к Спискам Последовательностей, в данном Списке Последовательностей используются коды, которые указывают расположение двуаллельных маркеров в данных последовательностях и которые идентифицируют каждый из аллелей, представленный по полиморфному основанию. Код "r" для данной последовательности свидетельствует о том, что один аллель по данному полиморфному основанию представляет собой гуанин, а другой аллель представляет собой аде-нин. Код "у" для данных последовательностей свидетельствует о том, что один аллель по данному полиморфному основанию представляет собой тимин, тогда как другой аллель представляет собой цитозин. Код "m" для данных последовательностей свидетельствует о том, что один аллель по данному полиморфному основанию представляет собой аденин, а другой аллель представляет собой цитозин. Код "k" для данных последовательностей свидетельствует о том, что один аллель по данному полиморфному основанию представляет собой гуанин, а другой аллель представляет собой тимин. Код "s" для данных последовательностей свидетельствует о том, что один аллель по данному полиморфному основанию представляет собой гуанин, а другой аллель представляет собой цитозин. Код "w" для данных последовательностей свидетельствует о том, что один аллель по данному полиморфному основанию представляет собой аденин, а другой аллель представляет собой тимин. Нуклеотидный код исходного аллеля для каждого двуаллельного маркера следующий: Двуаллельный маркер Исходный аллель 5-124-273 А (например) - 4 - 008252 В некоторых случаях полиморфные основания двуаллельных маркеров изменяют идентичность аминокислот в кодируемом полипептиде. Это указано в прилагаемом Списке Последовательностей с использованием элемента VARIANT, размещения Хаа в положении данной полиморфной аминокислоты и определения Хаа в виде двух альтернативных аминокислот. Например, если один аллель двуаллельного маркера представляет собой кодон САС, который кодирует гистидин, а другой аллель двуаллельного маркера представляет собой САА, который кодирует глутамин, то Список Последовательностей для кодируемого полипептида будет содержать Хаа в положении полиморфной аминокислоты. В данном случае Хаа должен определяться в виде гистидина или глутамина. В других случаях Хаа может обозначать аминокислоту, идентичность которой неизвестна, вследствие неопределенности нуклеотидной последовательности. В данном случае использование элемента UNSURE размещает Хаа в положение неизвестной аминокислоты и значение Хаа считается любым из 20 аминокислот или ограниченным числом аминокислот, подсказанных генетическим кодом. Подробное описание изобретения Ассоциация шизофрении и биполярного расстройства в семьях, доказанная при изучении близнецов и усыновления, хотя изменчивость по всему миру отсутствует, свидетельствует о том, что шизофрения и биполярное расстройство в первую очередь представляют собой генетические состояния, хотя факторы риска окружающей среды также до некоторой степени рассматриваются в качестве необходимых, достаточных или воздействующих друг на друга причин. Например, шизофрения встречается у 1% от общей численности населения. Но при наличии шизофрении у одного из дедушек или бабушек риск возникновения данной болезни возрастает до примерно 3%; если же шизофренией страдает один из родителей, этот риск возрастает до примерно 10%. Если оба родителя больны шизофренией, тогда риск возрастает приблизительно до 40%. Идентификация генов, связанных с характерным признаком, таким как специфическое нарушение центральной нервной системы, подобное шизофрении, может быть осуществлена с помощью двух основных подходов, используемых в настоящее время для генетического картирования: анализа сцепления и ассоциативные исследования (исследование сообществ). Анализ сцепления требует изучения семей, в которых заболеванием поражены многие их члены, и в настоящее время пригоден для выявления моно- и олигогенных наследственных признаков. И наоборот, в ассоциативных исследованиях анализируют частоту маркерных аллелей признака (Т+) у несвязанных родством индивидов, сравниваемых с негативным по данному признаку контролем (Т-) и обычно используемых для выявления полигенного наследования. Генетическая связь или сцепление базируется на анализе, в котором две соседствующие последовательности на хромосоме содержат, по меньшей мере, рекомбинации в результате кроссинговера во время мейоза. Чтобы осуществить это хромосомные маркеры, подобные микросателлитным маркерам, тщательно локализуют в данном геноме. В анализе генетического сцепления рассчитывают вероятность рекомбинаций в данном гене-мишени с использованием хромосомных маркеров, в соответствии с генеалогическим древом, передачей данного заболевания и передачей данных маркеров. Таким образом, если отдельный аллель данного маркера передается вместе с заболеванием более часто, а не случайно (уровень рекомбинации находится между 0 и 0,5), то можно сделать вывод о том, что ген-мишень, о котором идет речь, обнаруживается поблизости от данного маркера. При использовании этого метода можно локализовать несколько генов, демонстрирующих генетическую предрасположенность семейных злокачественных опухолей. Для того чтобы иметь возможность включиться в изучение генетического сцепления, семьи, пораженные наследственной формой данного заболевания, должны удовлетворять критерию "информативности": несколько человек (конституциональная ДНК которых доступна) на поколение, должны быть поражены данным заболеванием, и в лучшем случае, обладать большим числом сибсов. Результаты исследований по сцеплению подтверждают гипотезу о том, что хромосома 13, по-видимому, содержит локус предрасположенности к шизофрении на 13q32 (Blouin J.L. и соавт., 1998, Nature Genetics, 20:70-73; Lin M.W. и соавт., Hum. Genet., 99(3) (1997):417-420; Brzustowicz и соавт., Am. J. Hum. Genet. 65:1096-1103 (1999)). Однако, несмотря на то, что анализ сцепления представляет собой мощный метод выявления генов и связанного с ними признака, решение часто невозможно без уровня данной мегабазы данных, а соответствующие исследования часто нуждаются в уточнении анализа участков, первоначально идентифицированных с помощью данного способа. ВАС-контиг, покрывающий предполагаемую геномную область локуса хромосомы 13q-31-q33, конструированы с использованием общедоступных STS, расположенных в данной области хромосомного локуса 13q31-q33, для скринирования запатентованной отдельной ВАС-библиотеки, эквивалентной 7-геномам. По данным материалам получали новые STS, позволяющие конструировать компактную физическую карту данной области. Определяли размер всех ВАС и картировали с помощью хромосомной гибридизации in situ для верификации. Идентифицировали минимальную группу ВАС и полностью сек-венировали по нескольким контигам, что приводило к получению конечного контига, превышающего 4 млн.п.н. Создание данной карты позволяет идентифицировать CanIon-ген, который локализуется в геномной области, демонстрирующей существенное сцепление с шизофренией. Аминокислотная последовательность CanIon характерна для CACHANNEL, 7-элементного фингер-принта, который характеризует альфа-1-субъединицы кальциевых каналов (Ref. PR00167, BLOCK+база - 5 - 008252 данных). Данный фингерпринт получали, исходя из первичного выравнивания 6 последовательностей: мотивы извлекали из консервативных областей петли, поддающихся различению между этими и другими катионными каналами; мотивы 1 и 2 кодируют последовательности между трансмембранными сегментами 4 и 5, а также 5 и 6 (первый внутренний повтор); мотив 3 соответствует последовательности между сегментом 6 повтора 1 и сегментом 1 повтора 2; мотив 4 кодирует последовательность между сегментами 5 и 6 повтора 2; мотив 5 соответствует последовательности между сегментом 6 повтора 3 и сегментом 1 повтора 4; а мотивы 6 и 7 кодируют последовательности между сегментами 4 и 5, а также между сегментами 5 и 6 повтора 4. На фиг. 1 представлены ВАС-контиги, покрывающие представляющую интерес область хромосомы 13, которая включает CanIon-ген, и показано геномное расположение CanIon-гена относительно генетических маркеров, демонстрирующих наивысшую значимость в исследованиях по сцеплению. В частности, Blouin и соавт. (1998) осуществили широкое исследование генома на предмет выявления локусов предрасположенности к шизофрении, в 54 комплексных родословных, с использованием 452 микроса-теллитных маркеров. Наиболее значимое сцепление между шизофренией в семьях обнаруживалось на хромосоме 13q32 вблизи маркера D13S174. Brzustowicz и соавт. (1999) дали оценку микросателлитным маркерам, охватывающим хромосомы 8 и 13 в 21 обширных Канадских семьях. Маркеры для хромосомной области 13q дают позитивные оценки LOD в каждой используемой аналитической модели: аутосом-но-доминантной и аутосомно-рецессивной, с узким или широким определением шизофрении. Максимальные трехточечные LOD-оценки были получены с маркером D13S793 в рецессивно-широкой модели: 3,92 в рекомбинантной фракции (8) 0,1 при гомогенности и 4,42 с а=0,65 и 8=8 при гетерогенности. На указанной фиг. 1 CanIon-ген локализован частично в контиге, помеченном 'Region E' и частично в конти-ге, помеченном С0001А10. Данный CanIon-ген фланкирован двумя маркерами, демонстрирующими наивысшую значимость в исследованиях по сцеплению. Маркер D13S174 также находится в контиге С0001А10, а маркер D13S793 локализован, приблизительно 3,5 млн.п.н. центромернее к CanIon-гену. Существует очень большая потребность в идентификации генов, связанных с шизофренией, биполярным расстройством и другими ЦНС, а также сердечно-сосудистыми заболеваниями и состояниями. Существует также необходимость в идентификации новых ионных каналов, связанных с заболеваниями. Такие гены и белки могут открыть новые перспективы в лечении шизофрении, биполярного расстройства или иных ЦНС-состояний, а также иных состояний, таких как, например, состояния сердца и гипер-тензия, и позволяют, кроме того, изучать и характеризовать CanIon-ген и связанные с ним биологические пути. Познание таких генов и связанных с ними биологических путей, вовлеченных в данные заболевания и состояния, позволит исследователям понять этиологию, например, шизофрении и биполярного расстройства, и приведет к разработке лекарственных средств и лекарственных препаратов, нацеленных против причины данных заболеваний. Например, соединения, которые блокируют CanIon-каналы, могут использоваться для лечения любых заболеваний и состояний, предпочтительно шизофрении или биполярного расстройства, и включающих также болевые нарушения, эпилепсию и различные сердечнососудистые нарушения, такие как сердечная аритмия, стенокардия и гипертензия. Существует также большая необходимость в новых способах выявления предрасположенности к шизофрении, биполярному расстройству и иным состояниям, а также для предотвращения или для наблюдения за развитием любого из этих заболеваний. Диагностические средства могли бы также оказаться в высшей степени полезными. В самом деле, в качестве примера, ранняя идентификация у людей с риском развития шизофрении сделает возможным раннее и/или профилактическое осуществление лечения. Кроме того, точные оценки эффективности лекарственного средства, а также толерантности пациента, даст возможность врачу-клиницисту улучшить соотношение между выгодой и риском, что касается схем лечения шизофрении и биполярного расстройства. Определения Перед более подробным описанием настоящего изобретения формулируются следующие ниже определения, которые иллюстрируют и задают значение и рамки терминов, используемых для описания представленного в данном описании изобретения. Используемый в данном описании термин "CanIon-ген" включает в себя геномные, мРНК и кДНК последовательности, кодирующие CanIon-белок, в том числе и нетранслируемые регуляторные области геномной ДНК. Используемый в данном описании термин "гетерологичный белок" предназначен для обозначения любого белка или полипептида, отличающегося от CanIon-белка. Например, гетерологичный белок может представлять собой соединение, которое можно использовать в качестве маркера в дополнительных экспериментах с регуляторной областью CanIon. Термин "выделенный" требует, чтобы материал был удален из своей первоначальной окружающей среды (например, природной окружающей среды, если он является встречаемым в природе). Например, встречаемый в природе полинуклеотид или полипептид, представленный в живом организме животного, не является выделенным, но тот же полинуклеотид или ДНК, или полипептид, отделенный от некоторых или от всех сосуществующих веществ в данной природной системе, является выделенным. Такой поли-нуклеотид может являться частью вектора и/или такой полинуклеотид или полипептид может являться - 6 - 008252 частью композиции, и все же являться выделенным, вектор или композиция не являются частью его природного окружения. Например, встречаемый в природе полинуклеотид, представленный в живом организме животного, не является выделенным, но тот же полинуклеотид, отделенный от некоторых или из всех сопутствующих веществ в природной системе, является выделенным. В частности, исключенными из определения " выделенный" являются: встречаемые в природе хромосомы (такие как хромосомные распределения), библиотеки искусственных хромосом, геномные библиотеки и кДНК-библиотеки, которые существуют либо в виде in vitro препарата нуклеиновой кислоты, либо в виде препарата трансфицирован-ной/трансформированной клетки-хозяина, где данные клетки-хозяева представляют собой либо гетерогенный in vitro препарат, либо посеянные в виде гетерогенной популяции из единичных колоний. Исключаются также, в частности, и указанные выше библиотеки, в которых конкретный полинуклеотид составляет менее 5% от числа вставок нуклеиновых кислот в векторные молекулы. Кроме того, исключаются, в частности, препараты геномной ДНК целых клеток или РНК целых клеток (включая указанные выше препараты целых клеток, которые механически нарезаются или энзиматически расщепляются). Кроме того, исключаются, в частности, препараты указанных выше целых клеток в виде либо in vitro препарата, либо в виде гетерогенной смеси, разделенной с помощью электрофореза (включая блотинг), где полинуклеотид настоящего изобретения не был дополнительно отделен от гетерологичных полинук-леотидов в электрофоретической среде (например, дополнительное отделение путем вырезания одиночной полосы из гетерогенной популяции полос в агарозном геле или в найлоновом блоте). Термин "выделенный очисткой" не требует абсолютной степени очистки; скорее он предназначается в качестве относительного определения. Очистка исходного вещества или природного вещества до по меньшей мере одного порядка величины, предпочтительно двух или трех порядков, и более предпочтительно четырех или пяти порядков, определенно предполагается. В качестве примера, очистка с 0,1% концентрации до 10% концентрации является очисткой на два порядка. В качестве примера, индивидуальные кДНК-клоны, выделенные из кДНК-библиотеки, были традиционно очищены до электрофорети-ческой гомогенности. Последовательности, полученные из данных клонов, невозможно было бы получить непосредственно из данной библиотеки или из тотальной ДНК человека. Данные кДНК-клоны не являются встречаемыми в природе как таковые, но скорее их получают с помощью операции частично выделяемого очисткой встречаемого в природе вещества (информационная РНК). Преобразование мРНК в кДНК-библиотеку включает в себя создание синтетического вещества (кДНК), а очищенные индивидуальные кДНК-клоны могут быть выделены из данной синтетической библиотеки с помощью клональной селекции. Таким образом, создание кДНК-библиотеки из информационной РНК и последовательное выделение индивидуальных клонов из данной библиотеки приводит, приблизительно, к 104-106-кратной очистке нативной РНК. Кроме того, термин "выделенный очисткой" используют в данном описании для описания полипептида или полинуклеотида настоящего изобретения, который был отделен от других соединений, включая, но, не ограничиваясь ими, полипептиды или полинуклеотиды, углеводы, липиды и т.п. Термин "выделенный очисткой" можно использовать для конкретного отделения мономерных полипептидов настоящего изобретения от олигомерных форм, таких как гомо- или гетеродимеры, димеры, тримеры и т.п. Термин "выделенный очисткой" можно также использовать в отношении специфического отделения ко-валентнозамкнутых полинуклеотидов от линейных полинуклеотидов. Любой полинуклеотид является в значительной степени чистым, если, по меньшей мере, около 50%, предпочтительно 60-75% образца обнаруживает единственную полинуклеотидную последовательность и конформацию (например, линейную в сравнении с ковалентнозамкнутой). В значительной степени чистый полипептид или полинуклео-тид обычно включает около 50%, предпочтительно 60-90% масса/масса полипептидного или полинук-леотидного образца, соответственно, предпочтительно около 95% и более предпочтительно около 99% чистого продукта. Беспримесность полипептида или полинуклеотида или гомогенность показывают с помощью ряда способов, хорошо известных в данной области техники, таких, например, как электрофорез образца в геле агарозы или в геле полиакриламида, с последующей визуализацией единственной полосы после окрашивания геля. Для некоторых целей наибольшее разрешение можно получить с использованием ВЭЖХ или иных способов, хорошо известных в данной области техники. В качестве альтернативного варианта осуществления настоящего изобретения, очистка полипептидов и полинуклеотидов настоящего изобретения может быть выражена, по меньшей мере, в виде процентов чистого вещества по отношению к гетерологичным полипептидам и полинуклеотидам (ДНК, РНК или обоим). В качестве предпочтительного варианта осуществления настоящего изобретения, полипептиды и полинуклеотиды настоящего изобретения обладают, по меньшей мере, 10, 20, 30, 40, 50, 60, 70, 80, 90, 95, 96, 98, 99 или 100% чистоты (беспримесности) относительно, соответственно, гетерологичных полипептидов и поли-нуклеотидов. В качестве еще более предпочтительного варианта осуществления настоящего изобретения данные полипептиды и полинуклеотиды обладают степенью очистки, колеблющейся от любого числа до тысячной доли, между 90 и 100% (например, полипептид или полинуклеотид, по меньшей мере, 99,995% чистоты) по отношению, соответственно, к их гетерологичным полипептидам и полинуклеотидам, или в виде соотношения масса/масса по отношению ко всем соединениям и молекулам, отличным от сущест - 7 - 008252 вующих в данном носителе. Каждое число, представляющее процент чистоты, до тысячной доли, можно заявлять в качестве индивидуального вида чистоты. Термин "полипептид" относится к полимеру аминокислот, не принимая во внимание длину данного полимера; поэтому пептиды, олигопептиды и белки включены без определения полипептида. Данный термин также не определяет или исключает постэкспрессионные модификации полипептидов, например, полипептидов, которые имеют в своем составе ковалентно присоединенные гликозильные группы, ацетильные группы, фосфатные группы, липидные группы и им подобные, четко подпадаемых под термин полипептид. В данное определение включены также полипептиды, которые содержат один или несколько аналогов аминокислот (включая, например, не встречаемые в природе аминокислоты, аминокислоты, которые встречаются в природе только в неродственной биологической системе, модифицированные аминокислоты из систем млекопитающих и т.п.), полипептиды с замещенными связями, а также иные модификации, известные в данной области техники, как встречаемые в природе, так и не встречаемые в природе. Используемый в данном описании термин "рекомбинантный полипептид" относится к полипептиду, который искусственно создают и который включает, по меньшей мере, две полипептидные последовательности, которые не обнаруживаются в виде смежных полипептидных последовательностей в их природной среде, или относится к полипептидам, которые экспрессируются из рекомбинантного полинуклеотида. Используемый в данном описании термин "животное, не принадлежащее человеческому роду", относится к любому позвоночному, не принадлежащему человеческому роду, птицам и, более предпочтительно, млекопитающим, предпочтительно приматам, сельскохозяйственным животным, таким как свинья, козы, овцы, ослы и лошади, кролики или грызуны, более предпочтительно крысы или мыши. Используемый в данном описании термин "животное" относится к любому позвоночному, предпочтительно млекопитающему. Оба термина "животное" и "млекопитающее" определенно включают человеческие существа, если не предшествует термин "не принадлежащий человеческому роду". Используемый в данном описании термин "антитело" относится к полипептиду или к группе полипептидов, которые включают, по меньшей мере, один связывающий домен, отличающийся тем, что анти-телосвязывающий домен образуется из упакованных вариабельных доменов молекулы антитела с образованием трехмерного связывающего пространства с формой внутренней поверхности и распределением заряда, комплементарного особенностям антигенной детерминанты антигена, что позволяет иммунологи-чески реагировать с данным антигеном. Антитела имеют в своем составе рекомбинантные белки, включающие связывающие домены, а также фрагменты, включающие Fab, Fab', F(ab)2 и F(ab')2-фрагменты. Используемая в данном описании "антигенная детерминанта" представляет собой часть молекулы антигена, в данном случае CanIon-полипептида, которая определяет специфичность взаимодействия антиген-антитело. "Эпитоп" относится к антигенной детерминанте полипептида. Эпитоп может включать не менее 3 аминокислот для пространственной конформации, которая уникальна для данного эпитопа. Как правило, эпитоп включает по меньшей мере 6 таких аминокислот, но более обычно, по меньшей мере, 8-10 таких аминокислот. Способы определения аминокислот, которые составляют эпитоп, включают рентгеновскую кристаллографию, 2-мерный ядерный магнитный резонанс и эпитопное картирование, например, способом Pepscan, описанном Geysen и соавт., 1984; публикация РСТ No WO84/03564; и публикация РСТ No WO84/03506. На всем протяжении настоящего описания экспрессия "нуклеотидной последовательности" может употребляться для обозначения полинуклеотида или нуклеиновой кислоты. Более точно, экспрессия " нуклеотидной последовательности" охватывает собственно материал нуклеиновой кислоты и, следовательно, не ограничивается информацией о последовательности (т.е., последовательности букв, выбранной из четырех букв основания), которая биохимически характеризует специфичность молекулы ДНК или РНК. Используемые в данном описании попеременно термины "нуклеиновые кислоты", "олигонуклеоти-ды" и " полинуклеотиды" включают РНК, ДНК или гибридные последовательности РНК/ДНК с более чем одной последовательностью в одноцепочечной или дуплексной форме. Термин "нуклеотид" используют в данном описании в качестве прилагательного к описанию молекул, включающих РНК, ДНК, или гибридных последовательностей РНК/ДНК любой длины в одноцепочечной или дуплексной форме. Термин "нуклеотид", также используемый в данном описании в качестве существительного, относится к индивидуальным нуклеотидам или к множеству нуклеотидов, подразумевая молекулу или индивидуальный элемент в большей молекуле нуклеиновой кислоты, включающей пурин или пиримидин, сахарную составляющую рибозы или дезоксирибозы и фосфатную группу, или фосфодиэфирную связь в случае нук-леотидов в олигонуклеотиде или полинуклеотиде. Хотя используемый в данном описании термин "нук-леотид" охватывает также "модифицированные нуклеотиды", которые включают, по меньшей мере, одну из модификаций (а) альтернативную связывающую группу, (b) аналоговую форму пурина, (с) аналоговую форму пиримидина или (d) аналоговый сахар, примеры аналоговых связывающих групп, пурина, пиримидинов и cахаров см., например, в публикации РСТ No WO95/04064. Полинуклеотидные последовательности настоящего изобретения могут быть получены любым известным способом, включая синтетический, рекомбинантный, ex vivo преобразование или их сочетанием, а также используя любые способы очистки, известные в данной области техники. Последовательность, которую "функционально присоединяют" к регуляторной последовательности, - 8 - 008252 такой как промотор, означает, что указанный регуляторный элемент находится в правильном положении и ориентации по отношению к нуклеиновой кислоте, для контролирования РНК-полимеразной инициации и экспрессии представляющей интерес нуклеиновой кислоты. Используемый в данном описании термин "функционально присоединенный" относится к связыванию полинуклеотидных элементов в функциональной взаимосвязи. Например, промотор или энхансер функционально связывают с кодирующей последовательностью, если он влияет на транскрипцию данной кодирующей последовательности. Термины "признак" и "фенотип" используются в данном описании взаимозаменяемо и относятся к любому видимому, обнаруживаемому или иначе измеряемому свойству организма, такому, например, как симптомы или предрасположенность к заболеванию. Обычно термины "признак" или "фенотип", используемые в данном описании, относят к симптомам или к предрасположенности к заболеванию, к положительному ответу или к побочным эффектам, связанным с лечением. Предпочтительно, указанный признак может представлять собой, без ограничения, психические расстройства, такие как шизофрения или биполярное расстройство, другие ЦНС или нейрональные расстройства, такие как эпилепсия или нарушения болевых ощущений, а также сердечно-сосудистые состояния, такие как стенокардия, гипертензия и аритмия, а также любой аспект, особенность или характеристику любого из данных заболеваний или состояний. Термин "аллель", используемый в данном описании, относят к вариантам нуклеотидной последовательности. Двуаллельный полиморфизм имеет две формы. Диплоидные организмы могут быть гомозиготой или гетерозиготой по аллельной форме. Термин "уровень гетерозиготности", используемый в данном описании, относят к доле индивидуумов в популяции, которые являются гетерозиготами по отдельному аллелю. В двуаллельной системе уровень гетерозиготности равен, в среднем, 2Ра(1-Ра), где Ра соответствует частоте наименее распространенного аллеля. Для использования в генетических исследованиях генный маркер должен обладать адекватным уровнем гетерозиготности, допускающим приемлемую вероятность того, что случайно выбранный субъект будет гетерозиготой. Термин "генотип", используемый в данном описании, относят к тождественности аллелей, представленных у индивидуума или в образце. В контексте настоящего изобретения генотип предпочтительно относят к описанию двуаллельных маркерных аллелей, представленных у индивида или в образце, например, аллели двуаллельных маркеров в CanIon-гене или в геномной области. Термин "генотипиро-вание" образца или индивида по двуаллельному маркеру включает в себя определение конкретного алле-ля или конкретного нуклеотида, осуществляемое по индивидуальному двуаллельному маркеру. Термин "мутация", используемый в данном описании, относят к различию в ДНК-последовательности между или среди разных геномов или индивидов, которые обладают частотой ниже 1%. Термин "гаплотип" относят к сочетанию аллелей, представленных у индивида или в образце. В контексте настоящего изобретения гаплотип предпочтительно относят к сочетанию двуаллельных маркерных аллелей, обнаруживаемых у данного индивида, и которые могут быть ассоциированы с фенотипом. Термин "полиморфизм", используемый в данном описании, относят к встречаемости одной или нескольких альтернативных геномных последовательностей или аллелей среди или у разных геномов или индивидов. "Полиморфный" относится к состоянию, в котором два или более вариантов конкретной геномной последовательности могут быть обнаружены в популяции. "Полиморфный сайт" представляет собой локус, по которому наблюдается изменчивость. Полиморфизм единственного нуклеотида представляет собой замещение одного нуклеотида другим нуклеотидом в данном полиморфном сайте. Деле-ция единичного нуклеотида или вставка единичного нуклеотида также приводит к возрастанию полиморфизма единственного нуклеотида. В контексте настоящего изобретения "полиморфизм единственного нуклеотида" предпочтительно относится к единичной нуклеотидной замене. Обычно у разных индивидов данный полиморфный сайт может быть занят двумя разными нуклеотидами. Термины "двуаллельный полиморфизм" и "двуаллельный маркер" используются в данном описании взаимозаменямо и относятся к полиморфизму единственного нуклеотида, обладающего двумя аллелями с достаточно высокой частотой в популяции. "Двуаллельный маркерный аллель" относится к нуклеотид-ным вариантам, представленным по сайту двуаллельного маркера. Обычно частота малораспространенного аллеля двуаллельных маркеров настоящего изобретения, как оценка, составляет более чем 1%, предпочтительно его частота превышает 10%, более предпочтительно его частота составляет по меньшей мере 20% (т.е., уровень гетерозиготности составляет по меньшей мере 0,32), еще более предпочтительно его частота равна по меньшей мере 30% (т.е., уровень гетерозиготности достигает по меньшей мере 0,42). Двуаллельный маркер, в котором частота малораспространенного аллеля равна 30% или больше, называют "высококачественным двуаллельным маркером". Положение нуклеотидов в полинуклеотиде относительно центра полинуклеотида описывается в данном описании следующим образом. Если полинуклеотид обладает нечетным числом нуклеотидов, то считают, что нуклеотид, равноотстоящий от 3' и 5' концов в данном полинуклеотиде, находится "в центре" данного полинуклеотида, и любой нуклеотид, непосредственно примыкающий к нуклеотиду в центре, или же сам нуклеотид в центре, считают находящимся "в 1 нуклеотиде от центра". При нечетном числе нуклеотидов в полинуклеотиде любую из пяти нуклеотидных позиций в средине полинуклеотида следует считать находящейся в 2 нуклеотидах от центра, и так далее. Если полинуклеотид обладает чет - 9 - 008252 ным числом нуклеотидов, то в центре данного полинуклеотида будет находиться связь, а не нуклеотид. Таким образом, любой из двух центральных нуклеотидов следует считать находящимися "в 1 нуклеотиде от центра", а любой из четырех нуклеотидов в средине данного полинуклеотида следует считать находящимся "в 2 нуклеотидах от центра", и так далее. Что касается полиморфизма, который связан с заменой, вставкой или делецией 1 или более нуклеотидов, то данный полиморфизм, аллель или двуаллельный маркер находится "в центре" полинуклеотида, если разница между расстоянием от замещенных, вставленных или делетированных полинуклеотидов данного полиморфизма до 3'-конца данного полинуклео-тида, расстоянием от замещенных, вставленных или делетированных полинуклеотидов данного полиморфизма до 5'-конца данного полинуклеотида, равна нулю или одному нуклеотиду. Если данная разница равна 0-3, тогда данный полиморфизм считают находящимся "в 1 нуклеотиде от центра". Если эта разница составляет 0-5, данный полиморфизм считают находящимся "в 2 нуклеотидах от центра". Если же эта разница составляет 0-7, то данный полиморфизм считают находящимся "в 3 нуклеотидах от центра", и т.д. Термин "левее", используемый в данном описании, относится к месту, которое находится около 5'-конца данного полинуклеотида от определенной точки отсчета, или, в случае гена, в направлении от кодирующей последовательности к промотору. Термины "спаренные основания" и "спаренные основания Уотсона и Крика", используемые в данном описании взаимозаменяемо, относятся к нуклеотидам, которые могут быть связаны друг с другом водородной связью, благодаря идентичности их последовательностей, способом, подобным тому, который обнаруживает двухспиральная ДНК для остатков тимина или урацила, связанных с остатками аде-нина с помощью двух водородных связей, и остатков цитозина и гуанина, связанных с помощью трех водородных связей (См. Stryer, L., Biochemistry, 4th edition, 1995). Термины "комплементарный" или "его комплемент", используемые в данном описании, относятся к последовательностям полинуклеотидов, которые способны образовывать пары оснований Уотсона и Крика с другим конкретным полинуклеотидом по всей полностью комплементарной области. Для данной цели настоящего изобретения первый полинуклеотид признают комплементарным второму полинуклео-тиду, если каждое основание первого полинуклеотида спаривается со своим комплементарным основанием. Комплементарными основаниями являются, как правило, А и Т (или А и U), или С и G. "Комплемент" используется в данном описании в качестве синонима "комплементарного полинуклеотида", "комплементарной нуклеиновой кислоты" и "комплементарной нуклеотидной последовательности". Данные термины, применяемые к парам полинуклеотидов, базируются исключительно на их последовательностях, а не на какой-либо конкретной совокупности условий, при которых данные два полинуклеотида были бы фактически связаны. Варианты и фрагменты 1. Полинуклеотиды. Варианты полинуклеотидов, в виде используемого в данном описании термина, представляют собой полинуклеотиды, которые отличаются от исходного полинуклеотида. Вариант полинуклеотида может быть встречаемым в природе вариантом, таким, например, как встречаемый в природе аллельный вариант, или он может быть вариантом, который не известен в качестве встречаемого в природе. Такие не встречаемые в природе варианты данного полинуклеотида можно создать методами мутагенеза, в том числе и теми, которые применимы к полинуклеотидам, клеткам или организмам. Обычно, различия столь невелики, что нуклеотидные последовательности исходного и вариантного полинуклеотидов почти полностью совпадают, а по многим участкам идентичны. Варианты полинуклеотидов, в соответствии с настоящим изобретением, включают, без существенного ограничения, нуклеотидные последовательности, которые по меньшей мере на 95% идентичны по-линуклеотиду, выбранному из группы, состоящей из нуклеотидных последовательностей SEQ ID Nos 14, или полинуклеотидному фрагменту по меньшей мере из 12 последовательных нуклеотидов полинук-леотида, выбранного из группы, состоящей из нуклеотидных последовательностей SEQ ID Nos 1-4, и предпочтительно, по меньшей мере, идентичны на 99%, более предпочтительно, по меньшей мере, идентичны на 99,5%, и наиболее предпочтительны, по меньшей мере, идентичны на 99,8% полинуклеотиду, выбранному из группы, состоящей из нуклеотидных последовательностей SEQ ID Nos 1-4, или любому полинуклеотидному фрагменту, по меньшей мере, из 12 последовательных нуклеотидов полинуклеотида, выбранного из группы, состоящей из нуклеотидных последовательностей SEQ ID No 1-4. Нуклеотидные изменения, присутствующие в вариантном полинуклеотиде, могут быть молчащими, что означает, что они не изменяют аминокислоты, кодируемые данным полинуклеотидом. Однако нукле-отидные изменения могут также приводить и к аминокислотным заменам, добавлениям делециям, слияниям и укорочениям в данном полипептиде, кодируемом начальной последовательностью. В этих заменах, делециях или добавках могут участвовать один или более нуклеотидов. Данные варианты могут являться результатом изменений в кодирующей или некодирующей областях или в обоих. Изменения в кодирующих областях могут создавать консервативные или неконсервативные замены, делеции или добавки. В контексте настоящего изобретения особенно предпочтительными вариантами осуществления настоящего изобретения являются те, в которых данные полинуклеотиды кодируют полипептиды, которые фактически сохраняют ту же биологическую функцию или активность, что и зрелый CanIon-белок, или те, - 10 - 008252 в которых данные полинуклеотиды кодируют полипептиды, которые сохраняют или повышают основную биологическую активность, снижая в то же время вспомогательную (вторичную) биологическую активность. Полинуклеотидный фрагмент представляет собой полинуклеотид, обладающий последовательностью, которая совершенно такая же, что и часть, но не вся данная нуклеотидная последовательность, предпочтительно нуклеотидная последовательность CanIon-гена, и его варианты. Данный фрагмент может представлять собой участок интрона или экзона CanIon-гена. Он может также представлять собой какую-нибудь часть из регуляторных областей CanIon. Предпочтительно, такие фрагменты включают, по меньшей мере, один из двуаллельных маркеров А1-А17 или его комплементов или двуаллельный маркер, сцепленный при нарушении равновесия с одним или несколькими двуаллельными маркерами А1-А17. Такие фрагменты могут быть "автономными", т.е., не являться частью или слившимися с другими полинуклеотидами, или они могут быть включены в отдельный большой полинуклеотид, в котором они образуют сегмент или участок. Действительно, несколько таких фрагментов могут быть представлены в отдельном большом полинуклеотиде. Факультативно, такие фрагменты могут состоять или состоять в основном из непрерывного участка последовательности, по меньшей мере, из 8, 10, 12, 15, 18, 20, 25, 35, 40, 50, 70, 80, 100, 250, 500 или 1000 нуклеотидов в длину. Совокупность предпочтительных фрагментов содержит, по меньшей мере, один из двуаллельных маркеров А1-А17 CanIon-гена, который описан в данном описании, или его комплементы. 2. Полипептиды. Данный вариант может представлять собой 1) вариант, в котором один или несколько аминокислотных остатков заменены консервативным или неконсервативным аминокислотным остатком, и такой замещенный аминокислотный остаток может или может не кодироваться с помощью генетического кода, или 2) вариант, в котором один или несколько аминокислотных остатков включают замещенную группу, или 3) вариант, в котором мутантный CanIon сливается с другим соединением, таким, например, как соединение, которое увеличивает время полужизни данного полипептида (например, полиэтиленгликоль), или 4) вариант, в котором дополнительные аминокислоты сливаются с мутантным CanIon, такой, например, как лидерная или секреторная последовательность или последовательность, которую употребляют для выделения очисткой мутантного CanIon, или препротеиновая последовательность. Такие варианты считаются находящимися в компетенции специалистов в данной области техники. Полипептидный фрагмент представляет собой полипептид, обладающий последовательностью, которая идентична части, но не всей данной полипептидной последовательности, предпочтительно полипептид, кодируемый CanIon-геном и его вариантами. Что касается аминокислотной замены в аминокислотной последовательности полипептида, в соответствии с настоящим изобретением, одну или несколько аминокислот можно заменить "эквивалентными" аминокислотами. Выражение "эквивалентная" аминокислота используют в данном описании для обозначения любой аминокислоты, которая может быть заменена одной или несколькими аминокислотами, обладающими аналогичными свойствами, при условии, что специалисты в области химии пептидов могут предполагать вторичную структуру и гидропатичную природу данного полипептида, которые будут оставаться практически неизмененными. Большей частью следующие группы аминокислот соответствуют эквивалентным изменениям: (1) Ala, Pro, Gly, Glu, Asp, Gln, Asn, Ser, Thr; (2) Cys, Ser, Tyr, Thr; (3) Val, Ile, Leu, Met, Ala, Phe; (4) Lys, Arg, His; (5) Phe, Tyr, Trp, His. Специфичный вариант осуществления представляющей интерес модифицированной пептидной CanIon-молекулы, в соответствии с настоящим изобретением, включает, но не ограничивается указанным, пептидную молекулу, которая устойчива к протеолизу, в которой пептидная связь -CONH- модифицирована и заменена восстановленной связью (CH2NH), связью retro inverso (NHCO), метиленоксис-вязью (QH2-O), тиометиленовой связью (CH2-S), карбосвязью (СН2СН2), ацетометиленовой связью (СО-СН2), гидроксиэтиленовой связью (СЖШ-СН2), N-N-связью, Е-ак^-связью или -СН=СН-связью. В настоящее изобретение включен CanIon-полипептид человека или его фрагмент, или его вариант, в котором по меньшей мере одна пептидная связь была модифицирована, как описано выше. Такие фрагменты могут быть "автономными", т.е., не являться частью или слившимися с другими полипептидами, или они могут быть включены в отдельный большой полипептид, в котором они образуют сегмент или участок. Вместе с тем, несколько фрагментов можно включить в один большой полипептид. В качестве репрезентативных примеров полипептидных фрагментов настоящего изобретения в данном описании можно указать на фрагменты, которые обладают длиной от примерно 5, 6, 7, 8, 9 или 1015, 10-20, 15-40 или 30-55 аминокислот. Предпочтительными являются фрагменты, содержащие, по крайней мере, мутацию одной аминокислоты в белок CanIon. Идентичность нуклеиновых кислот или полипептидов Термины "процент идентичности последовательностей" и "процент гомологии", используемые в данном описании взаимозаменяемо, применяют для сравнения полинуклеотидов или полипептидов, и определяют путем сравнения двух оптимально выровненных последовательностей в окне сравнения, где часть полинуклеотидной или полипептидной последовательности в окне сравнения может включать добавки или делеции (т.е. пробелы) при сравнении с исходной последовательностью (которая не включает добавки или делеции) для оптимального выравнивания двух данных последовательностей. Указанный - 11 - 008252 процент вычисляют путем определения числа позиций, по которым у обеих последовательностей наблюдаются идентичные основания нуклеиновых кислот или аминокислотные остатки, с получением соответствующих позиций, делением числа соответствующих позиций на общее число позиций в данном окне сравнения и умножением полученного результата на 100 с получением процента идентичности последовательностей. Гомологию оценивают с использованием любого из множества разнообразных алгоритмов сравнения последовательностей и программ, известных в данной области техники. Такие алгоритмы и программы включают, но никоим образом не ограничиваются, TBLASTN, BLASTP, FASTA, TFASTA и CLUSTALW (Pearson and Lipman, 1988; Altschul и соавт., 1990; Thompson и соавт., 1994; Higgins и соавт., 1996; Altschul и соавт., 1990; Altschul и соавт., 1993). В особенно предпочтительном варианте осуществления настоящего изобретения гомологии последовательностей белков и нуклеиновых кислот оценивают с использованием Basic Local Alignment Search Tool ("BLAST"), который хорошо известен в данной области техники (см., например, Karlin and Altschul, 1990; Altschul и соавт., 1990, 1993, 1997). В частности, пять специфичных BLAST-программ используются для выполнения следующей задачи: (1) BLASTP и BLAST3 сравнивают запрашиваемую аминокислотную последовательность с последовательностью белка базы данных; (2) BLASTN сравнивает запрашиваемую нуклеотидную последовательность с нуклеотидной последовательностью базы данных; (3) BLASTX сравнивает воображаемые продукты трансляции шести рамок запрашиваемой нуклео-тидной последовательности (обе цепи) с последовательностью белка базы данных; (4) TBLASTN сравнивает запрашиваемую последовательность белка с нуклеотидной последовательностью базы данных, транслированную со всех шести открытых рамках считывания (обе цепи); и (5) TBLASTX сравнивает трансляцию шести рамок запрашиваемой нуклеотидной последовательности с трансляцией шести рамок нуклеотидной последовательности базы данных. BLAST-программы идентифицируют гомологичные последовательности путем идентификации сходных сегментов, которые именуются в данном описании как "высокооцениваемые пары сегментов", между представляющей интерес аминокислотной или нуклеотидной последовательностью и тест-последовательностью, которую предпочтительно получают из последовательности белка или нуклеотид-ной последовательности базы данных. Высокооцениваемые пары сегментов предпочтительно идентифицируют (т.е. выстраивают) с помощью способов оценки матрицы, многие из которых хорошо известны в данной области техники. Предпочтительно используемая матрица оценок представляет собой ВLOSUM62-матрицу (Connet и соавт., 1992; Henikoff and Henikoff, 1993). С меньшей предпочтительностью можно также использовать матрицы РАМ или РАМ250 (см., например, Schwartz and Dayhoff, eds., 1978). BLAST-программы оценивают статистическую значимость идентифицируемых высокооцениваемых пар сегментов и предпочтительно отбирают те сегменты, которые удовлетворяют порогу значимости, устанавливаемому пользователем, такому как устанавливаемый пользователем процент гомологии. Предпочтительно статистическая значимость высокооцененной пары сегментов оценивается с использованием формулы статистической значимости Карлина (см., например, Karlin and Altschul, 1990). BLAST-программы могут использоваться со значениями параметров по умолчанию или с модифицированными параметрами, созданными пользователем. Жесткие условия гибридизации С целью охарактеризовать такую гибридизующуюся нуклеиновую кислоту, в соответствии с настоящим изобретением, жесткие условия гибридизации являются следующими: стадию гибридизации осуществляют при 65°С в присутствии 6 х SSC-буфера, 5 х раствора Ден-хардта, 0,5% SDS и 100 мкг/мл ДНК спермы лосося. Стадию гибридизации осуществляют с последующими четырьмя стадиями отмывки: две отмывки по 5 мин, предпочтительно при 65°С в 2 х SSC и 0,1% SDS-буфере; одна отмывка 30 мин, предпочтительно при 65°С в 2 х SSC и 0,1% SDS-буфере, одна отмывка 10 мин, предпочтительно при 65°С в 0,1 х SSC и 0,1% SDS-буфере, данные условия гибридизации пригодны для молекулы нуклеиновой кислоты длиной около 20 нук-леотидов. Нет необходимости говорить о том, что описанные выше условия гибридизации адаптируют в соответствии с длиной требуемой нуклеиновой кислоты, следуя методам, хорошо известным специалистам в данной области техники. Подходящие условия гибридизации можно, например, адаптировать в соответствии с указаниями, изложенными в книге Hames и Higgins (1985). Геномные последовательности CanIon-гена Настоящее изобретение включает в себя CanIon-ген или геномные CanIon-последовательности, состоящие, или в основном состоящие из, или включающие последовательность SEQ ID Nos 1-3, комплементарную ей последовательность, а также ее фрагменты и варианты. Данные полинуклеотиды могут представлять собой полинуклеотиды, выделенные очисткой, выделенные или рекомбинантные. Настоящее изобретение включает в себя также выделенный очисткой, выделенный или рекомби-нантный полинуклеотид, включающий нуклеотидную последовательность, обладающую по меньшей мере 70, 75, 80, 85, 90 или 95% нуклеотидной идентичностью с нуклеотидной последовательностью SEQ - 12 - 008252 ID Nos 1-3 или с ее комплементарной последовательностью, или с ее фрагментом. Нуклеотидные различия, что касается нуклеотидной последовательности SEQ ID Nos 1-3, могут быть, как правило, распределены случайно по всей длине нуклеиновой кислоты. Тем не менее, предпочтительными нуклеиновыми кислотами являются те из них, в которых нуклеотидные различия, что касается нуклеотидной последовательности SEQ ID Nos 1-3, преимущественно локализованы за пределами данных кодирующих последовательностей, содержащихся в экзонах. Данные нуклеиновые кислоты, а также их фрагменты и варианты, могут использоваться в качестве олигонуклеотидных праймеров или зондов с целью обнаружения наличия копии CanIon-гена в тест-выборке или, альтернативно, с целью амплифицировать нуклеотидную последовательность-мишень в CanIon-последовательностях. Другой объект настоящего изобретения состоит из выделенной очисткой, выделенной или реком-бинантной нуклеиновой кислоты, которая гибридизуется с нуклеотидной последовательностью SEQ ID Nos 1-3 или с комплементарной ей последовательностью, или с ее вариантом, в жестких гибридизацион-ных условиях, как указано выше. В предпочтительных вариантах осуществления настоящего изобретения указанная выделенная очисткой, выделенная или рекомбинантная нуклеиновая кислота специфически гибридизуется с полинуклеотидами CanIon-гена человека, более предпочтительно, если указанная нуклеиновая кислота способна гибридизоваться с нуклеотидами из CanIon-гена человека, но практически неспособна гибридизоваться с нуклеотидной последовательностью CanIon-гена крысы. Особенно предпочтительная нуклеиновая кислота настоящего изобретения включает выделенные, выделенные очисткой или рекомбинантные полинуклеотиды, включающие непрерывный участок последовательности, по меньшей мере, из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 3000, 4000, 5000 или 10000 нуклеотидов из SEQ ID Nos 1-3 или из ее комплементов. Следует отметить, что нуклеотидные фрагменты любого размера и последовательности могут также включаться в полинуклеотиды, описанные в данном разделе. Геномная нуклеиновая кислота CanIon включает 44 экзона. Расположение экзонов в SEQ ID Nos 1-3 подробно представлено ниже в табл. В. Таблица В Экзон Положение в SEQ ID Nol Интрон Положение в SEQ ID Nol Начало Конец Начало Конец 2001 2026 2027 19187 19188 19334 19335 22995 22996 23178 23179 39730 39731 39814 39815 41336 41337 41476 41477 41564 41565 41693 41694 73012 73013 73167 Экзон Положение в SEQ ID No2 Интрон Положение в SEQ ID No2 1 Начало Конец Начало Конец 43726 43868 43869 43997 43998 44102 44103 52092 52093 52179 52180 77567 77568 77699 77700 98225 98226 98393 98394 106566 106567 106758 106759 144108 144246 144246 144247 159793 159794 159868 159869 191291 191292 191428 191429 192966 192967 193108 193109 211539 211540 211613 211614 225005 225006 225107 225108 225543 225544 225613 225614 228449 228450 228541 228542 228629 228630 228752 228753 231288 231289 231345 231346 231588 231589 231709 231710 231812 231813 231944 231945 232899 232900 233067 233068 235354 235355 235459 - 13 - 008252 Экзон Положение в SEQ ID No3 Интрон Положение в SEQ ID No3 Начало Конец Начало Конец 3895 4001 4002 9610 9611 9731 9732 9815 9816 9914 9915 15775 15776 15869 15870 16381 16382 16488 16489 16696 16697 16771 16772 17933 17934 18053 18054 23643 23644 23712 23713 24927 24928 25076 25077 25912 25913 26006 26007 30766 30767 30899 30900 31560 31561 31676 31677 34073 34044 34201 34202 37492 37493 3763 37644 39651 39652 39801 39802 41562 41563 41680 41681 44130 44131 45841 Таким образом, настоящее изобретение содержит выделенные очисткой, выделенные или рекоби-нантные полинуклеотиды, включающие нуклеотидную последовательность, выбранную из группы, состоящей, каждая, из 44 экзонов CanIon-гена и, каждая, - из комплементарной ей последовательности. В настоящем изобретении обеспечиваются также выделенные очисткой, выделенные или рекомбинантные нуклеиновые кислоты, включающие сочетание, по меньшей мере, из двух экзонов CanIon-гена, в котором данные полинуклеотиды размещены в нуклеиновой кислоте, от 5'-конца до 3'-конца указанной нуклеиновой кислоты, в том же порядке, что и в SEQ ID No 1-3. Интрон 1 относится к нуклеотидной последовательности, локализованной между Экзоном 1 и Эк-зоном 2, и так далее. Положение интронов детально представлено в табл. А. Таким образом, настоящее изобретение содержит выделенные очисткой, выделенные или рекобинантные полинуклеотиды, включающие нуклеотидную последовательность, выбранную из группы, состоящей из 43 интронов CanIon-гена, или комплементарную ей последовательность. Хотя данный раздел называется "Геномные Последовательности CanIon", следует иметь ввиду, что нуклеотидные фрагменты любого размера и последовательности могут также включаться в полинуклеотиды, описанные в данном разделе, фланкируя данные геномные последовательности CanIon на любой стороне или между двумя или более такими геномными последовательностями. кДНК-последовательности CanIon Показано, что экспрессия CanIon-гена приводит к продуцированию по меньшей мере одного из видов мРНК, последовательности нуклеиновой кислоты, которая приведена в SEQ ID No 4. Другой объект настоящего изобретения представляет собой выделенную очисткой, выделенную или рекомбинантную нуклеиновую кислоту, включающую нуклеотидную последовательность SEQ ID No 4, комплементарные ей последовательности, а также аллельные варианты, и ее фрагменты. Кроме того, предпочтительные полинуклеотиды настоящего изобретения включают выделенные очисткой, выделенные или рекомбинантные кДНК CanIon, состоящие, в основном состоящие из или включающие последовательность SEQ ID No 4. Особенно предпочтительные нуклеиновые кислоты настоящего изобретения включают выделенные, выделенные очисткой или рекомбинантные полинуклеотиды, включающие непрерывный участок последовательности, по меньшей мере, из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 3000, 4000, 5000 или 6000 нуклеотидов из SEQ ID No 4 или из ее комплементов. В предпочтительных вариантах осуществления настоящего изобретения указанный непрерывный участок последовательности включает связанный с CanIon двуаллельный маркер; предпочтительно избранный из группы, состоящей из А12 или А16. К настоящему изобретению относится также выделенная очисткой или выделенная нуклеиновая кислота, включающая полинуклеотид, обладающий по меньшей мере 95% нуклеотидной идентичностью с полинуклеотидом SEQ ID No 4, преимущественно 99% нуклеотидной идентичностью, предпочтительно 99,5% нуклеотидной идентичностью и наиболее предпочтительно 99,8% нуклеотидной идентичностью с полинуклеотидом SEQ ID No 4, или комплементарная ей последовательность или ее биологически активный фрагмент. Другая цель настоящего изобретения имеет отношение к выделенным очисткой, выделенным или рекомбинантным нуклеиновым кислотам, включающим полинуклеотид, который гибридизуется в сформулированных в данном описании жестких условиях гибридизации с полинуклеотидом SEQ ID No 4, или с комплементарной ему последовательностью или с его вариантом, или с его биологически активным - 14 - 008252 фрагментом. Дополнительная цель настоящего изобретения имеет отношение к выделенному, выделенному очисткой или рекомбинантному полинуклеотиду, который кодирует CanIon-полипептид, включающий непрерывный участок последовательности по меньшей мере из 6 аминокислот SEQ ID No 5, в котором указанный непрерывный участок последовательности включает, по меньшей мере, 1, 2, 3, 5 или 10 аминокислотных позиций 277, 338, 574, 678, 680, 683, 691, 692, 695, 696, 697, 894, 1480, 1481, 1483, 1484, 1485, 1630, 1631, 1632, 1636, 1660, 1667, 1707, 1709 SEQ ID No 5. Включен также выделенный, выделенный очисткой или рекомбинантный полинуклеотид, который кодирует CanIon-полипептид, включающий аминокислотную последовательность SEQ ID No 5, или производные или ее биологически активные фрагменты, а также выделенный, выделенный очисткой или рекомбинантный полинуклеотид, который кодирует CanIon-полипептид, идентичный по меньшей мере на 80, 85, 90, 95, 98, 99, 99,5 или 99% аминокислотной последовательности SEQ ID No 5. кДНК SEQ ID No 4 включает 5'-UTR-область, начинающуюся от нуклеотида в положении 1 и заканчивающуюся нуклеотидом в положении 65 SEQ ID No 4. кДНК из SEQ ID No кДНК включает 3'-UTR-область, начинающуюся от нуклеотида в положении 5283 и заканчивающуюся нуклеотидом в положении 6799 SEQ ID No 4. Следовательно, в настоящем изобретении рассматривается выделенная очисткой, выделенная или рекомбинантная нуклеиновая кислота, включающая нуклеотидную последовательность 5'UTR из кДНК CanIon, комплементарную ей последовательность, или ее аллельный вариант. В настоящем изобретении рассматривается также выделенная очисткой, выделенная или рекомбинантная нуклеиновая кислота, включающая нуклеотидную последовательность 3'UTR из кДНК CanIon, комплементарную ей последовательность, или ее аллельный вариант. Хотя данный раздел называется "кДНК-последовательности CanIon", следует иметь ввиду, что нук-леотидные фрагменты любого размера и последовательность могут быть также включены в полинуклео-тиды, описанные в данном разделе, фланкируя геномные последовательности CanIon на любой стороне или между двумя или несколькими такими геномными последовательностями. Кодирующие области Открытая рамка считывания CanIon содержится в соответствующей мРНК из SEQ ID No кДНК. Точнее, эффективная CanIon-кодирующая последовательность (CDS) включает область между нуклеотидной позицией 66 (первый нуклеотид ATG-кодона) и нуклеотидной позицией 5282 (концевой нуклео-тид TGA-кодона) из SEQ ID No 4. Настоящее изобретение включает также выделенные, выделенные очисткой и рекомбинантные полинуклеотиды, которые кодируют полипептиды, включающие непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно по меньшей мере 8 или 10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50, 100, 200, 300, 400, 500, 700 или 1000 аминокислот SEQ ID No 5. Раскрытый выше полинуклеотид, который содержит кодирующую последовательность CanIon-гена, может экспрессироваться в требуемой клетке-хозяине или в требуемом организме-хозяине, если данный полинуклеотид поместить под контроль соответствующих экспрессионных сигналов. Экспрессионные сигналы могут представлять собой или экспрессионные сигналы, содержащиеся в регуляторных районах CanIon-гена настоящего изобретения, или в противоположность сигналы могут представлять собой экзогенные регуляторные последовательности нуклеиновых кислот. Такой полинуклеотид, помещенный под соответствующие экспрессионные сигналы, можно также встроить в вектор для его экспрессии и/или амплификации. Регуляторные CanIon-последовательности Как отмечено, геномная последовательность CanIon-гена содержит регуляторные последовательности как в некодирующей 5'-фланкирующей области, так и в некодирующей 3'-фланкирующей области, которые ограничивают CanIon-кодирующую область, содержащую 44 экзона данного гена. Полинуклеотиды, полученные из 5'- и 3'-регуляторных областей, пригодны для того, чтобы выявить наличие, по меньшей мере, копии нуклеотидной последовательности CanIon или ее фрагмента в тестируемом образце. Промоторную активность 5'-регуляторных областей, содержащихся в CanIon, можно оценить, как описано ниже. Для того чтобы идентифицировать релевантные биологически активные полинуклеотид-ные фрагменты или варианты SEQ ID No 1, специалистов в данной области техники можно отослать к Sambrook и соавт., 1989, которые описывают использование рекомбинантного вектора, несущего маркерный ген (т.е. бета-галактозидазу, хлорамфениколацетилтрансферазу и т.п.), экспрессию которого можно обнаружить при помещении его под контроль биологически активных полинуклеотидных фрагментов или вариантов SEQ ID No 1. Геномные последовательности, локализованные "левее" первого эк-зона CanIon-гена, клонируют в соответствующий промоторный репортерный вектор, такие как доступные от Clontech Промоторные репортерные векторы pSEAP-Basic, pSEAP-Enhancer, pPgal-Basic, pPgal-Enhancer или pEGFP1, или беспромоторный репортерный вектор гена люциферазы от Promega pGL2-basic или pGL3-basic. Вкратце, каждый из этих промоторных репортерных векторов включает множество сайтов клонирования, позиционированных "левее" репортерного гена, кодирующего легко поддающийся - 15 - 008252 анализу белок, такой как секретируемая щелочная фосфатаза, люцифераза, в-галактозидаза или зеленый флуоресцентный белок. Последовательности, расположенные "левее" от кодирующей CanIon-области, встраивают в сайты клонирования "левее" данного репортерного гена в обоих ориентациях и вводят в соответствующую клетку-хозяина. Анализируют уровень репортерного белка и сравнивают с уровнем, полученным для вектора, у которого отсутствует вставка в сайт клонирования. Наличие повышенного уровня экспрессии в векторе, содержащем вставку, по сравнению с уровнем для контрольного вектора, свидетельствует о наличии промотора в данной вставке. При необходимости расположенные слева последовательности могут быть клонированы в векторы, которые содержат энхансер для увеличения уровней транскрипции под слабыми промоторными последовательностями. Существенный уровень повышенной экспрессии, которая наблюдается для вектора, не имеющего вставки, свидетельствует о том, что промоторная последовательность присутствует во встроенной слева последовательности. Промоторную последовательность, расположенную "левее" геномной ДНК, можно также определить путем создания сгруппированных 5' и/или 3' делеций в расположенной слева ДНК с использованием традиционных методик, таких как обработка экзонуклеазой III или обработка соответствующей эндонук-леазой рестрикции. Полученные делеционные фрагменты можно встроить в данный промоторный ре-портерный вектор, чтобы определить, действительно ли данная делеция снижает или уничтожает промо-торную активность, как это описано, например, у Coles и соавт. (1998), раскрытие которой включено в данное описание путем ссылки во всей ее полноте. Таким способом можно определить границы промоторов. При желании можно идентифицировать потенциальные индивидуальные регуляторные сайты внутри промотора с использованием сайт-направленного мутагенеза или линкерного сканирования, которые аннулируют сайты связывания потенциальных транскрипционных факторов внутри промотора, индивидуально или в сочетании. Влияние таких мутаций на уровень транскрипции можно определить путем встраивания мутаций в сайты клонирования промоторных репортерных векторов. Данный вид анализа хорошо известен специалистам в данной области техники и описан в WO 97/17359, патенте США № 5374544; ЕР 582796; патенте США № 5698389; патенте США № 5643746; патенте США № 5502176; и патенте США № 5266488; раскрытие которых включено в данное описание путем ссылки во всей их полноте. Силу и специфичность промотора CanIon-гена можно оценить через уровни экспрессии детектируемого полинуклеотида, функционально присоединенного к данному CanIon-промотору, в разных типах клеток и тканях. Данный детектируемый полинуклеотид может представлять собой полинуклеотид, который специфически гибридизуется с заранее определенным олигонуклеотидным зондом, или поли-нуклеотид, кодирующий детектируемый белок, в том числе CanIon-полипептид или фрагмент или его вариант. Данный вид анализа хорошо известен специалистам в данной области техники и описан в патенте США № 5502176; и патенте США № 5266488; раскрытие которых включено путем ссылки во всей их полноте. Некоторые способы обсуждаются более подробно ниже. Полинуклеотиды, несущие регуляторные элементы, локализованные на 5' или на 3'-конце CanIon-кодирующей области, можно преимущественно использовать для контроля транскрипционной и трансляционной активности представляющего интерес гетерологичного полинуклеотида. Таким образом, в настоящем изобретении рассматривается также выделенная очисткой или выделенная нуклеиновая кислота, включающая полинуклеотид, который выбран из группы, состоящей из 5'-и 3'-регуляторных областей, или комплементарную ему последовательность, или ее биологически активный фрагмент или вариант. В предпочтительных вариантах осуществления настоящего изобретения "5'-регуляторная область" локализована в нуклеотидной последовательности, расположенной между позициями 1 и 2000 SEQ ID No 1. "3'- регуляторная область" локализована в нуклеотидной последовательности, расположенной между позициями 45842 и 47841 SEQ ID No 3. Настоящее изобретение относится также к выделенной очисткой или выделенной нуклеиновой кислоте, включающей полинуклеотид, обладающий, по меньшей мере, 95% нуклеотидной идентичности с полинуклеотидом, выбранным из группы, состоящей из 5' и 3' регуляторных областей, преимущественно 99% нуклеотидной идентичности, предпочтительно 99,5% нуклеотидной идентичности и наиболее предпочтительно 99,8% нуклеотидной идентичности с полинуклеотидом, выбранным из группы, состоящей из 5' и 3' регуляторных областей, или комплементарной ему последовательностью или его вариантом, или его биологически активным фрагментом. Другой объект настоящего изобретения включает выделенные очисткой, выделенные или рекомби-нантные нуклеиновые кислоты, включающие полинуклеотид, который гибридизуется в установленных в данном описании жестких условиях гибридизации с полинуклеотидом, выбранным из группы, состоящей из нуклеотидных последовательностей 5' и 3' регуляторных областей, или с комплементарной ему последовательностью или с его вариантом, или с его биологически активным фрагментом. Предпочтительные фрагменты 5'-регуляторной области обладают длиной около 1500 или 1000 нук-леотидов, предпочтительно около 500 нуклеотидов, более предпочтительно около 400 нуклеотидов, еще более предпочтительно около 300 нуклеотидов и наиболее предпочтительно около 200 нуклеотидов. Предпочтительные фрагменты 3'-регуляторной области обладают, по меньшей мере, 50, 100, 150, - 16 - 008252 200, 300 или 400 п.н. в длину. "Биологически активные" полинуклеотидные производные SEQ ID Nos 1-3 представляют собой по-линуклеотиды, включающие или же состоящие из фрагмента указанного полинуклеотида, который является функционирующим в качестве регуляторной области для экспрессирования рекомбинантного полипептида или рекомбинантного полинуклеотида в рекомбинантной клетке-хозяине. Он должен действовать либо в качестве энхансера, либо как репрессор. Для цели настоящего изобретения нуклеиновая кислота или полинуклеотид являются "функциональными" в качестве регуляторной области для экспрессии рекомбинантного полипептида или реком-бинантного полинуклеотида, если указанный регуляторный полинуклеотид содержит нуклеотидные последовательности, которые содержат информацию о регуляции транскрипции и трансляции, и такие последовательности "функционально связаны" с нуклеотидными последовательностями, которые кодируют требуемый полипептид или требуемый полинуклеотид. Регуляторные полинуклеотиды настоящего изобретения можно получить из нуклеотидной последовательности SEQ ID Nos 1 и 3 путем расщепления с использованием соответствующих ферментов рестрикции, что описано, например, в книге Sambrook и соавт. (1989). Регуляторные полинуклеотиды можно также получить путем обработки SEQ ID Nos 1 и 3 ферментом экзонуклеазой, такой как Ва131 (Wabiko и соавт., 1986). Такие регуляторные полинуклеотиды можно также получить с помощью химического синтеза нуклеиновых кислот, как описано в другом месте в данном описании. Регуляторные полинуклеотиды, в соответствии с настоящим изобретением, могут представлять собой часть рекомбинантного экспрессионного вектора, который можно использовать для экспрессии кодирующей последовательности в требуемой клетке-хозяине или организме-хозяине. Рекомбинантные экспрессионные векторы, в соответствии с настоящим изобретением, описаны в другом месте настоящего описания. Предпочтительный 5'-регуляторный полинуклеотид настоящего изобретения включает 5'-нетранс-лируемую область (5'-UTR) кДНК CanIon, или ее биологически активный фрагмент или ее вариант. Предпочтительный 3'-регуляторный полинуклеотид настоящего изобретения включает 3'-нетранс-лируемую область (3'-UTR) кДНК CanIon, или ее биологически активный фрагмент или ее вариант. Дополнительная цель настоящего изобретения включает выделенную очисткой или выделенную нуклеиновую кислоту, включающую: a) нуклеиновую кислоту, включающую регуляторную нуклеотидную последовательность, выбранную из группы, состоящей из: (i) нуклеотидной последовательности, включающей полинуклеотид из 5'-регуляторной области или комплементарную ей последовательность; (ii) нуклеотидной последовательности, включающей полинуклеотид, обладающий по меньшей мере 95% нуклеотидной идентичности с нуклеотидной последовательностью из 5'-регуляторной области или с комплементарной ей последовательностью; (iii) нуклеотидной последовательности, включающей полинуклеотид, который гибридизуется в жестких гибридизационных условиях с нуклеотидной последовательностью из 5'-регуляторной области или с комплементарной ей последовательностью; и (iv) биологически активного фрагмента или варианта полинуклеотидов в (i), (ii) и (iii); b) полинуклеотид, кодирующий требуемый полипептид или представляющую интерес нуклеиновую кислоту, функционально присоединенную к нуклеиновой кислоте, указанной выше в (а); c) факультативно, нуклеиновую кислоту, включающую 3'-регуляторный полинуклеотид, предпочтительно 3'-регуляторный полинуклеотид CanIon-гена. В конкретном варианте осуществления настоящего изобретения указанная выше нуклеиновая кислота отличается тем, что указанная нуклеиновая кислота включает 5'-нетранслируемую область (5'-UTR) кДНК CanIon, или ее биологически активный фрагмент или ее вариант. В другом конкретном варианте осуществления настоящего изобретения указанная выше нуклеиновая кислота отличается тем, что указанная нуклеиновая кислота включат 3'-нетранслируемую область (3'-UTR) кДНК CanIon, или ее биологически активный фрагмент или ее вариант. Регуляторный полинуклеотид 5'-регуляторной области или его биологически активные фрагменты или варианты, функционально присоединяют по 5'-концу полинуклеотида, кодирующего требуемый полипептид или полинуклеотид. Регуляторный полинуклеотид 3'-регуляторной области, или его биологически активные фрагменты или варианты, преимущественно функционально присоединяют по 3'-концу полинуклеотида, кодирующего требуемый полипептид или полинуклеотид. Требуемый полипептид, кодируемый указанной выше нуклеиновой кислотой, может иметь разную природу или происхождение, включая белки прокариотического или эукариотического происхождения. Полипептиды, экспрессируемые под контролем CanIon-регуляторной области, включают антигены бактерий, грибов или вирусов. Включены также эукариотические белки, такие как внутриклеточные белки, подобные белки "домашнего хозяйства", связанные с мембраной белки, подобные рецепторам, и секре-тируемые белки, подобные эндогенным медиаторам, таким как цитокины. Требуемый белок может быть - 17 - 008252 CanIon-белком, в частности, белком с аминокислотной последовательностью SEQ ID No 5, или фрагментом или его вариантом. Требуемые нуклеиновые кислоты, кодируемые указанным выше полинуклеотидом, обычно РНК-молекулой, могут быть комплементарны требуемому кодирующему полинуклеотиду, например, CanIon-кодирующей последовательности, и таким образом использоваться в качестве антисмыслового полинук-леотида. Такой полинуклеотид может включаться в рекомбинантный экспрессионный вектор, чтобы экс-прессировать требуемый полипептид или требуемую нуклеиновую кислоту в клетке-хозяине или в организме-хозяине. Соответствующие рекомбинантные векторы, которые содержит полинуклеотид, такой как описано в данном описании, рассматриваются в другом месте настоящего описания. Полинуклеотидные конструкции Термины "полинуклеотидная конструкция" и "рекомбинантный полинуклеотид", используемые в данном описании взаимозаменяемо, относятся к линейным или кольцевым, выделенным очисткой или выделенным полинуклеотидам, которые были искусственно созданы и которые включают, по меньшей мере, две нуклеотидные последовательности, которые не обнаруживаются в виде непрерывных нуклео-тидных последовательностей в их исходном природном окружении. ДНК-конструкция, которая обеспечивает прямую временную и пространственную экспрессию CanIon-гена в рекомбинантных клетках-хозяевах и в трансгенных животных Для того чтобы исследовать физиологические и фенотипические последствия отсутствия синтеза CanIon-белка, как на клеточном уровне, так и на уровне мультиклеточного организма, в настоящее изобретение включены также ДНК-конструкции и рекомбинантные векторы, обеспечивающие обусловленную экспрессию конкретного аллеля CanIon-геномной последовательности или кДНК, а также копии данной геномной последовательности или кДНК, содержащие замены, делеции или добавки из одного или более оснований, что касается CanIon-нуклеотидной последовательности SEQ ID Nos 1-4, или ее фрагмента, причем данные замены, делеции или добавки оснований локализованы в экзоне, интроне или регуляторной последовательности, но предпочтительно в 5'-регуляторной последовательности или в эк-зоне CanIon-геномной последовательности, или в CanIon-кДНК SEQ ID No 4. В предпочтительном варианте осуществления CanIon-последовательность включает двуаллельный маркер настоящего изобретения. В предпочтительном варианте осуществления настоящего изобретения CanIon-последовательность включает двуаллельный маркер настоящего изобретения, предпочтительно один из двуаллельных маркеров А1-А17. Настоящее изобретение содержит рекомбинантные векторы, включающие любой один из полинук-леотидов, описанных в настоящем изобретении. В частности, полинуклеотидные конструкции, в соответствии с настоящим изобретением, могут включать любой из полинуклеотидов, описанных в разделе "Геномные Последовательности CanIon-гена", в разделе "кДНК-последовательности CanIon", в разделе "Кодирующие Области" и в разделе "Олигонуклеотидные Зонды и Праймеры". Первую предпочтительную ДНК-конструкцию создают на основе оперона устойчивости к тетрациклину tet из транспозона Tn10 E. coli для контроля экспрессии CanIon-гена так, как описано у Gossen и соавт. (1992, 1995) и у Furth и соавт. (1994). Такая ДНК-конструкция содержит семь tet-операторных последовательностей из Tn10 (tetop), которые сливаются с минимальным промотором или с 5'-регуляторной последовательностью CanIon-гена, причем указанный минимальный промотор или указанную CanIon-регуляторную последовательность функционально присоединяют к представляющему интерес полинуклеотиду, который кодирует смысловой или антисмысловой олигонуклеотид или полипептид, включая CanIon-полипептид или его пептидный фрагмент. Данная ДНК-конструкция является функциональной в качестве условной экспрессионной системы представляющей интерес нуклеотидной последовательности, где одна и та же клетка включает нуклеотидную последовательность, кодирующую репрес-сор дикого типа (tTA) или мутантный (ГТА) репрессор, слитый с активирующим доменом вирусного белка VP16 из вируса простого герпеса, помещенного под контроль промотора, такого как HCMVIE1 энхан-сер/промотор или MMTV-LTR. Фактически предпочтительная ДНК-конструкция настоящего изобретения включает как полинуклеотид, содержащий tet-операторную последовательность, так и полинуклеотид, содержащий последовательность, кодирующую репрессор tTA или ГТА. В конкретном варианте осуществлении настоящего изобретения условная экспрессионная ДНК-конструкция содержит последовательность, кодирующую мутантный тетрациклиновый репрессор ГТА, причем экспрессия представляющего интерес полинуклеотида является молчащей в отсутствие тетрациклина и индуцируется в его присутствии. ДНК-конструкции, обеспечивающие гомологичную рекомбинацию: замещающие векторы Вторая предпочтительная ДНК-конструкция включает, от 5'-конца до 3'-конца: (а) первую нуклео-тидную последовательность, которая включена в CanIon-геномную последовательность; (b) нуклеотид-ную последовательность, включающую позитивный селективный маркер, такой как маркер устойчивости к неомицину (шо); и (с) вторую нуклеотидную последовательность, которая включена в CanIon-геномную последовательность и локализована в геноме справа от первой CanIon-нуклеотидной последовательности (а). - 18 - 008252 В предпочтительном варианте осуществления настоящего изобретения данная ДНК-конструкция включает также негативный селективный маркер, расположенный слева от нуклеотидной последовательности (а) или справа от нуклеотидной последовательности (с). Предпочтительно, если негативный селективный маркер включает ген тимидинкиназы (tk) (Thomas и соавт., 1986), ген бета-гигромицина (Те Ricle и соавт., 1990), hprt-ген (Van der Lught и соавт., 1991; Reid и соавт., 1990) или фрагмент гена дифтерийного токсина A (Dt-A) (Nada и соавт., 1993; Yagi и соавт., 1990). Предпочтительно, если позитивный селективный маркер локализован в экзоне CanIon-последовательности так, что разрывает данную последовательность, кодирующую CanIon-белок. Данные замещающие векторы описаны, например, у Thomas и соавт. (1986; 1987), Mansour и соавт. (1988) и у Koller и соавт. (1992). Безразлично где расположить первую и вторую нуклеотидные последовательности (а) и (с) - в CanIon-регуляторной последовательности, в интронной последовательности, в экзонной последовательности или в последовательности, содержащей и регуляторную и/или интронную и/или экзонную последовательности. Размер нуклеотидных последовательностей (а) и (с) колеблется от 1 до 50 т.п.н., предпочтительно от 1 до 10 т.п.н., более предпочтительно от 2 до 6 т.п.н. и наиболее предпочтительно от 2 до 4 т.п.н. ДНК-конструкции, обеспечивающие гомологичную рекомбинацию: Cre-LoxP-система Такие новые ДНК-конструкции используют в сайт-специфичной рекомбинационной системе фага Р1. Фаг Р1 обладает рекомбиназой, именуемой Сге, которая специфически взаимодействует с 34 парами нуклеотидов loxP-байта. Данный loxP-сайт составлен из двух палиндромных последовательностей из 13 п.н., разделенных 8 п.н. консервативной последовательности (Hoess и соавт., 1986). Рекомбинация между двумя loxP-сайтами, обладающими идентичной ориентацией, с помощью Cre-фермента приводила к удалению ДНК-фрагмента. Cre-loxP-система, используемая в сочетании с методом гомологичной рекомбинации, впервые была описана Gu и соавт. (1993, 1994). Вкратце, представляющую интерес нуклеотидную последовательность встраивают в намеченное место генома, содержащего по меньшей мере два loxP-сайта в одной и той же ориентации и расположенных на соответствующих концах нуклеотидной последовательности, которую вырезают из данного рекомбинантного генома. Данное экцизионное событие требует присутствия в ядре рекомбинантной клетки-хозяина фермента рекомбиназы (Сге). Фермент рекомбиназу можно внести в нужное время либо с помощью (а) инкубации данных рекомбинантных клеток-хозяев в культуральный среде, содержащей данный фермент, с помощью инъекции Cre-фермента прямо в нужную клетку так, как описано у Araki и соавт. (1995), или путем введения данного фермента в клетки-хозяева с помощью ли-посом так, как описано у Baubonies и соавт. (1993); (b) трансфекции клетки-хозяина с помощью вектора, включающего Сге-кодирующую последовательность, функционально присоединенную к функциональному в данной рекобинантной клетке-хозяине промотору, который необязательно является индуцируемым, причем указанный вектор внедряют в данную рекомбинантную клетку-хозяина так, как описано у Gu и соавт. (1993) и у Sauer и соавт. (1988); (с) введения в геном клетки-хозяина полинуклеотида, включающего Сге-кодирующую последовательность, функционально присоединенную к функциональному в данной рекомбинантной клетке-хозяине промотору, который необязательно является индуцируемым, а указанный полинуклеотид встраивается в геном клетки-хозяина либо в результате случайного встраивания, либо в результате гомологичной рекомбинации так, как описано у Gu и соавт. (1994). В конкретном варианте осуществления настоящего изобретения векторную последовательность, содержащую последовательность, которую встраивают в CanIon-ген с помощью гомологичной рекомбинации, конструируют таким образом, чтобы селективные маркеры фланкировались бы loxP-сайтами в одной и той же ориентации в результате обработки Cre-ферментом возможно было осуществить удаление селективных маркеров, оставляя представляющие интерес CanIon-последовательности, которые были встроены с помощью гомологичной рекомбинации. Снова необходимы два селективных маркера: позитивный селективный маркер - для отбора по рекомбинационному событию, и негативный селективный маркер - для отбора по событию гомологичной рекомбинации. Векторы и способы с использованием Cre-loxP-системы описываются у Zou и соавт. (1994). В соответствии с этим третья предпочтительная ДНК-конструкция настоящего изобретения включает, от 5'-конца до 3'-конца: (а) первую нуклеотидную последовательность, которая включается в CanIon-геномную последовательность; (b) нуклеотидную последовательность, включающую полинуклеотид, кодирующий позитивный селективный маркер, причем указанная нуклеотидная последовательность дополнительно включает две последовательности, определяющие сайт, узнаваемый рекомбиназой, такой как loxP-сайт, и данные два сайта размещаются в той же самой ориентации; и (с) вторую нуклеотидную последовательность, которая включается в CanIon-геномную последовательность и локализуется в данном геноме справа от первой CanIon-нуклеотидной последовательности (а). Последовательности, определяющие сайт, узнаваемый рекомбиназой, такой как loxP-сайт, предпочтительно расположены в нуклеотидной последовательности (b) в соответствующих местах, граничащих с нуклеотидной последовательностью, которую необходимо вырезать. В одном из конкретных вариантов осуществления настоящего изобретения два loxP-сайта расположены с каждой стороны последовательности позитивного селективного маркера для того, чтобы обеспечить ее вырезание в нужное время после проявления события гомологичной рекомбинации. - 19 - 008252 В предпочтительном варианте осуществления способа с использованием описанной выше третьей ДНК-конструкции, вырезание полинуклеотидного фрагмента, ограниченного двумя сайтами, узнаваемыми рекомбиназой, предпочтительно двумя ^хР-сайтами, осуществляют в нужный момент, обусловленный наличием в геноме рекомбинантной клетки-хозяина последовательности, кодирующей Cre-фермент, функционально присоединенный к промоторной последовательности, предпочтительно индуцируемому промотору, более предпочтительно к тканеспецифичной промоторной последовательности и наиболее предпочтительно к промоторной последовательности, которая одновременно является и индуцируемой, и тканеспецифичной, как это описано у Gu и соавт. (1994). Наличие Cre-фермента в геноме рекомбинантной клетки-хозяина может явиться следствием скрещивания двух трансгенных животных, где первое трансгенное животное несет представляющую интерес созданную CanIon-последовательность, содержащую описанные выше loxP-сайты, а второе трансгенное животное несет Сге-кодирующую последовательность, функционально присоединенную к соответствующей промоторной последовательности, так как это описано у Gu и соавт. (1994). Пространственно-временной контроль экспрессии Сге-фермента можно также осуществить на основе аденовирусного вектора, который содержит Cre-ген, обеспечивая тем самым заражение клеток или in vivo заражение органов, для доставки Cre-фермента, так как это описано у Anton and Graham (1995) и у Kanegae и соавт. (1995). Описанные выше ДНК-конструкции можно использовать для введения требуемой нуклеотидной последовательности настоящего изобретения, предпочтительно CanIon-геномной последовательности или кДНК-последовательности CanIon, но более предпочтительно - для введения измененной копии геномной или кДНК-последовательности CanIon, в заранее определенное местоположение целевого генома, приводящее либо к образованию измененной копии целевого гена (нокаутная гомологичная рекомбинация), либо к замене копии целевого гена другой копией, достаточно гомологичной, чтобы обеспечить прохождение гомологичной рекомбинации (гомологичная рекомбинация с вбиванием). В конкретном варианте осуществления настоящего изобретения описанные выше ДНК-конструкции могут использоваться для встраивания CanIon-геномной последовательности или кДНК-последовательности CanIon, включающей по меньшей мере один двуаллельный маркер настоящего изобретения, предпочтительно по меньшей мере один двуаллельный маркер, выбранный из группы, состоящей из А1-А17. Ядерные антисмысловые ДНК-конструкции Другие композиции содержат вектор настоящего изобретения, включающий олигонуклеотидный фрагмент нуклеотидной последовательности SEQ ID No 4, предпочтительно фрагмент, включающий старт-кодон CanIon-гена, в качестве антисмыслового средства, которое подавляет экспрессию соответствующего CanIon-гена. Предпочтительные способы с использованием антисмыслового полинуклеотида, в соответствии с настоящим изобретением, представлены методиками, описанными у Sczakiel и соавт. (1995) или методиками, описанными в РСТ-заявке № WO 95/24223, раскрытие которых включено в данное описание путем ссылки во всей их полноте. Предпочтительно, антисмысловые средства выбраны среди полинуклеотидов (длиной 15-200 п.н.), которые комплементарны 5'-концу мРНК CanIon. В одном из вариантов осуществления настоящего изобретения используется сочетание различных антисмысловых полинуклеотидов, комплементарных разным частям требуемого целевого гена. Предпочтительные антисмысловые полинуклеотиды, в соответствии с настоящим изобретением, комплементарны последовательности многих мРНК CanIon, которые содержат либо кодон инициации трансляции ATG, либо сайт сплайсинга. Кроме того, предпочтительные антисмысловые полинуклеоти-ды, в соответствии с настоящим изобретением, комплементарны сайта сплайсинга мРНК CanIon. Предпочтительно, если антисмысловые полинуклеотиды настоящего изобретения обладают 3'-сиг-налом полиаденилирования, который заменен саморасщепляющейся рибозимной последовательностью так, чтобы транскрипты РНК-полимеразы II получались без поли(А) на их 3'-концах, причем эти антисмысловые полинуклеотиды оказываются неспособными выйти из ядра, что описано у Liu и соавт. (1994). В предпочтительном варианте осуществления настоящего изобретения эти CanIon-антисмысло-вые полинуклеотиды включают также, в рибозимной кассете, гистоновую шпилечную структуру, которая стабилизирует расщепляемые транскрипты от 3'-5'-экзонуклеолитической деградации, такая структура описана у Eckner и соавт. (1991). Олигонуклеотидные зонды и праймеры Полинуклеотиды, полученные из CanIon-гена, пригодны для выявления присутствия в тестируемом образце, по меньшей мере, копии нуклеотидной последовательности SEQ ID Nos 1-4 и 6, или ее фрагмента, комплемента, или варианта. Особенно предпочтительные зонды и праймеры настоящего изобретения включают выделенные, выделенные очисткой или рекомбинантные полинуклеотиды, включающие непрерывный участок последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 5000, 10000 или 20000 нуклеотидов SEQ ID Nos 1-3 или их комплементов. Кроме того, предпочтительные зонды и праймеры настоящего изобретения включают выделенные, выделенные очисткой или - 20 - 008252 рекомбинантные полинуклеотиды, в которых указанный непрерывный участок последовательности включает двуаллельный маркер, выбранный из группы, состоящей из А1-А17. Другая цель настоящего изобретения заключается в получении выделенной очисткой, выделенной или рекомбинантной нуклеиновой кислоты, включающей нуклеотидную последовательность SEQ ID No 4, комплементарные ей последовательности, а также аллельные варианты и их фрагменты. Кроме того, предпочтительные зонды и праймеры настоящего изобретения включают выделенную очисткой, выделенную или рекомбинантную CanIon-кДНК, состоящую из, фактически состоящую из или включающую последовательность SEQ ID No 4. Особенно предпочтительные зонды и праймеры настоящего изобретения включают выделенные, выделенные очисткой или рекомбинантные полинуклеотиды, включающие непрерывный участок последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 3000, 4000, 5000 или 6000 нуклеотидов SEQ ID No 4 или их комплементов. Кроме того, предпочтительные зонды и праймеры настоящего изобретения включают выделенные, выделенные очисткой или рекомбинантные полинуклеотиды, включающие непрерывный участок последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 3000, 4000, 5000 или 6000 нуклеотидов SEQ ID No 4 или их комплементов, где указанный непрерывный участок последовательности включает двуаллельный маркер, выбранный из группы, состоящей из А12 и А16. В дополнительных вариантах осуществления настоящего изобретения зонды и праймеры настоящего изобретения включают выделенные, выделенные очисткой или рекомбинантные полинуклеотиды, включающие непрерывный участок последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300 или 400 нуклеотидов SEQ ID No 6 или их комплементов. В предпочтительных вариантах осуществления настоящего изобретения указанный непрерывный участок последовательности SEQ ID No 6 включает двуаллельный маркер А18. Таким образом, настоящее изобретение относится также к зондам нуклеиновых кислот, отличающимся тем, что они специфически гибридизуются в описанных выше жестких условиях с нуклеиновой кислотой, выбранной из группы, состоящей из CanIon-нуклеотидных последовательностей SEQ ID No 13 человека, ее вариантов или последовательности, комплементарной ей. В одном из вариантов осуществления настоящего изобретения включены выделенные, выделенные очисткой и рекомбинантные полинуклеотиды, состоящие или в основном состоящие из непрерывного участка последовательности из 8-50 нуклеотидов какой-либо одной SEQ ID Nos 1-4 или 6, или его комплементов, где указанный участок включает связанный с CanIon двуаллельный маркер в указанной последовательности; где необязательно указанный связанный с CanIon двуаллельный маркер выбран из группы, состоящей из А1-А18, и ее комплементов, или, необязательно, данные двуаллельные маркеры сцеплены при нарушении равновесия. Где, необязательно, указанный участок составляет в длину 18-35 нуклеотидов, а указанный двуаллельный маркер находится в пределах 4 нуклеотидов от центра указанного полинуклеотида; где необязательно указанный полинуклеотид состоит из указанного непрерывного участка последовательности и указанный непрерывный участок последовательности составляет 25 нук-леотидов в длину, а указанный двуаллельный маркер находится в центре указанного полинуклеотида; где необязательно 3'-конец указанного участка непрерывной последовательности представлен на 3'-конце указанного полинуклеотида; и где необязательно 3'-конец указанного непрерывного участка последовательности расположен на 3'-конце указанного полинуклеотида, а указанный двуаллельный маркер представлен на 3'-конце указанного полинуклеотида. В предпочтительном варианте осуществления настоящего изобретения указанные зонды включают, состоят или в основном состоят из последовательности, выбранной из следующих последовательностей: Р1-Р18 и комплементарных им последовательностей. В другом варианте осуществления настоящее изобретение включает в себя выделенные, выделенные очисткой и рекомбинантные полинуклеотиды, включающие, состоящие, в основном состоящие из непрерывного участка последовательности 8-50 нуклеотидов SEQ ID Nos 1-4, или их комплементов, где 3'-конец указанного непрерывного участка последовательности расположен на 3'-конце указанного по-линуклеотида, и где 3'-конец указанного полинуклеотида расположен в пределах 20 нуклеотидов слева от связанного с CanIon двуаллельного маркера в указанной последовательности; где, необязательно, указанный двуаллельный маркер, связанный с CanIon, выбран из группы, состоящей из А1-А18, и их комплементов, или необязательно двуаллельные маркеры, к тому же, сцеплены при нарушении равновесия; где необязательно 3'-конец указанного полинуклеотида расположен в указанной последовательности 1 нуклеотидом "левее" от указанного двуаллельного маркера, связанного с CanIon; и где, необязательно, указанный полинуклеотид состоит, в основном, из последовательности, выбранной из следующих последовательностей: D1-D18 и Е1-Е18. Еще в одном варианте осуществления настоящее изобретение включает в себя выделенные, выделенные очисткой или рекомбинантные полинуклеотиды, включающие, состоящие, в основном состоящие, из последовательности, выбранной из следующих последовательностей: В1-В17 и С1-С17. В дополнительном варианте осуществления настоящее изобретение включает в себя полинуклеоти-ды, используемые в гибридизационном анализе, секвенирующем анализе и в анализе ферментного выявления дефекта комплементарного спаривания для определения нуклеотидной идентичности двуаллель - 21 - 008252 ного маркера, связанного с CanIon, в SEQ ID Nos 1-4 и 6 или в их комплементах, а также полинуклеоти-ды, используемые для амплификации нуклеотидных сегментов, включающих двуаллельный маркер, связанный с CanIon, в SEQ ID Nos 1-4 и 6 или в их комплементах; где, необязательно, указанный двуаллель-ный маркер, связанный с CanIon, выбран из группы, состоящей из А1-А18 и их комплементов, или необязательно двуаллельные маркеры сцеплены к тому же при нарушении равновесия. В настоящем изобретении рассматривается использование полинуклеотидов, в соответствии с настоящим изобретением, для определения нуклеотидной идентичности двуаллельного маркера, связанного с CanIon, предпочтительно в гибридизационном анализе, секвенирующем анализе, микросеквени-рующем анализе или методом ферментного выявления дефекта комплементарного спаривания и амплификацией нуклеотидных сегментов, включающих двуаллельный маркер, связанный с CanIon. Зонд или праймер, в соответствии с настоящим изобретением, обладает около 8-1000 нуклеотидов в длину или задан по крайней мере 12, 15, 18, 20, 25, 35, 40, 50, 60, 70, 80, 100, 250, 500 или 1000 нуклеоти-дами в длину. Более предпочтительно, длина таких зондов и праймеров может колебаться от 8, 10, 15, 20 или 30-100 нуклеотидов, предпочтительно от 10 до 50, более предпочтительно от 15 до 30 нуклеотидов. У коротких зондов и праймеров обычно недостаточно специфичности для целевой последовательности нуклеиновой кислоты и, как правило, нужны пониженные температуры для образования достаточно стабильных гибридных комплексов с матрицей. Более длинные зонды и праймеры являются дорогостоящими в изготовлении и могут иногда самогибридизоваться с образованием шпилечных структур. Надлежащую длину для праймеров и зондов под конкретный комплекс аналитических условий может эмпирически определить специалист в данной области техники. Предпочтительный зонд или прай-мер состоит из нуклеиновой кислоты, включающей полинуклеотид, выбранный из группы нуклеотидных последовательностей Р1-Р18 и их комплементарных последовательностей, В1-В17, С1-С17, D1-D18, Е1-Е18, для которых соответственные местоположения в списке последовательностей представлены в табл. 1, 2 и 3. Образование стабильных гибридов зависит от температуры плавления (Тт) ДНК. Тт зависит от длины праймера или зонда, ионной силы данного раствора и содержания G+C. Чем выше содержание G+C праймера или зонда, тем выше температура плавления, потому что G:C пары удерживаются тремя Н-связями, тогда как А:Т пары удерживаются только двумя такими связями. GC-содержание в зондах настоящего изобретения, как правило, колеблется между 10 и 75%, предпочтительно между 35 и 60%, и наиболее предпочтительно между 40 и 55%. Праймеры и зонды можно изготовить любым подходящим способом, включая, например, клонирование и рестрикцию соответствующих последовательностей, а также способом прямого химического синтеза, например, таким как фосфодиэфирный способ Narang и соавт. (1979), фосфодиэфирный способ Brown и соавт. (1979), диэтилфосфоамидитный способ Beaucage и соавт. (1981) и твердофазный способ, описанный в ЕР 0707592. Зонды для детекции представляют собой, как правило, последовательности нуклеиновых кислот или незаряженные аналоги нуклеиновых кислот, такие, например, как пептидонуклеиновые кислоты, которые раскрыты в международной патентной заявке WO 92/20702, морфолиновые аналоги, которые описаны в патентах США №№ 5185444; 5034506 и 5142047. Зонд может оказаться "не удлиняемым", поскольку дополнительные dNTP невозможно добавить к данному зонду. Внутри себя аналоги, как правило, не удлиняемы, и зонды нуклеиновых кислот могут оказываться неудлиняемыми из-за модификации 3'-конца данного зонда, так что гидроксильная группа уже не способна участвовать в элонгации. Например, 3'-конец данного зонда может быть функционализирован захватом или детекцией метки, чтобы таким образом истратить или иначе блокировать данную гидроксильную группу. С другой стороны, 3'-гидроксильная группа может попросту отщепиться, заместиться или модифицироваться, патентная заявка США, порядковый № 07/049061, поданная 19 апреля 1993 г., описывает модификации, которые могут использоваться для придания зонду неудлиняемости. Любой полинуклеотид настоящего изобретения можно пометить при желании путем включения любой метки, известной в данной области техники, которая обнаруживается спектроскопическим, фотохимическим, биохимическим, иммунохимическим или химическим способом. Например, используемые 32 35 3 125 метки включают радиоактивные вещества (в том числе, Р32, S35, Н3, I125), флуоресцентные красители (в том числе, 5-бромдезоксиуридин, флуоресцеин, ацетиламинофтор, дигоксигенин) или биотин. Предпочтительно, полинуклеотиды метят по их 3'- и 5'-концам. Примеры нерадиоактивного мечения фрагментов нуклеиновых кислот описаны во французском патенте № FR-7810975 или у Urdea и соавт. (1988), или у Sanchez-Pescador и соавт. (1988). Кроме того, зонды, в соответствии с настоящим изобретением, могут иметь структурные характеристики, которые делают возможным усиление сигнала, причем такими структурными характеристиками являются, например, разветвленные ДНК-зонды, как те, что описаны у Urdea и соавт. (1991) или в европейском патенте № ЕР0225807 (Chiron). Метку можно также использовать для захвата данного праймера, с тем, чтобы обеспечить иммобилизацию на твердом носителе либо данного праймера, либо удлиненного продукта, такого как амплифи-цированная ДНК. Введенная метка присоединяется к праймерам или к зондам и может представлять собой связывающий член, который образует связывающую пару вместе со специфичным связывающим - 22 - 008252 членом твердофазного реагента (например, биотин и стрептавидин). Поэтому, в зависимости от типа метки, переносимой полинуклеотидом или зондом, ее можно использовать для улавливания или для обнаружения ДНК-мишени. Кроме того, следует иметь в виду, что созданные в данном описании полинук-леотиды, праймеры или зонды могут, сами по себе, служить в качестве захватной метки. Например, в том случае, когда связывающий член твердофазного реагента представляет собой последовательность нуклеиновой кислоты, его можно выбрать так, чтобы он связывал комплементарную часть праймера или зонда с тем, чтобы иммобилизовать данный праймер или зонд на данную твердую фазу. В случае если полинуклеотидный зонд сам служит в качестве связывающего члена, специалисты должны знать, что данный зонд будет содержать последовательность или "хвост", который не комплементарен данной мишени. В том случае, если полинуклеотидный праймер сам служит в качестве захватной метки, по меньшей мере, часть данного праймера будет свободна для гибридизации с нуклеиновой кислотой на твердой фазе. Методы меченья ДНК хорошо известны квалифицированным специалистам. Зонды настоящего изобретения пригодны для многих целей. Они могут, в частности, использоваться в Саузерн-гибридизации с геномной ДНК. Зонды могут также использоваться для обнаружения продуктов ПЦР-амплификации. При использовании других методик они могут также использоваться для обнаружения несоответствий в CanIon-гене или мРНК. Они могут также использоваться для обнаружения экспрессии CanIon-гена, например, в Нозерн-блоте. Любой полинуклеотид, праймер или зонд настоящего изобретения можно удобно иммобилизовать на твердый носитель. Твердые носители хорошо известны специалистам в данной области техники и включают стенки лунок реакционной кюветы, пробирки, шарики из полистирола, магнитные шарики, полоски нитроцеллюлозы, мембраны, микрочастицы, такие как латексные частицы, эритроциты барана (или другого животного), duracytes и другие. Твердый носитель не является ключевым и его может выбрать специалист в данной области техники. Таким образом, латексные частицы, микрочастицы, намагниченные и ненамагниченные шарики, мембраны, пластмассовые пробирки, стенки лунок для микротитрования, стеклянное или силиконовое крошево, эритроциты барана (или иных подходящих животных) и duracytes (мозговые оболочки), все, представляют собой подходящие модели. Соответствующие способы иммобилизации нуклеиновых кислот на твердых фазах включают ионное, гидрофобное, ковалентное взаимодействие и т.п. Используемый в данном описании твердый носитель относится к любому нерастворимому материалу, или его можно сделать нерастворимым в последующей реакции. Твердый носитель можно выбрать в соответствии с его способностью притягивать и иммобилизовать захватный реагент. Или же, данная твердая фаза может удерживать дополнительный рецептор, который обладает способностью притягивать и иммобилизовать захватный реагент. Дополнительный рецептор может включать заряженное вещество, которое противоположно заряжено по отношению к собственно захватному реагенту или к заряженному веществу, конъюгированному с данным захватным реагентом. Согласно еще одной альтернативе, рецепторная молекула может представлять собой любой специфический связывающий член, который иммобилизуется на (присоединяется к) твердом носителе, и который обладает способностью иммобилизовать захватный реагент посредством специфичной реакции связывания. Рецеп-торная молекула делает возможным непрямое связывание захватного реагента с материалом твердого носителя перед осуществлением анализа или в течение осуществления данного анализа. Твердая фаза, таким образом, может быть представлена пластмассой, производными пластмассы, намагниченным или ненамагниченным металлом, стеклянной или силиконовой поверхностью пробирки, лункой для микротитрования, полоской, шариком, микрочастицей, крошкой, эритроцитами барана (или иных подходящих животных), duracytes(r) и другими формами, хорошо известными среднему специалисту в данной области техники. Полинуклеотиды настоящего изобретения можно присоединять или иммобилизовать на твердом носителе индивидуально или группами по меньшей мере из 2, 5, 8, 10, 12, 15, 20 или 25 разных по-линуклеотидов настоящего изобретения с единственным твердым носителем. Кроме того, полинуклеоти-ды, отличающиеся от полинуклеотидов настоящего изобретения, можно присоединять к тому же твердому носителю, к которому присоединены один или несколько полинуклеотидов настоящего изобретения. Следовательно, настоящее изобретение включает также способ для обнаружения присутствующей нуклеиновой кислоты, включающей нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID Nos 1-4 и 6, ее фрагмента или варианта, и комплементарную ей последовательность, в образце, причем указанный способ включает следующие стадии: a) приведение в контакт зонда нуклеиновой кислоты или совокупности зондов нуклеиновой кислоты, которые могут гибридизоваться с нуклеотидной последовательностью, включенной в нуклеиновую кислоту, выбранную из группы, состоящей из нуклеотидных последовательностей SEQ ID Nos 1-4 и 6, их фрагмента или варианта, и комплементарную ей последовательность, и анализируемого образца; и b) выявление гибридного комплекса, образованного между зондом и нуклеиновой кислотой данного образца. Кроме того, в настоящем изобретении рассматривается набор для детектирования присутствующей нуклеиновой кислоты, включающий нуклеотидную последовательность, выбранную из группы, состоящей SEQ ID Nos 1-4 и 6, ее фрагмента или варианта, и комплементарную ей последовательность, в образце, причем указанный набор включает: - 23 - 008252 a) зонд нуклеиновой кислоты или множество данных зондов нуклеиновой кислоты, которые могут гибридизоваться с нуклеотидной последовательностью, включенной в нуклеиновую кислоту, выбранную из группы, состоящей из нуклеотидных последовательностей SEQ ID Nos 1-4 и 6, ее фрагмента или варианта, и комплементарной ей последовательности; и b) необязательно, реагенты, необходимые для осуществления реакции гибридизации. В первом предпочтительном варианте осуществления настоящего изобретения данный способ детекции и набор отличаются тем, что указанный зонд нуклеиновой кислоты или множество данных зондов нуклеиновой кислоты метят с помощью детектируемой молекулы. В другом предпочтительном варианте осуществления настоящего изобретения указанный способ и набор отличаются тем, что указанный зонд нуклеиновой кислоты или множество данных зондов нуклеиновой кислоты были иммобилизованы на субстрате. В третьем предпочтительном варианте осуществления настоящего изобретения данный зонд нуклеиновой кислоты или множество данных зондов нуклеиновой кислоты включают либо последовательность, которая выбрана из группы, состоящей из нуклеотидных последовательностей Р1-Р18 и комплементарной им последовательности, В1-В17, С1-С17, D1-D18, Е1-Е18, либо двуаллельный маркер, выбранный из группы, состоящей из А1-А18 и их комплементов. Олигонуклеотидные наборы Любой субстрат, включающий множество олигонуклеотидных праймеров или зондов настоящего изобретения, может использоваться либо для детекции или амплификации последовательностей-мишеней CanIon-гена, либо может также использоваться для детекции мутаций в кодирующих и некоди-рующих последовательностях CanIon-гена. Любой созданный в данном описании полинуклеотид можно присоединить в области перекрывания или в случайные места данного твердого носителя. Альтернативно, полинуклеотиды настоящего изобретения можно присоединить в виде упорядоченного набора, где каждый полинуклеотид присоединяется к определенной области твердого носителя, которая не перекрывается с участком присоединения любого другого полинуклеотида. Предпочтительно, такой упорядоченный набор полинуклеотидов создают "адресным", где отдельные участки регистрируются и могут быть доступны как часть аналитического метода. Адресные полинуклеотидные наборы обычно включают множество различных олигонуклеотидных зондов, которые связаны с поверхностью субстрата в разных известных местах. Знание точного расположения каждого полинуклеотида делает эти "адресные" наборы особенно пригодными для методов гибридизации. Любая технология адресного набора, известная в данной области техники, может использоваться для полинуклеотидов настоящего изобретения. Один из конкретных вариантов осуществления данных полинуклеотидных наборов известен в качестве Genechips(tm), и в основном описан в патенте США №5143854; публикациях РСТ WO 90/15070 и 92/10092. Данные наборы обычно можно получить с использованием методов автоматизированного синтеза или методов светонаправленного синтеза, которые включают сочетание фотолитографических способов и твердофазного олигонуклеотидного синтеза (Fodor и соавт., 1991). Иммобилизация наборов олигонуклеотидов на твердых носителях представляется возможной благодаря развитию технологии, обычно идентифицируемой как "Very Large Scale Immobilized Polymer Synthesis" (Крупномасштабный синтез иммобилизованного полимера) (VLSIPS(tm)), в которой, как правило, зонды иммобилизуются в виде набора с высокой плотностью на твердой поверхности чипа. Примеры технологий VLSIPS(tm) представлены в патентах США №№ 5143854; и 5412087, а также в публикациях РСТ WO 90/15070, WO 92/10092 и WO 95/11995, в которых описываются способы создания олигонуклеотидных наборов с помощью таких методов, как методы по светонаправленному синтезу. В разработанных методиках, предназначенных для создания наборов нуклеотидов, иммобилизованных на твердых носителях, дополнительно были разработаны методики представления для приведения в порядок и представления олигонуклеотидных наборов на чипах в попытке извлечь максимальную пользу из паттернов гибридизации и информации о последовательности. Примеры таких методик представления раскрыты в публикациях РСТ WO 94/12305, WO 94/11530, WO 97/29212 и WO 97/31256, раскрытие которых включено путем ссылки во всей их полноте. В другом варианте осуществления олигонуклеотидных наборов настоящего изобретения матрица олигонуклеотидного зонда может преимущественно использоваться для обнаружения мутаций, встречаемых в CanIon-гене и предпочтительно в его регуляторной области. Для этой конкретной цели специально создают зонды, которые обладают нуклеотидной последовательностью, обеспечивающей их гибридизацию с генами, которые несут известные мутации (в результате делеции, вставки или замены одного или нескольких нуклеотидов). Для известных мутаций это означает, что мутации в CanIon-гене были идентифицированы в соответствии, например, с методом, используемым Huang и соавт. (1996) и Samson и соавт. (1996). В другом методе, который используется для обнаружения мутаций в CanIon-гене, используют набор ДНК высокой плотности. Каждый олигонуклеотидный зонд, составляющий единичный элемент набора ДНК высокой плотности, создается для сопоставления специфичной субпоследовательности геномной ДНК или кДНК CanIon. Таким образом, набор, состоящий из олигонуклеотидов, комплементарных субпоследовательностям из последовательности гена-мишени, используют для определения идентичности целевой последовательности с последовательностью природного гена, измерения его количества, обна - 24 - 008252 ружения различий между целевой последовательностью и стандартной последовательностью природного гена из CanIon-гена. Одна такая конструкция, названная 4L-"плиточный" набор (4L tiled array), представляет собой группу из четырех зондов (А, С, G, Т), предпочтительно 15-нуклеотидных олигомеров. В каждой группе из четырех зондов полный комплемент будет гибридизоваться более интенсивно, чем комплементарно несогласованные зонды. Следовательно, целевая нуклеиновая кислота длиной L сканируется по мутациям с помощью "плиточного" набора, содержащего 4L-зонды, причем весь набор зондов содержит все возможные мутации в известной стандартной природной последовательности. Гибридизаци-онные сигналы группы 15-мерного зонда "плиточного" набора нарушаются из-за единственной замены основания в целевой последовательности. Вследствие этого происходит характерная утрата сигнала или " футпринта" для зондов, фланкирующих положение мутации. Данный метод описан Chee и соавт. (1996). Следовательно, в настоящем изобретении рассматривается набор молекул нуклеиновых кислот, включающих, по меньшей мере, один полинуклеотид, описанный выше в качестве зондов и праймеров. Предпочтительно, в настоящем изобретении рассматривается набор нуклеиновой кислоты, включающей, по меньшей мере, два полинуклеотида, описанных выше в качестве зондов и праймеров. Дополнительная цель настоящего изобретения заключается в создании набора последовательностей нуклеиновых кислот, включающих, по меньшей мере, одну из последовательностей, выбранную из группы, состоящей из Р1-Р18, В1-В17, С1-С17, D1-D18, Е1-Е18, комплементарных ей последовательностей, ее фрагментов по меньшей мере из 8, 10, 12, 15, 18, 20, 25, 30 или 40 последовательных нуклеотидов либо, по меньшей мере, одну последовательность, включающую двуаллельный маркер, выбранный из группы, состоящей из А1-А18, и его комплементов. Настоящее изобретение относится также к набору последовательностей нуклеиновых кислот, включающих либо, по меньшей мере, две последовательности, выбранные из группы, состоящей из Р1-Р18, В1-В17, С1-С17, D1-D18, Е1-Е18, комплементарных ей последовательностей, ее фрагмента, по меньшей мере, из 8 последовательных нуклеотидов, либо по меньшей мере две последовательности, включающие двуаллельный маркер, выбранный из группы, состоящей из А1-А18, и его комплементов. CanIon-белки и полипептидные фрагменты Термин "CanIon-полипептиды" используется в данном описании для охвата всех белков и полипептидов настоящего изобретения. Кроме того, образующей частью настоящего изобретения являются полипептиды, кодируемые полинуклеотидами настоящего изобретения, а также слитые полипептиды, включающие такие полипептиды. В настоящем изобретении содержатся CanIon-белки людей, включая выделенные или выделенные очисткой CanIon-белки, состоящие, состоящие в основном или включающие последовательность SEQ ID No 5. В настоящем изобретении рассматривается полипептид, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID No 1-4 и 6, комплементарной ей последовательностью или ее фрагментом. В настоящем изобретении содержатся выделенные, выделенные очисткой и рекомбинантные полипептиды, включающие непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50, 100, 150, 200, 300, 400, 500, 700, 1000, 1200, 1400, 1600 или 1700 аминокислот SEQ ID No 5. В других предпочтительных вариантах осуществления настоящего изобретения непрерывный участок из аминокислот включает участок с мутацией или функциональной мутацией, включающий деле-цию, добавку, перестановку, укороченные аминокислоты в белковой последовательности CanIon. В предпочтительных вариантах осуществления настоящего изобретения, настоящее изобретение содержит выделенные, выделенные очисткой и рекомбинантные полипептиды, включающие непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно, по меньшей мере, 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50, 100, 150, 200, 300, 400, 500, 700, 1000, 1200, 1400, 1600 или 1700 аминокислот SEQ ID No 5, где непрерывный участок включает по меньшей мере 1, 2, 3, 5 или 10 аминокислотных позиций 277, 338, 574, 678, 680, 683, 691, 692, 695, 696, 697, 894, 1480, 1481, 1483, 1484, 1485, 1630, 1631, 1632, 1636, 1660, 1667, 1707, 1709 из SEQ ID No 5. Предпочтительно, указанный непрерывный участок SEQ ID No 5 включает остаток Аланина в положении 277; Серин в положении 338; Валин в положении 574; Лейцин в положении 678; Серин в положении 680; Треонин в положении 683; Гистидин в положении 691; Серин в положении 692; Серин в положении 695; Аланин в положении 696; Изолейцин в положении 697; Изолейцин в положении 894; Лизин в положении 1480; Аргинин в положении 1481; Глицин в положении 1483; Валин в положении 14 84; Изолейцин в положении 1485; Аспарагин в положении 1630; Серин в положении 1631; Метионин в положении 1632; Треонин в положении 1636; Аланин в положении 1660; Фенилаланин в положении 1667; Треонин в положении 1707; и/или Аланин в положении 1709. Обеспечены также полинуклеотиды, кодирующие любой из этих полипептидов. Настоящее изобретение включает также в себя выделенные очисткой, выделенные или рекомби-нантные полипептиды, включающие аминокислотную последовательность, обладающую, по меньшей мере, 70, 75, 80, 85, 90, 95, 98 или 99% аминокислотной идентичностью с аминокислотной последовательностью SEQ ID No 5 или с ее фрагментом. - 25 - 008252 CanIon-белки предпочтительно выделяют из тканевых образцов человека или млекопитающих или экспрессируют из генов человека или млекопитающих. CanIon-полипептиды настоящего изобретения могут быть получены с использованием стандартных способов экспрессии, известных в данной области техники. Полинуклеотид, кодирующий требуемый полипептид, лигируют в экспрессионный вектор, приемлемый для любого удобного хозяина. Эукариотическая и прокариотическая хозяйская система, та и другая, используются для создания рекомбинантных полипептидов, при этом кратко излагаются некоторые наиболее распространенные системы. Затем полипептид выделяют из лизированных клеток или из культуральной среды и выделяют очисткой до степени, необходимой для его предполагаемого использования. Очистку осуществляют с помощью любого метода, известного в данной области техники, например, дифференциальным экстрагированием, солевым фракционированием, хроматографией, центрифугированием и т.п. См., например, Methods in Enzymology для разнообразных способов по выделению очисткой белков. Кроме того, короткие белковые фрагменты получают химическим синтезом. Альтернативно, белки настоящего изобретения экстрагируют из клеток или тканей людей или животных, не принадлежащих человеческому роду. Способы выделения очисткой белков известны в данной области техники и включают использование детергентов или хаотропных агентов, которые дезинтегрируют (разобщают) частицы, за которым следует дифференциальное экстрагирование и разделение полипептидов с помощью ионообменной хроматографии, хроматографии по сродству, осаждения в соответствии с плотностью и гель-электрофореза. Любая кДНК CanIon, включая SEQ ID No 4, может использоваться для экспрессии CanIon-белков и полипептидов. Нуклеиновую кислоту, кодирующую экспрессируемый CanIon-белок или полипептид, функционально присоединяют к промотору в экспрессионном векторе с использованием традиционной техники клонирования. CanIon-вставка в данный экспрессионный вектор может включать полную кодирующую последовательность для CanIon-белка или его части. Экспрессионный вектор представляет собой любую экспрессионную систему млекопитающего, дрожжей, насекомого или бактерии, известную в данной области техники. Коммерчески доступные векторы и экспрессионные системы можно получить от разных поставщиков, включая Genetics Institute (Cambridge, MA), Stratagene (La Jolla, California), Promega (Madison, Wisconsin) и Invitrogen (San Diego, California). При желании, для усиления экспрессии и обеспечения надлежащей упаковки белка, для конкретного экспрессионного организма, в который интродуцирован экспрессионный вектор, оптимизируют кодоновый фон и спаривание кодонов данной последовательности, в соответствии с разъяснениями Hat-field и соавт., патент США № 5082767, раскрытие которого включено в данное описание путем ссылки во всей его полноте. В одном из вариантов осуществления настоящего изобретения полную кодирующую последовательность кДНК CanIon, включая полиА-сигнал кДНК, функционально присоединяют к промотору данного экспрессионного вектора. Альтернативно, если нуклеиновая кислота кодирует часть CanIon-белка, в котором отсутствует метионин, служащий в качестве сайта инициации, инициирующий метионин можно встроить рядом с первым кодоном данной нуклеиновой кислоты с использованием традиционных методов. Также, если в данной вставке из кДНК CanIon отсутствует полиА-сигнал, данную последовательность можно добавить в конструкцию, например, путем вырезания полиА-сигнала из pSG5 (Stratagene) с использованием эндонуклеаз рестрикции BglI и SalI и встраивания в экспрессионный вектор pXT1 (Stratagene) млекопитающего. pXT1 содержит LTR и часть gag-гена вируса лейкоза мышей Молони. Положение LTR в данной конструкции делает возможной эффективную стабильную трансфекцию. Данный вектор включает тимидинкиназный промотор вируса простого герпеса и селективный ген неомицина. Нуклеиновую кислоту, кодирующую CanIon-белок или его часть, получают с помощью ПЦР из бактериального вектора, содержащего CanIon-кДНК SEQ ID No 5 с использованием олигонуклеотидных праймеров, комплементарных CanIon-кДНК или ее части и содержащих последовательность для рестрициро-ванной эндонуклеазы PstI, встраиваемую в 5'-праймер, и BglII-последовательность, встраиваемую в 5'-конец соответствующего 3'-праймера кДНК, обеспечивая данной последовательности, кодирующей CanIon-белок или его часть, надлежащее расположение относительно полиА-сигнала. Выделенный очисткой фрагмент, полученный в окончательной ПЦР-реакции, расщепляют с помощью PstI, затупляют по концам нуклеазой, расщепляют с помощью BglII, выделяют очисткой и лигируют с ХТ1, содержащем по-лиА-сигнал и расщепляют с помощью BglII. Данный лигированный продукт можно трансфицировать в мышиные №Н3Т3-клетки с использованием Lipofectin (Life Technologies, Inc., Grand Island, New York) в условиях, вкратце изложенных в описании к данному продукту. Позитивные трансфектанты отбирают после выращивания трансфицирован-ных клеток в 600 ug/мл G418 (Sigma, St. Louis, Missouri). Приведенные выше протоколы можно также использовать для экспрессии мутантного CanIon-белка, ответственного за обнаруживаемый фенотип, или его части. Экспрессированный белок можно выделить очисткой с использованием традиционных методов очистки, таких как осаждение сульфатом аммония или хроматографическим разделением по размеру или заряду. Белок, кодируемый встроенной нуклеиновой кислотой, можно также выделить очисткой с ис - 26 - 008252 пользованием стандартных иммунохроматографических методов. В таких протоколах раствор, такой как клеточный экстракт, содержащий экспрессированный CanIon-белок или его часть, наносят на колонку, содержащую антитела к CanIon-белку или его части, которые присоединяют к хроматографическому матриксу. Экспрессированному белку дают возможность связаться иммунохроматографической колонкой. Затем колонку промывают для удаления неспецифически связавшихся белков. После чего специфически связавшийся экспрессированный белок высвобождают из колонки и извлекают с использованием стандартных методов. Для подтверждения экспрессии CaIon-белка или его части, белки, экспрессированные из клеток-хозяев, содержащих экспрессионный вектор, содержащий вставку, кодирующую CanIon-белок или его часть, можно сравнить с белками, экспрессированными из клеток-хозяев, содержащих экспрессионный вектор без вставки. Наличие полосы в образцах клеток, содержащих экспрессионный вектор со вставкой, которая отсутствует в образцах из клеток, содержащих экспрессионный вектор без вставки, свидетельствует о том, что CanIon-белок или его часть экспрессируются. Как правило, данная полоса будет обладать подвижностью, ожидаемой для CanIon-белка или его части. Однако данная полоса может обладать подвижностью, отличающейся от ожидаемой из-за модификаций, таких как гликозилирование, убихитини-зиция или ферментативное расщепление. Антитела, способные специфически распознавать экспрессированный CanIon-белок или его часть, описаны ниже. Когда образование антител невозможно, нуклеиновые кислоты, кодирующие CanIon-белок или его часть, встраиваются в экспрессионный вектор, созданный для использования в схемах очистки, использующих химерные полипептиды. В таких методиках нуклеиновая кислота, кодирующая CanIon-белок или его часть, встраивается в открытую рамку считывания вместе с геном, кодирующим другую половину данной химеры. Другая половина химеры представляет собой последовательность в-глобина или последовательность, кодирующую полипептид, связывающий никель. Хроматографический матрикс, содержащий антитело к в-глобину или присоединенный к нему никель, используется для выделения очисткой химерного белка. Сайты протеазного расщепления создают генно-инженерным способом между геном в-глобина или никельсвязывающим полипептидом и CanIon-белком или его частью. Таким образом, два полипептида химеры отделяются друг от друга с помощью протеазного расщепления. Пригодным экспрессионным вектором для образования в-глобиновых химерных белков является pSG5 (Stratagene), который кодирует в-глобин кролика. Интрон II кроличьего гена в-глобин обеспечивает сплайсинг экспрессируемого транскрипта, а сигнал полиаденилирования, встраиваемый в данную конструкцию, повышает уровень экспрессии. Эти методы хорошо известны в данной области молекулярной биологии. Стандартные способы опубликованы в руководствах, как например, Davies и соавт. (1986), но многие способы доступны от Stratagene, Life Technologies, Inc., или Promega. Полипептид можно дополнительно получить из конструкции с использованием in vivo трансляционных систем, таких как In vitro Express(tm) Translation Kit (Stratagene). Антитела, которые связывают CanIon-полипептиды настоящего изобретения Любой CanIon-полипептид или целый белок можно использовать для получения антител, способных специфически связываться с экспрессируемым CanIon-белком или его фрагментами, которые описаны. Антительная композиция настоящего изобретения способна специфически или селективно связываться с вариантом CanIon-белка SEQ ID No 5. Для антительной композиции, которая специфически связывается с первым вариантом CanIon, необходимо продемонстрировать в ELISA, RIA или ином анализе по связыванию антител, по меньшей мере, на 5, 10, 15, 20, 25, 50 или 100% более высокое связывание по сродству для полноразмерного первого варианта CanIon-белка, чем для полноразмерного второго варианта CanIon-белка. В предпочтительном варианте осуществления настоящего изобретения антительная композиция способна специфически связывать CanIon-белок человека. В предпочтительном варианте осуществления настоящего изобретения рассматриваются антительные композиции, поликлональные или моноклональные, способные селективно связывать или селективно связываться с эпитопсодержащим полипептидом, включающим непрерывный участок последовательности, по меньшей мере, из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно, по меньшей мере, 12, 15, 20, 25, 30, 40, 50, 100, 200, 300, 400, 500, 700 или 1000 аминокислот SEQ ID No 5. В предпочтительных вариантах осуществления настоящего изобретения указанный непрерывный участок последовательности включает, по меньшей мере, 1, 2, 3, 5 или 10 аминокислотных позиций 277, 338, 574, 678, 680, 683, 691, 692, 695, 696, 697, 894, 1480, 1481, 1483, 1484, 1485, 1630, 1631, 1632, 1636, 1660, 1667, 1707, 1709 SEQ ID No 5. Любой CanIon-полипептид или целый белок можно использовать для получения антител, способных, как описано, специфически связываться с экспрессируемым CanIon-белком или его фрагментами. Эпитоп может включать не менее 3 аминокислот для пространственной конформации, которая является уникальной для данного эпитопа. Как правило, эпитоп состоит, по меньшей мере, из 6 таких аминокислот, но чаще, по меньшей мере, из 8-10 таких аминокислот. В предпочтительном варианте осуществления настоящего изобретения антигенные эпитопы включают ряд аминокислот, которые составляют - 27 - 008252 любое целое число между 3 и 50. Фрагменты, которые функционируют в качестве эпитопов, можно получить с помощью любого традиционного способа. Эпитопы можно определить в антигеном анализе Jameson-Wolf, например, применяя компьютерную программу PROTEAN с использованием значений параметров по умолчанию (Version 4.0 Windows, DNASTAR, Inc., 1228 South Park Street Madison, WI). В настоящем изобретении рассматривается также выделенное очисткой или выделенное антитело, способное специфически связаться с мутантным CanIon-белком, его фрагментом или вариантом, включающим эпитоп мутантного CanIon-белка. В другом предпочтительном варианте осуществления настоящего изобретения рассматривается антитело, способное связаться с полипептидом, включающим, по меньшей мере, 10 последовательных аминокислот CanIon-белка и включающего, по меньшей мере, одну из аминокислот, которая может кодироваться мутациями, определяющими данный признак. Животные или млекопитающие, не принадлежащие человеческому роду, природные или трансгенные, которые экспрессируют разные виды CanIon, отличные от CanIon, к которому требуется связывание антитела, а также животные, которые не экспрессируют CanIon (т.е. CanIon-нокаутное животное, которое описано в данном описании), особенно пригодны для получения антител. CanIon-нокаутные животные будут распознавать все или большинство находящихся на поверхности участков CanIon-белка в качестве чужеродных антигенов и, вследствие этого, производить антитела с более широким набором CanIon-эпитопов. Кроме того, меньшие полипептиды, всего лишь с 10-30 аминокислотами, могут быть пригодны для получения специфичного связывания с одним из CanIon-белков. К тому же, гуморальная иммунная система животных, которая продуцируют виды CanIon, имеющие сходство с антигенной последовательностью, будет предпочтительно распознавать различия между нативными CanIon-видами животного и данной антигенной последовательностью, и продуцировать антитела к этим уникальным сайтам в последовательности данного антигена. Такой метод особенно пригоден для получения антител, которые специфически связываются с любым отдельным CanIon-белком. Препараты антител, полученные в соответствии с любым протоколом, пригодны для количественного иммуноанализа, в котором определяют концентрации несущих антиген веществ в биологических образцах; их также используют, полуколичественно или количественно, для идентификации антигена, присутствующего в биологическом образце. Антитела можно также использовать в терапевтических композициях для уничтожения клеток, экспрессирующих данный белок или для снижения уровня данного белка в данном организме. Антитела настоящего изобретения можно метить радиоактивной, флуоресцентной или ферментной меткой, известной в данной области техники. Поэтому, настоящее изобретение относится также к способу детектирования специфически присутствующего в биологическом образце CanIon-полипептида в соответствии с настоящим изобретением, причем указанный способ включает следующие стадии: a) приведения в контакт данного биологического образца с поликлональным или моноклональным антителом, которое специфически связывает CanIon-полипептид, включающий аминокислотную последовательность SEQ ID No 5, или его пептидный фрагмент или вариант; и b) детектирования образованного комплекса антиген-антитело. В настоящем изобретении рассматривается также диагностический набор для детектирования в биологическом образце, в соответствии с настоящим изобретением, наличия in vitro CanIon-полипептида, где указанный набор включает: a) поликлональное или моноклональное антитело, которое специфически связывает CanIon-полипептид, включающий аминокислотную последовательность SEQ ID No 5, или его пептидный фрагмент или вариант, необязательно меченый; b) реагент, обеспечивающий детекцию образованных комплексов антиген-антитело, причем указанный реагент необязательно несет метку или способен распознавать самого себя с помощью меченного реагента, в частности, в случае, если указанное выше моноклональное или поликлональное антитело само не является меченым. Настоящее изобретение относится, таким образом, к антителам и Т-клеточным антигенным рецепторам (TCR), которые специфически связывают полипептиды и, в частности, эпитопы полипептидов настоящего изобретения, включая, но, не ограничиваясь ими, IgG (в том числе IgG1, IgG2, IgG3 и IgG4), IgA (в том числе IgA1 и IgA2), IgD, IgE или IgM и IgY. В предпочтительном варианте осуществления настоящего изобретения данные антитела, представляющие собой антительные фрагменты настоящего изобретения, связывающие антиген человека, включают, но не ограничиваются ими, Fab, Fab', F(ab)2 и F(ab')2, Fd, одноцепочечные Fvs (scFv), одноцепочечные антитела, связанные дисульфидной связью Fvs (sdFv) и фрагменты, включающие домен VL или VH. Антитела можно получить у любого животного, в том числе у птиц и млекопитающих. Предпочтительны антитела человека, мыши, кролика, козы, морской свинки, верблюда, лошади или обезьяны. Антигенсвязывающие антительные фрагменты, включая одноцепочечные антитела, могут включать вариабельную область(и), отдельно или в сочетании с полной или частичной: шарнирной областью, доменами CH1, CH2 и СН3. В настоящее изобретение включены также любые сочетания вариабельной об-ласти(ей) и шарнирной области, доменов CH1, CH2 и СН3. Кроме того, настоящее изобретение включает - 28 - 008252 химерные, гуманизированные, а также моноклональные и поликлональные антитела человека, которые специфически связывают полипептиды настоящего изобретения. Настоящее изобретение к тому же включает антитела, которые являются антиидиотипическими к антителам настоящего изобретения. Антитела настоящего изобретения могут быть моноспецифичными, биспецифичными, триспеци-фичными или обладать более высокой мультиспецифичностью. Мультиспецифичные антитела могут быть специфичны для разных эпитопов полипептида настоящего изобретения или могут быть специфичны для полипептида настоящего изобретения, а также для гетерологичных композиций, таких как гете-рологичный полипептид или вещество твердого носителя. См., например, WO 93/17715; WO 92/08802; WO 91/00360; WO 92/05793; Tutt и соавт. (1991) J. Immunol. 147:60-69; патенты №№ 5573920, 4474893, 5601819, 4714681, 4925648; Kostelny и соавт. (1992) J. Imminol. 148:1547-1553. Антитела настоящего изобретения могут быть описаны или охарактеризованы в пределах эпито-па(ов) или эпитоп-несущей части(ей) полипептида настоящего изобретения, которые распознаются или специфически связываются с данным антителом. В случае белков настоящего изобретения, секретируе-мых белков, антитела могут специфически связывать полноразмерный белок, кодируемый нуклеиновой кислотой настоящего изобретения, зрелый белок (т.е., белок, образованный в результате отщепления сигнального пептида), кодируемый нуклеиновой кислотой настоящего изобретения, сигнальный пептид, кодируемый нуклеиновой кислотой настоящего изобретения, или любой другой полипептид настоящего изобретения. Поэтому, эпитоп(ы) или эпитоп, несущий часть(и) полипептида, можно точно определить, как описано в данном описании, например, по N-концевому и С-концевому положению, по размеру в контиге аминокислотных остатков или описать иначе (включая список последовательностей). Антитела, которые специфически связывают любой эпитоп или полипептид настоящего изобретения, можно также исключить в качестве индивидуальных видов. Таким образом, настоящее изобретение включает антитела, которые специфически связывают определенные (заданные) полипептиды настоящего изобретения, и позволяют исключить одинаковые. Антитела настоящего изобретения можно также описать или точно определить в пределах их перекрестной реактивности. В настоящее изобретение включены антитела, которые специфически не связывают любой другой аналог, ортолог или гомолог полипептидов настоящего изобретения. В настоящее изобретение включены также антитела, которые не связывают полипептиды менее чем с 95%, менее чем с 90%, менее чем с 85%, менее чем с 80%, менее чем с 75%, менее чем с 70%, менее чем с 65%, менее чем с 60%, менее чем с 55%, менее чем с 50% идентичностью (что рассчитано с использованием способов, известных в данной области техники и описанных в данном описании) с полипептидом настоящего изобретения. Кроме того, в настоящее изобретение включены антитела, которые связывают лишь полипептиды, кодируемые полинуклеотидами, которые гибридизуются с полинуклеотидом настоящего изобретения в жестких гибридизационных условиях (как описано в данном описании). Антитела настоящего изобретения могут быть также описаны или точно определены в терминах их связывания по сродству. Предпочтительное связывание по сродству проявляют антитела с константой диссоциации или Kd меньше 5Х10-6М, 10-6М, 5Х10-7М, 10-7М, 5Х10-8М, 10-8М, 5Х10-9М, 10-9М, 5Х10-10М, 10-10М, 5Х10-11М, 10-11М, 5Х10-12М, 10-12М, 5Х10-13М, 10-13М, 5Х10-14М, 10-14М, 5Х10-15М и 10-15М. Используемые антитела настоящего изобретения включаются, но не ограничиваются ими, в известные в данной области техники способы по выделению очисткой, обнаружения и мишенирования полипептидов настоящего изобретения, в том числе in vitro и in vivo диагностические и терапевтические способы. Например, в иммуноанализе антитела используют для качественного и количественного измерения уровня полипептидов настоящего изобретения в биологических образцах. См., например, Harlow и соавт., ANTIBODIES: A LABORATORY MANUAL, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988) (включенный в данное описание путем ссылки во всей его полноте). Антитела настоящего изобретения могут использоваться отдельно или в сочетании с другими композициями. Данные антитела можно, кроме того, рекомбинантно сливать с гетерологичным полипептидом по N- или С-концу или химически конъюгировать (включая ковалентную и нековалентную конъюгацию) с полипептидами или с другими композициями. Например, антитела настоящего изобретения можно рекомбинантно слить или конъюгировать с молекулами, пригодными в качестве метки для детек-ционных анализов, и с эффекторными молекулами, такими как гетерологичные полипептиды, лекарственные средства или токсины. См., например, WO 92/08495; WO 91/14438; WO 89/12624; патент США № 5314995; и ЕР О 396387. Антитела настоящего изобретения можно получить любым подходящим способом, известным в данной области техники. Например, полипептид настоящего изобретения или его антигенный фрагмент можно вводить животному для того, чтобы индуцировать образование сыворотки, содержащей поликло-нальные антитела. Термин "моноклональное антитело" не ограничен антителами, получаемыми с помощью гибридомной техники. Термин "антитело" относится к полипептиду или к группе полипептидов, которые включают, по меньшей мере, один связывающий домен, где связывающий домен образуется из упакованных вариабельных доменов антительной молекулы с образованием трехмерного связывающего пространства, причем форма и заряд внутренней поверхности являются комплементарными элементам антигенной детерминанты антигена, что обеспечивает иммунологическое взаимодействие с антигеном. - 29 - 008252 Термин " моноклональное антитело" относится к антителу, которое получают из одиночного клона, в том числе эукариотического, прокариотического или фагового клона, но не к способу, с помощью которого его получают. Моноклональные антитела можно получить с использованием весьма разнообразных методов, известных в данной области техники, в том числе с использованием гибридомной, рекомбинант-ной и фаговой дисплейной технологии. Гибридомные методы включают методы, известные в данной области техники (см., например, Harlow и соавт. (1998)); Hammerling и соавт. (1981) (указанные ссылки включены путем ссылки во всей их полноте). Фрагменты Fab и F(ab')2 можно получить, например, из гибридомно-полученных антител в результате протеолитического расщепления, с использованием ферментов, таких как папаин (для получения Fab-фрагментов) или пепсин (для образования F(ab')2-фрагментов). Альтернативно, антитела настоящего изобретения можно получить путем применения техники ре-комбинантных ДНК или с помощью химического синтеза с использованием способов, известных в данной области техники. Например, антитела настоящего изобретения можно получить с использованием разнообразных способов фагового дисплея, известных в данной области техники. В способах фагового дисплея функциональные антительные домены представлены на поверхности фаговой частицы, которая несет кодирующие их полинуклеотидные последовательности. Фаг с требуемой связывающей способностью выбирают из набора или из комбинаторной библиотеки антител (например, человека или мыши) путем прямого отбора по антигену, как правило, антигену, связанному или захваченному твердой поверхностью или шариком. Используемый в этих способах фаг обычно представляет собой нитчатый фаг, включающий fd и М13 с Fab, Fv или стабилизированные дисульфидной связью Fv-домены антитела, ре-комбинантно слитые с фаговым геном III фага или с белком гена VIII. Примеры способов фагового дисплея, которые можно использовать для создания антител настоящего изобретения, включают способы, которые описаны у Brinkman и соавт. (1995); Ames и соавт. (1995); Kettleborough и соавт. (1994); Persic и соавт. (1997); Burton и соавт. (1994); PCT/GB91/01134; WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; W095/15982; WO 95/20401; и патенты США №№ 5698426, 5223409, 5403484, 5580717, 5427908, 5750753, 5821047, 5571698, 5427908, 5516637, 5780225, 5658727 и 5733743 (указанные ссылки включены путем ссылки во всей их полноте). Как описано в указанных выше ссылках, после фаговой селекции кодирующие антитело области можно выделить из данного фага и использовать для образования целых антител, в том числе и антител человека, или любого другого требуемого антигенсвязывающего фрагмента и экспрессировать в любом желаемом хозяине, включая клетки млекопитающих, клетки насекомых, растительные клетки, дрожжевые и бактериальные. Например, методы для рекомбинантного производства фрагментов Fab, Fab', F(ab)2 и F(ab')2, можно также употребить, используя способы, известные в данной области техники, способы, раскрытые в WO 92/22324; Mullinax и соавт. (1992); а также Sawai и соавт. (1995); а также Better и соавт. (1998) (указанные ссылки включены путем ссылки во всей их полноте). Примеры методик, которые можно использовать для получения одноцепочечных Fvs и антител, включают методики, описанные в патентах США №№ 4946778 и 5258498; Huston и соавт. (1991); Shu и соавт. (1993); и Skerra и соавт. (1988). Для определенного использования, включая in vivo использование антител у людей и in vitro детекционные анализы, предпочтительным может оказаться использование химерных, гуманизированных или антител человека. Способы получения химерных антител хорошо известны в данной области техники. См. например, Morrison (1985); Oi и соавт. (1986); Gillies, S.D. и соавт. (1989); и патент США № 5807715. Антитела можно гуманизировать с использованием разнообразных методов, включая CDR-трансплантацию (ЕР 0239400; WO 91/09967; патент США № 5530101; и № 5585089), облицовку или перекладку (ЕР 0592106; ЕР 0519596; Padlan E.A. (1991); Studnicka G.M. и соавт. (1994); Roguska M.A. и соавт. (1994)) и перестановку цепи (патент США № 5565332). Антитела человека можно создать разнообразными способами, известными в данной области техники, включая описанные выше способы фагового дисплея. См. также патенты США №№ 4444887, 4716111, 5545806, и 5814318; WO 98/46645; WO 98/50433; WO 98/24893; WO 96/34096; WO 96/33735 и WO 91/10741 (указанные ссылки включены путем ссылки во всей их полноте). В настоящее изобретение дополнительно включены рекомбинантно слитые антитела или химически конъюгированные (включая ковалентную и нековалентную конъюгацию) с полипептидом настоящего изобретения. Данные антитела могут оказаться специфичными для антигенов, отличных от полипептидов настоящего изобретения. Например, антитела можно использовать для мишенирования полипептидов настоящего изобретения в отдельных типах клеток либо in vitro, либо in vivo, путем слияния или конъюгации полипептидов настоящего изобретения с антителами, специфичными к отдельным рецепторам клеточной поверхности. Антитела, слитые или конъюгированные с полипептидами настоящего изобретения, можно также использовать в иммуноанализе in vitro и в способах очистки с использованием способов, известных в данной области техники. См., например, Harbor и соавт. выше и WO 93/21232; ЕР 0439095; Naramura, H. и соавт. (1994); патент США № 5474981; Gillies S.O. и соавт. (1992); Fell H.P. и соавт. (1991) (указанные ссылки включены путем ссылки во всей их полноте). Настоящее изобретение содержит также композиции, включающие полипептиды настоящего изобретения, слитые или конъюгированные с доменами антитела, отличными от вариабельных областей. - 30 - 008252 Например, полипептиды настоящего изобретения можно соединить путем слияния или конъюгировать с Fc-областью антитела или ее частью. Данная антительная часть, соединенная путем слияния с полипептидом настоящего изобретения, может включать шарнирную область, ОгИ-домен, СН2-домен и СН3-домен или любое сочетание целых доменов или их частей. Полипептиды настоящего изобретения можно соединить путем слияния или конъюгировать с указанными выше частями антитела для повышения in vivo времени полужизни полипептидов или для использования в иммунном анализе с использованием способов, известных в данной области техники. Полипептиды можно также соединить путем слияния или конъюгировать с указанными выше частями антитела с образованием мультимеров. Например, Fc-сегменты, слитые с полипептидами настоящего изобретения, могут образовывать димеры в результате дисульфидного связывания между Fc-сегментами. Большие мультимерные формы можно создать путем слияния полипептидов с участками IgA или IgM. Способы слияния или конъюгации полипептидов настоящего изобретения с участками антител хорошо известны в данной области техники. См., например, патенты США №№ 5336603, 5622929, 5359046, 5349053, 5447851, 5112946; ЕР 0307434, ЕР 0367166; WO 96/04388, WO 91/06570; Ashkenazi А. и соавт. (1991); Zheng Х.Х. и соавт. (1995); и Vil H. и соавт. (1992) (указанные ссылки включены путем ссылки во всей их полноте). Кроме того, настоящее изобретение относится к антителам, которые действуют в качестве агони-стов или антагонистов полипептидов настоящего изобретения. Например, настоящее изобретение включает антитела, которые частично или полностью нарушают рецептор/лигандные взаимодействия с полипептидами настоящего изобретения. Включены также рецепторспецифичные антитела и лигандспеци-фичные антитела. Включены также рецепторспецифичные антитела, которые не предотвращают лиганд-ное связывание, но предотвращают рецепторную активацию. Рецепторную активацию (т.е. сигнализацию) можно определить описанными в данном описании методами или другими методами, известными в данной области техники. Включены также рецепторспецифичные антитела, которые предотвращают как лигандное связывание, так и рецепторную активацию. Более того, включены нейтрализующие антитела, которые связывают данный лиганд и предотвращают связывание данного лиганда с данным рецептором, а также антитела, которые связывают данный лиганд и тем самым предотвращают рецепторную активацию, но не предохраняют данный лиганд от связывания данным рецептором. Кроме того, включены антитела, которые активируют данный рецептор. Эти антитела могут действовать в качестве агонистов в отношении либо всех или не всех биологических активностей, порождаемых лиганд-опосредованной рецепторной активацией. Антитела могут быть специализированными в качестве агонистов или антагонистов биологических активностей, включая описанные в данном описании специфические активности. Указанные выше антительные агонисты можно создать с использованием способов, известных в данной области техники. См., например, WO 96/40281; патент США № 5811097; Deng В. и соавт. (1998); Chen Z. и соавт. (1998); Harrop J.A. и соавт. (1998); Zhu, Z. и соавт. (1998); Yoon, D.Y. и соавт. (1998); Prat, M. и соавт. (1998); Pitard, V. и соавт. (1997); Liautard, J. и соавт. (1997); Carlson, N.G. и соавт. (1997); Taryman, R.E. и соавт. (1995); Muller, Y.A. и соавт. (1998); Bartunek, P. и соавт. (1996) (указанные ссылки включены путем ссылки во всей их полноте). Как указано выше, антитела полипептидов настоящего изобретения могут, в свою очередь, утилизироваться с образованием антиидиотипических антител, которые "имитируют" полипептиды настоящего изобретения, с использованием методов, хорошо известных специалистам в данной области техники. См., например, Greenspan and Bona (1989); Nissiniff (1991). Например, антитела, которые связывают и конкурентно ингибируют полипептидную мультимеризацию или связывание полипептида настоящего изобретения с лигандом, могут использоваться для получения антиидиотипов, которые "имитируют" полипептидную мультимеризацию или связывающий домен и, как следствие, связывают и нейтрализуют полипептид или его лиганд. Такие нейтрализующие антиидиотипические антитела можно использовать для связывания полипептида настоящего изобретения или для связывания его лигандов/рецепторов, и таким образом блокировать их биологическую активность. Двуаллельные маркеры, связанные с CanIon Преимущество двуаллельных маркеров настоящего изобретения CanIon-связанные двуаллельные маркеры настоящего изобретения обеспечивают ряд существенных преимуществ по сравнению с другими генетическими маркерами, такими как RFLP (Полиморфизм длины фрагментов рестрикции) и VNTR (Вариабельность числа тандемных повторов) маркеры. Первым поколением маркеров явились RFPL, которые представляют собой вариации, модифицирующие длину рестрикционного фрагмента. Однако способы, используемые для идентификации и типи-рования RFLP, являются достаточно неэкономичными по материалам, усилиям и времени. Вторым поколением генетических маркеров были VNTR, которые можно распределить по категориям в качестве ми-нисателлитов или микросателлитов. Минисателлиты представляют собой тандемно повторяющиеся последовательности ДНК, представленные блоками из 5-50 повторов, которые распределяются по областям хромосом человека в диапазоне длины от 0,1 до 20 т.п.н. Так как они представляют много возможных аллелей, их информативное содержание очень высоко. Минисателлиты оценивают, осуществляя Сау-зерн-блоттирование для идентификации числа тандемных повторов, присутствующих в образце нуклеиновой кислоты из отдельного тестируемого организма. Однако существует лишь 104 потенциальных - 31 - 008252 VNTR, которые можно типировать Саузерн-блоттингом. Кроме того, создание и анализ RFLP-маркеров и VNTR-маркеров в больших количествах являются дорогостоящим и трудоемким. Полиморфизм единственного нуклеотида или двуаллельные маркеры можно использовать таким же образом, что и RFLP и VNTR, но он обладает некоторыми преимуществами. SNP густо распределены в геноме человека и представляет наиболее частый тип изменчивости. По имеющимся оценкам в геноме человека рассеяно более 107-сайтов по 3х109 пар оснований. Таким образом, SNP встречается с большей частотой и с большей однородностью, чем RFLP-маркеры и VNTR-маркеры, и это означает, что существует большая вероятность обнаружения такого маркера в непосредственной близости от представляющего интерес генетического локуса. SNP менее вариабельны, чем VNTR-маркеры, и мутационно более стабильны. Кроме того, различные формы охарактеризованного полиморфизма единственного нуклеотида, такие как двуаллельные маркеры настоящего изобретения, часто легко различимы, в результате чего легче типируются стандартными методами. Двуаллельные маркеры обладают аллелями, отличающимися единственным нуклеотидом, и обладают только двумя общими аллелями, что обеспечивает параллельную детекцию и получение автоматизированной оценки. Двуаллельные маркеры настоящего изобретения обеспечивают возможность быстрого, высокопроизводительного генотипирования большого числа индивидов. Двуаллельные маркеры густо распределены в геноме, достаточно информативны и могут анализироваться в большом количестве. Объединенный эффект этих преимуществ делает двуаллельные маркеры чрезвычайно ценными в генетических исследованиях. Двуаллельные маркеры могут использоваться при изучении сцепления в семьях, в способах изучения общих аллелей, в исследованиях сцепления при нарушении равновесия в популяциях, в исследовании ассоциаций в популяциях заболевших и здоровых индивидов или в популяциях с позитивным признаком и в популяциях с негативным признаком. Существенный аспект настоящего изобретения заключается в том, что двуаллельные маркеры позволяют осуществлять ассоциативные исследования по идентификации генов, участвующих в комплексных признаках. В ассоциативных исследованиях анализируют частоту маркерных аллелей в несвязанных (неродственных) популяциях заболевших и здоровых индивидов и, как правило, используют для обнаружения полигенных или единичных признаков. Ассоциативные исследования можно осуществлять в общей популяции и они не ограничиваются исследованиями, выполняемыми на родственных индивидах в пораженных заболеванием семьях (исследования по сцеплению). Двуаллельные маркеры в разных генах можно скринировать параллельно для прямой ассоциации заболевания или ответа на лечение. Данный подход по множественным генам является мощным инструментом в разных генетических исследованиях человека, поскольку он определяет необходимость создания статистического показателя для проверки синергического эффекта множественных генетических факторов на отдельный фенотип, лекарственный ответ, спорадический признак или болезненное состояние со сложной генетической этиологией. Ген-кандидат настоящего изобретения Различные подходы можно применить для осуществления ассоциативных исследований: широкое ассоциативное исследование генома, ассоциативные исследования кандидатной области и ассоциативные исследования гена-кандидата. Широкое ассоциативное исследование генома опирается на скрининг равномерно распределенных и покрывающих весь геном генетических маркеров. Изучение гена-кандидата основано на исследовании генетических маркеров, специфически локализованных в генах, потенциально участвующих в биологическом пути, связанным с интересующим признаком. В настоящем изобретении ген-кандидат представлен CanIon. Анализ данного кандидатного гена отчетливо предоставляет упрощенный метод для идентификации генов и генных полиморфизмов, связанных с отдельными конкретными признаками при наличии определенной информации, касающейся биологии данного признака. Однако следует отметить, что все двуаллельные маркеры, раскрытые в настоящей заявке, можно использовать как часть или как часть ассоциированных исследований кандидатной области широких ассоциативных исследований генома, и такое использование специально рассматривается в настоящем изобретении и формуле изобретения. Двуаллельные маркеры, связанные с CanIon, и связанные с ними полинуклеофиды В настоящем изобретении рассматриваются также двуаллельные маркеры, связанные с CanIon. Используемый в данном описании термин "двуаллельный маркер, связанный с CanIon", относится к группе двуаллельных маркеров, сцепленных при нарушении равновесия с CanIon-геном. Термин "двуаллельный маркер, связанный с CanIon", включает двуаллельные маркеры, обозначенные А1-А17. Часть двуаллельных маркеров настоящего изобретения раскрыты в табл. 2. Они также описаны в качестве полиморфизма единственного нуклеотида для признаков, связанных SEQ ID Nos 1-4 и 6. Пары праймеров, обеспечивающих амплификацию нуклеиновой кислоты, содержащей полиморфный нуклео-тид из одного CanIon-двуаллельного маркера, приведены в табл. 1 примера 2. 17 Двуаллельных маркеров, связанных с CanIon, A1-A17, локализованы в геномной последовательности CanIon. Двуаллельные маркеры А12 и А16 локализованы в экзонах CanIon. Двуаллельный маркер А18 фланкирует CanIon-ген. Настоящее изобретение относится также к выделенной очисткой и/или выделенной нуклеотидной - 32 - 008252 последовательности, включающей полиморфное основание двуаллельного маркера, связанного с CanIon. В предпочтительных вариантах осуществления настоящего изобретения данный двуаллельный маркер выбран из группы, состоящей из А1-А18, и их комплементов. Данная последовательность содержит около 8-1000 нуклеотидов в длину и предпочтительно включает, по меньшей мере, 8, 10, 12, 15, 18, 20, 25, 35, 40, 50, 60, 70, 80, 100, 250, 500 или 1000 нуклеотидов непрерывной нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID Nos 1-4 и 6, или ее варианта, или комплементарной ей последовательности. Эти нуклеотидные последовательности включают полиморфное основание либо из аллеля 1, либо из аллеля 2 рассматриваемого двуаллельного маркера. Указанный двуаллельный маркер может находиться, необязательно, в пределах 6, 5, 4, 3, 2 или 1 нуклеотидов от центра указанного поли-нуклеотида или в центре указанного полинуклеотида. 3'-Конец указанного непрерывного участка последовательности может присутствовать, необязательно, на 3'-конце указанного полинуклеотида. Двуал-лельный маркер может быть представлен, необязательно, на 3'-конце указанного полинуклеотида. Указанный полинуклеотид может дополнительно включать, необязательно, метку. Указанный полинуклео-тид можно присоединить, необязательно, к твердому носителю. В дополнительном варианте осуществления настоящего изобретения указанные выше полинуклеотиды можно использовать отдельно или в любом сочетании. Настоящее изобретение относится также к выделенной очисткой и/или выделенной нуклеотидной последовательности, непрерывно включающей около 8-1000 нуклеотидов и/или предпочтительно, по меньшей мере, 8, 10, 12, 15, 18, 20, 25, 35, 40, 50, 60, 70, 80, 100, 250, 500 или 1000 нуклеотидов непрерывной нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID Nos 1-4 и 6, или ее варианта, или комплементарной ей последовательности. 3'-Конец указанного полинуклеотида может быть локализован, необязательно, в пределах по меньшей мере 2, 4, 6, 8, 10, 12, 15, 18, 20, 25, 50, 100, 250, 500 или 1000 нуклеотидов "левее" от связанного с CanIon двуаллельного маркера в указанной последовательности. Указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А17. 3'-Конец указанного полинуклеотида может быть локализован, необязательно, в пределах по меньшей мере 2, 4, 6, 8, 10, 12, 15, 18, 20, 25, 50, 100, 250, 500 или 1000 нуклеотидов "левее" от связанного с CanIon двуаллельного маркера в указанной последовательности. 3'-Конец указанного полинуклеотида может быть локализован, необязательно, на 1 нуклеотид "левее" от связанного с Can-Ion двуаллельного маркера в указанной последовательности. Указанный полинуклеотид может дополнительно включать, необязательно, метку. Указанный полинуклеотид может быть присоединен, необязательно, к твердому носителю. В дополнительном варианте осуществления настоящего изобретения указанные выше полинуклеотиды можно использовать отдельно или в любом сочетании. В предпочтительном варианте осуществления настоящего изобретения последовательности, включающие полиморфное основание одного из двуаллельных маркеров, перечисленных в табл. 2, выбраны из группы, состоящей из нуклеотидных последовательностей, которые обладают непрерывным участком, который состоит из полинуклеотида, который включен в полинуклеотид или который включает полинук-леотид, выбранный из группы, состоящей из последовательностей нуклеиновых кислот, изложенных в виде ампликонов, перечисленных в табл. 1, или его вариант, или комплементарную ему последовательность. Кроме того, в настоящем изобретении рассматривается нуклеиновая кислота, кодирующая CanIon-белок, где указанная нуклеиновая кислота включает полиморфное основание двуаллельного маркера, выбранного из группы, состоящей из А12 и А16 и их комплементов. Настоящее изобретение включает также использование любого полинуклеотида для определения идентичности одного или более нуклеотидов в двуаллельном маркере, связанном с CanIon, или любой полинуклеотид для использования в указанном определении. Кроме того, полинуклеотиды настоящего изобретения, используемые для определения идентичности одного или нескольких нуклеотидов в двуал-лельном маркере, связанном с CanIon, включают полинуклеотиды с любым дополнительным ограничением, описанным в данном описании, или следующие ниже полинуклеотиды, указанные отдельно или в любом сочетании. Указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А18 и их комплементов, или двуаллельных маркеров необязательно сцепленных к тому же при нарушении равновесия; указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А17 и их комплементов, или двуаллельных маркеров, необязательно сцепленных к тому же при нарушении равновесия; указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А12 и А16 и их комплементов, или двуаллельных маркеров, необязательно сцепленных к тому же при нарушении равновесия. Указанный полинуклеотид может включать, необязательно, последовательность, раскрытую в настоящем описании. Указанный по-линуклеотид может состоять, необязательно, или в основном состоять из любого полинуклеотида, описанного в настоящем описании. Указанное определение можно осуществить, необязательно, в гибриди-зационном анализе, секвенирующем анализе, микросеквенирующем анализе или в анализе методом ферментного выявления дефекта комплементарного спаривания. Указанный полинуклеотид можно присоединить, необязательно, к твердому носителю, матрице или адресной матрице. Указанный полинуклеотид, необязательно, может быть меченным. Предпочтительный полинуклеотид можно использовать в гибри-дизационном анализе для установления идентичности определенного нуклеотида в связанном с CanIon - 33 - 008252 двуаллельном маркере. Другой предпочтительный полинуклеотид можно использовать в секвенирую-щем или микросеквенирующем анализе для определения идентичности нуклеотида в связанном с CanIon двуаллельном маркере. Третий предпочтительный полинуклеотид можно использовать в анализе методом ферментного выявления дефекта комплементарного спаривания для определения идентичности нук-леотида в связанном с CanIon двуаллельном маркере. Четвертый предпочтительный полинуклеотид можно использовать для амплификации полинуклеотидного сегмента, включающего связанный с CanIon двуаллельный маркер. Любой из описанных выше полинуклеотидов можно присоединить, необязательно, к твердому носителю, матрице или к адресной матрице. Указанный полинуклеотид может быть, но необязательно, меченным. Настоящее изобретение дополнительно включает в себя использование любого полинуклеотида для использования для амплификации нуклеотидного сегмента, включающего связанный с CanIon двуал-лельный маркер. Кроме того, полинуклеотиды настоящего изобретения, используемые для амплификации нуклеотидного сегмента, включающего связанный с CanIon двуаллельный маркер, включают в себя полинуклеотиды с любым дополнительным ограничением, описанным в данном описании, или следующие ниже полинуклеотиды, указанные отдельно или в любом сочетании. Указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А18 и их комплементов, или двуаллельных маркеров необязательно, к тому же, находящихся в состоянии сцепления при нарушении равновесия; указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А17 и их комплементов, или указанные двуаллельные маркеры необязательно, к тому же, сцеплены при нарушении равновесия; необязательно указанный двуаллельный маркер, связанный с Can-Ion, выбран из группы, состоящей из А12 и А16 и их комплементов, или двуаллельные маркеры необязательно, к тому же, сцеплены при нарушении равновесия; указанный полинуклеотид может включать необязательно последовательность, раскрытую в настоящем описании; указанный полинуклеотид может необязательно состоять или в основном состоять из полинуклеотида, описанного в настоящем описании; указанная амплификация необязательно может осуществляться с помощью ПЦР или ЛЦР. Указанный полинуклеотид необязательно может присоединяться к твердому носителю, матрице или адресной матрице. Указанный полинуклеотид необязательно может быть меченным. Праймеры для амплификации или реакции секвенирования полинуклеотида, включающего двуал-лельный маркер настоящего изобретения, может быть создан из раскрытых последовательностей с помощью любого способа, известного в данной области техники. Предпочтительный набор праймеров приспособлен так, что 3'-конец непрерывного участка последовательности, идентичной последовательности, выбранной из группы, состоящей из SEQ ID Nos 1-4 и 6, или из последовательности, комплементарной им, или из ее варианта, присутствует на 3'-конце данного праймера. Такая конфигурация позволяет 3'-концу данного праймера гибридизоваться с выбранной последовательностью нуклеиновой кислоты и резко увеличивать эффективность данного праймера в амплификации или в реакциях секвенирования. Аллельспецифичные праймеры можно создать так, что полиморфное основание двуаллельного маркера находится на 3'-конце непрерывного участка последовательности, а непрерывный участок последовательности находится на 3'-конце данного праймера. Такие аллельспецифичные праймеры стремятся избирательно начать процесс амплификации или реакцию секвенирования, если они используются с образцом нуклеиновой кислоты, который содержит один из двух аллелей, представленный в двуаллельном маркере. 3'-Конец праймера настоящего изобретения может быть локализован в пределах или, по меньшей мере, 2, 4, 6, 8, 10, 12, 15, 18, 20, 25, 50, 100, 250, 500 или 1000 нуклеотидами "левее" от связанного с CanIon двуаллельного маркера в указанной последовательности или в любом другом месте, которое соответствует их предполагаемому использованию в секвенировании, амплификации или расположению новых последовательностей или маркеров. Поэтому другой набор предпочтительных амплификационных праймеров включает выделенный полинуклеотид, в основном состоящий из непрерывного участка последовательности в 8-50 нуклеотидов в последовательности, выбранной из группы, состоящей из SEQ ID Nos 1-4 и 6, или комплементарной ей последовательности, или ее варианта, где 3'-конец указанного непрерывного участка последовательности локализован по 3'-концу указанного полинуклеотида, и где 3'-конец указанного полинуклеотида локализован "левее" связанного с CanIon двуаллельного маркера в указанной последовательности. Предпочтительно, эти амплификационные праймеры включают последовательность, выбранную из группы, состоящей из последовательностей В1-В17 и С1-С17. Праймеры с 3'-концами, локализованными на 1 нуклеотид "левее" от двуаллельного маркера CanIon, особенно полезны в микросеквенирующем анализе. Предпочтительные микросеквенирующие праймеры описаны в табл. 4. Указанный двуаллельный маркер, связанный с CanIon, выбран из группы, состоящей из А1-А18 и их комплементов, или двуал-лельные маркеры необязательно, к тому же, сцеплены при нарушении равновесия; указанный двуаллель-ный маркер, связанный с CanIon, выбран из группы, состоящей из А1-А17 и их комплементов, или дву-аллельные маркеры необязательно, к тому же, сцеплены при нарушении равновесия; указанный двуал-лельный маркер, связанный с CanIon, выбран из группы, состоящей из А12 и А16 и их комплементов, или двуаллельные маркеры необязательно, к тому же, сцеплены при нарушении равновесия. Микросек-венирующие праймеры выбраны, но необязательно, из группы нуклеотидных последовательностей D1- - 34 - 008252 D18 и Е1-Е18. Зонды настоящего изобретения можно создать из раскрытых последовательностей любым способом, известным в данной области техники, в частности, с помощью способов, которые обеспечивают тестирование при наличии раскрытого в данном описании маркера. Предпочтительный набор зондов можно создать для использования в гибридизационном анализе настоящего изобретения любым способом, известным в данной области техники, так что они избирательно связываются с одним аллелем дву-аллельного маркера, но не с другим, в соответствии с любой конкретной совокупностью условий анализа. Предпочтительные гибридизационные зонды включают полиморфное основание из аллеля 1 или ал-леля 2 рассматриваемого двуаллельного маркера. Указанный двуаллельный маркер может находиться, необязательно, в пределах 6, 5, 4, 3, 2 или 1 нуклеотида(ов) от центра гибридизационного зонда или в центре указанного зонда. В предпочтительном варианте осуществления настоящего изобретения данные зонды выбирают из группы, содержащей каждую из последовательностей Р1-Р18 и каждую из комплементарных им последовательностей. Следует отметить, что полинуклеотиды настоящего изобретения не ограничиваются имеющими точные фланкирующие последовательности, окружающие полиморфные основания, которые перечислены в Списке Последовательностей. Следует иметь ввиду, что фланкирующие последовательности, окружающие двуаллельные маркеры, могут быть удлинены или укорочены до любого размера, сопоставимого с их предполагаемым использованием, и в настоящем изобретении, в частности, такие последовательности рассматриваются. Для фланкирующих областей вне непрерывного участка последовательности не требуется быть гомологичными нативным фланкирующим последовательностям, которые действительно встречаются у людей. Специально рассматривается добавление любой нуклеотидной последовательности, которая сопоставима с нуклеотидами, предназначенными для использования. Праймеры и зонды можно пометить или иммобилизовать на твердом носителе, как описано в "Оли-гонуклеотидных зондах и праймерах". Полинуклеотиды настоящего изобретения, которые присоединяют к твердому носителю, включают в себя полинуклеотиды с любым дополнительным ограничением, описанным в данном описании, или следующие ниже полинуклеотиды, указанные отдельно или в любом сочетании. Указанные полинуклео-тиды могут быть определены, необязательно, как индивидуально присоединенные или в группах, по меньшей мере, из 2, 5, 8, 10, 12, 15, 20 или 25 отдельных полинуклеотидов настоящего изобретения, к единственному твердому носителю. Полинуклеотиды, отличающиеся от полинуклеотидов настоящего изобретения, необязательно могут присоединяться к одному и тому же твердому носителю в качестве полинуклеотидов настоящего изобретения. Если к твердому носителю присоединяется множество поли-нуклеотидов, они могут присоединяться необязательно случайным образом или в виде упорядоченного набора. Указанный упорядоченный набор может быть необязательно адресным. Настоящее изобретение включает в себя также диагностические наборы, включающие один или более полинуклеотидов настоящего изобретения вместе с частью или со всеми необходимыми реагентами и инструкциями для генотипирования обследуемого индивида для определения идентичности нуклеоти-да в связанном с CanIon двуаллельным маркере. Полинуклеотиды набора могут необязательно присоединяться к твердому носителю или являться частью набора или адресным набором полинуклеотидов. Данный набор можно создать для определения идентичности нуклеотида в позиции маркера любым способом, известным в данной области техники, включающим, но не ограничивающимся указанным, способ аналитического секвенирования, способ аналитического микросеквенирования, способ аналитической гибридизации или способ ферментного выявления дефекта комплементарного спаривания. Способы идентификации двуаллельных маркеров de novo Для скринирования геномного фрагмента на полиморфизм единственного нуклеотида можно использовать различные способы, такие как дифференциальная гибридизация с олигонуклеотидными зондами, обнаружение изменений в подвижности, измеряемой с помощью гель-электрофореза, или прямым секвенированием амплифицированной нуклеиновой кислоты. Предпочтительный способ идентификации двуаллельных маркеров включает сравнительное секвенирование фрагментов геномной ДНК у соответствующего числа не связанных родством индивидов. В первом варианте осуществления настоящего изобретения образцы ДНК от не связанных родством индивидов объединяют вместе, после чего представляющую интерес геномную ДНК амплифицируют и секвенируют. Затем, полученные таким образом нуклеотидные последовательности анализируют, чтобы идентифицировать достоверный полиморфизм. Одно из основных преимуществ данного способа заключается в том, что объединение образцов ДНК в значительной степени снижает количество реакций амплификации и реакций секвенирования ДНК, которые следует осуществить. Кроме того, данный способ достаточно чувствителен, так что полученный таким образом двуаллельный маркер обычно демонстрирует достаточную частоту своего менее распространенного аллеля, который полезен для проведения ассоциативных исследований. В другом варианте осуществления настоящего изобретения образцы ДНК не объединяют и поэтому их амплифицируют и секвенируют индивидуально. Данный способ, как правило, предпочтителен, если двуаллельные маркеры нуждаются в идентификации, чтобы осуществить ассоциативные исследования в - 35 - 008252 рамках кандидатных генов. Предпочтительно, в случае двуаллельных маркеров можно скринировать высокозначимые генные области, такие как промоторные области или экзонные области. Двуаллельный маркер, полученный с использованием данного способа, может проявить меньшую информативность для проведения ассоциативных исследований, например, если частота его наименее частого аллеля составляет менее чем около 10%. Вместе с тем, такой двуаллельный маркер будет достаточно информативным для проведения ассоциативных исследований и, кроме того, ценным из-за того, что включение менее информативных двуаллельных маркеров в исследования настоящего изобретения по генетическому анализу может обеспечить в некоторых случаях прямую идентификацию каузальных (определяющих данный признак) мутаций, которые могут, в зависимости от их пенетрантности, оказаться редкими мутациями. Ниже следует описание различных параметров предпочтительного способа, используемого заявителями для идентификации двуаллельных маркеров настоящего изобретения. Образцы геномной ДНК Образцы геномной ДНК, из которых создают двуаллельные маркеры настоящего изобретения, предпочтительно получают от не связанных родством индивидов, соответствующих гетерогенной популяции с известным этническим фоном. Число индивидов, от которых получают образцы ДНК, может существенно варьировать, предпочтительно от около 10 до около 1000, предпочтительно от около 50 до около 200 индивидуумов. Как правило, предпочтительно собирать образцы ДНК, по меньшей мере, от около 100 индивидуумов, для того, чтобы обладать достаточно полиморфным разнообразием в данной популяции, чтобы идентифицировать столько маркеров, сколько возможно, и чтобы получить статистически значимые результаты. Что касается источника геномной ДНК, которую подвергают анализу, то анализируемым образцом может быть любой образец без какого-либо особого ограничения. Такие анализируемые образцы включают биологические образцы, которые можно проанализировать описанными в данном описании способами настоящего изобретения, и включают жидкости организма человека и животного, такие как цельная кровь, сыворотка, плазма, спинно-мозговая жидкость, моча, лимфатическая жидкость и различные внешние выделения при дыхании, выделения кишечного и мочеполого трактов, слезы, слюну, молоко, лейкоциты, миеломы и тому подобное; биологические жидкости, такие как супернатанты клеточных культур; фиксированные тканевые образцы, включая злокачественную и незлокачественную ткань и ткани лимфатических узлов; пункции костного мозга и образцы фиксированных клеток. Предпочтительным источником геномной ДНК, используемой в настоящем изобретении, является периферическая венозная кровь каждого донора. Методы получения геномной ДНК из биологических образцов хорошо известны квалифицированным специалистам. Подробности предпочтительного варианта осуществления настоящего изобретения приведены в примере 1. Специалист в данной области техники может выбрать для амплификации объединенные или необъединенные ДНК-образцы. Амплификация ДНК Идентификации двуаллельных маркеров в образце геномной ДНК может способствовать использование способов амплификации ДНК. Для стадии амплификации образцы ДНК можно объединить или не объединять. Методы амплификации ДНК хорошо известны специалистам в данной области техники. Методы амплификации, используемые в контексте настоящего изобретения, включают, но не ограничиваются ими, лигазную цепную реакцию (LCR), описанную в ЕР-А-320308, WO 9320227 и ЕР-А-439182, полимеразную цепную реакцию (PCR, RT-PCR) и такие методы, как амплификация на основе последовательности нуклеиновой кислоты (NASBA), описанная у Guatelli J.С. и соавт. (1990) и у Compton J. (1991), Q-бета амплификация, которая описана в европейской патентной заявке №4544610, амплификация с замещением цепи, как описано у Walker и соавт. (1996) и ЕР А 684315, и опосредованная мишенью амплификация, как описано в публикации РСТ WO 9322461. LCR и Gap LCR являются экспоненциальными амплификационными методами, обе зависят от ДНК-лигазы, которая соединяет смежные праймеры, отжигаемые к молекуле ДНК. В лигазной цепной реакции (LCR) используют пару зондов, которые включают два первичных [основных] (первый и второй) и два вторичных [вспомогательных] (третий и четвертый) зонда, которые все используются в молярном избытке к мишени. Первый зонд гибридизуется с первым сегментом данной цепи-мишени, а второй зонд гибридизуется со вторым сегментом данной цепи-мишени, причем первый и второй сегменты являются смежными, так что первичные зонды примыкают друг к другу в 5'фосфат-3'-гидроксильной взаимосвязи, и так что лигаза может ковалентно сочетать или лигировать два зонда в соединенный путем слияния продукт. Кроме того, третий (вторичный) зонд может гибридизоваться с частью первого зонда, а четвертый (вторичный) зонд может гибридизоваться с частью второго зонда путем аналогичного примыкания. Конечно, если данная мишень исходно состоит из двух цепей, то в первом случае вторичные зонды также будут гибридизоваться с комплементом-мишенью. После того, как лигированная цепь первичных зондов отделена от цепи-мишени, она будет гибридизоваться с третьим и четвертым зондом, которые могут быть лигированы с образованием комплементарного, вторичного лигированного продукта. Важно осознавать, что данные лигированные продукты функционально эквивалентны данной мишени или ее комплементу. В результате повторения циклов гибридизации и лигирования достигается амплификация последовательности-мишени. Описан также способ мультиплексной LCR (WO 9320227). Gap - 36 - 008252 LCR (GLCR) представляет собой вариант LCR, в котором зонды не соседствуют, а разделены 2-3 основаниями. Что касается амплификации мРНК, то в рамках настоящего изобретения мРНК обратно транскрибируют в кДНК с последующей полимеразной цепной реакцией (RT-PCR); или с использованием в обоих стадиях единственного фермента, как описано в патенте США №5322770, или с использованием Асимметричной Gap LCR (RT-AGLCR), как описано у Marshall и соавт. (1994). AGLCR представляет собой модификацию GLCR, которая делает возможной амплификацию РНК. Техника ПЦР является предпочтительной техникой амплификации, используемой в настоящем изобретении. Разнообразные методы ПЦР хорошо известны специалистам в данной области техники. Обзор техники ПЦР см. у White (1997) и в публикации, озаглавленной "PCR Methods and Applications" (1991, Cold Spring Harbor Laboratory Press). В каждой из этих методик ПЦР-праймеры для каждой из сторон амплифицируемой последовательности нуклеиновой кислоты добавляются к соответствующим образом приготовленному образцу нуклеиновой кислоты вместе с dNTP и термостабильной полимеразой, такой как Taq-полимераза, Pfu-полимераза или Vent-полимераза. Нуклеиновую кислоту данного образца денатурируют и ПЦР-праймеры специфически гибридизуются с комплементарными нуклеотидными последовательностями в данном образце. Гибридизующиеся праймеры удлиняются. Поэтому инициируется очередной цикл денатурации, гибридизации и удлинения. Такие циклы повторяются много раз с образованием амплифицируемого фрагмента, содержащего нуклеотидную последовательность между праймер-ными сайтами. ПЦР дополнительно была описана в нескольких патентах, в том числе в патентах США №№4683195; 4683202 и 4965188, раскрытие которых включено в данное описание путем ссылки во всей их полноте. Техника ПЦР представляет собой предпочтительную технику амплификации, используемую для идентификации новых двуаллельных маркеров. Типичный пример реакции ПЦР, подходящей для целей настоящего изобретения, представлен в примере 2. Одним из аспектов настоящего изобретения является способ амплификации CanIon-гена человека, в частности, фрагмента геномной последовательности SEQ ID No 1-3 или кДНК-последовательности SEQ ID No 4, ее фрагмента или ее вариант в анализируемом образце, предпочтительно с использованием ПЦР. Данный способ включает следующие стадии: a) контактирования анализируемого образца с реагентами реакции амплификации, включающими пару праймеров амплификации, как описано выше, и локализованных на любой стороне амплифицируе-мого полинуклеотидного участка, и b) необязательного детектирования продуктов амплификации. В настоящем изобретении рассматривается также набор для амплификации последовательности CanIon-гена, в частности участка геномной последовательности SEQ ID No 1-3 или кДНК-последователь-ности SEQ ID No 4 или ее варианта в анализируемом образце, где указанный набор включает: a) пару олигонуклеотидных праймеров, локализованных на любом участке амплифицируемого района CanIon; b) необязательно реагенты, необходимые для выполнения реакции амплификации. В одном из вариантов осуществления указанного выше способа амплификации и набора реагентов к нему продукт амплификации детектируют путем гибридизации с меченым зондом, обладающим последовательностью, которая комплементарна амплифицированному участку. В другом варианте осуществления указанного выше способа амплификации и набора реагентов к нему праймеры включают последовательность, которая выбрана из группы, состоящей из нуклеотидных последовательностей В1-В17, С1-С17, D1-D18, и Е1-Е18. В первом варианте осуществления настоящего изобретения двуаллельные маркеры идентифицируют с использованием информации о геномной последовательности, которая создана заявителями. Фрагменты секвенированной геномной ДНК используют для создания праймеров для амплификации (500 п.н.)-фрагментов. (500 п.н.)-фрагменты амплифицируют из геномной ДНК и сканируют на наличие двуаллель-ных маркеров. Праймеры можно создать с использованием программного обеспечения OSP (Hillier L. and Green P., 1991). Все праймеры могут находиться "левее" специфичных оснований-мишеней, общий оли-гонуклеотидный хвост которых служит в качестве секвенирующего праймера. Специалистам в данной области техники хорошо известно удлинение праймеров, которое можно использовать для этих целей. Предпочтительные праймеры, пригодные для амплификации геномных последовательностей, кодирующих CanIon-ген, фокусируются на промоторах, экзонах и сайтах сплайсинга генов. Двуаллельный маркер обнаруживает более высокую вероятность возможного появления каузальной мутации, если он локализован в этих функциональных участках данного гена. Предпочтительные праймеры амплификации настоящего изобретения включают нуклеотидные последовательности В1-В17 и С1-С17, дополнительно детализированные в примере 2, табл. 1. Секвенирование амплифицированной геномной ДНК и идентификация полиморфизма единственного нуклеотида Продукты амплификации, полученные как описано выше, затем секвенируют с использованием любого способа, известного и доступного квалифицированным специалистам. Способы секвенирования - 37 - 008252 ДНК с использованием дидезоксиопосредованного метода (метод Сэнгера) или метода Максам-Гилберта, широко известны специалистам в данной области техники. Такие методы раскрыты, например, у Sambrook и соавт. (1989). Альтернативные способы включают гибридизацию с набором зондов ДНК высокой плотности, как описано у Chee и соавт. (1996). Предпочтительно, амплифицированную ДНК подвергают автоматизированным дидезокситермини-рующим секвенирующим реакциям с использованием протокола циклического секвенирования с прай-мером, меченым красителем. Продукты реакций секвенирования разделяют на секвенирующих гелях и полученные последовательности определяют с использованием анализа визуализации геля. Поиск полиморфизма основан на наличии накладывающихся пиков в электрофоретическом паттерне, полученном для разных оснований, встреченных в одной и той же позиции. Поскольку каждый дидезокситерминатор помечен отличающейся флуоресцентной молекулой, то два пика, соответствуя двуаллельному сайту, представлены разным цветом, соответствуя двум отличающимся нуклеотидам по одной и той же позиции в данной последовательности. Однако присутствие двух пиков может оказаться артефактом, обусловленным фоновым шумом. Чтобы исключить такой артефакт, секвенируют две цепи ДНК и сравнивают их по двум полученным пикам. Последовательность квалифицируют как полиморфную в том случае, если полиморфизм обнаруживается на обоих цепях. Указанная выше методика позволяет идентифицировать те продукты амплификации, которые содержат двуаллельные маркеры. Граница выявления частоты двуаллельного полиморфизма, обнаруженного путем секвенирования пулов из 100 индивидов, составляет приблизительно 0,1 для минорного алле-ля, что подтверждается пулами секвенирования для известных аллельных частот. Однако более 90% дву-аллельного полиморфизма, обнаруженного методом объединения, характеризуется частотой минорного аллеля, превышающей значение 0,25. Поэтому, двуаллельные маркеры, выбранные с помощью данного метода, обладают частотой, по меньшей мере, 0,1 для минорного аллеля и менее 0,9 для основного алле-ля. Предпочтительно, если частота составляет, по крайней мере, 0,2 для минорного аллеля и менее 0,8 для основного аллеля, более предпочтительно, по меньшей мере, 0,3 для минорного аллеля и менее 0,7 для основного аллеля, таким образом уровень гетерозиготности превышает 0,18, предпочтительно выше 0,32, более предпочтительно выше 0,42. В другом варианте осуществления настоящего изобретения двуаллельные маркеры детектируется путем секвенирования индивидуальных образцов ДНК, где частота минорного аллеля, такого как двуал-лельный маркер, может составлять менее 0,1. Проверка достоверности двуаллельных маркеров настоящего изобретения Полиморфизм оценивают по его пригодности в качестве генетических маркеров путем подтверждения того, что оба аллеля присутствуют в популяции. Проверка достоверности двуаллельных маркеров осуществляется путем генотипирования группы индивидов способом настоящего изобретения и демонстрации того, что оба аллеля присутствуют. Микросеквенирование является предпочтительным методом генотипирования аллелей. Проверка достоверности с помощью операции генотипирования осуществляют на индивидуальных образцах, полученных от каждого индивида данной группы или путем генотипи-рования объединенной выборки, полученной от более чем одного индивида. Данная группа может равняться одному индивиду, если этот индивид является гетерозиготой по аллелю, о котором идет речь. Предпочтительно группа содержит по меньшей мере три индивида, более предпочтительно данная группа содержит пять или шесть индивидов, так что единственный тест на достоверность, вполне вероятно, приведет к подтверждению существования большинства тестируемых двуаллельных маркеров. Следует, однако, иметь ввиду, что при осуществлении теста на достоверность в небольшой группе можно столкнуться с ложным отрицательным результатом, если в качестве результата ошибки выборки ни один из проверяемых индивидов не несет один из двух данных аллелей. Таким образом, метод проверки достоверности менее пригоден для демонстрации того, что отдельный исходный результат представляет собой артефакт, а не демонстрации того, что в конкретной позиции последовательности существует подлинный двуаллельный маркер. Методы изучения генотипирования, гаплотипирования, ассоциации и взаимодействия настоящего изобретения можно необязательно осуществлять исключительно в отношении достоверности двуаллельных маркеров. Оценка частоты двуаллельных маркеров настоящего изобретения Проверенные на достоверность двуаллельные маркеры далее оценивают на их пригодность в качестве генетических маркеров определения частоты менее распространенного аллеля в сайте двуаллельно-го маркера. Чем больше частота менее распространенного аллеля, тем выше пригодность двуаллельного маркера для исследований по ассоциации и взаимодействию. Определение наименее распространенного аллеля осуществляется путем генотипирования группы индивидов способом настоящего изобретения и демонстрацией присутствия обоих аллелей. Это определение частоты с помощью операции генотипиро-вания осуществляют на индивидуальных образцах, полученных от каждого индивида данной группы или путем генотипирования объединенной выборки, полученной от более чем одного индивида. Данная группа должна быть достаточно большой, чтобы представлять данную популяцию как целое. Предпочтительно данная группа содержит, по меньшей мере, 20 индивидов, более предпочтительно данная группа содержит, по меньшей мере, 50 индивидов, более предпочтительно данная группа содержит, по меньшей - 38 - 008252 мере, 100 индивидов. Разумеется, большая группа увеличит точность определения частоты, вследствие снижения ошибки выборки. Двуаллельный маркер, у которого частота менее распространенного аллеля составляет 30% или больше, называют "высококачественным двуаллельный маркером". Методы изучения генотипирования, гаплотипирования, ассоциации и взаимодействия настоящего изобретения можно необязательно осуществлять лишь с двуаллельными маркерами высокого качества. Способы индивидуального генотипирования по двуаллельным маркерам Разработаны способы определения генотипа биологического образца по одному или нескольким двуаллельным маркерам настоящего изобретения, которые все могут осуществляться in vitro. Такие способы генотипирования включают определение идентичности нуклеотида в сайте CanIon-двуаллельного маркера любым способом, известным в данной области техники. Такие способы находят применение при генотипировании больных и здоровых популяций в ассоциативных исследованиях, а также при геноти-пировании индивидов в контексте выявления аллелей двуаллельных маркеров, которые, как установлено, ассоциированы с данным признаком, и в этом случае обе копии двуаллельного маркера, представленные в геноме индивида, определяются так, что индивид можно классифицировать как гомозиготу или гетеро-зиготу по отдельному аллелю. Данные способы генотипирования осуществляют на образцах нуклеиновых кислот, полученных от отдельного индивида или на объединенных образцах ДНК. Генотипирование можно осуществлять с использованием аналогичных способов, которые описаны выше для идентификации двуаллельных маркеров, или с использованием других способов генотипиро-вания, таких как описано ниже. В предпочтительных вариантах осуществления настоящего изобретения сравнение последовательностей амплифицированных геномных фрагментов от разных индивидов используют для идентификации новых двуаллельных маркеров, а микросеквенирование используется при генотипировании известных двуаллельных маркеров для применения в диагностическом и ассоциативном исследовании. Один из вариантов осуществления настоящего изобретения включает в себя способы генотипиро-вания, включающие определение идентичности нуклеотида в CanIon-связанном двуаллельном маркере или в его дополнении в биологическом образце; где указанный CanIon-связанный двуаллельный маркер необязательно выбран из группы, состоящей из А1-А18 и их комплементов, или необязательно двуал-лельные маркеры при этом неравновесно сцеплены; где указанный CanIon-связанный двуаллельный маркер необязательно выбран из группы, состоящей из А1-А17 и их комплементов, или необязательно дву-аллельные маркеры при этом неравновесно сцеплены; где указанный CanIon-связанный двуаллельный маркер необязательно выбран из группы, состоящей из А12 и А16 и их комплементов, или необязательно двуаллельные маркеры при этом неравновесно сцеплены; где указанный биологический образец необязательно получен от отдельного индивида; где идентичность нуклеотидов в указанном двуаллельном маркере необязательно определяют по обоим копиям указанного двуаллельного маркера, представленного в указанном геноме индивида; где указанный биологический образец необязательно получен от многих индивидов. Способы генотипирования настоящего изобретения необязательно включают в себя способы с любым дополнительным ограничением, описанным в данном описании, или следующие ниже способы, указанные отдельно или в любом сочетании. Указанный способ необязательно осуществляют in vitro; кроме того, необязательно включение амплификации части указанной последовательности, включающей двуаллельный маркер, перед указанной стадией определения. Где указанную амплификацию необязательно осуществляют с помощью ПЦР, ЛЦР или репликацией рекомбинантного вектора, включающего ориджин репликации и указанный фрагмент, в клетке-хозяине; где указанное определение необязательно осуществляют с помощью гибридизационного анализа, секвенирующего анализа, микросеквенирующего анализа или метода ферментного выявления дефекта комплементарного спаривания. Происхождение нуклеиновых кислот для генотипирования Любую нуклеиновую кислоту, выделенную очисткой или не выделенную очисткой, можно использовать в качестве исходной нуклеиновой кислоты, если содержит или предположительно содержит требуемую специфичную последовательность нуклеиновой кислоты. ДНК или РНК можно экстрагировать из клеток, тканей, жидкостей организма и тому подобное. Хотя нуклеиновые кислоты, используемые в способах генотипирования настоящего изобретения, можно получить от любого млекопитающего, под обследуемыми субъектами и индивидами, у которых взяты образцы нуклеиновой кислоты, подразумевают человека. Амплификация фрагментов ДНК, включающих двуаллельные маркеры Обеспечены способы и полинуклеотиды для амплификации сегмента нуклеотидов, включающие один или несколько двуаллельных маркеров настоящего изобретения. Следует иметь в виду, что амплификация фрагментов ДНК, включающих двуаллельные маркеры, может осуществляться разными способами и для разных целей и не ограничивается генотипированием. Тем не менее, многие способы геноти-пирования, хотя и не все, требуют предварительной амплификации участка ДНК, несущего интересующий двуаллельный маркер. Такие способы специфически увеличивают концентрацию или общее количество последовательностей, которые включают двуаллельный маркер или включают такой участок, и последовательности, локализованные вблизи или на расстоянии от него. Диагностический анализ может - 39 - 008252 быть также основан на амплификации сегментов ДНК, несущих двуаллельный маркер настоящего изобретения. Амплификацию ДНК можно осуществить любым способом, известным в данной области техники. Методы амплификации описаны выше в разделе, озаглавленном "Амплификация ДНК". Некоторые из таких методов амплификации особенно подходят для обнаружения полиморфизма единственного нуклеотида и делают возможной одновременную амплификацию последовательности-мишени и идентификацию полиморфного нуклеотида, как это описано ниже. Идентификация описанных выше двуаллельных маркеров позволяет создать соответствующие оли-гонуклеотиды, которые можно использовать в качестве праймеров для амплификации фрагментов ДНК, включающих двуаллельные маркеры настоящего изобретения. Амплификацию можно осуществить с использованием праймеров, первоначально используемых для открытия описанных в данном описании новых двуаллельных маркеров, или любого набора праймеров, обеспечивающих амплификацию фрагмента ДНК, включающего двуаллельный маркер настоящего изобретения. В некоторых вариантах настоящего изобретения созданы праймеры для амплификации фрагмента ДНК, содержащего один или несколько двуаллельных маркеров настоящего изобретения. Предпочтительные амплификационные праймеры перечислены в примере 2. Следует иметь ввиду, что данные перечисленные праймеры являются лишь иллюстративными, и что также используют любой другой набор праймеров, который производит продукты амплификации, содержащие один или несколько двуаллель-ных маркеров настоящего изобретения. Расстояние между праймерами определяет длину амплифицированного сегмента. В контексте настоящего изобретения амплифицированные сегменты, несущие двуаллельные маркеры, можно расположить по размеру по меньшей мере от около 25 п.н. до 35 т.п.н. Фрагменты амплификации с 25-3000 п.н. являются обычными, фрагменты с 50-1000 п.н. являются предпочтительными, а фрагменты со 100-600 п. н. являются высокопредпочтительными. Следует иметь в виду, что праймеры амплификации двуал-лельных маркеров могут представлять собой любую последовательность, которая делает возможной специфичную амплификацию любого фрагмента ДНК, несущего данные маркеры. Праймеры амплификации можно метить или иммобилизовать на твердом носителе, как описано в разделе "Олигонуклеотидные зонды и праймеры". Способ генотипирования образцов ДНК по двуаллельным маркерам Для идентификации нуклеотида, присутствующего в сайте двуаллельного маркера, можно использовать любой способ, известный в данной области техники. Поскольку данный детектируемый аллель двуаллельного маркера был идентифицирован и точно определен в настоящем изобретении, детекция оказывается простой для среднего специалиста в данной области техники, с применением любой из имеющихся методик. Многие способы генотипирования требуют осуществления предварительной амплификации области ДНК, несущей интересующий двуаллельный маркер. Хотя в настоящее время амплификация мишени или сигнала часто предпочтительна, сверхчувствительные способы детекции, которые не требуют амплификации, также включены в современные способы генотипирования. Способы, хорошо известные специалистам в данной области техники, которые могут использоваться для выявления двуаллельного полиморфизма, включают такие способы, как стандартный дот-блот анализ, анализ однонитевого конформационного полиморфизма (SSCP), описанного Orieta и соавт. (1989), денатурирующий градиентный гель-электрофорез (DGGE), гетеродуплексный анализ, обнаружение расщепления ошибочного спаривания и другие традиционные методики, которые описаны у Sheffield и соавт. (1991), White и соавт. (1992), Grompe и соавт. (1989 и 1993). Другой способ определения идентичности нуклео-тида, присутствующего в конкретном полиморфном сайте, использует специализированное, устойчивое к экзонуклеазе, нуклеотидное производное, как описано в патенте США № 4656127. Предпочтительные способы подразумевают прямое определение идентичности нуклеотида, присутствующего в сайте двуаллельного маркера, с помощью анализа секвенирования, анализа ферментного выявления дефекта комплементарно спаривания или гибридизационного анализа. Далее следует описание некоторых предпочтительных способов. Особо предпочтительным способом является метод микро-секвенирования. Используемый, в основном, в данном описании термин "секвенирование" относится к полимеразному удлинению дуплексных комплексов праймер/матрица и включает традиционное секве-нирование и микросеквенирование. 1) Секвенирующий анализ. Нуклеотид, присутствующий в полиморфном сайте, можно определить с помощью способов секве-нирования. В предпочтительном варианте осуществления настоящего изобретения образцы ДНК перед секвенированием подвергают ПЦР-амплификации, как описано выше. Способы секвенирования ДНК описаны в разделе "Секвенирование амплифицированной геномной ДНК и идентификация полиморфизма единственного нуклеотида". Предпочтительно, амплифицированную ДНК подвергают автоматизированному дидезокситерми-нирующему секвенированию с использованием протокола цикла секвенирования с праймером, меченным красителем. Секвенирующий анализ делает возможной идентификацию основания, присутствующего в сайте двуаллельного маркера. 2) Микросеквенирующий анализ. - 40 - 008252 В микросеквенирующих способах нуклеотид полиморфного сайта в ДНК-мишени выявляют с помощью реакции однонуклеотидного праймерного удлинения. Данный способ включает в себя соответствующие микросеквенирующие праймеры, которые гибридизуются как раз "левее" от представляющего интерес полиморфного основания в нуклеиновой кислоте-мишени. Используют полимеразу, которая специфически удлиняет 3'-конец праймера на один-единственный ddNTP (терминатор цепи), комплементарный нуклеотиду полиморфного сайта. Затем идентичность включенного нуклеотида определяют любым подходящим образом. Обычно реакции микросеквенирования осуществляют с использованием флуоресцентных ddNTP и достроенные микросеквенирующие праймеры анализируют с помощью электрофореза на секвенаторе ABI 377 для определения идентичности встроенного нуклеотида, как описано в ЕР 412883, раскрытие которого включено в данное описание путем ссылки во всей его полноте. Альтернативно, можно использовать капиллярный электрофорез, для того, чтобы одновременно проанализировать большое число продуктов микросеквенирования. Пример типичной микросеквенирующей методики, которую можно использовать в контексте настоящего изобретения, представлен в примере 4. Для меченья и детекции ddNTP можно применить разные методы. Способ гомогенно-фазной детекции основан на флуоресцентном переносе энергии резонанса, описанный Chen and Kwok (1997) и Chen и соавт. (1997). В данном способе амплифицированные фрагменты геномной ДНК, содержащие полиморфные сайты, инкубируют с праймером, меченным флуоресцеином по 5'-концу в присутствии аллель-ных меченных красителем дидезоксирибонуклеозидтрифосфатов и модифицированной Taq-полимеразы. Меченный красителем праймер удлиняется на одно основание с помощью красителя-терминатора, специфичного для данного аллеля, представленного в данной матрице. В конце реакции генотипирования интенсивность флуоресценции двух красителей в данной реакционной смеси анализируют непосредственно без разделения или выделения очисткой. Все эти стадии можно осуществить в одной и той же пробирке, а изменения флуоресценции отслеживать в реальном времени. Альтернативно, удлиненный прай-мер можно анализировать с помощью MALDI-TOF-масс-спектрометрии. Данное основание в полиморфном сайте определяют по массе, добавляемой к праймеру микросеквенирования (см. Haff and Smirnov, 1997). Микросеквенирование можно осуществить с помощью ставшего традиционным или усовершенствованного способа микросеквенирования или с помощью его вариантов. Альтернативные способы включают несколько твердофазных методик микросеквенирования. Основным протоколом по микросеквени-рованию является тот же протокол, который описан выше, за исключением того, что данный способ осуществляют в виде анализа в гетерогенной фазе, в котором данный праймер или молекулу-мишень иммобилизуют или улавливают на твердый носитель. Чтобы упростить анализ отделения праймера и добавления терминирующего нуклеотида, олигонуклеотиды присоединяют к твердому носителю или модифицируют таким образом, чтобы позволить разделение по сродству, а также полимеразное удлинение. 5'-Концы и внутренние нуклеотиды синтетических олигонуклеотидов можно модифицировать несколькими разными путями, позволяющими применять разные методы разделения по сродству, например, биотинилирование. При использовании в олигонуклеотидах единственной аффинной группы, данные олигонуклеотиды можно отделить от встроенного терминирующего реагента. Это исключает необходимость физического разделения или разделения по размеру. При использовании более одной аффинной группы от данного терминирующего реагента можно отделить и анализировать одновременно более одного олигонуклеотида. Это позволяет проводить анализ нескольких видов нуклеиновой кислоты или получать больше информации о последовательности нуклеиновой кислоты на реакцию удлинения. Аффинная группа не является необходимой в праймировании олигонуклеотида, но могла бы быть альтернативно представлена на данной матрице. Например, иммобилизацию можно осуществлять путем взаимодействия между биотинилированной ДНК и микротитрационными лунками, покрытыми стрептавидином или частицами из полистирола, покрытыми авидином. Таким же образом, олигонуклеотиды или матрицы можно присоединить к твердому носителю в высокоплотном виде. В таких твердофазных микросеквени-рующих реакциях встроенные ddNTP могут быть радиоактивно меченными (Syvanen, 1994) или связаны с флуоресцеином (Livak and Hainer, 1994). Детекцию радиоактивно меченных ddNTP можно осуществлять с помощью сцинтилляционных методов. Детекцию флуоресцеинсвязанных ddNTP можно строить на связывании антифлуоресцеинового антитела, конъюгированного со щелочной фосфатазой, с последующей инкубацией с хроматографическим субстратом (таким как паранитрофенилфосфат). Другие возможные пары детекции репортера включают: ddNTP, связанный с динитрофенилом (DNP), и конъюгат анти-DNP щелочная фосфатаза (Harju и соавт., 1993) или биотинилированный ddNTP и стрептавидин, конъюгированный с пероксидазой хрена, с ортофенилендиамином в качестве субстрата (WO 92/15712, раскрытие которого включено в данное описание путем ссылки во всей его полноте). Еще в одном альтернативном твердофазном методе микросеквенирования Nyren и соавт. (1993) описывают способ, основывающийся на детекции ДНК-полимеразной активности в энзиматическом люминометрическом анализе детекции неорганического пирофосфата (ELIDA). Pastinen и соавт. (1997) описывают способ многократной детекции полиморфизма единственного нуклеотида, в котором принцип твердофазного микросеквенирования применяется к формату олигонук - 41 - 008252 леотидного набора. Высокоплотные наборы ДНК-зондов, присоединенные к твердому носителю (ДНК-чип), описаны ниже. Одним из аспектов настоящего изобретения является создание полинуклеотидов и способов гено-типирования одного или нескольких двуаллельных маркеров настоящего изобретения, осуществляемых с помощью анализа микросеквенирования. Предпочтительные праймеры микросеквенирования включают нуклеотидные последовательности D1-D18 и Е1-Е18. Следует иметь ввиду, что микросеквенирующие праймеры, перечисленные в примере 4, являются всего лишь иллюстративными, и что можно использовать любой праймер, обладающий 3'-концом, непосредственно примыкающим к данному полиморфному нуклеотиду. Подобно этому, следует иметь в виду, что микросеквенирующий анализ можно осуществлять для любого двуаллельного маркера или для любого сочетания двуаллельных маркеров настоящего изобретения. Одним из аспектов настоящего изобретения является твердый носитель, который включает один или несколько микросеквенирующих праймеров, перечисленных в примере 4, или их фрагменты, включающие, по крайней мере, 8, 12, 15, 20, 25, 30, 40 или 50 их последовательных нуклеотидов, пока такие совместимые с описанным праймером и обладающие 3'-концом непосредственно "левее" от соответствующего двуаллельного маркера, для определения идентичности нуклеотида в сайте двуаллельного маркера. 3) Основанный на полимеразах и лигазах анализ выявления ошибочного спаривания. Одним из аспектов настоящего изобретения является обеспечение полинуклеотидов и способов для определения аллеля одного или нескольких двуаллельных маркеров настоящего изобретения в биологическом образце с помощью анализа по выявлению ошибочного спаривания, основанного на полимеразах и/или лигазах. Проведение таких анализов основывается на специфичности полимераз и лигаз. Реакция полимеризации определяет особенно жесткие требования к правильному спариванию оснований 3'-конца праймера для амплификации, а соединение двух олигонуклеотидов, гибридизующихся с последовательностью ДНК-мишени, является весьма чувствительным к несоответствиям вблизи сайта лигирования, особенно на 3'-конце. Кроме того, способы, праймеры и различные параметры для амплификации фрагментов ДНК, включающих двуаллельные маркеры настоящего изобретения, описаны выше в разделе "Амплификация фрагментов ДНК, включающих двуаллельные маркеры". Аллельная специфичность праймеров амплификации Дифференциацию аллелей двуаллельного маркера можно осуществлять с помощью специфической амплификации, селективной стратегии, при помощи которой один из аллелей амплифицируется в отсутствие амплификации другого аллеля. Для аллельспецифичной амплификации по меньшей мере один член пары праймеров является достаточно комплементарным области CanIon-гена, включающей полиморфное основание двуаллельного маркера настоящего изобретения, который к тому же гибридизуется и начинает амплификацию. Такие праймеры способны различать эти два аллеля двуаллельного маркера. Это осуществляется путем помещения полиморфного основания на 3'-конец одного из праймеров амплификации. Поскольку удлинение начинается с 3'-конца праймера, ошибочное спаривание в данном положении или вблизи него обладает ингибирующим действием на амплификацию. Поэтому в соответствующих условиях амплификации праймеры направляют амплификацию только на комплементарный к нему аллель. Определение точного расположения ошибочного спаривания и соответствующих условий исследования находятся в пределах компетенции среднего специалиста в данной области техники. Способы, основанные на лигировании/амплификации "Метод олигонуклеотидного лигирования" (OLA) использует два олигонуклеотида, которые создают способными гибридизоваться с примыкающими последовательностями однонитевых молекул-мишеней. Один из олигонуклеотидов биотинилирован, а другой поддается обнаружению благодаря метке. При обнаружении в молекуле-мишени точно комплементарной последовательности, данные олиго-нуклеотиды будут гибридизоваться так, что их концы соединятся впритык и создадут субстрат для лиги-рования, который можно захватить и детектировать. OLA способен детектировать полиморфизм единственного нуклеотида и может быть преимущественно объединен с ПЦР, как описано у Nickerson и соавт. (1990). В данном способе ПЦР используют для осуществления экспоненциальной амплификации ДНК-мишени, которую затем детектируют с использованием OLA. Другие способы амплификации, которые особенно пригодны для обнаружения полиморфизма единственного нуклеотида, включают LCR (лигазную цепную реакцию), Gap LCR (GLCR), которые описаны выше в разделе "Амплификация ДНК". LCR использует две пары зондов для экспоненциальной амплификации конкретной мишени. Последовательности каждой пары олигонуклеотидов выбраны таким образом, чтобы дать возможность паре гибридизоваться с примыкающими последовательностями одной и той же цепи данной мишени. Такая гибридизация образует субстрат для матрично-зависимой лигазы. В соответствии с настоящим изобретением LCR можно осуществить с олигонуклеотидами, обладающими проксимальной и дис-тальной последовательностями одной и той же цепи сайта двуаллельного маркера. В одном из вариантов осуществления настоящего изобретения создают один из двух олигонуклеотидов, который включает сайт двуаллельного маркера. В таком варианте осуществления настоящего изобретения реакционные условия выбраны так, чтобы данные олигонуклеотиды могли быть лигированы вместе лишь в том случае, если - 42 - 008252 данная молекула-мишень либо содержит, либо не содержит специфичный нуклеотид, который комплементарен двуаллельному маркеру в олигонуклеотиде. В альтернативном варианте осуществления настоящего изобретения данные олигонуклеотиды не включают данный двуаллельный маркер, так что, когда они гибридизуются с молекулой-мишенью, создается "gap" (пробел), как описано в WO 90/01069, раскрытие которого включено в данное описание путем ссылки во всей его полноте. Данный пробел "заполняется" комплементарными dNTP (при посредничестве ДНК-полимеразы) или с помощью дополнительной пары олигонуклеотидов. Таким образом, в конце каждого цикла, каждая одиночная цепь обладает комплементом, способным служить в качестве мишени в течение следующего цикла и получают экспоненциальную аллельспецифичную амплификацию желательной последовательности. Ligase/Polymerase-mediated Genetic Bit Analysis(tm) (Анализ небольшого генетического фрагмента при посредничестве лигазы/полимеразы)(tm) представляет собой еще один способ определения идентичности нуклеотида в заранее выбранном сайте молекулы нуклеиновой кислоты (WO 95/21271). Данный способ включает в себя введение нуклеозидтрифосфата, который комплементарен нуклеотиду, присутствующему в заранее выбранном сайте на конце молекулы праймера, и их последующее лигирование со вторым олигонуклеотидом. Данную реакцию отслеживают путем детектирования специфической метки, присоединенной к твердой фазе данной реакции или путем детектирования в растворе. 4) Способы гибридизационного анализа. Предпочтительный способ определения идентичности нуклеотида, присутствующего в сайте двуал-лельного маркера, включает гибридизацию нуклеиновых кислот. Гибридизационные зонды, которые удобно использовать в таких реакциях, предпочтительно включают указанные в данном описании зонды. Можно использовать любой гибридизационный анализ, включая Саузерн-гибридизацию, Нозерн-гибридизацию, дот-блот-гибридизацию и твердофазную гибридизацию (см. Sambrook и соавт., 1989). Гибридизация относится к образованию дуплексной структуры двумя однонитевыми нуклеиновыми кислотами, вследствие комплементарности спариваемых оснований. Гибридизация может происходить между точно комплементарными цепями нуклеиновой кислоты или между цепями нуклеиновой кислоты, которая содержит минорные области ошибочного спаривания. Можно создать специфичные зонды, которые гибридизуются с одной формой двуаллельного маркера, но не с другой, и поэтому способны различать разные аллельные формы. Аллельспецифичные зонды часто используются парами, причем один член такой пары демонстрирует полное совпадение с последовательностью-мишенью, содержащей первоначальный аллель, а другой демонстрирует полное совпадение с последовательностью-мишенью, содержащий альтернативный аллель. Условия гибридизации должны быть достаточно жесткими, чтобы существовала существенная разница в интенсивности гибридизации между аллелями, и, в основном, предпочтителен по существу бинарный ответ, в силу чего зонд гибридизуется только с одним из таких аллелей. Жесткие, специфичные для последовательности, гибридизационные условия, в которых зонд гибридизуется только с точно комплементарной последовательностью-мишенью, хорошо известны в данной области техники (Sambrook и соавт., 1989). Жесткие условия зависят от последовательности и будут отличаться при разных обстоятельствах. Как правило, жесткие условия выбирают таким образом, чтобы они были примерно на 5°С ниже температуры плавления (Tm) специфичной последовательности при определенной ионной силе и рН. Хотя такую гибридизацию можно осуществить в растворе, предпочтительно использовать твердофазный метод гибридизации. ДНК-мишень, включающую дву-аллельный маркер настоящего изобретения, можно амплифицировать перед реакцией гибридизации. Присутствие специфичного аллеля в данном образце определяют путем детектирования наличия или отсутствия стабильных гибридных дуплексов, образованных между данным зондом и данной ДНК-мишенью. Детекцию гибридных дуплексов можно осуществлять многими способами. Хорошо известны разные виды метода детекции, в которых используют детектируемые метки, связанные с данной мишенью или с данным зондом для детекции образованных гибридных дуплексов. Обычно, гибридизацион-ные дуплексы отделяют от негибридизованных нуклеиновых кислот, после чего метки, связанные с образовавшимися дуплексами, детектируются. Специалистам в данной области известно, что стадии отмывки можно использовать для отмывки избытка ДНК-мишени или зонда, а также несвязавшегося конъюгата. Кроме того, стандартные гетерогенные виды анализа являются подходящими для детекции полученных гибридов с использованием меток, присутствующих в праймерах и зондах. Два усовершенствованных в последнее время метода позволяют дискриминировать аллели на основе гибридизации без необходимости разделений или отмывок (см. Landegren U. и соавт., 1998). Преимущество метода TaqMan в том, что в нем используется 5'-нуклеазная активность ДНК-полимеразы Taq для расщепления ДНК-зонда, специфически отжигаемого с накапливающимся продуктом амплификации. TaqMan-зонды метят с помощью донор-акцепторной пары красителей, которые взаимодействуют путем флуоресцентного переноса энергии. Расщепление TaqMan-зонда, в результате продвижения полимеразы в течение амплификации, разъединяет донорный краситель от подавляющего красителя-акцептора, существенно увеличивая флуоресценцию донора. Все реагенты, необходимые для выявления двух аллель-ных вариантов, можно собрать к началу реакции, а результаты отследить в реальном времени (см. Livak и соавт., 1995). В альтернативном методе, основанном на гомогенной гибридизации, для различения аллелей используют молекулярные маяки. Молекулярные маяки представляют собой олигонуклеотидные - 43 - 008252 зонды шпилечной формы, которые сообщают о наличии специфичной нуклеиновой кислоты в гомогенных растворах. При связывании со своими мишенями они подвергаются конформационной реорганизации, которая восстанавливает флуоресценцию внутренне потушенного флуорофора (Tyagi и соавт., 1998). Созданные в данном описании полинуклеотиды можно использовать для получения зондов, которые можно использовать в гибридизационном анализе для детекции аллелей двуаллельного маркера в биологических образцах. Данные зонды характеризуются тем, что они предпочтительно включают 8-50 нуклеотидов и тем, что они достаточно комплементарны последовательности, включающей двуаллель-ный маркер настоящего изобретения, с тем чтобы гибридизоваться с ней, и предпочтительно достаточно специфичны для различения последовательности-мишени всего с одним нуклеотидным изменением. Особенно предпочтительный зонд составляет 25 нуклеотидов в длину. Предпочтительно двуаллельный маркер находится в 4 нуклеотидах от центра данного полинуклеотидного зонда. В особенно предпочтительных пробах двуаллельный маркер находится в центре указанного полинуклеотида. Предпочтительные зонды включают нуклеотидную последовательность, выбранную из группы, состоящей из амплико-нов списка табл. 1 и комплементарных им последовательностей, или ее фрагмент, причем указанный фрагмент включает, по меньшей мере, около 8 последовательных нуклеотидов, предпочтительно 10, 15, 20, более предпочтительно 25, 30, 40, 47 или 50 последовательных нуклеотидов и содержит полиморфное основание. Предпочтительные зонды включают нуклеотидную последовательность, выбранную из группы, состоящей из Р1-Р18 и последовательностей, комплементарных им. В предпочтительных вариантах осуществления настоящего изобретения полиморфное(ые) основание(я) находятся в пределах 5, 4, 3, 2, 1 нуклеотидов от центра указанного полинуклеотида, более предпочтительно - в центре указанного поли-нуклеотида. Предпочтительно зонды настоящего изобретения являются меченными и иммобилизованными на твердом носителе. Метки и твердые носители описаны также в разделе "Олигонуклеотидные зонды и праймеры". Данные зонды могут быть не удлиняемыми, как описано в разделе "Олигонуклеотидные зонды и праймеры". При анализе гибридизации аллельспецифичного зонда можно обнаружить присутствие или отсутствие аллеля двуаллельного маркера в данном образце. Высокопроизводительная параллельная гибридизация в виде набора специфически включена в "методы гибридизации" и описывается ниже. 5) Гибридизация с адресными наборами олигонуклеотидов. Методы гибридизации, основанные на олигонуклеотидных наборах, зависят от различий в гибриди-зационной стабильности коротких олигонуклеотидов, которые полностью совпадают, и несовпадающих вариантов последовательности-мишени. Эффективный доступ к информации о полиморфизме получают через основную структуру, включающую высокоплотные наборы олигонуклеотидных зондов, присоединенных на выбранной позиции к твердому носителю (например, чип). Каждый ДНК-чип может содержать от тысяч до миллионов индивидуальных синтетических ДНК-зондов, выстроенных в подобную сетке структуру и миниатюризированную до размера монеты в 10 центов. Технология чипа уже с успехом применяется во многих случаях. Например, в мутантных штаммах S. cerevisiae скринированию на наличие мутаций был подвергнут ген BRCA 1, протеазный ген в HIV-1-ви-русе (Hacia и соавт., 1996; Shoemaker и соавт., 1996; Kozal и соавт., 1996). Чипы разного вида для обнаружения двуаллельного полиморфизма можно получить на основании индивидуальных заказов у Affymetrix (GeneChip(tm)), Hyseg (HyChip and HyGnostics) и Protogene Laboratories. В общем, в данных способах используют наборы олигонуклеотидных зондов, которые комплементарны сегментам последовательности-мишени нуклеиновой кислоты индивида, последовательности-мишени которого включают полиморфный маркер. ЕР 785280, раскрытие которого включено в данное описание путем ссылки во всей его полноте, описывает "плиточную" стратегию для обнаружения полиморфизма единственного нуклеотида. Вкратце, наборы в основном могут быть "плиточными" для большого числа конкретных полиморфизмов. Под "плиточным" покрытием в основном подразумевается синтез определенного набора олигонуклеотидных зондов, которые создают последовательность, комплементарную представляющей интерес последовательности-мишени, а также заданную изменчивость этой последовательности, например, замена одной или нескольких данных позиций одним или несколькими членами базисного набора нуклеотидов. "Плиточные" стратегии описаны также в заявке РСТ № WO 95/11995. В конкретном аспекте наборы являются "плиточными" для ряда специфичных, идентифицированных последовательностей двуаллельного маркера. В частности, данный набор делают "плиточным" для включения ряда детекционных блоков, каждый из которых является специфичным для конкретного двуаллельного маркера или набора двуаллельных маркеров. Например, детекционный блок может быть " плиточным" для включения ряда зондов, которые охватывают участок последовательности, включающий специфичный полиморфизм. Чтобы обеспечить получение зондов, которые комплементарны каждому аллелю, зонды синтезируют парами, отличающимися по двуаллельному маркеру. Кроме зондов, отличающихся по полиморфному основанию, однозамещенные зонды также, как правило, укладывают " плитками" внутри данного детекционного блока. Такие монозамещенные зонды обладают определенным числом оснований или вплоть до определенного числа оснований в любом направлении от данного - 44 - 008252 полиморфизма, замещенных на остающиеся нуклеотиды (выбранными из А, Т, G, С и U). Обычно зонды в "плиточном" детекционном блоке будут включать замены позиций последовательности вплоть до позиций и, включая позиции, которые находятся в 5 основаниях от двуаллельного маркера. Монозамещен-ные зонды обеспечивают внутренние контроли "плиточного" набора для распознавания действительной гибридизации от артефактной перекрестной гибридизации. По окончании гибридизации с последовательностью-мишенью и отмывки полученной матрицы, данную матрицу сканируют для определения в данной матрице позиции, с которой гибридизуется искомая последовательность-мишень. Затем данные гибридизации сканированной матрицы анализируют, чтобы идентифицировать наличие того или иного аллеля данного двуаллельного маркера в данном образце. Гибридизацию и сканирование можно осуществить так, как описано в заявке РСТ № WO 92/10092 и WO 95/11995 и в патенте США № 5424186. Таким образом, в некоторых вариантах осуществления настоящего изобретения чипы могут включать матрицу последовательностей нуклеиновых кислот с фрагментами длиной около 15 нуклеотидов. В дополнительных вариантах осуществления настоящего изобретения чип может включать матрицу, включающую по меньшей мере одну из последовательностей, выбранных из группы, состоящей из амплико-нов, представленных в табл. 1, и комплементарных им последовательностей, или их фрагментов, причем указанный фрагмент включает по меньшей мере около 8 последовательных нуклеотидов, предпочтительно 10, 15, 20, более предпочтительно 25, 30, 40, 47 или 50 последовательных нуклеотидов и содержит полиморфное основание. В предпочтительных вариантах осуществления настоящего изобретения указанное полиморфное основание находится в пределах 5, 4, 3, 2, 1 нуклеотидов от центра указанного полинуклеотида, более предпочтительно - в центре указанного полинуклеотида. В некоторых вариантах осуществления настоящего изобретения чип может включать матрицу, по меньшей мере, из 2, 3, 4, 5, 6, 7, 8 или более из данных полинуклеотидов настоящего изобретения. Твердые носители и полинуклеоти-ды настоящего изобретения, присоединенные к твердым носителям, описаны также в разделе "Олиго-нуклеотидные зонды и праймеры". 6) Интегрированные системы. Другой метод, который может использоваться для анализа полиморфизмов, включает многосоставные интегрированные системы, которые миниатюризируют и компартаментализуют такие технологические процессы, как ПЦР и реакции капиллярного электрофореза в одном функциональном устройстве. Например, такой метод раскрыт в патенте США № 5589136, раскрытие которого включено в данное описание путем ссылки во всей его полноте, который описывает интеграцию ПЦР-амплификации и капиллярного электрофореза в чипах. Интегрированные системы можно предусматривать, в основном, при использовании микрофлюи-дальных систем. Эти системы включают набор микроканалов, созданных в стекле, силиконе, кварце или в пластиковом тонком диске, включенным в микрочип. Передвижение образцов контролируется электрическими, электроосмотическими или гидростатическими силами, прикладываемыми к разным областям микрочипа для создания функциональных микроскопических волн и насосов в отсутствие движущихся частей. Что касается генотипирования двуаллельных маркеров, то микрофлюидальная система может интегрировать амплификацию нуклеиновых кислот, микросеквенирование, капиллярный электрофорез и способ детекции, такой как флуоресцентная детекция, созданная лазерным излучением. Способы генетического анализа с использованием двуаллельных маркеров настоящего изобретения Для генетического анализа сложных признаков доступны разные способы (см. Lander and Schork, 1994). Поиск генов, чувствительных к заболеванию, осуществляют с использованием двух основных способов: методом сцепления, в котором ищут доказательство совместной сегрегации локуса и предполагаемого локуса, контролирующего признак, с использованием исследования семей, и методом ассоциации, в котором ищут доказательство статистически значимой ассоциации между аллелем и признаком или аллелем, определяющим признак (Knoury и соавт., 1993). В целом, двуаллельные маркеры настоящего изобретения находят использование для любого способа, известного в данной области техники, для демонстрации статистически значимой корреляции между генотипом и фенотипом. Такие двуаллельные маркеры могут использоваться в параметрических и непараметрических способах анализа сцепления. Предпочтительно, двуаллельные маркеры настоящего изобретения используются для идентификации генов, ассоциированных с детектируемыми признаками, с использованием ассоциативных исследований, подхода, который не требует использовать пораженные семьи, и который позволяет идентифицировать гены, ассоциированные с комплексными и отдельными признаками. Генетический анализ с использованием двуаллельных маркеров настоящего изобретения можно осуществлять в любом масштабе. Можно использовать целую совокупность двуаллельных маркеров настоящего изобретения или любую подгруппу двуаллельных маркеров настоящего изобретения, соответствующую гену-кандидату. Кроме того, можно использовать любую совокупность генетических маркеров, включая двуаллельный маркер настоящего изобретения. Совокупность двуаллельных полиморфизмов, которую можно было бы использовать в качестве генетических маркеров в сочетании с двуаллель-ными маркерами настоящего изобретения, описана в WO 98/20165. Как указано выше, следует отметить, что двуаллельные маркеры настоящего изобретения могут быть включены в любую полную или непол - 45 - 008252 ную генетическую карту генома человека. Эти различные применения конкретно рассматриваются в настоящем изобретении и формуле изобретения. Анализ сцепления Анализ сцепления основывается на установлении корреляции между переносом (передачей) генетических маркеров и тем, как конкретный признак передается в поколениях в пределах семьи. Таким образом, целью анализа сцепления является обнаружение маркерного локуса, который демонстрирует одновременное распределение представляющего интерес признака в родословных. Параметрические способы Если данные доступны в ряду последовательных поколений, существует благоприятная возможность исследовать степень сцепления между парами локусов. Оценки доли рекомбинаций делают возможным располагать и размещать локусы на генетической карте. Для локусов, которые представляют собой генетические маркеры, можно создать генетическую карту, после чего можно рассчитать силу сцепления между маркерами и признаками и использовать в качестве указания относительного расположения маркеров и генов, влияющих на эти признаки (Weir, 1996). Классический способ анализа сцепления представляет собой способ логарифмической оценки вероятности (lod) (см. Morton, 1955; Ott, 1991). Вычисление lod-оценок требует детализации схемы наследования для данного заболевания (параметрический способ). Как правило, длина идентифицируемой кандидатной области, с использованием анализа сцепления, составляет между 2 и 20 млн. п. н. После идентификации кандидатной области, как описано выше, анализ рекомбинантных индивидов, с использованием дополнительных маркеров, позволяет дополнительно очертить кандидатную область. Исследование анализа сцепления, в общем, рассчитано максимально на использование 5000 микросателлитных маркеров, ограничивая таким образом максимальную теоретически достигаемую разрешающую способность анализа сцепления в среднем до около 600 т. п. н. Анализ сцепления был успешно применен для картирования простых генетических признаков, которые демонстрируют примеры четкого Менделевского наследования, и которые обладают высокой пе-нетрантностью (т.е. соотношение между числом позитивных носителей аллеля а признака и общим числом носителей а в данной популяции). Однако параметрический анализ сцепления страдает от различных недостатков. Во-первых, он ограничен надежностью в выборе генетической модели, подходящей для каждого исследуемого признака. Более того, как уже упоминалось, достигаемая разрешающая способность, с использованием анализа сцепления, ограничена, и исследование комплементарности требует усовершенствования анализа типичных областей в 2-20 млн.п.н., исходно идентифицированных с помощью анализа сцепления. Кроме того, доказано, что параметрические методы анализа сцепления трудны для применения к комплексным генетическим признакам, таким, которые связанные с объединенным действием множества генов и/или факторов окружающей среды. Очень трудно адекватно моделировать данные признаки методом подсчета lod-балла. В таких случаях требуются чрезмерно большое усилие и затраты для комплектования адекватного количества пораженных болезнью семей, необходимых для анализа сцепления, применительно к этим ситуациям, что недавно обсуждали Risen and Merikangas (1996). Непараметрические способы Преимущество так называемых непараметрических способов для анализа сцепления заключается в том, что они не требуют детализации схемы наследования для данной болезни, они способствуют большему использованию в анализе комплексных признаков. В непараметрических способах предпринимаются попытки доказать, что картина наследования хромосомной области не согласуется со случайным Менделевским расщеплением, демонстрируя, что пораженные болезнью родственники наследуют идентичные копии данной области чаще, чем при случайном ожидании. Пораженные болезнью родственники должны демонстрировать избыток "совместно используемого аллеля" даже в присутствии неполной пе-нетрантности и полигенном наследовании. В непараметрическом анализе сцепления уровень совпадения по маркерному локусу у двух индивидов можно измерить либо с помощью числа аллелей, идентичных по состоянию (IBS), либо с помощью числа аллелей, идентичных по происхождению (IBD). Анализ пораженных болезнью пары сибсов представляет собой хорошо известный специальный случай и является простейшей формой этих способов. Двуаллельные маркеры настоящего изобретения можно использовать и в параметрическом и в непараметрическом анализе сцепления. Предпочтительно двуаллельные маркеры можно использовать в непараметрических способах, которые позволяют картировать гены, участвующие в комплексных признаках. Двуаллельные маркеры настоящего изобретения могут использоваться в IBD- и IBS-способах для картирования генов, воздействующих на комплексный признак. В таких исследованиях, используя преимущество высокой плотности распределения двуаллельных маркеров, несколько смежных локусов дву-аллельных маркеров можно объединить для достижения эффективности, достигаемой мультиаллельны-ми маркерами (Zhao и соавт., 1998). Популяционные ассоциативные исследования Настоящее изобретение включает способы идентификации с использованием двуаллельных маркеров настоящего изобретения в том случае, когда CanIon-ген ассоциирован с выявляемым признаком. В одном из вариантов осуществления настоящее изобретение включает способы выявления ассоциации между аллелем двуаллельного маркера или двуаллельным маркерным гаплотипом и признаком. Кроме - 46 - 008252 того, настоящее изобретение включает способы идентификации аллеля, определяющего признак при неравновесии по сцеплению с любым аллелем двуаллельного маркера настоящего изобретения. Как указано выше, для осуществления ассоциативных исследований можно использовать альтернативные подходы: широкое ассоциативное исследование генома, ассоциативное исследование кандидат-ной области и ассоциативное исследование гена-кандидата. В предпочтительном варианте осуществления настоящего изобретения двуаллельные маркеры настоящего изобретения используются для осуществления ассоциативных исследований кандидатного гена. Указанный метод гена-кандидата несомненно дает быструю оценку идентификации генов и генного полиморфизма, связанного с отдельным признаком, если доступны некоторые сведения по биологии данного признака. Кроме того, двуаллельные маркеры настоящего изобретения можно встроить в любую карту генетических маркеров генома человека для того, чтобы осуществить широкие ассоциативные исследования генома. Способы создания высокоплотной карты двуаллельных маркеров описаны в предварительной патентной заявке США с порядковым номером 60/082614. Двуаллельные маркеры настоящего изобретения можно, кроме того, встроить в любую карту конкретной кандидатной области генома (например, конкретной хромосомы или конкретного хромосомного сегмента). Как указано выше, ассоциативные исследования можно осуществить в генеральной совокупности и не ограничиваться исследованиями, осуществляемыми на родственниках в пораженных болезнью семьях. Ассоциативные исследования представляются чрезвычайно ценными, поскольку позволяют анализировать отдельные или мультифакторные признаки. Более того, ассоциативные исследования представляют собой мощный способ картирования на малом участке гена, обеспечивая более тонкое картирование аллелей, контролирующих признак, чем исследования по сцеплению. Исследования, базирующиеся на родословных, часто лишь сужают местоположение аллеля, контролирующего данный признак. Следовательно, ассоциативные исследования с использованием двуаллельных маркеров настоящего изобретения можно применять для уточнения местоположения аллеля, контролирующего признак, в кан-дидатной области, идентифицируемой способами анализа по сцеплению. Более того, в том случае, если интересующий хромосомный сегмент, представляющий ген-кандидат, такой как ген-кандидат настоящего изобретения, был идентифицирован, для данной представляющей интерес области можно использовать упрощенную идентификацию аллеля, контролирующего данный признак. Двуаллельные маркеры настоящего изобретения можно использовать для демонстрации ассоциации гена-кандидата с признаком. Такое использование, в частности, подразумевается в настоящем изобретении. Определение частоты аллеля двуаллельного маркера или гаплотипа двуаллельного маркера в популяции Ассоциативными исследованиями выясняются взаимосвязи частот аллелей между локусами. Определение частоты аллеля в популяции Аллельные частоты двуаллельных маркеров в популяциях определяют с использованием одного из способов, описанного выше под заголовком "Способы индивидуального генотипирования по двуаллельным маркерам", или любым методом генотипирования, подходящим для данной обозначенной цели. Ге-нотипирование объединенных образцов или индивидуальных образцов может определить частоту аллеля двуаллельного маркера в популяции. Один из путей по уменьшению числа требуемых операций геноти-пирования заключается в использовании объединенных образцов. Основным препятствием для использования объединенных образцов является ограничение точности и воспроизводимости при определении точных концентраций ДНК в собранных пулах. Индивидуальное генотипирование образцов предусматривает более высокую чувствительность, воспроизводимость и точность и является предпочтительным способом настоящего изобретения. Целесообразно генотипировать каждого индивида отдельно, и для определения частоты аллеля двуаллельного маркера или генотипа в данной популяции применяют простой генный подсчет. Настоящее изобретение относится также к способам оценки частоты аллеля в популяции, включающим: а) генотипирование индивидов указанной популяции по указанному двуаллельному маркеру, в соответствии со способом настоящего изобретения; b) определение пропорционального представительства указанного двуаллельного маркера в указанной популяции. Кроме того, способы настоящего изобретения оценки частоты аллеля в популяции включают способы любого дополнительного ограничения, описанного в данном сообщении, или следующие ниже способы, указанные отдельно или в любом сочетании; где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А18 и их комплементов, или, к тому же, двуаллельные маркеры необязательно обнаруживают нарушение равновесия по сцеплению; где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А17 и их комплементов, или, к тому же, двуаллель-ные маркеры необязательно неравновесно сцеплены; где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А12 и А16 и их комплементов, или, к тому же, двуаллельные маркеры необязательно обнаруживают нарушение равновесия по сцеплению. Определение частоты аллеля двуаллельного маркера в популяции можно осуществить, необязательно, в результате определения идентичности нуклеотидов обоих копий указанного двуаллельного маркера, присутствующего в данном геноме каждого индивида в указанной популяции, и вычисления для данной популяции пропорционального представительства указанного нуклеотида по указанному двуаллельному маркеру, - 47 - 008252 связанному с CanIon. Определение пропорционального представительства можно, необязательно, осуществить в результате осуществления способа генотипирования настоящего изобретения в объединенном биологическом образце, полученным от представительного числа индивидов, или от каждого индивида, в указанной популяции, и вычисления пропорционального количества указанного нуклеотида в сравнении с общим количеством. Определение частоты гаплотипа в популяции Гаметическая фаза гаплотипов неизвестна, если диплоидные организмы являются гетерозиготами более чем по одному локусу. Иногда, при использовании генеалогических данных для семей, можно предположить гаметическую фазу (Perlin и соавт., 1994). Если генеалогические данные недоступны, можно применить разные методики. Одна из таких возможностей заключается в том, что гетерозигот-диплоидов по многим сайтам можно исключить из данного анализа, оставляя лишь гомозиготных и гетерозиготных по одному сайту индивидов, однако, данный метод может привести к возможной погрешности относительно структуры образца и недооценке низкочастотных гаплотипов. Другая возможность заключается в том, что одиночные хромосомы можно изучать самостоятельно, например, с помощью асимметричной ПЦР-амплификации (см. Newton и соавт., 1989; Wu и соавт., 1989) или в результате выделения отдельной хромосомы путем предельного разбавления с последующей ПЦР-амплификацией (см. Ruano и соавт., 1990). Кроме того, образец можно гаплотипировать по достаточно близким двуаллельным маркерам с помощью двойной ПЦР-амплификации специфичных аллелей (Sarkar, G. and Sommer S.S., 1991). Такие методы не являются вполне удовлетворительными из-за их технической сложности, влекущей за собой удорожание, их недостаточного обобщения в большом масштабе или из-за возможных смещений. Чтобы преодолеть данные трудности, можно использовать алгоритм для определения фазы генотипов ПЦР-амплифицированных ДНК, предложенный Clark, A.G. (1990). Вкратце, основная идея заключается в начальном заполнении списка гаплотипов, присутствующих в данном образце, в результате обследования индивидов, которые однозначно являются полными гомозиготами и гетерозиго-тами по одному сайту. Затем скринируют других индивидов в том же самом образце на возможную встречаемость ранее определенных гаплотипов. При каждой позитивной идентификации в список установленных гаплотипов добавляют комплементарный гаплотип до тех пор, пока информация по данной фазе для всех индивидов не будет разложена или не будет идентифицирована как далее неразлагаемая. Данным способом для каждого мультигетерозиготного индивида определяют единственный гаплотип, хотя, при наличии более чем одного гетерозиготного сайта, возможны и другие гаплотипы. Альтернативно, можно использовать способы, оценивающие частоты гаплотипов в популяции без определения гаплотипов каждого индивида. Предпочтительно, если способ основывается на алгоритме максимизации математического ожидания (ЕМ) (Dempster и соавт., 1977), лежащего в основе оценок максимального правдоподобия частот гаплотипов при использовании допущения равновесия Харди-Вайнберга (случайное скрещивание) (см. Excoffier L. and Slatkin М. 1995). Данный ЕМ-алгоритм обобщает итерационный метод максимального правдоподобия для оценки, пригодной для противоречивых данных и/или неполных. ЕМ-алгоритм используют для разложения гетерозигот на гаплотипы. Оценки гаплотипов дополнительно описываются ниже под заголовком "Статистические методы". Можно использовать любой другой метод, известный в данной области техники, для определения частоты гаплотипа в популяции. Настоящее изобретение включает также способы определения частоты гаплотипа для совокупности двуаллельных маркеров в популяции, включающие следующие стадии: а) генотипирование по меньшей мере одного двуаллельного маркера, связанного с CanIon, в соответствии со способом настоящего изобретения для каждого индивида в указанной популяции; b) генотипирование второго двуаллельного маркера в результате определения идентичности нуклеотидов по указанному второму двуаллельному маркеру для обоих копий указанного второго двуаллельного маркера, представленного в геноме каждого индивида в указанной популяции: и с) применение способа определения гаплотипа к идентичным нуклео-тидам, установленным на стадиях а) и b) для получения оценки указанной частоты. Кроме того, способы определения частоты гаплотипа настоящего изобретения включают способы с любым дополнительным ограничением, описанным в данном сообщении, или следующие ниже способы, указанные отдельно или в любом сочетании: где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А18 и их комплементов, или двуаллельные маркеры необязательно, к тому же, сцеплены при нарушении равновесия; где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А17 и их комплементов, или двуаллельные маркеры необязательно, к тому же, демонстрируют нарушение равновесия по сцеплению; где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А12 и А16 и их комплементов, или двуаллельные маркеры необязательно, к тому же, неравновесно сцеплены. Указанный способ определения гаплотипа необязательно осуществляют с помощью асимметричной ПЦР-амплификации, двойной ПЦР-амплификации специфичных аллелей, алгоритма Clark или алгоритма максимизации математического ожидания. Анализ сцепления при нарушении равновесия Нарушение равновесия по сцеплению представляет собой неслучайную ассоциацию аллелей по двум или более локусам и является мощным инструментом для картирования генов, участвующих в кон - 48 - 008252 троле признаков заболевания (см. Ajioka R.S. и соавт., 1997). Двуаллельные маркеры, поскольку они компактно расположены в геноме человека и могут быть генотипированы в большем числе случаев по сравнению с другими видами генетических маркеров (такими, как RFLP- или VNTR-маркеры), особенно пригодны для генетического анализа, основанного на изучении равновесия по сцеплению. Когда мутация заболевания впервые внедряется в популяцию (посредством новой мутации или иммиграции носителя мутации), она непременно находится в отдельной (одной) хромосоме и, поэтому, в отдельной генетической "среде" или "анцестральном" (предковом, родовом) гаплотипом связанных маркеров. Следовательно, между такими маркерами и мутацией, контролирующей заболевание, существует полное нарушение равновесия: мутация, контролирующая заболевание, обнаруживается только в присутствии специфичной группы маркерных аллелей. В течение жизни последовательных поколений между мутацией, контролирующей заболевание, и таким маркерным полиморфизмом происходят рекомби-нантные события, и указанное нарушение равновесия постепенно исчезает. Скорость этого исчезновения является функцией частоты рекомбинации, так что маркеры, ближе всего расположенные к гену, контролирующему заболевание, будут обнаруживать более высокую степень нарушения равновесия, чем отдаленные маркеры. Если не происходит нарушения в результате рекомбинаци, то "анцестральные" гапло-типы и нарушение равновесия по сцеплению между маркерными аллелями разных локусов могут прослеживаться не только в родословных, но также и в популяциях. Нарушение равновесия по сцеплению обычно наблюдается в виде ассоциации между одним специфичным аллелем одного локуса и другим специфичным аллелем второго локуса. Паттерн или кривая нарушения равновесия между заболеванием и маркерным локусом, как ожидается, демонстрирует максимум, который наблюдается по локусу, контролирующему заболевание. Следовательно, степень сцепления при нарушении равновесия между аллелем, контролирующим заболевание, и тесно связанными генетическими маркерами может давать ценную информацию, относящуюся к местоположению гена, контролирующего данное заболевание. Для тонкого картирования локуса, контролирующего заболевание, полезно обладать определенным знанием картины сцепления при нарушении равновесия, которое существует между маркерами в исследуемой области. Как указано выше, уровень картирования, осуществляемый посредством анализа сцепления при нарушении равновесия, представляется более высоким, чем в исследованиях по сцеплению. Высокая плотность распределения двуаллель-ных маркеров, сочетаемая с анализом сцепления при нарушении равновесия, создает мощный инструмент для тонкого картирования. Разные способы, которые рассчитывают сцепление при нарушении равновесия, описаны ниже под заголовком "Статистические методы". Популяционные исследования ассоциаций признак-маркер у больных и здоровых Как указано выше, встречаемость пар специфичных аллелей разных локусов в одной и той же хромосоме не является случайным, а отклонение от случайности называют нарушением равновесия по сцеплению или сцеплением при нарушении равновесия. Ассоциативные исследования сосредоточены на по-пуляционных частотах и опираются на феномен нарушении равновесия по сцеплению. Если специфичный аллель данного гена непосредственно контролирует конкретный признак, его частота будет статистически выше в пораженной болезнью популяции индивидов (признак позитивный), по сравнению с частотой в популяции с негативным признаком или в случайной контрольной (здоровой) популяции. Из-за наличия нарушения равновесия по сцеплению частота всех других аллелей, представленных в данном гаплотипе, несущем аллель, контролирующий признак, будет также повышенной у индивидов с позитивным признаком по сравнению с индивидами с негативным признаком или в рандомизированной контрольной выборке. Поэтому, ассоциация между данным признаком и любым аллелем (особенно, аллелем двуаллельного маркера) при нарушении равновесия по сцеплению с аллелем, определяющим признак, будет достаточной, чтобы предположить наличие гена, контролирующего признак, в этой конкретной области. Популяции больных и здоровых можно генотипировать по двуаллельным маркерам для идентификации ассоциаций, которые точно определяют местонахождение аллеля, определяющего признак. Так любой маркер при нарушении равновесия по одному данному маркеру, связанному с признаком, будет ассоциирован с данным признаком. Нарушение равновесия по сцеплению позволяет соответствующие частоты в популяциях больных и здоровых с ограниченным числом генетических полиморфизмов (особенно двуаллельных маркеров) анализировать в качестве альтернативных при скринировании всех возможных функциональных полиморфизмов для того, чтобы найти аллели, определяющие признак. Ассоциативные исследования сопоставляют частоту маркерных аллелей в несвязанных родством популяциях больных и здоровых, и представляют мощный инструмент для изучения комплексных признаков. Популяции больных и здоровых (критерии включения) Популяционные исследования, основанные на ассоциации, не затрагивают наследование в семьях, а сопоставляют распространение отдельного генетического маркера, или группы маркеров, в популяциях больных и здоровых. Они представляют собой исследования больных и здоровых, основанные на сравнении не связанных родством больных (пораженных заболеванием или позитивные по признаку) индивидов и не связанных родством здоровых (непораженных, отрицательных по признаку или случайных) индивидов. Предпочтительно, если контрольная группа (здоровых) составлена из непораженных заболеванием или негативных по признаку индивидов. Кроме того, данная контрольная группа этнически со - 49 - 008252 поставима с популяцией людей, пораженных заболеванием. Более того, данная контрольная группа предпочтительно сопоставима с популяцией больных по основному известному фактору смешения для данного признака в исследовании (например, подобрана по возрасту или по зависимому от возраста признаку). Теоретически, индивидов по двум образцам объединяют таким образом, чтобы между ними ожидались различия только по своему статусу заболевания. Термины "популяция с позитивным признаком", "популяция больных" и "популяция пораженных заболеванием" используются в данном описании как взаимозаменяемые. Существенной ступенью в изучении комплексных признаков с использованием ассоциативных исследований является подбор популяций больных и здоровых (см. Lander and Schork, 1994). Основное действие при подборе популяций больных и здоровых заключается в клиническом определении данного признака или фенотипа. Любой генетический признак можно проанализировать ассоциативным способом, предложенным в данном описании, путем тщательного подбора индивидов, которые фенотипически включаются в группы с позитивным признаком и негативным признаком. Чаще используют четыре критерия: клинический фенотип, возраст при возникновении заболевания, история семьи и отягощенность. Процедура отбора непрерывных или количественных признаков (таких как, например, артериальное давление) включает отбор индивидов на краях распределения фенотипа при исследовании данного признака так, чтобы включить в данные позитивные и негативные популяции неперекрывающиеся фенотипы. Предпочтительно, если популяции больных и здоровых составляют фенотипически гомогенные популяции. Позитивные и негативные популяции по данному признаку включают фенотипически однородные популяции индивидов, соответствующие в исследовании, каждая, 1-98%, предпочтительно 1-80%, более предпочтительно 1-50%, еще более предпочтительно 1-30%, наиболее предпочтительно 1-20% от общей популяции, и предпочтительно выбраны из индивидов, демонстрирующих неперекрывающиеся фенотипы. Чем четче разница между двумя фенотипами по данному признаку, тем больше вероятность обнаружения ассоциации с двуаллельными маркерами. Отбор таких резко отличных и, вместе с тем, относительно однородных фенотипов делает возможным эффективное сравнение в ассоциативных исследованиях и вероятное обнаружение четких различий на генетическом уровне, при условии, что размеры по-пуляционных выборок для исследования довольно значительны. В предпочтительных вариантах осуществления настоящего изобретения первая группа из 50-300 индивидов с позитивным признаком, предпочтительно около 100 индивидов, набрана в соответствии с их фенотипами. Аналогичное количество контрольных индивидов также включены в такие исследования. Ассоциативный анализ Настоящее изобретение включает также способы выявления ассоциации между генотипом и фенотипом, включающие следующие стадии: а) определение частоты, по меньшей мере, одного двуаллельно-го маркера, связанного с CanIon, в популяции с позитивным признаком, в соответствии со способом ге-нотипирования настоящего изобретения; b) определение частоты указанного двуаллельного маркера, связанного с CanIon, в контрольной популяции, в соответствии со способом генотипирования настоящего изобретения; и с) определение существования статистически значимой ассоциации между указанным генотипом и указанным фенотипом. Кроме того, указанные способы выявления ассоциации между генотипом и фенотипом в настоящем изобретении включают способы для любого дополнительного ограничения, описанного в данном описании, или следующие ниже способы, указанные отдельно или в любом сочетании: где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А18 и их комплементов, или, к тому же, двуаллельные маркеры необязательно неравновесно сцеплены; где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А17 и их комплементов, или, к тому же, двуаллельные маркеры необязательно неравновесно сцеплены; указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А12 и А16 и их комплементов, или, к тому же, двуаллельные маркеры необязательно неравновесно сцеплены. Указанная контрольная популяция может необязательно представлять собой популяцию с негативным признаком или произвольную популяцию. Каждую из указанных стадий генотипирования а) и b) необязательно осуществлять на объединенном биологическом образце, полученном от каждой указанной популяции. Каждую из указанных стадий генотипирования а) и b) необязательно осуществлять отдельно на биологических образцах, полученных от каждого индивида в указанной популяции или в ее субвыборке. Общая методология осуществления ассоциативных исследований с использованием двуаллельных маркеров, полученных из области, несущей ген-кандидат, заключается в обследовании двух групп индивидов (популяции больных и здоровых) с тем, чтобы измерить и статистически сравнить аллельные частоты двуаллельных маркеров настоящего изобретения в обоих группах. Если статистически значимая ассоциация с признаком идентифицируется, по меньшей мере, для одного или более анализируемых двуаллельных маркеров, то следует предположить, что, либо ассоциируемый аллель непосредственно ответствен за определяемый им признак (т. е. ассоциированный аллель представляет собой аллель, определяющий данный признак), либо, что более вероятно, данный ассоциированный аллель неравновесно сцеплен с аллелем, определяющим данный признак. Специфические характеристики ассоциированного аллеля относительно функции гена-кандидата, как правило, дают до - 50 - 008252 полнительное представление о взаимосвязи ассоциированного аллеля и данного признака (причинно-следственную взаимосвязь или сцепление при нарушении равновесия). Если данные факты указывают, что данный ассоциированный аллель в данном гене-кандидате, вероятнее всего, не является аллелем, определяющим данный признак, но вместе с тем, неравновесно сцеплен с подлинным аллелем, определяющим данный признак, тогда аллель, определяющий данный признак, можно обнаружить путем сек-венирования вблизи ассоциированного маркера и осуществить дальнейшие ассоциативные исследования полиморфизма, который обнаружен итеративным образом. Ассоциативные исследования осуществляют, как правило, в две последовательные стадии. В первой фазе (на первой стадии) частоты приведенных двуаллельных маркеров гена-кандидата определяют для популяций с позитивным признаком и для контрольных популяций. Во второй фазе (на второй стадии) анализа позицию генетических локусов, ответственных за данный признак, дополнительно уточняют с использованием маркеров повышенной плотности из значимой области. Однако, если исследуемый ген-кандидат относительно невелик по длине, как в случае с CanIon, одной стадии может оказаться достаточно для установления значимых ассоциаций. Анализ гаплотипов Как указано выше, когда хромосома, несущая аллель заболевания, впервые появляется в популяции, то ли в результате мутации, то ли миграции, мутантный аллель непременно находится на хромосоме, обладающей группой сцепленных маркеров: анцестральный гаплотип. Данный гаплотип можно проследить в популяциях и проанализировать его статистическую ассоциацию с данным признаком. Исследования комплементирующей одиночной (аллельная) ассоциации вместе с мультиточечными ассоциативными исследованиями, именуемыми также гаплотипными исследованиями, повышают статистическую мощность ассоциативных исследований. Поэтому, изучение гаплотипной ассоциации позволяет определить частоту и тип анцестрального носителя гаплотипа. Анализ гаплотипа важен тем, что он повышает возможность статистического анализа рассматриваемых индивидуальных маркеров. На первой стадии частотного анализа гаплотипов определяют частоту возможных гаплотипов на основе различных сочетаний идентифицированных двуаллельных маркеров настоящего изобретения. Затем данную частоту гаплотипов сравнивают в отдельных популяциях индивидов с позитивным признаком и контрольными. Количество индивидов с позитивным признаком, в большинстве случаев между 30 и 300, с предпочтительной численностью индивидов, колеблющейся между 50 и 150, подвергают данному анализу для получения статистически значимых результатов. Эти же соображения применяют к используемым в данном исследовании численностям индивидов, непораженных заболеванием (или случайно выбранным здоровым индивидам). По результатам этого первого анализа получают частоты гап-лотипа для популяций больных и здоровых, для каждой определяют частоту гаплотипа, р-значение и рассчитывают соотношение баллов. При обнаружении статистически значимой ассоциации относительный риск для индивида, несущего данный гаплотип, который соответствует пораженному данным исследуемым признаком, может быть аппроксимирован (приблизительно точным). Дополнительный вариант осуществления настоящего изобретения включает способы обнаружения ассоциации между гаплотипом и фенотипом, включающими стадии: а) оценки частоты, по меньшей мере, одного гаплотипа в популяции с позитивным признаком, в соответствии со способом настоящего изобретения оценки частоты гаплотипа; b) оценки частоты указанного гаплотипа в контрольной популяции, в соответствии со способом настоящего изобретения оценки частоты гаплотипа; и с) определения статистически значимой ассоциации, существующей между указанным гаплотипом и указанным фенотипом. Кроме того, данные способы обнаружения ассоциации между гаплотипом и фенотипом настоящего изобретения включают способы с любым дополнительным ограничением, где указанный двуал-лельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А18 и их комплементов, или данные двуаллельные маркеры необязательно, к тому же, неравновесно сцеплены; где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А1-А17 и их комплементов, или данные двуаллельные маркеры необязательно, к тому же, обнаруживают нарушение равновесия по сцеплению; где указанный двуаллельный маркер, связанный с CanIon, необязательно выбран из группы, состоящей из А12 и А16 и их комплементов, или данные двуаллельные маркеры необязательно, к тому же, неравновесно сцеплены. Указанная контрольная популяция необязательно представляет собой негативную по признаку популяцию или произвольную популяцию. Указанный способ необязательно включает дополнительные стадии определения фенотипа в указанной позитивной по признаку популяции и в указанной контрольной популяции перед стадией с). Анализ взаимодействия Двуаллельные маркеры настоящего изобретения можно также использовать для идентификации паттернов двуаллельных маркеров, ассоциированных с выявляемыми признаками, полученными в полигенном взаимодействии. Анализ генетического взаимодействия между аллелями несцепленных локусов требуют индивидуального генотипирования с использованием описанных в данном описании методов. Анализ аллельного взаимодействия в выбранной совокупности двуаллельных маркеров с соответствующим уровнем статистической значимости можно рассматривать в качестве гаплотипного анализа. Анализ взаимодействия включает стратификацацию популяций больных и здоровых относительно данного гаплотипа - 51 - 008252 для первого локуса и осуществление гаплотипного анализа по второму локусу в каждой субпопуляции. Статистические методы, используемые для исследования ассоциации, дополнительно описаны ниже. Проверка сцепления при наличии ассоциации Двуаллельные маркеры настоящего изобретения могут использоваться, кроме того, в TDT (тест передача/нарушение равновесия). TDT проверяет сцепление и ассоциацию и не изменяется при популяци-онной стратификации. TDT нуждается в данных по пораженным заболеванием индивидам и их родителям или в данных от непораженных заболеванием сибсов, но не их родителей (см. Spielman S. и соавт., 1993; Schaid D.J. и соавт., 1996, Spielman S. and Ewens W.J., 1998). Такие объединенные тесты уменьшают, как правило, количество ложно-позитивных ошибок при раздельном анализе. Статистические методы Вообще, любой способ, известный в данной области техники, можно использовать для проверки статистически значимой корреляции признака и генотипа. 1) Способы анализа сцепления. Статистические методы и компьютерные программы, пригодные для анализа сцепления, хорошо известны специалистам в данной области техники (см. Terwilliger J.D. and Ott J., 1994; Ott J., 1991). 2) Способы оценки частот гаплотипов в популяции. Как указано выше, при подсчете генотипов часто невозможно отличить гетерозиготы так, чтобы можно было легко подсчитать частоты гаплотипов. Если гаметическая фаза неизвестна, то оценить гап-лотипные частоты можно из мультилокусных генотипических данных. Любой способ, известный специалисту в данной области техники, можно использовать для оценки гаплотипных частот (см. Lange К., 1997; Weir, B.S., 1996). Предпочтительно, максимально-правдоподобные частоты гаплотипов подсчитывают с использованием алгоритма максимизации математического ожидания (ЕМ) (см. Dempster и соавт., 1977; Excoffier L. and Slatkin M., 1995). Данная операция представляет собой итерационный процесс, нацеленный на получение максимально правдоподобных оценок частот гаплотипов по данным мультило-кусного генотипа, когда гаметическая фаза неизвестна. Гаплотипные оценки осуществляют, как правило, путем применения ЕМ-алгоритма с использованием, например, EM-HAPLO-программы (Hawley M.E. и соавт., 1994) или Arlequin-программы (Schneider и соавт., 1997). ЕМ-алгоритм представляет собой обобщенный итерационный метод максимального правдоподобия для оценивания и кратко описан ниже. Следует учесть, что в представленном разделе "Способы оценки частот гаплотипов в популяции" фенотипы относятся к мультилокусным генотипам неизвестной гаплотипной фазы. Генотипы относятся к мультилокусным генотипам известной гаплотипной фазы. Предположим, имеется выборка из N не связанных родством индивидов, типированных по К маркерам. Данные исследования представлены К-локусными фенотипами неизвестной фазы, которые можно распределить в качестве F разных фенотипов. Далее, допустим, что мы имеем H возможных гаплотипов (для случая К двуаллельных маркеров мы имеем максимальное число возможных гаплотипов Н=2К). Для фенотипа] с Cj возможных генотипов имеем: Р, =%РК*енотип(0) = i> (**A) м Уравнение 1 где Pj означает вероятность фенотипа j, a P(hk,hi) означает вероятность генотипа i, составленного гап-лотипами hk и hi. При случайном скрещивании (т.е. равновесие Харди-Вайнберга), P(hk,hi) выражено в виде: P(hk,hl) = P(hk)2для Ьь = Ь P(hk ,h,) = 2P(hk )P(ht) для hk Ф h{ Уравнение 2 Е-М-алгоритм состоит из следующих стадий. Первая, генотипические частоты оценивают из совокупности исходных значений гаплотипных частот. Эти гаплотипные частоты обозначают как P1(0), Р2(0), Р3(0), Рн(0). Исходные значения гаплотипных частот можно получить с помощью генератора случайных чисел или каким-либо иным образом, хорошо известным в данной области техники. Данная стадия называется стадией Ожидания. Следующая стадия данного способа, названная стадией Максимизации, заключается в использовании оценок генотипических частот, которые пересчитаны для гаплотипных частот. Первая итерация оценок гаплотипной частоты обозначается при помощи P1(1), Р2(1), Р3(1), Рл(1). В итоге стадия Ожидания на итерации s состоит из вычисления вероятности размещения каждого фенотипа в различные вероятностные генотипы, основанные на гаплотипных частотах в предшествующей итерации: Уравнение 3 где nj равно числу индивидов с фенотипом j, a Pj(hk,hi)(s) является вероятностью генотипа hk,hi в фенотипе j. На стадии Максимизации, которая эквивалентна методу подсчета генов (Smith, АПП. Hum. Genet., 21:254-276, 1957), гаплотипные частоты переоценивают на основании генотипических оценок: - 52 - 008252 Jmi Уравнение 4 где 5it представляет собой показатель изменчивости, в котором подсчитывается величина встречаемости, с которой гаплотип t представлен в генотипе i; он имеет значения 0, 1 и 2. ЕМ-итерации прекращают при достижении следующего критерия. При использовании расчетных оценок по методу максимального правдоподобия (MLE) можно допустить, что фенотипы j распределяются полиномиально. В каждой итерации s можно вычислить одну L-функцию правдоподобия. Сходимость достигается при разнице лог-правдоподобия между двумя последовательными итерациями, равной некоему малому числу, предпочтительно 10-7. 3) Способы вычисления сцепления между маркерами при нарушении равновесия. Ряд методов могут использоваться для вычисления сцепления при нарушении равновесия по между любыми двумя генетическими позициями, практически сцепление при нарушении равновесия оценивают, используя статистический тест на ассоциацию для гаплотипных данных, взятых из популяции. Сцепление при нарушении равновесия между любой парой двуаллельных маркеров, включающей по меньшей мере один из двуаллельных маркеров настоящего изобретения (Mi, Mj), имеющих аллели (ai/bi) по маркеру Mi и аллели (ajbj) по маркеру Mj, можно вычислить для каждой аллельной комбинации (a;,aj; a;,bj; и b;,aj), в соответствии с формулой Piazza: Amj= V04 - V (94 + 63) (04 +92) где 64=__=частоте генотипов, не обладающих аллелем ai для Mi и не обладающих аллелем aj для Mj 63=_+=частоте генотипов, не обладающих аллелем ai для Mi и обладающих аллелем aj для Mj 62=+_=частоте генотипов, обладающих аллелем ai для Mi и не обладающих аллелем aj для Mj Сцепление при нарушении равновесия (LD) между парами двуаллельных маркеров (Mi, Mj) можно также вычислить для каждой аллельной комбинации (ai, aj; ai, bj; bi,aj и bi,bj) в соответствии с оценкой максимального правдоподобия (MLE) для дельта (объединенный коэффициент нарушения генотипиче-ского равновесия), как описано у Weir (Weir B.S., 1996). MLE для объединенного коэффициента сцепления при нарушении равновесия равна: Daiaj= (2nj + n2 + п3 + iu/2)/N - 2(рг(а;). pr(aj)) где n1=E фенотипа (ai/ai, aj/aj). n2=E фенотипа (ai/ai, aj/bj), n3=E фенотипа (ai/bi, aj/aj), n4=E фенотипа (ai/bi, aj/bj) и N равно числу индивидов в данной выборке. Данная формула позволяет оценить нарушение равновесия по сцеплению между аллелями, только при наличии данных по генотипу, а не по гаплотипу. Другой способ вычисления сцепления при нарушении равновесия между маркерами заключается в следующем. Для сцепленных двуаллельных маркеров, Mi(ai/bi) и Mj(aj/bj), аппроксимирующих равновесие Харди-Вайнберга, можно оценить частоты четырех возможных гаплотипов в данной популяции в соответствии с методом, описанным выше. Оценка гаметического нарушения равновесия между ai и aj проста: Daiaj = pr( womun (а,-, aj )) - pr (a f- ).pr(a j ) где pr(ai) означает вероятность аллеля a pr(aj) означает вероятность аллеля a, и где РГ( гаплотип (ai,aj)) оценивается, как указано выше в уравнении 3. Для сцепленного двуаллельного маркера необходимо провести только одно измерение нарушения равновесия, чтобы описать ассоциацию между Mi и Mj. Затем нормируют полученное выше значение и рассчитывают следующим образом: D'aiaj = Daiaj / ШЭХ (-prfa). prfcj) , -pr(bj). pr(bj)) при Daiaj <0 D'aiaj = Daiaj / max (pr(bj). pr(aj) , pr(aj). pr(bj)) при Daiaj> 0 Специалистам легко понять, что можно использовать и другие способы вычисления сцепления при нарушении равновесия. Сцепление при нарушении равновесия в группе двуаллельных маркеров, обладающих адекватным уровнем гетерозиготности, можно определить путем генотипирования 50-1000 не связанных родством индивидов, предпочтительно 75-200, более предпочтительно около 100. 4) Проверка на ассоциацию. Способы определения статистической значимости корреляции между фенотипом и генотипом, в данном случае аллеля в двуаллельном маркере или гаплотипа, слагаемых из таких аллелей, можно найти с помощью любого статистического теста, известного в данной области техники при любом приемлемом пороге статистической значимости, который требуется. Применение конкретных способов и порогов значимости находится в компетенции среднего специалиста в данной области техники. Проверку на ассоциацию осуществляют путем определения частоты аллеля двуаллельного маркера в больной и контрольной популяциях и сравнения данных частот с помощью статистического критерия, чтобы определить является ли их различие частот статистически значимым, что должно указывать на - 53 - 008252 корреляцию между исследуемым данным признаком и аллелем двуаллельного маркера. Подобным образом осуществляют анализ гаплотипа путем определения частот всех возможных гаплотипов для данной группы двуаллельных маркеров в больной и контрольной популяциях, и сравнивают эти частоты с помощью статистического критерия, чтобы определить, соответствует ли их статистически значимое различие корреляции между исследуемым гаплотипом и фенотипом (признаком). Можно использовать любой статистический инструмент, пригодный для проверки статистически значимой ассоциации между генотипом и фенотипом. В качестве статистического критерия предпочтительно использовать критерий хи-квадрат с одной степенью свободы. Вычисляют Р-значение (Р-значение является вероятностью того, что статистический результат большой или больше, чем наблюдаемая величина для случайного события). Статистическая значимость В предпочтительных вариантах осуществления настоящего изобретения, значимость для целей диагностики, либо в качестве позитивного основания для дальнейших диагностических тестов или в качестве предварительной начальной точки для ранней превентивной терапии, р-значение, связанное с ассоциацией двуаллельного маркера, составляет предпочтительно около 1 х 10-2 или меньше, более предпочтительно около 1 х 10-4 или меньше, для анализа одного двуаллельного маркера, и около 1 х 10-3 или меньше, еще более предпочтительно 1 х 10-6 или меньше и наиболее предпочтительно около 1 х 10-8 или меньше, для гаплотипного анализа, рассматривающего два или более маркера. Считают, что данные значения приложимы к любым исследованиям по ассоциации, рассматривающим одиночный маркер или их множественное сочетание. Специалисты могут использовать область значений, изложенных выше, в качестве исходной точки для осуществления исследований по ассоциации с двуаллельными маркерами настоящего изобретения. При этом могут обнаружиться значимые ассоциации между двуаллельными маркерами настоящего изобретения и признаком, которые используются для целей диагностики и лекарственного скринирования. Фенотипическая перестановка Чтобы подтвердить обнаруженную выше статистическую значимость гаплотипного анализа на первой стадии, можно было бы осуществить последующий анализ, в котором данные по больным и здоровым индивидам объединяются и рэндомизируются в отношении данного фенотипического признака. Данные каждого индивидуального генотипирования распределяются случайным образом на две группы, которые содержат одинаковое число индивидов по больной и здоровой популяциям, используемых для составления данных, полученных на первой стадии. Вторая стадия гаплотипного анализа предпочтительно осуществляется на этих искусственных группах, предпочтительно по маркерам, включенным в данный гаплотипный анализ первой стадии, демонстрируя наивысший относительный коэффициент риска. Данный эксперимент повторяют, предпочтительно, по меньшей мере, 100-1000 раз. Повторяемые итерации позволяют определить вероятность случайного тестирования гаплотипа. Определение статистической ассоциации Обращаясь к проблеме ложных позитивов, можно осуществить сходный анализ для случайных геномных областей у тех же больных и контрольных популяциях. Результаты для случайных областей и кандидатной области сравнивают, как описано в находящейся на рассмотрении предварительной патентной заявке США с названием "Способы, программное и аппаратное обеспечение для идентификации геномных областей гена, ассоциированного с обнаруживаемым признаком", порядковый номер 60/107986, поданной 10 ноября 1998г., содержание которой включено в данное описание путем ссылки. 5) Оценка факторов риска. Ассоциация между фактором риска (в генетической эпидемиологии фактор риска подразумевает наличие или отсутствии некоего аллеля или гаплотипа в маркерных локусах) и заболеванием измеряется соотношением шансов (OR) и относительным риском (RR). Когда P(R+) представляет собой вероятность развития заболевания у индивидов с R, a P(R-) представляет собой вероятность для индивидов в отсутствие фактора риска, тогда относительный риск равен просто соотношению двух вероятностей, то есть: RR=P(R+)/P(R) В исследованиях заболевание-контроль прямое измерение относительного риска осуществить невозможно из-за выборочного обследования. Однако соотношение шансов обеспечивает достаточно хорошую аппроксимацию относительного риска для редко встречающихся заболеваний и может быть вычислено: " F+ 1-F\ OR=(F+/(l-F+))/(FV(l-F-)) F+ представляет собой частоту выявляемости фактора риска в популяции больных, a F- представляет собой частоту выявляемости фактора риска в контроле. F+ и F- вычисляют с использованием аллельных или гаплотипных частот в данном исследовании и зависят от лежащей в основании генетической модели (доминантной, рецессивной, аддитивной...). Можно также оценить атрибутивный риск, который описывает пропорцию индивидов в популяции, обнаруживающей признак, связанный с данным риском. Данное измерение представляется важным для - 54 - 008252 количественного определения роли специфичного фактора в этиологии заболевания и в рамках влияния фактора риска на здравоохранение. Важность данного измерения для здравоохранения лежит в определении доли случаев заболевания в данной популяции, которые можно было бы предупредить, если бы не отсутствие проявления интереса. AR определяют следующим образом: AR=PE (RR-1) / (РЕ (RR-1) +1) AR означает риск, свойственный аллелю двуаллельного маркера или гаплотипу двуаллельного маркера. РЕ означает частоту проявления аллеля или гаплотипа в популяции в целом; a RR означает относительный риск, который, являясь аппроксимированным для соотношения шансов, при исследовании данного признака, обладает относительно низкой частотой в генеральной совокупности. Идентификация двуаллельных маркеров при нарушении равновесия для двуаллельных маркеров настоящего изобретения После идентификации первого двуаллельного маркера в представляющей интерес геномной области, специалисты в данной области техники, используя указания настоящего изобретения, могут легко идентифицировать дополнительные двуаллельные маркеры, сцепленные при нарушении равновесия с данным первым маркером. Как указано ранее, любой маркер, сцепленный при нарушении равновесия с первым маркером, ассоциированным с признаком, будет ассоциироваться с данным признаком. Поэтому, после ассоциации, продемонстрированной между данным двуаллельным маркером и признаком, обнаружение дополнительных двуаллельных маркеров, ассоциированных с данным признаком, представляет большой интерес для того, чтобы увеличить плотность двуаллельных маркеров в данной конкретной области. Каузальный ген или мутация будут обнаруживаться вблизи данного маркера или группы маркеров, демонстрируя наивысшую корреляцию с данным признаком. Идентификация дополнительных маркеров в сцеплении при нарушении равновесия с данным маркером включает: (а) амплификацию геномного фрагмента, включающего первый двуаллельный маркер, у множества индивидов; (b) идентификацию вторых двуаллельных маркеров в данной геномной области, несущей указанный первый двуаллельный маркер; (с) проведение анализа нарушения равновесия сцепления между указанным первым двуаллельным маркером и вторыми двуаллельными маркерами; и (d) отбор указанного второго двуаллельного маркера, который сцеплен при нарушении равновесия с указанным первым маркером. Рассматриваются также субкомбинации, включающие стадии (b) и (с). Способы идентификации двуаллельных маркеров и проведения описанного в данном описании анализа сцепления при нарушении равновесия может осуществить специалист без чрезмерного экспериментирования. Настоящее изобретение рассматривает также двуаллельные маркеры, которые находятся в состоянии нарушения равновесия по сцеплению с двуаллельными маркерами А1-А18 и которые, как ожидается, представлены аналогичными характеристиками в рамках их соответствующей ассоциации с данным признаком. Идентификация функциональных мутаций Мутации по CanIon-гену, которые ответственны за выявляемый фенотип или признак, могут быть идентифицированы путем сравнения последовательностей СаnIоn-гена индивидов с позитивным признаком и контрольных индивидов. После того, как позитивная ассоциация подтверждена с помощью двуал-лельного маркера настоящего изобретения, идентифицированный локус можно просканировать в отношении наличия мутаций. В предпочтительном варианте осуществления настоящего изобретения, функциональные области, такие как экзоны и сайты сплайсинга, промоторы и другие регуляторные области CanIon-гена, сканируют на наличие мутаций. В предпочтительном варианте осуществления настоящего изобретения последовательность CanIon-гена сравнивают у индивидов с позитивным признаком и у контрольных индивидов. Предпочтительно, если показаны индивиды с позитивным признаком, несущие гаплотип, который ассоциирован с данным признаком, и индивиды с негативным признаком, не несущие гаплотип или аллель, ассоциированный с данным признаком. Обнаруживаемый признак или фенотип может включать различные проявления изменений CanIon-функции. Метод выявления мутации в сущности аналогичен методу, используемому для идентификации дву-аллельного маркера. Способ, используемый для обнаружения таких мутаций, включает, как правило, следующие стадии: амплификации области CanIon-гена, включающей двуаллельный маркер или группу двуаллельных маркеров, ассоциированных с данным признаком в образцах ДНК пациентов с позитивным признаком и образцах ДНК людей в контроле; секвенирования амплифицированной области; сравнения последовательностей ДНК индивидов с позитивным признаком и индивидов в контроле; определения мутаций, специфичных для пациентов с позитивным признаком. В другом варианте осуществления настоящего изобретения указанный двуаллельный маркер выбран из группы, состоящей из А1-А18 и их комплементов. Затем, путем скринирования целесообразно проверить более крупные популяции больных и здоровых с помощью любого метода генотипирования, который описан в данном описании, предпочтительно с использованием метода микросеквенирования в индивидуальном тест-варианте. Полиморфизм рассматривают в виде кандидатных мутаций при наличии - 55 - 008252 частот у больных и в контроле, сопоставимых с ожидаемыми результатами по ассоциации. Полиморфизм рассматривают в виде кандидатных "признак-определяющих" мутаций, если они обнаруживают статистически значимую корреляцию с выявляемым фенотипом. Двуаллельные маркеры настоящего изобретения в способах генетического диагностирования CanIon-последовательность нуклеиновой кислоты и двуаллельные маркеры настоящего изобретения можно также использовать для создания диагностических тестов (диагностикумов), способных идентифицировать индивиды, генотипы которых несут риск развития выявляемого признака в последующее время. Такой диагноз может быть пригоден для определения, мониторинга, прогноза и/или профилактики или лечения многочисленных заболеваний или состояний, включая шизофрению, биполярное расстройство и другие нарушения ЦНС, такие как эпилепсия и нарушения болевых ощущений, сердечнососудистые состояния, такие как порок сердца, гипертензия, аритмия, и многочисленные другие заболевания и состояния. Методы диагностики настоящего изобретения могут использовать различные методологии для определения присутствия у обследуемого человека паттерна двуаллельного маркера, ассоциированного с повышенным риском развития выявляемого признака, или у индивида, страдающего от выявляемого признака, как результата конкретной мутации, включая способы, которые делают возможным анализ индивидуальных хромосом для гаплотипирования, такие как исследование семей, ДНК-анализ отдельного спермия или соматических гибридов. В настоящем изобретении созданы диагностические способы определения индивида с риском развития заболевания или страдающего от заболевания, возникшего в результате мутации или полиморфизма в CanIon-гене. В настоящем изобретении созданы также способы определения индивида, обладающего восприимчивостью к шизофрении и биполярному расстройству, или к любому другому состоянию, связанному с кальциевыми каналами, известному в данной области техники и описанному в данном описании. Данные способы включают получение образца нуклеиновой кислоты от данного индивида и определение в образце нуклеиновой кислоты наличия по меньшей мере одного аллеля или по меньшей мере одного гаплотипа двуаллельного маркера, свидетельствующего о риске развития данного признака, или свидетельствующего о том, что у данного индивида проявляется данный признак из-за обладания конкретным CanIon-полиморфизмом или конкретной мутацией (аллельопределяющий признак). Предпочтительно, для таких диагностических способов образец нуклеиновой кислоты получают от данного индивида и данный образец генотипируют с использованием способов, описанных выше в разделе "Способы генотипирования образцов ДНК по двуаллельным маркерам". Данная диагностика может основываться на одиночном двуаллельном маркере или на группе двуаллельных маркеров. В каждом из этих способов образец нуклеиновой кислоты получают от обследуемого субъекта, после чего определяют паттерн двуаллельного маркера одного или нескольких двуаллельных маркеров А1-А18. В одном из вариантов осуществления настоящего изобретения на образце нуклеиновой кислоты осуществляют ПЦР-амплификацию, чтобы амплифицировать области, в которых идентифицируют полиморфизм, ассоциированный с выявляемым фенотипом. Полученные продукты амплификации секве-нируют для определения индивидуального обладания одним или несколькими CanIon-полиморфизмами, ассоциированными с выявляемым фенотипом. Праймеры, используемые для получения продуктов амплификации, могут включать праймеры, приведенные в табл. 1. Альтернативно, данный образец нуклеиновой кислоты подвергают реакциям микросеквенирования, как описано выше, для определения индивида, обладающего одним или несколькими CanIon-полиморфизмами, ассоциированными с выявляемым фенотипом, возникшим в результате мутации или полиморфизма в CanIon-гене. Праймеры, используемые в реакциях микросеквенирования, могут включать праймеры, приведенные в табл. 4. В другом варианте осуществления настоящего изобретения данный образец нуклеиновой кислоты контактирует с одним или несколькими аллельспецифичными олигонуклеотидными зондами, которые специфически гиб-ридизуются с одним или несколькими CanIon-аллелями, ассоциированными с выявляемым фенотипом. Зонды, используемые для гибридизационного анализа, включают зонды, приведенные в табл. 3. В другом варианте осуществления настоящего изобретения, данный образец нуклеиновой кислоты контактирует со вторым CanIon-олигонуклеотидом, способным произвести продукт амплификации при использовании с аллельспецифичным олигонуклеотидом в реакции амплификации. Присутствие продукта амплификации в данной реакции амплификации свидетельствует о том, что данный индивид обладает одним или несколькими CanIon-аллелями, ассоциированными с выявляемым фенотипом. В предпочтительном варианте осуществления настоящего изобретения определяется идентичность определенного нуклеотида, присутствующего по меньшей мере в одном двуаллельном маркере, выбранном из группы, состоящей из А1-А18 и их комплементов, и выявляемым признаком, является шизофрения и биполярное расстройство. Диагностический набор включает любой из полинуклеотидов настоящего изобретения. Такие диагностические способы чрезвычайно полезны и при некоторых обстоятельствах могут использоваться для инициации превентивного лечения или для учета индивида, несущего значимый гапло-тип, который предвосхищает диагностические знаки, такие как минорные симптомы. Диагностика, которая анализирует и предсказывает реакцию на лекарственное средство или побоч - 56 - 008252 ные эффекты на лекарственное средство, может использоваться для определения индивида, нуждающегося в лечении с помощью конкретного лекарственного средства. Например, если диагноз указывает на вероятность того, что индивид будет положительно реагировать на лечение конкретным лекарственным средством, то данное лекарственное средство можно назначать данному индивиду. Напротив, если диагноз свидетельствует, что индивид, вероятно, реагирует отрицательно на лечение конкретным лекарственным средством, можно предписать альтернативный курс лечения. Отрицательную реакцию можно определить как отсутствие эффективной реакции или наличие токсических побочных эффектов. Клинические испытания лекарственного средства представляют собой другое применение для маркеров настоящего изобретения. Один или несколько маркеров, показывающих реакцию на агент, действующий против шизофрении или биполярного расстройства или иного состояния, связанного с кальциевыми каналами, или показывающих реакцию в виде побочных эффектов на агент, действующий против шизофрении или биполярного расстройства, или иного состояния, связанного с кальциевыми каналами, можно идентифицировать с использованием описанных выше способов. Соответственно, потенциальных участников клинических испытаний такого агента можно скринировать для идентификации индивидов, которые вероятнее всего положительно отреагируют на данное лекарственное средство, и для исключения тех, которые, вероятно, испытают побочные эффекты. Таким образом, эффективность лекарственного лечения можно измерить у индивидов, которые положительно реагируют на данное лекарственное средство, без снижения измеряемости в результате включения индивидов, которые едва ли отреагируют положительно в данном исследовании, и без риска возникновения нежелательных проблем, связанных с безопасностью. В конкретных вариантах осуществления настоящего изобретения данный признак, который представляет собой шизофрению или биполярное расстройство, анализируют с использованием представленных диагностик. Вместе с тем, настоящее изобретение также включает в себя любой из описанных в данном описании превентивных, диагностических, прогностических и терапевтических способов с использованием двуаллельных маркеров настоящего изобретения в способах, предупреждающих, диагностирующих, управляющих или лечащих родственные заболевания, особенно связанные с расстройствами ЦНС. В примере осуществления родственные заболевания могут включать психотические нарушения, расстройства эмоционального состояния, аутизм, зависимость от химических веществ и алкоголизм, нарушение болевых ощущений, эпилепсию, задержку психического развития и другие психические заболевания, включая когнитивную функцию, страх, нарушение функции принятия пищи, нарушение функции мотивации и расстройство личности, которые определяются в соответствии с четвертым изданием классификации Diagnosis and Statistical Manual of Mental Disorders (DSM-IV). Другие нарушения включают сердечно-сосудистые расстройства, такие как стенокардия, гипертензия или аритмии. Рекомбинантные векторы Используемый в данном описании термин "вектор" подразумевает кольцевую или линейную молекулу ДНК или РНК, которая является двухцепочечной либо одноцепочечной, и которая включает, по меньшей мере, один полинуклеотид, представляющий интерес, который требуется перенести в клетку-хозяина или в одноклеточный, или в многоклеточный организм-хозяин. Настоящее изобретение включает в себя семейство рекомбинантных векторов, которые включают регуляторный полинуклеотид, полученный из CanIon-геномной последовательности, и/или кодирующий полинуклеотид либо из CanIon-геномной последовательности, либо из кДНК-последовательности. Как правило, рекомбинантный вектор настоящего изобретения может включать любой из описанных в данном описании полинуклеотидов, включая регуляторные последовательности, кодирующие последовательности и полинуклеотидные конструкции, а также любой CanIon-праймер или зонд, как указано выше. В частности, рекомбинантные векторы настоящего изобретения могут включать любой из полинуклеотидов, описанных в разделе "Геномные последовательности CanIon-гена", в разделе "CanIon-кДНК-последовательности", в разделе "Кодирующие области", в разделе "Полинуклеотидные конструкции" и в разделе "Олигонуклеотидные зонды и праймеры". В первом предпочтительном варианте осуществления настоящего изобретения рекомбинантный вектор настоящего изобретения используют для амплификации встраиваемого полинуклеотида, полученного из геномной последовательности SEQ ID No 1-3 или 6 или CanIon-кДНК, например, кДНК с SEQ ID No 4, в соответствующей клетке-хозяине, данный полинуклеотид амплифицируется каждый раз, когда происходит репликация данного рекомбинантного вектора. Другой предпочтительный вариант осуществления рекомбинантных векторов, в соответствии с настоящим изобретением, включает экспрессионные векторы, включающие либо регуляторный полинук-леотид, либо кодирующую нуклеиновую кислоту настоящего изобретения, либо и тот, и другую. В некоторых вариантах осуществления настоящего изобретения экспрессионные векторы используют для экспрессии CanIon-полипептида, который затем можно выделить очисткой и, например, использовать в анализе по скринированию лиганда или в качестве иммуногена с тем, чтобы вызвать продукцию специфичных антител, направленных против CanIon-белка. В других вариантах осуществления настоящего изобретения данные экспрессионные векторы используют для создания трансгенных животных, а также для генотерапии. Экспрессия требует, чтобы в векторах создавались соответствующие сигналы, причем ука - 57 - 008252 занные сигналы включают различные регуляторные элементы, такие как энхансеры/промоторы из источников вируса и млекопитающих, которые приводят в действие в клетках-хозяевах экспрессию интересующих генов. Доминантные лекарственные селективные маркеры для создания долговременных стабильных клеточных клонов, экспрессирующих данные продукты, как правило, включены в экспрессион-ные векторы настоящего изобретения, так как они являются элементами, которые связывают экспрессию данных лекарственных селективных маркеров с экспрессией данного полипептида. Более подробно, настоящее изобретение относится к экспрессионным векторам, которые включают нуклеиновые кислоты, кодирующие CanIon-белок, предпочтительно CanIon-белок аминокислотной последовательности SEQ ID No 5, или его варианты или фрагменты. Настоящее изобретение относится также к рекомбинантному экспрессионному вектору, пригодному для экспрессии CanIon-кодирующей последовательности, где указанный вектор включает нуклеиновую кислоту SEQ ID No 4. Рекомбинантные векторы, включающие нуклеиновую кислоту, содержащую двуаллельный маркер, связанный с CanIon, также являются частью настоящего изобретения. В предпочтительном варианте осуществления настоящего изобретения указанный двуаллельный маркер выбран из группы, состоящей из А1-А18 и их комплементов. Некоторые элементы, которые можно найти в векторах настоящего изобретения, описаны более подробно в нижеследующих разделах. Настоящее изобретение включает также первичные, вторичные, иммортализованные гомологично рекомбинантные клетки-хозяева позвоночных, предпочтительно млекопитающих и, в частности, человека, которые были созданы биотехнологически для: а) встраивания экзогенных (гетерологичных) поли-нуклеотидов в эндогенную хромосомную ДНК гена-мишени, b) удаления эндогенной хромосомной ДНК, и/или с) замены эндогенной хромосомной ДНК экзогенными полинуклеотидами. Вставки, делеции и/или замены полинуклеотидных последовательностей могут представлять собой кодирующие последовательности гена-мишени и/или регуляторные области, такие как промоторные и энхансерные последовательности, функционально присоединенные к гену-мишени. Кроме того, настоящее изобретение относится к способу создания гомологичной рекомбинантной клетки-хозяина in vitro или in vivo, в котором экспрессия гена-мишени, аномально экспрессируемого в данной клетке, изменяется. Предпочтительно изменение вызывает экспрессию данного гена-мишени в нормальных условиях роста или в условиях, пригодных для продуцирования полипептида, кодируемого данным геном-мишенью. Данный способ включает следующие стадии: (а) трансфицирования данной клетки in vitro или in vivo при помощи полинуклеотидной конструкции, причем данная полинуклеотид-ная конструкция включает: (i) нацеливающую последовательность; (ii) регуляторную последовательность и/или кодирующую последовательность; и (iii) неспаренный донорный сайт сплайсинга, если требуется, в результате чего продуцируется трансфицированная клетка; и (b) сохранения данной трансфи-цированной клетки in vitro или in vivo в условиях, подходящих для гомологичной рекомбинации. Настоящее изобретение относится, кроме того, к способу изменения экспрессии гена-мишени в клетке in vitro или in vivo, где данный ген экспрессируется в данной клетке аномально, включающему следующие стадии: (а) трансфицирования данной клетки in vitro или in vivo полинуклеотидной конструкцией, причем данная полинуклеотидная конструкция включает: (i) нацеливающую последовательность; (ii) регуляторную последовательность и/или кодирующую последовательность; (iii) неспаренный донорный сайт сплайсинга, если требуется, в результате чего продуцируется трансфицированная клетка; и (b) сохранения данной трансфицированной клетки in vitro или in vivo в условиях, подходящих для гомологичной рекомбинации, посредством которой продуцируется гомологично рекомбинантная клетка; и (с) сохранения гомологично рекомбинантной клетки in vitro или in vivo в условиях, подходящих для экспрессии данного гена. Кроме того, настоящее изобретение имеет отношение к способу создания полипептида настоящего изобретения путем изменения экспрессии целевого эндогенного гена в клетке in vitro или in vivo, где данный ген экспрессируется в данной клетке аномально, включающему следующие стадии: а) трансфи-цирования данной клетки in vitro полинуклеотидной конструкцией, включающей: (i) нацеливающую последовательность; (ii) регуляторную последовательность и/или кодирующую последовательность; и (iii) неспаренный донорный сайт сплайсинга, если требуется, продуцируя тем самым трансфицированную клетку; (b) сохранения данной трансфицированной клетки in vitro или in vivo в условиях, подходящих для гомологичной рекомбинации, продуцируя таким образом гомологично рекомбинантную клетку; и (с) сохранения гомологично рекомбинантной клетки in vitro или in vivo в условиях, подходящих для экспрессии данного гена, создавая таким образом данный полипептид. Настоящее изобретение имеет также отношение к полинуклеотидной конструкции, которая изменяет экспрессию целевого гена в типичной клетке, в которой данный ген экспрессируется аномально. Это происходит в том случае, если данная полинуклеотидная конструкция встраивается в хромосомную ДНК клетки-мишени, где данная полинуклеотидная конструкция включает: а) нацеливающую последовательность; b) регуляторную последовательность и/или кодирующую последовательность; и с) неспаренный донорный сайт сплайсинга, если требуется. Дополнительно включаются полинуклеотидные конструк - 58 - 008252 ции, как описано выше, где данная конструкция дополнительно включает полинуклеотид, который кодирует полипептид, и находится в рамке считывания с эндогенным геном-мишенью после гомологичной рекомбинации с хромосомной ДНК. Композиции можно получить и способы осуществить с помощью методов, известных в данной области техники, которые описаны в патентах США №№: 6054288; 6048729; 6048724; 6048524; 5994127; 5968502; 5965125; 5869239; 5817789; 5783385; 5733761; 5641670;5580734;в международных публикациях №№: WO96/29411, WO94/12650; и научных статьях, включающих Roller и соавт., 86:8932-8935 (1989). 1. Основные характерные особенности экспрессионных векторов настоящего изобретения. В соответствии с настоящим изобретением рекомбинантный вектор включает, но не ограничивается ими, YAC (искусственная хромосома дрожжей), ВАС (искусственная хромосома бактерий), фаг, фагеми-ду, космиду, плазмиду или даже линейную молекулу ДНК, которая может включать хромосомную, нехромосомную, полусинтетическую и синтетическую ДНК. Такой рекомбинантный вектор может включать транскрипционный элемент, включающий совокупность из: (1) генетического элемента или элементов, играющих регуляторную роль в экспрессии гена, например, промоторы или энхансеры. Энхансеры являются цис-действующими элементами ДНК, обычно длиной от около 10 до 300 п.н., которые действуют на промотор для увеличения транскрипции. (2) структурной или кодирующей последовательности, которая транскрибируется в мРНК и, в конечном счете, транслируется в полипептид, причем указанная структурная кодирующая последовательность функционально присоединяется к регуляторным элементам, описанным в (1); и (3) соответствующих последовательностей инициации транскрипции и терминации. Структурные единицы, предназначенные для использования в дрожжевой или эукариотической экспрессионных системах, предпочтительно включают лидерную последовательность, делающую возможной внеклеточную секрецию транслируемого белка клеткой-хозяином. Альтернативно, если рекомбинантный белок экс-прессируется без лидерной или транспортной последовательности, он может включать N-концевой остаток. Данный остаток может или не может быть впоследствии отщеплен от экспрессированного рекомби-нантного белка для создания конечного продукта. В большинстве случаев рекомбинантные экспрессионные векторы будут включать ориджины репликации, селективные маркеры, позволяющие трансформировать клетку-хозяина, и промотор, полученный из высокоэкспрессируемого гена, который контролирует транскрипцию расположенного "правее" структурной последовательности. Гетерологичную структурную последовательность собирают в соответствующей фазе последовательностями инициации трансляции и терминации, и предпочтительно с лидерной последовательностью, способной направлять секрецию транслируемого белка в периплазмати-ческое пространство или во внеклеточную среду. В специфичном варианте осуществления настоящего изобретения, в котором данный вектор адаптирован для трансфицирования и экспрессирования требуемых последовательностей в клетках-хозяевах млекопитающих, предпочтительные векторы будут включать ориджин репликации требуемого хозяина, подходящий промотор и энхансер, а также любые необходимые сайты связывания рибосом, сигнал полиаденилирования, донорный и акцепторный сайты сплайсинга, последовательности терминации транскрипции и нетранскрибируемые 5'-фланкирующие последовательности. Последовательности ДНК, полученные из генома вируса SV40, например, SV40-ориджина, раннего промотора, энхансера и сигналов сплайсинга и полиаденилирования, можно использовать для создания требуемых нетранскрибируемых генетических элементов. In vivo-экспрессия CanIon-полипептида SEQ ID No 5, или его фрагментов или его вариантов, может оказаться пригодной для того, чтобы исправить генетический дефект, связанный с экспрессией нативно-го гена в организме-хозяине, или для получения биологически неактивного CanIon-белка. Следовательно, настоящее изобретение включает также рекомбинантные экспрессионные векторы, созданные, главным образом, для in vivo-получения CanIon-полипептида SEQ ID No 5, или его фрагментов или его вариантов, путем введения соответствующего генетического материала в организм лечащегося пациента. Данный генетический материал можно вводят in vitro в клетку, которую раньше извлекли из данного организма, затем модифицированную клетку вновь ввели в указанный организм, непосредственно in vivo в соответствующую ткань. 2. Регуляторные элементы. Промоторы Подходящие промоторные области, используемые в соответствии с настоящим изобретением, в экспрессионных векторах, выбраны с учетом клетки-хозяина, в которой экспрессируется данный гетеро-логичный ген. Конкретный промотор, используемый для контроля экспрессии представляющей интерес последовательности нуклеиновой кислоты, не считается существенным, до тех пор, пока он способен управлять экспрессией нуклеиновой кислоты в данной клетке-мишени. Поэтому, если клетка человека является мишенью, предпочтительно поместить нуклеотидную кодирующую область по соседству и под контролем промотора, такого как, например, промотор человека или вирусный промотор, который способен экспрессироваться в клетке человека. Подходящий промотор может быть гетерологичным по отношению к нуклеиновой кислоте, экспрессию которой он контролирует, или, альтернативно, он может быть эндогенным к данному нативному - 59 - 008252 полинуклеотиду, содержащему кодирующую экспрессируемую последовательность. Кроме того, данный промотор, как правило, гетерологичен по отношению к рекомбинантным векторным последовательностям, в которые встраивается данная конструкция промотор/кодирующая последовательность. Промоторные области можно выбрать из любого желаемого гена с использованием, например, CAT (хлорамфениколтрансферазных) векторов и более предпочтительно с использованием векторов рКК232-8 и рСМ7. Предпочтительными бактериальными промоторами являются промоторы LacI, LacZ, РНК-полимеразные промоторы бактериофага Т3 или Т7, промоторы gpt, лямбда PR, PL и trp (ЕР 0036776), промотор гена полиэдрина или промотор белка р10 бакуловируса (Kit Novagen) (Smith и соавт., 1983; O'Reilly и соавт., 1992), промотор лямбда PR или также trc-промотор. Эукариотические промоторы включают предранний CMV, тимидинкиназный HSV, ранний и поздний SV40, LTR из ретровируса и металлотионеин-L мыши. Выбор удобного вектора и промотора определяется на уровне специалиста в данной области техники. Выбор промотора определяется квалификацией специалиста в области генной инженерии. Для этого, например, можно обратиться к Sambrook и соавт. (1989) или к методам, описанным также у Fuller и соавт. (1996). Другие регуляторные элементы Если используют кДНК-вставку, то, как правило, желательно включить сигнал полиаденилирова-ния, который осуществляет надлежащее полиаденилирование генного транскрипта. Природа сигнала полиаденилирования не считается ключевой для успешного осуществления настоящего изобретения, и можно использовать такую последовательность, как последовательность гормона роста человека и сигнал полиаденилирования SV40. В качестве элемента экспрессионной кассеты рассматривают также терминатор. Эти элементы могут служить для повышения уровней мРНК и для минимизации сквозного прочтения генетической информации из данной кассеты в другие последовательности. 3. Селективные маркеры. Такие маркеры должны обеспечивать клетку идентифицируемым изменением, позволяющим легкую идентификацию клеток, содержащих данную экспрессионную конструкцию. Селективные маркерные гены для отбора трансформированных клеток-хозяев, обладают предпочтительно дигидрофолатре-дуктазной или неомициновой устойчивостью в культуре эукариотических клеток, TRPI для С. cerevisiae, или тетрациклиновой, рифампициновой или амплицилиновой устойчивостью для Е. coli, или левансаха-разной устойчивостью для микробактерий, данный последний маркер является негативным селективным маркером. 4. Предпочтительные векторы. Бактериальные векторы В качестве представительного, но не ограничивающего примера, пригодные экспрессионные векторы для бактериального применения могут включать селективный маркер и бактериальный ориджин репликации, полученный из коммерчески доступных плазмид, включающих генетические элементы pBR322 (ATCC 37017). Такие коммерческие векторы включают, например, рКК223-3 (Pharmacia, Uppsala, Швеция) и GEMI (Promega Biotec, Madison, WI, США). Специалистам в данной области техники известно большое количество других подходящих и коммерчески доступных векторов, таких как следующие бактериальные векторы: pQE70, pQE60, pQE-9 (Qiagen), pbs, pD10, phagescript, psiX174, pbluescript SK, pbsks, pNH8A, pNH16A, pNH18A, pNH46A (Stratagene); ptrc99a, pKK223-3, pKK233-3, pDR540, pRIT5 (Pharmacia); pWLNEO, pSV2CAT, pOG44, pXT1, pSG (Stratagene); pSVK3, pBPV, pMSG, pSVL (Pharmacia); pQE-30 (QIAexpress). Векторы бактериофага Вектор бактериофага Р1 может содержать большие вставки в диапазоне от около 80 до около 100 т.п.н. Конструкция векторов бактериофага Р1, таких как р158 или р158/nео8, полнее всего описана Sternberg (1992, 1994). Рекомбинантные клоны Р1, включающие CanIon-нуклеотидные последовательности, можно создать путем встраивания больших полинуклеотидов из более чем 40 т.п.н. (Linton и соавт., 1993). Для получения ДНК Р1 для трансгенных экспериментов предпочтительным протоколом является протокол, описанный McCormick и соавт. (1994). Вкратце, Е. coli (предпочтительно штамм NS3529), несущих плазмиду Р1, выращивают в течение ночи в соответствующей бульонной среде, содержащей 25 мкг/мл канамицина. ДНК Р1 получают из Е. coli путем щелочного лизиса с использованием набора Qiagen Plas-mid Maxi (Qiagen, Chatsworth, CA, США), в соответствии с инструкциями производителя. ДНК Р1 выделяют очисткой из полученного бактериального лизата на двух колонках Qiagen-tip 500 с использованием промывочного и элюирующего буферов, содержащихся в данном наборе. Затем, перед осаждением ДНК 70%-м этанолом, проводят экстрагирование смесью фенол/хлороформ. После солюбилизации осажденной ДНК в ТЕ (10 мМ Трис-HCl, рН 7,4, 1 мМ ЭДТА), концентрацию солюбилизированной ДНК определяют на спектрофотометре. Если целью является экспрессия клона Р1, включающего CanIon-нуклеотидные последовательно -сти, в трансгенном животном, обычно в трансгенных мышах, желательно удалить векторные последовательности из ДНК-фрагмента Р1, например, путем расщепления ДНК Р1 по редкоразрезаемым сайтам - 60 - 008252 внутри Р1-полилинкера (SfiI, NotI или SalI). Затем Р1-вставку выделяют очисткой из векторных последовательностей, подвергнутых гель-электрофорезу в пульсирующем поле, с использованием методов, аналогичных методам, которые первоначально предлагались для выделения ДНК из YAC (Schedl и соавт., 1993а; Peterson и соавт., 1993). На этой стадии полученная, выделенная очисткой вставка ДНК может быть сконцентрирована, при необходимости, на фильтровальной установке Millipore Ultrafree-MC (Milli-pore, Bedford, MA, США - в пределах молекулярной массы 30000) и затем диализована против буфера для микроинъекций (10 мМ Трис-HCl, рН 7,4; 250 мкМ ЭДТА), содержащего 100 мМ NaCl, 30 мкМ спермина, 70 мкМ спермидина, в мембране для микродиализа (тип VS, 0,025 мкМ от Millipore). Целостность выделенной очисткой ДНК-вставки Р1 определяют с помощью электрофореза в пульсирующем поле в 1%-м агарозном (Sea Kem GTG; FMC Bio-products) геле и окрашивают этидийбромидом. Бакуловирусные векторы Подходящим вектором для экспрессии CanIon-полипептида SEQ ID No 5 или его фрагментов или вариантов является бакуловирусный вектор, который можно размножить в клетках и в клеточных линиях насекомых. Специфичной подходящей хозяйской векторной системой является бакуловирусный трансфер-вектор pVL1392/1393 (Pharmingen), который используют для трансфицирования клеточной линии SF9 (АТСС N°CRL 1711), которую получают из Spodoptera frugiperda. Другие подходящие векторы для экспрессии CanIon-полипептида SEQ ID No 5 или его фрагментов или вариантов в бакуловирусной экспрессионной системе включают векторы, описанные у Chai и соавт. (1993), Vlasak и соавт. (1983) и Lenhard и соавт. (1996). Вирусные векторы В одном из специфичных вариантов осуществления настоящего изобретения вектор получают из аденовируса. В соответствии с настоящим изобретением предпочтительные аденовирусные векторы представляют собой векторы, которые описаны у Feldman and Steg (1996) или у Ohno и соавт. (1994). Другой предпочтительный рекомбинантный аденовирус, в соответствии с осуществлением данного специфичного варианта, представленного в настоящем изобретении, представляет собой аденовирус человека тип 2 или 5 (Ad 2 или Ad 5) или аденовирус животного происхождения (см., например, французская патентная заявка № FR-93.05954). Под ретровирусными векторами и аденоассоциированными вирусными векторами подразумевают, главным образом, рекомбинантные системы доставки генов, выбранные для переноса экзогенных поли-нуклеотидов in vivo, в частности, млекопитающим, включая людей. Данные векторы обеспечивают эффективную доставку генов в клетки, а перенесенные нуклеиновые кислоты стабильно интегрируются в хромосомную ДНК данного хозяина. Особенно предпочтительные препараты ретровирусов или ретровирусные конструкции-переносчики настоящего изобретения для доставки генов in vitro или in vivo включают ретровирусы, выбранные из группы, состоящей из вируса, индуцирующего очаг клеток норки, вируса саркомы мышей, вируса рети-кулоэндотелиоза и вируса саркомы Рауса. Особенно предпочтительные вирусы лейкоза мышей включают вирусы 4070А и 1504А, Abelson (ATCC No VR-999), Friend (ATCC No VR-245), Gross (ATCC No VR-590), Rauscher (ATCC No VR-998) и вирус лейкоза мышей Молони (ATCC No VR-190; Заявка PCT № WO 94/24298). Особенно предпочтительные вирусы саркомы Рауса включают Bryan с высоким титром (ATCC Nos VR-334, VR-657, VR-726, VR-659 и VR-728). Другие предпочтительные ретровирусные векторы представляют собой ретровирусы, которые описаны у Roth и соавт. (1996), заявка РСТ № WO 93/25234, заявка PCT № WO 94/06920, Roux и соавт., 1989, Julan и соавт., 1992 и Neda и соавт., 1991. Еще одна вирусная векторная система, которая рассматривается в настоящем изобретении, включает аденоассоциированный вирус (AAV). Аденоассоциированный вирус представляет собой встречаемый в природе дефектный вирус, который нуждается в другом вирусе, таком как аденовирус или герпес-вирус, в качестве вируса-помощника для эффективной репликации и продуктивного жизненного цикла (Muzyczka и соавт., 1992). Это один из немногих вирусов, который может интегрировать свою ДНК в неделящиеся клетки и проявляют высокую частоту устойчивой интеграции (Flotte и соавт., 1992; Samul-ski и соавт., 1989; McLaughlin и соавт., 1989). Отличительной чертой AAV является его уменьшенная эффективность в отношении трансдуцирования первичных клеток по сравнению с трансформированными клетками. ВАС векторы Клонирующая система бактериальной искусственной хромосомы (ВАС) (Shizuya и соавт., 1992) была создана для устойчивого сохранения в Е. coli больших фрагментов геномной ДНК (100-300 т.п.н.). Предпочтительный ВАС-вектор включает вектор pBeloBAC11, который был описан Kim и соавт. (1996). С помощью данного вектора получают ВАС-библиотеки, а с использованием отобранной по размеру геномной ДНК, которую частично расщепляют ферментами, их можно лигировать в сайты Ват HI или HindIII данного вектора. Фланкирование этих клонирующих сайтов осуществляют Т7 и SP6 РНК-полимеразными сайтами инициации транскрипции, которые можно использовать для создания концевых зондов с помощью методов РНК-транскрипции или ПЦР. После создания ВАС-библиотеки в Е. coli, ДНК ВАС выделяют очисткой из клетки-хозяина в виде сверхспирализованной кольцевой молекулы. Преобразованию таких кольцевых молекул в линейную форму предшествует определение размера и введение - 61 - 008252 ВАС в реципиентные клетки. Клонирующие сайты фланкируют с помощью двух Not I-сайтов, что позволяет вырезать клонированные сегменты из данного вектора в результате расщепления Not I. Альтернативно, ДНК-вставку, содержащую вектор pBeloBAC11, можно линеализировать путем обработки ВАС-вектора коммерчески доступным ферментом лямбда-терминаза, что приводит к расщеплению уникального cosN-сайта, но данный способ расщепления дает полноразмерный ВАС-клон, содержащий последовательности и ДНК-вставки и ВАС. 5. Доставка рекомбинантных векторов. Для того чтобы эффективно экспрессировать полинуклеотиды и полинуклеотидные конструкции настоящего изобретения, данные конструкции необходимо доставить в клетку. Такую доставку можно осуществить in vitro, как в лабораторных методах трансформирования клеточных линий или in vivo, или ex vivo, так и при лечении некоторых болезненных состояний. Одним из механизмов является вирусная инфекция, при которой экспрессионная конструкция инкапсулируется в вирусные частицы, вызывающие заражение. В настоящем изобретении рассматривается также несколько невирусных способов переноса поли-нуклеотидов в культивируемые клетки млекопитающих, которые включают, без ограничения, осаждение кальцийфосфатом (Graham и соавт., 1973; Chen и соавт., 1987), DEAE-декстраном (Gopal, 1985), электро-порацией (Tur-Kaspa и соавт., 1986; Potter и соавт., 1984), прямой микроинъекцией (Harland и соавт., 1985), нагруженными ДНК липосомами (Nicolau и соавт., 1982; Fraley и соавт., 1979) и рецептор-опосредованной трансфекцией (Wu and Wu, 1987; 1988). Некоторые из таких методов можно успешно адаптировать для in vivo или ex vivo использования. После доставки в клетку экспрессионный полинуклеотид можно стабильно интегрировать в геном реципиентной клетки. Данная интеграция может произойти в соответствующем месте и ориентации с помощью гомологичной рекомбинации (замещение гена) или его можно интегрировать произвольно, не помещая в конкретное место (аугментация гена). Еще в одних вариантах осуществления настоящего изобретения нуклеиновую кислоту можно стабильно сохранять в клетке в виде отдельного эписомного сегмента ДНК. Такие сегменты нуклеиновых кислот или "эписомы" кодируют последовательности, достаточные для сохранения и репликации синхронно с циклом клетки-хозяина или независимо от указанного цикла. Один из специфичных вариантов способа доставки in vivo белка или пептида внутрь клетки позвоночного включает стадию введения препарата, включающего физиологически приемлемый носитель и " голый" полинуклеотид, оперативно кодирующий представляющий интерес полипептид во внутритканевом пространстве ткани, включающей данную клетку, посредством чего "голый" полинуклеотид помещается внутрь клетки, и обладающей физиологическим действием. Это особенно применимо для переноса in vitro, но можно также применить и для in vivo. Композиции для использования in vitro и in vivo, включающие "голый" полинуклеотид, описаны в заявке РСТ № WO 90/11092 (Vical Inc.), а также в заявке РСТ № WO 95/11307 (Institut Pasteur, INSERM, Universite d'Ottawa), а также в статьях Tacson и соавт. (1996) и Huygen и соавт. (1996). Еще в одном варианте настоящего изобретения перенос "голого" полинуклеотида настоящего изобретения, включающего полинуклеотидную конструкцию настоящего изобретения, в клетки, можно задействовать с помощью бомбардировки микрочастицами (биолистика), причем указанные частицы, покрытые ДНК, ускоряются до высокой скорости, что позволяет им пронзать клеточные мембраны и проникать в клетки без их повреждения так, как описано Klein и соавт. (1987). В дополнительном варианте осуществления настоящего изобретения полинуклеотид настоящего изобретения может быть включен в липосому (Ghosh and Bacchawat, 199;: Wong и соавт., 1980; Nicolau и соавт., 1987). В специфичном варианте осуществления настоящего изобретения создана композиция для получения in vivo описанного в данном описании CanIon-белка или полипептида. Она включает "голый" поли-нуклеотид, оперативно кодирующий данный полипептид, в растворе физиологически приемлемого носителя, и подходящего для введения в ткань, что определяет появление клеток данной ткани, которые экс-прессируют указанный белок или полипептид. Количество вектора, которое инъецируется в желаемый организм-хозяин, варьирует в соответствии с местом инъекции. В качестве примерной дозы, в тело животного, предпочтительно тело млекопитающего, например тело мыши, следует инъецировать 0,1-100 мкг вектора. В другом варианте осуществления настоящего изобретения вектор в соответствии с настоящим изобретением можно вводить в клетку-хозяина in vitro, предпочтительно в клетку-хозяина, предварительно взятую у животного, подвергающегося лечению, и более предпочтительно в соматическую клетку, такую как клетка мышц. На следующей стадии клетку, которую трансформировали вектором, кодирующим требуемый CanIon-полипептид или требуемый его фрагмент, реинтродуцируют в тело животного для того, чтобы доставить рекомбинантный белок в данное тело либо местно, либо системно. Клетки-хозяева Другая цель настоящего изобретения включает клетки-хозяева, трансформированные или трансфи-цированные описанными в данном описании полинуклеотидами, и, в частности, полинуклеотидом, - 62 - 008252 включающим CanIon-регуляторный полинуклеотид или кодирующую последовательность CanIon-полипептида, выбранную из группы, состоящей из SEQ ID Nos 1-4 или их фрагментов или вариантов. Включены также клетки-хозяева, которые трансформируют (прокариотические клетки) или которые трансфицируют (эукариотические клетки) рекомбинантным вектором, таким как один из тех, которые описаны выше. В частности, клетки-хозяева настоящего изобретения могут включать любой из полинук-леотидов, описанных в разделе "Геномные последовательности CanIon-гена", в разделе "CanIon-кДНК-последовательности", в разделе "Кодирующие области", в разделе "Полинуклеотидные конструкции" и в разделе "Олигонуклеотидные зонды и праймеры". Кроме того, рекомбинантная клетка-хозяин, в соответствии с настоящим изобретением, включает полинуклеотид, содержащий двуаллельный маркер, выбранный из группы, состоящей из А1-А18 и их комплементов. Дополнительная рекомбинантная клетка-хозяин, в соответствии с настоящим изобретением, включает любой из описанных в данном описании векторов, в частности, любой из векторов, описанных в разделе "Рекомбинантные векторы". Предпочтительными клетками-хозяевами, используемыми в качестве реципиентов для экспресси-онных векторов настоящего изобретения, являются следующие: a) прокариотические клетки-хозяева: штаммы Escherichia coli (штамм I.E.DH5-a), Bacillus subtilis, Salmonella typhimurium и штаммы видов, подобных Pseudomonas, Streptomyces и Staphilococcus; b) эукариотические клетки-хозяева: HeLa-клетки (АТСС N°CCL2; N°CCL2.1; N°CCL2.2), Cv 1-клетки (АТСС N°CCL70), COS-клетки (АТСС N°CRL1650; N°CRL1651), Sf-9-клетки (АТСС N°CRL1711), C127-клетки (АТСС N°CRL-1804), 3T3 (ATCCC N°CRL-6361), CHO (ATCC N°CCL-61), 293-клетки почек человека (АТСС N°45504; N°CRL-1573) и ВНК (ЕСАСС N° 84100501; N° 84111301); c) другие клетки-хозяева млекопитающих. Экспрессия CanIon-ген в клетках млекопитающих, прежде всего человека, может оказаться дефектной или же она может осуществляться в результате вставки геномной CanIon-последовательности или кДНК-последовательности CanIon с заменой копии CanIon-гена в геноме клетки животного на CanIon-полинуклеотид согласно настоящему изобретению. Такие генетические изменения можно осуществить путем гомологичных рекомбинационных событий с использованием специфических ДНК-конструкций, которые были описаны выше. Одним из видов клеток-хозяев, которые можно использовать, являются зиготы млекопитающих, такие как мышиные зиготы. Например, мышиные зиготы можно подвергнуть микроинъекции выделенной очисткой, представляющей интерес молекулой ДНК, например, выделенной очисткой молекулой ДНК, которая предварительно доведена до концентрации 1 нг/мл для ВАС-вставок до 3 нг/мкл для вставок бактериофага Р1 в 10 мМ Трис-HCl, рН 7,4, 250 мкМ ЭДТА, содержащего 100 мМ NaCl, 30 мкМ спермина и 70 мкМ спермидина. При микроинъекции ДНК большого размера можно использовать полиамины и высокие концентрации соли для того, чтобы исключить механическое повреждение данной ДНК, как описано Schedl и соавт. (1993b). Любой из полинуклеотидов настоящего изобретения, включая описанные в данном описании ДНК-конструкции, можно ввести в эмбриональную стволовую (ES) клеточную линию, предпочтительно мышиную ES-клеточную линию. ES-клеточные линии получают из плюрипотентных, некоммитированных клеток внутренней клеточной массы преимплантированных бластоцист. Предпочтительными ES-кле-точными линиями являются следующие: ES-E14TG2a (ATCC n°CRL-1821), ES-D3 (ATCC n°CRL-1934 и n°CRL-11632), YS001 (ATCC n°CRL-11776), 36,5 (ATCC n°CRL-11116). Для сохранения ES-клеток в не-коммитированном состоянии, их культивируют в присутствии фидерных клеток с подавленным ростом, которые создают соответствующие сигналы, предохраняющие этот эмбриональный фенотип и служат в качестве матрикса для адгезии ES-клетки. Предпочтительными фидерными клетками являются первичные эмбриональные фибробласты, которые создаются из ткани 13-14-дневных эмбрионов практически любого мышиного штамма, которые поддерживаются в культуре так, как это описано у Abbondanzo и соавт. (1993), и рост которых подавляется в результате облучения так, как описано у Robertson (1987), или в присутствии ингибирующей концентрации LIF так, как описано у Pease and Williams (1990). Конструкции для клеток-хозяев можно использовать традиционным образом для получения генного продукта, кодируемого рекомбинантной последовательностью. После трансформации соответствующего хозяина и роста данного хозяина до надлежащей клеточной плотности, выбранный промотор индуцируют подходящими средствами, такими как температурный сдвиг или химическая индукция, и клетки культивируют в течение дополнительного периода. Клетки собирают, как правило, центрифугированием, разрушением с помощью физических или химических средств, и полученный неочищенный экстракт оставляют для последующей очистки. Микробные клетки, используемые для экспрессии белков, можно разрушить любым удобным способом, включая циклическое замораживание-оттаивание, ультразвуковую обработку, механическое разрушение или используя агенты, лизирующие клетки. Такие способы хорошо известны специалистам в данной области техники. - 63 - 008252 Трансгенные животные Термины "трансгенные животные" или "животные-хозяева" используют в данном описании для обозначения животных, которые, обладая своим геном, подвергаются генетическому и искусственному воздействию так, что включают одну из нуклеиновых кислот настоящего изобретения. Предпочтительными животными являются млекопитающие, не относящиеся к человеческому роду, и включают млекопитающих, принадлежащих роду, выбранному из Mus (например, мыши), Rattus (например, крысы) и Oryctogalus (например, кролики), геном которых искусственно и генетически изменяют встраиванием нуклеиновой кислоты настоящего изобретения. В одном из вариантов осуществления настоящее изобретение включает в себя млекопитающих-хозяев, не принадлежащих человеческому роду, и животных, включающих рекомбинантный вектор настоящего изобретения или CanIon-ген, разрушенный гомологичной рекомбинацией с помощью нокаутного вектора. Трансгенные животные настоящего изобретения, все, включают во множестве своих клеток клонированную рекомбинантную или синтетическую ДНК-последовательность, одну из выделенных очисткой или выделенных нуклеиновых кислот, включающих CanIon-кодирующую последовательность, CanIon-регуляторный полинуклеотид, полинуклеотидную конструкцию или ДНК-последовательность, кодирующую антисмысловой полинуклеотид, такой как описанный в настоящем подробном описании. Как правило, трансгенное животное, в соответствии с настоящим изобретением, включает любой один из полинуклеотидов, рекомбинантных векторов и клеток-хозяев, описанных в настоящем изобретении. В частности, трансгенные животные настоящего изобретения могут включать любой из полинук-леотидов, описанных в разделе "Геномные последовательности CanIon-гена", в разделе "CanIon-кДНК-последовательности", в разделе "Кодирующие области", в разделе "Полинуклеотидные конструкции", в разделе "Олигонуклеотидные зонды и праймеры", в разделе "Рекомбинантные векторы" и в разделе "Клетки-хозяева". Кроме того, трансгенные животные, в соответствии с настоящим изобретением, содержат в своих клеточных телах и/или в своих линиях зародышевых клеток полинуклеотид, включающий двуаллельный маркер, выбранный из группы, состоящей из А1-А18 и их комплементов. В первом предпочтительном варианте осуществления настоящего изобретения трансгенные животные могут представлять собой хорошие экспериментальные модели с целью изучения разнообразных патологий, связанных с клеточной дифференциацией, в частности, трансгенные животные, в геном которых встраивают одну или несколько копий полинуклеотида, кодирующего нативный CanIon-белок, или, альтернативно, мутантный CanIon-белок. В другом предпочтительном варианте осуществления настоящего изобретения трансгенные животные могут экспрессировать представляющий интерес требуемый полипептид под контролем регулятор-ных полинуклеотидов CanIon-гена, приводя к хорошему выходу в синтезе данного, представляющего интерес белка и, в конечном счете, к тканеспецифичной тканевой экспрессии данного, представляющего интерес белка. Создание трансгенных животных настоящего изобретения можно осуществить в соответствии с общепринятыми методиками, хорошо известными специалистам в данной области техники. Дополнительные подробности, относящиеся к получению трансгенных животных и специфически трансгенных мышей, можно найти, например, в патентах США №№ 4873191, 5464764 и 5789215, каждый из которых включен здесь путем ссылки. Трансгенные животные настоящего изобретения получают путем применения методов, которые дают животное с геномом, в который введен экзогенный генетический материал. Данный метод включает получение генетического материала или его части, который кодирует CanIon-кодирующую последовательность, CanIon-регуляторный полинуклеотид либо ДНК-последовательность, кодирующую CanIon-антисмысловой полинуклеотид, такой как описано в настоящем описании. Рекомбинантный полинуклеотид настоящего изобретения встраивают в эмбриональную или в ES-стволовую клеточную линию. Данную вставку предпочтительно осуществляют с использованием элек-тропорации так, как описано у Thomas и соавт. (1987). Указанные клетки подвергают электропорации и скринируют (например, путем отбора с помощью селективных маркеров, ПЦР или с помощью Саузерн-блот-анализа), чтобы обнаружить позитивные клетки, которые интегрировали экзогенный рекомбинант-ный полинуклеотид в свой геном, предпочтительно с помощью гомологичного рекомбинантного события. В соответствии с настоящим изобретением можно использовать иллюстративный позитивно-негативный метод отбора, описанный Mansour и соавт. (1988). Затем позитивные клетки выделяют, клонируют и инъецируют в 3,5-дневные бластоциты мышей так, как описано у Bradley (1987). Данные бластоцисты вводят затем самке животного-хозяина и позволяют расти до условного момента. Альтернативно, позитивные ES-клетки приводят в контакт с 2,5-дневными эмбрионами на 8-16-клеточной стадии (морула) так, как описано у Wood и соавт. (1993) или у Nagy и соавт. (1993), ES-клетки, которые интернализуются с обширной колонизацией бластоцисты, включают клетки, которые дадут начало зародышевой линии. Потомство хозяйской самки тестируют, чтобы определить, какие животные являются трансгенны - 64 - 008252 ми, например, включают экзогенную ДНК-последовательность, а какие представляют собой дикий тип. Таким образом, настоящее изобретение включает также в себя трансгенное животное, содержащее нуклеиновую кислоту, рекомбинантный экспрессионный вектор или рекомбинантную клетку-хозяина, в соответствии с настоящим изобретением. Рекомбинантные клеточные линии, полученные от трансгенных животных настоящего изобретения Дополнительная цель настоящего изобретения включает рекомбинантные клетки-хозяева, полученные от описанных в данном описании трансгенных животных. Один из вариантов осуществления настоящего изобретения включает в себя клетки, полученные от млекопитающих-хозяев, не принадлежащих человеческому роду, и от животных, содержащих рекомбинантный вектор настоящего изобретения или CanIon-ген, разрушенный гомологичной рекомбинацией с помощью нокаутного вектора. Рекомбинантные клеточные линии можно основать in vitro из клеток, полученных из любой ткани трансгенного животного в соответствии с настоящим изобретением, например, путем трансфекции культуры первичных клеток с помощью векторов, экспрессирующих оnс-гены, таких как большой Т-антиген на основе SV40, как описано у Chou (1989) и Shaw и соавт. (1991). Способы скринирования CanIon-модуляторов и CanIon взаимодействующих соединений Для многих вариантов осуществления настоящего изобретения созданы соединения, которые взаимодействуют, связывают, активируют или подавляют экспрессию или активность CanIon-полипептидов, каналов и полинуклеотидов. Такие соединения могут представлять собой какое-либо органическое или неорганическое соединение, включая, но, не ограничиваясь ими, полипептиды, полинуклеотиды, липи-ды, углеводы, нуклеотиды, аминокислоты или малые молекулы ингибиторов или активаторов. Как указано в другом месте данной заявки, такие соединения пригодны для лечения или предупреждения любого из большого числа заболеваний или состояний. Для лечения или предупреждения психических нарушений, таких как шизофрения или биполярное расстройство, предпочтительно использовать ингибиторы CanIon-активности или экспрессии. Способы скринирования модуляторов CanIon-канала Соединения, способные связываться с CanIon, и соединения, способные модулировать CanIon-функцию, имеют важное применение в лечении заболевания. Хорошо установлено, что потенциал-зависимые воротные ионные каналы, как правило, являются лекарственными мишенями, потому что они фармакологически доступны, кодируются разными генами и, как правило, функционируют в виде сообществ мультимерных белков, обеспечивающих высокую степень функциональной и анатомической специфичности. Более того, поскольку ионный канал, открываясь и закрываясь, двигает заряженные потен-циалчувствительные аминокислоты, что приводит к изменению конформационного состояния, ионные каналы позволяют создать состояние, зависящее от молекул, которые, например, свяжут лишь каналы, находящиеся в состоянии токопроведения (активированные) или нетокопроведения (инактивированные). Кроме того, было показано, что множество модуляторов кальциевых каналов эффективны для лечения или предупреждения многих заболеваний и состояний. Например, было показано, что ингибиторы кальциевых каналов эффективны против сердечно-сосудистых заболеваний и состояний (например, стенокардии, аритмий, гиперетензии), а также расстройств ЦНС и нейрональных нарушений (например, мигреней, неврологических последствий инсульта, мании, индуцированной нейролептиками поздней дискинезии, шизофрении, биполярного расстройства, нарушении болевых ощущений, эпилепсии и других). Кроме того, было показано, что агонисты кальциевых каналов эффективны для различных применений, таких как уменьшение продолжительности и иным способом истощающего действия местной анестезии. Антагонисты и агонисты CanIon-каналов аналогичным образом пригодны для лечения или предупреждения таких и иных заболеваний и состояний. Например, CanIon-антагонисты пригодны для лечения или предупреждения шизофрении и биполярного расстройства. Поскольку активация потенциалзависимых воротных ионных каналов не требует связывания с аго-нистами, соединения предпочтительно скринируют против функциональных CanIon-каналов. Анализ может включать методы функционального и радиолигандного связывания, применяемые к клеткам (пузырькам или мембранам), экспрессирующим нативные или клонированные каналы, или проверку целых клеток. Для анализа функциональных целых клеток можно использовать электрофизиологические методы, такие как метод фиксации потенциала. В анализе можно использовать любой тип потенциал-зависимого воротного канала, предпочтительно каналы типов L, N и Т. Можно измерить также кинетику протекания ионов через данный канал, например, с использованием методов флуоресценции, конечного радиоактивного индикатора или методов клеточной выживаемости. Анализ можно осуществить с использованием различных токсинов, ядов или соединений, которые, связываясь, открывают или закрывают каналы (Denyer и соавт., Drug Disc. Today 3(7):323-332 (1998), включенный в данном описании путем ссылки во всей его полноте). Один из вариантов осуществления настоящего изобретения включает в себя использование любого из большого числа известных агонистов и антагонистов кальциевых каналов, например, в качестве позитивного или негативного контроля. Примеры подходящих известных антагонистов кальциевых каналов включают фенилалкиламины (например, верапамил), бензотиазепины (например, дилтиазем) и дигидропиридины (например, нифедипин); агони - 65 - 008252 сты кальциевых каналов включают FPL-64176 и BAY К 8644; агонисты натриевых каналов включают Batrachotoxin; а антагонисты натриевых каналов включают спирадолин, мексилетин, U-54494A (+/-)-цис-3,4-дихлор-N-метил-N-[2-(1-пирролидинил)циклогексил]бензамид. Такие соединения можно также использовать в качестве "проводящих" соединений, т.е. служащих в качестве стартовых молекул для создания или обнаружения производных молекул, которые специфически связываются с CanIon-каналами или модулируют их. В предпочтительных вариантах осуществления настоящего изобретения анализ настоящего изобретения включает способ скрининга кандидатного вещества, включающий следующие стадии: a) создания (i) образца или клетки-хозяина, содержащих полипептид, включающий, в основном состоящий, или состоящий из CanIon-белка или его фрагмента, или (ii) рекомбинантной клетки-хозяина, экспрессирующей полинуклеотид, кодирующий полипептид, включающий, в основном состоящий или состоящий из CanIon-белка или его фрагмента; b) получения кандидатного вещества; c) приведения в контакт указанной клетки-хозяина с указанным кандидатным веществом; d) определения эффекта указанного кандидатного вещества на CanIon-активность. Определение эффекта кандидатного вещества на CanIon-активность можно осуществить в соответствии с хорошо известными способами. Предпочтительно, эффект кандидатного вещества на CanIon-активность представляет собой агонистический или антагонистический эффект. Как правило, соединение подавляет CanIon, если его способность транспортировать ионы (например, Са2+ или Na+) снижается. Соединение стимулирует CanIon, если повышается его способность транспортировать ионы. CanIon-активность можно обнаружить при использовании любого подходящего способа. В предпочтительных примерах CanIon-активность выявляют путем измерения сигнального события. Хотя сигнальное событие может включать любое соответствующее изменение молекулярной характеристики или параметра клетки, неограничивающие примеры сигнального события включают изменения потока ионов, такого как изменение образования потока Са2+ или Na+, или К+, или ферментной активности. Один из аспектов состоит в отслеживании ионного потока путем измерения электрофизиологических свойств CanIon-канала с использованием, например, методов измерения потока в одной клетке или на небольшом участке мембраны. В других примерах используют флуоресцентную или радиоактивную метки для выявления перемещения известного CanIon-связывающего соединения или для выявления ионного потока через клетку (например, меченный Са2+ или Na+). Можно использовать индикатор физиологических параметров клетки, такой, например, как флуоресцентный индикатор клеточной жизнеспособности. В других примерах можно выявлять изменение физического положения индикатора с тем, чтобы использовать активированный флуоресценцией клеточный сортинг для идентификации исключения или поглощения физиологического индикатора. Образец, используемый для анализа настоящего изобретения, содержит полипептид или клетку-хозяина, экспрессирующую полипептид, включающий, в основном состоящий, состоящий из CanIon-белка или его фрагмента, или (ii) рекомбинантную клетку-хозяина, экспрессирующую полинуклеотид, кодирующий полипептид, включающий, в основном состоящий, состоящий из CanIon-белка или его фрагмента. Предпочтительно, CanIon-анализ настоящего изобретения включает использование рекомбинантной клетки-хозяина, экспрессирующей функциональный CanIon-полипептид. Клетки-хозяева могут экспрессировать или включать функциональную альфа-субъединицу CanIon-канала или могут экспрес-сировать или включать одну или несколько дополнительных субъединиц ионного канала, или комплексный ионный канал, включающий CanIon. Предпочтительно используется клетка-хозяин, которая обладает низкой эндогенной экспрессией ионного канала или обладает низкой фоновой проводимостью, в частности Са2+ и/или Na+. Радиолигандное связывание Одним из аспектов настоящего изобретения является скринирование CanIon-канала путем идентификации высокоаффинного лиганда, который связывается с представляющим интерес сайтом CanIon и предпочтительно обладает требуемым модулирующим эффектом, и выявления способности испытываемого соединения замещать указанный меченный лиганд. Список токсикологических/фармакологических агентов, используемых в анализах потенциал-зависимых воротных (Са2+, Na+ и К+) каналов приведен у Denyer и соавт. (выше). Данный способ в основном пригоден для выявления соединений, которые связываются с одним и тем же сайтом или аллостерически соединяются с указанным сайтом, в виде меченного лиганда, однако, не предоставляют информацию о свойствах данного соединения как об агонисте или антагонисте. Анализы, основанные на флуоресцентной и радиоактивной индикации клетки В другом анализе CanIon-функцию можно проследить путем измерения изменения внутриклеточной концентрации проницаемого иона с использованием флуоресцентно индикаторных ионов или радиоактивно индикаторных ионов. Обычно ионные каналы, такие как №+-каналы инактивируются в течение миллисекунд после стимуляции потенциала. Са2+-каналы не инактивируются или в меньшей степени инактивируются и могут открываться при высокой К+-деполяризации. В анализах, основанных на флуоресценцентной и радиоак - 66 - 008252 тивной индикации клетки, CanIon-канал можно, как правило, активировать с помощью токсина или любого испытываемого соединения, или высокой К+-деполяризацией, так, чтобы данный канал оказался открытым в течение длительного периода времени (вплоть до нескольких минут). В анализах с флуоресценцией используют доступные флуоресцентные Са2+-красители (например, Fluo-3, Calcium green-1, Молекулярные Зонды, OR, США) . Са2+-каналы можно активировать деполяризацией мембраны с помощью изотоничного раствора, a №+-каналы - с помощью токсина или иного соединения, и полученное в результате нестационарное передвижение флуоресценции в данной клетке можно измерить на протяжении 20-60 с. Системы по измерению флуоресценции и приборы дополнительно описываются у Denyer и соавт. (выше). Радиоактивные индикаторы 22Na+ и 14С-гуанидин обычно используют для анализа №+-каналов, а 45Са2+ используют для анализа Са2+-каналов. В предпочтительном варианте, описанном у Denyer и соавт. (выше), высокопроизводительный клеточный анализ Can-Ion осуществляют с использованием сцинтилляционных Cytostar-T-микроплашек (Amersham International, U.K.). В последующих анализах функцию Са2+-ионного канала отслеживают путем измерения мембранного потенциала с помощью индикатора мембранного потенциала. Высокое электрическое сопротивление биологических мембран позволяет малым ионным потокам через плазматическую мембрану вызывать большие изменения мембранного потенциала. Таким образом, анализ потенциала удобно использовать для выявления общего ионного потока через мембраны. Клеточные линии, как правило, выбирают так, чтобы минимизировать действие эндогенных ионных каналов. Ряд красителей, доступных в качестве индикаторных красителей мембранного потенциала, делятся на красители быстрого и медленного ответа, а также на потенциал-сенсорные красители, основанные на FRET (Aurora Biosciences, CA, США; обзор Gonzalez и соавт., Drug Disc. Today 4(9):431-439 (1999)). Жизнеспособность клеток В анализах жизнеспособности клеток активность ионных каналов и поток ионов непосредственно связаны с жизнеспособностью клетки. Клеточные системы дрожжей и млекопитающих пригодны для тестирования ионного канала-мишени. Например, используют дрожжевую систему, представленную ион-специфичной, дефицитной по поглощению К+ клеточной линией Saccharomyces cerevisiae, в которой представляющий интерес функциональный К+-канал экспрессируется в данной клеточной линии, восстанавливая тем самым поглощение К+ и способствуя клеточному выживанию (Anderson и соавт. Symp. Soc. Exp. Biol. 48:85-97 (1994)). Такой анализ для Са2+ или №+-каналов можно применять для идентификации соединений, способных блокировать CanIon-функцию. Доступны также клеточные системы млекопитающих, такие как №+-каналы, анализируемые с использованием колориметрических данных по жизнеспособности нейробластомных клеток млекопитающих. Клетки обрабатывают с помощью открывателя №+-каналов и ингибитора №+/К+-насоса, чтобы способствовать созданию летального избытка внутриклеточного Na+. Обработка с помощью испытываемого соединения, способного блокировать данные каналы, будет увеличивать клеточную жизнеспособность, а соединения, которые усиливают открывание каналов, в последующем будут способствовать гибели клеток (Manger и соавт., Anal. Biochem. 214:190-194 (1993)). Электрофизиология Электрофизиологические методы фиксации потенциала включают измерение ионного тока, протекающего через один или множество каналов. Единственным микроэлектродом контролируют мембранный потенциал, когда электрический ток измеряют в отдельной клетке или на небольшом участке мембраны (Hamill, Pfugers Arch. 391, 85-100 (1981)). Ионный ток можно, следовательно, измерить в присутствии или в отсутствие представляющего интерес испытываемого соединения. Была создана система для скрининга большого числа соединений (Neurosearch A/S, Glostrup, Дания; Olesen и соавт., Voltage gated ion channel modulators, 7-8 December, Philadelphia PA, США (1995); Denyer и соавт., выше). Способы скринирования веществ, взаимодействующих с CanIon-полипептидом Для цели настоящего изобретения под лигандом подразумевают молекулу, такую как белок, пептид, антитело или любое синтетическое химическое соединение, способное связаться с CanIon-белком или с одним из его фрагментов или вариантов, или модулировать экспрессию полинуклеотида, кодирующего CanIon, его фрагмент или вариант. В соответствии с настоящим изобретением в способе скринирования лиганда биологический образец или указанную молекулу, которую испытывают в качестве предполагаемого лиганда CanIon-белка, приводят в контакт с соответствующим выделенным очисткой CanIon-белком, например, с соответствующим очищенным рекомбинантным CanIon-белком, полученным с помощью рекомбинантной клетки-хозяина, как описано выше, с тем, чтобы создать комплекс между данным белком и испытываемой предполагаемой лигандной молекулой. В качестве иллюстративного примера, для изучения взаимодействия CanIon-белка или фрагмента, включающего непрерывный участок последовательности, по меньшей мере, из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50, 100, 200, 300, 400, 500 или 1000 аминокислот SEQ ID No 5, с лекарственным средством или с малыми молекулами, такими как молекулы, полученные с помощью методов комбинаторной химии, могут - 67 - 008252 быть использованы метод микродиализа, соединенный с ВЭЖХ, описанный Wang и соавт. (1997), или метод капиллярного электрофореза по сродству, описанный Bush и соавт. (1997), раскрытие которых включено путем ссылки. В дополнительных способах пептиды, лекарственные средства, жирные кислоты, липопротеины или малые молекулы, которые взаимодействуют с CanIon-белком или с фрагментом, включающим непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно, по меньшей мере, 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50 или 100 аминокислот SEQ ID No 5, могут быть идентифицированы с использованием анализов, таких как следующие ниже. Испытываемую на связывание молекулу метят детектируемой меткой, такой как флуоресцентная, радиоактивная или ферментная метка, и помещают для контактирования с иммобилизованным CanIon-белком или его фрагментом в условиях, позволяющих происходить специфическому связыванию. После удаления неспецифически связавшихся молекул, связавшиеся молекулы детектируют с использованием подходящих средств. Другая цель настоящего изобретения включает способы и наборы для скринирования кандидатных веществ, которые взаимодействуют с CanIon-полипептидом. Настоящее изобретение относится к способам скринирования представляющих интерес веществ, которые взаимодействуют с CanIon-белком или с каким-либо его фрагментом или вариантом. Исходя из их способности связываться ковалентно или нековалентно с CanIon-белком или с его фрагментом или вариантом, данные вещества или молекулы могут преимущественно использоваться in vitro и in vivo. Указанные взаимодействующие молекулы могут использоваться in vitro в качестве детекционных средств идентификации присутствия CanIon-белка в образце, предпочтительно в биологическом образце. Способ для скринирования кандидатного вещества включает следующие стадии: a) создания полипептида, включающего, в основном состоящего, состоящего из CanIon-белка или фрагмента, включающего непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50 или 100 аминокислот SEQ ID No 5; b) получения кандидатного вещества; c) приведения в контакт указанного полипептида с указанным кандидатным веществом; d) выявления комплексов, образованных между указанным полипептидом и указанным кандидат-ным веществом. Кроме того, в настоящем изобретении рассматривается набор для скринирования кандидатного вещества, взаимодействующего с CanIon-полипептидом, где указанный набор включает: a) CanIon-белок, обладающий аминокислотной последовательностью, выбранной из группы, состоящей из аминокислотных последовательности SEQ ID No 5 или ее пептидного фрагмента, включающего непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50 или 100 аминокислот SEQ ID No 5; b) факультативно, средства, пригодные для выявления комплекса, образованного между CanIon-белком или его пептидным фрагментом или вариантом и кандидатным веществом. В предпочтительном варианте осуществления настоящего изобретения в описанном выше наборе, детекционные средства включают моноклональные или поликлональные антитела, направленные против CanIon-белка или его пептидного фрагмента или варианта. Можно проанализировать разнообразные кандидатные вещества на взаимодействие с CanIon-полипептидом. Данные вещества или молекулы включают, без ограничения, природные или синтетические органические соединения или молекулы биологического происхождения, такие как полипептиды. Если кандидатное вещество или молекула включает полипептид, то полипептид может представлять собой экспрессионный продукт, полученный из фагового клона, относящегося к произвольно созданной пептидной библиотеке на основе фага, или, альтернативно, данный полипептид может представлять собой экспрессионный продукт, полученный из кДНК-библиотеки, клонированной в вектор, пригодный для осуществления дигибридного скринирующего анализа. Настоящее изобретение относится также к наборам, пригодным для осуществления описанного выше способа скринирования. Предпочтительно, такие наборы включают CanIon-полипептид или его фрагмент или вариант и, факультативно, средства, пригодные для выявления комплекса, образованного между CanIon-полипептидом или его фрагментом или вариантом и данным кандидатным веществом. В предпочтительном варианте осуществления настоящего изобретения детекционные средства включают моноклональные и поликлональные антитела, специфичные к соответствующему CanIon-полипептиду или его фрагменту или варианту. А. Лиганды-кандидаты, полученные из произвольных пептидных библиотек. В конкретном варианте осуществления способа скринирования настоящего изобретения, предполагаемый лиганд представляет собой экспрессионный продукт ДНК-вставки, содержащейся в фаговом векторе (Parmley and Smith, 1988). В данном случае используются произвольные пептидные библиотеки на основе фагов. Произвольные ДНК-вставки кодируют пептиды длиной 8-20 аминокислот (Oldenburg K.R. - 68 - 008252 и соавт., 1992; Valadon P., и соавт., 1996; Lucas А.Н., 1994; Westerink M.A.J., 1995; Felici F. и соавт., 1991). В соответствии с этим конкретным вариантом, рекомбинантные фаги, экспрессирующие белок, который связывается с иммобилизованным CanIon-белком, сохраняется, а комплекс, образованный между CanIon-белком и рекомбинантным фагом, можно впоследствии иммунопреципитировать с помощью поликлонального или моноклонального антитела, специфичного к CanIon-белку. После того как лигандную библиотеку в рекомбинантных фагах сконструировали, фаговую популяцию приводят в контакт с иммобилизованным CanIon-белком. Затем полученные комплексы отмывают для того, чтобы удалить неспецифически связавшиеся рекомбинантные фаги. Фаги, которые специфически связались с CanIon-белком, элюируют буфером (кислый рН) или иммунопреципитируют монокло-нальным антителом, продуцированным в гибридомой анти-CanIon, и данную фаговую популяцию впоследствии размножают с помощью сверхинфицированных бактерий (например, Е. coli). Селекционную стадию можно повторить несколько раз, предпочтительно 2-4 раза, для того, чтобы отобрать более специфичные рекомбинантные фаговые клоны. Последняя стадия включает характеристику данного пептида, продуцированного рекомбинантным фаговым клоном, либо путем экспрессии в инфицированных бактериях и выделения, экспрессируя фаговую вставку в другой системе "хозяин-векторная ДНК", либо секвенированием данной вставки, содержащейся в отобранных рекомбинантных фагах. B. Лиганды-кандидаты, полученные с помощью конкурентных экспериментов. Альтернативно, пептиды, лекарственные средства или малые молекулы, которые связываются с CanIon-белком или фрагментом, включающим непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50 или 100 аминокислот SEQ ID No 5, можно идентифицировать в конкурентных экспериментах. В таких анализах CanIon-белок или его фрагмент, иммобилизуют на поверхности, такой как пластмассовый планшет. Увеличивающиеся количества пептидов, лекарственных средств или малых молекул контактируют с иммобилизованным CanIon-белком или его фрагментом в присутствии обнаруживаемого с помощи метки известного лиганда CanIon-белка. Например, CanIon-лиганд можно пометить меткой с помощью флуоресцентной, радиоактивной или ферментной метки. Способность испытываемой молекулы связывать CanIon-белок или его фрагмент, определяют измерением количества обнаруживаемой метки в известном лиганде, связавшемся в присутствии данной тест-молекулы. Уменьшение количества известного лиганда, связавшегося с CanIon-белком или его фрагментом, при наличии тест-молекулы, свидетельствует о том, что данная тест-молекула способна связываться с CanIon-белком или его фрагментом. C. Лиганды-кандидаты, полученные с помощью аффинной хроматографии. Белки или иные молекулы, взаимодействующие с CanIon-белком или фрагментом, включающим непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно по меньшей мерее 12, 15, 20, 25, 30, 40, 50 или 100 аминокислот SEQ ID No 5, можно индуцировать с помощью аффинной хроматографии. В таком анализе CanIon-белок или его фрагмент можно присоединить к колонке с использованием традиционных методов, в том числе химически связать с соответствующим связующим веществом колонки, таким как агароза, Affi Gel(r) или с иными матрицами, хорошо известными специалистам в данной области техники. Для некоторых вариантов осуществления данного способа аффинная колонка содержит химерные белки, в которых CanIon-белок или его фрагмент соединен путем слияния с глутатион^-трансферазой (GST). Смесь клеточных белков или пул экспрессированных белков, которые описаны выше, наносят на аффинную колонку. Белки или другие молекулы, взаимодействующие с CanIon-белком или его фрагментом, присоединяемые к данной колонке, можно затем выделить и проанализировать 2D-гель-электрофорезом, как описано у Ramunsena и соавт. (1997), раскрытие которого включено путем ссылки. Альтернативно, белки, удерживаемые на данной аффинной колонке, можно выделить очисткой способами, основанными на электрофорезе, и секвенировать. Тот же самый способ можно использовать для выделения антител, для скринирования продуктов фагового дисплея или для скринирования антител человека фагового дисплея. D. Лиганды-кандидаты, полученные с помощью способов оптического биосенсора. Белки, взаимодействующие с CanIon-белком или с фрагментом, включающим непрерывный участок последовательности по меньшей мере из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50 или 100 аминокислот SEQ ID No 5, можно также скринировать с использованием Оптического Биосенсора, как описано у Edwards and Leatherbarrow (1997), а также у Szabo и соавт. (1995), раскрытие которых включено путем ссылки. Данный способ позволяет обнаруживать взаимодействие между молекулами в реальном времени, без необходимости метить молекулы. Данный метод основан на явлении поверхностного плазменного резонанса (SPR). Вкратце, тестируемую молекулу лиганда-кандидата присоединяют к поверхности (такой как карбоксиметилдекстрановая матрица). Луч света, направленный на сторону поверхности, которая не содержит тестируемого образца, отражается указанной поверхностью. Эффект SPR вызывает уменьшение интенсивности отраженного света, что взаимосвязано с определенным углом и длиной волны. Связывание молекул лиганда-кандидата обуславливает изменение коэффициента преломления на поверхности, изменение которого обнаруживается в виде изменения SPR-сигнала. Для скринирования молекул лиган - 69 - 008252 да-кандидата или веществ, которые способны взаимодействовать с CanIon-белком или его фрагментом, CanIon-белок или его фрагмент иммобилизуют на поверхности. Данная поверхность включает одну сторону клетки, через которую протекает анализируемая молекула-кандидат. Связывание молекулы-кандидата к CanIon-белку или его фрагменту обнаруживается в виде изменения SPR-сигнала. Тестируемые молекулы-кандидаты могут представлять собой белки, пептиды, углеводы, липиды или малые молекулы, образованные с помощью комбинаторной химии. Данный метод можно также осуществлять путем иммобилизации эукариотических или прокариотических клеток или липидных везикул, демонстрирующих на своей поверхности эндогенно или рекомбинантно экспрессируемый CanIon-белок. Основное преимущество данного способа заключается в том, что он позволяет определить скорость связывания CanIon-белка и молекул, взаимодействующих с CanIon-белком. Таким образом представляется возможным специально выбрать лигандные молекулы, взаимодействующие с CanIon-белком или с его фрагментом, благодаря константам сильной или, наоборот, слабой ассоциации. Е. Лиганды-кандидаты, полученные с помощью дигибридного скринирующего анализа. Для изучения in vivo взаимодействий белок-белок создана дрожжевая дигибридная система (Fields and Song, 1989), основывающаяся на слиянии белка-затравки с ДНК-связытающим доменом дрожжевого Gа14-белка. Данный метод описан в патентах США №№ 5667973 и 5283173 (Fields и соавт.), представленные там методические указания включены путем ссылки. Основную операцию скринирования библиотеки с помощью дигибридного анализа можно осуществить как описано у Harper и соавт. (1993), у Cho и соавт. (1998) или у Fromont-Racine и соавт. (1997). Затравочный белок или полипептид включает, состоит в основном или состоит из CanIon-полипептида или фрагмента, включающего непрерывный участок последовательности, по меньшей мере, из 6 аминокислот, предпочтительно по меньшей мере 8-10 аминокислот, более предпочтительно по меньшей мере 12, 15, 20, 25, 30, 40, 50 или 100 аминокислот SEQ ID No 5. Более точно, нуклеотидную последовательность, кодирующую CanIon-полипептид, его фрагмент или вариант, соединяют путем слияния с полинуклеотидом, кодирующим ДНК-связывающий домен GAL4-белка, причем полученную слитую нуклеотидную последовательность встраивают в соответствующий экспрессионный вектор, например, pAS2 или рМ3. Затем в специально созданном векторе конструируют кДНК-библиотеку человека так, чтобы кДНК-вставка человека сливалась с нуклеотидной последовательностью в векторе, который кодирует транскрипционный домен GAL4-белка. Предпочтительно, данный используемый вектор представляет собой рАСТ-вектор. Полипептиды, кодируемые нуклеотидными вставками из кДНК-библиотеки человека, являются полипептидами-"добычами". Третий вектор содержит детектируемый маркерный ген, такой как бета-галактозидазный ген или САТ-ген, который помещают под контроль регуляторной последовательности, которая ответственна за связывание полноразмерного Gа14-белка, содержащего домен активации транскрипции и ДНК-связывающий домен. Можно использовать, например, pG5EC-вектор. Используются также два разных дрожжевых штамма. В качестве иллюстративного, но не ограничивающего примера, два разных дрожжевых штамма могут являться следующими штаммами: Y190, фенотип которого представляет собой (МАТа, Leu2-3, 112 ura3-12, trp1-901, his3-D200, ade2-101, ga14Dgal180D URA3 GAL-LacZ, LYS GAL-HIS3, cyh'); Y187, фенотип которого представляет собой (МАТа gа14 gа180 his3 trp1-901 ade2-101 ura3-52 leu2-3, -112URA3 GAL-lacZmet-), который представляет собой тип спаривания противоположный Y190. Вкратце, 20 мкг pAS2/CanIon и 20 мкг рАСТ-кДНК-библиотеки котрансформируют в дрожжевой штамм Y190. Полученные трансформанты отбирают по росту на минимальной среде без гистидина, лейцина и триптофана, но содержащей ингибитор синтеза тистидина 3-АТ (50 мМ). Позитивные колонии скринируют на бета-галактозидазу с помощью анализа отпечатков на фильтре. Затем двойные позитивные колонии (His+, beta-gal+) выращивают на чашках без гистидина, лейцина, но с триптофаном и цикло-гексимидом (10 мг/мл) для отбора колоний с утраченными pAS2/CanIon-плазмидами, но сохранившими плазмиды рАСТ-кДНК-библиотеки. Полученные Y190-штаммы спаривают с Y187-штаммами, экспрес-сирующими CanIon или неродственные контрольные белки; такие как циклофилин В, ламин или SNF1, в виде GaM-химер, как описано у Harper и соавт. (1993) и у Bram и соавт. (Bram R.J. и соавт., 1993), и скринируют на бета-галактозидазу с помощью анализа скринирования колоний на фильтре. Дрожжевые клоны, которые представляют собой beta gal- после спаривания с контрольными GaW-химерами, считают ложнопозитивными. В другом варианте осуществления дигибридного способа, в соответствии с настоящим изобретением, взаимодействие между CanIon или его фрагментом или вариантом с клеточными белками можно оценить с помощью Matchmaker Two Hybrid System 2 (Catalog No K1604-1, Clontech). Как описано в инструкции, прилагаемой к Matchmaker Two Hybrid System 2 (Catalog No K1604-1, Clontech), раскрытие которой включено в данное описание путем ссылки, нуклеиновые кислоты, кодирующие CanIon-белок или его часть, встраивают в экспрессионный вектор так, чтобы они находились в рамке считывания с ДНК, кодирующей ДНК-связывающий домен дрожжевого транскрипционного активатора GAL4. Требуемая кДНК, предпочтительно кДНК человека, встраивается во второй экспрессионный вектор так, чтобы они - 70 - 008252 находились в рамке считывания с ДНК, кодирующей активаторный домен GAL4. Две данные экспресси-онные плазмиды трансформируют в дрожжи и трансформированные дрожжи высевают на селективную среду, которая селектирует по экспрессии селективных маркеров в каждом экспрессионном векторе, а также GAL4-зависимой экспрессии HIS3-гена. Трансформанты, способные расти на среде без гистидина, скринируют на GAL4-зависимую lacZ-экспрессию. Те клетки, которые оказываются позитивными при гистидиновом отборе и lacZ-анализе, удерживают взаимодействие между CanIon и белком или пептидом, кодируемым исходно выбранной кДНК-вставкой. Способ скринирования веществ, взаимодействующих с регуляторными последовательностями CanIon-гена В настоящем изобретении рассматривается также способ скринирования веществ или молекул, которые способны взаимодействовать с регуляторными последовательностями CanIon-гена, такими как, например, промоторная или энхансерная последовательности. Нуклеиновые кислоты, кодирующие белки, которые способны взаимодействовать с регуляторными последовательностями CanIon-гена, более предпочтительно с нуклеотидной последовательностью, выбранной из группы, состоящей из полинуклеотидов 5'- и 3'-регуляторной области или ее фрагмента или варианта, и предпочтительно варианта, включающего один из двуаллельных маркеров настоящего изобретения, могут быть идентифицированы с использованием моногибридной системы, такой, которая описана в проспекте, прилагаемом к набору Matchmaker One-Hybrid System от Clontech (Catalog Ref n° K1603-1), методические указанная которого, представленные в данном описании, включены путем ссылки. Вкратце, нуклеотидную последовательность-мишень клонируют слева от селективной репортерной последовательности, а полученную ДНК-конструкцию интегрируют в дрожжевой геном (Saccharomyces cerevisiae). Дрожжевые клетки, содержащие в своем геноме данную репортерную последовательность, трансформируют затем с помощью библиотеки, включающей кДНК, кодирующие белки-кандидаты для связывания с регуляторными последовательностями CanIon-гена, слитые с последовательностями, кодирующими активаторный домен дрожжевого фактора транскрипции, такого как GAL4. Рекомбинантные дрожжевые клетки высевают в культуральный бульон для селекции клеток, экспрессирующих указанную репортерную последовательность. Отобранные таким образом рекомбинантные дрожжевые клетки содержат слитый белок, который способен связываться с регуляторной последовательностью-мишенью CanIon-гена. После чего кДНК, кодирующие слитые белки, секвенируют и их можно клонировать in vitro в экспрессионный или транскрипционный векторы. Связывание кодируемых полипептидов с регулятор-ными последовательностями-мишенями CanIon-гена можно подтвердить методами, известными специалистам в данной области техники, такими как метод задержки в геле или метод защиты от ДНКазы. Метод задержки в геле можно также осуществить независимо для того, чтобы скринировать молекулы-кандидаты, которые способны взаимодействовать с регуляторными последовательностями CanIon-гена так, как описано у Fried and Crothers (1981), у Garner and Revzin (1981) и у Dent and Latchman (1993), методические указания представленных там публикаций включены путем ссылки. Данные методы основываются на принципе, согласно которому фрагмент ДНК, который связывается с белком, мигрирует медленнее, чем такой же несвязавшийся ДНК-фрагмент. Вкратце, метят нуклеотидную последовательность-мишень. Затем меченную нуклеотидную последовательность-мишень приводят в контакт либо с тотальным ядерным экстрактом из клеток, содержащим факторы транскрипции, либо с разными тестируемыми молекулами-кандидатами. Взаимодействие между регуляторной последовательностью-мишенью CanIon-гена и молекулой-кандидатом или фактором транскрипции выявляют после гелевого или капиллярного электрофореза благодаря задержке миграции. Способы скринирования лигандов, которые модулируют экспрессию CanIon-гена Другим объектом рассмотрения настоящего изобретения является способ скринирования молекул, которые модулируют экспрессию CanIon-белка. Такой способ скринирования включает следующие стадии: a) культивирования прокариотической или эукариотической клетки, которую трансформировали нуклеотидной последовательностью, кодирующей CanIon-белок, его вариант или фрагмент, помещенной под контроль своего собственного промотора; b) приведения в контакт данной культивируемой клетки и тестируемой молекулы; c) количественной оценки экспрессии CanIon-белка или его варианта или фрагмента. В варианте осуществления настоящего изобретения нуклеотидная последовательность, кодирующая CanIon-белок или его вариант или фрагмент, включает аллель, по меньшей мере, одного из двуаллельных маркеров А12 или А16 и их комплементов. Используя методы рекомбинантных ДНК, хорошо известные специалистам в данной области техники, последовательность ДНК, кодирующую CanIon-белок, встраивают в экспрессионный вектор, справа от его промоторной последовательности. В качестве иллюстративного примера, промоторная последовательность CanIon-гена содержится в нуклеиновой кислоте 5'-регуляторной области. Количественную оценку экспрессии CanIon-белка можно осуществить либо на уровне мРНК, либо на белковом уровне. В последнем случае можно использовать поликлональные или моноклональные антитела для измерения количества синтезированного CanIon-белка, например, в ELISA или RIA-анализом. В предпочтительном варианте осуществления настоящего изобретения количественное измерение - 71 - 008252 CanIon-мРНК осуществляют с помощью количественной ПЦР-амплификации кДНК, полученной с помощью обратной транскрипции тотальной мРНК из культивированных клеток-хозяев, трасфицирован-ных CanIon, с использованием пары праймеров, специфичных для CanIon. В настоящем изобретении рассматривается также способ скринирования веществ или молекул, которые способны повысить или понизить уровень экспрессии CanIon-гена. Такой способ может позволить специалисту в данной области техники выбрать вещества, проявляющие регулирующее действие на уровень экспрессии CanIon-гена, и которые могут быть пригодны в качестве активных ингредиентов, включаемых в фармацевтические композиции для лечения пациентов, страдающих от любой описанной в данном описании болезни. Таким образом, в настоящем изобретении создан также способ скринирования кандидатного вещества или молекулы, которые модулируют экспрессию CanIon-гена, причем данный способ включает следующие стадии: создания рекомбинантной клетки-хозяина, содержащей нуклеиновую кислоту, где указанная нуклеиновая кислота включает нуклеотидную последовательность 5'-регуляторной области или ее биологически активный фрагмент или вариант, локализованную "левее" полинуклеотида, кодирующего выявляемый белок; получения кандидатного вещества; и определения способности кандидатного вещества модулировать уровень экспрессии полинуклеоти-да, кодирующего выявляемый белок. В дополнительном варианте осуществления настоящего изобретения нуклеиновая кислота, включающая нуклеотидную последовательность 5'-регуляторной области или ее биологически активный фрагмент или вариант, включает также 5'UTR-область CanIon-кДНК SEQ ID No 4 или ее биологически активные фрагменты или варианты. Среди предпочтительных полинуклеотидов, кодирующих выявляемый белок, можно привести по-линуклеотиды, кодирующие бета-галактозидазу, зеленый флуоресцентный белок (GFR) и хлорамфени-колацетилтрансферазу (CAT). Настоящее изобретение относится также к наборам, пригодным для осуществления описанного в данном описании способа скринирования. Предпочтительно, такие наборы включают рекомбинантный вектор, который обеспечивает экспрессию нуклеотидной последовательности 5'-регуляторной области или ее биологически активного фрагмента или варианта, локализованной "левее" и функционально присоединенной к полинуклеотиду, кодирующему выявляемый белок или CanIon-белок или его фрагмент или вариант. В другом способе скринирования кандидатного вещества или молекулы, которые модулируют экспрессию CanIon-гена, данный способ включает следующие стадии: a) создания рекомбинантной клетки-хозяина, содержащей нуклеиновую кислоту, где указанная нуклеиновая кислота включает 5' UTR-последовательность CanIon-кДНК SEQ ID No 4 или один из ее биологически активных фрагментов или вариантов, причем 5'UTR-последовательность или ее биологически активный фрагмент или вариант функционально присоединен к полинуклеотиду, кодирующему выявляемый белок; b) получения кандидатного вещества; и c) определения способности кандидатного вещества модулировать уровни экспрессии полинуклео-тида, кодирующего выявляемый белок. В конкретном варианте осуществления указанного выше способа скринирования, нуклеиновая кислота, которая включает нуклеотидную последовательность, выбранную из группы, состоящей из 5' UTR-последовательности CanIon-кДНК SEQ ID No 4 или из ее биологически активных фрагментов или вариантов, включает промоторную последовательность, которая является эндогенной относительно Can-Ion-5'UTR-последовательности. В другом конкретном варианте осуществления указанного выше способа скринирования нуклеиновая кислота, которая включает нуклеотидную последовательность, выбранную из группы, состоящей из 5'UTR-последовательности CanIon-кДНК SEQ ID No 4 или одного из ее биологически активных фрагментов или вариантов, включает промоторную последовательность, которая является экзогенной по отношению к указанной в данном описании CanIon-5'UTR-последовательности. В дополнительном предпочтительном варианте осуществления настоящего изобретения нуклеиновая кислота, включающая 5'UTR-последовательность CanIon-кДНК или SEQ ID No 4 или ее биологически активные фрагменты, включает двуаллельный маркер, выбранный из группы, состоящей из А12 или А16 или их комплементов. Кроме того, настоящее изобретение включает набор для скринирования кандидатного вещества, модулирующего экспрессию CanIon-гена, где указанный набор включает рекомбинантный вектор, который включает нуклеиновую кислоту, включающую 5'UTR-последовательность CanIon-кДНК SEQ ID No 4 или один из ее биологически активных фрагментов или вариантов, причем данная 5'UTR-последовательность или ее биологически активный фрагмент или вариант функционально присоединен к полинуклеотиду, кодирующему выявляемый белок. - 72 - 008252 Уровни экспрессии и паттерны CanIon можно анализировать в гибридизационном растворе с помощью длинных зондов, как описано в международной патентной заявке № WO 97/05277, полное содержание которой включено в данное описание путем ссылки. Вкратце, CanIon-кДНК или геномная ДНК Can-Ion, описанные выше, или их фрагменты встраивают в клонирующий сайт сразу "правее" от промотора РНК-полимеразы бактериофага (Т3, Т7 или SP6) для получения антисмысловой РНК. Предпочтительно, CanIon-вставка включает, по меньшей мере, 100 или более последовательных нуклеотидов из геномной ДНК-последовательности или из кДНК-последовательностей. Полученную плазмиду линеаризуют и транскрибируют в присутствии рибонуклеотидов, включающих модифицированные рибонуклеотиды (т.е. биотин-UTP и DIG-UTP). Избыток такой вдвойне меченной РНК гибридизуют в растворе с мРНК, выделенной из представляющих интерес клеток или тканей. Гибридизацию осуществляют в стандартных жестких условиях (40-50°С в течение 16 ч в 80% формамиде, 0,4 М NaCl-буфере, рН 7-8). Непрогибриди-зовавшийся зонд удаляют путем расщепления рибонуклеазами, специфичными для одноцепочечной РНК (т.е. РНКазы CL3, T1, Phy M, U2 или А). Наличие биотин-UТР-модификации делает возможным поглощение гибрида на планшете для микротитрования, покрытого стрептавидином. Наличие DIG-модификации делает возможным выявление гибрида и его количественную оценку в ELISA с использованием антитела к DIG, соединенного со щелочной фосфатазой. Количественный анализ экспрессии CanIon-гена можно также осуществить с использованием наборов. Используемый в данном описании термин "набор" означает одномерное, двухмерное или мульти-мерное расположение множества нуклеиновых кислот достаточной длины, которые позволяют специфически обнаруживать экспрессию мРНК, способных гибридизоваться с ними. Например, наборы могут содержать множество нуклеиновых кислот, полученных из генов, для которых оценивают уровни экспрессии. Данные наборы могут включать геномную ДНК CanIon, кДНК-последовательности CanIon или последовательности, комплементарные им или их фрагментам, в частности, последовательности, включающие по меньшей мере один из двуаллельных маркеров, в соответствии с настоящим изобретением, предпочтительно по меньшей мере один из двуаллельных маркеров А1-А17. Предпочтительно, фрагменты составляют по меньшей мере 15 нуклеотидов в длину. В других вариантах осуществления настоящего изобретения данные фрагменты содержат по меньшей мере 25 нуклеотидов в длину. В некоторых вариантах осуществления настоящего изобретения данные фрагменты содержат по меньшей мере 50 нуклео-тидов в длину. Более предпочтительно, если данные фрагменты содержат по меньшей мере 100 нуклео-тидов в длину. В других предпочтительных вариантах фрагменты содержат более 100 нуклеотидов в длину. В некоторых вариантах осуществления настоящего изобретения данные фрагменты могут содержать более 500 нуклеотидов в длину. Например, количественный анализ экспрессии CanIon-гена можно осуществлять с помощью микронабора комплементарной ДНК, как описано у Schena и соавт. (1995 и 1996). Полноразмерные кДНК CanIon или их фрагменты амплифицируют с помощью ПЦР и выкладывают из 96-луночного планшета для микротитрования на силилированные предметные стекла для микроскопа с использованием высокоскоростной роботехники. Отпечатанные наборы инкубируют во влажной камере, чтобы обеспечить обводнение элементов набора, и ополаскивают однократно в 0,2% SDS в течение 1 мин, дважды в воде в течение 1 мин и однократно в течение 5 мин в растворе борогидрида натрия. Данные наборы погружают в воду на 2 мин при 95°С, переносят в 0,2% SDS на 1 мин, дважды ополаскивают водой, сушат на воздухе и хранят в темноте при 25°С. Клеточную или тканевую мРНК выделяют или закупают, а зонды получают с помощью обратной транскрипции за один цикл. Зонды гибридизуют с 1 см2 микронабором под стеклянным покровным стеклом с размером 14 х 14 мм в течение 6-12 ч при 60°С. Наборы отмывают в течение 5 мин при 25°С в нежестком промывочном буфере (1 х SSC/0,2% SDS), затем в течение 10 мин при комнатной температуре в очень строгом промывочном буфере (0,1 х SSC/0,2% SDS). Наборы сканируют в 0,1 х SSC с использованием флуоресцентного лазерного сканирующего устройства, совпадающего с обычным фильтровальным набором. Точные измерения дифференциальной экспрессии получают благодаря средним соотношениям двух независимых гибридизаций. Количественный анализ экспрессии CanIon-гена можно также осуществить с помощью полноразмерных кДНК CanIon или их фрагментов в наборе комплементарных ДНК, как описано у Pietu и соавт. (1996). Полноразмерную CanIon-кДНК или ее фрагменты амплифицируют ПЦР и распределяют на мембранах. Затем мРНК, происходящие из разных тканей или клеток, метят радиоактивными нуклеотидами. После гибридизации и отмывки в контролируемых условиях с помощью фосфорной визуализации или радиоавтографией выявляют гибридизованные мРНК. Проводят повторные эксперименты и затем осуществляют количественный анализ по разному экспрессируемых мРНК. Альтернативно, экспрессионный анализ с использованием геномной ДНК CanIon, кДНК CanIon или ее фрагментов можно сделать с помощью нуклеотидных наборов высокой плотности, как описано у Lockhart и соавт. (1996) и Sosnowsky и соавт. (1997). Олигонуклеотиды в 15-20 нуклеотидов из последовательностей геномной ДНК CanIon или кДНК-последовательностей CanIon, особенно тех из них, которые включают по меньшей мере один из двуаллельных маркеров, в соответствии с настоящим изобретением, предпочтительно по крайней мере один из двуаллельных маркеров, выбранных из группы, состоя - 73 - 008252 щей из А1-А17 или комплементарных им последовательностей, синтезируют непосредственно на чипе (Lockhart и соавт., выше) или синтезируют и затем переносят на чип (Sosnowski и соавт., выше). Предпочтительно, длина олигонуклеотидов составляет около 20 нуклеотидов. Зонды кДНК CanIon, меченые с помощью соответствующего соединения, такого как биотин, дигок-сигенин или флуоресцентный краситель, синтезируют из соответствующей мРНК-популяции и затем фрагментируют случайным образом на куски размером 50-100 нуклеотидов. Затем данные зонды гибри-дизуют с чипом. После отмывки, как описано у Lockhart и соавт., выше, и приложения разных электрических полей (Sosnowski и соавт., 1997) красители или меченные соединения выявляют и количественно определяют. Осуществляют повторные гибридизации. Сравнительный анализ интенсивности сигнала, идущего от кДНК-зондов на одном и том же олигонуклеотиде-мишени в разных образцах кДНК, свидетельствует о разной экспрессии мРНК CanIon. Способы ингибирования экспрессии CanIon-гена Другие терапевтические композиции, в соответствии с настоящим изобретением, включают олиго-нуклеотидный фрагмент нуклеиновой последовательности CanIon в качестве антисмыслового инструмента или трехспирального инструмента, который ингибирует экспрессию соответствующего CanIon-гена. Предпочтительный фрагмент нуклеиновой последовательности CanIon включает аллель по меньшей мере одного из двуаллельных маркеров А1-А17. Антисмысловой способ Предпочтительные способы использования антисмысловых полинуклеотидов, в соответствии с настоящим изобретением, представлены методами, описанными у Sczakiel и соавт. (1995). Предпочтительно, антисмысловые инструменты выбраны из полинуклеотидов (длиной 15-200 т.п.н.), которые комплементарны 5'-концу CanIon-мРНК. В другом варианте осуществления настоящего изобретения используют сочетание разных антисмысловых полинуклеотидов, комплементарных разным частям требуемого гена-мишени. Предпочтительные антисмысловые полинуклеотиды, в соответствии с настоящим изобретением, комплементарны последовательностям мРНК CanIon, которые содержат кодон инициации трансляции ATG либо донорный, либо акцепторный сайт сплайсинга. Антисмысловые нуклеиновые кислоты должны обладать длиной и температурой плавления, позволяющими образовать внутриклеточный дуплекс достаточной стабильности для ингибирования экспрессии CanIon-мРНК в этом дуплексе. Методология создания антисмысловых нуклеиновых кислот, пригодных для использования в генной терапии, приводится у Green и соавт. (1986) и Izant and Weintraub (1984), раскрытие которых включено в данное описание путем ссылки. В некоторых методах антисмысловые молекулы получают путем изменения ориентации CanIon-кодирующей области относительно промотора так, чтобы транскрибировать цепь, противоположную той, которая нормально транскрибируется в данной клетке. Антисмысловые молекулы можно транскрибировать с использованием in vitro систем транскрипции, таких как те, которые используют Т7- или SР6-полимеразу для создания транскрипта. Другой подход включает в себя транскрипцию антисмысловых нуклеиновых кислот CanIon in vivo в результате функционального присоединения ДНК, содержащей антисмысловую последовательность, к промотору в соответствующем экспрессионном векторе. Альтернативно, подходящие антисмысловые методы представляют собой те методы, которые описаны у Rossi и соавт. (1991), в международных заявках №№ WO 94/23026, WO 95/04141, WO 92/18522 и в европейской патентной заявке № ЕР 0572287А2. Альтернатива антисмысловой технологии, которая используется в соответствии с настоящим изобретением, включает использование рибозимов, которые связываются с последовательностью-мишенью с помощью их комплементарных полинуклеотидных хвостов, которые расщепляют соответствующую РНК в результате гидролиза ее сайта-мишени (а именно рибозимов в форме "головки молотка"). Вкратце, упрощенный цикл рибозима в форме "головки молотка" включает (1) последовательность, специфически связывающуюся с РНК-мишенью с помощью комплементарных антисмысловых последовательностей; (2) сайт-специфичный гидролиз расщепляемого мотива цепи-мишени; и (3) высвобождение расщепляемых продуктов, которые дают начало другому каталитическому циклу. Действительно, использование длинноцепочечного антисмыслового полинуклеотида (по меньшей мере, длиной 30 оснований) или ри-бозимов с длинными антисмысловыми плечами дает преимущество. Предпочтительная система доставки для антисмыслового рибозима осуществляется в результате ковалентного связывания таких антисмысловых рибозимов с липофильными группами или с использованием липосом в качестве удобного вектора. Предпочтительные антисмысловые рибозимы, в соответствии с настоящим изобретением, получают как описано у Sczakiel и соавт. (1995), причем специфические методы получения, на которые ссылаются в указанной статье, включены здесь путем ссылки. Трехспиральный метод Геномную ДНК CanIon можно также использовать для подавления экспрессии CanIon-гена на основе внутриклеточного трехспирального образования. Трехспиральные олигонуклеотиды используются для подавления транскрипции из генома. Они осо - 74 - 008252 бенно пригодны для изучения изменений клеточной активности при их ассоциации с конкретным геном. Аналогично, часть геномной ДНК CanIon можно использовать для изучения эффекта ингибирова-ния транскрипции CanIon в клетке. Традиционно гомопуриновые последовательности считают наиболее пригодными для трехспиральных методов. Но и гомопиримидиновые последовательности также могут подавлять экспрессию гена. Такие гомопиримидиновые олигонуклеотиды связываются с большой бороздкой в гомопурин: гомопиримидиновых последовательностях. Поэтому, в рамках настоящего изобретения рассматриваются оба типа последовательностей геномной ДНК CanIon. Для осуществления методов генной терапии с использованием трехспирального подхода, последовательности геномной ДНК CanIon прежде всего сканируют для того, чтобы идентифицировать 10-мерные - 20-мерные гомопиримидиновые или гомопуриновые участки, которые могли бы использоваться в методах, основанных на трех спиралях, для подавления экспрессии CanIon. После идентификации кандидатных гомопиримидиновых или гомопуриновых участков, их эффективность в подавлении экспрессии CanIon определяют путем введения в клетки тканевой культуры, в которой экспрессируется Can-Ion-ген, изменяющихся количеств олигонуклеотидов, содержащих последовательности-кандидаты. Данные олигонуклеотиды можно вводить в клетки с использованием способов, известных специалистам в данной области техники, включающих, но не ограниченных ими, кальций-фосфатное осаждение, DEAE-Декстран, электропорацию, липосом-опосредованную трансфекцию или нативное поглощение. Обработанные клетки отслеживают по измененной клеточной функции или сниженной CanIon-экспрессии с использованием таких методов, как Нозерн-блоттинг, метод защиты от РНКазы или методы на основе ПЦР для определения уровней транскрипции CanIon-гена в клетках, которые были обработаны данным олигонуклеотидом. Олигонуклеотиды, которые оказываются эффективными в подавлении генной экспрессии в клетках тканевой культуры, можно затем ввести in vivo с использованием описанных выше методов антисмыслового подхода в дозе, рассчитанной на основе результатов in vitro, как описано для антисмыслового подхода. В некоторых вариантах осуществления настоящего изобретения природные (бета) аномеры олиго-нуклеотидных элементов можно заменить альфа-аномерами для придания данному олигонуклеотиду большей устойчивости к нуклеазам. Кроме того, интеркалирующий агент, такой как бромистый этидий и т.п., можно присоединить к 3'-концу альфа-олигонуклеотида, чтобы стабилизировать трехнитевую структуру. Информацию об образовании олигонуклеотидов, подходящих для трехнитевого образования, см. в работе Griffin и соавт. (1989), которая таким образом включена с помощью данной ссылки. Фармацевтические композиции и рецептурные составы Соединения, модулирующие CanIon С использованием описанных в данном описании способов можно идентифицировать соединения-агонисты или антагонисты Canlon, которые селективно модулируют активность Canlon in vitro и in vivo. Настоящее изобретение включает, таким образом, способы лечения шизофрении, биполярного расстройства или любые иные описанные в данном описании заболевания или состояния пациентов, включающие введение эффективного количества соединения, модулирующего CanIon. Предпочтительно, указанное соединение представляет собой селективно CanIon-модулирующее соединение. Соединения, идентифицируемые способом настоящего изобретения, включают, например, антитела, обладающие связывающей специфичностью для CanIon-полипептида человека. Предполагается, что гомологи CanIon могут быть пригодны для модулирования CanIon-опосредованной активности и связанного с ней физиологического состояния, ассоциированного с шизофренией или биполярным расстройством. Кроме того, вообще предполагается, что способы анализа настоящего изобретения, базирующиеся на роли CanIon в нарушении центральной нервной системы, могут быть пригодны для идентификации соединений, способных воздействовать на каскадный анализ настоящего изобретения. В предпочтительном варианте осуществления настоящего изобретения пациента, страдающего шизофренией или биполярным расстройством, лечат путем введения данному пациенту фармацевтической композиции, включающей терапевтически эффективное количество антагониста CanIon. Показания Несмотря на то, что CanIon связан с геномной областью, ассоциированной с шизофренией и биполярным расстройством, показания на использование CanIon могут включать различные нарушения центральной нервной системы. Предполагается, что нарушения нервной системы обладают комплексной генетической основой и часто характеризуются общими симптомами. В частности, как описано в данном описании, показания могут включать шизофрению и другие психические расстройства, социальные расстройства, аутизм, зависимость от химических веществ и алкоголизм, нарушения болевых ощущений, эпилепсию, задержку психического развития, и другие психические заболевания, включая когнитивную функцию, страх, нарушение функции принятия пищи, нарушение функции мотивации и расстройство личности, которые определяются в соответствии с четвертым изданием классификации Diagnosis and Statistical Manual of Mental Disorders (DSM-IV). Кроме того, много сердечно-сосудистых расстройств, такие как стенокардия, гипертензия и аритмии, можно также лечить с использованием модуляторов Can-Ion, предпочтительно антагонистов. - 75 - 008252 Фармацевтические рецептурные составы и способы введения Соединения, идентифицированные с использованием способов настоящего изобретения, можно вводить млекопитающему, в том числе больному человеку, отдельно или в виде фармацевтических композиций, в которых они смешаны с подходящими носителями или наполнителем(ями) в терапевтически эффективных дозах, для лечения или улучшения состояния больного шизофренией или биполярным расстройством и близкими расстройствами. Терапевтически эффективная доза относится еще и к такому количеству соединения, которое достаточно для улучшения симптомов, как определено описанными в данном описании способами. Предпочтительно, терапевтически эффективная дозировка применима для продолжительного периодического использования или введения. Методы подбора состава и введения соединений настоящей заявки можно найти в "Remington's Pharmaceutical Sciences", Mack Publishing Co., Easton, PA, последнее издание. Способы введения Подходящие способы введения включают пероральное, ректальное, трансмукозальное или инте-стинальное введение, парентеральную доставку, в том числе внутримышечные, подкожные, внутримоз-говые инъекции, а также интратекальные, прямые внутрижелудочковые, внутривенные, внутрибрюшин-ные, интраназальные или внутриглазные инъекции. Особенно пригодный способ введения соединения для лечения заболевания центральной нервной системы включает хирургическую имплантацию устройства для доставки соединения в течение продолжительного периода времени. В частности, рассматривается длительное высвобождение рецептурных составов заявляемых лекарственных средств. Композиция/Рецептурная смесь Фармацевтические композиции и лекарственные средства для применения в соответствии с настоящим изобретением формируют традиционным способом с использованием одного или нескольких физиологически приемлемых носителей, включая наполнители и вспомогательные вещества. Оптимальный рецептурный состав зависит от выбранного способа введения. Для инъекции, агенты настоящего изобретения можно ввести в водные растворы, предпочтительно физиологически совместимые буферы, такие как раствор Хэнкса, раствор Рингера или физиологический солевой раствор, такой как фосфатный или бикарбонатный буфер. Для трансмукозального введения в рецептурной смеси можно использовать смачивающие вещества, подходящие для прохождения через буфер. Такие смачивающие вещества в большинстве случаев известны в данной области техники. Фармацевтические препараты, которые можно использовать перорально, включают сделанные из желатины раздвижные капсулы, а также сделанные из желатины и пластификатора, такого как глицерин или сорбит, герметичные капсулы. Раздвижные капсулы могут содержать активные ингредиенты в смеси с наполнителями, такими как лактоза, связывающими веществами, такими как крахмалы, и/или лубри-кантами, такими как тальк или стеарат магния, и, необязательно, стабилизаторами. В мягких капсулах данные активные соединения могут находиться в растворенном состоянии, или быть суспендированными в подходящих жидкостях, таких как жирные масла, жидкий парафин или жидкие полиэтиленовые гликоли. Кроме того, можно добавить стабилизаторы. Все рецептурные смеси для перорального введения должны быть дозированы для такого введения. Для трансбуккального введения данные композиции можно создавать в виде таблеток или лепешек традиционным способом. Для введения путем ингаляции используемые соединения настоящего изобретения удобно доставлять в виде распыляемого аэрозоля, находящегося в герметичной упаковке или в аэрозольном аппарате, с использованием соответствующего газа-вытеснителя, например, диоксида углерода. В случае герметизированного аэрозоля дозированную единицу можно определить с помощью клапана, обеспечивающего получение любого контролируемого количества аэрозоля. Капсулы и ампулы, например, из желатины, для использования в ингаляторе или инсуффляторе, можно создать в виде рецептурной смеси, содержащей порошок данного соединения и соответствующую порошковую основу, такую как лактоза или крахмал. Можно создать составные соединения для парентерального введения путем инъекции, например, путем болюсной инъекции или непрерывного вливания. Рецептурные смеси для инъекции можно представить в виде дозированной формы, например, в ампулах или в виде упаковки лекарственного средства для многократного приема с добавлением консерванта. Данные композиции могут принимать такие формы, как суспензии, растворы или эмульсии в водных носителях, и могут содержать фармакологические агенты, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. Фармацевтические рецептурные смеси для парентерального введения включают водные растворы данных активных соединений в водорастворимой форме. Водные суспензии могут содержать вещества, которые увеличивают вязкость данной суспензии, такие как карбоксиметилцеллюлоза, сорбит или декст-ран. Факультативно указанная суспензия может также содержать соответствующие стабилизаторы или агенты, которые повышают растворимость данных соединений, обеспечивая получение высококонцентрированных растворов. Альтернативно, данный активный ингредиент может находиться в порошковой или лиофилизован-ной форме для связывания перед использованием с соответствующим растворителем, таким как стерильная апирогенная вода. - 76 - 008252 Кроме того, помимо описанных выше рецептурных смесей, данные соединения можно составить в виде препаратов-депо. Такие рецептурные смеси длительного действия можно вводить путем имплантации (например, подкожно или внутримышечно) или путем внутримышечной инъекции. Поэтому, например, данные соединения можно включить в препарат вместе с полимерным или гидрофобным веществом (например, в виде эмульсии в соответствующем масле) или с ионообменными смолами, или с малорастворимыми производными, например, в виде малорастворимой соли. Кроме того, данные соединения можно доставлять с использованием системы с замедленным высвобождением, такой как полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих данный терапевтический агент. Были созданы различные вещества с замедленным высвобождением, которые хорошо известны специалистам в данной области техники. Капсулы с замедленным высвобождением, в зависимости от их химической природы, могут высвобождать данные соединения в течение нескольких недель вплоть до 100 дней. В зависимости от химической природы и биологической стабильности терапевтического агента можно применять дополнительные методы для стабилизации белка. Фармацевтические композиции могут также включать соответствующую твердую или гелевую фазу-носитель или фазу-наполнитель. Примеры таких носителей или наполнителей включают, но не ограничиваются ими, карбонат кальция, фосфат кальция, различные сахара, крахмалы, производные целлюлозы, желатину и полимеры, такие как полиэтиленгликоли. Эффективное дозирование Фармацевтические композиции, пригодные для использования в настоящем изобретении, включают композиции, в которых активные ингредиенты содержатся в эффективном количестве для достижения их предполагаемой цели. В частности, терапевтически эффективное количество означает количество, эффективное для предотвращения развития или смягчения существующих симптомов подвергнутого лечению больного. Определение эффективных количеств находится в рамках возможностей специалиста в данной области техники, особенно в свете представленного в данном описании подробного описания. Для любого соединения, используемого в данном способе настоящего изобретения, терапевтически эффективную дозу можно исходно определить в клеточной культуре и дозу можно разработать для животных моделей. Такие сведения можно использовать для более точного определения пригодных доз для человека. Терапевтически эффективная доза имеет отношение к такому количеству данного соединения, которое приводит к ослаблению симптомов у больного. Токсичность и терапевтическую эффективность таких соединений можно определить с помощью стандартных фармацевтических процедур на клеточных культурах или экспериментальных животных, например, для определения LD50 (летальная доза для 50% тестируемой популяции) и ED50 (терапевтически эффективная доза для 50% тестируемой популяции). Дозовое соотношение между токсическим и терапевтическим эффектами соответствует терапевтическому индексу лекарства и может быть выражено в виде соотношения между LD50 и ED50. Соединения, которые проявляют высокие терапевтические индексы, предпочтительны. Данные, полученные в этих исследованиях клеточных культур и животных, можно использовать для определения диапазона дозирования применительно к человеку. Дозирование таких соединений находится предпочтительно в пределах диапазона циркулирующих концентраций, которые включают ED50, с небольшой или в отсутствие токсичности. Дозировка может варьировать в зависимости от вида используемой дозы и способа ее введения. Точный состав, способ введения и дозировку может выбрать каждый врач, обследующий состояние пациента (См., например, Fingl и соавт. 1975, в "Pharmacological Basis of Therapeutics", Ch.1). Варианты осуществления настоящего изобретения, связанные с применением компьютера Используемый в данном описании термин "коды нуклеиновых кислот настоящего изобретения" включают в себя нуклеотидные последовательности, включающие, в основном состоящие, состоящие из любого нижеследующего: а) непрерывного участка последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 3000, 4000, 5000 или 10000 нуклеотидов SEQ ID No 1-3; b) непрерывного участка последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 3000, 4000, 5000 или 6000 нуклеотидов SEQ ID No 4 или ее комплементов; с) непрерывного участка последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, где указанный непрерывный участок последовательности включает двуаллельный маркер, выбранный из группы, состоящей из А1-А17; d) непрерывного участка последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300 или 400 нуклеотидов SEQ ID No 6; е) непрерывного участка последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300 или 400 нуклеотидов SEQ ID No 6, где указанный непрерывный участок последовательности включает двуаллельный маркер А18; и f) нуклеотидной последовательности, комплементарной любой из предшествующих нуклеотид-ных последовательностей. Кроме того, "коды нуклеиновых кислот настоящего изобретения" включают нуклеотидные последовательности, гомологичные: а) непрерывному участку последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 3000, 4000, 5000 или 10000 нуклео - 77 - 008252 тидов SEQ ID No 1-3; b) непрерывному участку последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 500, 1000, 2000, 3000, 4000, 5000 или 6000 нуклеотидов SEQ ID No 4; с) непрерывному участку последовательности по меньшей мере из 12, 15, 18, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300 или 400 нуклеотидов SEQ ID No 6; и d) последовательностям, комплементарным любой предшествующей последовательности. Гомологичные последовательности относятся к последовательности, обладающей по меньшей мере 99, 98, 97, 96, 95, 90, 85, 80 или 75% гомологией с данными непрерывными участками последовательности. Гомологию можно определить с использованием любого описанного в данном описании способа, в том числе BLAST2N со значениями параметров по умолчанию или с любыми модифицированными параметрами. Гомологичные последовательности могут также включать РНК-последовательности, в коде нуклеиновых кислот которых настоящего изобретения уридины заменяют тимином. Следует иметь ввиду, что коды нуклеиновых кислот настоящего изобретения могут быть представлены традиционным односимвольным форматом (См. внутреннюю заднюю сторону книги Stryer, Lubert. Biochemistry, 3rd edition. W.H.Freeman & Co., New York) или любым иным форматом или кодом, который регистрирует идентичность нуклеотидов в последовательности. Используемый в данном описании термин "полипептидные коды настоящего изобретения" включает в себя полипептидные последовательности, включающие непрерывный участок последовательности, по меньшей мере, из 6, 8, 10, 12, 15, 20, 25, 30, 40, 50, 100, 150, 200, 300, 400, 500, 700, 1000, 1200, 1400, 1600 или 1700 аминокислот SEQ ID No 5. В предпочтительных вариантах осуществления настоящего изобретения указанный непрерывный участок последовательности включает, по меньшей мере, 1, 2, 3, 5 или 10 аминокислотных позиций 277, 338, 574, 678, 680, 683, 691, 692, 695, 696, 697, 894, 1480, 1481, 1483, 1484, 1485, 1630, 1631, 1632, 1636, 1660, 1667, 1707, 1709 SEQ ID No 5. Следует иметь ввиду, что полипептидные коды настоящего изобретения могут быть представлены в традиционном односимвольном формате или трехбуквенном формате (См. внутреннюю заднюю сторону книги Stryer, Lubert. Biochemistry, 3rd edition. W.H. Freeman & Co., New York) или в любом ином формате или коде, который регистрирует идентичность полипептидов в последовательности. Для специалистов в данной области техники является понятным, что коды нуклеиновых кислот настоящего изобретения и полипептидные коды настоящего изобретения могут сохраняться, регистрироваться и обрабатываться на любом носителе, который может быть прочитан с помощью компьютера и предоставлен ему. Используемые в данном описании термины "регистрировать" и "сохранять" относятся к способу сохранения информации на компьютерном носителе. Специалистам в данной области техники легко усвоить любой из ныне известных способов записи информации на компьютерном носителе в пригодном для чтения виде, созданном производителями, включающим один или несколько кодов нуклеиновых кислот настоящего изобретения или один или несколько полипептидных кодов настоящего изобретения. Другой аспект настоящего изобретения заключается в том, что компьютерный носитель, пригодный для чтения, обладает записью на нем, по меньшей мере, 2, 5, 10, 15, 20, 25, 30 или 50 кодов нуклеиновых кислот настоящего изобретения. Другой аспект настоящего изобретения заключается в том, что компьютерный носитель, пригодный для чтения, обладает записью на нем, по меньшей мере, 2, 5, 10, 15, 20, 25, 30 или 50 полипептидных кодов настоящего изобретения. Компьютерный носитель, пригодный для чтения, включает магнитный носитель, пригодный для чтения, оптический носитель, электронный носитель, пригодный для чтения, и магнитнооптический носитель. Например, компьютерный носитель, пригодный для чтения, может быть представлен жестким диском, дискетой, магнитной лентой, CD-ROM, цифровым универсальным диском (DVD), запоминающим устройством прямого доступа (RAM) или постоянными запоминающими устройствами (ROM), а также другими видами носителей, известных специалистам в данной области техники. Варианты осуществления настоящего изобретения включают системы, в частности компьютерные системы, которые сохраняют и контролируют описанную в данном описании информацию о последовательности. Один из примеров компьютерной системы 100 иллюстрируется в виде блока диаграмм на фиг. 2. Используемый в данном описании термин "компьютерная система" относится к компонентам технического обеспечения, а данные компонентов сохранения используются для анализа нуклеотидных последовательностей кодов нуклеиновых кислот настоящего изобретения или аминокислотных последовательностей полипептидных кодов настоящего изобретения. В одном из вариантов осуществления настоящего изобретения компьютерная система 100 представляет собой сервер Sun Enterprise 1000 (Sun Microsystems, Palo Alto, CA). Компьютерная система 100 предпочтительно включает процессор для обработки данных, а также доступа и контроля данных о последовательности. Процессор 105 может представлять собой любой из хорошо известных типов процессорных элементов, такой как Pentium III от Intel Corporation, или аналогичный процессор от Sun, Motorola, Compaq or International Buisness Machines. Предпочтительно, компьютерная система 100 представляет собой основную целевую систему, которая включает процессор 105 и один или несколько внутренних компонентов 110 хранения данных для сохранения данных и одно или несколько поисковых устройств для поиска данных, сохраняемых в компонентах хранения данных. Специалистам в данной области следует иметь ввиду, что годится любая из ныне существующих компьютерных систем. - 78 - 008252 В одном из конкретных вариантов осуществления настоящего изобретения компьютерная система 100 включает процессор 105, который соединен шиной, которая соединяется с основным запоминающим устройством 115 (предпочтительно представленным в виде RAM) и одним или несколькими внутренними устройствами 110 сохранения памяти, такими как накопитель на жестких дисках и/или иные компьютерные носители для чтения, обладающие записанными данными. В некоторых вариантах осуществления настоящего изобретения компьютерная система 100, кроме того, включает одно или несколько поисковых устройств 118 для чтения сохраненных данных на внутренних устройствах 110 сохранения данных. Поисковое устройство данных 118 может представлять собой, например, дисковод для дискеты, дисковод для компакт-диска, накопитель для магнитной ленты и т.д. В некоторых вариантах осуществления настоящего изобретения внутреннее устройство 110 для сохранения данных представляет собой съемный компьютерный носитель для чтения, такой как дискета, компакт-диск, магнитная лента и т. д., содержащий логическую схему управления и/или записанные данные. Компьютерная система 100 может преимущественно включать или программироваться с помощью соответствующего программного обеспечения для прочтения логической схемы управления и/или данных из компонента хранения данных, однажды введенных в устройство поиска данных. Компьютерная система 100 включает дисплей 120, который используется для выведения данных на экран пользователя компьютера. Следует также иметь в виду, что компьютерную систему 100 можно подсоединить к другим компьютерным системам 125а-с в сеть или в глобальную сеть для создания централизованного доступа в компьютерную систему 100. Программное обеспечение для доступа и обработки данных по нуклеотидным последовательностям кодов нуклеиновых кислот настоящего изобретения или по аминокислотным последовательностям полипептидных кодов настоящего изобретения (таких как инструменты поиска, инструменты сравнения и инструменты моделирования и т.п.) могут постоянно храниться в основной памяти 115 в период исполнения. В некоторых вариантах осуществления настоящего изобретения компьютерная система 100 может, кроме того, включать программу сравнения последовательности для сравнения указанных выше кодов нуклеиновых кислот настоящего изобретения или полипептидных кодов настоящего изобретения, сохраняемых на компьютерном носителе для прочтения, со стандартной нуклеотидной или полипептидной последовательностями, сохраняемыми на компьютерном носителе для прочтения. "Программа сравнения последовательности" относится к одной или нескольким программам, которые выполняются на компьютерной системе 100 для сравнения нуклеотидной или полипептидной последовательностей с другими нуклеотидными или полипептидными последовательностями и/или соединениями, включающими, но не ограничивающимися ими, пептиды, пептидомиметики и химические агенты, хранимые в средствах хранения данных. Например, программа сравнения последовательности может сравнить нуклеотидные последовательности кодов нуклеиновых кислот настоящего изобретения или аминокислотные последовательности полипептидных кодов настоящего изобретения, сохраняемые на компьютерном носителе для прочтения, со стандартными последовательностями, сохраняемыми в компьютерном носителе для прочтения, чтобы идентифицировать гомологи, мотивы, вовлеченные в биологическую функцию, или структурные мотивы. Разнообразные программы сравнения последовательности, идентифицированные в данном патентном описании, подразумевают в частности использование данного аспекта настоящего изобретения. На фиг. 3 представлена схема последовательности операций, иллюстрирующая один из вариантов процесса 200 для сравнения новой нуклеотидной или полипептидной последовательности с последовательностями базы данных с тем, чтобы определить уровень гомологии между данной новой последовательностью и последовательностями из базы данных. Последовательности из базы данных могут представлять собой собственную базу данных, сохраняемую в компьютерной системе 100, или общедоступную базу данных, такую как GENBANK, PIR OR SWISSPROT, которые доступны через Интернет. Процесс 200 начинают со стартового положения 201 и затем перемещают в положение 202, в котором сравниваемая новая последовательность хранится в памяти компьютерной системы 100. Как указано выше, память может быть представлена памятью любого типа, в том числе RAM или внутренним запоминающим устройством. Затем процесс 200 перемещается в положение 204, в котором последовательности базы данных открыты для анализа и сравнения. Процесс 200 затем перемещается в положение 206, в котором первая последовательность, хранимая в базе данных, прочитывается в память данного компьютера. Затем осуществляют сравнение в положении 210, чтобы определить является ли первая последовательность той же, что и вторая последовательность. Важно отметить, что данный шаг не ограничивается осуществлением точного сравнения между новой последовательностью и первой последовательностью базы данных. Хорошо известные способы для сравнения двух нуклеотидных или полипептидных последовательностей, даже если они не идентичны, известны специалистам в данной области техники. Например, в одну из последовательностей можно ввести пробелы для того, чтобы повысить уровень гомологии между двумя тестируемыми последовательностями. Параметры, которые контролируют ввод пробелов и иных особенностей в последовательность во время сравнения, обычно вводит пользователь данной компьютерной системы. После осуществления сравнения двух последовательностей в положении 210, определение произ - 79 - 008252 водят в положении 210, являются ли две данные последовательности одинаковыми. Понятно, что термин " одинаковые" не ограничивается абсолютно идентичными последовательностями. Последовательности, которые находятся в гомологичных параметрах, введенных пользователем, будут помечены в процессе 200 как "одинаковые". Если сделано определение, что данные две последовательности являются одинаковыми, процесс 200 перемещают в положение 214, в котором название последовательности из базы данных выводится пользователем на дисплей. Данное положение извещает пользователя о том, что последовательность с выведенным названием удовлетворяет введенной ограниченной гомологии. После того как наименование сохраняемой последовательности демонстрируется пользователю, процесс 200 перемещается в положение 218, в котором определяют, имеются ли еще в базе данных последовательности. Если в базе данных нет больше последовательностей, тогда процесс 200 завершают в конце положения 220. Однако, если в базе данных имеются еще последовательности, то процесс 200 перемещают в положение 224, где курсор передвигают к следующей последовательности базы данных так, что ее можно сравнить с новой последовательностью. Таким образом, новую последовательность выравнивают и сравнивают с каждой последовательностью базы данных. Следует отметить, что если определение выполнено в положении 212, в котором последовательности не является гомологами, тогда процесс 200 следует двигать сразу в положение 218 для того, чтобы определить, доступны ли другие последовательности базы данных для сравнения. Таким образом, один из аспектов настоящего изобретения представляет собой компьютерную систему, включающую процессор, устройство сохранения данных, обладающее сохраненным в нем кодом нуклеиновых кислот настоящего изобретения или полипептидным кодом настоящего изобретения, устройство для сохранения данных, обладающее восстановимо сохраненными стандартными нуклеотидны-ми последовательностями или полипептидными последовательностями, сравниваемыми с кодом нуклеиновых кислот настоящего изобретения или с полипептидным кодом настоящего изобретения, и программой сравнения последовательности для осуществления сравнения. Программа сравнения последовательности может показывать уровень гомологии между сравниваемыми последовательностями или идентифицировать мотивы, вовлеченные в биологическую функцию, а также структурные мотивы кода нуклеиновых кислот настоящего изобретения и полипептидных кодов настоящего изобретения, или она может идентифицировать структурные мотивы в последовательностях, которые сравниваются с данными кодами нуклеиновых кислот и с полипептидными кодами. В некоторых вариантах осуществления настоящего изобретения устройство сохранения данных может хранить в себе последовательности, по меньшей мере, из 2, 5, 10, 15, 20, 25, 30 или 50 кодов нуклеиновых кислот настоящего изобретения или полипептидных кодов настоящего изобретения. Другой аспект настоящего изобретения представляет собой способ определения уровня гомологии между кодом нуклеиновых кислот настоящего изобретения и стандартной нуклеотидной последовательностью, включающей стадии прочтения кода нуклеиновых кислот и стандартной нуклеотидной последовательности путем использования компьютерной программы, которая определяет уровень гомологии, и определяет гомологию между кодом нуклеиновых кислот и стандартной нуклеотидной последовательностью с помощью компьютерной программы. Компьютерная программа может представлять собой любую из числа компьютерных программ по определению уровней гомологии, включая те, которые перечислены в данном описании, включая BLAST2N со значениями параметров по умолчанию или с любыми модифицированными параметрами. Данный способ можно реализовать с использованием описанных выше компьютерных систем. Данный способ можно также осуществить путем прочтения 2, 5, 10, 15, 20, 25, 30 или 50 описанных выше кодов нуклеиновых кислот настоящего изобретения через использование данной компьютерной программы и определения гомологии между кодами нуклеиновых кислот и стандартными нуклеотидными последовательностями. На фиг. 4 представлена схема последовательности операций, иллюстрирующая один из вариантов осуществления процесса 250 в компьютере для определения гомологичности двух последовательностей. Процесс 250 начинают с запуска в положении 252, а затем перемещают в положение 254, в котором первую сравниваемую последовательность сохраняют в память. Затем и вторую сравниваемую последовательность сохраняют в память в положении 256. После этого процесс 250 перемещают в положение 260, в котором прочитывают первый символ первой последовательности, и перемещают в положение 262, в котором прочитывают первый символ второй последовательности. Следовало бы иметь в виду, что если данная последовательность представляет собой нуклеотидную последовательность, то, как правило, символ должен быть либо А, Т, С, G, либо U. Если же данная последовательность представляет собой белковую последовательность, то она должна находиться в однобуквенном аминокислотном коде с тем, чтобы можно было легко сравнить первую и последующие последовательности. Затем определяют тождественность двух символов в положении 264. Если они тождественны, то процесс 250 перемещают в положение 268, в котором прочитывают очередные символы в первой и второй последовательностях. Затем определяют тождество следующих символов. Если они тождественны, то процесс 250 продолжает этот цикл до двух символов, которые не тождественны. Если определят, что - 80 - 008252 эти следующие два символа не тождественны, процесс 250 перемещают в положение 274, чтобы еще определить наличие символов для прочтения в любой последовательности. Если символы для прочтения отсутствуют, то процесс 250 перемещают в положение 276, в котором уровень гомологии между первой и второй последовательностями выводится данному пользователю. Уровень гомологии определяют путем вычисления доли символов в последовательностях, которые были одинаковыми из общего числа символов для первой последовательности. Таким образом, если каждый символ первой 100 нуклеотидной последовательности совместить с каждым символом второй последовательности, то уровень гомологии составит 100%. Альтернативно, компьютерная программа может представлять собой компьютерную программу, которая сравнивает нуклеотидные последовательности кодов нуклеиновых кислот настоящего изобретения со стандартной последовательностью нуклеиновой кислоты по одной или нескольким позициям. Факультативно такая программа регистрирует длину и идентичность встроенных, делетированных или замещенных нуклеотидов по отношению к последовательности стандартного полинуклеотида, либо кода нуклеиновых кислот настоящего изобретения. В одном из вариантов осуществления настоящего изобретения компьютерная программа может представлять собой программу, которая определяет в нуклеотид-ных последовательностях кодов нуклеиновых кислот настоящего изобретения присутствие одного или нескольких полиморфизмов единственного нуклеотида (SNP) по отношению к стандартной нуклеотид-ной последовательности. Каждый полиморфизм единственного нуклеотида может включать замену, вставку или делецию единственного основания. Другим аспектом настоящего изобретения является способ определения уровня гомологии между полипептидным кодом настоящего изобретения и последовательностью стандартного полипептида, включающий стадии прочтения полипептидного кода настоящего изобретения и последовательности стандартного полипептида путем использования компьютерной программы, которая определяет уровни гомологии, и определения гомологии между полипептидным кодом и последовательностью стандартного полипептида, используя компьютерную программу. Соответственно, другой аспект настоящего изобретения представляет способ определения кода нуклеиновых кислот, отличающегося по одному или нескольким нуклеотидам от стандартной нуклео-тидной последовательности, включающий стадии прочтения кода данной нуклеиновой кислоты и стандартной нуклеотидной последовательности путем использования компьютерной программы, которая идентифицирует различия между последовательностями нуклеиновой кислоты, и идентификации различий между кодом данной нуклеиновой кислоты и данной стандартной нуклеотидной последовательностью с помощью компьютерной программы. В некоторых вариантах осуществления настоящего изобретения данная компьютерная программа представляет собой программу, которая идентифицирует полиморфизмы единственных нуклеотидов. Данный способ можно осуществить с помощью описанных выше компьютерных систем и способа, проиллюстрированного на фиг. 4. Данный способ можно также осуществить в результате прочтения, по меньшей мере, 2, 5, 10, 15, 20, 25, 30 или 50 кодов нуклеиновых кислот настоящего изобретения и стандартных нуклеотидных последовательностей, через использование данной компьютерной программы, и идентификации различий между кодами данной нуклеиновой кислоты и данными стандартными нуклеотидными последовательностями с помощью данной компьютерной программы. В других вариантах осуществления настоящего изобретения компьютерная система, кроме того, может включать идентификатор, идентифицирующий элементы в нуклеотидных последовательностях кодов нуклеиновых кислот настоящего изобретения или в аминокислотных последовательностях полипептидных кодов настоящего изобретения. "Идентификатор" относится к одной или нескольким программам, которые идентифицируют определенные элементы в описанных выше нуклеотидных последовательностях кодов нуклеиновых кислот настоящего изобретения или в аминокислотных последовательностях полипептидных кодов настоящего изобретения. В одном из вариантов осуществления настоящего изобретения идентификатор может включать программу, которая идентифицирует открытую рамку считывания в кДНК-кодах настоящего изобретения. На фиг. 5 представлена схема последовательности операций, иллюстрирующая один из вариантов идентификатора процесса 300 по выявлению присутствия элемента в последовательности. Процесс 300 начинается с начального положения 302 и затем перемещается в положение 304, в котором первая последовательность, проверенная по элементам, сохраняется в памяти 115 компьютерной системы 100. Затем процесс 300 перемещается в положение 306, в котором открываются элементы последовательности базы данных. Такая база данных должна включать список символов каждого элемента в соответствии с наименованием данного элемента. Например, названный элемент мог бы представлять собой "Кодон Инициации", а символом мог бы быть "ATG". Другой пример элемента мог бы именоваться "ТААТАА Box", а символ данного элемента был бы "ТААТАА". Пример такой базы данных выпускается University of Wisconsin Genetics Computer Group (www.gcg.com). После того, как базу данных элементов открыли в положении 306, процесс 300 перемещается в положение 308, в котором первый элемент прочитывается из данной базы данных. Сравнение символа первого элемента с первой последовательностью затем осуществляют в положении 310. Затем осуществляют определение в положении 316 по нахождению символа элемента в первой последовательности. Если - 81 - 008252 символ обнаруживается, тогда процесс 300 перемещается в положение 318, в котором наименование найденного элемента выводится на экран дисплея пользователя. Затем процесс 300 перемещается в положение 320, в котором определяют движущиеся элементы, существующие в данной базе данных. Если элементов больше нет, то процесс 300 завершают в конце положения 324. Однако, если в данной базе данных имеются еще элементы, то процесс 300 прочитывает элемент следующей последовательности в положении 326 и цикл возвращают в положение 310, в котором символ следующего элемента сравнивают с символом элемента первой последовательности. Следует отметить, что если символ элемента не обнаруживается в первой последовательности в положении 316, то процесс 300 перемещается сразу в положение 320 для того, чтобы определить существование других элементов в данной базе данных. В другом варианте осуществления настоящего изобретения идентификатор может включать программу молекулярного моделирования, которая определяет 3-мерную структуру полипептидов и кодов настоящего изобретения. В некоторых вариантах осуществления настоящего изобретения программа молекулярного моделирования идентифицирует последовательности-мишени, которые в большинстве своем совместимы с профилями, представляющими структурную конфигурацию остатков в известных трехмерных белковых структурах (см., например, патент США № 5436850). В другом методе известные трехмерные структуры белков данного семейства накладывают одну на другую, чтобы определить структурно консервативные области в этом семействе. Эта белок-моделирующая методика также использует известную трехмерную структуру гомологичного белка для аппроксимации структуры полипептидных кодов настоящего изобретения (см., например, патент США №5557535). Традиционные методы моделирования гомологии использовали, как обычно принято, для построения моделей протеаз или антител (Sowdhamini и соавт. (1997)). Сравнительные подходы могут также использоваться для построения трехмерных моделей белка, если представляющий интерес белок обладает последовательностью, плохо идентифицирующейся с матричными белками. В некоторых случаях белки свернуты в сходные трехмерные структуры, несмотря на то, что их последовательности характеризуются очень низкой идентичностью. Например, трехмерные структуры ряда спиральных цитокинов скручены в сходную трехмерную топологию, несмотря на слабую гомологию последовательностей. Недавно созданные способы поточной обработки данных в настоящее время делают возможной идентификацию паттернов вероятностной упаковки для ряда ситуаций, где структурная связанность между мишенью и матриц(ей)ами не выявляется на уровне последовательностей. В гибридных способах, в которых распознавание укладки осуществляется с использованием Multiple Sequence Threading (Поточная обработка данных для множества последовательностей), структурная эквивалентность выводится по данным поточной обработки с использованием программы метрической геометрии DRAGON для создания модели с низким разрешением, а полностью атомизированное отображение создают с использованием пакета молекулярного моделирования, такого как QUANTA. В соответствии с данным 3-стадийным методом матрицы-кандидаты сначала идентифицируют с использованием нового алгоритма распознавания упорядоченной укладки MST, который способен осуществлять одновременную поточную обработку множества совмещенных последовательностей в одну или несколько 3-D структур. На второй стадии структурные эквиваленты, полученные по выходным данным MST, преобразуют в ограничения промежутков между остатками и вводят в программу метрической геометрии DRAGON, вместе с вспомогательной информацией, полученной из прогноза о вторичной структуре. Данная программа объединяет ограничения для несмещенного способа и быстро создает большое число подтверждений модели с низким разрешением. На третьей стадии эти подтверждения модели с низким разрешением преобразуют в полностью атомизированные модели и подвергают энергетической минимизации с использованием пакета молекулярного моделирования QUANTA (см., например, Aszodi и соавт. (1997)). Результаты анализа молекулярного моделирования используют затем в методах по рациональному созданию лекарственного средства, чтобы идентифицировать агенты, которые модулируют активность полипептидных кодов настоящего изобретения. В соответствии с этим другой аспект настоящего изобретения представляет собой способ идентификации элемента в кодах нуклеиновых кислот настоящего изобретения или в полипептидных кодах настоящего изобретения, включающий прочтение кода(ов) или полипептидного кода(ов) нуклеиновых кислот через использование компьютерной программы, которая идентифицирует элементы в ней, и идентификацию элементов в коде(ах) нуклеиновых кислот или в полипептидном(ых) коде(ах) с помощью данной компьютерной программы. В одном из вариантов осуществления настоящего изобретения компьютерная программа включает компьютерную программу, которая идентифицирует открытые рамки считывания. В другом варианте осуществления настоящего изобретения компьютерная программа идентифицирует структурные мотивы полинуклеотидной последовательности. В дополнительном варианте осуществления настоящего изобретения данная компьютерная программа включает программу молекулярного моделирования. Данный способ можно осуществить путем прочтения отдельной последовательности или, по меньшей мере, 2, 5, 10, 15, 20, 25, 30 или 50 кодов нуклеиновых кислот настоящего изобретения, либо полипептидных кодов настоящего изобретения через использование данной компьютерной - 82 - 008252 программы и идентификации элементов в данных кодах нуклеиновых кислот или полипептидных кодах с помощью данной компьютерной программы. Коды нуклеиновых кислот настоящего изобретения или полипептидные коды настоящего изобретения можно сохранять и управлять разнообразными программами процессорных данных в разных форматах. Например, их можно сохранять в виде текста в файле обработки текста, таком как Microsoft-WORD или WORDPERFECT или в виде ASCII-файла для разнообразных программ базы данных, хорошо известных специалистам в данной области техники, таких как DB2, SYBASE или ORACLE. Кроме того, многие компьютерные программы и базы данных можно использовать в качестве программ сравнения последовательностей, идентификаторов или поставщиков стандартных нуклеотидных или полипептидных последовательностей для сравнения с кодами нуклеиновых кислот настоящего изобретения или с полипептидными кодами настоящего изобретения. Нижеследующий список не следует рассматривать как ограничение настоящего изобретения, но как руководство к программам и базам данных, которые пригодны для кодов нуклеиновых кислот настоящего изобретения или для полипептидных кодов настоящего изобретения. Программы и базы данных, которые можно использовать, включают, но не ограничиваются перечисленным: MacPattern (EMBL), Dis-coveryBase (Molecular Application Group), GeneMine (Molecular Application Group), Look (Molecular Application Group), MacLook (Molecular Application Group), BLAST и BLAST2 (NCBI), BLASTN и BLASTX (Altschul и соавт., 1990), FASTA (Pearson and Lipman, 1988), FASTDB (Brutlag и соавт., 1990), Catalyst (Molecular Simulations Inc.), Catalyst/SHAPE (Molecular Simulations Inc.), Cerius2.DBAccess (Molecular Simulations Inc.), HypoGen (Molecular Simulations Inc.), Insight II (Molecular Simulations Inc.), Discover (Molecular Simulations Inc.), CHARMm (Molecular Simulations Inc.), Felix (Molecular Simulations Inc.), Del-Phi (Molecular Simulations Inc.), QuanteMM (Molecular Simulations Inc.), Homology (Molecular Simulations Inc.), Modeler (Molecular Simulations Inc.), ISIS (Molecular Simulations Inc.), Quanta/Protein Design (Molecular Simulations Inc.), WebLab (Molecular Simulations Inc.), WebLab Diversity Explorer (Molecular Simulations Inc.), Gene Explorer (Molecular Simulations Inc.), SeqFold (Molecular Simulations Inc.), the EMBL/Swissprotein database, the MDL Available Chemicals Directory database, the MDL Drug Data Report data base, the Comprehensive Medicinal Chemistry database, Derwents's World Drug Index database, the BioByteMasterFile database, the Genbank database и Genseqn базы данных. Многие другие программы и базы данных будут очевидны специалистам в данной области техники с учетом настоящего описания. Мотивы, которые можно выявлять с использованием указанных выше программ, включают последовательности, кодирующие лейциновые застежки, мотивы спираль-поворот-спираль, сайт гликозилиро-вания, сайты убихитинизации, альфа-спирали и бета-конфигурации, сигнальные последовательности, кодирующие сигнальные пептиды, которые управляют секрецией кодируемых белков, последовательности, вовлеченные в регуляцию транскрипции, такие как гомеобоксы, кислотные участки, энзиматически активные сайты, связывающие субстрат сайты и ферментативно расщепляемые сайты. В данной заявке приводятся ссылки на различные публикации, патенты и опубликованные патентные заявки. Раскрытие данных публикаций, патентов и опубликованных описаний изобретений к патентам, на которые ссылаются в данной заявке, включены путем ссылки в представленном изложении для наиболее полного удовлетворительного описания области техники, к которой относится настоящее изобретение. Примеры Пример 1. Идентификация двуаллельных маркеров - экстрагирование ДНК. Доноры не связаны родством и здоровы. Они были представлены достаточным разнообразием для типичной Французской гетерогенной популяции. От 100 индивидов была получена ДНК и проверена на наличие двуаллельных маркеров. От каждого донора брали 30 мл периферической венозной крови с добавлением ЭДТА. Клетки (осадок) собирали центрифугированием в течение 10 мин при 2000 об./мин. Эритроциты лизировали с помощью лизирующего раствора (50 мл конечный объем: 10 мМ Трис рН 7,6; 5 мМ MgQ2; 10 мМ NaCl). Данный раствор центрифугировали (10 мин, 2000 об./мин) столько раз, сколько необходимо для удаления остаточных эритроцитов в супернатанте, после ресуспендирования клеточного осадка в данном ли-зирующем растворе. Осадок лейкоцитов лизировали в течение ночи при 42°С 3,7 мл лизирующего раствора, состоящего из 3 мл ТЕ 10-2 (Трис-НО 10 мМ, ЭДТА 2 мМ)/№0 0,4M; 200 мкл SDS 10%; 500 мкл K-протеиназы (2 мг К-протеиназы в ТЕ 10-2/NaCl 0,4М). Для экстрагирования белков добавляли 1 мл насыщенного NaCl (6 М) (1/3,5 v/v). После энергичного перемешивания данный раствор центрифугировали в течение 20 мин при 10000 об./мин. Для осаждения ДНК к полученному супернатанту приливали 2-3 объема 100% этанола и раствор центрифугировали в течение 30 мин при 2000 об./мин. Полученный раствор ДНК трижды промывали 70%-м этанолом для удаления солей и центрифугировали в течение 20 мин при 2000 об./мин. Полученный осадок сушили при 37°С и ресуспендировали в 1 мл ТЕ 10-1 или в 1 мл воды. Концентрацию ДНК определяли путем измерения ОП при 260 нм (1 единица ОП=50 мкг/мл ДНК). Чтобы определить присутствие белков в данном растворе ДНК, определяли отношение ОП 260/ОП - 83 - 008252 280. В последующих описанных ниже примерах использовали только препараты ДНК, обладающие ОП 260/ОП 280 между 1,8 и 2. Объединенную ДНК получали смешиванием эквивалентных количеств ДНК от каждого индивида. Пример 2. Идентификация двуаллельных маркеров: амплификация геномной ДНК с помощью ПЦР. Амплификацию образцов специфичных геномных последовательностей ДНК примера 1 осуществляли на объединенной ранее полученной ДНК. Кроме того, аналогичным образом амплифицировали 50 индивидуальных образцов. ПЦР-анализ осуществляли с использованием следующего протокола: Конечный объем 25 мкл ДНК 2 нг/мкл MgСl2 2 мМ dNTP (каждого) 200 мкМ Праймер (каждого) 2,9 нг/мкл Ampli Taq Gold ДНК-полимераза 0,05 единиц/мкл ПЦР-буфер (10х=0,1М ТрисНС! рН 8,3 0,5М Ка) 1х Каждую пару первых праймеров создавали с использованием информации о последовательности из описанного в данном описании CanIon-гена и OSP-программного обеспечения (Hillier & Green, 1991). Длина первой пары праймеров составляла около 20 нуклеотидов и обладала последовательностью, раскрытой в табл. 1 в колонке, помеченной PU и RP. Таблица 1 Ампликон Диапазон позиций ампликона в SEQ ID 1 Название праймера Диапазон позиций амплификационного праймера в SEQ ID No 1 Название праймера Комплементарный диапазон позиций амплификационного праймера в SEQ ID No 1 99-62626 12343 12810 12343 12363 12793 12810 99-62632 13814 14296 13814 13832 14279 14296 99-62633 24863 25396 24863 24881 25378 25396 99-62611 69198 69650 69198 69218 69632 69650 99-62605 73005 73483 73005 73022 73466 73483 99-62635 79808 80334 79808 79826 80314 80334 Ампликон Диапазон позиций ампликона в SEQ ID 2 Название праймера Диапазон позиций амплификационного праймера в SEQ ID No 2 Название праймера Комплементарный диапазон позиций амплификационного праймера в SEQ ID No 2 99-79335 51031 51559 51031 51051 51539 51559 99-79336 60925 61374 60925 60945 61354 61374 99-79338 80271 80720 80271 80290 80700 80720 99-79339 91037 91486 В10 91037 91056 91466 91486 99-79314 100285 100784 В11 100285 100305 100764 100784 99-79316 106568 107020 В12 106568 106585 С12 107000 107020 99-79322 165864 166401 В13 165864 165884 С13 166381 166401 99-79306 235713 236210 В14 235713 235732 С14 236190 236210 Ампликон Диапазон позиций ампликона в SEQ ID 3 Название праймера Диапазон позиций амплификационного праймера в SEQ ID No 3 Название праймера Комплементарный диапазон позиций амплификационного праймера в SEQ ID No 3 99-79310 31618 32100 В15 31618 31635 С15 32080 32100 99-79311 42324 42723 В16 42324 42344 С16 42704 42723 Ампликон Диапазон позиций ампликона в SEQ ID 6 Название праймера Диапазон позиций амплификационного праймера в SEQ ID No 6 Название праймера Комплементарный диапазон позиций амплификационного праймера в SEQ ID No 6 99-62617 В17 С17 435 453 Предпочтительно, праймеры содержали общий олигонуклеотидный хвост "левее" от специфичных оснований, намеченных для амплификации, который пригоден для секвенирования. Праймеры PU содержат следующую дополнительную PU 5'-последовательность: TGTAAAACGACGGCCAGT; праймеры RP содержат следующую RP 5'-последовательность: CAGGAAACAGCTATGACC. Праймер, содержащий дополнительную PU 5'-последовательность, приведен в SEQ ID No 7. Прай - 84 - 008252 мер, содержащий дополнительную RP 5'-последовательность, приведен в SEQ ID No 8. Синтез данных праймеров осуществляли с помощью фосфорамидитного способа в синтезаторе GENSET UFPS 24.1. Амплификацию ДНК осуществляли в амплификаторе Genius II. После нагревания до 95°С в течение 10 мин проводили 40 циклов. Каждый цикл включал: 30 с при 95°С, 54°С в течение 1 мин и 30 с при 72°С. Конечное удлинение, 10 мин 72°С, завершало амплификацию. Количества полученных продуктов амплификации определяли в 96-луночных планшетах для микротитрования с использованием флуоро-метра и Picogreen в качестве интеркалирующего агента (Molecular Probes). Пример 3. Идентификация двуаллельных маркеров - секвенирование амплифицированной геномной ДНК и идентификация полиморфизмов. Секвенирование амплифицированной ДНК, полученной в примере 2, осуществляли на секвенаторе ABI 377. Последовательности продуктов амплификации определяли с использованием автоматизированных реакций дидезокситерминирующего секвенирования согласно протоколу цикла секвенирования, терминированного красителем. Продукты реакций секвенирования разделяли в секвенирующих гелях и последовательности определяли с использованием анализа визуализацией геля (ABI Prism DNA Sequencing Analysis software (2.1.2 version)). Затем оценивали данные о последовательностях, чтобы в полученных амплифицированных фрагментах обнаружить присутствие двуаллельных маркеров. Поиск полиморфизма основан на присутствии наложенных пиков в электрофоретической картине, полученных для разных оснований, встреченных в одной и той же позиции, как описано раньше. В 17 амплифицированных фрагментах выявлено 18 двуаллельных маркеров. Локализация этих дву-аллельных маркеров показана в табл. 2. Таблица 2 Ампликон Название маркера Полиморфизм Позиция ВМ в SEQ ID Ал1 Ал2 Nol No 4 99-6226 99-62626-168 12642 99-62632 99-62632-275 14088 99-62633 99-6263-409 24981 99-62611 99-62611-51 69248 99-62605 99-62605-56 73428 99-62635 99-62635-443 80250 Ампликон Название маркера Полиморфизм Позиция ВМ в SEQ ID Ал1 Ал2 No2 No4 99-79335 99-79335-60 51090 99-79336 99-79336-369 61293 99-79338 99-79338-332 80602 99-79314 А10 99-79314-201 100845 99-79314 99-79314-225 100509 99-79316 А12 99-79316-158 106725 1658 99-7932 А13 99-79322-224 166087 99-79322 А14 99-79322-473 166336 99-79306 А15 99-79306-182 235894 Ампликон Название маркера Полиморфизм Позиция ВМ в SEQ ID Ал1 Ал2 No3 No 4 99-79310 А16 99-79310-29 31646 4481 99-79311 А17 99-79311-50 42373 Ампликон Название маркера Полиморфизм Позиция ВМ в SEQ ID Ал1 Ал2 No 6 99-62617 А18 99-62617-105 105 - 85 - 008252 ВМ относится к "двуаллельному маркеру". Ал1 и ал2 относятся, соответственно, к аллелю 1 и алле-лю 2 данного двуаллельного маркера. Таблица 3 Название маркера Диапазон позиций Зондов в SEQ ID No 1 Зонды 99-62626-168 12630 12654 99-62632-275 14076 14100 99-62633-409 24969 24993 99-62611-51 69236 69260 99-62605-56 73416 73440 99-62635 80238 80262 Название маркера Диапазон позиций Зондов в SEQ ID No 2 Зонды 99-79335-60 51078 51102 99-79336-369 61281 61305 99-79338-332 80590 80614 А10 99-79314-201 100473 100497 Р10 99-79314-225 100497 100521 Р11 А12 99-79316-158 106713 106737 Р12 А13 99-79322-224 166075 166099 Р13 А14 99-79322-473 166324 166348 Р14 А15 99-79306-182 235882 235906 Р15 Название маркера Диапазон позиций Зондов в SEQ ID No 3 Зонды А16 99-79310-29 31634 31658 Р16 А17 99-79311-50 42361 42385 Р17 Название маркера Диапазон позиций Зондов в SEQ ID No 6 Зонды А18 99-62617-105 117 Р18 Пример 4. Проверка достоверности полиморфизма с помощью микросеквенирования. Двуаллельные маркеры, идентифицированные в примере 3, в последующем получали подтверждение, а их соответствующие частоты были определены с помощью микросеквенирования. Микросеквенирование осуществляли для каждого индивидуального образца ДНК, описанного в примере 1. Амплификацию геномной ДНК индивидов осуществляли с помощью ПЦР, как описано выше, для выявления двуаллельных маркеров с одним и тем же набором ПЦР-праймеров (табл. 1). Предпочтительные праймеры, используемые для микросеквенирования, составляли в длину около 19 нуклеотидов и гибридизовались непосредственно слева от рассматриваемого полиморфного основания. В соответствии с настоящим изобретением, данные праймеры, используемые для микросеквениро-вания, подробно представлены в табл. 4. - 86 - 008252 Таблица 4 Наименование маркера Двуалле льный маркер Mis. 1 Диапазон позиций микросеквенирующего праймера mis 1 в SEQ ID No 1 Mis. 2 Диапазон комплементарных позиций микросеквенирующего праймера mis. 2 в SEQ ID No 1 99-62626-168 12623 12641 12643 12661 99-62632 14069 14087 12642 12661 99-62633-409 24962 24980 24982 25000 99-62611-51 69229 69247 69249 69267 99-62605-56 73409 73427 73429 73447 99-62635-443 80231 80249 80251 80269 Наименование маркера Двуалле льный маркер Mis. 1 Диапазон позиций микросеквенирующего праймера mis 1 в SEQ ID No 1 Mis. 2 Диапазон комплементарных позиций микросеквенирующего праймера mis. 2 в SEQ ID No 2 99-79335-60 51071 51089 51091 51109 99-79336-369 61274 61292 61294 61312 99-79338-332 80583 80601 80603 80621 99-79314-201 А10 D10 100466 100484 100486 100504 99-79314-225 Dll 100490 100508 Ell 100510 100528 99-79316-158 А12 D12 106706 106724 E12 106726 106744 99-79322-224 А13 D13 166068 166086 E13 166088 166106 99-79322-473 А14 D14 166317 166335 E14 166337 166355 99-79306-182 А15 D15 235875 235893 E15 235895 235913 Наименование маркера Двуалле льный маркер Mis. 1 Диапазон позиций микросеквенирующего праймера mis 1 в SEQ ID No 3 Mis. 2 Диапазон комплементарных позиций микросеквенирующего праймера mis. 2 в SEQ ID No 3 99-79310-29 А16 D16 31627 31645 E16 31647 31665 99-79311-50 А17 D17 42354 42372 E17 42374 42392 Наименование маркера Двуалле льный маркер Mis. 1 Диапазон позиций микросеквенирующего праймера mis 1 в SEQ ID No 6 Mis. 2 Диапазон позиций микросеквенирующего праймера mis. 2 в SEQ ID No 6 99-62617-105 А18 D18 104 E18 106 124 Mis 1 и Mis 2 относятся, соответственно, к микросеквенирующим праймерам, которые гибридизу-ются с некодирующей цепью CanIon-гена или с кодирующей цепью CanIon-гена. Реакцию микросеквенирования осуществляли следующим образом. После выделения очисткой продуктов амплификации реакционную микросеквенирующую смесь получали путем добавления, в конечном объеме 20 мкл: 10 пмолей микросеквенирующего олигонуклео-тида, 1 Ед. Термосеквеназы (Amersham E79000G), 1,25 мкл термосеквеназного буфера (260 мМ Трис НС1 рН 9,5, 65 мМ MgСl2) и два соответствующих ddNTP (Perkin Elmer, Dye Terminator Set 401095), комплементарных нуклеотидам по полиморфному сайту каждого тестируемого двуаллельного маркера, следуя рекомендациям производителя. Через 4 мин при 94°С в амплификаторе Tetrad РТС-225 (MJ Research) осуществляли 20 ПЦР-циклов из 15 с при 55°С, 5 с при 72°С и 10 с при 94°С. Затем с помощью этаноль-ного осаждения удаляли невключенный терминирующий краситель. Образцы окончательно ресуспенди-ровали в формамид-ЭДТА-буфере для насыщения и нагревали в течение 2 мин при 95°С перед загрузкой в полиакриламидный секвенирующий гель. Данные получали с помощью ДНК-секвенатора ABI PRISM 377 и обрабатывали с использованием программного обеспечения CanIonSCAN (Perkin Elmer). После гелевого анализа данные автоматизированно обрабатывали с помощью программного обеспечения, которое позволяет определять аллели двуаллельного маркера, представленные в каждом ам-плифицированном фрагменте. Данная программа оценивает такие факторы, как насколько интенсивность сигналов, получаемых в осуществленных выше операциях микросеквенирования, является слабой, нормальной или интенсивной или же данные сигналы являются неоднозначными. Кроме того, данная программа идентифицирует существенные пики (по форме и высоте). В числе существенных пиков, пики, соответствующие сайту-мишени, идентифицировали по их положению. Если два существенных пика обнаруживаются в одном и том же положении, каждый образец относят к категории гомозиготного или гетерозиготного типа на ос - 87 - 008252 нове соотношения высоты пика. Пример 5. Получение антительных композиций к CanIon-белку. Из трансфицированных или трансформированных клеток, содержащих экспрессионный вектор, кодирующий CanIon-белок или его часть, выделяют практически чистый белок или полипептид. Концентрацию белка конечного препарата доводят, например, путем концентрирования на устройстве с Amicon-фильтром, до уровня нескольких микрограмм/мл. Затем моноклональные или поликлональные антитела к данному белку можно получить следующим образом. A. Получение моноклональных антител путем гибридомного слияния. Моноклональные антитела к эпитопам CanIon-белка или к его части можно получить из мышиных гибридом в соответствии с классическим способом Kohler, G. and Milstein, С. (1975) или производными от него способами. См. также Harlow, E. and D. Lane, 1988. Вкратце, мышь периодически инокулируют несколькими микрограммами CanIon-белка или его части на протяжении нескольких недель. Затем мышь умерщвляют и выделяют клетки селезенки, продуцирующие антитела. Выделенные клетки селезенки сливают с помощью полиэтиленгликоля с мышиными миеломными клетками, а излишек неслившихся клеток разрушают выращиванием на селективной среде, включающей аминоптерин (НАТ-среда). Успешно слившиеся клетки разбавляют и аликвоты разбавления помещают в лунки планшета для микротитрования, где данная культура продолжает расти. Ан-тителопродуцирующие клоны идентифицируют путем выявления антител в супернатантной жидкости лунок с помощью методов иммунологического анализа, таких как ELISA, впервые описанного Engvall (1980), и его производного способа. Отобранные позитивные клоны можно увеличить в объеме и собрать для использования их продукт в виде моноклональных антител. Подробные методы получения монокло-нальных антител описаны у Davis L. и соавт. Basic Methods in Molecular Biology (Основные методы молекулярной биологии), Elsevier, New York, Section 21-2. B. Получение поликлональных антител с помощью иммунизации. Поликлональную антисыворотку, содержащую антитела к гетерогенным эпитопам CanIon-белка или его части, можно получить путем иммунизации животного, не принадлежащего человеческому роду, CanIon-белком или его частью, который может быть немодифицированным или модифицированным для усиления иммуногенности. Подходящим животным, не принадлежащим человеческому роду, предпочтительно является выбранное млекопитающее, обычно, мышь, крыса, кролик, коза или лошадь. Альтернативно, неочищенный препарат, который обогащали по концентрации CanIon, можно использовать для образования антител. Такие белки, фрагменты или препараты вводят не принадлежащему человеческому роду млекопитающему в присутствии соответствующего адъюванта (например, гидроксида алюминия, RIBI и т. п.), который известен в данной области техники. Кроме того, белок, фрагмент или препарат может быть предварительно обработан агентом, который повышает антигенность, такие агенты известны в данной области техники и включают, например, метилированный бычий сывороточный альбумин (mBSA), бычий сывороточный альбумин (BSA), поверхностный антиген гепатита В и гемоцианин улитки фиссурелии (KLH). Собирают сыворотку иммунизированного животного, обрабатывают и тестируют в соответствии с известными методами. Если собранная сыворотка содержит поликлональные антитела с нежелательными эпитопами, тогда поликлональные антитела можно выделить очисткой с помощью им-муноаффинной хроматографии. На получение эффективных поликлональных антител влияют многие факторы, связанные с антигеном и видовым статусом хозяина. Кроме того, животные-хозяева по-разному отвечают на место инокуляции и дозу, причем несоответствующие или чрезмерные дозы антигена приводят к снижению титра антисыворотки. Небольшие дозы (на уровне нг) вводимого антигена во многие интрадермальные места оказываются более надежными. Методы получения и обработки поликлональной антисыворотки известны в данной области техники, см. например, Mayer and Walker (1987). Протокол эффективной иммунизации кроликов можно найти у Vaitukaitis J. и соавт. (1971). Повторные инъекции можно осуществлять с регулярными интервалами и собирать антисыворотку, когда титр ее антител, определяемый полуколичественно, например, с помощью двойной иммунодифузии в агаре против известных концентраций данного антигена, начинает падать. См., например, Ouchterlony О. и соавт. (1973). Плато кривой концентрации антител обычно находится в пределах от 0,1 до 0,2 мг/мл сыворотки (около 12 мкМ). Сродство антисыворотки по антигену определяют путем получения конкурентных кривых связывания, как описано, например, Fisher D. (1980). Препараты антител, полученные в соответствии с моноклональным или поликлональным протоколом, пригодны для количественного иммунологического анализа, в котором определяют концентрации веществ, несущих антиген, в биологических образцах; их также используют полуколичественно или количественно для идентификации наличия антигена в биологическом образце. Данные антитела могут также использоваться в терапевтических композициях для уничтожения клеток, экспрессирующих данный белок или для снижения уровня данного белка в данном организме. Хотя предпочтительные варианты осуществления настоящего изобретения были проиллюстрированы и описаны, следует иметь ввиду, что специалисты в данной области техники могут внести в него различные изменения, не выходя за рамки существа и объема настоящего изобретения. - 88 - 008252 Ссылки Abbondanzo S.J. et al., 1993, Methods in Enzymology, Academic Press, New York, pp. 803-823. Ajioka R.S. et al., Am. J. Hum. Genet., 60:1439-1447, 1997. Altschul et al., 1990, J. Mol. Biol. 215 (3):403-410. Altschul et al., 1993, Nature Genetics 3:266-272. Altschul et al., 1997, Nuc. Acids Res. 25:3389-3402. Ames R.S. et al. J. Immunol. Methods 184:177-186 (1995). Anderson et al., Symp. Soc. Exp. Biol. 48: 85-97 (1994). Anton M. et al., 1995, J. Virol., 69 : 4600-4606. Araki K. et al. (1995) Proc. Natl. Acad. Sci. USA. 92(1): 160-4. Ashkenazi, A. et al. PNAS 88:10535-10539 (1991). Aszodi et al., Proteins:Structure, Function, and Genetics, Supplement 1:38-42 (1997). Ausubel et al. (1989) Current Protocols in Molecular Biology, Green Publishing Associates and Wiley In-terscience, N.Y. Bartunek P. et al. Cytokine 8(l):14-20 (1996). Baubonis W. (1993) Nucleic Acids Res. 21(9):2025-9. Beaucage et al., Tetrahedron Lett 1981,22: 1859-1862. Better, M. et al. Science 240:1041-1043 (1988). Blouin J.L. et al. Nature Genetics, 20: 70-73 (1998). Bradley A., 1987, Production and analysis of chimaeric mice. In: E.J. Robertson (Ed.), Teratocarcinomas and embryonic stem cells: A practical approach. IRL Press, Oxford, pp.113. Bram R.J. et al., 1993, Mol. Cell Biol., 13 : 4760-4769. Brinkman U. et al. J. Immunol. Methods 182:41-50 (1995). Brown E.L., Belagaje R., Ryan M.J., Khorana HG, Methods Enzymol. 1979; 68:109-151. Brutlag et al. Согпр. Арр. Biosci. 6:237-245, 1990. Brzustowicz et al., Am. J. Hum. Genet. 65:1096-1103 (1999). Bulman et al., Hum. Mol. Gen. 6(10) 1679-1685 (1997). Burton D.R. et al. Advances in Immunology 57:191-280 (1994). Bush et al., 1997, J. Chromatogr., 777 : 311-328. Carlson, N.G. et al. J. Biol. Chem. 272(17): 11295-11301 (1997). Chai H. et al. (1993) Biotechnol. Appl. Biochem. 18:259-273. Chee et al. (1996) Science. 274:610-614. Chen and Kwok, Nucleic Acids Research 25:347-353, 1997. Chen et al. (1987) Mol. Cell. Biol. 7:2745-2752. Chen et al. Proc. Natl. Acad. Sci. USA 94/20 10756-10761, 1997. Chen, Z. et al. Cancer Res. 58(16): 3668-3678 (1998). Cho R.J. et al., 1998, Proc. Natl. Acad. Sci. USA, 95(7):3752-3757. Chou J.Y., 1989, Mol. Endocrinol., 3: 1511-1514. Clark A.G. (1990) Mol. Biol. Evol. 7:111-122. Coles R., Caswell R., Rubinsztein D.C., Hum. Mol. Genet. 1998; 7:791-800. Compton J. (1991) Nature. 350(6313):91-92. Davis L.G., M.D. Dibner, and J.F. Battey, Basic Methods in Molecular Biology, ed., Elsevier Press, NY, 1986. Dempster et al., (1977) J. R. Stat. Soc., 39B:1-38. Deng B. et al. Blood 92(6): 1981-1988 (1998). Dent D.S. & Latchman D.S. (1993) The DNA mobility shift assay. In: Transcription Factors: A Practical Approach (Latchman D.S., ed.) pp.1-26. Oxford: IRL Press. Denyer et al., Drug Disc. Today 3(7): 323-332 (1998). Eckner R. et al. (1991) EMBO J. 10:3513-3522. Edwards et Leatherbarrow, Analytical Biochemistry, 246, 1-6 (1997). Engvall, E., Meth. Enzymol. 70:419 (1980). Excoffier L. and Slatkin M. (1995) Mol. Biol. Evol., 12(5): 921-927. Feldman and Steg, 1996, Medecine/Sciences, synthese, 12:47-55. Felici F., 1991, J. Mol. Biol., Vol. 222:301-310. Fell, H.P. et al. J. Immunol. 146:2446-2452 (1991). Fields and Song, 1989, Nature, 340 : 245-246. Fisher, D., Chap. 42 in: Manual of Clinical Immunology, 2d Ed. (Rose and Friedman, Eds.) Amer. Soc. For Microbiol., Washington, D.C. (1980). Flotte et al. (1992) J. Respir. Cell Mol. Biol. 7:349-356. Fodor et al. (1991) Science 251:767-777. Fraley et al. (1979) Proc. Natl. Acad. Sci. USA. 76:3348-3352. Fried M., Crothers D.M., Nucleic Acids Res. 1981; 9:6505-6525. - 89 - 008252 Fromont-Racine M. et al., 1997, Nature Genetics, 16(3): 277-282. Fuller S.A. et al. (1996) Immunology in Current Protocols in Molecular Biology, Ausubel et al. Eds, John Wiley & Sons, Inc., USA. Furth P.A. et al. (1994) Proc. Natl. Acad..Sci. USA. 91:9302-9306. Garner M.M., Revzin A., Nucleic Acids Res. 1981; 9:3047-3060. Geysen H. Mario et al. 1984. Proc. Natl. Acad. Sci. U.S.A. 81:3998-4002. Ghosh and Bacchawat, 1991, Targeting of liposomes to hepatocytes, IN: Liver Diseases, Targeted diagnosis and therapy-using specific receptors and ligands. Wu et al. Eds., Marcel Dekeker, New York, pp. 87-104. Gillies, S.D. et al. J. Immunol. Methods 125:191-202 (1989). Gillies, S.O. et al. PNAS 89:1428-1432 (1992). Gonnet et al., 1992, Science 256:1443-1445. Gonzalez et al., Drug Disc. Today 4(9): 431:439 (1999). Gopal (1985)Afol. Cell. Biol., 5:1188-1190. Gossen M. et al. (1992) Proc. Natl. Acad. Sci. USA. 89:5547-5551. Gossen M. et al. (1995) Science. 268:1766-1769. Graham et al. (1973) Virology 52:456-457. Green et al., Ann. Rev. Biochem. 55:569-597 (1986). Greenspan and Bona, FASEB J. 7(5):437-444 (1989). Griffin et al. Science 245:967-971 (1989). Grompe, M. (1993) Nature Genetics. 5:111-117. Grompe, M. et al. (1989) Proc. Natl. Acad. Sci. U.S.A. 86:5855-5892. Gu H. et al. (1993) Cell 73:1155-1164. Gu H. et al. (1994) Science 265:103-106. Guatelli J.C et al. Proc. Natl. Acad. Sci. USA. 35:273-286. Hacia J.G., Brody L.C., Chee M.S., Fodor S.P., Collins F.S., Nat. Genet. 1996; 14(4):441-447. Hall L.A. and Smirnov I.P. (1997) Genome Research, 7:378-388. Hames B.D. and Higgins S.J. (1985) Nucleic Acid Hybridization: A Practical Approach. Hames and Hamill, Pfugers Arch. 391, 85-100 (1981). Hammerling, et al., in: MONOCLONAL ANTIBODIES AND T-CELL HYBRIDOMAS 563-681 (Elsevier, N.Y., 1981). Harju L., Weber T., Alexandrova L., Lukin M., Ranki M., Jalanko A., Clin. Chem. 1993; 39(1 IPt 1):2282- 2287. Harland et al. (1985) J. Cell. Biol. 101:1094-1095. Harlow, E. and D. Lane. 1988. Antibodies A Laboratory Manual. Cold Spring Harbor Laboratory, pp. 53242. Harper J.W. et al., 1993, Cell, 75 : 805-816. Harrop J.A. et al. J. Immunol. 161(4): 1786-1794 (1998). Hawley M.E. et al. (1994) Am. J. Phys. Anthropol. 18:104. Hell et al., Annals N.Y. Acad. Sci. 747:282-293 (1994). HenikoffandHenikoff, 1993, Proteins 17:49-61. Higgins et al., 1996, Methods Enzymol. 266:383-402. Higgins Ed., IRL Press, Oxford. Hillier L. and Green P. Methods Appl., 1991, 1: 124-8. Hoess et al. (1986) Nucleic Acids Res. 14:2287-2300. Huang L. et al. (1996) Cancer Res. 56(5):1137-1141. Huston et al. Methods in Enzymology 203:46-88 (1991). Huygen et al. (1996) Nature Medicine. 2(8):893-898. Izant J.G., Weintraub H., Cell 1984 Apr; 36(4):1007-15. Julan et al. (1992) J. Gen. Virol. 73:3251-3255. Kanegae Y. et al., Nucl. Acids Res. 23:3816-3821 (1995). Karlin and Altschul, 1990, Proc. Natl. Acad. Sci. USA 87:2267-2268. Kettleborough, C.A. et al. Eur. J. Immunol. 24:952-958 (1994). Khoury J. et al., Fundamentals of Genetic Epidemiology, Oxford University Press, NY, 1993. Kim U-J. et al. (1996) Genomics 34:213-218. Klein et al. (1987) Nature. 327:70-73. Koch et al., J. Biol. Chem. 265 (29): 17786-17791 (1990). Kohler, G. and Milstein, C., Nature 256:495 (1975). Koller et al. (1992) Annu. Rev. Immunol. 10:705-730. Kozal M.J., Shah N., Shen N., Yang R., Fucini R., Merigan T.C., Richman D.D., Morris D., Hubbell E., Chee M., Gingeras T.R., Nat. Med. 1996; 2(7):753-759. Lander and Schork, Science, 265, 2037-2048, 1994. Landegren U. et al. (1998) Genome Research, 8:769-776. - 90 - 008252 Lange K. (1997) Mathematical and Statistical Methods for Genetic Analysis. Springer, New York. Lee et al., FEBS Lett. 445 231:236 (1999). Lenhard T. et al. (1996) Gene. 169:187-190. Liautard, J. et al. Cytokinde 9 (4):233-241 (1997). Lin M.W. et al. Hum. Genet., 99(3): 417-420 (1997). Linton M.F. et al. (1993) J. Clin. Invest. 92:3029-3037. Liu Z. et al. (1994) Proc. Natl. Acad. Sci. USA. 91:4528-4262. Livak et al., Nature Genetics, 9:341-342, 1995. Livak K.J., Hainer J.W., Hum. Mutat. 1994; 3(4):379-385. Lockhart et al. Nature Biotechnology 14: 1675-1680, 1996. Lucas A.H., 1994, In : Development and Clinical Uses of Haempophilus b Conjugate. Manger et al., Anal. Biochem. 214: 190-194 (1993). Mansour S.L. et al. (1988) Nature. 336:348-352. Marshall R.L. et al. (1994) PCR Methods and Applications. 4:80-84. McCormick et al. (1994) Genet. Anal. Tech. Appl. 11:158-164. McLaughlin B.A. et al. (1996) Am. J. Hum. Genet. 59:561-569. Morena et al., Annals NY Acad. Sci. 102-117 (1999). Mori, et al., Nature 350: 398-402 (1991). Morrison, Science 229:1202 (1985). Morton N.E., Am. J. Hum. Genet., 7:277-318, 1955. Muller, Y.A. et al. Structure 6(9): 1153-1167 (1998). Mullinax, R.L. et al. BioTechniques 12 (6) :864-869 (1992). Muzyczka et al. (1992) Curr. Topics in Micro, and Immunol. 158:97-129. Nada S. et al. (1993) Cell 73:1125-1135. Nagy A. et al., 1993, Proc. Natl. Acad. Sci. USA, 90: 8424-8428. Naramura, M. et al. Immunol. Lett. 39:91-99 (1994). Narang S.A., Hsiung H.M., Brousseau R., Methods Enzymol. 1979; 68:90-98. Neda et al. (1991) J. Biol. Chem. 266:14143-14146. Newton et al. (1989) Nucleic Acids Res. 17:2503-2516. Nickerson D.A. et al. (1990) Proc. Natl. Acad. Sci. U.S.A. 87:8923-8927. Nicolau С. et al., 1987, Methods Enzymol., 149:157-76. Nicolau et al. (1982) Biochim. Biophys. Acta. 721:185-190. Nissinoff, J. Immunol. 147(8): 2429-2438 (1991). Nyren P., Pettersson B., Uhlen M., Anal. Biochem. 1993; 208(1): 171-175. O'Reilly et al. (1992) Baculovirus Expression Vectors: A Laboratory Manual. W.H. Freeman and Co., New York. Ohno et al. (1994) Science. 265:781-784. Oi et al., BioTechniques 4:214 (1986). Oldenburg K.R. et al., 1992, Proc. Natl. Acad. Sci., 89:5393-5397. Olesen et al., Voltage gated ion channel modulators, 7-8 December, Philadelphia PA, USA (1995). Orita et al. (1989) Proc. Natl. Acad. Sci. U.S.A. 86: 2776-2770. Ott J., Analysis of Human Genetic Linkage, John Hopkins University Press, Baltimore, 1991. Ouchterlony, O. et al., Chap. 19 in: Handbook of Experimental Immunology D. Wier (ed) Blackwell (1973). Padlan E.A., Molecular Immunology 28(4/5): 489-498 (1991). Parmley and Smith, Gene, 1988, 73:305-318. Pastinen et al., Genome Research 1997; 7:606-614. Pearson and Lipman, 1988, Proc. Natl. Acad. Sci. USA 85 (8):2444-2448. Pease S. and William R.S., 1990, Exp. Cell. Res., 190: 209-211. Perlin et al. (1994) Am. J. Hum. Genet. 55:777-787. Persic, L. et al. Gene 187 9-18 (1997). Peterson et al., 1993, Proc. Natl. Acad. Sci. USA, 90: 7593-7597. Pieru et al. Genome Research 6:492-503, 1996. Pitard, V. et al. J. Immunol. Methods 205 (2):177-190 (1997). Potter et al. (1984) Proc. Natl. Acad Sci. U.S.A. 81 (22):7161-7165. Prat, M. et al. J. Cell. Sci. 11 l(Pt2): 237-247(1998). Ramunsen et al., 1997, Electrophoresis, 18 : 588-598. Reid L.H. et al. (1990) Proc. Natl. Acad. Sci. U.S.A. 87:4299-4303. Risch, N. and Merikangas, K. Science, 273:1516-1517, 1996. Robertson E., 1987, Embryo-derived stem cell lines. In: E.J. Robertson Ed. Teratocarcinomas and embrionic stem cells: a practical approach. IRL Press, Oxford, pp. 71. Roguska M.A. et al. PNAS 91:969-973, (1994). Rossi et al., Pharmacol. Ther. 50:245-254, (1991). - 91 - 008252 Roth J.A. et al. (1996) Nature Medicine. 2(9):985-991. Roux et al. (1989) Proc. Natl. Acad. Sci. U.S.A. 86:9079-9083. Ruano et al. (1990) Proc. Natl Acad. Sci. U.S.A. 87:6296-6300. Sambrook, J., Fritsch, E.F., and T. Maniatis. (1989) Molecular Cloning: A Laboratory Manual. 2ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York. Samson M., et al. (1996) Arotore, 382(6593):722-725. Samulski et al. (1989) J. Virol. 63:3822-3828. Sanchez-Pescador R. (1988) J. Clin. Microbiol. 26(10): 1934-1938. Sarkar G. and Sommer S.S. (1991) Biotechniques. Sauer B. et al. (1988) Proc. Natl. Acad. Sci. U.S.A. 85:5166-5170. Sawai, H. et al. AJRI 34:26-34 (1995). Schedl A. et al., 1993a, Nature, 362: 258-261. Schedl et al., 1993b, Nucleic Acids Res., 21: 4783-4787. Schena et al. Science 270:467-470, 1995. Schena et al., 1996, Proc. Natl. Acad. Sci. USA, 93(20):10614-10619. Schneider et al.( 1997) Arlequin: A Software For Population Genetics Data Analysis. University of Geneva. Schwartz and Dayhoff, eds., 1978, Matrices for Detecting Distance Relationships: Atlas of Protein Sequence and Structure, Washington: National Biomedical Research Foundation. Sczakiel G. et al. (1995) Trends Microbiol. 3(6):213-217. Shay J.W. et al., 1991, Biochem. Biophys. Acta, 1072: 1-7. Sheffield, V.C. et al. (1991) Proc. Natl. Acad. Sci. U.S.A. 49:699-706. Shizuya et al. (1992) Proc. Natl Acad. Sci. U.S.A. 89:8794-8797. Shoemaker D.D., et al., Nat . Genet. 1996;14 (4):450-456. Shu L. et al. PNAS 90:7995-7999 (1993). Skerra A. et al. Science 240:1038-1040 (1988). Smith (1957) Ann. Hum. Genet. 21:254-276. Smith et al. (1983) Mol. Cell Biol. 3:2156-2165. Sosnowski R.G., et al., Proc. Natl. Acad. Sci. USA 1997; 94:1119-1123. Sowdhamini et al., Protein Engineering 10:207, 215 (1997). Spielmann S. and Ewens W.J., Am. J. Hum. Genet., 62:450-458, 1998. Spielmann S. et al., Am. J. Hum. Genet., 52:506-516, 1993. Steinberg N.L. (1992) Trends Genet. 8:1-16. Sternberg N.L. (1994) Mamm. Genome. 5:397-404. Stryer L., Biochemistry, 4th edition, 1995. Studnicka G.M. et al. Protein Engineering 7(6):805-814 (1994). Syvanen A.C., Clin. Chim. Acta 1994; 226 (2):225-236. Szabo A. et al. Curr. Opin. Struct. Biol. 5, 699-705 (1995). Tacson et al. (1996) Nature Medicine. 2(8):888-892. Taryman, R.E. et al. Neuron 14(4):755-762(1995). Schaid D.J. et al., Genet. Epidemiol.,13:423-450,1996. Те Riele et al. (1990) Nature. 348:649-651. Terlau et al., Naturwissenschaften 85: 437-444 (1998). Terwilliger J.D. and Ott J., Handbook of Human Genetic Linkage, John Hopkins University Press, London, 1994. Thomas K.R. et al. (1986) Cell. 44:419-428. Thomas K.R. et al. (1987) Cell. 51:503-512. Thompson et al., 1994, Nucleic Acids Res. 22 (2):4673-4680. Tur-Kaspa et al. (1986) Mol. Cell. Biol. 6:716-718. Tyagi et al. (1998) Nature Biotechnology. 16:49-53. Urdea M.S. (1988) Nucleic Acids Research. 11:4937-4957. Urdea M.S. et al. (1991) Nucleic Acids Symp. Ser. 24:197-200. Vaitukaitis, J. et al. J. Clin. Endocrinol. Metab. 33:988-991 (1971). Valadon P., et al., 1996, J. Mol. Biol., 261:11-22. Van der Lugt et al. (1991) Gene. 105:263-267. Vil H. et al. PNAS 89:11337-11341(1992). Vlasak R. et al. (1983) Eur. J. Biochem. 135:123-126. Wabiko et al. (1986). D № 4.5(4):305-314. Walker et al. (1996) Clin. Chem. 42:9-13. Wang et al. (1997), Chromatographia, 44: 205-208. Weir B.S. (1996) Genetic data Analysis II: Methods for Discrete population genetic Data, Sinauer Assoc, Inc., Sunderland, MA, U.S.A. Westerink M.A.J. (1995) Proc. Natl. Acad. Sci., 92:4021-4025. White M.B. et al. (1992) Genomics. 12:301-306. - 92 - 008252 White M.B. et al. (1997) Genomics. 12:301-306. Williams, et al. Science 257: 389-395 (1992). Wong et al. Gene. 10:87-94 (1980). Wood S.A. et al., 1993, Proc. Natl. Acad. Sci. USA, 90: 4582-4585. Wu and Wu (1987) J. Biol. Chem. 262:4429-4432. Wu and Wu (1988) Biochemistry. 27:887-892. Wu et al. (1989) Proc. Natl. Acad. Sci. U.S.A. 86:2757. Yagi T. et al. (1990) Proc. Natl. Acad. Sci. U.S.A. 87:9918-9922. Yoon, D.Y. et al. J. Immunol. 160(7): 3170-3179 (1998). Zhang et al., Neuropharmacology 32 (11): 1075-1088 (1993). Zhao et al., Am. J. Hum. Genet., 63:225-240, 1998. Zheng, X.X. et al. J. Immunol. 154:5590-5600 (1995). Zhu Z. et al. Cancer Res. 58(15): 3209-3214 (1998). Zou Y. R. et al. (1994) Curr. Biol. 4:1099-1103. Список последовательностей <110> GENSET <120> Связанный с шизофренией ген и белок потенциал-зависимого воротного ионного канала <130> 93.W01 <141> 2001-12-04 <150> US 60/251,317 <151> 2000-12-05 <160> 8 <170> Patent.pm <210> 1 <211> 82293 <212> ДНК <213> Homo sapiens <220> <221> признак ошибки копирования <222> 1..2000 <223> 5'регуляторная область <220> <221> экзон <222> 2001..2026 <223> экзон 1 <220> <221> экзон <222> 19188..19334 <223> экзон 2 <220> <221> экзон <222> 22996..23178 <223> экзон 3 <220> <221> экзон <222> 39731..39814 <223> экзон 4 <220> <221> экзон <222> 41337..41476 <223> экзон 5 <220> <221> экзон <222> 41565..41693 <223> экзон 6 <220> <221> экзон <222> 73013..73167 <223> экзон 7 <220> - 93 - 008252 <221> аллель <222> 12642 <223> 99-62626-168 : полиморфное основание А или Т <220> <221> аллель <222> 14088 <223> 99-62632-275 : полиморфное основание А или G <220> <221> аллель <222> 24981 <223> 99-62633-409 : полиморфное основание А или Т <220> <221> аллель <222> 69248 <223> 99-62611-51 : полиморфное основание А или С <220> <221> аллель <222> 73428 <223> 99-62605-56 : полиморфное основание А или G <220> <221> аллель <222> 80250 <223> 99-62635-44 3 : полиморфное основание А или G <220> <221> связанный праймер <222> 12343..12363 <223> 99-62626.гр <220> <221> связанный праймер <222> 12793..12810 <223> 99-62626.ри комплемент <220> <221> связанный праймер <222> 13814..13832 <223> 99-62632.ри <220> <221> связанный праймер <222> 14279..14296 <223> 99-62632.гр комплемент <220> <221> связанный праймер <222> 24863..24881 <223> 99-62633.гр <220> <221> связанный праймер <222> 25378..25396 <223> 99-62633.ри комплемент <220> <221> связанный праймер <222> 69198..69218 <223> 99-62611.ри - 94 - 008252 <220> <221> связанный праймер <222> 69632..69650 <223> 99-62611.гр комплемент <220> <221> связанный праймер <222> 73005..73022 <223> 99-62605.гр <220> <221> связанный праймер <222> 73466..73483 <223> 99-62605.ри комплемент <220> <221> связанный праймер <222> 79808..79826 <223> 99-62635.ри <220> <221> связанный праймер <222> 80314..80334 <223> 99-62635.гр комплемент <220> <221> связанный праймер <222> 12623..12641 <223> 99-62626-168.mis <220> <221> связанный праймер <222> 12643..12661 <223> 99-62626-168.mis комплемент <220> <221> связанный праймер <222> 14069..14087 <223> 99-62632-275.mis <220> <221> связанный праймер <222> 14089..14107 <223> 99-62632-275.mis комплемент <220> <221> связанный праймер <222> 24962..24980 <223> 99-62633-409.mis <220> <221> связанный праймер <222> 249Э2..25000 <223> 99-62633-409.mis комплемент <220> <221> связанный праймер <222> 69229..69247 <223> 99-62611-51.mis <220> <221> связанный праймер <222> 69249..69267 - 95 - 008252 <223> 99-62611-51.mis комплемент <220> <221> связанный праймер <222> 73409..73427 <223> 99-62605-56.mis <220> <221> связанный праймер <222> 73429..73447 <223> 99-62605-56.mis комплемент <220> <221> связанный праймер <222> 80231..80249 <223> 99-62635-443.mis <220> <221> связанный праймер <222> 80251..80269 <223> 99-62635-443 .mis комплемент <220> <221> ошибка связывания <222> 12630..12654 <223> 99-62626-168.зонд <220> <221> ошибка связывания <222> 14076..14100 <223> 99-62632-275.зонд <220> <221> ошибка связывания <222> 24969..24993 <223> 99-62633-409.зонд <220> <221> ошибка связывания <222> 69236..69260 <223> 99-62611-51.зонд <220> <221> ошибка связывания <222> 73416..73440 <223> 99-62605-56.зонд <220> <221> ошибка связывания <222> 80238..80262 <220> <221> признак ошибки копирования <222> 2375, 2377, 4041..4042, 5180, 7138, 7174 ,10118 , 12577, 12637, 1264 8, 13556 15322,15341,15357,15383,15409,15994,16064..16065,16077,16237 16264, 17751, 19182, 19424,20735, 20840,21865, 21885,21900, 22814, 23954 24969,24985,25001,25005.-25006,25017,25040, 25057,26646,27478 274 96,29278, 30793, 31122, 31125, 34 515, 34541, 34 563,34572, 34884, 34 915 36809,36834,3694 4, 37011,39862,39881,39895, 39903,39921, 39948 39954..39955, 41108, 4 2572,4 34 23, 44122, 53546, 53634, 55777,56140 56160,56182,56185, 56200,56206,56219,56234,56258,56268,56280,56290 56324,56561, 57259, 58256, 58297, 58910, 59321, 60724 , 60743,68789, 68803 - 96 - 008252 68871, 69936, 77190, 774 84,78780,78891, 7 934 3, 80394, 81773,817 95, 82232 <223> n=a, g, cmt <400> 1 catcaatgca aacaaaagaa gttttttaaa tgtgctggat attgggattg aggacagaga atcactgcct atttactaac tatatatata aaagccgcaa gtagatttag ttgtttaaaa tctcgcatga aaacgtacta gattaattaa acttctatac tcttgtagag gatgtctaaa attccagacc gcatggtgct tcagatgcag ctgaatagac cctatttaga acttgaggtg gcacagtctg gtaccccgcg agacccccga tggcagccgg cagcctcctc ttaaaaattt cccccagcct ccccagccgg ccgcccgccc ccgccgcccg ccggtgcccg cgcgctcgtt agaggggctg cgcctccacc gcgatgcgcc tgggctggct agcaggtgtt acctgccccc gggaggtggg cccaccccca cttgttaagc aacaccattt ctgggggccc gaatgctgaa gtacaatttt tcagaaggtt ggtggtgaaa tctaataggg atatatatat gagaggagcg tttaaattca tgcaacgaga tcgatttttc gaagacaaaa ttctgtgttc ctccaggcta gagagatact atgattgtct tccatgactt tttggctcat aaggtctgaa tgaccactfcc agataattcg gctatatatt ataaagatga cattgtttaa cctggctttt ctctgtattt atatggatta atatctgtaa cttgccacat aaagcttcat tcactggtca caaagtaaac taggaggcca tctgctgtcc tttgtctcat aaactataag aacggtttca acctcacgct tgtgcccggc gaggagctcg caagaaggaa tgcggtgccc aaataaaata gcccggcggc gccgagcgcg tgggcgccgc cgggcaccat ggaggctgcg agttccttgg ctagaggggc cgagagcggg ctccctattc tgggaagccc ggggggcagg tcggcaccag ggtgggcagc tgctcctgtg agccgtcatt tgtcactgaa agatcgcagt ctcagaccct tcagacactg ttactcttaa tgtgggttga gatcaaaaat gtgtgtgatt gtttcactat aagataaggt aacatttact gttcttctgt tccagctttg atagaagaca ggcaacggag gagcaggtag actccaatac ccttgaagcc gagcatgaat atgcctgttg aaaacattat aacgaatgtc tcagtttgtg tgaaacaagt attactgtca gacacagttt catgaaatac tgaatgtgaa ccagaaaata aacattttgt tcagttgttt ggagttccct ctttcagaga gatttcccta aggtatactc ccacacgaca tgtgactgat ccctttcata cgcacgctgg ggactcccgt cagatgagag ctgccagtac gcccttcggc ataatttttt ggcggcgctc ctgcctgagc gcctggcact gacagctctg gcgcggccgg ccggtccggg tgcgggcggg gcggtggggt ctgcgcgtgg agggccggcg tggctgagga ggccgctgcc ctggctccag gattgctcct agggtggcta tctttaagac cattttgctg tagtgcttga tattattaag tctggtgttc ggaccagtga taacaggtgg tgtatgttca ggtagttacc ttccacagtt ccctaactgg agtatacttt gagaaaaact taatctgttt ataggccttg aaggggtact acagtgaatc catgaattct tatagagact ggtgaggagt aaataaatgt aaggaataac aaataaacta tggtaaatac attttttctt ttctgaagtc caatatttta caaattgtga catttttaaa gatagacaaa ctttcccttt ggggattttc gtgggaatga aacctgctct acagggtatg acatatttta gcattcatta ctcaggaatg gctcccggcc gcacaccgcc cctagctgtt cgaccctggc atcactgcag aaaggcccga agtgagtgac tgagccgccg gatcctgccg .agcgctgggg ctccgtgccc ccaggacggc ggccccgcgc ccttgccggc gtgtcggggg acacntnggg cgaggccact tgggtaaacc caggtctccg tgctaaatgt caccgaacgc accagcaagg acttaatgtt aataccttag gatgtgtgaa gtgtggtcat ccttcgtgtt cgattttgaa taactacaac cctgaggcag ctaaataaaa ttggctgtca gacctagcca taaaaagtga gcagaataaa tctctaaaaa atgatatttt cctatgcatt ctgagactgt taacagaggt ggcactgaga t taaatattt acactatcat gtatttagaa tttttctaag atgaggagtg tttagaaatt acatttccct atggactttt atgtaggttt tgctctttgt atttatatgc gtatctgact gtgggaggat gctgggagcc gggtggggga attttaaagg tttaataact gaactagtcc tcctggcagt gccgtgctgt gctgtgagac gctccccacc tccctgtagc aaactccggc acggcgagcg taggtgaggg gtcgcccact ttacaggttg tgcgcccgag tcaggggcgc gggggagggg ctggccgggg cgcctgccgc cctgccccgc gggcatcact tctgggccaa cgggggctcc cccttatgat aaaacagtgt atggcctggc cctctttcag aataaatctt tacacgctca cagctaggcc agtagttggt atatatatgt tgtgatttga atgcatctca atcatggctt ttctgcgtgc ctgctaaaac aaaggttcag tgtgccgtca taacaaagaa ggatgtacct ataggatgag ctccatcatc agttcaacca ttagtaaaaa tcagctacca caagcaattg caaactggtt atttcatcat tgaaaaatta gtaaacaata taagaaagta gtgtagtttt ttagaatttc ctgtgtgcat acatggcaag gtcgccctgg ctaggaagcc ctgtcaattt cgaacgtcaa ccggggtgaa tcggtaccgt gcacttaagt tgagtacgaa tgactgcggg aactgcggcg aaagctaact tcccacccct tgagccgcgg gcccgcgtcc gtcgccgccg gtcaccgccg gcccgcgatg tggggctgcg tggggagggg cgcgggagcc gggatgcccc ctgggcatct acagcgctgc cgcgcgggtg gcggccttgc attcattggc cctgccaagc tgggatccct gatggtgtga tcagtttcag ctttcaggcc cgactgccct ggataacacc atgtatatat tatggcctgg ggactggctt cccaaacatc ttttctagag cagacagtgt ttggatcctc ctgataagga - 97 - 008252 ctatgctaaa cacttataac tacttaaaaa ttgagtagca ttatcattcc tttatcattt cccaggtcag tatatcctaa taaactgaga tgagatgtct agaatggtcc ccaaggcagg ccccgtctct agttacttgg aggctatgat ttaaaaataa catgttatac tatgcaaatg agagatgata atgaccacaa tcatcgtggc ttaaaatatt cccagtaaat cagtttttca ttatgttttt ttcaaattgg tatgtctttg tgatttccta taaatctacc agaaggtaac caaaaatcac = V "nf ¦"+• = tggctgttgt atgatcaaaa atctttcttt caggactgat aaaaatacgg tggaagcagg agccataccg ggggtaggcc gccaagagaa aagagatgaa gagacacacc tgttttaaag gatgaaaata agcctgataa tgatcaaatg tcctctctaa atatcagtag agaaatacat tcagaaaaca agttgctatc attatagtat tcccagcact tggccaacat ggtgggcacc acccaggagg agggcaagac aagcggaagc attgggagtc ggataaagtt atctgatagc gtatatttaa atctctacag tttggatagg aagtctactc ttttcttctt ggcatgcttt atcgatgatg agtcacatgt gataagctaa gcctaagaat tattttctag ccaaggccac cctattgctt acaaaaaaaa gaggctgagg ggcaccactg aaaaagaatg aattctcgcc cttaaatttt gaatgataga tatctgtgtg attggacaac tctatataat acagcctcat tcattataaa cagtataaat tttcctgaaa aaaaaagaag aagaattcan cttttacaaa ataataattt ctgaaaacca attcaggagt gggcttacat cctaggctgc aggaaaagct aagagagacc cttttgtaaa gaatatgttc tccgcagcag tcatttatga aagaggatgt aataaatgtt tgagataaga agttatcatg aaactttgga cttcagaaat aattatcagc acaattctct actaggcaag gttcaaaaaa tttaatacat gcacagtcaa ttgggaggcc ggtgaaaccc tgtaatctca cagaggttgc tccgtctcaa atgcccacag agcatttcat ttaagaatga aaaaactgag ggtaaattag aactctagga acttgcaaga aaaatagtgt cttgggtatg gtgtgtgcat ttcagataat ggatttcaag gcactgggcc nnttcaggaa ttttgctgtg acacggtggc gaacccaaga aaaaaattca tggaaggatc tactctagaa cccctgtccc tgtctcaagg tttaattgat gaaaaaagag ttttttgtag ttttatactg tttcttactt atatttaacc cagtgtcctg atccaaatat aattcagatt tatctgaatt gattataatt aagtgttggt ggatgtttga gatgctaaaa ctaattctgc agtgagaaat agaatccaag aaagttcagt ctggaagagc acagaattgc tgaagacagg gagtggaaga agagatcaag tgtttggaga tttcctgaca ttcttttttt taccaaaaca tttgaaaacc tacttatcca aaacataaaa tacgttgtaa tgatatcagt ttgtttgatt ttataactac aagcataatg gagatgggtg cgtccctgct gctactcggg agcgagccga aaaaaaaaaa actcaatatc cttaagaagt aaacatgaat agctacagca aatgccaaca gaaagcatgt tattcaaagg catttagttg gcatttcttt gtatgtgtat ttcagaattt tttgtgaatg agaagctggg agacaaatat gaaactatga tcatgcctat gtccagacca ccaagtgtag gcgtgagccc tgggcgactg aaaatgttat accagcacag tttttgaatt taaacagcta tctttttaag tttttaccac aaaaattgta atacctctat atagttataa gtccatatct ataatttatt attatagaga ttgaataaat ggatcacatt cctttgtgct tatacctctt tttttgcttt ctgtaccccc gcttttctgg tcacaatttc cccttagtat ttgtttctcc gactgacccc attagaagta agtgagaatt gagaatgata gagaggatca cttttagatt attaataaaa aataaaaaga caaagagaaa taacgtcctg atgagtcaaa aaaaaaaaat aaatgagaaa aggcttccca ctggccgggc gatcacgagg aaaaatacaa aggggaggct gatcacgcat aaagtctgat tttttaaaga acaaaaaaaa gaanttcact ttccaatctg aattggaatg cttataaaat agaaatgtac caatttagtg gaaatggttt acacaaatgt ccatagctgt aaaattcaca tggtgagtga gtaagcaggt tagtgagctt aatcccagca gcctgggcaa tggtgtacac agagatgtca ggcacagagt caacatttta attttcatac ttaaagtaac gttttaataa ggcttgactt tgaatattta aatggctgta cattggacat tgtagagatt gtcacctatt tatgataaat tatattgtat tgcttaatct ccctctttta atataaaaac agagtaaagc +- лпэ о "-i rir> ^ w" " ^ u LWV cctactcaga acaggtaaag tagattcagt ctttctaaga gaaggagcga atgagatccc acaggtccag ggatagacca atttccaatt tagatggaag cagtgcatgg aaaaacaatc cttagccaac aaaatatttc atttagatta aaaatctgta agtgaaaaat gaggtagaag aaagaccgta tatataccaa gcggtggctc tcaggagttt aaaaattagc gaggcaggcg tacactctag gcttatttta gttttgtgac aagaaaaaaa atgacataga ggaggaatat atgacaagga agtggttgta ttaaaaaatg aactgaagat ccaggtgatt acacatttgt ataaatttat tctacacctc gaaaaagaga ggatatacag aaacatatta ctttgggagg catggtgaaa ctgtagcccc aggctgcagc gagactctgt gagcttcact cttcaaactc caaagttgat ccgaagacag caggtagttg ttataaatat taacaccata ttgctgtttc tatttctatt tttccatatc ttccagtaat gtctttaaat gcttttgagg cagaaataaa ttgaccttta caaaatagga +-+•+¦ => /Т =t 9 a s *> cttcaagtaa gaagaagagg atagatggaa tcagtgtaca agttgaaagt agggcaaggc ggtgggggca gaacactgaa atgcaaagaa acagccaaca aacacagcca caaatcttca atctacatat accagcattg atgttatact gtcctgaaaa aaataggtga tgtgtaaata aagtcataaa cgatcagagc acgcctgtaa gagaccagcg taggtgtgat aatcatttga cctgggcgac ggggaaatgg ctaaaatggt aaagaaanga taaactctaa tgatctacag ggttagtgaa - 98 - 008252 gaaatgcact ggagaaattg gatatacaat tccaagattg aagaactgga atcagattgc 7380 agtaatggaa atgaagaggt ggatggtaaa gatatatcca caaaaactaa tgctaccacg 7440 tcttagtgat ggaaggaaga gaatgaaagg gagtcctagg ttaaaaataa tttccgatca 7500 tcgagcttgg gtgattgtgc tattgtattc tattaagtct agttaaaaga aattgttacg 7560 gctttatctt tcagttaata gtatctacag ggtaaatcct ttgcttcatt cccttttcta 7620 gttgttttat tgttattttg aaattcacat gagattactt ccagaggcaa tatcgttttg 7680 cttcctttct ttgaaggcat tgaatggcaa tttaaaggtg tagtctagac tagaaaatca 7740 taacagaaac cccaacaaaa actcaaatat cttagttatc cttttataaa acaaaaattt 7800 aattcctgta tacaatgcaa gttagttatt ttaaaagtaa attctgaaga tttctgagga 7860 aagaatagaa tatcctgtat atacatacaa ttataaatca aaatttttgt ctcagacata 7920 ttttgcctgt gttgcttata attagactct cttattcttt tccacacaga gccatgacta 7980 tgcaatgtga actctctaag tgttctcttc tgagcatcta ggctagtgat gcctcattaa 804 0 tttggaaaca gtgatgggag aggtgtgcag ggatcacctg aagatgtttt taaaaaaata 8100 caatgtcctg ttttccaccc ttaaagattc tgatttagta tgtctgaggt gaaggaggcc 8160 tcagcttgag tgtcttagtt agcttgggct gctgtaacaa atagcacaga ctaggtggct 8220 taaactccag actttggttt tatacagtct ggagactaag aagtccaaga tcaaggtgct 8280 ggctgattcc tgtgagggct gtcttcctgg tccacagacc attcagctgc ctccgttctt 8340 cttatagatt cgctagtccc attcatgggg tctccacctt catggcctca tgacctccca 8400 aaggccccac ctccaaatac attagagata agagcttcaa catatgactt gaggaaggca 8460 gaaccattca gtttatagtg ttgagaatgt ttctgaacct cccttgataa ttcatatgtt 8520 aagcagtctt taggaattac tattttaaac catccccttt tgagcaaatt tgtggattca 8580 tattgcaaaa tttaacaaac acaaaaacat aatctccctt tgatcttctg ttatgtttaa 8640 atatagtagc aaattttgaa aaccactgtt ttaaaccatc caccttcttt atctaacctt 8700 tacagattca tattataaga ttaaacaacc acaatcaaaa taagtatccc ctttctggtt 8760 tttcataata tataaatgat agtagcagct catcaaaata aaatatccaa cactggataa 8820 ttaatataga gtaggatata tactcagcac agggtaggac ggtggtggcc aggggctggg 8880 aggaggggaa aatggggagt tgttaattac tgggtagaac gtttcagtta tgcaagatga 8940 gtgagctcga gagatctgtg ttatgacatt gtgcctgtag ttaacatttt atagtgaatg 9000 taaaaatctg ttaagacagt tctcgtgtta aatgctccta ctacaatgaa acaaaaataa 9060 aaattactgt gatgtaggtg acaaaaatac gacccagttc cctctcatac tgcttgacag 9120 ttttagtttg ggcatgtagg tgcaatgaca gttttagttt gggcgtgcag ggtgcaatga 9180 ^-i-,-^-!^^^,^-. nfnn(*Mf/Tf ^4-^9 -" +¦ /-.^^4- ^-i QOitft u" и ua <^ <^ ч^ч"са u ^. аааддссада gaaaattcaa ttttcacaat taaaaaataa aacaaaacag aatccttgga 9300 gaagtaatga cagttttgga gtttttctag aaataaccag agaaaaataa cctgctgttt 9360 tactttggga tttaagaaat agacttataa aaagtgggga accatttttt atttctaatc 9420 agtaccttga caaaaggaaa acagaagagt ctgtttttgg agatggttct ttgaaacatt 9480 tgccttcccc tatgtggtca ccaccttggg tagcccttga agtctgctgg gactggacca 9540 tgcatgcaaa attctgtttt ttgcagagct acattattta gtgagggcca agacacagat 9600 gaatgagtac cccaaaagaa aagtggcagg tggcatttgg aggagagaga gagaaggaga 9660 gaggcagaaa gaaaaagaga aggaagaaca aagaaaacca ggcagccaaa tcagggtctt 9720 gacttttgta caaaattaga ttgaaaatca accacaacaa tccactacct gcaggggcag 9780 gggagtgggc cgctgttctg ttgtggagac acgcattgcc gctgtgatgc ccagggtgtc 9840 agttttttga tgctgtctta gttctcctgc taacatgttc tctgctgtgt tgctggggga 9900 gacgcaggac tccaggctaa gtttaagaga gaaaacatag agctgctgcc tttcctctga 9960 gcagaggcca gattacaggg aatgatgcat ccagctcatt gaagttcaca agctaaggat 10020 acgcggagga gaggcttgct tgagagggcc cggaaggact ggcattgtaa gtctagtggc 10080 aggcatgctc taatttggtt ttattactta gacgtggnaa taggcaggaa cttggagatt 10140 cccgtctctg ttctctggga aagaagactg ttctatttac aaagacttga aagggtcaga 10200 ctgaaagaag acatggacag caaagtattc ctggggtggc aatcagaaag acctggctcc 10260 ctcgtggttt atcacagcta accatgtgac ctcagacaag gcacttatct tctccaaaac 10320 tcagtttcac ctgtaaaatc agaataacag agagagagac acttacctca ttgaggcatg 10380 gtgagaaata ggtaacaatt agcctcacac ttaataaacc ctcagcaaat gttaggtgta 10440 art +¦ rrt- t-+• = = = <~+- rr = = = = *• t- t- -t- f = = 4- rrb bffafH-rrtt fraaanaaat- t- a t- a a a f- a a a 1 ПЧПП acacatcatc tttgtgaaca tgtacagaag ttcgtgtttg tgaaatgtac aaaagttctt 10560 aaggttaaaa agtgaaagtc aaatgcttca ccatttctct ccctccctct gttctcccaa 10620 caaatagact acttggattc taaacttgct tccaattcat gtgggtctgg gcatgtttgc 10680 ctccaatttt taatctctgt agtgggaata atactaaaaa ctacttcaga gaggtgtcaa 10740 atgcttagaa tagtgcctgg catggagtaa gctttcaata atatttgttc tcatctttca 10800 agttaaaaaa atgagtacat aatgtctaag cttgcttcta ttctataatt atgcacagta 10860 gtttatggac tttcattcta ccctgagaaa tgatcaaatg cataaaaatc tgagcagtgt 10920 caagagtatt aatcactttt atcattggaa gtcactttgg atatcatcta ttgcaaagtc 10980- ccagacagag gagaaatctt ttctgcaaac tctaaaatgt gtagtcattc cctctgagag 11040 tatccataaa ggagtttacc accttttggg atattccaac atattatgat tggaaactta 11100 - 99 - 008252 tagagtagat atttctattt taacaactgt ccttttaatt tcataggtgg gagactaaaa gtctcttcac tctttaccaa cctcaaggaa gatagattta gtgtggtgag tgagaaatgt ggctatatga atacttaatt taagtttcct ttttcttttt tatgatcttg ctcctgagta tttttttttt actcaagcga cacatggcct aacatttttg caaccagtga tggggtatat aagcattctg cccttaaatt ggagacagca ggaggccgtg atgatctggg tcagctgggc gttcttttta tgtctgcatt gttctgggct gctcttgcac aatctatttt ctaactctag tacatggggt atttagagaa aaaggaataa atgagaggtt tcaaataaac ttccttgcta ttggattttg aatctctgta ttagttttca gaagtgaaga gagattatca aaaaataatg taaatggata tgatgatttc agtcacctct tgataaactt gaagaacaga gagcactaca ggtatgacag aaaaactttc gaaaattaca tacattcttt tattcttagc ctagtcagtc aatctgtcat tttaggaaat aatgggagtt agtaaatatg aaggtataat agaatatttc ataccttcag aaacttgaaa taccagaaaa aaattgacat ttgttagaga aaacaaatca tcaatacata tagccatgta aagaaattac ggagagatct tttttctcat taggattcat gggttattta ttttataagt ?aaaaL^aua aa ctgtactatt tttaaatata tttgattcaa catctgtaaa ttctttttga gctcattgca gctgggacta tgtagagatg tccactgcct gaaaatgtat ttagaaggac gctcactatt gtgtgtgtgc agtaggagat gtcgtgatga gagttgctgg agaggtgacc ctcagagaaa agagggaggt attggaggac ggcagagtgg tgcacaaaaa ttgaaaaagt tagttctgtc atggttcttg gacagagtct gcctctactt caggtgcatg ggttttctcc cagcttccct gatattatta tctgtttttg ttaacatgga aagtgtgtgt aacattcatt ttgaaatctc attctggtct tcatggaggg gaggggaagt gcaggtatca gcagggatgg jQuaQLQQaa a l. ataattgatg ccaagagttt cattttctca tttcctgtcc aggcacattg tcccgtcttg tgaaaatgga atgacatagt cactctgtca cccaggctga cccaggctca agtgagcctc ccagtactcc cagctaattt atgttgccca ggctggtctc aaatgctagg attacaagtg aacagactgg cacataataa catgacagta aacgtaattt aactctcatg tatcccaaag gagctactga aggctatttt tttacangca acttctgttt aaggcangtc awtgagcnag ggaaagtgcc cagtgcgtgt agaggaagag cgtgagcatg tcatacactc aagatgcaca gtagctctga ggctcaaatg tgacacatga ggcaggtgga gtaaacacca attctgtaat tctctgtgcc gcatattctt ggggcagtag ccacctcaga tttgtatttt gaactcctgg ggagccacca cccctcaacc gggttgaaat cactttttca cctttggaga ttaacacttt cccctaaatg gagaggctgt caggacacag gagaggacac ggccctggga tggatgttgg atgtatgatg agcaagcaaa ccttttgtcg atatagggaa agttctgtgg atattgtaat gggtaatgcc tatatcagct gaaagacaag ctgcacatgt atctggttac atatgtggca cctaaagaaa tggcagacac tggaatccct ttctacaaca cttctaactt ctcctggatt actgctcaat ataagaaaac ----------3 gggaaaagac ggatagtcaa atgctggttg gttattgaac agagataaaa gataaacaat agagtggttg gtcaatgact caagttgtat caaatcacac ctacattcat attgacagtc tcaagttttc aggacaggaa ctattggaac gcctacatta tttctcagat caccttgtaa gaaggggcac gtatttattg aaaaccatgt aagaaagtcc attccacaag aatgtatgtg gatggctttc aacctgtatt aatttaaggt tttgcatgaa attccttgcc cactagatgg ttgctctgaa aaactttaca actgtggtgt gctcttgttc taaaaagagg acaaattggt acaaatttaa ggccttgcac agatgtttga acagaggatg gtaagcagcc taaaagagat agaccataac ctcaacaaat atttttgttt acttgtgtta ttaagtattt tctcattatg tagggaggta acgttgaaga tggacatgtc ttctgtctga gaaacaaggc catcttgctg attgaggtaa aactgaagat aaatgtagat tttagtggtt aagaaataaa aacatttgaa aacatcaaat acatgtgtct cactgggaac ctaatctgga gtataagcat gtaaatgagc aggtagtgtt tacttacagt atgttcacag aatatggcta atttaaaaat aaatgtatgc acaattcata tcctcttgga taattacttc atctgcagcc tctttgcaaa aatgattgag cttattttgt ttatatagtt atgaaaataa aataagaata gagatctagg caaagaaaga cccgatagca cagaaacrtg gtatggagcc agcagtaagg gaaggaactt ttaagcatcg gtaaatgtgc tgttgttttg ataagaatta tcaaaatatg aagctgtgct tatgggacaa taggantttt gatttaaaaa atggaaaaat gagtaagcac ccttgttcgg tctctagaag tttctaattt gaattatttt ccagtcagct ttgggaatcc tactgaagga gaaagtgcgc cacactagag gctctctttt rt* a^t* nt nrr tftaaaf ara rrt я t* rrt" +• rrt-r ttrrAtnfnfl ctagtatgtt aaaacaaaac gtttcccaaa attttctttt acccaccatt ttcacttgca ttggtattga tctgtcttct gtttatcttc acatagacaa ttgttcaatg aattatgcaa gacatgaaag cttaatttcc aagtgtgtat ctcctaaaga tgtattgaca ttggggcttt tcacatcttt agtcttcatg ttaaatccac gtcacttaac acatgaatga atgagaaact tcagagtttg aggtgaagag ttctcaaact gtgccaggga gtgtgtgtct gtgtctgcaa ccccacagtt tatgcacaaa accttaataa cacctaatga ttagaatgaa atagatgctt attcccaaat attaagtttt agttaagtga gggaattagg aaaatggtca tctgctagca tcccggagat tactaagcaa tagcgcggtt agttatcaga ttaaaaatga tcattttata tcagttataa aaagactgat - 100 - 008252 gtgggattgg cattggataa caaagaggac atggcttgga taaagggttt gaattagttg 14 940 agtaaaggtt gcaggatggt gaataatggt atgtttagcc aacctgagtg cagcatgtgg 15000 tatgagtgtg aaaggaaggg caggtacaaa tcagtgggac tcagggtcct ggaaacataa 15060 ggttaaataa agtaggcagt tcgtaagctt caagaatgcc ttcatattca gtgctgtata 15120 cccagcacat accgcagtgc ctgataccac agtggatgca taatatttgc tgtgtgaatg 15180 aatgaatgaa caaatgagtg agtgaatgag aaataagaag atacactgag gttttctggg 15240 ctacaaagga gacctacagt gtgtttggaa gctgattttt ttttatataa aagagggaac 15300 attttgaaag gcatcccaga cntgagggcc catggcctac ntagaaagga gtctagnaaa 15360 tggggatagg tgaaaaaggc cgnttgtaca tatatggaat gaatgagcna agaaaacaat 15420 ccactgctga gagtgggtgc aaagagaacc tgctgggcct tcccgctgca catggggaac 15480 ctgaagagtt tagagcagta gggtctggga catgagagga aggagaaaag caggatgcct 15540 ggctcatgtc gctctctcag gctgaatctt cattcatgat ggtgttggca ccgtcttcct 15600 acatctgcgg gggaagcagc accgatacct ctctaaatta gtagctggac acagatggta 15660 atggaaactg taacaacagg tgatcactta aggggttaag tagagaagac aggcaagaag 15720 ggcatgctgt gaaccataat ttggaagaca ggagaagatt caaaagactt taatactgaa 15780 cccaagagag ctgcaaggga aagaaatgac ccatagtgtt gcaagggcgc tgcagttaaa 1584 0 gaaaatggaa tattttaatt ttaatactgt gaagacgggg ctggaatgaa gtagggaaag 15900 tatataccaa tagaaatgaa attgctatca cagaatacaa aaactgggtc ttaaccattt 15960 aaaacagcaa tgaacatggg atgaatgagt ttcntgagtt gctataagct gtttctgagt 16020 aaaagattta ttaatggata cattgtgaac tgtgtagcct ctcnnccgat attgaantat 16080 ttaaatacca actactttta ataaatgtcg gaaacctttt gaaagccatg tttttcaaac 1614 0 tttgtgtact gtcttatgaa ggacactttg gtaaacagaa aaaagagata ttggcttata 16200 aaaacataat aatgataagg ctcagataca caggaancaa tttcctatca aaataataaa 16260 cttnacctag gatacttaaa tatccctact ggtttttatc agagaaataa taggtctgct 16320 gtatgcagtt ttattgattc aacatgaatg aaggcataaa acaatatttc tgtttgcaaa 16380 ggaaaattat tacaatcaac cacctcaaat taattttgaa ttcctcatgt gtttgtaact 16440 taaatttatt ttactacttg aatgctttat ataatattta atagtattaa attaaatcta 16500 gagatccagg agctagttgt tgagttttta tgtatgtgtg gattagccag gtatctttgg 16560 agcttagatc atttcttggg cactgtgaga tttagtggca cctgggagca ggtaaagtga 16620 aggtaaatgt gagtgacagc taatgtcatt cagttataat tgcactattt tggttggtca 16680 gatataacca aagtgggaat tttttaaaaa acaataaatt ctggacatgt atttttctag 16740 ataaacaaat ccataatcaa aaatgatttt tgctttaggt ggcgtattga taataaaggt 16800 gcatggaatt catatgccca ttgtctccac tgctcagtga aagaggaatt gtggacagat 16860 ctctaaaatt ccttatataa tctttgaagt gaaaccactc ttttacttgc tcctttgtat 16920 gtgtcatctt gaaggatttc taaaagattg tatctacact cagtatttgg gtggatagga 16980 gactgtcatt gaagaggcat ctcaaattga ggccaccaga tttccttccc cactaccagt 17040 acatcagaga aaaaaaaaac actttaaata ctctgtatac attaacatgg gatatcaaag 17100 agtcccaaac tccaattata taacataaat .taattagtgc agcctactgc aaagcccata 17160 ttcaaactca atattcagag caataacagc ctggaaatac ccagcatgtc ttcaagccag 17220 ttagcttgtt ttcttctggg aataggaggt gaatccgtcc gttttcacac ttctataaag 17280 aaatacctga cgacgctggc taatttataa aggaaagagg tttaattgac tccattctgc 17340 atggctgggg aggcctcagg aaacttacaa tcatggtcga agacaccttc ttcacaaggc 17400 ggcaggagaa agagtgtgtg aagtaggaac tgtcaaacac ttataaaacc atcagatctt 17460 gtgagaactc actccctatc atgagaacag catgggggaa accgccacca taattcaacc 17520 cctctctcca cacgtgggga ttgtggggat tataattcaa gatgaaattt tggggggtgg 17580 gaggacacag ccaacccata tcaggagagt taggcagaga tacctgccca ggtagataaa 17640 ggagcacata gcgatgggga taggtgctca atacaggcaa tcagattcgg agctgctttg 17700 ttctgagttt ctgtgtttta ttctcttttc cagtaaaaac tgcatctgtg ngcgcgcgca 17760 cacacgcgcg cgcacacaca cacacacaca cacacacaca cacacactgc aaacatgttc 17820 ttgtactcca gtgaggacta agtggcaccc attgtgcttg gttaggtttc tttctctaga 17880 cccatggagc accagatagg agagccagtg aacccgcctg atgttaaaac tgcagtgcta 17 94 0 cgttttcctg actcagcctc ttttttttgc ctgaagcgct cttttttttc tgttacccgc 18000 rftnanaaat' tntrrfaafa ntrtnranrt- t- г-д гтг"э t-nrrt- raartrfaaa rrrrtt-anar ----7 - -?---------------n----эч, - ww.--, v~ *2 ^ -----• ----v ~ .----" ----- attgtagtca aacagagctt attttaaatt tcaagtttgc ttggatggcc ttggtcaagt 18120 tattacctgc cttgaggcta gcttttctca tttttctcat gcatgtgtaa gaagatgagt 18180 tttttgtttt ttgttttttt ttggttaggg ctgttgtaaa gatcaaataa aatatcattg 18240 ctaagtgtcc caaacatagt atatgcccat agctgttagt tcctcacctt cctttaagac 18300 tcaatccagg tgctaatcaa atgggaagtc tcacagtcga ctcagggatc atcaggtacc 18360 acctgccttt ataacaattc ttctcacatt gtattgcact tatttgctat ttgtgtcctg 18420 agggctgaca tataatagcc ccccttcaag tgactgctga gttgtggaag gcagtagcaa 18480 tggccatagg gaggtaacgt ctatcacagg atgtagatgt ttcagaattg tggaaattta 18540 agcagttagc aggcatctca tctcccacct caatgtaatt tcatgaacaa tttacttcag 18600 agtagctgga agaatttaga taatcactgc attcataaaa caacatctcc atttagggga 18660 - 101 - 008252 ggttcttccc ctactgttgg tggggaagat tatgtggaga tctgccctaa cttttgaagt attaatttat tagaacacat agggcaagat gcttgctcac ccattctttc aagtaagaga ctgtgcattt cagctttagg gaagcaaccc acataaaata ctcatccttt atatgcctta ggtttagatg caaagctaac tgacctccct cgtctgtgaa catctgtatt tataagagca attttactct atccattgct taaagtttct caaataagta acaaatacaa ctcagcatgc i-n-ayvtayL ^at.LijautLL yytui-iyaLy ggattaacaa aaatgtgatt gtagaatatg tgtcgctatg cacgctgtgc gatccaaagc tcaacgttcc atttgcaaaa tcaccttcac actgtgtgcc tgctggaatc atgctgaagt ccatgataca aatctgccct attcacatca taaaatacgt ctgctccttt tctaggctct gaccctttcc tgtcacctct actgtatttt gtgtgtatat tgttatatac attacacata aaatatcatg tgtatgtata gtatatatgt catgttttat cctaaccatg tctttctcta cataatatat tgtgtgtata aggtgtttta ttctccattc ggaatagaga tacaaactta ataattcttc tatttggaag ttcacacacc aagtaaactg accagtaagt gacaaattca gatgaggttc agttactcac accctgtcct tggggtccaa tgaatccgtg tttgagtaaa cgcctaatta agaaagcata tcagctctgc aacaaaagga cacatgatct ccaggttact cgcttctgct tagtgtgaca cctgtgtcag ctggattgtt cagctcctct ggcatcctcc ^ -i ^ +- f- A-. tatatgtatc ttatataata acatacatat cacatatatg tgtatataan atgtgtacat tatatacgtg aatgttatat atacatgcag atctgccttc acatacatat tatatatata cttcttggac ccagcattct atgaagcacc tgaagaaggt attaaaatta ctcttttgta aatgtttatt aatcatcagt ttcttctttg aatatcatgt agagtctgga ataggaagat cagagtcaca gatgagcaac agacaatatg aggtcaatga agttgatgag gtaaccagaa aaaaccaagc aatggcctgt gcttggactc ttccctgcct ttataatcat ttcctgcttc cacactgtcc cacattgctt gatgagtcaa tctgaaaggc 4- 4-4- -a 4- 9 4-4-^wwUw"LCiww atatattatg tacatatatt aaacatataa ttatatgtgt catatataca atatttatat agactataca aatgtgacat gatctgcatg cagactacat aacatgctat tatatatata tgcaaagcag gacacatagt cctaactttt agtataaaag tgtctttaaa taattagatt gaagtgcttc ggcttatgtg cccctcccaa tgtcgtatta actcagcagt tacatctaaa agctattcta ttgggagaaa attttcatat gatatatgtg cttttccttt tcaaaaggaa agtctctgtc agtatggaga acctttggct agtagagaac gcactgggta tggtgtagag ctgttctgaa cacatgtttg gataatggat gatttgagaa ctaagcctga aggatcaact tccctcttgc tttgctgtcc gctcacattt ggaagcatcc ttttcattca acttgagccc tccctgctcg atcctcctca ttatttccgc fnf^b^f & a tgtatattta tatggatcta caggnttata taacatatat tgtcgcatat gtgcatgcat tgatatatgt gtttttgcct gactctttgt gtgtttacat gtgtatatgc tatatatata gagtcagaag aggccttcat tcttagtgtc atgtggcctt ctaaaaacac ctttatgttg caaattgcat gtatggaatt gttgggagat aattaacact gttctagcac aagattttcc aatagctagc aaaatcaagc gtttattata gggaactcaa tncccagact gcagagttcc ggataatgct agcatatgtg tcgnaaatat aatccaatga gtctgatccc atgataccac tgaacctcct gaattaaagg ggtatctctg gaactgctag caggcatgca tcttactcat agtgcttgtc ttcccagtct ctttctatat ctattacctt ggacctaacc catcagcagc tactgctccc tccattagga cacagacaga 4- s <- = = = nfn? cattatatac ttgtcttagg tgtatatcac gcatattacc gcatatacat ataacatgta atattatgtc cttgcagtgc tgtccttccc gtatgtgtat acacattata tatatatata tgtgttgctt tcatacttag catcttatta ggaggcactt ttttgttgtt aaagtccaga gaaaaataca agcagcaaaa caacttgggg ggggaaacca aaagcacctc caagaatatg ctctaaactg ttctgttggt cactatggag ttttttgatg gtggttttgt agggtggaag +- яя at- a+- +- a +- aaf^trrfTan r*+" nra a rrt n-t" tgaaagttct ctgcagaatc tcataatttc ttgtatacta ttaagtattt tgtatgtcct tttggggtta ttttgtggaa aagagaaaaa ctttgtattt gacagtcttt catctactac atatatgtcc attgattaac ttttaatcct gtatatatat aagaaaatct ttttgaaaga tctatgcatt ttatacaaag tctanttttt aagactaaaa cagatagtca tggtaatatt agatactagt tcatgttaac tcagacagca tgtctgggct tttttcaaat gcattatata catcttgagt tttttttcag ataaaaacgt aattgtggtg tttcataaaa acacacttat gacatgcctg aaatttgtat gtaggatgag tcagatacaa gtttngctag agtattatat atttggatta ggtttagcat aaatattgag ctttaattct tagtgttgca aaaatgtgtt accagatttt atagaaagga gatgtgagat ctgagctgca aatagtgaga ctcccctccc ttctcaggct ttacttgttg tctccttctg aagaactttg gccaggcacg tatccatgag ctcatggttg cttcccccaa ctcttcctct ttctattttt cattgcaaat tgtccagcaa ctcctactcc tggccccagt ctggatgaga atgtacctct V r**r I-.4- a a ^4- a attgtaggtc tatatataca atatatacac cacacatata atatattata tatgtaatat ttatctagaa ttgtctttcc agtctgtctt ttacatgaaa tatatgtatg atacattcat ccatattttg agaattgaat cagagctttc actaaaatta gaagaactta tggggctaaa tcaaccttta tatcttcatg nf-г?эгт+- r"t-r"r- - - -"www tatatatagn taaagaatta tgtttaagaa ggagatgcag tatgagtgat ttcttcttat cagaaagtta ctccaaaata taaaaagttt aattctaaat - 102 - 008252 atgggatgtg ttgtgctaga tagtacaaaa cacatatttt agatgtttgg cacagaaatg aattaaatga gatttcttct acttagtgtg ggaccatcat tctgtttgta ttcactttgg atccggggca tgaaagtctt tttatcttat ttgttgcatg atctgagaag catgccactt ttatttgttg tattgcagca aggacatgag agtaccaatt aaaggaagaa ggttaaacat cagtaatagg tttcacttac attttgtaca atttcattgc acagacaatg cccaaagtat atttagccaa apf *— r*-t- +-+-¦*-/* -- - --r> aaatataata taatgtaggc tatggttggt ttgttttcgt gatctcagct ccaagtagct agagatgggg ggcctcccac cttttcacat ttgtcttgtg aatagtttnc tgcacacagg tttgccagtc ttgagagtga tcattcaatt caattgtttt acaattctta gagttttgta attttattta ttttaaatga gttctagatt tgatcatatc agttgctttt taaatactag atctctagtg attggattaa ttaatttgtc aagagaagag tgggtcaaca ttccttattt gtgggtggcg ttatatatca tgctagggaa atgaaaaaaa aattgaacca gagagcctga cctccacctt tatnaaaaac tcttcacctc gttcataata tgcagtgggt tgaatacgcc atacattatt ttgtcaaggt aatcccttta acatatggaa agactatttt gaacaagatg agggctgagc acttagggag cgatgtcatt atggaaaaaa ggaaacaaaa aatttctgtc atgtagatga agtgcagatt tggnttgtat aggattttga ttccctttta cattaatagg attacagtta ttctgtttca +¦ a i" +¦ nrr-r+- мал atattttacg taatacttaa ttcttcaact ttttgggagg cactgcaacc gggattacaa cttctccatg ttatgtttct cctgcaaaaa gaagattcta tgtgtactcc tgcttctttn tgaagggtgg ggttagatgg ttggccactc tcccagtttg tccaaatcat ggtttcctca atcattttgt aggctgttgg cttgtattag atattcaaat acttacttaa tagaggtagt tttcttcatt gtgaataaac aaatgtttat atttttaaat tacattttta taaatttggt ggcatgtatg attcatttaa gttatggtaa agtaatggca gaatcttcct tcaggttccg tctgtcttag gtgaaaatat catattagta caagtttgag tcactctttg aatgaccttc gatgtttctc gagcacttcc atcatatttt gtccttttta aaaagttaaa gtttgactaa ccacaaccca tgcagaatca tacttgatgt tgtctggggc tagatataaa actggatatc tgctgtccct ttagagtttg cagcatctca gaatctactg tacataatta ttcttgttta taatgtgaat ttttaaagca tatgataaaa cccaccgttc caagactcag gaggttcact tctgcctcct ggcctaagcc ttggtcttga taagtctcct attttcctat catttgagat wtgtncctct cccacatggt gaaacggtaa gattttatgt tgtgtgtgtt ccattgtcat ggacaaactt tacagatttt tatttgtatg tttctataca tcataatagt aaagaaagat agtaaaataa gaacattttt aactagttca tttaatttca ttttaatttg attgagttat ttctatttta ctaatctttg cacactataa aaagcatttc atatagtatg catcaagttc atattgtagt tctatgcttt tgtgtttatg ttgtagcaca aactatttta atatcagtta ctgcgcatct gagcactatc tacacggcag atgtcattta cttgttcatt aaaaggcata taaattttgt tgactgtgtg gcacaggggc gagtcccatg gttttgaaaa caaaaggaaa atgttttaaa ttttccttcc agagtattta acaaatgaca agttagctaa agacaatata attgtgtgcc actttttatt ccctgtaact aatctgagaa ttaactacta cccctaattc ctatgtgact ctgatgccca gggtctttgt tggaaaatag ttatatgcac acctcagtat tggtgccgtg gtgccatcat ctccacttca agatgatagc aactaagaac tcagtaaaac actccagaga aagtcacaaa aagtctcgag tggcccagag atagagttag tacaaaagga attatgtgtc agggtaaaga atagcactaa aattgtacta gacgactcag ttttggtgag taacacataa atccttgaaa gtggacttta acacatcaac accaagtcat tgataggccc taggtactta aaattacaaa ggaggttatc aacgcatggc gtgccttgag ttagatgtta tatgttttcc tagtaactct cagcgtcatt gtatgtgacc aaaaatgcac ctaaatgtgt aaaacatcat ctgggagaaa ctcttggaat tagggttatt aagcacctta agcatgatat taaaaaaaaa atgtgaaatt aagggaaaga tctttacata tttggggaag atcttagttc aaagtgtttt ttatttgtca gacagagatc aaaaatttac aattatcaat ttcagaaata tatttgtaac atatttccta acaaggttta tttgttgccc gcattcaagc accaccccgg actcccgacc ttaagtctct gtcattgatc ttggctgatt gaactgcttt cacctcnaaa tccactgtag atcaagagcc agaagtatca tggctttgtt tctattgtct gcacattctt aggtcttcta ttaatattat tttcctgcta ttgtcttttt atgaattgac attttgtttt aaaaattggg cattttgaga tcaaattatt ccttccattc atgtgttgtt tcaggttttg aatagtctga aagacttagc atattgtcac catagtatac aggctggagt aattctcctg gctaattttg tcaggtgatc gtaagtccac cattggaaga gcttccagat ngtanntttt atatgtcaga ttttaatttg atttttaagt gctgtttgtg tgtggttatt gcattttatg ggttatattt ttacatgttc attctgctac attcttttga ctttgtaatt taacactgcc gtttttggtt atttgaagta gttgtagcac attttttcct cgataaagaa ttgttatata atattagatt gcactctcag aataattcct aaattttttt ttgattgttt gcaatggcac cctcagcctc tattttcagt tgcccgcctc taagtttcag catgcgtctt ggtgctcttg gaccagncaa cttctggata catttccctt tactattttt atataagttg tgtaccatac tcatacagga atgctttggt taacttgttc tttactaaat tgcttccata tttatgccct ggtaaaattt tatctagaac tgtgtacatt aactagcttt tagtgcttgt aaatccctac tgttattatt tttcatgtgt aattgttttc - 103 - 008252 actttaaaag gggttccttt tattttttat ctcttgtcac ccaggttcaa caccacgcct ggatggtctc ttacanggcg ttgctctgtt tcttgggatc ccaccatgtg aggctggtct ggattacagg tattggggtc taattatttg ctaattttgt gtattttttg ttttttttaa aaaaatattt acatatgtat gtatatttcc tgatgttccc tttactttat gagactctta gttaaccagc ctcttttctc gagaatgtat gatatttatt tatggtttgg tttctacatt tgtatttcac gtttttgttc tttctagttt tcatattcga tattatttta cggagtacag ctcctgcctc tttttgtatt tgacctcagg accatgcccg tcatcatgat ccacacattg ttatcctgat ctgcctctgg tgccctactg tgtcctttgg ttggtagtca taccagttta tgctcagcat tggaagaggt gtctgtatct ataggttcag gagaatgcta acattctgtg gtgtgtagtg tctgggtgaa gaatgaaaca ataaagactt ctaatgataa acagagcagt taaaagagaa tccagtattt tttttttttt tttggtatca gcagtatttc tttttgtttt ccaggctgga gcgattctcc ggctaacttt aatctcctga tgagccactg gcccaggctg aagagatctc tggctgattc ccaactccca cgtgagtcac tatttggtga gtaacataga atttttttgt tttgttccat aaagaatcag ttattatact acatgtgcca taatgctatc cttccntgtg gggtctgttt aatctgctgc agcataaaat ctgaatacgg gttatctttt ttattgcatt tctttccttt ctctttccct tcctctttct tttctttttt tgtgattctt ttatagtctt tttttacttt tggtgcaatc agcctcctga tttagtagag tgattcgcct gcctatgtat ccgtttgttt ggtagagtag gggtaaacat actgctggcc aattgccatt agtttctaat tttgtttttg catttctaca tccacacgta gggcatcctc tccacaatgt aaaactggaa acatctctgt tagttgcccg gttattgggg gcattttatg aataagtgac agatttcaca aaagaacaga gagagaacac agaaatgact tcacatttcc tattttcaat gttcccggtg ctatttcttc ttttttgttt gtgcagtggt tgccttagcc ttgtgtgttt ccttgtgatc tggcagacct gagtgtagtg ccacctcatc ttaaaacttt gactcaagcg agtgttcagt agtatatttt cctgtgtaaa tattttggtt tttgttttag cttaattaac ttaagttcta tgttggtgtg cctcctccct tccaagtgca ctaacttttc ttccttgcct gttactgctg aaagagactt tgttgatagt gttttcagga cctttgtttt tgcttcactc caaatacttt cagtctctta ttggagcttc catctgcttt tattattttt ttggctcact gtagctgaga atggggtttc gccttggcct tatttcattc ggttgttctt cctctgcata ggagcttttc caattatcga ctaaacatgg attccattgc ttttgttcct aatgttgcca tttattagtg catagcttca ggctaggcta cctgtctcag tattcacagt gccctagctc ccatggcaac attgaaattg atgttcctac tcactaaaat ggcatactta aagttttaat taggaccaaa atcaacattt ctgtaagagg aaatcatctg tatggaaatt ttgtttttgt tcattttcgg tcccgagtag agtagagatg cacctgtctt atgtcttcat gtgtgatcat ctcccatgta ttttagagac atcctcccac caaaactgtc cttaggaaat tagcttttct agcattttgt gttagccaat ttattaattt gggtacatgt ctgcacccat cccccgaccc tttattagtt gaaggtgttt cgtggttaaa agaatgtttt tccctttatt tacggatata ttccttttgt ctttccatgt ctcatgctgt tttcatttct tcatttagac tatttgttta gagtgtttat tgagacagaa gcaacctcca ttacaggcac accacgttgg cccaaggtgc atcttattct cttgttgttc ttgtcagtac tgtgtaccga atatttctta ggctgtggtc ctctgatctt ttgttttaca tttattttaa attttggggt atgaacagat gtttttataa gaggccatct actcccctgg actagagggc ctttgcccat ccatgtgttg aagaaaactt gatagtaggc atatataaat aatgtaaatg tcgttttcaa gggtgatgag tgaaatttaa gtcctgttgc gtctgtttta tttttcgaga ctcactgcaa ctgggactac cggtttcact ggcttcccaa tttttagtag agctcactgc gctgggacta agggtcttgc ctcggcctcc tgtttcagac tatcccctat ttcctctgtt tgtcatcagg tggtttgctc ttgtttaatt gcataacatg taacttgtca catgacaggc tctttctttg gtctcacatt atgttaacca ttttccctcg gttcaagtca ttttcctagt attctaatta atttccctct cccctcactc ttttcatcat atcatttctg aatttctttt tttctgttta cctcgctctg ccttctgggt gcaccaccac ccaggccagt tgggattcca gtaatgtttt acatgaaatg gaggggtccc ccgtcaggtt cccactggcc tggaaagggt tctgcagttt ttaacctgca ggttcacctt tttggaaatc ttcaggatga gcactaggtt tgggagaaag acgttgcccg catgaatggc gcattgaaag acttcttaag ttgttttttt attctgctag tatatactca ttagaagttg aatgctgtaa ctatttacta aatgaaaatc gtttttgtgg tttttatttt tggagtctca cctctgtccc aagcgtgtgc gtgttggcca agtgctggga agacaagatc aacctccagc caggtgtgtg tatgttgccc caaagtgctg ttgcttggtc ttattttgtt tgattttctc ttggctagca attttgttga tttttccttt caggtttgtt tttacattag cccggttntg ctttatttgg ttcatttctt caaggataat aagtggcttg gcagctctat ttcatcatgt taccaggatc gtttttatgt ctctgtcccc ttcctgaaca tgttttctta ggtcatgttg tacgtttatg tcacccaggc tcaagcaatt acccggctaa ctcagactcc ggcgtgagcc tacaagggct ggagtttttc tccactgttg ctgagctctg ccagtgtccc ttttgttggg cttggggaca acttgcagcc ctgagatatc tactggttta ttccaaaatt tagataangc ttggtcttgg gccctagctc cctgtcactt atctgaatat ggttccattt aagtcattaa ttgtcacagt ttccaagtga aatcaatagt attagccatt caaccaaaaa aaataaggtt - 104 - 008252 ttctggtttt ttgcactgtt attctatgtt tatcctgaac tattgttaga agccacagtt acagctcaaa aataagactt attttattag aaatttgtac tcaactctac tttttcatac ctttagtccc taagcttgta tcttcaggtt atgaaatgtt agttattgag tgatgtacac ttttttttga aatctcggct ccgagtagct tagagacggg cccgcctcgg tgttattttt tatagaagca cctagccaat tgttatccaa attttttcaa gacctagtaa atatgaaaat aaacccaagt tacaatgagc gtcctcagag attttcaaca cacttctctt tttctccaaa gcaaaataaa tggatgtttt aatacattaa aaaaatggca tactttgact tccagaattg gataaaatgg ggggaaaggt acaatgctgg actatacctt tcattatggt aaatagcaaa aaagggcttt accaaaactg tgccttgaag agggatctca atgccgagct ggagtttgtg tctgactttg atctgatttg gagaaactga aatgcatttg gcccatcaat ggaagcattt tctgtgttgt gttgtctatt cattcagaaa aacattatct taaatgctag caagctttaa gggtaataat ttattatttt gagtatattg tattttagca ctttgcattt atattgtagg cclllcaggg cataattttt tttttctacc ttncacccag ttttttcata acacttttca taatattcac tgagcagtac atatgccagt gacggagtct cactgcaagc gggactacag gtttcacctt cctcccaaag taaagagtct aaccaatgta tttggcaatg tgatacctag aacaaacgtg tttgccaaat gtttaagata acagtttcca tcatctctgt aaaacctggt tccaatatat tcactaattt tatcactgca aagcatttaa atatgtgccc agtaccatcc tcacttcaca gtaagatgta tgcaactgtg gcctgggttg aagacagagg tacaagattg tggcagaatc ggttcttcta tattagtata aatagtggtc tctgcttggg gatgagtaag cggagaaaga caaagtagtc aaactagacc ttctgtggtc tgtttgaata agcagaaagt ttttaagtgt tcccactcct tcactcacca tgcagagcac tcacatctgg taattggatc tgaagtagtc ctaacgtttt atgtatcaac tatatcagta tatactacag ggcttgatat aaatgtcctt ttgtatttgt tgtttaagtt aaatacaact agctcaactt agttaaatat tcactaaatt agtactgtca acttcttcaa tattttctta tgtcacatta aaatttgttt acatcataat gtaatgcaat catagctcag agttaaatat tttaccatca ttttcatgat acatgcatgt tattcaaaac aatgtatgtt tgcttttata aaaaacacac aaaacttttg ttaatacatt aataatttgc cataaatagg tagtaagttt tgatttccaa ctgttagcca gatttgcgaa cgctctgtcg tccgcttccc gcgcccgcca gttagccagg tgctgggatt tttccatgca ttttttgccc ttatggtgta tttataaaat tcaaagctgt atattaaaat tttaagatat agttttcctt ccttggagaa tcactctgtg ttctgaaaac atatgggaaa ttaccatcta aacatccctt aacaataaca accatatttt gattcacggg agattttaat ttggagcagg gtaaactcaa aagatatcaa ggcccaaaag cactggcaac ggttcatagt atgagaatct tgtaaaagct gatatcagag tgtggtctgt aaaggaatat agaagtggca tggctgcatc agtaaagatg gttcactctg ccttccagca tttccatctt caacagcttt aatccctaga ttcatgcatt caatagatac ttaagcactt tgccaatagc aaatatgtgc aacgttatga tcacattgtt ctaaaatatt tgtgttctgg atgctaacct cccaggccgg gggttcacgc ccgcgcccgg atggtctcga acaggcgtga taaaactatg agctgtcatg cctaagagtc tatgcacgac ggcttcctag atgtataaat aaatgtaata tagtcatcca gggaaacaga ggacagactt atggtggata actaataatg atacctatac acccaattat ggaagtttct attgaattgt tctctaagaa tgaagcatca gtagggtgtg tttttggatc tgcactagta gtgaacaaca tttaatgtct gattctgggg acatagagag atgggggagg aagtcttgaa ggcaggggtg gaggcacaaa gatgggctgt atggagggcc catggaggat gaagttacat agaaatgtct ttataatatt cttctaattg ggtgaaaaga gcagactgtg aagtaaaagt gtctgctgaa actgcagtta tgcattttct atatacatgt tggaacagag tctaaggaat gtggaaaagt gntanttttt actgcggact cattctcctg ctaatttttt tctcctgacc gccaccgcgc ttagtccata cacagttctt atacctaatc aaaccaaacc cattgcctac ataaatataa taagaatgag gcaaccattt caaaaccaag agcacataca gcatattttc tattagggga aagtcaacct ttgcactaat gttattaagt tgatatcatc ataataaagg attagaagat ttttagaatt ttttgaaagg ttaaatagtt gagatacacc ccattggcac caaacatttt ggaccttctt caggcataaa gaaagaggaa atggaagtaa ggagtggaac gcacaggagg cagcaggctc tttcatctga gctggatgca ttaacaattt cccaaagctt cctccctaat aactatttgg agttctttag agcatgcatc tttagtaata taaaactggt gaaatctgtt tggcctctat attaaatcta cataacagat ttaaactttt cctaatttat cttacatgag tgtcacatcg waugCwtuCt aacctcatca tctctcaatt cactaacatt tctattttaa ttgtgcaaat agtcaaattt gtgtgtgagt tttttttttt gcagtggcgc cctcagcctc gtatttttag tcatgatcca ctggccaacc agaaaatatt taaaattata gttaaagtag cagaaacttt taagtttatt aatatgtaag ttatattctt attgaacact cagagctcct ggcactcttc agaaaatagg gctcgactga gcatatttct caaaccacag aaatttaaaa aattgtaaaa cttctaatta gcatgctgaa ggtgaaatat ctaactttca aagtaggtaa tgcattgcac acgtgcaaag ttgcagatat tttttttatg aaagaaaaac cgtgaaacaa gatagaagta agcaaggtac cagctggaga tgcaaggagt gagtggtaca cagctgggga ttgttggagg gaaggtgctt tcctcttgct atgatacatg accattctca ctttaagcac - 105 - 008252 tgttgaagta acaataaatc tatccctata tttcttacct tacagctctt aagcactctc atttaaggtg gcatatttta aaggatctgg tctgacgata gtgtcagatt tagcaattat tttacaatca ttnagttctg tctcatctgt gcatataaat ctgacatctt agacatggtt tactgggtgc catcttcaga tgtaactcca gatccaataa ggcatattct ttgttgcatg ggaaaggttg gaacattaaa gtatcttggg aacatgagcc tgggaaaata gcttgttggt tccaaacaaa atatataatg ctgttctcat aatgtgtttc tatcgttgat gaagcactcc tgccctccat ttaaaatatc gaattgggtt ccatgcttat atttgataaa tcaaggtctg tgataatgat tggtnaatgt cnttatgcaa gatctttcat aatttcagtg gctgtataca catgtaccta tgcaaacaca atacactcaa gagaatacgt gttggagaga gcgagtaaca acgcctgagt gcacagactt aagccaaaga tcataactct aaaaagtaat aaaagaggtc gacacaggaa acaattaaat atgctaagac gaaactgtac ctttttctgc tggcagtctt tctcctcagc ttcttgttga ttgagtacca gcaaatatta aaatggaatc tgtgccagat cattctactt taatatgata ataaaatatg actatgctaa ggttacaaag aatagatgga ttgtatatta tctcatcatt accnttaaat acatttacca tcagagaaga ccaaatataa caagatgcac taaggtgatc ggcagaattt ttttttacaa gcttataaaa acatgaaact aattctgatt tgccactata tacttcctat atagtgacag cagtgcatat ttgttaaata caagtgttta caccaaagtc tggaacaaac ctgggcattc taacttgagc aaagcaactt acaaaacata atttaaaagt gtaattttaa taaaccgtaa tccaggaagt gagccagaaa tggatggatg actgacatct catttaatat tatccgactt tgagagatag tttaagtagg attcatgaaa attatttata agaggaggct ttcgggaaga atggaaaaat gatgtttatg tatgcccaga gaccactgca tgtttctgaa ggaaaagaaa agg ucgu ucg aaaatgattt acaacattaa atagcttttc actggagaag atttttaaaa .agcatgtctc acacaaaggg taatggtgaa agcaattaaa gaaataaaat ccgtgtactc cccaatttta ttctttgcct gatacttttc gtccatttga tgaatatttt tttgttcacc gctgagggaa tttaaataat agtcaatttt ni- rrt- rrraaf л --~J w~~ -w ¦-- acacaactat cgttatttgt agcatcggta tgataatcac agtttgcttt gattaccttg ctaaactccc ttttgcttta ggttgttggt gcattatata atgggccctg cattttgtca cttgcatgat ccgttataaa ctgatttttg aaattaaaac atatgtgtta ctcaatacag tgactactat atccaagatt attataaact ttcctaccag nctgtatttg atccataagt ggaataattt cattcattaa tgctgtatac ctgctatatg aaagtctcag acttcttttt aaactcaaac taaccaatat tacatgagct gtagtcagat tggacagcat gctgagggtt aatattgaat ggattatttc taccatctag aagtccaagg gttttctaca gaaacttggg ctgcaaaata atgttgttac aattgacata cgcatagctc attttttcat gcttctggtt tagaatttta ggctagggct gtattgcaaa tattgtgcct ttcaattgta catggcatgt catctcataa ctttttagaa gaaatattaa actgtatcta tcacccactg ggtgnttact ttttttctaa ttcaccatca ggattacaaa gagttgtcag cactctggaa gaaaaacaat gctggccttt gactcttgga catacttcac catgtgctga cagaagacat ttacatggta ^aa " u u u uy \~ uacay vvay u u LLtauaouLij ttctaatttc tgatcaccac tggtgcttct atagtttgaa atgtagtgtg ggccagtttt gaaacaaaaa ttcagtcaca tctgttattg caatttttaa atttaaattt atttttgcca tgtccaactt aaaaaaaaaa ttgaaaagct tccaaaaaca tatcagaata cattgtgaaa gccattcact tgcccgcccc cctccccata atttgttaac attgtaaatg tacctcattg gagtttaatt catttctaaa gcaaattatg gctacatata acattgttaa tgacaaaaat gtacaatata aattcatcct atttgggtgc ttcctgccta atatgtactc attcattgct cccataaaat ttcagttttc gtttttggaa tctgcctagt tcttaaaata aagatgaagt aagtaaaaaa catttgttca tttaataaaa cttcatgaca cttgtcaaca tataacataa tgtaaattgc ttaaagacag aagcgtttgt acactgccta ggttaaaggg atgcttatga gagagggagg aaggaatgaa gggcacaanc tgggtattaa tcaaatttag gctcagaaaa дт-т-1-дгтлИ-д +¦ да(*+ a a t-+- f- arar't-htraa tatagttaaa aaagttgaag tgcgtttatg tctcttcctt tccttcttgt tcccaaaccc ccgttcattt ctttcccacc acatctgttt tgtgtgtgtg gatcctctgt atgactataa tgtagcacat gggtataaat cacccagagc aggagtcacc gggaattaac atgtttatag ttctacaaga tgtgtgtgtg gagatcttta aaaatcctta aaacaccagt cttcatatga caagcttcag aagtttggtt attggtttct ctttaagtta ctgttctttt tgttagctag aaaataattc tttccttagc attctgaatt tgattttgtt atttaacatg tcctccacct aagtatcact ctctcattca aagagggtaa agtaatactg aacgatctat acatagttct ttttaaaaac tttttttttc aaatatacac atattttatt ctcgtatccc agaaggaagt tcatcttcta atatgggcta tacnaggcaa taaaaaatgc tatatatgta gcatcctaag acaatttaat catatctctt cccctgaaaa catagaatta tatactagct ggtggtgatg tcctctaatg attattttat aaagaaatca ggtgaggccc aaagctaaaa taagcattgg ttggctcctg gtttatcctc atgttccacc ttgtagtgct ttttgaaaaa aattactctt ctgctgagtc aagtgacaac cctaacacac ttttattttt tcattcagaa tagttcttga aagggaggga ttanccttct tgttatctgt rrt* at- nrat- rrr-h ~D---~D--~3-- nactataaaa acctttgtga atgcagcttt aaggccattg ttgtgagtct cattatttgt atcttaccat ggatgtgttc gtggtgtgtg taaaacaggc - 106 - 008252 acttacccag taagtgtttt aaggagatat acatgtttta aggtttaatc gattacagaa agtttctaat attcccatgt ggtacatctt ttttccttgc tttttattac tgagagatgt gaaaaaatgt tgtagatgtc gtcaaggtgt agaacttgct gggtcttttt cacgtgccat atgctatccc tgtgtccata ttttctgttc tcaagaaaca acattttctt ttgtgaatag aatcctttgg gatccttgag aacagtgtaa ttttaatgat ttctctaatg tgaatgcctt tgttgaccac aaaaatagtg ttttataaat aaagtctgtt attttcctcc ttatttttag ggtcttttgc gcaggtattc canaaatata tattttaaca tttaattgat caaggtgtaa gttagaaaag ctcaatcaga tatattccct attgaactta aaatactata agacttacag gacatgattt tgttttgatt aattgtacaa tgttcattaa atcatagtgt ttggatactt tcatttgatt agaggaggta ttcatgattt ttttaacctc tattaaagca tatctttctt aaattaagac actgtggtcc tttatgtctg atatatatat tatcaggtgc tgaaatctcc ttataaattt ttattattat ggtggtttgc tctcctagcc tgttctcatt ctgtgttagt tgaactcatc taaccagttt tgctgcaata gtatatatcc gaatcgccac aagtgttcct caccattaaa accagtgatg gttagttttc agtgcactct acaatggctt atattttccc tatttttctt ctattatgat acagattgtt ggggatagtt ctttgggttt tccaaaatgg tatcaaatgt gatgtgatta acgtaatact tgatcaaatc tcaatagctg aacgaaaatc gaaatgaagg aaaatttaaa ttgcaaatat gacagatttg tgccttttaa tggttgtggt taaattattg catcatagtg aatttaattt acattaatgt ttattttgcc actttatttt catacttact agacaggnaa agatagctat tggctaaatt ctataagcaa wt-wUdwU-acv tttgttgata catcactcat gttcttttcc tgttagtaaa ctagatattc atatattcta atgaataaga agcttcaaaa cagagtagag tgctaaggga gttgagttta tagttttgca atttgttact tgaaatcttg atgatatatg aatgggatag atgaagtttt ttctaatgtc ttacatatgc aagtttcttc aagagaatgg attaagtgtt atgtgtatat atttgatctt caatattatg gggtgttcct aaaaaatttg tgcaccatca ccccacctct gttcgactcc ttggctgaga cttttttatg atcattgatg aacatacgtg agtaatggga actgtctttc agttctctac ctggcgtgag atgagctttt ctactaacgg tggttcttgt u -у у - к.V.U u - tttgcatgaa ataagtaatg acaaaaattg ctttaaaata cctatgtgaa ctttggtgct gnaattattt ngattgtgta gtctgtttac tgtatgtatt tgcataatta taatatctta aatactatag aaacaatgtg gtctagtctc taaatgcttt attggaaaat ttcaactaaa aagaatgcct ttaactgtag 333 v_ u С 3 Cf S 3 agctttcctt atattttttt atttaataac tagtgcaatt aaaagcctcc agttggcact ttaatgttgt tatactcaaa gtgcacctac aaatccaacc tacttcattt tacaccataa aaaggtagag acattttatt tgactgaatt tcctaaaaaa ctgttttatt agaatagata aatcccaatt ttcacttttt ttgttttgat gatgttagaa atacatccac tgggggtttt cagaactgta gtgtaaagtg cagaatgtgc acctgtcatc tgacaggccc cacttatgag gtgatggttt gctgcatagt ggcatttgga tgcatgtgtc ttgctaggtc acaatggttg atcctctcca atggtatctc tttcttcttc tgctaaacct aggcagggag aagt.gut.aga ttaaatattt ctacttaaaa tagatagctt tttgctttgc agatcgctgg acaggtaatt tgcattaaaa attgcctaca tctgacatat tatgggggac gcaattttat attattgtca aaaattatta gagcaatatt cccatatttg attataagtg aagtagaatt gttactatat aacatgaaat gtacaatgtt tcatattttt aatcagcttt tattatgtga cttttttaat actaaagaac tcttgtaact tcttattagt agaaagtgga ttatgctgct tcatttaccc ttgggtcaaa actatttcca gcttatgtca tctattttca cagaattgac gcaaaggctt attccaaagg ccctattaaa aaaattaaag tttgttagtc ttcagttatt tatgtgctat atatatacag tgccttaatg agtctctttg catatatagt agttttgtta tacgttaggt gtgtgtgatg tgagaacatg ccagcttcat attccatagt ttgattccaa cttgtactag aaatggtatt aactaattta gcatctgttg attgtggttt actttttaat agcagtttta ctggcttagg atttgtgtgg tacctgcaaa tgctaaactt ttattgaaat ctaataatgc tgtgtttttg aaatattttt ngtctaaata acattttnat ttgaaatttg tgagtgatat atcacctcaa gttatgattg gctgttaatc tctgtatttg gataggtatt atggggaacg aatttctgtg tctattgatt ctactgtttt gtacaggtct CaCayCCCat ataaagttca gtcaggttga ttgacaaaca attttaaccc gttattaaca aaaaactgtc ttatgtattt ttttggtgga aacatctgta atcttttaat taacttccat gaaacctctc atatcctgca tttccctagc ttacatggat ttctgtttaa cattttttaa aaatgaaagt tcaatatgtg ttgaattcta ttgcattttc gtgacaatga tggaaagttc atctaatact aacatctcta taggatagtt cacaggtata gtttctccta ttcccctccc cagtgtttgg ccatgtccct atattgggcc gtctttgcta aatgatttgt tctggttcta cactcccacc tttcctgact tgatttgcat cagcttgaag aagaccttcc acctacatag ctgagaaaaa gttgcttgtc attccttgtg aatatttgac catgtaaaat atggatttat taaattacaa gaaantatta tttnnatttt aaaaattaat ttgatgtata acattttatt tgaattattt agatataaaa taaataaaaa agccaacata tacattaaaa cattcattca tgatttgatc aatgaatttt ctagagttca cctctcccac attgtttctc tttcaaatta tttctctatt agaaagacag ttaacgaacg gtttctgtta aaataatgta tgttccatga ttgtaggtgt - 107 - 008252 ttgaaattgc cactgattat ttacaaatat ttttttgtaa atatggagtg cagatgtttg tatctattaa gaataacctc gcttcctcgg atggagtctc ctccgcatcc ggcgtgcacc aagacaggac gtacatttag tagagcttca tcatttttat aacatcattt gtaagataga tcttctactt cagagtggga tagagtttgg cagggcagca ataggaattt tgattcaaat gcagccattc tatataagga gaggatttct ataagtgaat tattttatga gacatacttt tgatatagtt gatccgagca tttaaagtga gttttatgta tttccatttt gaacatttac ctgcatttta tatgtcagat tattgttttt gctctgtcat cgggttcaag accatgcccg agattggaaa aaaccttagt ctgtggaagc tattatttac ctagactaaa atgtatgtac ggctgatcat tgttggcccc ggttccaagc gggattgggg gtctatttat cttagaaatt atcttcaaaa tcattgagtt ttttcataaa atctctgtga gtttgcataa ctagaggcat gatcagatgt ttccggattt gcgctgcttt ctttacctac tctacttttc ttatcactgt atctagttga tttttaaaca gtccaatttt gaggctgggg cgtttctcct actaattttt atgtaacctg gtggcaaaat ttttcatatt ttttgtcatc cactattatc tttaattacc tgttaatata tggatggtcc acaacccaga ataatacaat aaatgaagaa agatttaaag tatttttaat taaaaacatt gtctatatac gtatagacat attatttagg caggaagagc caccttgggg atttccgatt acaataagtt ttttgtttct tttctacttc gttgtaaatg ttaacaaaga gtgcttatgt tattttattt tgcagtggcg tcctcagcct gttttgtcca gtggaaaagt ggggagtgtg agaatgagtt attccctaaa tgttgcattt tattatgtat caactccagg agaagtgtcc angtgcaaca tgattttata ctcatccata cactttgtaa atatcaatat gaaagggtta ttgtatatat catgataagc ttactcctca aaatgaggga ggcattcact gtatgccagc agtaaccagc acatagtaaa tagagataac atggttaaaa aaaaggaaag agcaataaag taagagtgtg aaaggaaaac atctatgagt aactcatggc gtaactaaat taattctgag tttatttcac atttataatg ctcacagaaa taaatttaat aattgaattt ctaattcatg tatgcaactc ctaataatta tataatgttc taaaaactaa tgttgcacca ccccaaagaa tgtcaactcc aaaaaggtag ttcagaccag gaggcatgtg caagtttcta ggaaaatatg aacatcagta taatttcact ggttaaatgt taaacagtaa gccttaatta attttatttt tatttagaag gcatgcatct ccagaggggg ctgtgcacag tggctcacac ctgtaatctc agggtagata acctgaggtc aggagttcca gaccagcctg tctctactaa aaatatgaaa attagccggg catggtggca aggcacctgt gatcgggagg ctgagacaag agaattgctt gcagtgagct gagatcgcgc cactgcactc cagcctggac caaaaaaaaa aaaaaaaaaa angaaagaaa tttaccttag ataccatgcc ttccttgatt atccatagta ttatgttgaa tgttttttaa tataactttt atttttgcat gatgtgagtg ttttaaaaga aaaatactgt tgctactaac cccactccta aaatagcctt ggaaaataac tttttgataa ccattttctg tattattgaa attatgtctc tactttaacc actttattga tctttacata tttattatat atgtcttgag ggaactattt atccagagag taagttattg catttattat tctacttcct aattaacttg tatatttctt atgaggtact tcctgaaaat tttgtgtatt tcttatgagg tacgggtcaa gatgtagtct acatcggtat cttaggagaa gctctcagta tttccgttaa ttttcaaagt gaggaagtta cctttaattc ctatagtgcc gtatgctaaa ttttgtccag tgcttttccg ttttgttcag attgactttt taaatagaaa ccaatcttgc atttctggta tgtattcctc tttatacatt ggaagatttg atttgctaat catctttgtt cacaaggagc attattttta ggttttttaa tcttgaaatc cgggtaatgc tgtcctcata aaacaagtgg catgttgcgg tgaactgcca aagaggaaaa ctaggcgatc tttatggaat acacaaagcc tatctctgta cctctttgtt tattttattt cgatctcggt gccaagtagc atttttaaag agattaagca ctaacataat acatgttcgg atagtttcct gactgttgta tatgcaatag gatgtgatgg cattaggtgt gatatgacta ccttatattc aattaccgat aattcaaaaa ctacttacaa tgttgtgttt acaaagtaaa ttggctcaat atgaagtaga atgaatgtaa tgtcattatt aagaccaaaa tatgttattt gtncatttaa cacaaacagt aagtgaattt agattttgtt aatgtattca agcataatct catacctact agcactttag accaatacgg caggcacctg gaatccagaa cacagagcaa gttcacagaa gtaaagatct aataatctat atcttcatta gacatattat gattttggtg cctttttctg gaaaattatg aatgatcctc taacttagtg gtgggatgtg aagagttttt ttaatgtaat tattgctaat gtttttgtta attgaaatat aacatgtttc attccacggc 41400 aggaccagaa 41460 gtttatgaag 41520 gggagaacaa 41580 tttaggagtt 41640 agggtaagtt 41700 attttcataa 41760 ctgtgtctct 41820 tatttttgag 41880 tcactgcaac 41940 tgggactaca 42000 acaagtattc 42060 aatgcaaagt 42120 aacataaacc 42180 gaaaaaaagc 42240 aaatgattat 42300 gggttggtgg 42360 agcacctgcc 42420 caggagagag 42480 tagctagata 42540 tgccagattt 42600 aaatcttcac 42660 tgaaaaatct 42720 gagattacaa 42780 accaattggg 42840 tctgcagttg 42900 taaaccacaa 42960 tccaacactt 43020 ttaataacag 43080 agacaagcaa 43140 ttgttgtcat 43200 ^ /-> a a a a 43260 tttaaacttg 43320 tactgacaaa 43380 ttattagtct 43440 atatgtatat 43500 agttgtttgc 43560 tccacgtgtt 43620 gtaacttcaa 43680 attagtcaag 43740 tagaaattta 43800 gaggctgagg 43860 tgaaaccccc 43920 taatgccagg 43980 ggcggaggtt 44040 gactccgtct 44100 gtgtggacag 44160 cattttttct 44220 tcaaaattat 44280 ttattgttca 44340 tattttcccc 44400 aatataaaaa 44460 tcagaattaa 44520 atccttctca 44580 ctcaaattaa 44640 gttggaatgg 44700 agctgtagag 44760 gatcattgat 44820 gaattacatg 44880 ttggtcatat 4 4 940 caaattattg 45000 ctttaccggg 45060 caagggttaa 45120 - 108 - 008252 tataagattc gcctgaaatt agctatggag cttacctatt tcttttgaat tcctgacatt ttcttttcac ttttcatgtt ctttttcatc taatttaaaa atattctttc ttctttgtga ttgttatctt tttataattt gtctgattat gataatttgt acacacacaa gattttttat tttcataaaa atataatttt tttcttaaat gaacgttttt tcttgttctg gtcaatattg ctttttaaca acctttctct gaatctttgc cttcattatt tgaaggtata cctctaagca tgttgtatuu acctgtaggt gggtgcataa gaaattcatt gagaatttga accaaagtca atcagggttg attttcaatc gcgcataacg aatttttttc ttcaaatttt atttaatcat tttagaccat catgggctgc gcgtaaatga atcagcaact gaagtgaatc caacattaac tacaaatatc tttaaaaaaa ctcagcactt gtgagctatg agaagtaaaa ccactccagg gaatgcccac aacagataga ataatgaact aataaagcca aaatgcaaac gtggtgggaa ttcattatct tatgtgcagt gaagtagaaa ctattaccca gtctgttcct atcacactct gctttcagct aggcattagc tactgctttc atgcggagtt ccatcttgcc gtccttgagg ccttactacc gttgagaaca atgaaataaa aaaaaataga aagaaaattg atgcatggaa aatatgccta gagacaatca agtactgcag atgtgagagt aaagctcata gtttgataga cattacaaat ttaagttggt cataatgttt tctgtagaat ccattttttt cctatttaat tccttatttc tagacatctt ctgttttagc attcatgagt taaatgtatt aaattttgct atacaaatgc agtggtaata atgtcttctc gttctctatt tgctttttag ttaacatttt ctctaagcac aaaactcagg caacttacag tttcaaagag cctctgatta agccattttt aattgtatta gttagtctgt tttttgtttt tctaatacaa tgttttagct tatatgtgat attttctaaa ctgtgaaatt tgaattggtg gtataatatc tatccttctt attttctaga taattttcaa ttttatcgtc atgggtcaag actctaaaat ccacaattac acaaattctt aattaagccc ccaactggtc tgtatgtttt ctgcatcagt tttgggggaa caaaacacag cagtggcaca cccaggagtt agagtgagac cctgccacat ccatagaaag tgacaaccca atactgttta caaaataatt catttacata gttgcctagt ctttggggtt tatcaaactg tgcttaaaat cctgcactat aattccctgt tggactacct cacatggggt tcacatgggc tccttctgtt gttttgtatt ggagttgaat tttttctatt tgttatattc aactatttat atgacaaatt gcaggaaagg aattagttca gtgggtgagg ttagtgtatg aaaagatatt ggataagaaa tttattaaaa aggcattatg ecu. у u uya ua ttctcctttg aatgttattt teagectttt aaaattccat tataaatgtc ggtttttaaa catgacagaa ctttaattag atgaattttg aacatgacgt aatttatttt ttacgtgtac agttttagag agtactaaga agatgetcta ttaaataggt gcgtggcata tagaggatgg tttggcagtt cacctgtaat tgagactaga ccagtctcta ctcatttatt atttacacat aatttccatc ataataataa atgatgactg aaattctaga gataaaggtt tattgacatg tacactttcg gtctcagtgt ttattgtgtg ctatacgtgg etttgecaat catatttcca attaaaaccc cctactcatt gtgagcctaa ttggtgtagt tttgtctaag atctttttat tgaaggcatg gataaagaag agaaatggct tataeggagt gaagagacaa taggctgtag gtgtgggttg gcagctgtgt ccaagtcaga ctggcaaacg LLdLLdLLdL u i-'ctct u - .add uai.i_i_L.wUaa tatttagaag ttttattctg aggaatcatt aaagaaaaaa agttggctga ttctagcaat tctcttattt tgtcattgta ctaattagct tcagtaatca cttttatctt tcctggattg acaatcctga agaagaaact ctcattccac aagtggtgcc aatgagaagc acaactttac taaaaataaa tgggagactg attgegtcac atgaaagaaa tatttaacca agcaacttta taaactaatg attgatgtat gatttaaaaa ttatctttaa ttaagggatt tgactgtgct ttatgttatg tctaattttt ctgagtttag gtggggctct gaaagtcttt ttacatgggc tcccattcat cttcttatgt tttaatgeta acaaccagga acaegtatga tagtatccta tattctattt aaagaaacag aaaacaaagt aagcaagtgg gaaaatctgt gattgttcta atatggagat tgggtacaga tataaaactt ttggcacata tgcattttta ttgtggtcag ttaaggacca gtgaaatctg taatttgttc tgctgagaga ttgtcagttt tctttcagat tegggatgee taactctaaa attaatcaat acagtaaaag gcccagcctg cagtagtaag tgtcaaaacc taccaaattt aactaactcc agcaaatatt agagttgatc aggtgggatg tgcactccag gaaaaaaagt aaataaataa ttcatagtac atggtatatc acagcatctt gaggacatgt tgatagaaaa acaaaggggc tctgtacatg tcagttacac catatggtta tttcacctgt gttgtgcttc cagattacaa tcaagacctg aacactatgt tttgacattt tctatttgtt ctcacatcca gaaaggtaaa acacagaata agacagataa agaaaagtaa ttaagcagaa tgagtaaggg aggtagttgg tgagtgaata gaagaacagg ggctggcttc gtaaatgcag aatgatcgea gaaggatgag acaatagtcc gtatatggtc cagtttttga tgggtatttc gtactattca ttgetttcta gggctgacct agggtatatg gtaatgtgta tgatttgttc ggagcacatg gtggtgatga agagtgtgga ttggagattt attttgtcct tataagctat acaactcctt aagctgggta atcacttgag cctgggtgat tggacaggca aaatgtatgt tccaaactgg catataataa ggataaatct tctattattc gaaatcagca ctcaagaaac tgecaaaact ccactgaaac agcaattgtg ctcattcact actggtctat tctggcttta gtcttccgcc caactcatgt tggaatgcag tgtagtggaa tagctatttt' gaggatattg aataattagt tattaacatg gttgctaaaa agaagcagtg agttgactgt atttattttt tgaaaatgga gaaatcttct aaattaccgt ctagaagatc gaaattgaga - 109 - 008252 acagatacac gaatgtcttc tgaactcatg accctagagt tgcttcagta agtattcttt gctagttcca cgctgtattt ggtaccaaga aaagaatgat actgaaaaaa tgcatgaaaa catgccttgt gatctaaata ttagctccag ctcattactt tattctaaaa atgaagttca ccatgaaaaa tttcataaat gttgacagaa tgcttagtta cctgagacat agccattatg gtgaaatggc tgcatttttc tctttcccat atgtgtaaaa cctgcctagt agcatacaca cacacacaca cacacacaca cacacacatt tattttgtct tctttttatg tgactctttg tttactattt aagaagaaaa tgcttaacta gccacatggt agtaatgttg ctcataatca caactgttat agctttgata attcatttca agaatctatc aataatatct tttttgttgt tgtttttttg agacagactc tcactctgac acccaggctg gagtgcagtg gcatgatctc agcttactga aacatccatc tcccaggttc aagagattct cctgcttcag ccccccaagt agctgggact acaggcgtgt gccaccacac ccgggtaatt tttgtatttt tagcagagac agggcttcat catgttggcc aggctggtct cgaactcctg acctcaagag atctgcctgc ctcagcctcc caaagtgctg ggattaaagg cctgagccac catgtccagc cttattttta aaaaaaaatt tcagttcaat attactgcat tttaaagccc cacacataag cagaaaaact gtcagcacca ttttaccgtt actattagcc cagacataaa tgtagtttta ctcttagaga ctcaagaaat atccaaactc aaaataaaac agaaaatgtt aattgatttg aaaattttta aacttttctg tttccttttg tagcctctaa actatgactt tttttgctgc tataaagaca catgcacacg tatgtttatt gcagcactat tcacaatagc aaagacttgg aaccaaccca aatgtccaac aatgatagac tggattaaga aaatgtggca catatacacc atggaatact atgcagccat aaaaaatgat gagttcatgt ccttggtagg gacgtggatg aaattggaaa tcatcattct cagtaaacta tcgcaagaac aaaaaaccaa acaccgcata ttctcactca taggtgggaa ttgaacaatg agaacacatg gatacaggaa ggagaacatc acactctggg gactgttgtg gggtgggggg aggggggagg gatagcttta ggagatatac ctaatgctaa atgatgagtt aatgggtaca gcacaccagc atggcacatg tatacatatg taactaacct gcacattgtg cacatgtacc ctaaaactta aagtataata ataataaaat aaaataaaat aaaataaaat aaatttcaaa aaagtcaaaa aaagaattcc ctgtgtaaga acattcaaag tgaaaggagc atcataggga aaagacaaac cgtgaaagca gagaaatagg agacgccaca tcaagacacc accatgctta caagttggca ttttattctg aagaccataa tgaactgcca aagaagctga aacctaaaat aaaattattg tttttacata taaagatgat ttttcacatc tatatttatg atctcacctt ttaaatcagt atctactttt gaaaatgtac atgtaacctt caacatatac acagaaaata agataaacta atttatatag ttttgtttca atcctacaca attagagttg atttttactc catatttaga tttccagaat cccagctcat taatttaata tttattaagc ataaaagctt taagacctag aatgcatggg acgtggaatt aagcactagg gataaaagag gaagaaaaaa ataggtcatg tcctggagga gctcacactt gaggcagcca aagattaatt accttaggca ttgattagta ataataataa taattagtta gaaagtgtta tcagatgtca aaaaagagag aaaatgtgag ttttatagtc tttgtaaaga atatgacctt tgagcaaact ctagaattag gaatggagtg tttgaattgg aaattaaggc aggataatac ctttcagatg aaacaatgta atagagtact.aggatgtata atagaatgct ggccccaaac tgggatggta gatttaagtt gtgagtttct ttctacaact gcagagtcca ttgaataaat acaactttat atattaaaat taaaaaaata aaaaaaaata aaaaaagaaa ttctgtcatt tggaataaca ggaatgaatc tagaggacat tatgctaagt gaaataagcc aggcacagaa agatcaatac tgcatgatct cacgcatatg tggaatccta accagttaat tcattgaagt agagaggaca atggtggtta tcaggggctg cggcgagaag gagatggagc tgcaagggaa tgaggagctg ctgatcaaag ggtacaaagt ttcagctagg cagtaagagt aagttttgag atccatgcac aacagggtga caatggtcaa taataatgcg ttatatatgt caacataact aagaaagtaa atttcaaatg tatcatcaca aaaaagttaa gtgatatgat ggatatatta attggcccga tgtaaccatt atgtcttgta tacatacatc aaaacatcac atggtgctgc attaaagtaa tacaattatg atttgtcaat taaaatacta ttatttaaaa aatgtattat acacatctcc gagtcacata ttaagctaag gtatgatggc tgtgttttca gcaatcagtg ttaccagcta ctgccgtgac ctcatttctc ttaatcattt aagagaaacg attaagagaa gttgttaaca attgttccaa gttctgccta attgatttgc atgtattata gcctaatcat gacagagatt tcagaccaga aacttgcata tcttgggttt catagttgac ctccagaact gattgtgttt aatatgtgag gcaaacaaga taaagaagca acacaatata tacagttttg cttttaaatg gaacttagtt cctgtaaggc ccaatagcag gaaatcttag ttatgtcttc agtagcagta ctagtaaagc tttagattct tctctgttaa tgaatggaga cctaaggcaa ttatgaaaag agtaatgaga tttgactaag caaagcagct catttcctaa agaaagttga ggagagcaat ggcttgtagc agaaacaggt gttgtgaaat accaactgtg atggcagaaa tctccccgaa gctggaacag gaatgtttaa aaatgagctg gcaggttaaa tagaaaagat gaagacttac ctgacacctt attatcattc ctgaaaaaga agggaaacat attcatagct cggtatcaaa taatgaaatc ttatttccaa gagtgagata attagaaaaa ctttaaggta agtaagtgta accattaaat atatgaaagt ttaatataga gagagcagga ttcaaacagg aactcaggaa catattcgcg atggctggac cactagcttt aagcccagag gttgggtgat cacacatcac agatgcccag gacaatcttg gcttatccct actgtttcaa agtaatcact - 110 - 008252 aacagcctgt tgcccagggg agataatata tttattaaaa ctttagggta gaggtttttt caacatgaat aataccgctt aaggcaacaa gaaaaactac cgctcgcatg aataaaagtt ttttaatttc ccccaaattc atatgtttat attgatgtaa ttttcttaat aattactgtt atctgactgt cttccccttc ggcctaaata aacataatga atggctgaat atttatccat agtcaacatg acccaatagt actgtttttc gtttctctac gtgacatgat catttttata ttttgcccat tatattctgg gtgggctgtc tgcaattcca aaaaatttgc atagattcag gaggtaggag gttgaagaga taaaagcatg tttaccagta gtgtgatgcc tgtgttttaa ttttgataga tattgattct ttctttcatc tattcctagg atttttcaga tgtatcctgc ggattttcta ccaatttgga agtatgttga aaggctttca ctttattgtt acatgttgaa ttcctggttt tcatttctgg tcaagtctaa tctacattca ataatatatg ccctatgtat tgatatattn ttgatctcca tattaataaa tattcgattt aagtttatag aagttggatt taaatacatt cttggcaaat catggaatgc gaagctggag gttctcactt cagacactgg ctatcgggtg acacgcagtt aaaagaaaaa taaggacttt atgcaacggt tgacacaata ggcataataa actgggaaca tactatagtc atttttgtac tctggtaaca tgagtgagat cctccagttc agtattaaca tggtgggcag tgagtgctgc gaaattgctg acagtggctg attctcacca atttcactgt tacctacttg ttcttaatca ttattaatcc acttcaattt tttatctatt ccagtccagt ttcttagatt cataatttta ctgtcctttc tatttatttc ccatgctggt tccagctttg tataaatttt gattgcattg tccaattcat agagtgttat catttttaat ttgttcactg aatttactga agtgtaagat tgtcgtttat ataaatctga atttttccgc ttgaggtata ttttaccaaa atttatgtaa gaggttaaat cttccaagcc taaacctata tagtataatt atgnttatgt atatattttt agatgtgaac gtaaatataa taaggaatct aacttggtat tttttgcagt aaaatttatc aagtgaagta tatgcagcca gccgttatcc atgaatgaga agcctacctg ctatgcttat caatgtagac tttataaaat ttgctcttgc aactaaatta ttgtcttata tgagaaagaa taagcaaact gctaaacact agggtggagg tagctgagtg acaaacctgc tggaaacata agttttgtag ttcttgattc atttttggag gcaattggca tctctactag tttatcaaac acctttcttc tttctatctt ttgtctttct gctgcaaacg gtatatatac tccgtatttg atatatagat ttgttccatg cattaccacc tcttttttat tgcatttccc gtcttctttt tttttcccat gaatcatttg tcctttgctg ttgcctgtgc atttacccaa atcctttatt catggatatc .ttgatacctt attccattcc tagctttgta acgagattgc tttatattcc ttgcttcggg tatctttcca gtagagatct atattgtaaa taaatgctat catttctaac gcaaatgagg tatctgattg tgtcctgatc atgttangcc actcaatttt gcatctattg ttattgattt gatggtaagt ttaccatttg gatccacatn aaagtttttn tcgacttnca atatatctct gccagaaacc cagaataaat aatgcatttt atcaccaaga tggtccaaat aaatttattt aattttgttg tgagactatg aacacaggaa gagtacatac atgagaggaa gcgaaacaat acatgtatcc ctggaattta aatatgtaaa ctatgtattt tacatatgat tatccatcac ctattttgaa gcttaggtct atccaaccct cataagatcc gtgcttggct tcaggatttc atatcacatt gctatttgga ttcctccctt tttagttttt aacagtgtat aaaagccatt tgatgattag gagaaatgtc tgagttattt cagacatttt tgccgaagtt ttttcagatt tgttttcttc atttgatttt catttttccc tgtcaagaat attgggatat gcatattttg tttggctatt tgtgaagaat tagtgttgac ttttttgtgt ttcacttctt tggcattgct taatttttgt agtttttggt ataattttat ctctggctag ttgttccaga atgggtttgt ttagtgtttt aaataataac ccatatgtta aagaaagcca agaaacttct ccatatgtat attaangtct tctaaatnct tttnaaaatt agcctttcat tgaactttca atggtcacct gttgaattag acattttaca gtcacgtatt acaaaataaa tcctttgtag gagaaaacca agacacaaag ggtgaggact ctgtacgcca ctgaatctaa gagcattaat atcataattg atttatttac atttcgatac ctgnaaacat atatgtaata tatttatttt ggcctttacc acttacttaa tatttctttt actcattttt ttctttatcc atagtgctgc ttggatatat ttcagtccgt gagtgttcct ttaacttggg tgatgttaac tattcagatt gagttcttta cccttattct ttgagcttaa ttgccccccc taggagtttc tatatgatga agcaccattt gaattggctg tttctgtttt aagtcagtta cagaatcttt gtcattggta attttaacaa ccacttcaat tagtcacaat ttcttgattt atgttggttt ggagtcttta ttcttccttc aacttccagt tcttagcaga caaatatggt gataataaag atggtttttg agccatcctt ggattttaac gtattttaat aaactgtttn ggtttctant ataaactagn gttctgcaca atgataataa gatgaaaatt gaaatacaaa atcaagagac ctatgacgtc gatgtnaagt tcaaatcagg tttgaattat accctactga ccattcacca accaatctac catgtgatat tgtccatgca ctgtgtgtgt ttaggttgat tatctcttaa gatcatagag tcagaatttg gcactcactg agcttttatt ccatttgtat gattattaga tttgtcagtt gttgattgtt tttgctttag gtcctggagc taagtcttta ttcttctgca cttaatgttc tgggttttct ttggttgcta ttctttttgc aggatagttt aaactgtaga gagcatcgaa agttttcatt tttttaataa ttggtgtata gtttgcttat catgtcgtct ttctttctct tgaaaatggg ctttggtatg ccccttttat tgctttttct tgtatcatgt aatatgtcca tatgtacgtt gacatataga tnctnacatt ttcatgggat agtactatct cacctgaggt gaatatattt - 111 - 008252 aataccagca aaagaaaacc cttttgatca tcttattttt aatacttata aaattgaagc aagactagac tgtatgagat ttcctttagt agtggttaat tgaacatcct gtttactgta acccaaccag gcaccaagag tcacctggca aggcaaggct tttgcaatta aaaaaaatag ctgcccaagg ggagcaccag ttttctggga ttctagagat gcattggcac tttgtaaatc ctcatgcctg agttcaagac agctgggaat tcgttggagc gcctgggtca agagagagag gaaaaaaaga gaaagaaaga cttatttctt tatttttcag caataaactg cagtttctga attttgtcaa tgtaaacaaa aaagaagggc cctaataatt taaccaagtg cacacagaag taaatttaac tgttctgtaa tcctaataat ttctactttt tactttaagt gccatggtgg tagctatttg tgtgttgtat tttctgatcc caaaggatat acattttctt tgtaaatagt ttcttttggg atccttgatg acagtgtaaa tttaataatc ttccctagtg cttttgagaa tcttgcaaat agattgcaaa ttcttttgct tgatttttac atgcagtttg tatgtttact agtggtaaat acaacccgat tgagtaaagt ccatttctcc aaccttcctt tacaaggagc tacgattgct ctgtgagtgt gccgcctcca cttggttttc taaattttng aaagttgtaa gagattctca aactccaaaa accttgagtc gtagcatgcc aggtaattag cggacaacca tgaattgggc ttttaaaatc cattctttca taatcccagc caacctgggc ggtactgtgc cgggaggtga cagagtgaaa agagagagag gagaaagaaa gaaagaaaga ttattacatt atggaaaaga gagatcgtgt atttttaaca aaattcagta gatgagagtt agctgccaga aagagaaaaa aatcataaac gacgcagcat cataaagaga tttgtaatgt ttttttcaga tttcctgctt tgtgggatac tttgctgcac tcctaatgct gttctcattg tgtgttacgt gatctcattt я nl" с гя rrt' r-t" -------I--- gttgcaagaa tgtataccca aatcatcaca agcgttccta gccgttctga atcagtggtg gagtctgttc ttgtttacat aattttctcc gtgcataagc cagaatagca aatgggggtc tcccaaaaag aacttttatt aagttatgtt taaacaaagt ctcacactgc caacatgtgt ctattcttaa tattttaaga ggaatgagga cagaatcgat cccatgatag aaaattaatt aaaatactga gaattggcca gatatgttat ttcaaagaaa tgtcctcttc agcctcctgt tctcaatttt tggtgtactt cctcactgac ttgctataaa actttgagag aatatggtgg acctgtgatc gggaggttgc ctccatcgaa aggaagagag gaaaaaagaa aagaaaaaga tttgggtcct aatattgact taatgtagat agctcaccag agtggaagag ttcatttacc ttaaatgtga taattaactc attaactgca cactgctgtg catcagtttt tgattttaag actttctggg tgtatccatt atgggcagaa ctgtcaaccc ctccctcccc ttcaactccc ttgctgaagg aaaatgtttg taaaaggaaa aagataatgt gaacaattgc ccatttttac ggctgagatg tttctatgct gtccaaaatg aagacaagtt ctgaattcta gctctaccta atttactatc aaaccataat tttaggatga ttccaagcca ggttagaaat agagtagaac tgttagtctt ccttaagact ctttcctgca atttatttag tgtggttgag tttacttttc gaaatacctg gtcaagacgg aaccccatct ccagctactt agtgagccga ggaaggaagg gaagagagga aggaaangaa aagagaaaaa atatttgtat taaaaggtac tattttttca taatgtcaat cctgtgtttc aaagacagat tggtttatgc tgtagtgcga ctaggacaaa atattgcccc atgcgaagct tagtctttag taataaatgc tattcacttg tgtgcaggtt atcatctatg ctgcccctca acttatgagt tgatggcttc ctttttatgg ctgcatagta atccttgatg acatacgtgt gtaatgggat ttgtcttcca tttctccaca ctggtgtgag ttgagctttt atatcctttg ttcttatagg tattctattc tctttagtac acatgtatct tgctgtgtca cagtggttga tcctcgccag atggtactgc tttcatgttt ttcacttttt ttgtggatat tgtaggttgc ttctagttta tgagagtctt ngtaacttat aatattatag tgtatgttat actgtatatt aaacagcttg ttcagttgta tttgaataaa ctcagtggga aaggcatgca tfcctgtccct acttttgtat aagtatgcat aatctctttg catttgtgat caatcgttta acaaaattac ctagctagct atttccccag aagaggcagg tgactcttac tgtggttgga gtgttcacat aaactggcag gcagatgact ctactgaaaa gggaggctga gatcatgcca aaggaaggag agaaagagaa aaagagaaag gaaaaactga catacagaat ctctcaaatc gaagatctgg gtttttggcc tttttctagt atatgcaaag acatcagctg cagtctgtca tttttatgan ctcaaaagta caagatacac caccctaggg catcctgtta tttgcttttt tgctacctag nttttcagcc ccccgcgaca gagaacatgc cagcttcatc ttccatggtg t nrtt-1 nr-rrt-n - - -л ----^--3 ttatagtaga aattttattt actaatttac catttattgt attgtggttt tttggctgca gatggggttg tagacctttg ctgttcactc attagatccc cagtatacca tttgatccat gttataagat tttctcactt acagccaagg tatcagaagc acttgttcag atatttagtt cgcatgagta ctggaacttc cacaaatagt ctcaggttcc aactacagca gtaaagaaaa tcgtattttt aaattttagt atggcttttt gcttactgcc aaatgtccaa tatgtctttg atgttcttat gtcgtcaggg cattggggag ggcacggtgg tgagactagg tacaagaaat ggcacaagaa ctgcattcta agagagagag agaaangaaa agaaagaaaa gactgggtat gagttctgta ttaatgtgta aagaaagcct aaagaagaat ttttctggct aaaacctaag cataaagata gagagctctt tgtgctgatg cattatagtc gtatctttca agcaagcatc aataagtatt aaaaaatttt gtatacatgt ccgcatgcat gaccccagta gctgtttggt catgtccctg tatatgtccc tctttgctat atgatgtcta ctggttctag attcctacca ttcttgactt tggtttgcat taaatatctt tttctttttt tcagatgggt tgatgatagt atttgtctgt - 112 - 008252 tttggctttt cttgccattg cttttagcat tttcatcatg aaatcactgt ccatgtctat gtcctgaata gtattgccta agttttcttc tagggttttt atggttatgg gtttaacatt taagtcttta atccatctcg atttaatttt tgtctaaggt ataaggggtc cagttttagt tttctgcaca tggctagcaa gttttctcag caccatttat taaataggga atcctttccc cactgcttgt ttttgtcagg tttgtcaaag atcagatggt tgtagatgcg tgatgtgatt tctgaggtct ctgttctgtt ccattggtct atatggctgt tttggtacca gtgccatgct gttttggtta ctctagcctt gtagtatagt ttgaagtcag gtagcatgat gcctccagct ttgttctttt tcctaaggat tgtcttggtt atacaggctc ttttttggtt ccatatgaaa tttnagggta gtttttttct aanttctgtg aagaatgtca atggtagttt gatgggaata gcattgaatc tataatttat tttgggcagt atggccattt tcatgatatt gattcttcct atccatgagg atggaatgtt tttccatttg tttgtgtcgt ctcttatttc cttgagcagt gatttgtagt tctccttgaa gaggtccttc acgtcctttg taagttgtat tcctaagtat tttattctct ttgtagcaat tgtgaatggg agttcattca tgatttggct ctctgcttgt gtatttttgg tgtataggaa tacttgtgat ttttgcacat ccattttgta tcctgagatt tgctgaagtt gcttatcagg ttaaggagtt ttcgggctga gatgatgggg ttttctaact atagaatcac gacatctgga aacagaaaat ttcacttcct ctcttcctat ttaaataccc tttatttctt tctcttgact gattgctctg gccagaacat ccaatactat gttgaatagg agtggtgaga gagggcatcc ttgtcttgtg ccagttttca aaaagaatgc ttccagcttt ttcccattca gtatgatact ggctgtgggt ttgtcacaaa tagcacttat tattttgaga tatgttccat caatacctag tttattgaaa gcttttaaca tgaagagttg ttgaatttta tcaaaggcct tttctgcatc tattgagaat catgtggctt ttgtctctgg ttctgtttat gtgatgaatt acgtttattg atttgtgtat gctgaaccag ccttacactt cagggatgaa gccgacttga ttgtggtggg taagcttttt gatgtgctgt tggattcggt ttgccagtat tttattgagg atttttgcat tgatgttcat cagggatatt ggcctgaagt tttatttttg ttgttgttgt gtctctgcca ggttttcgta tcgggatgat gctgacctca taaaatgagt taggaaggag tccctccttt tcaattcttt tggaatagtt tcagaaggaa tgataccagc tcctctttgt acctctggta gaatttggct gtgaatctgt ctggtcctgg gctttttttg gttggtaggc tattaattac tacctcaatt tcagaacttg ttattggttt atttagggag ccaacttctt gggagggtgc gtgtgtccaa gaatttatcc atttcttcta gattttctag tttatttagg tagacatgtt tatagtattc tgatggtatt ttgtatttct gtggggtcag tggtgatatc cccgttatca ttttttattg tgtctatttg attcttctct cttttcttct ttatcagttt agctagcagt ctgtctatct attttgttaa ttttttcaaa aactagctcc tgaattcatt aattttttga atggcttctt gtgtcactgt ctccttcaat tctgctctga tcttagttat ttcttgtctt ctgctagctt ttagattttt ttgctcttgc ttctctagtt cttttcattg tgatgttagg gtatccattt gagatatttc tagctttctg atgtgggcat ttagtactat aaatttcctt cttaacactg ctttagttcc cagatattct ggtacgttgt ctctttgttc tcattggttt caaagaactt cttgatttgt gccttaactt cattatttac tcaggagtca ttcagcagca ggttgttcaa tttccttgta attgtgttgt tttgagtgag tttcttaatc ctgagttcta atttgattgc actgtggtct gagagactgt ttgttatgag ttcagttctt ttgcatttgc tgagcagtgt tttacttcca attatgtggt tgattttaga ataagtgcca tgtcgcactg agaagaatgt atattctgtt gatttggggt tgagagttct atagatgtcc attaggtcca cttgatccag agctgagttc aagtcctgaa tatccttgtt aattttctgt ctcgttaatc tgtctaatat tgacagtggg gtgttaaatt ctcccactat tattttgtgg gagtctaagt ctctttgtgg atctctaaga acttgtttta tgaatctggg tgctcctgta ttgggtgcat atatatttag gatagttagc tcttcttgtt gaattgatcc ctttaccttt aagtaatgcc attctttgtc tttttttatc tttgtttgtt taaagtccgt tttctcagag gctaggattg caacccctga tttttttttt ttctttccat ttgcttggta ggtcttcctc catcccttta ttttgagcct atgtgtgtct ttgcacatga gattggtccc ctgaatacag cataccaatg gatcttaact ctttatccaa gttggagact gggtatttta taaagaagaa aggtttaatt ggctcatggt tctccaggct gtacaggaag catggcacca ggcacctgct cagcttctga tgaggcctca ggaagcttac aatcatggca gaaggcagac ggggagcagg cacatcacgt gacaagaatg aacacgagag agagatagtg gaggggaggt gccctacact tgtaaatgac cagatcccct gtgaactcag agcaggagct cattgatcac ctaggagttg gctcaagcca ttcatgaggg ttctgcccct gtgatccaat cacctcccac caggccccac ctccagcgct gaggagtata cttcgatgag atttgcgcaa ggacaaatat ccaaactata tcagggagtc gttagtatgt ctggccacct tgaaaggatg aaagcttatt ggagtggaaa aaaaattgcc tctctccagc catacaactc agtcaaacca cattcaatta acataaatat attttgacct ccaagtatgt aagaggtgat ttatattcac aatttctaat ttataataca gtttcataaa tttgatcttc acaagaaaga tgagcaaatt gctccttgcc aaagttaatt ggctgaatta ccaactgaag ttctgttatg cttggcccca aattctggtg gcccctttcc attatgtcaa ctggctctct ttacatatag attctcaccc tgtagtttcc agttcctaag gcaaaaggag aaaagttgct gcgtgaggat ctcttggccc agattttaaa agtcaaagac agtggactgc cttataacct caaacggttt cacatttcag tttggagctg - 113 - 008252 atcagtgcat atctcactct acccgctggg tacgctacca ccaggctggt tgggattgca ttcatgccct cactgtccct attttaaaaa atcatgtact ttaccatttt gaatgggtga agaaaaaatg tgcttcagaa agttttccaa ggattagtat aattaatcct agtgaagctg ggaaaacctc atatctgcca tctcttgagt ttctccatgg tatataaata gtgttatctg cttatggaat acaaatgggt taacaaaaaa tatatgtttt gacctttttt gtcacagtat taacagtgta atctcactaa ctctttcttt ttccagaatg atagtcactc gtagtcatag tcttaaagca gcagtaaagt gtggcataga acaattaagt taagcatagt cagaagccat gaggaatctt gaggagaact tatatccctt cagctggaaa tctcccaagg tatacagctc gaggtcctga attttcttag attaaaaaca ttagctgctc atttaaaaca caacaggatc ttaaataatg aaacatctac atgtatgtgt agatacagat aagaccaaaa ggctctgcca ctattaataa tcaatttaag caaccaaaca cctctgtgtc gttgcccagg ttcaagtgat aacctggcta ctcgaattcc ggcgtgagcc ctcattgcct tatccatact gctattcaga gcattgtcac tatgcatgat aaggagtggg tgccaggtta tatttatctt aatcaataaa tccaaagcat gaccagagga gaaggcggta tcaatctagg caaaaggcaa gcaaaagagt gtttatttag aaaatgaagc ttgcatgtgg gccgacttca caaatttcga aggtaataac gaaaatctag gtcctttctc ccatttaggg gtttaaaaat а с а у с t з с а с ctgtcactct ttatgctgtt caagcaacac ttgatgtttc ttcatcacac ccctgagtcc gtaggcttgt gcccccctac catattcatt gatggtagtt aacatgaagt atgcacagtg ttcttttata gtttctcatt taattaagtt agtgctctgc ggtattgttt atctcaaact aacaagcaaa ccagggattc ttQ323"tC3 cacttattaa gtagaaagac tatatattag atatacatat ataaagagat tcttcagggc aagatgtggt gatgagtccc tgttaatcac gccaggcacc ctgatgctta ctgtagttca tctcctgcct atttttgtat tgaccttagg actgcacccg tatcctaacc ggcaaagtag tataacaaga ttaaatttca ggctacaatt aaaatcatat acttgtttat gttttgcagt taatttatct ccagcattag agctaaataa acagccagac ggtcagcttt gacaagcacc ctgtgtctgt aattgtgttc cttacagatg atttgttggg gaaatgtatt taccaagaag aatgtaaaca caagtaaatt acctactagg tagcacgaat tgcttttgta ucaccaQa^a ctctctctca tattccatcc cctattgcaa cgctttgaac tctcctgtgg attttattcc caaaaataat gaatacagga aatgacagtg ccatatctgt cttaagctat aagcatatgt tgaaagcgtt tttcagatga ggttaatcac accgctagga gtggccatta cctacaaaat catatgctag tagtactcat rtt- ппе-яя h f +¦ •3"-3 — — — —~"- agagtttctg acattcattc gtttctccag atgtatgttg ttactttaag agcctggaaa aggccagtga actcacatta aactgtaaaa atactttagt acttttcttt gtggcatgat cagcctccca ttttagtaga tgatccgccc accaactttt caactgcagt ttgagggagt taactattta tagtcacaat ctttacaact taaaatactc taaaagaggt tccactaaaa ctgcaaataa aattagctag gagtgaaaat taagttttct tagcaagata acgtaaggta gtctatgtat ctctattttt caatcaaaaa ctacttattt gtagcttgga tcatccttag aaattgctat tagtaataat aacagtttat ccttgtcttt tgataacaac aaaggtgaaa tatgcaaatg ctagaactag ggttttccca ttcagttaac tcattccttt .ttttattcat tattatgaat aaatttttct attacagaag agagaaagac tgcctttgtc tgaattatgg atgaggtcct ggaaactgaa cgaactgaga gtggtgaata tcaacaattt gttatttgta gaggataatt tttgatacat П+ ПГТаа t* Y at" - -•Э'З — - - - tagtttacaa attcattcat agagaatata gcttagctca gaattggctc tttcagcaga cctctccctg tggagggtaa taccttcaca caagttgata tcttttcttt gttggctcag agtagctggg gatgggtttt gcctcggcct ctgatattat cttctggctt caacttgggg aaatattcct atttgaaaaa ttgcctacca agttttacaa gtgatgtcaa gtccaggcgg gactcttgct accaattcat gaagtcagat gtaagtacat atgtgtcttg tcaaaggcta gattgtgtgt gatgtatttc caaaatgctc atttacatca ctctgcaaat cctgtttgtc gtataaattc ttattttgtg aatcagctca taccctaatc tgcttgttaa -.+- ~*+- ayy UV.I.V.V.-W, caggctctga tacgtgtgca gtacccacca atcaggagca tttattctct ctccattcct taataaacag gaaaagaggc gccaacacta gatattgtga gttggtgagt ggcacatgct tctgggaatt gctcaaatgt tgatcatctt acttaaaacc tagtaacaag aatagaatgt gaaggctttt ggaagctact acaCwtaaaa ataaataatt ttattcagaa gaatagaata aaagatatgt atgtaatttt agctgatgtg cagagaagtc catgctttac gcaacatcta cataaattta ttgaggtgga 64080 r tgcaacctct 64140 attacaggca 64200 actgtgttgg 64260 cccaaagtgc 64320 tttgaatatg 64380 ctgtcaggtg 64440 aacatatatt 64500 ttaaaataaa 64560 atacttaatt 64620 accctttcag г t r- r* r\ OIOOU cttatttcct 64740 tattatttcc 64800 agccattgaa 64860 ttccagggtg 64920 aggagtttat 64980 gattgtctga 65040 gcaatgacca 65100 ctaaattttt 65160 tattcattat 65220 gggtgtaatt 65280 tagaacatac 65340 aagttgtata 65400 ctgacaattt 65460 gtcttttaca 65520 tttagctttt 65580 ctatcatagg 65640 agtatctttt 65700 tgctgcatca 65760 tataaatctc 65820 caatccttgt 65880 *- +- 65940 cattttcact 66000 tcctttaata 66060 tctgccaccc 66120 tatagatagc 66180 ctcatactaa 66240 agcatgctgt 66300 acattcaaat 66360 taagtcatca 66420 agtgcagtgc 66480 ataacgaaga 66540 cttgtggtga 66600 tttgcttaag 66660 gttttttaaa 66720 ttcaagtgaa 66780 tgtgttgtga 66840 acaggactct 66900 ttcagaacac 66960 tttgttaaga 67020 aaaaaaacaa 67080 aaagccagaa 67140 nhpiraf яяаа 67200 caccatagat 67260 aataattatt 67320 tattatatat 67380 ctatagatac 67440 gtaggctgtt 67500 gcaggccaaa 67560 aaaatctttc 67620 tcaaagtcta 67680 gactggtgtt 67740 actatcatag 67800 - 114 - 008252 acgctgtgtc gacatccaat caaaaagtga tggagtggct gttaaccaaa agctgggaga gggtgtttga ttgccactgt tgtatcagaa tgtgtagaga ggagaacgtg agacttcaag ggtttaggat aagtggagaa ggatgacaga aaaagagaaa tagaggagat gccagcagag atgatcactg ctcccaaatt gcaacattat tttttctcta ctaaaatttt ggagtcacag aagaaatmtg attaccactg tccaccccac atgactacgg ctgaaaaaaa aatttttaaa cttctaataa tcaagcctgc aaaatgagta aaaatactct taacttattt ccctacaaag cacagggctg taacatctag cttacgtctc taaagcatgc gctccaaagt atgtagacag taaaaatata cttcaagagt tgaacatgct ctgggattta tttgagtaca tacttaaccc aggaagctga gagctgggat tcttactgtt agtatttgta tfcggcctaca tatctttctg tacagatggg cttatttcct taaaggaata tgagaagtgg gtgtgtaaaa caattaggtg cataagaagg ctcacgtgac atgtagtggg agggacttca gaacagatga gggtaccttg cccccgagaa gaaaacagga aggccaacgt agtgtgggag taaggtagga aggctaagag gggagatgga acgattactt tttgctccac ttcaaattga atctaataag gaattgagaa ggtcaaagac ggaaaagcag ttgcatctgt ttagtgattc taatgtcttt ctactcacag ccacaaaaac tctgattggg aaatgggcta tgagtaagaa aaagaatcat ccccaactcc aaataattaa tgaaacaaag aacagtaatt atgtgtttct tgcctctgaa tcagcactaa aatatttata ttggcaccat aatgcaataa gagtttcaat cattgattgt accccaggct ttaagtcaca ccaatttctt agtgttatct tgtaattttg aaatgaagaa aaagtttctt tgctacattg gtacttgcaa tcacatgagc gcttagaggc caaacccaga atattgtagg ccacacattc -------J acccacccaa gtccaaagga gagaccagca gcttagggtg gcaaagtatc ctatgtacgt tcagacttta tattaaagaa tgaggctgtc aaattagtgt tggaattgga ccacacaaac agaagttata aatgatagtt gagacttcca ggtcagacac gagccagatc cagtgattat tgggaggcag gagaatggaa atgagggttg ggtttggaag ataagggacc cagttggaca atcatgaaag agagctcaga aatggcctna gtcttcctcc atggagttag agccttaatc gttccatatg aacaattcag tatatcatat aaaagttaaa atgctggtct tgtctatgtc agagagtcac ggttaatgaa attaacaaat accttgtggt aataaacatt gccaaagact tatgaaagaa aattaattta tctgagtgtt ctatcctcat ctaggaaatc ctatcaagta tatgaggtag catgtaataa ctccaaatat tagacatttc atgggctagt ctaggtgatc ctcctctttt tattattgta cacattaggt tttgctttta ccttagagcc aagcctcact taccaaatca tgaaacagag апаяаЬ nncin —-,---— zi ~ zi z> ttgtggtaat ccagcttaaa gacagatgag tttgaatcat accagccatg gtgtgtatgc gagaaagcat aaagagtcat ttagggttgt aacacaatta aattacacta atgaattgcc gcagggaaac aagctggggc agtggaggag tgggatactt cttcagtatc gaagggtctc ggtggccagt ttagctcaaa agggagaagt ctggtcctgt tattgagctg tgtagctcta tggatgagat gcagctcatg agaaatatgg catactcaca gttgggtcca acacctataa ttagaatggg agcactgctg ggaagtgaac agagggagat tgaaacccgt caggattttc ttgttctcta taaaaataaa atatattctg tttagataca tgctaataac tcaaaataat aaagcaaata aaaaatcata gcacattatc tttggnaagt tacaactttc cctgttttat acgttttcac gtaggtcaga ctgaaatatg tgctattgta agacccttcc catatagtat aacaagtctt atgattaata gctatgcaaa attatgatta ttatttaagt ctgaaactca ttgatgacaa tcagatcaga agatatacac ttccacaatt atatgactag ctcatctgct gggtgaagtt gggtgtgtgt gtggtggcag attataaaag ctggtaagtc tctccattta tggatttaga aggaacagtt tttgtctcaa ctggcccagt ctgaaggcta aagaccctgc gggggaaagg ttgcagacca tccatatgct gaggaggcca agatattttg aggtatcaag ttgttatgac gaggtacttg agactctaca tgcctgggag actaaaggtc acnaaaccag tctctctcca cctggataat ctttagttcc gacatcttta agaggtcaca aaaagtaatt tacaagtaga ttaaacatga ctggtggggc tccaaagtac ctctggaaga aaactcttga tgtgttccag ttgatgttgg atctagtgag taataggaag atcattaaca taatttaatc gggataactg aaactgtcaa acaatttaca accattatgg actcacagcc atttttttgg tttttaaaag ctctctgaac ctcttagctg tattgttctg atcatttaga gtgcattact ctctctgtac cacacaacca gaagcccagt gcatcgttga ctgaagcaaa nabnt- rb n agagcctttc agggagcaat gctgacaaag tctggagctt gtgtgtgtgt caaatagcta ggatggaagc tacctcaatt agaaattcct aggcttctga gagtttggaa gtgccagaaa ggaaggcact caaaaatggg attaggcagg gggtacaaag tgttacaggt ttgtgccaaa gataagagtg agaatatgtt catgattcta agggaactct tggtgtagcc acgatgtaaa agttatcatg agctcacgac caccttcaaa naccacagcc tcaggatcat cttttgctat ggggaccagt gaagataaaa tcattgtctt tactttattc gtggatattt aagaattcca ctgagggaaa ttcttgttaa cattcatttc ggcaattaat taaaaatgtt agttgtgctt aaaaaaggaa gttgagtgat attaaaatgc aggctataca taacagtagg gatgtaaata attggaaaac aagaagattg aaactttttc tatttggaat ttcaccttgc catagttcta tttttacatg tcaagcatta tggtttactt cttacagatg gtaaatggca ctccttacca tgaggttaat gatgaatcat tctaaaataa ccttgggttt gatttaacca ggcagaaccc tgtttatggc gtgtgtgtgt agggattgca agcaactgaa tggtcagaaa agaaacattg aatcatcaag - 115 - 008252 cccagacttt ccatttttaa ttgctgtcag ttcttaagaa agagctatca catttctcct aagtatcatt acattgctta tcacaatgac ttgcttatct aagagctatt tatgaatatc ttgcaaataa agagagagag ttaccacagt ctcatttctc tctatatttc gtgatttggt tctccctctc tagatagata tgctctgttg cctgggctca caacctcact tcaacattta cctggaatag agtgcccacc tgggctacag ctattttgtg ctggctgtag tattctaccc atggagttac ttaaaaatat tggtttcctt actgtaatac caattaatga tgtgcatgag taaaatgtgc ttgtattttt aattccaaac atttcttcaa tagtactgaa gttttcatct gcattcttcc caactgatgt tagtcttaaa cttttaggca ataggctgtg cttatcagag agtggtcacc atcatgtagc tcaaaatgca ttgcttaata ttgagaggac ttatcttact ggaagtgcat agccctccat taaccaggca ttgttctttt cttcctttct ctgtatgact aaatggtcgt tagatacaca aaaatgagga ggcgtctgct ggtcaaccca gaaagccatt ggataaaaat ctaaatcacc tatttgtctg gatgagtttc gaagtgggga tcctcagaca gggtctcagt gtggggatga atttatggat catcatatat agagagagag tgggtgggcc attcttatat aaagtgtgtg atctacaaca taaatatata gatagataga cccaggctag agggattctc tggccagata tgattgttga tttagctatt tggatttaaa tggctttaat atgtttatga tatagaagaa gacagtgcgt agatctctca ttgtttgttg tcatatgtaa taggtattga actacttagt aactatgtgc cattacatac ataattccat aagtatttta gtgtcaggcc cgtgggctga ttatgtttta atctcttttg gaagcaattt tccttccctc tgaggaagaa cggaataagc tctgcagctc attccagggt taaagcacct gcattatatt tagggtagaa cagttctagg atagctgtac cttttcgtct aaatatttac tgtttgtatg gcctgatttt atacaggctt aattgttaga tataatttag aactactata aaataaaata caatctgtgt gaatgagagt acatatctaa gggttatgtc tcataatctc gttatttcta gtagtattcg agtaaaatgt agttaacgtt ctgtaatggc atcagatatc tatgaatctt atatatatat agagagaaag agaaagatat gtaagccaaa gtacaaatat taagagcaag tatatatata tagatagata aatgcagtgg ctgcctatgc catagatatt aattgtctaa ccagacacac tgcatggacc gagataggtc agtggacaga atacaaatat agcatacaag tttttgacta aacagggttc ctatgtaact tgagtgcatt gaatatgcat tcaaatgtgt ataatgaaag gcaagttgcg tgccttgaga tgcgtgctaa tggtatcttt gtatttcctc accagtaata caattttcta tgtctcaaaa atcacagatg ttcaggagtt tttcttttag taaggttgcc cataccctca caagttgatt tctctttttg aggaaattgc tgttaattta tcaagtctcc taataggctc aggaggcaga gtctgttttt aatttcatcc taatgaaatg atcgaaattg tttaaaagtt ttttattaga gggatacctc ctacagtcag atcctttata attatccacc tctttcctat aactagacac tactgcaaag gtgcccacag ctttgcaaat aaaaaccgta atttgagaat aaaactatat atatatatag tctctcaagg gattttagac tgatccatct tattaacctc aaggtctctc tatatatatt gatagaaaga catgatcata ctcccaaagt aatgaagggt cactttcatg actgctcacc ttgaagatct tgtcttactg ataattaaat aaattttaat ctgtgtatgt tttcgattct tgggttgttg ataccctaat agagctgaaa caactaagta acattcttct aatttagcat catattatat attgttatca accataaagt ttaaagtcag atattttatt ttgtctccta tactctctta gtacagaggc tggtatggga ccaacagctg ggtctgagtg cttacagaaa ccgtttttaa catctttctt agagaaaaag cttggaggca tctcccaagg agggcacagg aacaaagcgt gccacaatag gttggcaata gtttgcttat ttgatcttgt atagctgctt cttatggaaa cacatgtgat agatagcagc cagtcctatg tcaagttgat taaaggattc gggatacctt cttccaacac aaaatgattt tgagacaaga ctgtagaaaa atggtgatag gtttaatatg tcagatacac agagagagag tctcatatat agacacgtca tcgcttcagt atcaacaatg aaggtctctc ctctttgctg gcaaaggtct gttcactgca gcaacgatta aattacatta tgttgttcac agagctagaa gggacttagc gtaaaatacg aattgatact tctatgccta atatgggcag caataacrgc actcaaagat aggcaaggta agtgctcctt ataaatgtat tggcttgcct ctgccattta tattaaagca gagtaatttt tacagctttt ttttgcaaat tcaagcatta tttcacaaaa aacttatcca atcatgaatt ggcgagatgt ggcatgacta tatccatgtc ataacctcca tgcaacaatt cactctgact tctccttgta gtcgccccct atttaagctc gctatgcctg atgtcttttt agaatttagt ttcattattc ttttcccaaa accataacca cctgatatta caaaagacaa catctcttca tatttggggt ttatctatac atccaaccta ttatattcct gagttccttc tcagaaggac tcatcatgca attttgtgaa attccttcct tattcttata LOCI U I. CI- - у t- atacacaatt agagagagag atattaccaa cgtatatctc cttaactgac cttgtcatca tttctctctc tactcttaga ctcaaggtct acctcgaacc caggcatgag ttcattgtaa aggaatgtaa gaaggctacc aggcaagagc ttttaagttt aatattttga ttttataata cagtgtttgt atttaagcat aaacaaaatt agtcttaaaa gattagaata gggaaatgct tattaagtcc gttgatatca aaaaatgtat ttaaactaaa atatagaata aacatctcag tagttagatg atccagaagc actttgtaat ataccaataa atataggctg gaatgcatgc ttctatggag attttttaaa gtacccaaat tacaatggct ctgtgctgac actaaagcaa cttgaaaaca gctcacaggg aattcattgg tgttttcttg ctcatgactc ttcatcatca aaacattttt tgtatctctg ttttttgccc tgcatgcatt - 116 - 008252 tgaatttaaa tatgtaagta accattcaaa ctcattaaga cagagtcatg catctttgcc atgaaatttt gggcatctaa agcaggatag ttgagtcagg tcacaaggca acaactgctt gttttcatgg caagcaccat ttttaaaggc aagaaataca atccattttg ttatttctca cctggtgaag gtagaagcag tgggctccac ccatgggggt cactatccta taacaaaaat aatgtacttt ttagtaggaa aacatagaac gaaaacattc agagaggcct aaattcttta atagagacta tatcatgagc atttacaaaa accgggggag aagtaacatt atgagaatgc taatattatc caattctttc taacctctga gactcctttg tagatcatga cctcaatttc aaagagcaac ctagaaattg ggtgtaataa atgattccct gaggcattgg tctcggtctg cctcctgggt ccatcaccat agccaggagg tgggattaca tgcctatgtt aaggcaagtt aatgtcctga gtcattgaat catatacatt caggtgctcc agccatgctg aattccctga tgcacctttc actgactcat ttgcaagctt acataattag attatggtta acatgaattt tttaacactg gagtagaaaa tctttgtgtc catgtttaag cagaaaactt caatattcta gaatcagagc gaggccctca tgttgctgct ccactataac cctcatgtca tctccaggtg cacatagaca gttgttgtaa cagttctaga gctcacttcc taagggttct cttcatgatc tagaaattca gaacattacc ttatttcaaa gaggtacttt acagagcact agtcaacact attaggcatc gaaacttcat gggaaagaat tattttgcat tagtctctag tacaatgtaa atagtgtgat tatacaatat atgataaact tatgaccacg taaatcttga atagagttat tggtaaagtt ctccacttta cccatctagg tgagaaacta ggatgcacgg taagggaggg ttgtttacag ggcaggggca tcgcccaagc tcaagcagtt gcctggctaa gtctcgatct ggcgtgagcc aactcatatc tttggtgatg cttctgtgct tttctagttt cgaatctgtg ttccacagag tgatgtgcag gccacgtgac catctttatt ccctgctaag cctgagtagt gtgcctaatt gttaaaaagg caattcactt atgatgttga aattacaagg cccaaaatca aactctgaaa cctgagaaga agataatgca aaaggctgat tttctgcagg gatctgggtt ttaggatgac attaaatcac attaatttat cacacacacc caaaatgtca gtctgggaag tggttcatag ctcttgaacc taatcacacc acaaatgaat ctgttcctcc gatgagtcat taaaactgaa gaattacaga gagacacatc cagaaaaaat gtcaatatgg gggtcaagct gaaatcttcn tatctatttg atattataaa tatgttgata atgaaatgta cagnaaatta tcccaaaaaa taatagttct gctatttgct tgcttgaagc tattagtgtg aaatggggta cacttagagt gcaaaatatg caggacatag acattaacta cggtgttttt tggagtgcag ctcctgcctc tttttttgta cctgacctcg actgcacccg tcccatttca tttggtcttt tgtaccaatc gatgaagcat acccctggat cctaagtaaa gggctccagc aactgagata tctggatagc acattagccc agacataatc atgcatatga agcccatttc ataaaatttc aaaattcaaa aacccctgtt tccaaatatc agctgtaact ttctgcctac tgttcctgga tgtggaggct cagttggtgg ggctagaatg ctggggaacc aaccttagag gtgcatgcag tgtattataa cagaatcagt tccaagagca aatggcacct tccttcataa caaaggtcct ttgggggaaa ccaaagtgaa aattgaaaga aaaaaatcat ttcgaaggat ttagtaaaat aaggtcaagt aactctagaa ctttctgaat ttgaaaacat ttttaagaac ttatgaatat ttcttaacat taatagagat ctttctttag tcatttcaat caaaagaaaa gtagcatgtt acagttgttc acagccttag agtgatagac acgtgtagcg aatgtatctc atattttcac tgcaggtttt tgtttgtttg tggcacaatt tgcctcccag tttttagtag tgatccaccc ccggggcacg ttttattcca attcatcaat tctctagaat catgcatatg tggagcctct gcaccagctg actaagtgat ctagttattt tttcctgtgg gtgcactcaa taaaggacat atttggctaa cttttcttac tggcagaagt ttttaaaata gtaagattga aagaagattg ataaaccaca tgtggatctc ggaacacaac tactaggacc ggagatagag ggataaaagc ttttaaaaca tgatccaagg agttatgaac aaaccttcaa gtctcataac agacagcagc tttcactgtg ggttactaat accttctggt catcagcatt acagtgacag atgaggacca gatatataac gcagcttttc tgttggaaat tagctataag tgtaatggca atttttgaag tcagttaatc aaagctaata ctagaagaga tcacatttta atgtgcatac aaataactca atgagtaata gtaacaaaca taaaattatt aaaagaatgg gaaagtaatt ctaccttata tagaacaaac tgtgttattt actcacacac ccagtatctt tttgtttgtt ttggctcact ttagctggga agacggagtt gcctcggcct gtgttcttaa atcaatgact tattcccctc ctgatcaaca tttaatttta ccctttcagc gggcattgag ttattttccg ctgtctcttt ctccttccta gatatcacac cttgcaaaat tatacttgaa taatcaaatt agctttatgt tattaaaagc agagtaaaga agaaacagaa tcacaggact agacagcacc tgtctcatat ccatgcagca tggaggggac taaacggaag aacagattct aatagatggc cattggacta ttccatcttg caagagaaat agatttcatg tcctcacgta accattaatg agcatcaccc ctgtccattg acatctggtc catctataaa tcataaaagc tcagtaagtg caaagctaaa gaggtacatc caatgcttac aggcaggaac aagaatttct taactgtagg taagatgaaa gcagttaaaa ttgtcatgag aaaatccaaa atattggaat gtatattttt aggggcacat ccctggggtt aactctcttg cagttaatgt gcagaatgaa gagggaaaat tgcatctgat attcaaggga tgagatggag gcaaactttg ctacaggcgc tcaccgtgtt ccaaaagtgc ccttgcatct agcattctag ccctctcaca tttagcccgt tttatttatc gcctttcaan cagaaggcac nacctcctct cttccattta aggaccccca actccccatg cgcaaaattt - 117 - 008252 atttagcaga acagtggtaa aatcacagca ctttcagttc gagacataaa agtttagagc gagtcagagt cttctccaga caaaatagtg ttttctagtg atccaaacag gaaataaatc aagaaagtaa cccttcattc ttaatgaatg tatcatgcat aaaaaaatcc gcaagataaa atgcccttaa aaagtcgaag atcagacagt ttatcgattg taatcctcaa agataagcaa tgctgccccg agagtagcca taaagtctca gcatatcctg aaaagaataa gctgaggcag aatcctgtct cage i. а с u eg aagecgagat aataaataaa aaggaatgtg gtctagtccc ttgaatgaaa tactgagctg ctacttttga tagcaagcaa ctaaattcaa attaatttct ttctaaatta aattaatttc gaaaaaatta caagtegtea atgatgactt agaaaatgea actcacttct aagaaatata gtattgacta tattgattta acccctgtat cattgagatt ggtttgggga acttacccta ctacttccgc agaggegaca tggaacagat gagtgggatt tttgcagatg tttaagaaga gaatgttagc aggtgatgtg acagcaaaaa aacatttggt attgtacagt tcattggatg tttttattag agaataaatg ctatgaatca cactacacga gtaaagccag cgttactttt catattatgt aatcagccca gaatgaactc tgtgctgagg gecatgagta ttatgatgat caacatggtg atgtgggctg gcagatcacc ctacagaaat ccgtgattgc ttgacaacat tttagaaaat tgnttcccac ctgtgatttt aatgeagtea agaattacag tccaccatac aaattcaaaa attatttccc ccagttagaa aacaccatcg acaagagatg caggtaaaaa ataaaactca tctttagaat aaattcaact agacattctg gctgtatatr gaaataataa gtaattactg gcctggcacc tggggtatca tcccagagcc ctgaagcctg accttcattc ggaagaaaca acctccctct ggcgtggtgg tgaggtcagg acaaaaatta agtatgggtg cctaaaagcc gtgtcttccc ttgctcataa ctgtaataac tacattttac agaactcegg ttgctgtttc acaatttgea tttagtgaaa agaaagcaac aactggtgat agaacaaaac ataaaaaatg tcctgaaaag catcagaata gaaacagctt aaaataacta gtggatcact acactgtatt tcattactta agtgaaaagg ttatccctca tcttaccagc tgctcttgat ctgtggcttg catcttccaa tgtcctgaag ctcaagcctg aattcaagac gctgggcatg gcaaggtggg acactgtcga ttttctgaat acatatgaga actggaaatg ccacacaggt gcagaggcag caccaaaccg aagcttagtt tgctgatggg acaaaatata tccttgtcat tattcaggtg ttcagtttgg tcacttagga aatgacaaaa atagcagagg atagaaaaga ttgcctgttc cttatggtga ggacaactgt ttttgeatec gctgatgagt cgcatttgga ctacaaacta tcaggacacc aagagtgacc tctttagaca taatccaagc cagcctgggc atggcgggtg tgggaggcag agcaagactc aataataaag gggttggtct tggtgcataa ggcaatgcag ccagaaaaaa tagagatgtg aagttttgtg tgtaatgtat tatgaatatt tgattttgga ccttattagt aatattagat atgctaagaa taaattaatt ccagatttcc ggactggtat ttcttctttg cataaatcag ggagttctgt taggaatcat agtgacacaa acttacccca cataaaagct aaattacaga ggggatctga gctgaactca tcttacaact ttattcctaa gaagaccgtt ecatatatge gaccttttaa gttatttgaa actaaacaac agtaaatege gcccattttc aagcccacaa catacaacat tatatattca ctaattgtcc tggntatcat attgtgtttt ggggagaatg agecatgtet ctacgttacc tggtccatta tgccatagtg agtcttagaa actttgggag aacatagtga ccagtaatcc agg \_acagu tgtctcaaaa agtaaatgtg ctggctttaa tatatgttag ttttataagc ttagtaaata ggagagaact tgtatttgaa agetacatge taaatttget aatccacatt ttntctttac aagaaaacta caaatttaag gggectttea tatattccca tttttacaag aatttgggta ttttattaaa getteggact ууаууи> -,уау tgagccactg taaataaata acgtccactg ttgtattctg gaatataaat taaacatctg tgtaaaatta aagaaaatat aatgccgtga gttgtgtatt tcacaaatta tttattagaa taaangcttt gctccacagg gtaatcttca gaaaactaag gaagaacgta ctgtagctgt taagctatgg gaaatatcag ettgetgaaa aactccagcc aataaataaa caaactgatg tattctgtga attaaacaat ggcaaggatt ggtggtagac gctatttggt taaggcatgt tgaatttact accaacaaat tgtgacataa tctatatata agttaacatg tgaattgett gtaaagggtg attatgagta tatttacttt actcagctcc ttattatttt taggattgac tgggcaacag taaataaata gccccttaag gacagagagt gtacctggga agagataatg acatcatggc atattttagt gtttgetata ttctttgaga ttatacaagg acagtattta tcctccttaa tggattttgg ttggcaacaa attgagtctt ttgaaatttg tgtggtattt ctgtgtgggt cntcactggt aat <210> 2 <211> 237961 <212> ДНК <213> Homo sapiens <220> <221> экзон <222> 43726..43868 <223> экзон 8 - 118 - 008252 <220> <221> экзон <222> 43998..44102 <223> экзон 9 <220> <221> экзон <222> 52093..52179 <223> экзон 10 <220> <221> экзон <222> 77568..77699 <223> зкзон 11 <220> <221> экзон <222> 98226..98393 <223> экзон 12 <220> <221> экзон <222> 106567..106758 <223> экзон 13 <220> <221> экзон <222> 144109..144246 <223> экзон 14 <220> <221> экзон <222> 159794..159868 <223> экзон 15 <220> <221> экзон <222> 191292..191428 <223> экзон 16 <220> <221> экзон <222> 192967..193108 <223> экзон 17 <220> <221> экзон <222> 211540..211613 <223> экзон 18 <220> <221> экзон <222> 225006..225107 <223> экзон 19 <220> <221> экзон <222> 225544..225613 <223> экзон 20 <220> <221> экзон - 119 - 008252 - 120 - 008252 <220> <221> <222> <223> <220> <221> <222> <223> <220> <221> <222> <223> аллель 166087 99-79322-224 : полиморфное основание G или Т аллель 166336 99-79322-473 : полиморфное основание А или G аллель 235894 99-79306-182 : полиморфное основание С или Т связанный праймер 51031..51051 99-79335.ри <220> <221> связанный праймер <222> 51539..51559 <223> 99-79335.гр комплемент <220> <221> связанный праймер <222> 60925..60945 <223> 99-79336.ри <220> <222> 61354..61374 <223> 99-79336.гр комплемент <220> <221> связанный праймер <222> 80271..80290 <223> 99-79338.ри <220> <221> связанный праймер <222> 80700..80720 <223> 99-79338.гр комплемент <220> <221> связанный праймер <222> 91037..91056 <223> 99-79339.ри <220> <221> связанный праймер <222> 91466..91486 <223> 99-79339.гр комплемент <220> <221> связанный праймер <222> 100285..100305 <223> 99-79314.ри <220> <221> связанный праймер <222> 100764..100784 - 121 - 008252 <223> 99-79314. гр комплемент <220> <221> связанный праймер <222> 106568..106585 <223> 99-79316.ри <220> <221> связанный праймер <222> 107000..107020 <223> 99-79316.гр комплемент <220> <221> связанный праймер <222> 165864..165884 <223> 99-79322.ри <220> <221> связанный праймер <222> 166381..166401 <223> 99-79322.гр комплемент <220> <221> связанный праймер <222> 235713..235732 <223> 99-79306.ри <220> <221> связанный праймер <222> 236190..236210 <223> 99-79306.гр комплемент <220> <221> связанный праймер <222> 51071..51089 <223> 99-79335-60.mis <220> <221> связанный праймер <222> 51091..51109 <223> 99-79335-60.mis комплемент <220> <221> связанный праймер <222> 61274..61292 <223> 99-79336-369.mis <220> <221> связанный праймер <222> 61294..61312 <223> 99-79336-369.mis комплемент <220> <221> связанный праймер <222> 80583..80601 <223> 99-79338-332.mis <220> <221> связанный праймер <222> 80603..80621 <223> 99-79338-332 .mis комплемент <220> - 122 - 008252 <221> связанный праймер <222> 100466..100484 <223> 99-79314-201.mis <220> <221> связанный праймер <222> 100486..100504 <223> 99-79314-201.mis комплемент <220> <221> связанный праймер <222> 100490..100508 <223> 99-79314-225.mis <220> <221> связанный праймер <222> 100510..100528 <223> 99-79314-225.mis комплемент <220> <221> связанный праймер <222> 106706..106724 <223> 99-79316-158.mis <220> <221> связанный праймер <222> 106726..106744 <223> 99-79316-158 .mis комплемент <220> <221> связанный праймер <222> 166068..166086 <223> 99-79322-224.mis <220> <221> связанный праймер <222> 166088..166106 <223> 99-79322-224 .mis комплемент <220> <221> связанный праймер <222> 166317..166335 <223> 99-79322-473.mis <220> <221> связанный праймер <222> 166337..166355 <223> 99-79322-473.mis комплемент <220> <221> связанный праймер <222> 235875..235893 <223> 99-79306-182.mis <220> <221> связанный праймер <222> 235895..235913 <223> 99-79306-182.mis комплемент <220> <221> ошибочное связывание <222> 51078..51102 <223> 99-79335-60.зонд - 123 - 008252 <220> <221> ошибочное связывание <222> 61281..61305 <223> 99-79336-369.зонд <220> <221> ошибочное связывание <222> 80590..80614 <223> 99-79338-332.зонд <220> <221> ошибочное связывание <222> 100473..100497 <223> 99-79314-201.зонд <220> <221> ошибочное связывание <222> 100497..100521 <223> 99-79314-225.зонд <220> <221> ошибочное связывание <222> 106713..106737 <223> 99-79316-158.зонд <220> <221> ошибочное связывание <222> 166075..166099 <223> 99-79322-224.зонд <220> <221> ошибочное связывание <222> 166324..166348 <223> 99-79322-473.зонд <220> <221> ошибочное связывание <222> 235882..235906 <223> 99-79306-182.зонд <220> <221> признак ошибки копирования <222> 6485,14 24 6,14248,14250,14252..14254,14256..14262,14 969..14 980 17712,18786,19477,194 96,21043..21064,21547,24 607,2494 6,24974 25072,25124,25136,25142,28843,31239,31372,37750..37751,4 0488 46542..46557,49309,49326,49357,49395..49398,49400..49402 4 9408..4 9409,56275..56280, 6104 8, 61050, 61059, 61856..61857 618 60..61861,64842,66307,66326..66327,69835..69840,70893,70905 73279,76154,76354..76355,76371..76372,76407..76408,77095 78933..78934,79363,89491..89492,95836,95838..95839,95841..95842 95958,96966, 9704 9,97067,97078, 97292, 98642,98712..98713,98716 98740..98741,101368,101373..101374,10137 6,101378,101401..101403 101406,110014, 129061, 129066,129973..129974,135336,135345,135359 139924,147368..147369,147469,147507..147510,147767,151694,151729 163111, 164480, 1697 64, 17334 0, 173390..173391,173398..173401,173407 173414,173419,173426,174800,1748 94,175041,175486,176256,17724 0 178864, 181149,181154, 181230,181287, 183433, 186606, 186648, 191065 191267,193117,193203,193223,193380,194051,194053,194057,196577 198388,198655, 198828, 19884 9,198862,199667, 199693, 199701, 199704 199747..199748,201117,201224,201226,201314,201410,202524,203887 204 396,204 939,205891,205930,205998,206908,207363..207364,210938 212018,214253..214254,217022,217333,219027,219909.-219912,220924 - 124 - 008252 222620,222882,223870,224 902..224 903,224911,224 914,226899,226915 227232.-227233,227415,228441,228446,229373,229471,233305,233405 233512,233601,233629,236096,237468,237471..237472,237478..237482 237486,237488..237490,237492,237495..237496,237498..237499 237504.-237507,237509 <223> n=a, g, сили- <400> 2 gatgttaaat aaggcgataa aatatgagga agaaaaatac ttttattcaa attataatgg 60 gatgtaaatt gaattttgtt gatgctgctg ctaaaaattg attcatcaga gttaaatgat 120 attaaagcat atttcatgct accatatgtg atatcagatg agaactttac aaataaatag 180 aaaacagaaa gttatatatc tacaaccatg atttattaca gaacgtatat ttttactgca 240 gtcataaaag aaaaagccca gtaaagacta atttgtaaaa tatgtaattg agatttataa 300 tgctttcaaa aaaaaaaaag caccaagggg tttttctgat gcatttatct tgatgtttac 360 tgttgccata ggaattactt attgattttt atgaaaacac atatggaaga gagaactaag 420 gcaatagaat tcaactaaaa gggcttcatc tatgataatt ctgtttgctc tgataaaaat 480 tcctttatca tggaacaagg aaaacctgac atatttttgt tgagtcgagg acaagatcaa 540 gaaacatcct atgaggaata tttgtgaaaa ctcttagcca aatgcaagga gttttatcct 600 tgtatcttga atcataagtg cccctaggtt acatgaaatg tgtgcttgag gatttttaaa 660 gggactgtct ttaggtttct caggttctca aatagggcca cagacttact taaactgaaa 720 ttaggtgtcc agatattctt cagaatgtta ggaagaaagg agtttaaagg cagaaattgc 780 aggcatgact ttttaaaaat aaggttgtcc attgtctgct gaaagatgtt taggagttct 840 tgttgaatgg atgagaaaag atagtgacta gaggagtttt tcatcagaca gtgaaaggaa 900 attttaaaag taaattttgg agagcataaa gtgatgattt atttcatgtt atatgtagaa 960 gacagtgttc attggaaata tctagtggat tccaagaaat tagcattttt atggttgtat 1020 tttaaacatt tataagttaa aaaagtaata cttcaaataa tgtaagagat ttaatgattt 1080 aagaaagtaa taaattctgc catatataat tccctccctc cttccctctc ttcctttctt 1140 ttctcatttc ttctcccctc tctgtaagtt tcatggtatg ttataacatg tttttcattc 1200 ttcccttact gatagacatt taagtggttt tcagtctttc cctgcaataa gtaatgctgt 1260 gttgaataca ggcatatata cattctgtgc tcatgtgtga ttatgcttta gaacagattc 1320 ctagaaacaa tttctgggtg aaatgatatg cacattttta atatgagata gctggcggtt 1380 4--,^4-4-*-+-> -+--* ^> ~ _, 4- 4- ~4- ~.t- ^--. *- I--.4- 4. -.-,,-,4- 4. -, 4- 4- 4- 4- 4- 4- Л А Л Pt " w i_ ъ" ъ. h. u ua сшауа.аауу a. - .atay La w UVJ \_-\_- U, L. ua fc. k.y -.y .yoy \-y aa i" w ь. u, v. а i_ а -л v aatacaaggt attctttttc aatatgcata aatattgaag aaaaaaaata atgtaggcat 1500 ttcgtattcc gtactctgta ctcttctgaa cactggcagt ccattctctt cttgaatgta 1560 tgtattcatc ccattcttcc tggaatgcct ctctccattc tcttcttgaa tgtatgtgtt 1620 catcccattc ttcctggatt gcctctctcc taccgctccc cctaccccca tctcctgtga 1680 ctcacctgtc aggacctact caggtaagtt tcccctaatc tgccagcagc actcatctcc 1740 cttctgtctg aactgtgata gcaaggtttg.tgcatcaaag tggacacttc gttacttcgt 1800 tctacatgtg tcccaccgtc tgggagtgtg tgtgtgtgtg tgcacacgtg cgtgcctgtg 1860 tgtgttttac ccttatgttc tggattcgtt cattagaccc agttgtctca ggtgggagca 1920 gagcctcagg cacttctttt gatgtctttt ctcgctctca ttgaactcgg tatatacttg 1980 aggactggct gatcacattt atgagaattt tatatttgat agattgcctt ttgaaaatca 2040 tcattctgca gaaatttagt aattgaaaat ttaaatcaaa ttcaaataca actctaggct 2100 ctaatttcac tgttcgtttg aatgtatgac ttctctgtta agccagatac tcacgtgaaa 2160 caaatgtgaa tagaatagca tggaaatcgt gtatggacag tcaaggaaac tttctctgct 2220 ggcgacagca gcctgcagaa cataattccc tggacactct atgaaatcaa atccgtttta 2280 tcaactaatc agttggagca aaaaataata atagtgttat ttaggaaagc cgtaacacag 2340 ctcttacctc aaggctaatg aagacctgac ccttgaactc taggggcttc catgttccag 2400 cttgtgagcc ctgctttgcc aaaaccctgc tgaagaggta agagcctcct tctctggaga 24 60 gaaagccccc agcctttgac aaagtgcttc tagcctgagc catgtgggct atgaaaaaga 2520 agttggtcat cccgaggagt cactggacag tggagagagc ccatccattt ttctccttta 2580 ttcttcccta cccctcagct ctcactccct taggacttgg gattctggaa tcataaaagc 2640 a a a a+-a 4-+-r-4* a a a +- a 4- a -.4- a a a a r* a 4- /-/-4- a a a 4- a a y-> +- +• a a a 4- a 4- -i/-"4- a a a 4-a a a a 4-4-a a a "Jl ПП -"-•-(-.-.UP-.-> w> h, U-BaUt-wUvV.-I -* -l"wt*V_ywbU Ы-*\-_ftU.WU-* -I-IW-fw-b-b-U Ub-ULwl-awWb. t- u _. г v v actaaaataa acgtattaga ataaatcatg aatggctaat aaatataagc tgccaccatt 2760 aaaacaaaaa taacaaccac taccacaatt gctactataa agattattat tgatcagctt 2820 tatttttagg taagtagaaa agtgagatgt atttctattt ttctgagaat gtaaccattc 2880 tgaaagaata ggagatactt tctcacagta agcagaagaa tgagatgtac tcataaataa 2940 tatattcaac aaacagttat tttgcactta ctctatatag gtactgaaaa tagaatgatg 3000 ttaataaata tgaacagccc tcaaacagag atattaaaat aaaacaagaa atgaaggtga 3060 aataaattta gaagaaacga tggatcatta acttgtagta gttcttacag ccatgtgctg 3120 gatgcaggca tccctgtctg agtgtgtgcg tgtgtacaga cattgtttcc aaactttttt 3180 actataactt tgttaaagtg aagcgtgagc actccacatt tgctgcaagt ctcacttccc 3240 ctatttcctt tatttctcac atttacgtaa cttgtctaac ctctgaaaca aatgttttta 3300 - 125 - 008252 ttagttctgg aataagatct tttctgcttc gcttcaatta cgttgtattc ctttcctttt tcgtttttct tatcctgtgc tgagactcat tctcttctcc catatttttt cttgacttct atgaccacaa acactgggga gggggctgct ttcatgcttc ggcatttagt ggatgcctgg tagaacactt cagtgtgtac caaggaatct attctttgct ttttcaattc cctgcctgtt atctcaccaa tctaactctc catttccact ttgtgtttgc tggcttaaag gtaatttata caatcatggc taatgcaaaa acagtagcat ctgtgtactt gtgcagaatt tggaatctat tttttttttt ctggtagttt ctaggcatgg gcctttcttc attctctcta attctatcct cttgttacca ccaaggagga tagttcccaa tagctgagtc ttcatatgtt atggtaaggt gaccagtttt ttaaacaacg ccaaaaaccc tctatcattc gaactcttaa caacatctat cttttatctt acattttaaa ctttacaata ggctatgagc tgcagtttca aatatattaa aagaaaaaga acaaggtgaa w -i .ywjyyy.a cgtgaggctt tacctcccac atgacatttg ttgcattatg ctgatttctc tctgcttttt cctttgtaac tgcctcccaa agtacatctg caaactctct aacttactct gaaaaaggtg gctgctaata ctctctctgt ctcccgggtt agccaccacg caggctggtt gagattacag gtaatgatgc atttttatac t-att-т- rr-t-i-n '------- --1 ttttngctgc gtcaatttgt tcagcaatga taagcttctt ttcagcaaat caagtaaatt ttgtgcatac aaattgaagt tgtaaggaaa ctaatggtac у у <-aay ay ay attcaccatc tgaatccctc ggtggaaaca ggggcactgt tgaggagact tctgagctgg acccagcttt ctcctctctc tgaaccattt gaccgctgga cttttatcta tctaaataag acattatata cacccaggct caagcaattc cccggctaat tagaactcct gtgtgagcca aattttcctt ttatatataa taatctttag aagttaagtg ttctgaaaat ctcctttggt tactgccaat tttttttttt ttaagattta atttttattt aattctggaa ttctttgttt ccatgtcttt cacaatgggc tttaaccaag agcagtgcct actgtgtgca gcccatacag gaagaggtct ttgtgtttgt gaaactcaag agggaccctg tccgtccctc ttttctctca aatttttata agattcctgt catgcttatt gttcattttc tcatttttca tggatatcaa tgtgttctca tttaatagat gaagtaaagg cctgtgcagg atgagaacag ccaggacaca cagctaaacc aaaattagac cttttgcccc aggccagggt gtagagacac tattgggacc ggcttcagct tagtttgctc tttccttgtg catagtccaa tatgtatata ggagtgcagt tcatgcctca ttttatattt gacctcaggt ctgtgtccag tctttgatat aatgcatttt tgaatagtta gtaaagacaa taattattga aatattaatt aagcaaaata gtcctaggca agatatatta tgaactgaaa gtcacctagt agatgttgag tgccaacctc aataatatta gtactgagaa gcagcaaata tgaaataaaa attcttacca tctcatttaa gaggctttcc cctcacatca atgcagagaa agaattacgt tgccaccttc ggctttaatt attgattgga aacaagccta tggctttgtg ccctttgtct tttatctact ggtttttagg tctggaatac aaatcgtgct gggttcagtt ggatttcatt ccccagacac gaggtggagt aaaatggcac ccaaagttca tctttgtttt gtgcaacagt ggttacactc caagagttgt gtgtgtctag cctaatattt tcttgctgtt ttagaatttg ttgtgtggtt ggggtactct ccgttcttac tgctgctaat tcacagttcc tatgtcttac gaaad.gccc cacaggaaag tggggattat atatcaggga tacacacttc gcatggatct tcttccagtc ctcagctcca cagaagccct gctgctcaca atgtagcttt ttgatttggc cacattgacc tatattatat ggcatgatct gcctcccaag ttagtagaga gatctgccca cctatgtttt gatgcatcta attaattaag ttCCatt ?C€f tgtgcagcct gagagcaaga caattccagt ttcaaatgct accctaattc tcttatgtcc aaactagcgg tgtaaagata accccaggtg cagaagcagc ttttattttt attttgtagc tgtcttattt cataaaacct ccgtgagtcg aatttgggtt ttgctgtctt gtgaactcat tattatcttc tgattaatat ttctaacttc gcttggcagg actggcaaca aggtggaaac gcaggcaggc ataagctctt tctccagctc gcaggggctc taaaagtgga ttgatccttg ttacgtattt actgctattt ctatctaatt ttctttctaa tttctaatgc tataatattg ggcctgaact tgatactcgt aaagacatac acatggctgg ttggtggcag иииаиаааач- actcgccccc tatgggagct aggagaggtc agttggccca ggacagaagt cagataaaat acttactgtc gaagatttat gctctgattt gagataattt atagggtgag taaagcccat ataatttttt cagctcactg tagctgggac cagggtttca cctcagcctc tatattttta tgattttata cattagttct gggaagacaa gaatgagaca ggattaagat ttcactttcc agaattacct agtaaaacaa tgattgtaaa cttaggatcc tgacatgtta tcttagggga gcttttgtta 3360 : tggctaaata 3420 ttgataatta 3480 gagatatgtt 3540 aattttaagg 3600 ttttttaaca 3660 atggtcttag 3720 tctctgcata 3780 tcaaaacagc 3840 ttgtttttat 3900 cctctatttt 3960 tgggagtcca 4020 agtaagtaga 4080 agggctttgt 4140 acagctgttg 4200 ccctgggcag 4260 aggcatcccc 4320 ggcttcttcc 4380 tagttcttca 4 440 attagttgca 4500 tcaagctcaa 4360 atttatcgaa 4620 ctgtttcaaa 4680 tcttttcttc 4740 ttccatattt 4800 gctatgagct 4860 gtgactgggg 4920 gttacttttg 4980 ttgagactgg 5040 ggaggcctca 5100 gcacatctta 5160 w укиауаu w _ J-.-, V atgattcaat 5280 acaattcaag - 5340 cttgaactat 54O0 ggtctggatt 54 60 caagcaacat 5520 gcagaacaat 5580 ttatattagg 5640 attgactcca 5700 tatattctcc 5760 atacattcaa 5820 gagggtaagg 5880 atatatatat 5940 ttgaaacagt 6000 cagcctcctc 6060 tacaggcgtg 6120 ccatttcggc 6180 ccaaagtgca 6240 ggattcccat 6300 ttctaaatgt 6360 taaaagttgg 6420 <- <-+¦ ? an+-a*--t- 6480 cacatcagca 6540 agactgcaaa 6600 ttgggatgct 6660 atataagtac 6720 aaatataaaa 6780 taacctatat 6840 tcttgacacc 6900 ttacattgta 6960 ttaacttatt 7020 cagagacagg 7080 - 126 - 008252 gctcagtgat gattatattc acctgtcatg gggtagagct acaatagtga gtattctgtg 714 0 gatggtccta tccagataaa agaaggaatt ggtttcagga catagtctgc atcttactag 7200 gtggggaatg ttggacaagt tacttaacct cttctgccct gtaaaatagg gataaaaatg 7260 gtacatatca cagagtgtca taggaagact aaagtagact attctcatcc tcacacctaa 7320 tgagttctca atatttaact gattataatt atctttacta gatacataat taattaaata 7380 aattaaggaa ttatagagca aatagtcata gggtcacaca gatagaaagt gggaaaaata 7 440 ggagacaaac ctatgtctat attcattcca tggcattaga tttagattac ctccttttag 7S00 tctggcaaaa cactcatgat tatacaatgt aatttaaaaa tatacagtac tttgctgggt 7560 gtggtggtgc atgcctgtgg tcccagctac ttgggaggct gaggtgggag gatggcttaa 7620 gcctgggaag cagaggttgc aatgagccga gatcacgcca ccgcactcta gcctgggctg 7 680 tagaacgaga tactgtctca aaaaagaaaa aaaaatacag tactcacatt ttactttgct 774 0 agtatatgct gtattcactg gtgtatgctc tcttgcatgg agactttcat tgggaataaa 7800 atgtttgaaa ctattttact atttcttgca ctgctttgag atactaccag caaagctatt 7860 tctaagctaa gacgctatgt cctggcagga cttctaaaga cttaaagcaa tgctgtagct 7920 gtgtatttac aactgactag agcagggatc agcaaaccat ggccagtggg tcaaatcctg 7980 cttactgcca gtttttgtaa acaaagtttt atcgcaactc cctgcccatt tgtttagaca 8040 tggtctgtgg ctgcttttct ccaacagcag aaaagttcag tagtttcaac agaaaccctg 8100 aggcccataa agcctaaaat gtttaccatc tggccattta cggaaagtta gccaaccccc 8160 agactagaat aaaggtagga aaaaccactc agcccaagct tcttttttga ttctatcttc 8220 ccacagctgc ctagaatcct ttgcagtctt tatcctagga ggctcagagg ctataacaca 8280 tctctaattt tgtctagatt tgtgagaatc catttctcac aaatccagtt ctggccctct 8340 ttccctgact cgatgttcct tgaactctaa acatgaattt ttaaaagaag gattacctgg 8400 gatgggtata tccctgaaag tcacctcctt tctgctgcct ctcacaaata cacaggctat 8460 gtgccttaaa cctgtcattc actttaagga atttttgaaa agtggataat aggttagaca 8520 taaaattcac tgaatctggg ttaaatttaa tcagtgagta ttctttagaa tggaagcatg 8580 agcctgaatg attaaagaaa ttgcttaaaa aattagaatc aaaatactaa atatcatccc 8640 ccccccaatg ttttagctag aaaaattact tgaaaatgag ccagagaagg agaaaaataa 8700 ttatcttaca gaaaactaat aaaaatacaa taaactatca tttatttaaa tacaataaac 8760 tatcatttgt taaaatacaa taaacatgca aaagcgctat cattgatagc acttttaatg 8820 tttaaataac ccactgatta ctttctctag tggattattt gcaagggtag agtaagaatc 8880 gttgcttctc tggacacata acccatttgc catgtgacat tgcctccctt ccaccaagaa 8940 gtggaatgta tttttcctcc ccttaaatct gatgcctata actttctttc cccttgagat 9000 aatggcagag tggcattgtg caagttttgg aactaaggtc ccacgaggcc ctgcggctct 9060 ctctcttgtt tatctgagta ctgccctaag atgaccatgg agagaagccc agtctagcct 9120 agaaaaggag aagaggccac atggagaact gaggtcagcc agccccagct gcttgacttc 9180 tgaacaaagc cttctccgac tatttggccc tgttagaccc agctgagatc agcagaagaa 9240 tcacccatct gaccctagcc caaattgcct tctgcagaac cacgggccaa taaatagcag 9300 atgctttaac ccaccatatt ttgcagtgat ttgttatatg gcaacatatc attgaaaaac 9360 cctcatttta ccaaagaagg ggggtcatga cattaagcaa ctacctaaac ccacaatgta 9420 aatggcaagt cagggattta gatgcaaatc attccaacta aaaatgtata ctctaaatca 9480 atgccctgta ctatactctg tattttccaa aaactaatta gattgtgtca tcagcatttc 954 0 tgaagacaga ggtagtcgtc tgggaagtag agttttctca accagtattt tgcacttgtt 9600 gggatttttg agatcaggaa tgttttggaa aacctgagat gtgcagtcac atctctgagc 9660 agtgttttaa gtttattcat tcactcattc attcattcag caaatattta ttgagatcta 9720 ctatatgtta ggcactatgt ttaccactgg aaggcaggca ccaggtaagt acaaacacag 9780 aaactgagat ggatggcatt gcattttcat ttgattacaa atttattctt gcttataagt 984 0 tgattaccta ccttattaag agctgatttg tttcaagttt tagaactcca attgtgtcct 9900 taaatttttg tctagcttaa aacatagtgc tagatttcaa gaagtaaatt gtcaaacagg 9960 tttgctgttt agtttttgta aaaaaggaaa cttgtaggca gaaacaaaaa ccaaaataaa 10020 taaaatctaa attatggtaa taactaaaga tagttgaggc attaacagag tttgactgaa 10080 ctctcaagtc tcaagcaaat ttctaaacta aactgttttc ttctgttaat cagtttaaaa 10140 gggaatacct tgtgctaatt tttcttggtg ttggtaacat taataaagct ctgtgaaatg 10200 aggccagaag gaacagaaat tgagtcaaaa taatgataac acctcttaca tattcttacc 10260 atcttgtctc ctagccacac acaccatgtt taaacctaaa aggagagaca cctggagggg 10320 ttaattagct tgcagctgtc tgtgactagt ctgcagagga taaaacggat atcagccgta 10380 agaaagcagc tagcactagc ctggatgatg aacaaagaaa agaagcagct gctgaggaaa 10440 agcacatttt gctgaccttt gaggagggct taagaggccc tttgccccct gaaaatggga 10500 cctgggaaga aagtcttgat ggcagacagt acaaaatcaa gggacagcag ctggctgttt 10560 tgtgtaaggg gcaaaattca agaggatttg tgaatcattg tgatttacag tgaccattaa 10620 ctattgtaat aaggagagaa aaatcaacat ccattcagtg cccacattgc aaattgccaa 10680 gcactggagg ggtactttca cctaggccaa caaacttaat aagcagagaa acccttcagt 10740 gttttggctc ctacagctca agtgtactag gttggcaagt ggaagttcct ttggtcatct 10800 ctgtttatac acttgtttga gatatttcat aaagtcagcc catttgttac tttatctgta 10860 - 127 - 008252 gtatgtgctg ctgtacggat ggaaccagct agcccttcat taagagctat cattcgcttt tagtgttgga taggaagaga accccatcgt gatacaaaat ccaaatcatg ctaacttaca ttgttaaaaa ctgataaaat caaaacagaa gttaaatact gttgagacgt ttatgaagta acttaaatag ctactaagtt tcagtgcctc atggcagaag gacaggcagt atgttagtac tatttttttc taaccaataa gactttatgt taatcatgca aggggacctg gcattgtgtc gccgtaaaat aatacaaatg attattcatc attaatagta gattataaga aatttttaaa ccaggcgcag cacctgaggt aaaatacaaa tgaggtagga actaaatttc aacaacgctt agtggtttct catcaagtcc accagatgat taaaaattgc atactgatta attgtacatt cgtactttgt tcacacagtt atacccacac aggagaaatt agaatggata gtgtctgcct cattaattct tccggtcctc aaaactttga tgttgtttac actatcacgt ttactaatac tctgtatttg aaattgttcc caatatttga tttcatgtgg ttaagccact tgtccagaag gctaaaaact ttgtgacaaa gaaaactggc agttctggcc ggaagtcaaa ctcagcccaa caatgtgcaa agtgaactcc agggatgtga ctaaagttgt attttaaatt acccactaaa tgttgcacag gttacgagga tacaagccat aattatacta aaacgttgct tctacataga aaaagttacc gcacaatatt atgtgactgt ttcctactgg ttgagcttac tctgctcttt ctgaggaaat tggaaataga gttcacatag ttgtaatttg gctaaggtct agtctctacc aactttttat tgttaataat ggtttattca tggctcacac cacaagtttg attagccggg gaattgtttg aacctgggcg agagattttt atttaaatca aaccttctgc ttctttgata tgttcttttg agttatacac gtcaaataat gtcatatata atagatctgt aaatacaccc cattagaaaa ttttattttt tacagttcat tttctttaaa tctccggtat aactcacttt atattattta attttatttg tggagtatta aaaatagaat tgcatactcc tataatgaaa aagatgcttg gagaagctat tttaatcctt ctcaataaat accacagcca acaagacagg agggcagtca ttgtccctgt aatctcctta aaatcacaag cattcacaat aggagctctt tggggaacaa ctggtgttac atgttaactg tagtatgagt ctcaaaatat tctaaccttc ataatgcata tcccatggtt ggaacctatc catttgaaaa tgccttccag ttccctgtct attccattat tatgttgagg ggattaggct acaatattaa atctaagaac caaattcagt tttggcaaat gtaattatga tactattcca attaagtata ggtaagtcat acaaaataga ctgtaatccc agaccagcct catggtggtg aacccgggag acagagtgag aaattaaatt caaatgatag tacgatatta gtctgtttaa gaattaatat tgcttagcct ttgcacacga tttgtgcatt gtgaattccc catgaaatga aaatcctata tccatgtttg atatttcttg ctattaagac ctccaagata catatngntn tatgaataaa gtcattctac atatacttgc tactgggtat caccagtact cttgtaaatt ccttttgata gaacagacag taaggaaaag taagtattga atatcatact gatggcctct ggcaggagaa ttgcagatga agctgatagg cattcctata tgcttcaaag caaggagaac attgtactta aaagatttag attgacatta tgtttctatc tctaattggc acagtaaatt ttttctgagt ttctgggtga tcagtaactc aataccttat aaaaggagat tatcctgaat aaaacctcta ctgtaggcag aattttcaaa aataaatttt atattgtaca acagattgtt cttgacaatg gttatttaag aattaattga catatgtgaa gactgttatt gttatcttac agcactttgg ggccaacatg ggcacctgta gcggaggttg actctgtctc agtttcaata aggttagaga gacttcaaat aaatattgaa aaaaacatat agcatgtgta tcatttacac atcagttgat tatattccac tatccagtta agcctctagt acttgtaaag aatttaaaaa agcaaagtct acatgattat tnnncnnnnn tatacaccta aattgattga atgagtctcc gcacattttt aagaatgtat ttagccaact gtattccaaa gtgaagtttt gatagcaagc tgggacgtat gaatgggcaa ctcaccactc agaaataaag catgattgta caacttcagc caccaataac agagtaaaat tacaaaccac aaatatattt tgtttctttt tattttttgc atttactctt ggtgaccaaa tttttaccct gttaatgtca tgggtgttta ctatactact tttccaaaca tgcataaatg gcatggaaag tttgtatagg aggataagat agttattgac gaagaagatg tatgctgaat tgttaatgtc ctgagacatt ctaagaaatt ctgttttata ataaaatcat ttgttgaagt ttaaaaatgg gaggccaagg gtgaaaccct attccagcta cagtgagccg aaaaaaataa cttctataat tgaaactaaa ggaaagcaca ttaataaact actttaaagt tgttatactt atctgtattg ttgcacattg atgcatggac atgaatacaa tttcccaatg taaaacttta actcaattac tttttatgat gagtttaatg nntacataga ctccaggtaa tgccttttgt ttgtgtacca aatttttaat ctgttttcct aaatgagtgt ctgaccaacc ggtattgcca ccaatgcaac ctcaaaataa aaactggaag ctattcaaca ggtattcagt tatctagaaa aaagtctcag agacagagag acctaggaat tgctagacaa attgtgtaat gttcttttta tttcgtcagt ctacaagcca tgtcatacaa ctggttgcat tgtgaaattg ttctgcttat ctacaacaaa gaagtcttca cataacttac tcatgagact ttataaattc ttacatgtca tgtaaaataa gaattatagc taatagagtt aacagattat tattctagtt tggcaagttc tactgataca attattattg gattttttag atgcttaggg cgggcagtat gtctctacta cttgggaggc agatcatgcc aaaataaaaa agttaaaaca catatgagat tcaaacgtta ggaaacattt aaggaatttt aaaaatgtgc ttcacattga cacaccagag atgttcatat aggtttttaa aattaatcta gtaataaaga ataagaaata tatctacaat tatgacattt atatatggag tttagtggtg gtgtgtgtga tgggaatagt atatacttcc cttagcatgt gatttctatt - 128 - 008252 cctttatagt cactcaggtt gacttgtgtg attttaaggt cagaatcgta gacaaaagaa atcaagaatg atcaagaaat cctgggaaag gttagaatca ccattccaag aacaaggaag catattccaa ggcagtaaac attctcctat ctgtactttg aatgacttca aggggaagca tagcttatat agttgtaatt atacgtgtaa agtacactaa aacttactgg aaagaacaag tgtgtttgag gaatagaata aattatcaac cattttcaga ttcttctatg ttaccttttt gaacgtatgt taatcaaata ttcagaaggt agaagtgtca cagtaggttt tgggagaggc gttaggacta ctggctgtac agattctgca caggtatttt acatccagca ggtttattga taatgaagtc attctaaaaa ttttgagttt agttccctgg ataaaaggga aagtcatttt cttcacatag ttccaaatgc acttgcagta ctgaccaccc actcactcat aggctggtag ggtgagaaaa caccctaagg aatttaaaac ccacaaagaa tcaatacgtt atgcatatat cattataata ctgaggcttt agagggcgtc aactgatgag tttttcttct tgtgtctttt tataggattt cataggattc aaatgtggaa actttgtcat taacggtgtt tattattaca aataaaagga aataaaaaac gatagagctc gaacctgaaa tgtctaatgg tttaaaaacc cagtattatt gaatggaaaa aaaacaacct tttatttcag tgaggaaact gacttgcttt ataatcatga gatgccagga attagttaac attgattttt gctatttttt tttatataaa gccatctaaa caacaaatgg ttctcatcat accaaaaaaa taacaaagta attgtgtttt ctccagcctg ggcacaggat tgcaggagga gacaagccct cttgggggac aaatatactg ccctccagtg gcactttacc gtttgaagag ttcttcacta tacatcatgc tagggctttc ggtggatttc catgaagtac cagaaattga ttaagatttc agcagcttca tgcctggtct cnaccgcagc gcaccccgca tgaaagtcaa acttgggcaa attctgtgac ccagttgagc aggatctccc aaaaaataag aaagaataaa aagcaaacac cagagtggct catgtttgag gttataatca acgaattgct tcttacatat tagagttaat taaacagaag tttcacagnn aacatcggca gctctgcaga atggaaaaag gggatagatg aaaaaaataa tgaaagctga aactggtaat gatataaaaa ttctttatat gagcatctca tgtcccagaa taatctaaaa aatgcttagc catggttttc gttattgttg tgttcattct agtgctctga tatcatggga aaaagaaaaa taattcataa agtaaaatga ttcagccttg ctgtatataa aaaggtaaaa tcacttcaaa tatttaggcc ctgagtattt ggtgtcctgt gaagcaagaa gcagagaggt gtgaccaaca ttcctccatt cagttctcat gtacccagtt cacaggtggg taggcaggga aaaaaaagat ttaattgtat ttattggtga cttagttgca acatatctgg tgtcaaatag cccctattca ctgtgttcta agccacagca cagcgtgtct ctgctcccca atgctgcctg acgtgccact aatgggaaaa aagtatatct tcctctatta tagataatgt tgcacataac tgatgagcca cttggcccat aaaccacaaa agttaagcat ggttcacatg attggtatat gaaaggataa ccatgttcta nnrmnrmnnn tcattgaatg ctggcaaatc atatgcgaca tagtaagaga taggcgagtt ctgcagtcac ggacccttcc tagagtcttt ggcactgcaa agtgtctgaa tttttaattt tccagcttaa aaatttcttg taagatcctt ttgtaaattg gtgctaatag tcacccagga aacataagaa gacctattta attttatatt taatttgata tttgttcatt aatatacatg cacatgcatt atcatggttt aatattttga cattatttaa gtggttgagt catcaaacta cctgataatg gaggaaagag .ttgtctgggc tcatgtgccc tattatttct tgtttattag tttctgtttt gtgaggacaa ccacaaaggt attatcaata tataattcta gaattaatta gtcttccata gaaatattag ataatgttca gggagccggc ggctgtgcac cctggctcaa ccaggccggg ggctacagag ggggtaatga gtaaataatg tcacagggtc ttttagcttt atttatgtgg ctggtgagga gtagaggtaa atcacaggag ttttttatat cttcgcccat ttgacttttg aatgctggag aataaggatt acatttatgg gtattgtcac tcattgccct agatattgta agcagatttt cattgtgggc taacaaaaac ttatactcgg ttttcctccc ataatagcac aggtacaaag tccttctaaa aatgacaacc actttgtatt tatacctgaa aatacctgaa gcaggaaaga gtgagacatg gaatttggct aaaatccaca ggttagtttt tatttcctga atacatatat acttgttgat tcataaatgt acgtatcttc ctatgataaa agcagaaggt caaatcagtg aaggggccaa aggttccctg tttctgagca tcatccacag cttgagatgc agtccattca gtttttattt tatgagtcac tcttgcgcag tttaaaaatt attgtagtgc gcctcttcag catttttctg gctgaaagtg tttctgatga tgcttaggga gcccatgcca catggccaga ccttggtggg tgggcagaac gtttccggct agggatagag tacttttgta atcccggaag aaaaaaaagt gttatgtaag aacataacct gggtgtgaaa tcgctaaagg gtttattggc tttccttctg tctattattt gcctagaatt tggcttgtgg gcaatagaga ctataccatg ttcttctaac ttaatagggt atagctatcc atctgtcagg ccttttgaga tgtcttgaaa ccactctaat acagcaagaa ctaacaacca acagaaatat agaggttttc aattttagtg aagttgtcgc ggtagatttt tggacaatag agatgggcag ttagggattg gatgcttagg cacagcagtg tatgagtcca aagaaaatcc ttaaaatctt ttgaaaatgg taaaaacatg atcgctttga ttcatctttc cttccactat gaacaatagg actatagcag ctttctatat tcctgctaag atttgagctg actcataaca cttttgtcca ttgtcagctg tgtcataatt taaacatcag tttcttgaaa cctactaagg ctcaagagcc ctaggtgtct caatggaaaa tcctgaaacc gttcccagag ctccatgctc catgggatcc gagcccagcg ggtgaagtga cagattaagg tactacaaaa aactgacatt ggaataatct cagaggtaag tcctaaatct gaacaggaag agtagtttaa - 129 - 008252 ttggttatat atcacagagg cataccttat tgtccttggt cagatcaaaa aaatgtcttg tatcanaaca gtaaaataag caatttcagg agaaaatcct ataaagaatt tatttttaaa gcatcatgaa ctgttctatg ggtcacctgg aattttctcc aaaagggtat agaaaaattt tattttcagt acatgaaata gaataaatct ataaaatgtc ggctgatcga aattagacca tgatgagaag atctaagtat acaactgtca acatggaaag ggagattttg ttggtagctt tgtttcattt gtgtttgtgt gatattaatc tttcttgaga gctgtttata atagataagc cctctaaaga ctagtgatcc aagaaatctg cgctctcttc tcgttttgac tatttctcat aataatgatt attccttaaa nnnnaagccc tcgccagcat gaacttaagt aagctttgct aaggctgaat tataatttgc aatagagaac gtggtatgta gtgactngaa aggaaaatgc catatgtttg tttaatcatt atgtgcacaa ccattaactc caccccacaa ttcaattccc tgctgagaat tttttatggc tcgttgttgg ggaggaaatc ccatcctgag gggtttaaac gtgcactttt ttcttccttg ccttcatctc ttattattta tggaagcaaa aaatagccat aagatgagga tacagaatag atgaaaaggc actacttaat ataatctaca tcccaacctt ttgcaatgta attcttgaca gatgtttgaa aatgacagaa aataggtaat ggaatcctag tgggtactta actgagaggg ttgtcatctg gacatttatt cttagtctat cttgagtatt aaggcaagga gcactgaaga aaaaggagga ttaccctctt ttactatatt attatgatta gattatatat tcttaaatta ttgtgcagtc ataaaatatg tgtaatatca gttggaactg tttaagggtg aatttttctt tttgaccaat aaacatatta gcctctctcc ttccttagca tgtcctcttc acaggcaagt cacctgcctt caaagcctta attttgtatt tgagtgattc cagctagtcc ctattcaggc cacctattcc ttttccgtgt ccctatatac tgtgcaggtt gtcatttagc cagtccccag acctatgagt gatggtttcc tgcatagtat acatttgggt tggagtgatg cagtacaata caatgatgct cagatggaga actttcatag ataaaattat tacaaataat cttaaatgat ttaatccctc gatgaagcta ctggaaaatt atttaacaat tatctgcacc ggaagacata tggaacattt tacaagaaat gggaatcaga cttaagaaaa agtcaggaaa ttagatacag gaggtcgccc ggactcctct attagtacat ttgctgtgtg gaatagcatt agactaagtt gggtgacttg agaggagaat cagatcagga gtcaagtctt agggtaaaag taagaaacac gacactagtt taaatgggtg atgtagaatt agctgtgaat aactatcttg atacatgcat atgccttccc ttcagattat tctaagttaa ttttccctag attaaacaaa atcgttaacc tctcactgga ccgccagtgg caacagattt tgtacaggag ttagtgggag catttgaaaa ctgggtttat ccaaatatga tgtaaaatat tacattagtc ctctgattga atttttttat tgttacatat attaggccta tgtgtgatgt gagaacatgc agcttcatcc tccatggtgt tggttccaag ttagaagatg ggaaattaat tacgagggag ttggcaagcc catttccaga ctctatgtcc tatattttct tcagaacaac gaaaacaaaa aaaaaaatat aaggttaagt ctaaaaacta tatcagtgaa attttaaaat gaaaagcaag ttaaaaaaga gtgctgacag tggaaanttt gatgaatgtg tgggtggtgc taaccagtga aggcaataaa aatttaatct ggagaggaag gtacacttta tgattctttc acttctcttc atatcttgtc gcatatcaga accaactctg agtctggtat cattagatca taaaagcata gtgtacacat tctcccttaa tcatctaggc ttgaaattaa gcatacatac agtgagcctt ctagactttc aaccacattg cattacagtt taatgattat cttttccttt ggtacgttca gaaccaacta catatgcatc tctgctgctg gatctttgac attgtcctgt gtgggtgata ttgtatttta aggaaccatc tggcccttgc tgcagctagc tttattatta gtatacatgt tctccaaatg tcgccttcct ggtgtttgat atgtccctac atatgtgcca tctttgctat caattgacca cagggatccc aagttagctt aatggctaag ggatgtctag tcacctgtaa taaaaagact tttctaaata tttggcttcg attgaacact gatcaggtgg ctatggttac ttctggacaa gagggtgaat tttatgagta agatgttcta tttttgtaca tttgactaaa gttaattttt gtgcatttca aiauaayyi- catcaaatac gcattttttt ttgtaaggat gaacatataa attttccaga atggctaaga ctttgctgga gaaacaaaat cagtatgctc ggtctctctc tgaccttcag atttgctaat aggacaaggc agagtggatt ctttcattag caagtcgaac ccatcactgg taggaacaaa aacactcacc ttctaagtat attcttatta gattagtttc tcttactaag tttctacata ctactaccat gtgctactag ataaatattt tgctggctct tagaattaat gcattttgtg cgttaattta ttatttttgt gtnnnnnnnn ggcagcatca acaagtacac caaatgacac ctgggtttta atgaaattct gcatttgttc ttgtggaaat ctagagatag tgccatagaa tccaaaattt taatgcctaa tactttaagt gccatgttag ctatccctcc gtgtccatgt tttttgtcct aaaggacatg cattttccta tgtgaatagt а L Laia Lac L. attatattta tctcataaat tagagtggat actgaacatt gtacaggtat tttccatcca cctcanataa gttcttgaac tgtaggtgaa acttatgaac ttagattgag tagcatgatg atttaagttt ctcatatatg tgacagagct acactgcata tcatttcagg aaaaacaggc tcagtagtga atgctttcta atgattactt attaaagact aaagttttct gaaacttagt ttcttcatct atctatactg taagaaggaa tgacagtttt gattatttta aatatgatca ctggtaatta attacgtttt nnnnnnnnnn ctttagtatc tctcaggtat gttaaataca atgaaactta ttttgttcag gaatatcgag aagaggtgct agaattcaca taagcatatt cacatgtact tgacagtttt tttagggtac tgtgctgcac cccctccccc gttctcattg tgcgatagtt aactcttcat atccagtctc gccacaataa - 130 - 008252 acatacatgt gtaatgggat ccaacttcca tttctccaca ggtatctcat tcatgtgttt cccacttttt ttctggatat cactctgatg gtcaattttg ~-/--. <"- 4- --4- aacacgtaag cagtttcagc atcctttccc tggcattatt gtaccatgct gcctgcagct ccatatgaac cacatagttg tgtctctcag cgctcaacta atgaaggcag aatctctggg aagagaaagc aagcaagagc gaactgaagg ttttttgaaa gagaagaatc gaaatgcaaa ctagaagaaa gttgaatctc gcatgtgtct ggctgggtca caagggttga tcctctccag tgtggttttg tttggctgca gatggggttg tagccgtttg acggtttctt gcttttgttg 4- -t -* 4- -*-.4> 4-tyaa .yy La u tctttaatcc tttctacata catttcttgt tctgagggct gttttggtta ttgctctttt tttaaagtag gaagtaaagc accacagtgc catggaaact aaataaagat acacattcaa aggaaagatc aaacacattc aaatagagac agatcaacaa aaatagacgc ctaccatcag tggataaatt tgaatagacc gaggaactgg cctaactcat aaaaaagaga atactggcaa ttcatccctg ataaacagaa gacaaaattc tatctcaaaa caaaaantgg ctcctattca aagggtattc atatatctag agcaaagtct aacagacaaa ataaaatacc aaaccactgc tgggtaggaa antgccatcc attcagttct gtttccttcc t-arrt-l-t-.-it-t-rf --="------J aaaatccttg taagtaagta agacaaagag gtgtcccttc agagtgtgca tgctctcctc agggaatacg aatgaccttg gcgagacaaa caatgacaga taccattcct tttatgaggc attttagacc accgaatcca ggatgcaagg ccaaagacaa aacagccctt taataagagc aagcattccc acataatgtt aattaggaaa aaaaccccat caggatacaa cagagagcca tagnaatcca tcaatgaaat gaatcaatat ccatcaagag gactttattt tggtcaattg annarnra r> f--ээ------- ctttcatgga aaataaatag aggggaaggt aaggaaaatg gggcatcggg ctcatatgag attggaaagg gctattcctt actgatcagg agagactgtc ttatagcagc aatggtattt actagtttat cacctgttct atttgcagtt taaatgtctt tttgtttttt tcagatgaaa ttgctgtgca ccattgcttt 4- ~ ,-.+- -.-.-,4.4. L.LjvvLCiyVJLL atcttgaatt tggctagcca ttttgtcagg ctgttctgtt ctgtagcctt ggcttaggat ttttttccag actcctcagc aatcaaacta gaacaacctg gttctttgaa agcagtgtgt caaaattgac aaaagctagc acaaaaaacc aattgataga aataaaaaat agaatattac cctcgacacg aataacaggc - > --.-чи- tctgaaacta cagcatcatc aatatctttg gcagcacatc ctggttcaac aaaccacatg. catgctaaaa tatctatgac tttgaaaact ggaagtcctg agaggaagtc tgtctcagcc aatcaatgtg aatcatgagt acttacgagg aaaagaggat cgtgaaaatg atttttttac ccattatatg actatatggg a? n? rtfb ancr --~Э " ~D--- gtttacaaat catgctacat gtgagcagga tagtagttgg agcagccctt gaggccaaca taacaaggaa ttaaatgaga acaactctgc ctaagagtcc atgatttata ctagttctag agtcctacca ttcctgacat ctctgatggc cttttgagaa tcttgtaaat aattttctcc gaagctcttt tggtgtttta atcctttggg atccctgagg acagtgtaaa tttaatgatt cagtgatgat gtgtctgttc ttgtttgagt cattttatag agtttaatta gacatgaagt aatttttgta gttttcccag tttgtcaaag ccattgatct gtagtatagt tgacttggtg ttctgtgaag aaatgtaaaa gaactcagga ctcctgaatg accaacgaga agagggaaat accctaacat agaaggcaag tttcaaaaaa cctctaacaa gataaagggg aaacacctct tacaccctcc tctgaaattg ttccaatcaa ctgataccaa atgaacattg aaaaagctta atacgcaaat attatctcaa actctcaata aaacccacag ggcacaagac gccagggcaa aaattgtccc caaaatctcc caaaaatcac gaactcccat gatgtgaagg acaaacaaat gccacactgc acattccata tgaatgtgtt aaaatatttt cstttgstcs tctaccaaag agtaataagt attgcagttt gaaggggcaa gcaaaggcct ggtcttgaat tggatcacgt tggaaagccc ttgctgggca aggcgagaga yy yV> LLLLOLLJkJ taaggtgtaa caccatttat aacagatggt atatctctgt ttgaagtcag atgcgggctc aaagtcattg gaacagaaat ttaagaaact aatactgggt acaaagacac ttatagcact cacaattaaa aaatagctaa ttaatgaatc gactaataaa atatcaccac acgcaaataa caagactaaa tggcaataat 331 Ct"CC3 tagaaaaaga agccgggcag atgcaaaaat tccaccatga caataaatgt tacatgcaga aattaggtat ccaatatcat agggatgccc ttaggcagga tgtttgcaga ttaagctgat aagcattctt tcacanttgc acctcttcaa ggaagaacat ccanggtaat tgtgaatatc tctttactat atccattcaa tatataccca aatcgccaca agtgttcctg ggtgtgagat gagcattttt atatccttca tcattgtaga gttccctatt aatcccattt ctttgcccat 4- 4- 4- 4- -i fr~-4- <-"+¦ ggaagggatc taaataggga tgtagatatg tttggtacca gtagcttgat ttttttggtt gtagcttgac tataacaaac cactcaaaac acataatgaa aacataccag aaatgcccac agaactagaa aatcagagca caggagctgg gaagaaaaga cgatcccaca actagaaaat ccaggaagaa caatagctta nai/-irf4" araart gggaatcctc agacacaacc cctcaataaa tcaggtgggc aatccagcat aaaggccttt tgatgggacg actgaatggg tctctcacca gaaggaaata tgacatgatt aagcaacttc atacaccaat ttcaaagaga ggagaactac tncatgctca ttacanattc ctattttctg ttttctatca tattcaaaaa raanant-iraa дддг-апаАаэ gaaacaggaa gtcaaggaga cagataaggt gtcacatggc tgagaaggaa gaagctatgg agagccttcc gtggagcatt gagaatagaa tggcagtggc ataatgaaaa aaaggaaagc ggccatgaag tctctgggga gcatgcccca agaaagagaa agggcaacgt ttgagtagag tcaagggaag ttggacaaag - 131 - 008252 gtggtagcag tggaggtcac agaagatgct gaatatagtg ttcatgttta ctttatggga 26040 tttactaaga gattggctgt gtgttaaaag gaaaagatca ttctaaagct tctggcctca 26100 gtgattagat agacttgcca ttaacttcca gaagagcatg tttgagggga aatatgagga 26160 tctcggtttc attacatgtt aagttccagt aatgtgatta gattgaaaat gctatagctg 26220 gtagaatttt ttcacttttt atgtcaaaac agaaaacatg attgaataat gaaaactaca 26280 catgctcaat ttttttaaaa ttgttttctt tgacacagta aaggatcaat gagacttgct 26340 taattcttgt caagaataca aagtcaacaa tggctgagat gaaaatattt tccataaagt 26400 aaaatgatca aatttgcaaa agcttggtta catgatgtga atttcactca aaccacaagg 26460 cattacacta gagtcttgct gaaatttaag acagactgtc ttaggaatta tttgccaata 26520 gacaaaatga atatggggtg gttgatgtgc tgaagtggcc aatgagcatg ctttgattga 26580 cagatgagga aatgtatccc tacattttat caaactatgt ggtgtctgat ttatatcact 26640 gaggtggcca ttcattatgg tcatgaactg tgaggtcaaa gcacttgcaa aaacttgcca 26700 aaatacttcc tattccagta ctaaatattt ctcatatttg tttgcctaga ataacaagat 26760 gcaggcttgg tagtcctttc aagaatgtga atttctttgg tatgtctgag tttagtaatg 26820 ttgaattgtg tgtgtgtgtg tattattctt aagactttgc ctgtatttta acattttatt 26880 tggtaaaaat agcattattt tggcaagatt tttaaagcct tgaaatggga caaattaaag 26940 tataatgctt tttcttgaga aataatttat tgctccatgt tgttgaatca taccattagt 27000 catgaaacca aaaccctata tttaaaaagt aacaatctca tattcaaagt atctcctttg 27060 aacatttcaa agaatcttaa gcaaatcttt atgtgacatt aaattgtaca ttttaccttg 27120 ttattttaga tctatgggtg gagttcattc ttcgggctaa gtgtaggtat gaagtgtgaa 27180 tttctgtgtc ttggtatgcc agtaacttag ggcaaggtta gtagagtggc ttttgtgaga 27240 aaaaaagagc tgttttatca gacatcagtt ggaactttgc cttgttttgt ttccctgaac 27300 cgtccaatct gcttcatata cttgtgtgaa atcctaaaat gtctgtatgt caataatttt 27360 tagttagcat attttttcca aacagacatc cttgtgaaca tcagccgttt ctgagaactg 27420 ggcccacgga ccatagctac tttgagtctc agtctatgta atagttgggg taaagtaagc 27480 tttaaagaga gaactcacaa cacaatggct caagcaagac acaagttgat ttctttctga 27540 agctgccatc ctgggtatgt gggtatttgg atgagctggg ccccatgaag tcatcccaaa 27600 gtgcaggcct ctgtggcatc tctgccaaac tcaacactta ctctccaagt ctgtccttgg 27660 cttctctatt tctagctggt ggaagggaag ggaaaaatga gtccagggaa ggagtttttg 27720 tagatggctt gggagttgca tacatcagga agaactttgt cacaaggcca caactccttg 27780 caggaaattc taggaaacat attccctcac tgagcagcca tgcgcccagg aagaagagaa 2784 0 gaatgagtct agagtcgcaa gcagcaatat ctactacagg cttaaataac caaaggacaa 27900 aatgcccagt ccattataaa tatattccaa aaatgaagcc atgaaatttg acttcattta 27960 ttgaagtcaa aacaaattta ttgttatgaa ctaatttctc tattttatat ttctcaaatg 28020 attacatttt taacacagaa caaagccata gttagaattt cagtaatcgt atggaatatc 28080 caaaaggtac acccaataca cccaaaagct aaaccagagt cactgctaaa ttttatattg 28140 ttctgattta cctaacagat tagtgaaaat aattaaaatc ttgctgcttt ttaaatttta 28200 aatatcggat tttggttaga actctaagaa .tatttaaaac tatctttacc tactcttgtt 28260 ggacaatcat ttgaggagct aagagtgcag agaaggcaga gaaatgttct cctcaaccca 28320 agcaaaagca cagaagaact attgctaatc caaaatgcag gaaggaaatt tcacccctcc 28380 tcccttccct tttcttgggc tagtctgttt aactttaaaa gcagaatttt tgctctcaca 28440 aagtatacca gacaggaatt tgttcaaatt cctcaaaacc aagccaagaa aagtttcaga 28500 aagttcagac atgcatggca gagtggaaag aacatagtct gtggaggcga gcacggattc 28560 attggaatgc cgctccagac atccactctg ggtgaactca gggatgtgtt tagtctcatt 28620 aagccacagt cttttcattt ttaatgtaag agtgataaga acgacttgcc agtgtgttgg 28680 tggagaactg aaatagagaa aggcctggtg tgtaacagga tgtttctctc catctgtgta 28740 atctgtgaca gtcctaggaa gttcaaagtt atgactgttc tgggtttgtg gccaacagtt 28800 gtctgtggct tccatgggtt ggagctgtct aatggggccc agnccaccat cgctctgctc 28860 agaagctaat cataccaaaa catgaggttt ttctccaagc gacaaagagg ggctaagcac 28920 ccaaggatct ccataatagt agcaggaaaa tgaagctgag accttaccct gacttagaag 28 980 ttcataacaa taatggcccc aaacaagatg ctcatgtcta ttggttgtgg ttaataatca 29040 tgcaacaaca tgaacttcaa acccagccta tgcttggctc ttccagggac tgaaggcatc 29100 tcagagcccc accccttacc tcagaaaccg caggaaggag cctgggttct cagagtccaa 29160 gaaaatgcac ctgataatag cacctgaact tcctccacac aaagaaagcc tgatagggac 29220 aggggattaa gcttgcatgg agggcaggat ccaacccaca gtagcacttc cttgccatgc 29280 gaccttagca aatggcttaa ctcgctttct gtatcagtaa aatggtgttt gtagtactaa 29340 ggtgtcagtg aggattaaat gagccaagcc tgacccacag caattcatga caagtctcag 29400 ccactttaat gaatgtgatg atattctaac gtttgggatg ctgaggtgga ggacgatggt 294 60 gtcccacctc ctgcaaatgc catgtgaaca ctgaagctac acaagcaatg gctctaaagt 29520 caaagccagc atcaggactg gtataaaaga atcaccagga caagggcagg gcagagggga 29580 ggccaaagca gctttgtcag tgcctgcacg gctggaccta gaggaatttc tcgacataga 29640 gggtaaaaca acccattggc ccacatttct gaggaccgac gtgggagaag ggtcacatga 29700 ggccatgaaa tgggcatctg cagaggaaat ggaagccagt gtctggagac aagtggagcc 29760 - 132 - 008252 attgagctag caagggaggt tctgcagcct catggaaatt ctggccggct actcttccct tctctctgct taacatacct caacacagag aactttaaac caatgcgtga ggctgagtct cgggagctct aatctggata ttcaaagtct tttacatgtg ttgttacttt gtgtttcatc tggctgcttt tttaaaattt tagatactgt ggcaatgagg ggcagctggt gtcccctcct gctgacctct tttcctcttt cctttgccca tgccaggcac catggagttt ggacaaactt gggaagggat gcctggggca gtgggtcacc ggagaccctc ataattgtgg gaatatttcc tttaatctac tggcccaggc acaatctaca ttataaatac gtggggaatg tccttagtca aactggcctc caggatggca ctcttcacat accagcccct attttatgtt cctaggaata aagggacaaa tgcatttttc cagcacctct gcaaagcagg gggcaccatt gatgagcaga ggtccaacac ctccctgacc ttgtccagat tggtggtgac aaggataaac caataaaatg aya t_a.aai_aa tgagataaaa aaccttgaat tctaggtcaa tgtcttgcta atttgtgctg agaattacga accagtgatg ttcaactcca aatcttgttg attttgtcaa gtgcggtggc ggtcaggaga caaaaatcag caggagaatc accctagcct aaattattcc aggtgggtgg tggtctctac ctcctcatcc ttccatcctg cctgtcaggg ggggcttaaa cctatgtaac ataaaaataa aaaattaaaa tcagatttaa gaacacctac agtgagttat gccaattgct ttctgtattc atgtctgatt ggccaaatga aagacatttt tgattaagtt ttttccttcc gttagttgtt agttgttggt ccaggagaat ctgtgctgtt agagaaccac atctgtacta ctgtttctgc ctgagcacac aattttctca aaagtaggaa tttttaattt aaaccagttg aatgtggata gaagttataa acattttcta catctttgct ggtatttatc Lyyuaitiai.. ggagacaaga attagccttt aacataaggg aaatgtctat gatattttat aacacaacta agaaggcaga tttcttcatg aatgaatcta caaacaataa tcatgcctgt tcaagaccag ctgggcgtgg actttaacac ggcaacagag aggccaggtg accgcttgag aagaaataca gcctgtaatc ggtgacagag ggtgggaggg acctagatga aaacctgcac taacataata ataaaattaa atggaggtca actctacaaa accttctgaa ccatatttct ctcccagtca gctctcttga gctctggcct attccaggca tgttgtgcta tctttcgcca aaaatcaagc ggggaaaatg cgccccatca agtgatgcat gtctttgtag tttttgatct ttgtaaagag tgaaagtcag tattcttctc tatacagtac tagtttaacc aacaaatttc gcaaatacag gacagaacaa ttcagtttta tttagtgctg gtaatacaaa ggtattgttg tgattgttgt ttaaaaggct gtagatgata aagcgtccat tcataggaga aagtgacaag tcccaataag gtacagaggt caataatttt aatcccagca tctggccaac tggcagggac gggaggcgga caagattccg tggtggctca ctcaggactt aaaattaacc ccaggcggag ccaaaccctg aaggggaggg tgggtcaata attctgcaca aaataacaaa agtacaaaaa ttgaagctca aagacttatg tagtgaatta ctgcatgtta ggaatagcca ggtcgttagc ggtgcagccg taagcaggcc ttatgtaatg ctattctaag agaaaagaaa acttataatg caattcagcc aagccaaatt acattaattt atatcttaaa cttggaagtt taactggaaa tcacctctgt agagctttct tttaaactgg aattttaaac gaaaaattct tttccccatg accattaagt tattataaat taagaacttt dyetelLwtgi-y gacttgaatt ctctttttgc ggaaaaatta aaatgtcctt taggcaatag tacttatttc attaaccaag agacaacgtt tcataaaaac ttctttaaaa ctttgggagg atggtgaaac ctgtagtcgc gattgcagng tctcaaaaaa cacctgtaat caaggccagc aggcgtggta attgaagtga tctcaaaaaa agagcatgag ggtgcagcaa tgtatcccag aaagtcttta cttaaaataa aacacctggc aataccctgc aataaaatac gagtttagct gtcagtatta ccagatactt tcttgccaat agatcacagc ggtacttgaa tatgttgtct acaggcaaat cattattctg tctctctgtg gtgaccaggc aaaactcgtt tcaaaacgaa atcaatcctg tcaacagtgt agagttctgg tggcagcata aatctgtagc atgttttcag gtgaatattt tacaagcttc tgtaatatta atttaaatta atttttagtt LL'l-.ULddd tgtcgtgcaa agtatgaggt aataggccaa gtgcctcttt caaatttatt cttccaacca taagagagtt tgacctgccc cgtcaggtat attattccag ctgaggcggg ccccatctct agctactcgg agctgagatc aaaaaaaaaa cccaacactt ctgagcaaca gtgagtgcct gccgagatgg taacaaaata gacaaatacc accatcatgc aacttaaatt catctgaaat aaatcatttt tgcaaagtga attggttgat tattaaagtt agtgtttcca ctccatcagc gctagaaagg gtgcctaatg tgcccaaagc ccatattccg gtatctaatg attaaaacgt acataatgtc ctaatgacgg agactagacc ctaatcaaat gcttttttga tcttcataac tttaaaagtt tctacattgg acttaataca acattggaaa gctttggaag ttattcacat aaatttttag ttacaaatga atgacattca aattccatgt a L. LdddlJUL L. ataagattaa agttgtgaat atgaaagaat aacttcaatg caactataag ataagtaacc ggatacaaca tctcattcct ataatgccgg gccaggccgg cggatcacga actaaaaata gaggctgagg cagccaccgc gaaaagaaaa tnggtggcca tggcgaaacc gtagtcccag caccactgca cacaccaggg tagtgcatgt cacatgcata taaaaataaa aagtgtactt agatatttcc tttgtaagac gaacaagaac ttaaataatt gggatgattt cacagcacaa tatgattgct agagcgcgga tctctggctt tgaatgaccg ttgccttgca aaatagaaaa ttggggtcca ccttttcatc acatgtaagc atgttcattt cttctggttt aagaaaaatg aattggttac aaatgctgga tgaaaaatga aaaaaaacct cttactgaag atgtaaacgt cagttagtta tattagaaca gtgtaaattt gttatttgta - 133 - 008252 tttcttatct ccctcccacg aaacgtcaat ttcattcagc ctgaggtaga tatgtatgat tagtactttt aactattatc atttccttcc catatttctg cacatttatt tagacctaat tatttatttc cttcctatta ctgaccaggt gatgcagata gccaaatgct ttaacattat aataaatctt gtatgacata agcgccttag tccagggtgt taggaactgt gtcttgtctt tgtgatctcg ctcccaagta agtagagaca cacctgcctt tcttttcttc ctcactgctc ctttgctacc gctgtagttg tcctgccaat gaaagccttc attttctaca acttagaaga tggcaagttg aattaaagta aaaagtcctt tttattttgt cacatgactt cagcacagtc ttatgcctgt gtgaaggctg actccatctc aaagaaagga gaaggaagga aggaagaaga agaggtttta tcgtttagta ccacatgatc ttgagtccag tatgtcacac ccacagaacc agaaaaacat ttaatatgtg gatgagagaa catcaaaagt ttaatctggt tgtattatct gtctgttttc aaaacctgaa tgtgtgtggt tttgcactat tttgtaatgg catgtataca cagtgcctag gtctactgtg ccttcaaaaa gtgaagagat taggcactgc tcaattggca gggttgcagg taaaattaca cggatttttt caatcttggt aaaataaata ctaccttatt tgaaccatgt tggttctaat gcgaacttct taaaatatgg ttttgtgtac tttggtccct aaaattggat tgatcttttt aataaataaa gttaattttt gcatatctca ttttggacta agcccttttg gtgacaaaac tctttaggac atcactcagg tttaatctgc attccgagat taattgctct gcattttcat gaataggagg ttatgaattt cttcaatttc taaaagaccc aattgaaaag tatgtttctt ctttataaac tatttactag tgtagctctt ttttattgaa aaaaatatct tttatgtcat ctttgaaaag tcagaggaaa cttctaatta dUdULLLL. L, L gaagttgagg actggaatgt tttttttctt agtgcggtgg ctgcctcagc ttgtattttt cctcatgatc gcccggcctt ttgacctgac gaccaggttt cgcagggagt acctacactt atactatagt aactttagaa taaaaaatta tttaccaata ccaaaatcaa ataattttaa tggtttttgc gtgtgtggag cacaatgtta gcgtggtggc aacccaggag acagggcgag aaaaaaagag aggaaggaag ggaaaagaag catggtttac agctgcttta gtctgctgat gttcattatg tttttttaat tttactctta aatttggtga aaaatttttt attattagag tagattttgc cactttgttc tttgcaagtt ttaagaatta attcagaaga aaaaaaaccc ttgggaacat ttaatgtact ctaagatatg cttttctttc ctattgagct aggcttttag ttgatctcaa gaccctgtgc aaagtgtttc ttttggttgg LyyLydLwdy actgactcac aaatgacagc ttttttttga gctcactgca gctgggatta aggtttcacc ggcctcccga ttaagacaac ctcaaaggca cttactctca gcgtgtggac gtcagcatcc acctctcctt tttaaggaaa tatatgtaaa tgccaaaaga gctcaaaaac ttatgtttgt tttattgcct gagtgtgtgg tatcagggaa aatcccagct cagtgagcca agaaaaagaa aggaaggagg aggaaagaaa agaaggaggg ttcactagga aatctctgca ttatttcatg atgtccacct taaatacatt agctctcagt tatagtaagt gctaaaatct actgagagtt gacctcaaat ggctgctgct gtctcatgat ttctgtgttt agcttaccca ttgtcttggg ttatttcatt cttattccgc gtcatcttac ttgttaaaac tggcaaatac tctttaaaga tcaattcatt agggcaaaag atttttccaa gactgttgga _dy LdL.dl.- acaggttacc taactgtggc gacagagtct ccctctgcct caggtacatg atgttggcca agtgctggaa taatggaaaa gaatgctgat gcctacctgg tgagcgtcct ggcttgtcaa tctttgaagt catatttcaa catgtgtatg gtgcttaagt ataaagttta cccatgttct tgatatgcca acaagttgtg aaatacttaa ccttgggagg agactgtacc aaaaaggaaa gaaggaagga gaaagaagaa aaggaaggaa tggtaacatc attcaggatg tatgtggatc taagagtaca tactttatga taagaagaaa tacagtcaat gcataacaga caccaaaaat gcaagtttgt taggtttcat ccagacagaa ttctatgaat aaaacaaaca acagttcaga caaaatatgt aagcagagat tacatagcag agcaaaatga tttatacaaa agaatgaaat tcaaaaatac ctgaagaatc caagggccac atttagtggg taggtaagaa aaagcgtagg tgctctgtcc cccaggttca ccaccaggcc ggttggtctc ttacaggcat taaacttcca tcacaggctg aaattgccga gggaactgtt cacttctttt attctctgtt ctatgattct gtttaaggta gtaaaaaaca taaatatttt ggaatagcaa cgtggcaagc ttgaagatta gtttattttt ctgaggcaga actgcactcc ggaagaaaga aggaaggaag aaagaaaaag ggaggatcgg ctcccaagta cacctttctt atccagccac cacccctcgc taactactgt gtgctgagat ctcactactg tggtatatct aaaaactaat agctgaattg tgtagaggct ccagagtcta atactgaatg aacaaacaaa aggcagtcat taatttattt agcctccctt gaattttaaa taaaaattta taagcaatgt gtcattttta ttaatataaa agagaggctg tggtgaatgt tagaggtcag Lyt_d LLLd Ly gtaatggatg gttttgtttt ccgaggctgg agcaattctc tggataattt aagctcctga gagccactgc agaagctctt aggggtatct gcgcaggggg ctgctgtagt ccatttgtga cagaacttgg ttattactta acaaagtgag aacaaagaaa taaggcattt ataaaaaatg ctcaggctgg ttattacagg ttgaggctga aaattggttg agcctgggtg aagagagaga gaaggaagga aaggaaggaa agggagaaaa atttagttct gctgttgctg cgtgacaaat tgctgctttt tggggtacaa ggtgtgggga acctttggct gagtgaagag aattgcaata aagaggtggc actgagctgt ccaacgtgga tctcatcttg caaacaaaca ttaatttcgt tcgattttaa cccccaaatt aagaaataag tgtaatggta cttaggatct agtcttgtat ttatatgcta tcaagttaga ctacaagtat gggtgtggct - 134 - 008252 acacatcttg caatgcatag gacagacctt cccattaaag catcatccac ctgcagatgt 37380 cactagtacc aaacttacat tttgtgtagc ttcagcaagt tctctcttaa actattcatc 37440 gctctctttt ttcattagtt ctgcagttta ataagaatat tgccctatgg agcagttcta 37500 gcactttggg aggccgaggt gagtggatca cctgaggtca ggagttcgag gccagcctgg 37560 ccaacatgac gaaaccctgt ctctactaaa aatacaaaac ttagccaggc gtggtggcat 37620 gtgcctgtag tcccagctac ttggggaggc tgaggcagga gaatcacttg aatctggagg 37680 cagaggttgc agtgcgttga gatcaagcca ctgcactcca gcctgggtga cagagcaaga 37740 caccgtctcn naaaaaaaaa aaatagctgg agattaggca agttgtatat atgtaaacat 37800 aaaaagacag aaacaaataa aaaccaatat agcacaaccc accccatctt gaacaagttg 37860 tacttctaat catgcacatg tgaaaacatc aagctcttat ttgaccacca ttcaggattg 37920 aatatttagt cattgcaaat gtagttgagc aaataagtat ctatttggag tttaggatta 37980 ggatcataca cttccacagt taaatctact gactgacagt agacagttca ttgagacaca 38040 gatacagcat agtcatccat gtatagtggt tgagtaacgt gtaagtttct tttcccttcc 38100 aactgaaggg gatgctcaaa ctgagattta accaggacaa agtcaatcag cttctctctg 38160 aaaagggtga tgtctgatca gagtcagctg gaggatcaga aagagtcatt tagaaaagtc 38220 tgagggaaac tgatgatctt gagtaacaag aaagttgcca ttttgttcac cacagatgaa 38280 agtctattcc cagacagtaa atgaagccgg ataatcaggc caatttcata aacagtagat 3834 0 cagattggaa gagttaaagg accagataag aaaacaagca cctgtagcag gatgacccga 38400 gtaacacaaa ggatgattca cgcaaagggg gtgcaaggac taattctgca gactcctata 384 60 gcttgttgtg ccttgcaagc ccaagagagg ttcccaaagg gactgaagcc tggcagcatc 38520 cagtgaatgg gctctagcat acctggtaag gagctgtctc agaagatggg gtgttccata 38580 tccctggagg ggttaaagtt caagaacatt gctgaggatc caaacactgg gagagagaaa 38640 gaaatggatg gtcataaaat aaatcctctt tgttctttta aaaattctga gtgtgggagt 38700 gaaggtgatt tttcagacca agcccttggg tactgactca ttcttattcc actgtctcgg 38760 gcttacatct ccaattctaa cttcccaact tctgtcatgg cagatcaggt attaccattg 38820 ttagaatagt actataggta ttttgtgttt ggagagaggt ttagtggtag gaccccaggt 38880 ttgtagtgtg ttgctagtta tcatcacaat tttatgaccc tctcacttga gagtggaatg 38940 acaatgtggt tatgtaacct aagactagaa gaaacaaatg ggagtactcc caaaacacaa 39000 atatctcacc tcttttttca tgggaaaaga gtcagcagaa gagggcaaag cggtaaataa 39060 aatcctaaag gtaatcaaaa cagatgggaa tataattttg ttcagagcta taaaaaatat 39120 cctctactat tctcaataca ggcagtgtgt agaaagtttc aaggttagta tttatcttct 39180 aaacatagac ttgtggtctt cttatttaat caattcaaac caccagtgga agaggtcaga 3924 0 aagaatgaat gaaatcagga gaactgatta tcatgagtat cattgttgtg attgcatttc 39300 ttgcacagct ttaatttgtg ccttgagagt ttggggtctg agaaagtagt gctattccaa 39360 gagctctcca gggatatttt tggaacaggc cttattgttc tctaaaaaat taatgcgtta 39420 tattcccagg tgctgttgta gcatcatttt tcatttaaca aaaatatggt cgtttagcca 39480 gaaattttag ttctataatt aaaatggact aaaaatattc tgtagcacat ctagaaaaaa 39540 tattttaaat gtatcaatgt tcttcctaga aattttccat gttggatcaa acagtagatg 39600 aagacttttc tgtgtgtata gcatgggttt ttatgtttat atatgagtgt gcaggtgttt 39660 taagagaatt ttgaaaatca gcaaagcaaa gtcaattcac tacaaaatag tttccaaatt 39720 atctgtgcag tattaatatt gaggaagaat taatcaaaaa gcaaatattt gagaaatgct 39780 ttcctatgtc tgtaggctac aaaatcattg tttaatttat caacaatctt tgagtgctcc 39840 tcagtgtctg gtacagaaat gaaagataca gttttaactt tagagtgctc acacctactg 39900 agggaaacaa acaaggtaga gaattcagta tgacgagtga catgagacag ttgaatggga 39960 ggctctctgg gagtactaag ggccattgcc tagtgtagct gagggcacag aggagctgag 40020 ctgggcagcc caggagtctg tcaagagggc cagagggaag cagtgcaggg ccagcagcga 40080 gagcgggact gtgcaaaaga agtttcagta gctacagttt ctttctcagg gtcctccact 40140 tcccaaagaa aatgaaagga acatacattt gatttttgtc agtttaaata ttggtcgacc 40200 attggaaaac catttttgaa aagtgatttc ttttgctatg tttggcaaag ccaggagtag 40260 taaagagatg aggaaggacc atcttcgtcc attgcctttg cctttgtaac tgttacttct 40320 tcacagcctt tctacccagt gtagttcacg tatgggaaac atttctttcc atcgtcagcg 40380 tcttcctggg gagtgttttg gttatacttt cacgaatgtc tcacctcttt gtgtcaacaa 404 4 0 acccttctct ttgagctgta ctgtgggttt ctgtttcttt cctttttntc ctttgtccaa 40500 ttagaaaaaa aaatggagta gaagtttcaa ggctactagt tcaatgtgct cttttttact 40560 tgactccaaa tgacatcttg cttgcagatg tgattaagaa agatctggac agggagaggg 40620 aggataattt tcaatgttcc tctcaaattt aatcttatag tgacaagctc catattctac 40680 atgaaaatgc tcagttaata cagctattaa acttgaatgg tagataatag tggatttctg 40740 tagccactgt ttggtttcat tcaaaaacct agaggaaaat aagggatata cattataaat 40800 tgtttgttta atgttaaaaa tatgtttcat attcagtgca cataaagcaa aggtattgtc 40860 cagtgttagt acctatgaat catcaataaa aacagttgct tctgtttgct gttgacattt 40920 tatgtatgtg ttgttcctct ctgaaccttt ttcttaaaag tcaactttta agaaacttat 40980 aagtttactg tggagagcct atgaacagat gcatgagggg gcgcctgttc atatggataa 41040 gatagggcta taaatgccct tatcttgcca gggctcttct aggcctcttt agggttaagg 41100 - 135 - 008252 catactccct tggattgttt ataggttatg aatattgtaa attacttcat aaactacccc gcatcacagg aaaaaaaaaa tttaaggata tacatattta tcaagatatt ttcaaggcct ccctcgtctg attaataatt aaaaaaattt ttggagacct tatctgccct atgttgtgtt gagttacagt gttatattct ctttgcttct gcggctggac ttctcatgca tgtttggata atgagtggag attgaggtac cttcaatggt tttatagtca taagtcatta gaaactaact gctttgtacc agacaagtca ttgttaggat aaaaaatcta tgaggtaatt gtgttcggta gatactttac tcattttggt tggcaagctt tgtttatttc tattttattg ttttaagaga tcattgagaa tcaaaagtca ccgtctatga gctttccccg ttgtgaaggt atcttttgtt tttgtgattt gagtacagtt aggtacgcta accatcagtg tctgagaatt gtaaccagct gaaagactgt attcttatct ccccatgtga caccctcact accagaaggc agaaacaata ataagagaaa tggggtacct taggatatcc ctctgctagc ccatggaaca tctttttaaa tttttgacct tcattaacca catccccact aaagaggcta accaatgtac atagaaagta ttctcttagc ttgccttctg caggctcaaa cttggtcata attggtttta aaatgtttac atatgtgtgt taaatgatac aatccttaca acagaaagac agaatgactg ctttatcttt taaacacatc tccacatatt ctcccagaaa acaccatgga tctccctgtg taaaactata aaatggtaca cttttccaca ttagatgttg acagtaaaat aataagtatg gctgtgactc ggccgcctca ttggcgttcc actgcgaaga tggttttggt ttttcagaac tcaacaaatg tcagcaagag gaccttgggg tgtgttgtaa tttgctgcaa tttctgtttt ctgtatactg ccatctcact aaacttaaca agtgatcccc aacttagcca ataatctctt gtgatatttt ataacctcaa tattttgaaa ttacaactta gccccagtct gatgatatgg gtgaccttca tcgccatttg gaaactcaac aaaatatgat tattagcaca ttttctttcc ttctgcatgc gaaacagtga tgcacattct tattctgtaa cagttttatg gtgttttctc ctgaaatatc acacagtaaa tattaaagag agttaattcc atatgcttta ttttccctac tttagctttc atacaaagtc atccaaagac agaaagtgat gcaggtgctt tatccttttt ttattatttg aaattaccct gtgttgcatg gataaatatt agtctgtcct caggaaggct tacttctatt attgtactaa tgccctctgc gtgtttattg tggggatcga gatgttataa aagctcttgc ccggttgtct ctgttactgc aaggctctgt tacttctgca tcataatcaa ataaatgctg ctggacccag ttataatccc tttaacatac gttttatgga gcatttatca tacacaatac ttccttctat caatcaagag gttcaatttc cttttctttt cctagctaat atgctttaca ttttaaaatg cattaatgca agcctctttt cttggtgttt ggctgcccca agtttaaatt taataaaact aataacaact acaaaagcat ttaagaacct gtctgtgtta gcaagatatc agttccctta gtttccctgt atggcatttt ctttctttcc ctatttggtg aagctccatc ttcaaatgaa ttcagcttat ttaattatac catttaacag ggcttgaagt tgattctaat gattgttttt cctctgtcat gggtgttcct tcatcactct gaaagtctat tttgtcattc ctgttatcat gaagcagcac cccattcgag atgatgggct agcttcacgt tgattaatac tagaaattgc tacagatatt atgactctaa gtatatccag ctttcactaa tcagaaaaac ttattgatac taaaatgtgt tttctatgtg attgttgtta ctaactctat aaatataaat tggttctacc ctgtaagatg aagacagaga aataccagct aaaggcagga taataggcga tctcccttgt gctgttgctc acctgtgcaa tctgtgtaac tatttataga tatatttatt attgagcaat accctgtgat tttcccattt atcacagttg attattatct ctgtaaaaca aggttctctt attactagaa attcactgcc atatcaattg actcagttct tgaatgtgtt atatatatac ctaaaggtat cagaaacaac caatttttga ctggaaaatg gacaggaact catgtacaga cattttcttc gttctgtgat ttagactttg tgaaacattt tacctcaaca catgcacatg tctatagtct cctgtttcta cttgctaatc tgatgcacac atgttaaaga atccctcact tacgttggcg aaaaaaaaaa attaactata ataataattg aattatcaaa ttgggaacat actataatca gtttgtaccc agcatggtaa actttccacc gtattgataa tttccttagc gtatgacaag catttatgaa taggactgca catcaagtct tgattctgct ctctcctctt taaaacatgt agtagaattt ttataaacaa tattttcaat ttacaggaat tacagatgga gatgtgtggg ttgcatcctc tcttatagtg aaccaatgtt gtactttcta ttacattctg tggagcctga cccatttatt aggctccaag ttggtttgac aatatcaaat tttattaaaa agcacttttg ctttcttatt agtatattca gcaattgaca ctcgcctggc agttctgcag gtaaatgtgg gcagaaatca gccaccaccc tctcaccctc gactcaggaa nr-t- n't- t-1- a a a aat-a, ttgggaggct gtagcaagac cttgtagtcc aaggtacagt tatccccaaa caatcaaatc tgggtggttt gacatcacaa tttatgggta ttttgtaatg gagggagagg cctgtctcta cagctactca gagctgcaat aaacataaaa agagcttcca tgatacaaaa atcctcaaat tttattcatg gaaaatatat gattgcttga caaaaaaatt gtaggctgag cacaacactg gaaaataaaa ggcagtgagg ccaagatcgt ggcttgtgga tacttgcaag ttcattcaga gcccaggagt ttaaaaatta atgggaggat caccccagcc taaaacaggt taatgacacc caaccacatt taaaatgcaa cctatgtaat acaaaccaca ttgagatcag gccaagagta cacttgagct tgggtgacag tcaaaggccc tatattttgt tattacatac ccactatgga gtctatgtgt gtacaataaa cctaggtaat gtggcatgag caggaggtca agcaagatcc aaacacagaa aacagctccg ctttgattct atttcttact aggattagaa ctatgaaaaa - 136 - 008252 gatccctcaa gattcccata gaaggtgggc agatccaatg aaaaaaagga gaaaatataa ttaaattcta gtatctatca ttaataatta tgaggcatct - 4- -"+. -, _ at.akjLijLk'aij tgaatatgct catagcaagt tcccggtcca acctaaatgg atgacctaag cacagctgga agggaaatgt cttaatcaag taatcataca gttgctaagc tagcaacgag gcttttatgt tatgaccaaa gttactaatt catggaagta gctattctca tgcagacttc atatatatat taggcagctc ctgtgatgag accaaatgcc tttaggagac tcaaattact tcttagacgt attcatacac acaacttttt aagtcctgag cttcagatac gagcttatcc caaggctact atatctaact aattaaatat ttgagcactc tgataaaggt tccatcaaca tcagtgctgg aagaatttct gcaaaaatgt aatatgtgag aataatgagt accataaatg aatcaagcca aaaaaccata tttctctgca ttcagctgct acctgttatt gcatttttag gacaggcctg agaggagagt tcttcttaca tcttatttaa tgtaaaaaat ccccaaatgc agttccatag tacagatttg cctttgtttt agggcagatt attccagaat aaggatattt aggagccgag tttgttaaaa ttatgatgct tCutattCoCj atccaggtta tctcatctcc ttccgaggtg aatatggaac attcctttaa aaaaatagtg gaagagactt aggaataaat gagaattgtg aaccaaagct cttcaggccc gtagagaagg ttccgtgaac tgaaggaaat cattcttgcc tcaaaggact ccataacagc atatatatat actaggaact aactgtttct tccaagacac cataatataa aaatattgac gggtaactat ccatgatata agtgatgtcc taccgtatct ctgttatgag caactttgcc tttctgttaa tttataaatg tctgattgca ttgctgacat agaactggtg ttcttagaat gttaaaaagt cctccaaaaa gtttactgtg aaaacaattc gccttctaat agaattaaat ggtatccact aattttcaaa ttactgtgta agtgattctc catgttgttg ctactttaca agagaattct gccatggaaa gtcactccca ttttcataat gacaatgatt atgtttgtga caatattgtt ctaaaagcag tatttttggt tgtgtatctt caggaataaa aacaactttg gtgtacaaat tgcaattaaa gtaaaagtct LULLuauaaa tatcagaaat tctctcctct gcagcctcta tatgatctca aaccatagta tttggaagca ggaacaaagt tggtaattca aaaaaccttc cctgctgtca cgcccaccct gagagacata agggacttct aaaataaaga tttctctagc aaagggtcct catgtgtata atgcacattt ggctctgaca ttaaaataac agcataggaa catgtaactg tctggcctct tgccttgtgc ataccctgct catgactgga cactaaccca ccccatggtc ttgagtagaa aatggtagga aatcataatc caaagacagc gtgttttctg tgggaagaat atgggttata agagaaaaaa gaaagctaca ttccaagacc aattaatgca aagaaaccat tttgtgaggt ttcactattt gaatccaaag tgtggatgtg cattaccaag atgatgatga aaccaaccga caaagttgaa ttcatttgtg tatattgaga aatactgtga ggaaagttaa ttttttaaag tccataaaac cagggcagtt tttgtttcac cttctagcgt atgatataat taaacactta tttacttttg cacactaatt gaattgtcca caac i"g t.ca gccatgtaga ctttttgcaa gaaggacatg gcccttaact tgtttatgga ccaccaagtc tgtacagaat gaggtaatgc attaagaaaa gcctctgggt ggctagccac atttgccatg cattcagtgg tgattattat atatggaaaa gggcttgttg gaaaatatgt tcagaacaaa gggaccattt gaagcaatat caaattacag gaaaacagaa gagagccttg tcagttgtgg ttgttaatcc tgttgggatg gacctttact ctctgatcta ctcaaagaac gacagaaagc caataggatc aatcatttct aattaatatc gggcactttt caatgcctaa tggatgaaaa tataacttaa atttatttcc tttcagggaa aataatagaa tgcatcattg tctgtcatag agcatgcaaa tgtgtgcctg attctttctc tattatgcct ccagttataa atctccatgc ttattggtga tctgcactag agtgggcact ttatctaacc aatccctata cataaggctc attacaaaag gtgtgtgttt gaatctttat gccttgtaat caaatactgt aaaaccctca tgggaaaagc gtcaattcat аасиисса_.д aaagccttca actatgagag ttttcagtta cacatgcagt gtaactctca aggcaatggt cattttgatg gtctagatat atgctatagc gaaacaccac ttgcgagtgg atgctattat tttaaaacac aatgcacaca tttgcagttg gacttgttga gnnnnnnnnn gaaggggctt caagggaggc ttttcaattg agacattaat ggaaaatgat tgatttcata attgggttga actatagggt tgcaggtgag cccattattt gactagctca tgctaaattt aggcttctga atattgaagg gggtctgtta ccctaattaa gagtttttct agcaacttgt gacacatgat tgctcaaaga tcatgaatat gggtgcttct tagtacaatc aatatattat tgactattag ttgcaaagta tcttgagttt ttaccactat ataatggaac aactgtagca gcttccaaat gattctacta tctaatggct attaatgtct caatgttata tattcagaag ctgagaattt agggcaattc ttgtggtgtt aaaagagaga tattataata taagacaaag agttgagctt ctggtggtgt tatagatgaa aaayvjLL.UL agaaatcaac ataagaaagt tgactttgag gttcctggag cgtataaaat tctaatagaa caaaattgta tcagcagtgg tgaggacctg tcaggcagcc gccctctgat gtttgtaaaa tagatcacaa gcttttcaaa ttaaatatag tgtctaagtc nnnnnnntat tctgttatat gaaaggaaat aatggtcacc gccaagggtc gtagaaaatt cctgcctgat aggataagtg tacccaggcc ctgaaagacg agatgcttta tgatggagat tagagccaca cagtcaatac tttggtaaat ttatctctga cagagaagac cctttgggtt gcataaatgt ccaaaatgga aaatacttta acatcattaa agcatggtgt atcataggaa ttcgtaatga ctattttttg aattttagat ggaattctaa ccattagata aatcccagaa gttgcaaatg gattgtgcta attcagaaca ttacaaactg ccactttgca taactggcta - 137 - 008252 gtggtaaaac gagatgctac ttctggcctc gggacattcg tttggcaata tagaaatgta tatgaggcat atcgattata attgatgtga gggatgcagg aactgtaaaa ctcatncctc aaaattacat atactagtta tgacgtatgc atggtatggt cagcccccat catccatatc catcctatag ctggcctttt aagagataca tgaggattct taaccaatct tcctttactt ttgattcttt ctgatcaaag gtatcaacag ccttaaaatg catctccact ccccaaaaag tgagatgagc taattttgtc gagttcaaaa aaaaattagt aggaaaattg ctccagcctg atacatatag aggcaagttt attacaaatc accccttcga atttatttct tatcattatt cacacacact gtgggaaatg aggagataat ttagtgggcc ttgttccctt aaaataatta agttctttcc taggtcagtg tggaagactg cacaatgctt taatggggat gcaaagcata ttatcattgt gacttcattc aagctggaga tttgtcttga gtcaacaagc aaaaatcgcc aagctctaga tctattccca ctgatggctg tgggactcaa cacctttcaa atgttaactt catacattgt tgtatccaaa tcctaagaaa tatggcaatt aattatgaaa agatatgctg tgtgtgtgtg tgttagtggt attttctaag cagtnnnntn agttacattc gatgaggaat tgtctgtgtt taccacagtc ccagagagca tcctgtatgt ttcttggtag aaggaaagac tggcttctgc aaatgttgta atttcataca tgattttggt tcaatatatg agggttttta aggcttccag gatgtctata agtttttgca aatttgcagt atttaaaaat ccaccctgga tgggcattgt cttgaacccg ggcgacagag ggactgtgat attttccaca ctaggtaatg atacagggtc gtaaaacagc tagaaaaaca cgctcctaag tcttccagct aggtacctag tgtagcatct cataatacgt cacaaagtgt ctatgccaca ctcctgttca tccactaaat ttaaattctg aaaatactaa tgcaagacag tcttttttca caaatataaa catagagttc agatgtttca cccagggacg cagcaaaggt tttgtccaaa ggacctagct caggcatgta aaccaggttg tttagccatt ttcttttctt tgccaagagt agtaaaacat aattgggggc ttttaaaaat ttggaataaa atgacatatt tgtgtgtgtg ggtgtctctg cctttaatag nnaaaaanna agttgcaagg cccattctct gccacggatc accttgtaga ggaagaagaa agttacatgg cctagtggcc tggcagtccc tagctagccc taattcaggg aataagtgac ctattgaaaa ccccagggat ttaataactg aattttgaca ttctcagaat gtttattcta tggcatcttt acatatagga caacaagctg ggcgggtgcc ggaggtggag caggactcta ttttggcaac aaggatctac• ggatctttgc agactaacag gacctcaggy aaaatgttta aactagaaag gggtgtaata gagataataa tgaatttcat acataaaatt attaagcaat cgcacacttc gccttgaggg aaaaataaaa attttcattt agtatttctc aaagtgcctg ttcatttaag tccataagaa tcaatgttcc tgaagatgct cgccccagcc ctttgaattt tttaaaatcc tttttttttt agagcatttc ctcagctgca acaagccctg aaaaaagaca atgaattaaa acatactatc agtagccaag ggaaacaatc tgcaaaccat agtacaaaaa tgtgtgtgtg gggaatgggt taaaatnatt aattaatgga gacaaaaaag ctcatataaa cagacacctt ctagcatggc ggaaatacag tcaagctcag ccatacagat tgccacgctc agcttctgtc gattaaagag cttgggagaa aaaaatttaa cctttatgcc gtgactggca gaaactttac gagctattca aagaaaatca gtcaaaggag ggcagaagct aaaccctctg tgtagtccca gttggagtga tctcccccca tgtctctcac caccactact atcatctcat gaggacataa aggaatgaaa attttgccct aaaaatatgt ttgacacata cctaggtatc cacttgggtc tctgctgagc gtggcaatct ttattttatg taagatacca atttgcaacc tctcccagat ttgtggagaa acatagcaag ctttcaatgt gacaaatttt atattaaatg gctggaggtt tgcctccagg tatttcatcc tgaagttaga taacatacac agtggtattg agagcctttt ctgtcaacat gtatttattg acatcttttc tgaccaagaa ggcatatgtt ttagtctcta taaaataaga aagatcctat tgtataatac ttaagctgnt taattataga cattctaaca cccaaaataa gttggaggtg ccttcctgtt tgctgtagct gaaaagaggg cttcaaaaaa tctctttgta cccgcaagga tcaaagatct aaattgaacg cacttttact gagagaaaat acaattagag atgctggttt aaggtcctat ggaagaatat gaatcctgta ctttatgtaa gggggattgc tctagtaaaa gctactcggg gtggagatca aaaataaata cacctcactc accagggatt tcgtatttcc ataagggtga cagtgagcac attcctacca tagtgggaaa ggtaatatgt taattcaatg tttaatccaa aggttaattg tctaccatta gatcctgaga tggtagtcca tatagtaaaa ctaatagata gaatatttga tagtcaataa taaagaacta tgataacata tctctgactc ggcagctggt tgcagtacaa aagaaaatcc gatggtattt aatagggatt aatcaatgaa tattcagcat gctgtctttg ctggctgaaa cagaaaaccg agtccgcttt tgaggatgtt gcaataagtt attgaacttt gtttgtctgt ctggaatgtt tcttttgcaa atgagaaagg gaaaccacgt gttgcttagg ctcattcagc ccactttgca ctaaccctca tttctgcttg agggcaagaa aaacagggaa tttgagtctc actgtaatat tttttgcatt gaaaattaaa tagtctgcta agtataggat tgtcttttat aacaggtttg gcttgattca aatggacagc atgtgcacct ctgagctcag aaaaaaatac aggctgagac cgcccctaca aataataaaa attttataga tgaaactcta atcagcagtt caaaggaaac ttgtagttag tacacacata atacgtgaaa gatgatacct gatttcagat aagaaactac ctgcctagtt aaataacacc atatttttat agccagggag acacaccaga tttatcaata gcaattatat actcaagtta taattgaaat aaaatataca atctcccttc agctgtggat tgacattttt acagctcttt cactccttcc tgataagttt gaattttgtt - 138 - 008252 gacatgtgaa aaattcattg ctgtttttct agtgcactga ttctcacagt catcttctcc taaactcctt ctccttgccc acccctaact accggagaat gtgacaaatc atttggagaa gtctttctcc tgtactctga acttcttttt aggtatacaa ctaattaaca gggtaacact ccattaatga ctaatatctc tgcttcaatg ctttctgtgt tcaatggcag acattttttt tgtgaataac ttcctttggg ttttttggga acagtgtaaa tgatggtgac ccctgataat ctttggaaaa atcttataaa tttcatacct gcatttggaa tagcaaacat gattagaaga aagctgccat tatctcttct cctaaatatc ttccataata tgattcttcc ctcttctcgc gtgttcacat agatagggac gccattactg gtttctattt tgtgatgatt cacccatcac taagatctaa atcactttgt cccatttcta aatttgactt ctagttcatt gatttttttc tatccattca gctgcagtaa catatatcca aacctccata agggttatct cattgtaaca tagggatcgt aaatctactc aatattcttt ccccgagtaa gctttcagag aaggattgca tggagtctca tgacaacaat tcagggggcc agtaactcat ttttcacttt acctggagaa aaggggtggt ctgccgtggg ttcttatgca tgggctatgt gtattgaggt tgatatatgt cgcatatatt tctcctagca ttattagatc ccccctgtcc tttgagattc tcacttagca tttattatgg tccatcagta acacggggct gaagtgggat ctgcgtttca tttctttatg gatatgaggt atgcactttt agaatttgct accgaaggac ctgggaaaaa gaacataatg ggtggggcct cgtccctgcc atggataaca atactgacat tcaaaataat cattttccct ttttattttc ttccatagtt atgagactgt actcaaggat ttatatggcc gtgattaaca acacagtgta cagtagttac aatctcatgt ctgagaactt ccagcccctg catatataaa taacgtcctc ctgagtaata gacacttagg gcagatgtct tgtgggatca gaatggctct cctttgccaa gatatctcaa tcatatacct catttataat tgagttatgt tatgttaata tagaggtgtt gagagtcctc tgtgaacttt ataataacaa cttctcttcc gtcaccttac ccaagcagcc tttagccttt gtttatatcc atctcttttc gggtacatga attttaccat aataaaattg aaacgattgt ctctgtgtgt atacaataca ggccatcttt gtaaccacaa tgagattatg caggcttatc tttcattgta ttgttccctg ctttgagata tatggtagtt accaatttaa atagttatct tgtggtttaa attggccatt cagattattt ggcagtgggc catagtctct tctttagcaa atgagataga ctggaaaaac taaggcccaa ttggtttccc caagagcagc cactcgtagg ttggttttca tgccctcata tttcttttgg aaaataaaat agagttattt taaatactta tgcaaccaag gtggggtggg gtattattaa tgactgataa tcctactctc cggtgtttgt catatagttg tgtctatacc tcttgaccat ctgatttcat ctatttttaa atccccacca tttgtctttt atttgtattt catatgtctt gcattttgtg cttttgaata ctgccttttc gcaataagta atcctatatt tactatcact taaaaagcac actatgtgct agcttacagt agtgatgtgg cgctttgaga tccagcattt ctggccaaca gtgcacgcct gaggtggagg gagactctgt ggatgaggtt taggagaaag acccaggact aatatgcaag ccaaatcaga gcctcaaggt gaataacaag aaaataaaga tctcagagac atgaatgtga acaagtttca caatgaagag aggagctgtg ttaaaagaaa ggctttcact cttccactcc ttatctcctt attttgttta gaaaagctta tgatgttgac actaatagta ttttattgac agacactaag atgcttgaga gttttgaaag ggtcagggaa tgggaggctg tggtgaaacc gtaatcccag ttgcagtgag ctcaaaaaaa tactacaacc tgctgattca aatggctggg taatcataag gtcgtatggt ggccatctga ggattaatga ttggaatggt ccctatatga ctcatatgct ttccactcaa tgcatggaag attagagaga agaattaagt aatgattaat ctggcttgga ttcagaaata ttgttagtta caatgaatat tatgctgcta ataacaataa attatttttt atagtcttga gtgggaggga aagtaatgca ggttttagtg aggcaggcag ccatctctac ctacttggga ctgagattgt aaaaaaaaga ctgtgatgta gagaacatag tcttatggct ttcgtgatag atccagagct aattaggagg cagggtgaag gtggggatag gatatgtgtt tagaaactgg cttcatgtgt aaaagtggga gaatacattt aaaaatatat gccatctaaa ctaccatggt tggtgttcca tttatttctg tgataagtat atataaaact tgacaaagat aatccatcaa aagaaacatg aaaaggcagc agatgcaatg cataatctta atcacctgag taaaaatcca ggctaaggca gccactgcaa gagagagaga gaaagaccta ggatgtgaat cctagcccca agaaataaac ttaaaatcta caagtcagcc tgtgatgaca atgttgggct atgaatatag ggggaaaggg ttggccctga tggccaagaa ctatgtctca tgaagataaa actgggtaac atcttaccct attttcccct tcagtttaaa tattttgtgg actacaacta tatagtacta ctgatattta cacagtctcc attgagctag agaatatata aaatcaaaac gtcatgagtt aatattaccc caagaatcac tgtagcctgg atgaggaaaa catgcctggt tacattcaac tgctttatcc tatcagggct ttaaatcctt tccttatcac aaatcgagca tgaagtgtat cagcatgctt gtcaggtaat ataaagcttt gttatctgtt gcaagaaaat atggtcctga aggaaaactt caaggctgaa gttccataga aaaatggagg tcctccttcc cctttactcc gcttcttacc ctgagtacca acactcatag caatcacaag aattggggtc cttgagtaat tgagaccagc aggtgttgtg ttgaacccgg gcaacagagt gaatgactaa cagccattta cagagggaga agctgaaaaa actctgagtg gttaatagat aaagatttga aggagatagt agatgtggca atgacatctt tcagtgtgac ccagaggcag tgtaccgggc aattaattaa aatacttgat tcatctcagt caggagcata - 139 - 008252 taccacttct tttatttatt cgggatctcg ctcccaagta ttagtagaga tccgcccgcc ctttttattt taaaattaaa ttccaatgag gcattttatc atgtttcctc cactttctta acacttttct ttgatgtctg gtcacttctg acagtccagt ctcgcttaga tagactgtgg ggagtctaaa taggactttg gagccttggc catcttgtaa catcattgaa gtctcactct gcctcccagg tgcgccacca gtcaggctgg gggattacag aaaacagagt tgaaataaat gagttcccat atttgaccca cttgtaaaag cagagctgag tcccaactct tcatacattt tggaaataaa gcatggatga aaaacaatga gtaaccctaa gggtgactca caattagtcc acattttaaa taaatgaaat accatatttt aacaagattg aaccaatgct tctctctcta catatataag gctggcataa tagcaactaa tatatcaaat gatagtcaat tctattgagc aatttatttt gaataggaac ctttctgttt agtttggagc gctctttaac aggacatttc agtttaataa ttaaaacgag gcctgcaatg ttttatttat tattttttga gctcactgca gctgggacta cggggtttca tcggcctccc ttaaaacaag aggaagagtt taacttggag tcacatgaaa ttcctacacc tcctcaaaaa tttatgtctg cttctccaac caactgtgct gaaagaagga agctgctctg caataacacc gcaagcagta gatatctctg aggacataac gcatataaga aagaacccaa cctgcccagg ttcaagtgat cgcctggctg tcttgaactc gtgtgagcca ttattggaat tagaaaaaat ttaactactg aagtttaaaa agaatgatct tggtggctgc ccatattgtc atttttctaa taaacaagta atgaataaag gaaatgaacc gcctagtttt cggaataaat acacatttca atgttaaaat gtactaaaaa ctactctgta attaaggtgg tctgtgatgc tgtgtgtgta tgcatatgtg aactattagg ttttaaattg atttccatta gaattagggc catttgtgga gtgttctgaa catatgcagg ttataataaa aggaggacta gccacttgtt tcatgtttat gatactgagc taagcgaatg tctttctggc ttatttattt gacggagtct agctccgcct caggcgcccg ccgttttagc aaagtgctgg caaaatgtga tacttgaaat gttaatgttg aagcatttct acttccttct ataagccagg tcataatttt caggggaaat ttagacatga ggcagtacct tttgtcctcc cttttccata cacttggagt gatagttttg cctgacttct taaatttgtc actcattctt ctggaatcca tctcctgcct atttttgtat ctgacctcat ctgtgcccag gaaacatttg gaggtatcaa ggaactgttg accagagtat ggcaataaag cacatgagca tccacagcac gagtagagaa ataaattcat aataaataca tagtatttta ctcattatgt gaaaataatg gtgattccaa cataactcag atcaaaactg tcctcatcca cttaattttc tgagtttttt tatatacaca tgtatatgta attttatata ttttcctaca ttatggtatt acataaaaga ggagaatata acacaaatta catgcaagga tgaaaatgca gcagaaaggt aaaatttttt ccagttatta atggtaaatg tcctagaacc tctttctcat atttatttat cgctctgtcg cccgggttca ccactacgcc cgggatggtc gattacaggc acatgttctc cttctttcaa agtcgattct gcctccagga cactcatcct tacctctgtt aataaaataa tccattactt gagatgatca tcttggctta tggtcttgtc cagaacactt ttgcttccag ccaggatgtc gttatgtttc ttatgtttag tttttttttt gtggcacaat cagcctccag ttttagtaga gatccgccca ctgaacccaa aacggctgta taatatctga attttgttca tttacataaa agctctcatt gaggcagaga tggaggtgag agatttctct aaatatataa aggcatttta agaaaaaagt tacttgctag taactttaga aatagtattt cataattgga tctccattgg gttactataa ctcaccatta acgagtaata aacatatata tatgtgtata tttcaatggg ttcccacatg tgtttcctct tcttcttact ggcatctttt caattacaat aacgattcaa catacttgta attgttagct gttaatcaac atgaaattta tgtgcaattc cctgaaaaac ctctgtgcat ttatttattt cccaggctgg cgccattctc cggctaattt tcgatctcct gtgagccacc ataaaaaatt tgtatccttc tctcagactc agcactgaca tcagaactca acatgttttc attaaggata tagagaccat attatgttta tgaaagtact tgtaagaact atttctatat aaagtgcctc tcctctaata acctctttca aaatatagtg tttttttttt cttggctcat agtagctggg aaccgggttt cctcggcctc actaattgta ctttgtaatt gagacagcaa aagtgggact aacccacacg tccacctggc ggctttattg tgttttacct ttgtccgtga atgtataaag aaagcaccct tattccgatg gtctcaggag tcacaaagat tcatatattt agctggctca tattatgtat gtaattaggt ttaaattaaa atttattctc tacacacgtt tacatatata cagtgattct tgcacataca tttcaaatat cttctacaga gtctgcagtt catagctata tctggtgccc aatttaactg tggtaagtta ttaattatat taccatgctt ctaataaatt cccctcccat tgctgatgtg atttnnnnnn agtgcagtgg ctgcctcagc ttttgtattt gacctcgtga gcgcccggcc aatatggaaa ctcattcttc ctgcatttat ttttcacaga gctcaaacat atatcatcct taattagttg accagtcatg ttaaccgaga tatttgacct agaaattgaa gcatgtcgtt gcagaggttg tgtggtggca gaactctttt ttaggaatag ttgagacaga tgcaacctcc actacaggca caccatgttg ccaaagtgct tgttaatgag ttgaaactgt actgatgttt ttgtttttaa tcctacccat tagagttggc tgcctcagtt acccaagaat ctctatcgtg atacacaaat gtgattacag tgttaaatat gcacttggtt tatataataa tatgttttta agttctctta tatacttttc aatttaataa tacatgaagt tctttctccc tttatatata tagaaacatt gatcacacat tattttacct tgcttatgga aaaaataggc tgaaaccaca attataatta ttgaagtgat caagaccaca tatgtggtga taatttgtct aataaggctg tatcctcacg cctctgtcat cttgtcactt - 140 - 008252 aggggcccct cgtgcctgtt cctttttttt tttttttttt tttttttttt ttgagaagga 60060 gtctcgctct gtcatcaggc tggagtgcat tggtgcgatc tcggctcact gcaacctcta 60120 cctcccaggt tcaagcgatt ctcctgcctc agcctcctga gtagctgaga ctacaggcgc 60180 ccgccaccat gcccagctaa ttttttgtat tttagtagag acggggtttc accacgttgg 60240 ccaggatgat ctcgatcttc tgacctcgtg atctgcctgc cttttcctgt ttttggtcca 60300 tactgtgggg acaagatgac tcgtggtgaa aaagaaaagc ctaggaacat agataagcca 60360 agccccatac ctggtttctt ttcctttttg tgctacaact gcctaaatgc gcatctccca 60420 acatgacctc ctcacctctc ccttcttcct ccccagctac agatctcctt atcttcacct 60480 gatgcataga cacccaacac aacatcttgt tttgagacta cacgctattg tggtctgtta 60540 gagataagac tctggaaaca tatttcagct acactaccta ccagtggtac gcgacaggac 60600 acgtttatgg aaaatgggga tgagaatgca gttttcctgg gtttgagaga attattgagt 60660 atttatgact tagtaaagaa ttatttggaa ggaacagacc atctgctcaa agggaacatt 60720 gattgcaagt gtcattcggc ctttctctgc ctgtctctca ctctccgttt gagccacatg 60780 atttctgtct gtgagcctgc ctctgttttc tcaccttgaa cactggcttt attctgtttt 60840 gttttggttt gtttctctcc ttgcatatta cccaagtatg gctgattgtg actgggtgta 60900 atgacctccc aatcaaaagc cctaacgaga ctccccagaa aacacccaaa ggtgaaggtg 60960 cactgggaaa aaatgaagtt gcgtgggggc atcagaaaca ggaggcgtgg ccgggagcag 61020 caggtcagga gagtgcaaat ggaagganan gagcaggtnc tgctgaagcc aagtattgga 61080 tttccctgca gagcaaaccc aaagtcgttt gaaaccgccc tttttaatcc tctctgaaag 61140 ccttgcccca tctctgctgg aaccactact caaccaccct ccttagcttc atggaatggg 61200 aggagagagg acacaactct ttggccatac taagtggtga gacttgactg cagtacttac 61260 aactcaggga tcatttttcc tgtcaaactt ctraatctta taggacttga gaagtttatt 61320 aagagctaaa aaaataggct attacagtct cacctgaagc tcaaatcatg ctgtataaga 61380 tagtttcttt agagaaacac cacgtagtta ctataaactt acaaatggac aatttgtaaa 61440 ttggtgctat tttacaacat aatattaagt cgatttcctt gcatttaaaa aatacatatt 61500 ctgaaataat ttagaaacac agataaaaaa tacctatgtg tctgtcagaa ataataatag 61560 ctaatattta tggaccactc actgtgtgtt ttattaggta ttaaattatt taatgctcac 61620 agcaccataa aggtaggtac taatacgatc tgttttacaa aagaggaggt tgaggcacag 61680 acagcataag gaatttgatg aaggtcacat agttactgaa tggggatttc atctcaggca 61740 gtccacgtta tacttttaac tacttcattt tcacccctgt gtattaccct actgtgaaaa 61800 taacccagtt tcttcattca tttattcatt ccactatcga tggacattga gcttgnnttn 61860 nattgttttg ctattgcaga ccttctaaga cctttcttat ccgtgccttt gtatgcacac 61920 gtgagagagg tgttctaaca tatatagcta ggacagcaat ttctgggtta aaggatatga 61980 gcacctttga ctctctgaga tatggcagat ggctctccgg atgatagggc tattttgtat 62040 gccttctaga aacgtgtaag ggtgctcgtt gtctacatct tcatctgcac ttgatgttat 62100 caggctttta caccagatga cacgaaatgg tgtcctgtaa tgtaaatttg catctccctg 62160 atctccctga ttaatggatg ttgaacattt ttaggttcaa gggccacaca ttttcttttt 62220 tttttttttt ttttttttga gatggagtct tgctctgtcg cccaggctgg agtgcggtgg 62280 cgtgatcttg gctcactgaa agctccgcct cctgggttca cgccattctc ctgccccagc 62340 ctcccgagtt gctgggtcta caggtgcctg ccaccacgcc cggctaattt tttgtatttt 62400 tagaagagac ggggtttcac tgtgttagcc aggatggtct cgatctcctg actgtagatt 624 60 gcttgtttat ctcatttgcc catttatctg ttgatttttc taagtgatgt tcaggaattc 62520 tgagtgtttc taaagggtat attgtttctt tctaaataaa aatttaagat attagatata 62580 aagacaaatc taaaatatta aatgcttgac acatgtgcct ttttcatctc tcccatgttt 62640 agtcaataag gcaattcttt tatgtggtat taaagaaacc gattagaaag ttgcttcgtt 62700 gtgtgcttaa gtgtgagtct tagaaaggtg aaaagttata attgactctt ttagcaccaa 62760 accaagcagg tatatagctg tttaaatact attttagcat aagccatagc tgtgggaagg 62820 gtcactccag tgcttgataa atagaagtct attcaattgg accctcattc ctcagcggct 62880 gctgtgacaa gccgttttcc cgctgctgct caggtaggag ttttcatctc ttgacacttt 62940 gacagatgtg gctgcacact tttgaaccat cttccaacac acccccgtgc tctattacca 63000 tgaggaaagt ctctgacaag agaatgaaag tacatatgca tgagacgtgg tacaatggat 63060 gtcagtttcc ctctggtagc atattccacc gcgtgcctcc tgtgaggaac agagggaggt 63120 aacaggtgct tgctgaaagt aggtcaaacg gtggcgaaat tattccttca gtgttttaat 63180 gctctttgaa aacatttcca ttgaaggcta agattgaggg gaggtggcct gtgtgatgtg 63240 agcatttcaa gtgacaaagt gacatcctag gcactcctga aactgtcttc ttcgtgaacc 63300 ttcccacaaa ctggcagatc atttcaagaa ttgatggagc ccatgagaat gtaggttccc 63360 tccagaacgc agctctctcc ccagcccatt cccacaaaga aactaggttt tccctgcatt 63420 tccagaaacc tccaaaccaa cccattatgc ttctcctttt gtcctctcca aattcaatct 63480 gatccttctt caattctgtc attacttcag tattttgata ttttttgact cattctctcc 63540 actccatctt catattcctg ccctcaaaat caggtcagtg cttcctcaac agagtgatct 63600 ctgattcctt gattatattt ggtcattttc ttttttgttt ctctttcttc tctattgctt 63660 cattcacgat aggcatttgc caaaaatttt ctgaattata cgtgattgta catatccagc 63720 tgcgttcacg tgcagaaaac atagcctcac acacagcaga cccaaattct tactctcccc 63780 - 141 - 008252 ttaggcaggt cacagtcaaa atgcatgcag caggctcaca acatgcacac acaactcaca ggcacacaca acagtgacac gcatgctcac acatgcatgc cagtgacaca caaacacact acacatacac aacaactgct attacctgga gctaaaatgc aacagagagg tatatattca gaatagacac atttttattt ttaaacctca tcaaactaaa ctcttgaatg ttaaaatgtc gcaaaaatga ctccaggtta attttctcag tatgcactct atcaggcagt caaaatgagt tttattaaat gtcaaacgat aacacagcat agacacacag catgctcaca tcgtgcacat tgtacataca catatctaca acaatcacac acacacctca acacagacat gtgacacaca tgtacataat tcacacacag tggaggaaaa ataaatatta tacttgaaaa ccaattgtat atatgtatct cagatgacca atttgtttat ctggaagaaa ttcctgtctc tttacaatta cattggagac aggatgagtt aatacacagc tgttcaacac aaagctgctg tcagtccttc ctttccttaa atcacaagtg acttccccca gcatgtgtac tcaaaaacac cgcacacaca acacatgcct cacacagtga cgcacatgct gcacacacat caatgcacac gcatacacac gacatttaca catgcatgct gcgcacacac gtttgtagga aaaccaggaa aggcctattg aaatcctcag aatatatcct ggcataggat ctgtcttact tatgcagaac ctattatatt tattaattag aaatttccat tttctgtttg cttctttaag aatctgcagt acaggtttag gcatttgttt agattctaca aggtcctgca aatgctcacg acacacatac agcatgcatg cctcaaacac acacacttat cacacaatcg catgcacaca atgcacacat tcacaaacgg gcttatacac tgcttatgca cacacataca attcattttc aaaagtgaat aaataaaata taattttgaa ttctaaatat gattatattt aataaaaaat ggcaattaaa attttaaaat ttcatcctcc gagacatgtg caccaaagat ttcgtttggt acttaaaccc agaagggttt acataatcta taatgttttt tagctcaaaa ttgcattaga catgcacgca acactcatac tgtacacaca acacagtgac acatgctcat cagacacaca cacttcacac gcatatggct tgacacactg acacacacct cacatgcaca cacgcacata aatttctcag ataaaaatgg gctacatgta gtcagtggcc cccatatata cnaaataata aagattaaaa cagttttgct tatttgcatg agtttgaatc ctaacttatg ttgttgcatt ttggttttgc agggtttgaa ggagcagcaa aaatgtaagt cctaaatact ttattttatt aatgcagtgt aacagagaca 63840 : aggctcacac 63900 cacactcata 63960 acagacacac 64020 gcacgctcac 64080 catgcacaca 64140 gtacatacac 64200 acacacacgt 64260 acacacacac 64320 cacgcatact 64380 cacacatgca 64440 cctcacacac 64500 gggaatataa 64560 ttatgcagta 64620 gaaaggggta 64680 aaataaacta 64740 tttatttaaa 64800 attatattta 64860 tatcagtgtt 64 920 tctttgattt 64980 ccacattatc 65040 agtcctataa 65100 ccaagcattt 65160 gtcacctttg 65220 aagtcttgat 65280 agtgctgtgt 65340 tatgatgcct 65400 cactttgtgt 65460 ctaaatattt 65520 tatgtatgca 65580 attagaaagg 65640 =. a _> /ff+- +• +• rr"- uuuu w w w <-) "- 65700 caatttgaag 65760 caaaaccatc 65820 ctcactaact 65880 agaaagcacc 65940 actagtgaga 66000 ccttttagga 66060 tttttctact 66120 attcttattt 66180 gaaaagccaa 66240 tgagtgtatg 66300 ggctgagact 66360 gtcttcttgt 66420 aagcaacaag 66480 taatacatgc 66540 agcatccctc 66600 ctggctcatg 66660 agatagagaa 66720 atggccaatt 66780 atgaatccag 66840 atatatatat 66900 gaaagaaaag 66960 gtaagcaaag 67020 ttttcttcca 67080 taggttagtt 67140 aatccataaa 67200 aagatagttt 67260 ctaaattgaa 67320 ttttattggg 67380 atttgtatct 67440 tcctagtgct 67500 agtctctaaa 67560 gttcaactgc tgattctaga atggcgtcgt tacccatgaa ttttgtcaaa aaaatataaa actcagtact ctttctagac ttattattac gattttagca tcagaantca tactgetgga gcccaaagca attgtattgc caaagatagg ttttaggagt gtccagttaa atgtttcacc gagtacatgg ecttgeagga atatatatat tgttgcggaa tgaacacttc agtcaataat aattgaaagg tgtaggecac aaattgtgaa gctttcataa caggatatag ctgtagtctt tagtaaggca ccaccgttta tgtaggaatt gtcacctgtc gacaaagata gtctgttgag acatcatgta gaagtctagg accagggaac ctatgtccag ttttcattct gagggtcatg acatgtatag gaatggtgtg ttgaactcta gaaggggatg ecttgetaag ctatgtcaac cccgaggcta aggaaattaa actgagtact atatgagctt taactaatat attgttgaaa gaagtcatgc tttttgtaca aaagtgtgag tatccattat agaaaatttc gaaaagttct agtggcagta tcatagctat atactaaaaa taattagctg tagcagccct aagaaaaact attattcagt tattcaaatt aacatcaggc atgagagaaa ccaagtaagc taatttttac aagaanngga tgaggcctcc gcttcctgga gtgtcctaat aaagggtctt acacctcagg agtaccaaca ctttcctcag gttatatcaa gaagcttagg tttctactct tttttatttt agecaagatt aggggtattg tgtcatcatt gaaaccagga ggcattgtac ccatgtttta gttttaaata tcccaatgcc gtgactgeca taaaagatga tgggaatgtc attgtagtcc cacaacaata aactatgtgg .gtttaatcta taaaaaaaat attattgtgg aaggtcaaca tctagtattt cagccctggt tttgttgacc ataggagatc ttgttttact gggaccacca ggaaactgee ttactttcat aaggaggcag aaattattgt aacaccaggc ttctggacac tcttacccat ttagcatgtt agttcttaat tacttttcct gattttaagt tctaggaaat aatcaattca tttcttactt cagaaatggc tggcattatg acataaaagt aatgacatct ttcctgactt caggtaacta ttggcggagt atgegtccat aatggttgga aaggactaat cttcaccgtt cactgacact gactgccaca atcaccctgg atattacttg cactttgtat tctaggttac tttctgaagc ttaatacagg aagatgetec tgaatctact taaaatatat caaggaacat gtccagccaa tattcttaat ccataaatta ttctcctact agtgttctat acatgttgtc agacagatga taactgatat ttttgtgtgc attgataagg - 142 - 008252 cagcagaaaa ggactatatg taatatatat gacccagact agagtaacaa agctaaacac 67620 cttagaaaag cccagaaaat agtctcccct tcccacaagg gttattattt atgatcaatg 67680 tatgagtttt caaataccct acagcagact cagagatgcc ataagagctg ggggagctgg 67740 gtggtgaaca atcaagttga agtccaggag tcctgactgt tagcattttc tgctttccat 67800 aatgtaaatg ctcccaccat ggccaatttc aagtcaccaa catgacatca gtaaacatga 67860 aggtggagag agatgtgatt gcagctccag cacaccacag ctggggcaag ctgcactccc 67920 actgcagtca gagatggcat atacatacat acatgtcata catgtctatg taaacacgtt 67980 gcaaaatttt gtttgcagaa agtggaattt ttgatgcctc catccaaagt acctagtttc 68040 caacatcact gtagagggaa agagagaggg agactttcga gaaggcacac cctttctcaa 68100 agtgacacat ccaaagtgat gcatcacttc catttctgtt ctggttgtga gaaccaatca 68160 cattgctcca cttcagtaat agtaagctga gtgatgtaga tgagctatat gccaggaaga 68220 ggaaaccagc ttggtaggta taatagcttt tgctaaacca caaggcatcc taaaacttaa 68280 tggcctggac agacactaac tcttgctcat gaatttgcgg atcagttgaa cagccctgct 68340 agtctgagcc aatctcagct gactacagct gagctcaccc ctgcctctgc ggtcagccac 68400 tcgtcagctg gcatctctga ttttgggtgt ggctcgccat taattggaat gaagagaatg 684 60 gcagggccca tggccacctg gccaccatgc aacaggctag gctgcagggt tgatgtggtc 68520 gctcggggtt ccaagagcag caagagtaat ttctaataca caagtatttt ccaagtctct 68580 gcttacttca cctttgctac catctcactg accaaaataa gtcacatggc tgagtctgga 68640 gttggtgtgg aggatactgc caaggatgtg tggatacagg gaggtgaatt caattggagg 68700 ccactgggtt ttttcacaca ggctaagagg aatttgaaaa tcactggctt ggcatcataa 68760 tcaatcctgc ttagaggtga tttctttcac accagccatt actggtgctt tttccttctt 68820 gttaaaatct agtttgtgtt gtaaagtcaa gaggagctgg tgattgaaaa aaataacatc 68880 cacatgtgtt tgttgatttg tgatttcaca tctaacctta ctcttacaga gtagagtgaa 68940 agctatactc acatggccta gagataaagg gtctgtccag gattccctta ctcggctctc 69000 cattccaata aagagagcca gccactgata gggcgtggca gctggcaggg tgtccactga 69060 aatcataggc tcttggtacc tttgtggaga tggacatgtg gattcccttc tgtcctgact 69120 ctctaaggac gcatcaggaa cagacagatg ggaaaggcag tctctaagga tatccactgg 69180 ttatgttttc agtgttgata taagtgttac ctttttcata taaattcttt gtattggaca 69240 gaataattgt gtacattgat agtagaagta atgaatatcc agatgtcttc taaataaggg 69300 aagcaaaaga tgtcccactt gcaaatggga gatttatttt aggttttatg catttggtct 69360 tttacattgt gtccttgttc ttatgttcat ggctcatatg caagatggtt tctgagaatt 69420 tcatcatgta attgcattgt tctccaaata agtttatttc aaaatccaat taaaacacaa 69480 gttttagtac atggctgatg cagtattcag tggaggagac tttgttttac taagttgagt 69540 tgttttactc actcaagcag caatttaaag acagatttca tttctgagct gtactattca 69600 ttatcctaat ttatatttat tgcaacctct gagaccacta ttttgtctct ttgcctacag 69660 tcttcagaat ctttaagtta ctgtttcata ggaaaaatta ttggagaaat ttaatagaaa 69720 gagcttttat gaagtactta agcatttata ccacaaaaaa agataagata gctttagatc 69780 attttgaaag tttataaatg ttcacggtcc atttcaagat tactaatctc ttccnnnnnn 69840 acctgatttc ttatgttaca ccttcaggcc ttccctgtaa gcaactgact gcatctctac 69900 tttcaaatcc ttagagtgct ttccaagact gaaataatga gagttacaga gcacacggtg 69960 tccactgcag gctagactct acactgaagt ctcatttatt ttcgttttac agccaaaaga 70020 tgtagatgtc atcatactca tttcctctgt gagaaaactg agtcacagaa aggtgaaatc 70080 cagcacagag acaagattta aactcagttc atcctgtctt caaaattgat attccattaa 70140 accatggcaa tcttaaatct ttctgtgaaa aacagattgt cttcagcttt cagatttagg 70200 gttcatgtac ttataccttc tatccaacca caaatacttg gtgaggaagc tgctgtcact 70260 cttttttaat atatagagca atgttgactg atgaattttg actgtctgat acaaggcaag 70320 agactacctg gtttgggctc acaatgtcta agatgtttct atctctgtgg gcctagaaat 70380 ctacttagac tttctgagcc tgtttctcca tatgtaaaag gggatgcaaa agtatctacc 70440 tcacagagtt gttgaggatt aaagatgtta ttacatgagc aagcaacatc tttggaatga 70500 atcttggcac aaaccaagca ctccagaaaa cagtagctct tatttcaata atccatcctg 70560 ttggtgtcca ttttaattgc gtgagaagcc ctccccgtgt ttggcttaga cttccacagt 70620 ctcctgggag gtttgctttc atggcaggac tcagaatcaa tgcgggaaaa agtgaccttt 70680 cttgttgttc tgcttgaggt tggactcagt cccattgttc cagactcatt gtctcattac 70740 acttttggac acaaagcaag atgaaagcgt tggttgctgt tatgagtttt atatctgtag 70800 ttgtcaaagt tacagagacc ccacgttggc aagtcacctt gtgtggaacc ctatggtcag 70860 gtaccatttt gtagccctgg gcacttttat acnaatcatg tttcnataga aggtgatttg 70920 agctggtata caagacagtt tgagctggtg tctgtcaaaa tcgttgacaa tgctgtgtta 70980 gtttcctgtt gctgctgtaa caaataacca caaattcagt gacttaaaac cacacacatg 71040 tattctctta gaactctgca gttagaagtc caaaatggac ttccttaggc taaacacaag 71100 gtatcaacaa ggctgcattg ctttgggaga ctctggagaa gaaatggctt ccttgctttt 71160 ttcacatcct ggaggccacc tgcatttgca tttcttaact ttatgagagt ctgtaatgat 71220 gtctttataa tgcctccaaa tggttgcaaa actaacacat gcttttgttt ttatgaagcc 71280 aaacagattt agttgatttt tgaccatgga gttggggtat ggggacaagg ttggcagcca 71340 - 143 - 008252 tttttatgtt tagtcattat cccagtgcag gctcacgaat tggggcctga acactttgag atagactctt tcaaagccaa tgacattgca ttatattaaa atgtactcac gagggataga tattttgtgg aaaaaattga atgtcatcag gaattgagtt taatatttaa agttactgct gtaagtgatt caacctaatt agtttcctga taagacataa cagatgaaat gaaaaggaat aaaaatttta agtcatttct tgatttttgt ggtgacagga ttagatcctt taattaattc tcacaaaatt K.^ w -.W-.-.UVfW gtactatttt acagtgccag aggataagga gaagaggcag atccacagct ccagacagta tgtgtggtgg tcctttccat tctctccatt atagactagg caccatatag ggattatgac aatgaatgtg tcatgctaaa aactggggat ctagtatcat caaggatgcc ataggacaag tgtttgcaga aagaactgat tagcatttct ttacggtagc cataatgaac atattccata cagtctacag aaaaaaattc acaaagctgg aaacgccgtg cagaaggaaa actggggaaa aactaacccc agtgtagtcc agacttttag cctgctatta cacctgggaa gatcctgcat taacaatgct tgtattttga actaaaaaca attaaaggaa atatataaat atacacatat tcatgcatcc tactgactca aaagtataaa cctttttaca gaagtgggcc atgtaggtac ttggagaaag attttaataa ataatgacag cagccatggc atccacagct ggaaatcagc aatagtaatg atgagcatag gcaagtaaat attacctgta agccttaata atcaatgtta ttcaacctcg gcattccttt aacatattac aatcatgaaa aggagtatcc ccgtgttaaa tcttaataat cattcctgac aaattccagc tatccaaaac actttaaaaa aaaaaagaat actagagatg atattaaata gctttaacat tttggacttt ttaaagaaac atgccactag tgcacatatt tcagctaagc ctctgaacca caccaaagtg taaaagcttt aaggaaaata agaaatattt gaatttaact aaatgtgcca tgatgtgcat aaataaagtc aaagtctact cttacctcac ggtaattaat tttctgtaaa attccagcca gcatctgtgg tattctattc ctgtgaagtt gcaaataatc ttataggtca cctacaacag tgcttctcaa aattcaacat gatacagatt ctgattcagt aggcctggga tgcttccagg ttgctgctgg tccagggacc agataatgct gccttgggga ttaatctagt tgtcatatac acagataagt aatttttgga tcctctgctt ttaaaaaatg taaaccaaaa atcaaatgtg tatgtggagt ctgaaggacc atgcacacac atatgtgaat ccacatatgt tatgtgttca tctgtctaca tagataggta atgcaattgt gaagacctca agtcatataa tttcctttat attaattttt attagaaaat cctgtagata caaggatgaa tctaaatgta tgatagccag tttcaagcac tggctgaagt ctctaacttt aaagctggag ttactgattt tatcacctta attcttattt aatatttgaa aatggtgatt atagtgtatc aaaatccatt acagggacat gtagacattc agacaaaaaa aagctgtgac agttacaaaa ccaatttaaa gtttggatct ttcagtaata acagtttact gaaaaagaaa gttgtggctt ttctgccatg ctaatagaac attatgctga aaggagtata ctcacaatgc aagaaagact aaataacaat gatgatttgc ctatctgtgt tatgaaggtc ataaacatgt tgattggcag ctctggaatt aaagggaaac ggatatgcat caatagccac ttcccgatgt cactggcatg tctcaaatta tagactatgg gctgagtctg atatgcaaca ttctcattcc cgaacagaaa aatgcttgat cctttgtaga atatttgtaa gtgtaatcac fff aaanff a a ?• а а г+л a rr +¦ r* t-1- +¦ rr /та a rr ctccacttna taagtggtgg agaccccatg ttcttgttct aaggaaaata agggcatgtg cgctagtccc aggtctgctt taagtattct gtaaaatgac agctatggtg caagtcggat atacatccta aaagcatttg agaaggaaca actgaatggg cactatcacc agaaagatta tgatatgatc gaacaaacct ataagccaac tacaaataaa actatcaaac ttcatggatt atgcaattga taaaattata gggaatcaca gtactgccat tccacacacc agagagttcc tatctctgta tcaatgagaa agttacggct aaacacatac cttaccccat gctctttgag ttaataaata tgggcagatg ctctctcctc ctaccccaca ttagtctctc gatattaaag ttatccctgg tcaacagaat gcaaaattca tacatcaaca gaaaatctga tctgttattc aagggcatcc ttatatttgg agtaatgttg agcaaacagt ataaaattcc agtgatgaaa ggaagaatca atccttatca tgaaaccaca ttacctgact aaaaacagac tacagtgaac ttcaataaat aggtacaaaa aacagaggtt ccaggcagtt atagagtcat tttagtaata ctgcaaggac cccagaagaa tggctcagtg tttctgctgc gtgggcagcg tgaactgtgt tagataaaac gatacaagga gaaggataaa acatctcttc taataaaagc aaagcctttc aacatagtac aaattggaac aaaaacctaa caggatacaa gtgaaaaaga ttgcaattaa gaaatcgaag atattgttaa aaataccaat gaagagccaa tcaatttata acatagacca tcatttttaa ggtcctggac atcacatcaa taagtaactc tggcccagag ggagacattg acagtttgct taaaggattc atgaaacctc gtgtgttgtt ccagggcctt ggccccagaa gaaatgtgag aaataacatt tgattcaaca agctatatga atggtaaaac tatatatgac ctctaatacc tggaagtccc ggaagaagtc agactccaca aatcaacata aatcaagaaa ccaaaaaatt aggacaccaa aatgttcata gatattcttc atctatccta attcaaagct acgaaacaga caaaggtgcc atccatatgc aatggattaa ccctggggcc cacctgatgt gcagtgctca taagacataa tcaggtcaga agaaagagga tgtgctcagt cagcctccac tggactttaa taatatatct tgaataatgt tatgcaaata tcatttcaat ccctcagcaa agacccacag tgcaacatga agctagaaaa aaattatcct aggaaactat caaaaattag gtaatcccat gaaggatctc aaatggaaaa ctacccaaag acagaaatag aacaaattga acagtaatca atagagaacc aagaacatac agaagaatga agacttaaag - 144 - 008252 ctatgacctc ggactgggca ggacaaatgg agtgaagaga attaataacc atcaaaaaat caaacaggca ctacaatgag ataaatgctg aattagtaca tgctgtaatt ctaagagaaa aacctaagtg agcactattc tggagatcat cacttatttg ttggttgcca ggaacaaaaa ctgcagtcaa ttgtaactca tcaaaacatc tgtgtgtata tgaataatat tcattatggc gggcggatca ctactaaaaa tcgggaggct gatcacgcca aataataata cagtagactt agtgggaaag ~--.4-r-.4-4--. 4- n -> yw-yLLOLVja atgtacatgt ataagtaaaa aaaatatttt ttactagctt accaaccaat agtcaaaagt ccttgcctgc tgcatagcag atgaggttta catccgtttt gcaactacta taagcagctc tcttagagat tcattgtctg accgcgggcc ctgggtctta tcagcgtaca aataagctgg taatgataaa aggtaggttt aaactatgaa aagattttat gattacatca caacacacag agaatatata gggcaaaaga tatgaaaagg acatcatatc gtgaggatgc atcactatgg tagcaatccc tcttcactcc tccatcaaca aaccatagta tatgttaagc tgggatcaca gaggctgaga aaantagaaa taataataac aaggataccc ttgtgtactc tatatatata ggcctggaac caggcatggt tgaggtcaga tacaaaaaaa gaagccggag ctgcactcta aataatactc ggaccagact ttacttaacc r- 4- 4- - -.---.-- - 4- cacttactag aataaatctc catctgcagt tttaaaagcc taaaaattaa ctctgaaacc ccagttcagc cacatcacaa tttctttgtt ccacatgttc caaaggagaa tccaccgtgc gcaaaagtga cttcttaagt tgagtcatct ctctagaatc ttagtttgtt tcagctcagc ttactcctct catgattaaa attactacaa gagcaatacc agttacaaag aattagagaa aggacctgaa tttgaataga tgttcaacat accccagtta gaagaaaagg agaacagttt actgctgggc catgtttgct gacgaataga aagaacaaga gaaataagcc aaatcaagat tgggtactgg gaatagataa tgtacatttt cattctccat cataaatata tatatannca atagtaaccg ggctcatgcc agatcgagac ttagccagac aatgacgtga gcctgggcga attatttttt tgggttccag tctttgagcc gtaaagcacc aagttgtagt aggtttctct attctcccta atttatccat gatgcaagcc atcttcctag agtgtgggac aggtgctgtc aagagtctgt atcctgagca aacttcagga agatcgctgg agtgtagcct ggttcctctg tcctcacagt ttgctgtttt tcctactaag aatacgtaga agaaacgggt taagcatctc gaaaacactg cccataagca cttctgcaca aatatttgca acaactctat catttctcaa cattgatcat aaatggttta gaacccttct gaacattcct atatacccaa gcagcactgt taaagaaaat tccagtcatt aggcacagaa aatcaaactc ggggacgggg gacctactat aaaataactt gatatgctta tatnngtgta cttactatgt tttaatacat tgtaatccca catcctggct gtggtggcgg atccaggaag gagagcgaga aatgtttaat tattgagtcc tcagtttcct tagaaaaact aaaattttca tacttttcca aacacagaat acagtcagaa atacacaaat tcactgttta atggatggct agccaaggac ctgtttcgcc tggtgaccgt ggcagtacga gcaaggagca gttcattact gtcacagtgt atgtcacgtt cctatccacc tacaagcagg agagggctgg gttacaacct cctagcactc gggaaaatct ccggcaacca gcaaaagata aactacccct aggaaaaatc aagaagacat cagagaaatg tattcaaaag acactgttgg caaaaaacta aagaaaggaa ttgcaataag acagtacata tgtaacaaca agacaaacat atggacatag ggaggcaggg ttgctagcat aaagagtgta ttccccattg tatatatata acccacaaat gataaccttt acactttggg aacgtggtga gcgcctgtag cagagcttgc ctctttctca tatctttatc atcttcaatc tattagaata ccaggtagag acaaatattt gttgtataat aaagaaatac ttaaggataa gatatatgtt gctgcctgaa ttatcttgga taagtcaatt ccctcagaaa ggacgtgatc cgagttctac ggtgaaccgc tgactttcac ggcctgattc gccagggtaa tcttattatc cacagattca aaggacatat acattaaaga tacatatcat ccaggacatt aagcaaaaat caattaacaa ctgacaaagg taataatctg acaaatgaca caaatgaaaa tcaggcaata tgggaaggta aaaattgagc ataagtatat atttggaagc tgcgtaatgg tggatggaac cacatgttct agagtagaga atggttaatg aatacagtga acataatcat catgcctgta nntttatgtg atttttaatg ataataatac aggccgaggc aaccctgtct tcccagctac agtgagccaa aaataataat ttaatgtagg cctgtttgac ggttcctagg -. ~-. - -.4- -.4- -" ay u> aay ua uy atggnaagta aatcatctct ttgttttgtg atactaccca ttccattttt agaaaggttc agccctgttt acacttaatc atgatgcggt gtggcggcta ctggcggagg ggacagtgcc caaagctgaa atttcctgtc cccttttccc tctccctttc gaagatcata agagagatgg caccaattaa attacagttt aatttttatt aagatctttg acatatttac tacatacaca acagaaaaaa atttgccata gatttggtta gcatgtacct tctgaattct ttttactggg tctttcaata tttttttaat aaaaaaattg gttgtacaac gtcacaattt taattaagtg ataaaaattt aacgagcata cctcttcatg ctacagtctc agcattttgc gtaaaaactt attgacaacg gcttcaataa tattattttt ttattacatt agtctaccca gacatctacg gaagtatgtc tgtgatgcaa atttatatta acctaaaata tagaaaccca atccaaccta accaaagcat cctagaattc cgcacaagct tggatatcaa tcctgaaaca catttgccca aagcatttta tatctcagaa ttcattgtgt tgaaaaagta ttttcacata tctcagtaca tttctgaaaa gaaactttta aaataccgtt atggccacct statttt ataattttcc ttcaatggtt gagtgaggta aactgaaaag aaaaaaatca tttaaaaatt agaattgatt agtagtcgct aatacaaaac ggtatatgga - 145 - 008252 gcttacaatc tagcaaaaaa aaaaaaaaaa aanncaataa acaataagag tggagaacta atgacagggg ctgatgaaga aattatatcc agcccagcct ggtataattt tgctgaaaat ccagcacacg atgaatggat tgtcaataaa aggtcgggta tggtggctta cgcccgtaat cccagaactt tgggaggcca atcacgaggt caggagttcg agacaagcct ggccaacatg gtgaaacccc aaaatacaaa aaattagctg ggtgtagtgt cgggcacctg taatcccagc gctgaggcag gagaatcact tgaacctggg aggcggaggt tgcagtgagc ccactgaatt ccagccaggg tgacagagtg agactccatc tcnaaaaaaa cagaaagaag cctagactga tactaatatt tgcaggtaaa tttctaacac caatgaagga agcctcatca agttatttct gtttttccca ggaagtagga gcatccagcc agatgatgag acagtgaagt gggaagttgg caaagaaagg gaggtgcctg gggttaggca aaaaattaac aatacagcaa atgcaacatt ttaggactat ggatttacag tgataccaat tcagatagct ctgcgacttt ataagaagag ggaaaagcaa catttaaatc aatccacctt tagggatttt ggggaactga gaaacaaggg gaaaagatgg attgctggca aaggagttga ctgtggtttc tgttggatgt ggaaatggga ggagaatgga ggagtgaagg atggtctcta tatggttaaa ggacgggtac caccaaagtg atggaataat gaggttatga tcagagagtg gagtccagga ttttacaatt ccagggtaat gaaatataaa atctagggtg tcacgtgaat gtgggtggct taaacatatt aggcagaaat ggaaacgact gttgaagcta aggcgctggc tgagtcttca gaagtcactc agagtgaggc cagttcttgg agagaagagt ggagtcaggg cggggaatga gcgagtgact gatgaacaga ttgacagaag gacagaagac agagtgggtg acagcatggt ttaacctcat ggaatacgtt gcaaaggagt aatggatggg cacctcttcc tcgtctacat gttctactag aaagggctgg aggggtaact catgtatttg agagagaatc agttgaggca cagacagaaa ggaacagaga ttgataatgt agaggacatt atttaacgta ggagagactc tcaaggctct gaggagggag aagtgggaag atcgatgaga cagaagaatg aatagcatca gtaggtaaag atttgctggc aaaaatgcca ggaaacaaaa ccacaagaga ttacagttat gyttatctct ctgggaggag gaatctgcat gggaatcttg acattagaga ttacagtaac attgcttttg tgatctagca aggtggtcac actaaatcag tcctggagca taagtgaata ggccctccac пгаппггз -а л/ч i+-+-+-+-a -.-+- •" -.-"+¦ 4- +¦ +• f/".*•-(¦ ttcctaatgt gtttatgtat ggtagatgat ttatcctcat accttaagaa aatgtcaaat aattgaatgt tgtagagttg actggttact atttaagtgt ccaggggtag ttggtaaatc cttgtctttt ttctttttta tataatatta caacctcaac aacagacaga caggattgga tggtggaatt cagtatatat ataattgttg tcccctgaca ataataataa gaaagggtgt gggaggaaac ggtggatatg tcaatggcat tgatggtaat .ggtttcacag gggtacactt tcatcacatt ggtacattac atgaatgcag cttttaacat gtcattcatt tttttttttt ttttgagaca gagtctcgct ctgtcgccca ggctggagtg atctcggctc actgcaagct ccgcctcccg ggttcacgcc attctcctgc caagtagctg ggactacagg cgcccgccac tacgcccggc taattttttg agagacgggg tttcaccgtt ttagccggga tggtctcgat ctcctgacct ccgcctcggc ctcccaaagt gctgggatta caggcgtgag ccaccgcgcc tcattcattc ttaacaaagc ggcaaatcag ttaaccagct aaaaactaac ggaaaaataa ataaataaat aaactaggta gtggtgattt tgttgactta tagaccccat cagtgtactc ttgaactact taagcattct gaatttgttc tttggggttt ttttgttgtt gtttttgtct cctttttttt tttttttttt gtcccactct gttgctcagg ctagagtgca gtggcaccat ctcggctccc gcctcccaag ttccagtggt tctcttgcct tagcatccca agaagctggg cacccagcta atttttgtat ttctagtaga gatggggttt cgccatgtta tctcaaactc ctgggctcga gcgatccatc catcttgacc tcccagagtg астПГУпЬпапг* rahr-rf r"r-t" rrnr^rrii- t- r"r"a" r*t- r ; ttttgtgcta 78960 : ctgaaaaata 79020 , agcagagaga 79080 . aggcgggtgg 79140 atctatacta 79200 : tactctggag 79260 caagagcgca 79320 aaaaaaaaag 79380 agggaaagaa 79440 agcaaggtct 79500 ggagagggaa 7 9560 aaagatccat 79620 ttagccctgt 79680 tttcaggtta 79740 aatgatgacc 79800 tgaaggatca 79860 agaaccagaa 79920 tctaagtaat 79980 gatggagtca 80040 catggacatt 80100 agcaaagtca 80160 agctacaggg 80220 atcaaggagt 80280 aaagggctgc 80340 gtagattctg 80400 aaagattagt 80460 atacaagagt 80520 gcagaaatga 80580 ctgctcctga 80640 cagggtgggc 80700 actggcaaca 80760 aatfnatrrf 80820 attccaacaa 80880 attgtagagt 80940 aaaataaaaa 81000 tatgtgtcca 81060 tttgagaggt 81120 attcccaagt 81180 cttttttttt 81240 cagtggcggg 81300 ctcagcctcc 81360 tatttttagt 81420 cgtgatccgc 81480 cggccacatg 81540 ttgaaatcct 81600 aacaaccaga 81660 cactctaggt 81720 ttttgagaca 81780 tgcaacctct 81840 ataacaggca 81900 gccaggctgg 81960 ctggaattac 82020 rrt-1- t-rrar-l-t-t- 82080 atcagaaact 82140 taattaaact 82200 agtttaggcc 82260 tttgtagacc 82320 gtgtctcaga 82380 gctatagaaa 82440 aatacaaata 82500 aagctatgat 82560 atttaattct 82620 gtatgtaagg 82680 - 146 - 008252 caatagcatt agcttttaaa tctaaaataa gaataaatta ttaattaacc ctatgcataa tgtgatctaa tacattttaa tattatatga cacaggagat acagatggga ctgtgtcacc aatggggagc tttccaatga taaattaact ccatatctcc aattccactc ctctgctatt ctcttagtcc ttcacatgga agaagggcct aaataaagta agaagctata ttgtggcaat acaatttcca taaatccgtg ttaagttgtc aagttacagc gagactaaat tacctatgat ttctttttct ctttntttts attataacca gtttgggtca gtaccacttc atgcatgctt tgggaaattg gggtatttca attagaattt ttgtgaattt aaagtgcttg cttttttttg tatacatttt atttcctctt agtagtatgc tcttttcttc tgattcagtg aaaggctaga gataaggact ttcattcttt acaattgtgt tgttgtcata fnbrnatat t-- -э - - -i- - - - w attatgaaaa ttcagttatt tcttccacat ataatttcaa ttcatagcca atttctatgt tcatatctct aatagaggca tgatttcatc gctcacttct atgtaatatt gaagttattt agcactttac agatacttag tacatttaga acagaaaaag gagaatatat aatatatatt atgtatttgt taaaagcttt gagatacaac ccctagcatt tcaatcctct agccagaggg agaactcctc ccaacttgtc cacccccaga aagaggatct cccacaggaa tataaaattt agtatccttt aataagatat gttctgtgag aacatgattt agtcagtgtt tttccaatag ttgtcggagg ttgagagaat taattttaca gtcagaagca atggttattt ttgaaaagtt tttaagttcc ccattaagag cttctgttgt attagttaag gtaaagttta tattattgca attttcttaa atccattaga atcttctttt aaggaaggca atccccagct tccacctcct gagaaagaag ttaccctcaa atcagccagg cttgcatctt agattcctca atatttattt ttcatctcat cttacatggc ggccatcgat attatctggt atcaaagaaa tgtctcaaca atctaaaaat gaatgagtca gttgtttcta caaataagat accattagcc ttccttcttt tattttaaaa cacatattcc ccaactcaca gaagctgggg aaacaggaaa ttttcctctg cattaaatca gaaataaaat ctgtacattt atctcaattc acggacctac atgttctctg ggtgttaagt tcctaggact ctctgtctat caggctcttc cacttgcttt gaaaagaaag taaatacatt ttcaattata agttgtgctt gtgtgactct gtaatcttga ttttagtgac catcatatcc cattctctat taaactgcca aaacagatta ttgggcagga tttgtgtatt ttgaaatttc atggatcaca agaataaatt gtctaaaaat acttgttttg cagtttacct taagaatttt tatgttataa ataagcaata aacaaaattt ttcttttgag agagaggccc atttcctcta attaatgagg ctgtgctaca ccctcctacc ccctgctact ttgttcagca tatcatctat cagagtttct aagtcagaga ggtgatgggg aaggacctct gtgaggtgta tctacaagaa atatgagcag tgtggtagac aggtgctagt ttgatgaagc ttagacttat aatatgatta tatcctttga <- V- С l"V_- "_ W- \-¦ V_ atatttgtgt ggtttttcaa ttgatctcct tataatagta attcatgtcg attgcttcca ctgtgccaca gtcagatatg tgtcaattta ttcaggattt cttggttacc ctataatttt agtgcagaca aatagaagtt ctttttgtgt gttttcaagt tgattttttc cctcactcct ttctggacac agtgctatcc r"t-1- r*t- i-пяпл tttaattcat ctaaccttcc tttttgttac ttacattttt gcaactttac agataattgt gatttttttc gtgatagtgg attaatgtca atttctaggt aat ua^c-aat-j gtccgtcacc tcagaaataa ccttctccct tgaagtatta attgaagcat actttttgct tttgggagag ctcactttca cattcctacc ttaagtttct atagagattt gcctgtttaa tctcccagcc attaatttta cttatctata tgtgcttttt cccatatata gtagtgcaat ctcagtctgt tttataataa tttgttttat tttagtaatg ttagtgtttt attcatgcct attgatgtag tacaattggt atcttctgaa ttcttttatt acatgctggg tttgtaggtt tgccaagagc cagattttta aacatcatta aatccacaga aggtgccact gaatttattt gttgaaaaca ctctttctaa actttgctaa aggatatatg tacaatctca gacctttttt tctgtaccct ttcagcttgt acactgtaag ccagagtcat caattttaga aacaattgtc ttctgtcata gtctatacat accttgagat aaaaattaat gtagagtaga gaactatatt ttttcaatcc gttctgcatg atacttcagt atatatacct atattgttaa agcagctcta tataatcatt tcgtattctg aattgatttt cttttaactt atatagtcat taaagacatt ctccaataca cttcgaagat tatcaggttt caaagcttta ctattagggc 1- <-1-t-rfi~-r- = 1-t- agattttaat ccaacaatgc acatatccta gaataatatt tgtttgtgat attaaggttt atccgggtgt tgtttttcta tttatagatg atttctcatt tgatgttaag tttataattt cagtttcttt aattttttat catccataga tgtctttcca gggtctctaa tattttgata cctttcatca atgagttgag acaggtcatg 82740 atttagaaaa 82800 caccctttaa 82860 ttactttgga 82920 cttttgtagc 82980 aacttttcat 83040 tgaacatgat 83100 tttttcttat 83160 atcagcacag 83220 aaactcacaa 83280 aaagttagct 83340 gacctatctg 83400 acagtcatgt 83460 taccccacag 83520 cagcctactc 83580 catgcacagc 83640 aaacaaaaag 83700 tacttaagcc 83760 ctgcaaagat 83820 acagagagga 83880 aatcagaggt 83940 ttctactctt 84000 tacctaaaat 84060 gaaattaaga 84120 ttttagttat 84180 taaatatgta 84240 tcgctgagag 84300 aaaacgtgct 84360 ttatgtcaga 84420 ttttgcagga 84480 aaatctgtga 84540 v-aijavjaHcia с 8 4600 agtttttcat 84660 agatccctgt 84720 catcatatca 84780 tcattatatt 84840 taagggcaat 84900 aaacattgtc 84960 agacttaaaa 85020 attgaacttc 85080 cttcatacta 85140 atagtcttaa 85200 tcagccctga 85260 tcagttatct 85320 ttatttgctg 85380 ctttaatttt 85440 ctcttatagt 85500 tttggaatga 85560 tcattgaaaa 85620 acacatgtgc 85680 gttgataaca 85740 aagctactga 85800 85860 aacatctcta 85920 tctccaaagg 85980 tgtgaatgaa 86040 attgacctaa 86100 cttgattgga 86160 cgtctaatta 86220 taaaatgttg 86280 cgaatgctta 86340 tgttaataaa 86400 tattcaatct 86460 - 147 - 008252 tcttctacct attagaataa atagcattac ataaggatag gaagttacac aagaatgaat ggtggagaaa ggtttgtcag tctcacattt aatccttttt ttctgatgct acaagattac acatatgaca atcaacaggc aatgactcac aggttgcaaa acatgtgaaa atggatgtga tgaggcttct cacaaggttg ccagggtcac ctacttccca cagcttttag ccaatatttt tttattattt tactggggtt gttcatcgca gatagactgg aaggaacgag caaactaaca gataatgaga gcctatcagg taatacctag taacaaacct taaaaaaata attttactga tttatttaaa ccctccctct ctggacccta atatgggtga tttttcagat gtcttagcct ctgtcttttc ccctatagga tgcatttctg tcctgttttt ctcattttgc gaagacattt aaatctgaca tcaatcttgc aaacttcaag ctggacatta catccatctc tgatttacag aatctgaatt ctgtaatctt tgaaattaga cactcatggg gttaaactct cgttttgaat aagcagagat agctcaaggg ttggccacct atactgagct tagatatttt ttttaataaa tagttctgtg ttcctttcac aaagtccctc gggagaagag tttcatggtt ccacagtttt gctgccaata aactacctgg ccttactcga tatcatatgg taaaaactcg agaaatcagg tcagagccag cttgctgttt aacaatattc gattgggatt gagtttctga cataaataac ggaacttgag tttggttgac gaactattaa tctctttttc ctctgggtgg gcactgttca ataaagaaaa atcacatcct caggaacaga acactgaata aggtgggggg gtcatgggtt gcatatcctg aaataatgaa gagtaaattc atctttgcat atgatcactt ccatccagag tatcacagtt aagctgataa tgggtagttg actatgaagg agcaattgag gtcaacaaag tcctttgatc cgtcttgcag ggttttatga cccacgtttg catagcagca taaataggtt tcttgttagg aaaatttgct acagctccag tcatgtaaga cctttgtgat aaacagagac caatgtccat gctgttgtta aaagaactgg ttattgaaag ccccagatac acttggtgtt gactacacag gatactgaaa caaatccaat attttatttt aaatggaata tggaacacca agatcttgat tattgcttgc ctctcaggat ttgtttattt attttagacc gatactaact ctaatatggt ggggtttcca aaagtgctat ccaaaagaag cctcttttca agagcgctga tccttaggta catggccagt aaagatacct gcaaaagcca actaacagtt gtccctttat atttatctct aatataaatc caatagcaaa tgtggtacgt ttgcagggac aaaccgaaca gctgaacatg aggggtagca tataggtgca cacatatatc aacaactaaa atagtttgaa gctgcctcat ctactttctt ccaggttttt cttgtcacta tttctcctcc gtctctgtcc ttctcatctt tccaacattg aggttaacct acagtatgtt ggagtcctct tatttgttaa agttcattaa gtctttgaca tcttctgaca taaaaatata ctttataaca actgatacag gaatgggatt aaatatgcat tgcaacacaa ctctaccatt ctggtagagg acaaaacgca tgaaagtaca agaatcttct caccccatgt actttccttt ctattttaga aagctgacat cttttacatt gacatatttt tctagggcta taatgactta tatttataga ttttataatc ataactttct aaacttcaca gtaagctcag tgatatgaca ctagccttct acttctgatt agatgcagag tggagtgagg gaaccaggga ggcatgatcg gctcatgaat atcaattggc gtaaaattct tctaatgtac ttctttgttt ttggtcatct attctattat gacatggaat atacactatg atggatagat ctgcatgttc gacacaggga tcaggataaa gcaaaccgcc ctggaacttt tgatggtgaa ttttagaatt ttggcttggg agggatattg aagtgtcatt gtctatattc tgcctctgga tttccttagc taccttatct cctgattctt cgtttagaat tgaaacagaa tttctagaaa aatgaagagt tcataaaaat ttattggtga nntacttttt atacaataat tactgatata gtaatcagta gcctgaactg ttttttctca atattagata ttatctcata gtctcgactg cagcaaagca cttcacagtg caggtccaaa aaatgaagtg ttaatttctt ttcctgaaac tatattagga gctagtctga tctctttttc ttgtttgcag ttttatggtt gtcattaatt tctattgaaa tttttataga gcacagaggc ggggttccag tatgacatat caagtttgat acatttttat agaaaggtct acatatcacc agctcacctg ctggaatcac gatctcattg aaattccaag ttatttttca ctgaaattat tttaaatttc taaacttctt aaagatacat caacccaaat gaatactagg cgggaagcca tcacttataa gaacaacaac tagctaatgc atggcacaca aaatttaatt aaaaaagttt ttgttttctt agattttctt cagttgcctt tggagaatgc cataaaatat ggcctgcagc agagaaaccc aaggtgcctt gacctagcat tccttcctgc ggggtttcat agtgatttct catttatttt tggccaaatg gtttaatttc ggggggtgtt gacaatctgg cactataagc atcacttttt attttcttgt tgatatccat acacatttca ggcctgtgtt caagttgtcc aggaaagaaa tgggagtgag taccccctag gttgcccgca agtgcagtat acttgatttt attttcaatc ttttgttagg tcttctgtga tgggatccct tgattattat ttaactaatg ctgaatgtat agtaaatgac acagtcctct ggcctccctg catatgacat aatttgctag taaagcaaaa ggagagtcct ctctcagcac agctttgtgc tggccatgca ccataaacta gatgtagagg ttcaaagaac tggttcgttc tcctctcacc tgaaaattct gcatgcatat gtccatcagg cagcgataaa tcatcctcag gtgggagctg acacactggg atgtggggct tttacttatg gaatttaatt gacaaaatta tctcattaga atttctctct aaccttgacc tagccccttg cttgttctga tgcctataag attctagtcc agtctcctca atgtttctgt ctctttggtt aaatcatgaa gcattcctct ggccccaaca aagaaaataa attttctggg tctaaaagat acttctcaac taagagtagg gttgttattg gagctttgtc catgattctt aatatctttc tacgagatta aggttcttga gaagcaagga cccaagcagc aggtttccca atcagaggct - 148 - 008252 gaggtgaagt tacgaagtta cattcctagg caagtgtctg gttgggaaaa gcaaccgatc agaggtactt tcaattatcc atctgccgca cagaaaaggt gggggtttgc aaagggagta gcccctggtc cttttgctac tcaggcatgg aaagttaggg tttttctttc aatttagttc taagaagtca gtgtgaaatg gacttaggtt ccctgcctcc agaccctatt ctcctgcctc attatggcga catcaaacca ggtatatcgg ttgagcatgc attctcaaca gggagtaaaa tcacctccaa aggggtgaaa atatgttctt agttgggggg tagaaaattc tctcttttat aatgcagatt gccttgtaga ccaaataacc aaagtagagt atgtaaaaag atatataata tggtagaggt atcaaaactc catgaggtgg tgattaagaa caaaatctct aaaagacttc ttagaagagg tgatttaaaa aaagattggg aaacactgct ttggggtatt tgtaagaaga aataaatatt tctcttcttt tctatgcatt gtttctaagt tttcttaaag taatgcaaaa aaaaagaaaa gtttattata ctcttacaaa ttttgtctta actcttaaaa agttaagttt taacttgtag gtactcacaa aggtggaatg cactgcagaa tatgaaacag aagaagggtc ccaatgagcc cttcttggtg caggggctgc agactgagga gggccaaagc agtggagggc tgccaggcta agagtgacaa atggaagcca ccgttccact tttccacagg ctgcctgctg aatgcctggg atttgcagaa agccttgaga ttggacacct gcacaggtga ctttggtggg agccctccag gcaactaggt tctgcagaga ccccagttgc cttctccctt ttagatggct cccagcatgc ccctggcaaa ttccaccctg tccagctacc atcaaggttg agttcactgc aataatctgg agctgcacat tgagaagtgg ttactagagc ttgcctccac tgcttctatt ttggttcttc tgaactatta tagtcctctg aatttaagtc ctgtaccaaa ccccataacc tacactgaaa tacttagtga tcatttaaac taatttgatt ttcctaatca ccttctttaa atcctgacac agttcttctc tatggcagga taacccatac cttcaagtta agttaggcat atttttatgt gcattgattt tccataaaat gttatcactt tggcatgaat atttcagaga gctagagtgt tggcttgtaa aagttaacaa attatgttca aagcaagcca tttgattcca agagaatctg gaatcaggaa ttctagggga aaatcagtga gactatagat atttaaatat aatcacataa caatttgcta aatcttttag tgccttttag tgtctttcaa aagaatatca ttttcccaca gagaaaattg taatgcaaac agatccagct gcatatttta ctataaatta tgctcattta aacagttcat tccatctgtc cttttgataa tatatgcagt tagtaaggtg attaattgta tgtatacata attagacaat ctattggaat atctttcagg cattcattag tagttttctt ttaaaaaaac actttaaaat ggatcaccac tctttttttt tggaatgaaa atctcagtct gaaaatacag gtgaagggag agatgatgta atgaatgcct attactgctt cctctgctaa tacagcaacc aggtcaggca tgtggatggg accttcatga cacatcaatt tcagtttctc ctgagattcc ctagtatcca taggagagaa aacaggttca agtcttcccc atatcattgc aaggccagaa ctaagaagcc tgtgctagag aggaagtacc tgtcgcagga cttgcagcaa agtctgagtc ctctggtccc ctgaggccac tgctcagggc agagagcctt tcatttcccg cagaggagct ggacctgcct gtatgctgct tgtggtccag gagcagtcca cagagtcctg tgggtctcag tgggccagga agggaagctc actagagact gaattactcc aggtggagaa gggggtcaat aaatgactcc actcatttct gggaaatcct gtcaccaaaa agccttcttc agatgttcag caaaggagct atggcttttc aggtctttcg cctccgtaaa ctatggtgcc ctctactacc cctcatttgc cactgcttaa gatagcataa aagacaaacg acaattacat caaaaacatg tatccatggt gaaggaaaga aaagttaact cacacacaaa gggagactca gaaactgagg atgaaactca gagaatcaag acaaatgtct agtctagaga tttcccaagc aagtagaatt gatgggcgag agaatccgtt gttgttgttg ttgttgttgt ctcaagaaaa agtaagcagg gcacaaaata caatttctgt aacttcaact aaaaatgtcc aaaaaatgag ttaaaaaagg gcatagtcct tgagagctag tttttaatat gggagatgaa gcaggaaaaa ataactcaaa aagtggtaga gatataataa taagggcaaa gtaaaatagc ctatccacac agctatgaat aataggagtc cctgaaggaa aaagcgaaca gatagaagaa aagttgtcat aaacaacgga agaaagctta cataaatgaa aaaaaaaaaa aaacataaat ctgcaaactg aaagagctca ctaaatttta gtgagattaa atgaaaatag atgcttactc atgtcctggt gaccttccta ggttgcaagc ataaagaagt aaccttgttc agcccctaaa caggctgaac ctaaacctga aaaagtaaaa taaatgtatt gtgcaatgga tgactcatct acaatactgg aaagtggaag gtaatgaatc aatatacata gactactgag agaatagaac tgcaactaca gaatctgaac acagacaaca tgttcatctg tcagtgttaa aaatgcattt tatatacaaa aggactaaga gagcataaaa ccctttgtat ttatttgaga aaactacttt aaagtgtgct aatcacaaga taagtaaatt gcaatctcaa tgactagaca aaagaagaga gaattgatgg gcattggatc tcaatagata ataataaata tgtcaaatag ggatctttga aaaaagagaa atgataacat taaagaaact agtaataaat ttgtaaatga tttcagcaaa accaggactt aggaaagggt gtgatatcaa aggagagtgg gatggtagaa gtgtactatg aagtttgcag atgtgtgggg tggagggatg agtgagccag caatttagtt gtgaagcaat aagaaaaaag cattggtttg atgtctaacc attgagaatt aagccaggag attagaaaaa ccaagtcgag cgtccaaatt aatgaatagg gaaagaaatt agcaaatagt tgaccaagtt aaaaaaagac aagataaggt taaaaatgaa acagcaatga cataaaatac acaggaagcg taagaccaag tataactttg gcacattttg tttgagcagg ttgaattttc ttatcaaaaa ccctagaaaa tctgagtatg tttaaacaca aaataaaaat tgtacactgc ttataggaag - 149 - 008252 cacatacaaa taagacgttt aaattaaatg ggacatttgc agtgactaga aggaaaaatc ggcagatcaa tttgggaggc catggtgaaa ctgtaattcc ggtcccagtg ctctgtctca cagcctgccc agcagccaaa agaaccaatt aatcaatgat atctgaacag tttcctggtg tttcatacaa aaacctgata gaaaattata agtttttttc gcctcagtga aaacatgata aactttcatt caaccattat atcttaaaaa aatcgtgaga gaaggtcaat agtaaaaaag atttaggata aggaaaagag atatgtaaaa ttatgtttgt tgtttgcagt aaggaacaca ctttaaaaag attacactga aatgatttct ctaatctgtc tcaataaata gtgcttaacc aaatgttgag ataggaccta agttgcagag taactttagg aaatacctgt taagtgtata tgctttcata ctgttnctgt tgacattaac ggacttaaga atggcaatta gctaatttcc aaaacactat cccttctgat ttcaataaat cctcatagtc tcctttacaa tatgtgtgtg gattaaaaga taaagtaaca ctcaatttct gtggccctgc taaatgttca tgacgtgagt aaatccttgt ggcctgtgca ccatccctca aaatgttatc cgaggtgggt ccccatctct agctactcgg agctaagact gaaaaagaaa actcgtgggg agttgattta ctttctggaa ttcctggcaa cactggacag ctttctgtct aagataaaaa atgatataca aataaaatat caaaaataaa attaaaggtt aaaattaatc gaaaacaagg ataactgtac tgaaataatc ctttatgaaa atagagaaat aaaaaataca tttatntnna tgcattcctg ggtaaagnaa agcactaaca atatatagca acataacacc aaattaaagt tactggcatg tataatattt tttgccccag ctggtattga accttcctga ttacataaac tatcccttaa tgatatataa taataccaag cacattattt gaaagtatga aattgtaaca gattgcatat atgaaatacc tcatttcaaa aaattaaaaa tcttcacatc tattaaaact actccacatt catgtagtgt tttaattaga aatgaaaatt ttttataata tcaaataaat tattgattta cttttgccat aaggtaacta gcagagtgtg ccagaatacc gaccagaact aagtgaagag gagggaaagg tctggccggg ggatcacttg actaaacata gaggctgagg gatcatgcca aaaaaaggtt ttggagctca agggggaccc aaatatgtac aatacaaact ctttgctcca acttttaaag ctgtacagaa agtaaaacta attacccaat taaatggtta aaatcctttg accaagactc agcaagttat ctttataact ttattttatt aatattagaa caatagcttt tagatagata nnaaacttat gatatgaaga aaaaatcata atatcaaagt aagattattg atataatagt gattaataaa gatttgagac tgaaagtaat caataccaac gggaggatgc tgattcctgc attgaacact tctggaggga tattcctgca aatgcctatt ttaacaactt acttgacaga attgttttta gtgtggatga tgagaaacnt tttaaaatat gcaactagaa taaattaact gcattttccc agtctagaag cttctcagag agttaatctt cttgttcata ttgttatttg gtgttaatat tttgcataag gtggcctaaa aaaatcattt ctaccaaaca caaaaaatta aggaaaattc aggtgggccc caaggcagtg tgcagttttc aggagtgtgt gcagaagaga ggccccacat atgggatcat cattctggac catggtggct cacgcctgta aggtcaggag ttcgagacca caaaaattag ccaggcatgg caggagaatc acttgaacct cttaaaatat ttgaggccaa gctgccctga ccaggatgga aacatcagga cttgacccct atcccagcac acctggccaa tggcatgtgc gggaggcaga cagagtaaga ttgtgtaaga tctgagaaac gttccctgga atctagagac aagaaacctg cccatgtggg tttcagaaat atcttagcca cagtaactgt tatcatctag atatttcatt gaaagtcttt acgctgaaga ttgtatttag tccaaaataa tttctataaa agtgggtaca gggtagaaat aagataggat tcaaataaat gttctatttt catgtaaatt agggaaaaaa caaaacgcat catttaatta aataagtttg ctgaagggca ttaaaacaca tatgaaagcc aagtagcaat ttagtagaga taaataaaac tcagaaaaca acagttcacg tttccagagt ttatacataa ttagaaatag aattgcaagc agaatataca cgtgtttngt aaggagggaa gagaagtgat ttcatttctt ggaagaaaat tattcctatc cagtagtcac agctgcaatc atgctgtctt gcatgtgaga ttattaatat ggtatgaaat ttttaacttt ctgcactcua gcictggg-gci atcatgttat ctgtactgaa aaatcccatt acgtccagaa aaatttctaa gatgacccag cctaaatgca ggtcttgaaa gccaaattga tggatgagat ccatcaccaa aaggcctgtg gtgtgctaat tcttatttta tgaatgataa agctagcata taggccaatt tcacataaaa aaaatccaac atgttagcat acatcaggag cttttgtgtt aaggagaata ggaagaaaag atttcaacca ctatatatga ggattcctat tattaccatt aatgcattca tatgattaac tccttatgaa attgtgcaca ttaatagaag aatctagtaa tcttcccact ggaactaact gagagagaga gagagaaaga taaagaacgt aaaatccagt ctagtattaa tcttcttaaa aagttgtaga catatgtgaa tagaaaccag actgttggct tcctcaatac caacaactcc gtcaaggtta cggacagtaa catataaaca gatgagcaac aagataactt tcctgtattg cactttaata ttttctaaat tttggaaatc tatttataga ttagtgcagc ctttttattg aaataccacc catcaatgaa atgggaaatt atgcagctga tatccacgat tgttagtgac tacccaaatc tttatcattc tgtctttaga attttttttc catagaactg tatgtaatat ggacaatgac atctttaaaa catctcatgt aaaaacatgt agaagcagat gatcaaggaa gggcaaagga gttcatncca attaccttgt tatgtttttt tctgcagaaa tggtagattt atctgcagtg cctttaatgt cncatgttcc atatatgagt aagagaagca aggtctgcta ggacatatta tgactaatcc ctattgcaaa ttttaattcc ggacatgtaa aggaaataac gaatagtaaa ttcatagaat actaatattg atcattaatt cacacataga tcaattgctt gcataaagga ataaatgata - 150 - 008252 taaattttag tttaattagg ttaataatat gtaagttgtt attttataat atgccatgta 97860 cattaaataa attttatctt aaaaaataca acatacaaaa aaattcagcc ttttgtgtac 97920 atagaaagtc tttaaataat aatggtttta aaataatatc aaatatagat aataaagaaa 97980 taatgccact cacaaacatt caaaatcata acatgttttt ataattgttt aagattgctt 98040 tggtctactg tgttctagtt ggtatgtttc aaatgcattc ttaaagtaaa atgtcattct 98100 tgaccttatg gggcacctta gtgtatatga tagtggcaaa aattcataca ctgacaaata 98160 tgtgaaataa tttagaataa attatagttc tgaatttcaa tttcaatgtt tctttgtttc 98220 tctaggtggc ttttacagta ctttttgatt tggaagcact tctgaagata tggtgtttgg 98280 gatttactgg atatattagc tcatctctcc acaaattcga actactactc gtaattggaa 98340 ctactcttca tgtataccca gatctttatc attcacaatt cacgtacttt caggtaataa 98400 aaataatgcc tatttgtctt tagaaattta tttatttcca gagtaaatac ctgtcacatt 7o4g. attttaacaa ccacatagaa ttatatataa tatttatata taataactat tatggtaaat 98520 atgaacttga gaaaggcaca atgacatctt acaaattaga aaagtgcatt cacaaattgt 98580 aacaatcgct ttttaaaact cctgaaaatt cacagaaaca atgtagatgt tttacttact 98640 tnttcctaaa ttagcagtat atagatacac atatttttat atatggataa catatatcca 98700 tacatgcgta tnnaantttt aaactacata agatgttggn ncaaacttta aagactagga 98760 aacattaatt taatcccaag tgtttctctt ccttgtcgat aaggctaagt gtttgctcaa 98820 caacggttta taatgaaaat attaaaggtt tataaaactt acctttacag attgtaatta 98880 ctcatacagt attatttata aatgcctgtt acttcatgat ggcacttata attcttattt 98940 aacttgtaat aattggatat ctttttttaa atgtgatata ttggatcaca ttgctttcta 99000 attatgcaat aaacagtaat tattagttac attctttgtt attattgaat atagcaacac 99060 gctcactgtc ctggtaaata cttagtcaaa atgataaaat atgtctacat ggtattctag 99120 tcaaaaacac ctcaaatgta gtcattttcc tgattcaaac ttggtcattt attttttatt 99180 ttattttatt ttattttata ttattttatt ttttatttta ttttatttta ttattattat 99240 actttaagtt ttagggtaca tgtgcacaat gtgcaggtta gttacatatg tatacatgtg 99300 ccatgctggt gtgctgcacc cactaactcg tcatctagca ttaggtatat ctcctaaagc 99360 tatccctgcc ccctcccccc accccacaac aggccccaga gtgtgatgtt ccccttcctg 99420 tgtccatgtg ttctcattgt tcaattccca cctatgagtg agaatatgcg gtgtttggtt 99480 ttttgttctt gcgatagttt actgagaatg atgatttcca atttcatcca cgtcccctac 99540 aaaggacatg aactcatcat tttttatggc tgcatagtat tccatggtgt atatgcgcca 99600 cattttctta atccagtcta tcattgttgg acatttgggt tggttccaag tctttcctat 99660 f* rrt" /та а ста /"+• глг*г*гтг*л з+- a a a г* а *л а *-"п+- /ч+* rrra (-/Tt/rr* rth t-t-afarf-anp at*rra+"h+'at*a Q Q ~7 9 П - WWW^w -.-WWW. W.W.W. WW. WW _y w WWwV.y WW W W w WWWwW.WW-.WW -.www. WW WW, w_> --.WW gtcctttggg tatataccca gtaatgggat ggctgggtca aatggtattt ctagttctag 99780 atccctgagg aatcgccaca ctgacttcca caagagttga actagtttac agtcccacca 99840 acagtgtaaa agtgttccta tttctccaca tcctctccag cacctgttgt ttcctgactt 99900 tttaatgatt gccattctaa ctggtgtgag atggtatctc attgtggttt tcatttgcgt 99960 ttctctgatg gccagtgatg gtgagcattt ttccatgtgt tttttggctg cataaatgtc 100020 ttcttttgag aagtgtctgt tcatgtcctt cgcatttata taatttatca tggaaacact 100080 agaatgtgca ttgattgttt tctaaataga tattgttaaa tatttgggaa ataaaatgac 100140 agtagaattt taaagaagta attttgagaa acaggactct aaccccttag gaaaaagaac 100200 atgcattgca actttattga ttttatttat tagtaatacc aaaatattat tttcagaaca 100260 atattcactt taactgccag atcacaggta actagacact tttagtacct gggggaaagg 100320 acatgcaaat ataattcaca aaaagtaaaa agcaaaaccc cctacaaacc caggcaaaaa 100380 gctgagccct tgtaatcaca gctaagcata ttgaaaccac agagggtttt catagttcaa 100440 gcgctcactt ccctcagcat ccgtggatgt aaatctctcc tcatktgttg agttatatcc 100500 tgaagcttrg ctatcataag tgacatggct ggggcaagag atatacatgt ttaaaactct 100560 tgatacaaaa tgtgctttcc aaacattcct gtgattataa gctcttgtgc actgctctct 100620 agattcacaa agttatcatg gccattgagt acctacctta gggcatcatt gcacataatt 100680 atagagaata tcctttatat caaaacaaca gcataccttt tttacaaagc tattaaagta 100740 gccaaattaa gaaaattggt gttgcctatt tcagcaaagg tacagtataa aatatattca 100800 gggccttaaa attgctatag tctctgacat gccatttttc agtaaaaagt tagaaaataa 100860 catttgtgtg caaaatattt attatagaat taatgagaga aaaaagaaaa ctgtttaaat 100920 gacaaacatt tgaggaatag cttaataagt cataacgcga atttaaccat aaaattataa 100980 ttgtgatgag ttttttataa cttagaaata tgactgtttt atttaaacga taaacagcat 101040 aaattatatt atatacataa ctacatatat gtatgtatat atttattata tacatatgta 101100 aaatacctat aaaatggaaa aaaatgacta gaaagaaacc aatttattac aaattattaa 101160 tcatctctag tttcctactc ttgtcatcat gtctgatttt cctactagtc tgttttttta 101220 caaaatatct aatattagcc tgcaaggctg taaccatttt atagatagac agatgataga 101280 tagatagata ggtagatgat agatagatag atagtagata gatagatagt agatagatag 101340 tcaaatcaca gtgaagtttg caatttcnag ttnngncntt gattttggat tattctgtaa 101400 nnngtnctgt ttaatttatg ttaatgatta ggtgattttc atttgtatct ctctatacat 101460 ggccatgctt ctctttttct aatgtattaa taatgaataa aaatgatctc agttaatttg 101520 tggctagaaa gtaaatctct tgagaaactt aggtgtcaca agggtttttg caaatttgca 1015B0 - 151 - 008252 tgtggagaat ccctattggt tctttcctta ggcttccttt ttcaagtgat gagcagtgga agttcacaaa gaagtgtgca tcatagaagc aaactgtaag ctgaggtggg tgcaaccaat gttcttacat gggtcgggca aggactgtat ctagcagatg cccagcactt ggctaacatg tgggcgcctg aggtggagct agactccgtc catattctta agagaaggac gcttatcttt tctttatagg ttagactcac ctgatcaaat cagttaggta tttccagatt aaagctgttt tgtatgttaa ccactgtacc cctcacactt gacaatttca aaaatattag atttctgagc caaagcagag gaagtacaca gggtaaaaag cctctctgaa gaggtctgag tgcatgattg gcagggcagc atcggtatca tcttgctgag gtgtaaattg atggagctaa tgggaggccg gtgaaacccc tagtcccagc tgcagtgagc tcacaaaaaa actatgcaat tcagaaatgt gagagtatgg gtagatagag tgacagggat caccaatgtg ctgaattatt attattctgg gcttgttctt gaattacagc agacagagcg ctcatttcaa tatttctcac ttatctgtta ttctactatg aggattccca aaggaatgtg catagagatt gaggtgaggt gggaagcatt a v_y Lyt i.aa atggcctgtg tcctgatgat tgcattgtct aggtctcaag atcttcttcc aggcgggtgg gtctctacta tacttgggag tgagattgcg aaaaaaaaaa ttctacagca gaccaggaaa tttacttcat tgaaaaacct ttttcctttc gaagtggtat tatgtccgaa aaattaatac tgtaaagatt aggatggccg gctctttgta tcaacctttc atttaaatga gactaactac tgtcagccac ttttatgaag aagtacaagt gtgtagtagg ttgagcagag ccaggtggag ggaatagcac ggaaatcatc attgactggt catttgtttc aaatctgttg cggctgggcg atcacgaggt aaaatacaaa gctgaggcag ccactgctct attcttccca tctcactaca atgtgtttca gttgtatctg gctgatccct taggaatgca taaatagttc aatagacaaa gaatgttaat atctgagagt ggtgcagtgg taaaggccta tttactgtgt agtaatgttt tgaagttatt tgttccagat ctgaaattct actggaaatg ggctatttta actgaatgaa agaatcacac aaaggccagc aggtcttgca gcgtcttgtg tcataacaag acttgcccaa cagtggctca caggagatca aaattagcca gagaatggtg ccagcctggg agaacatgaa aatcaacaga gagacaaaat taggaacaca gaataaatga ggtttaaccc catttttttc tctgataaaa actttaatag aatatgtcct ctcacacctg cattggtctt tggcaacaag agtgcaaagt catgtattta actgaagatg aaaggcaaac tcttaaggaa gattagttgt gtgaaggagc tgagcatgtc gtggctgggc aaacgctgtg aatttcctgt ccctaagata gaccgcatac cacctgtaat agactatccc ggcgtggtgg tgaacccggg ccacagaggg tttcagagtt gtgaccttta gttctgaagt aaatcaaaaa aaaggcacaa tttgaaaatg aagctgttta atgtgaaatc ttcttatgat gatttgggaa taatcccagc catggtgaaa gcctgtagtc gagcttgcag ccgtcacaaa ccatgtggta aattagcatg taaaccatat gttaaatcaa atggtgtcag cattcttaag gtttgtgtga aacatttctt actctggcca ctggggtagg ggaaagccaa aaaatgaatt tcaatctctt atgccaagaa tggtgtgtta gtgacatctt ^ \_# ". ^ у \-> -х у ccccgtctct ccagctactc tgagccgaga aaaaaaaaaa tgatgcttaa aaggaaacat catttgcatt agtaaatgca gggctcagtg ctggttttct ctcttacagc attaaaggca gatctggatc aatgatttgc aaagaaacaa ttctgtttct attgacatca tgcaattttc tactttaatt agacttaatg ftt-t-ai-.f 9. yV-yyaUWC-vy QVJVJLwQ^yoy ULVJ^U^ULLU actaaaatta caaaaaaatt agctgggtgt gggaggctga ggcaggagaa tggtgtgaac tcgctccact acactccagc ctgggcgaca gaaaagaaaa gaattacagc aggaaaaatt tttgaacaac ataatttctt acatatgttt gctttagtca ttttttttca attggtttac ctgcagacaa taggaataag actgaccact gtaaacgtgg tttcaggtat ggcaaaattc tatgtcttca cctacctttc ttctgttggt ctatgtgaag gcaaagatgg ctcccaggag agtacccgtt ggggattaaa aaaaaaaaaa tccagcaaaa gccctgagga tgactcagat atgacgggtg gggagtcctt cttgcacctt taaaggaaaa ccaggcagag ttgaagaagc atgttatttt agacatgtaa aaattaaaat tactgtgttc taaatacact attcatcttt aaaaatcttg gatgatcatt gtataaacta ctatgttaat atctctgaat ttgggagctg ggcagcattt ttttatttct tagagctaca aaaagttgta tttttcacac tatatacttc r-faafr-lfaf a aaf af> /rf ^r < +-+-а а а а n ggtggtgggc ccgggaggtg gagtgagact ggtagctgca aaaaattgca ttacatgctg gatacttcct agaggccgat gtgtcggtgt ctctgtaaga aaagaaatag gactgtgaat tgtactaagg cagaggcact ctaattttta aacagacaac tatttcattt ttataatcag tacaaaattg agcttttaat t^tcttt^t" agtttggcac ttaattactt tatttagatg attttgagaa caaacctaga ggctacaaac gtaagtattt catcctatac taaatatata atataacaaa ttatactcgc atagtaatca attttttttt tgtgtgtgtg atgcattgtt tgtatagccc ctgtacagtg gtatatctaa gtggcccata cacacacaca tttgtaatag ataaaggttt gcaaatgtga tgtgtataca aggcaatttt actatatacc tatgactaca acatagctaa cttgactcaa tatacatgcg tagccattga gttaaacaaa gtatgcaaaa cagtcatgca gttgtcgtgt taggctatat ttgaatactg atatagaaaa aagttatgcg tgtatatata atcctatttg tgagtgaata catctaaata tcatttaaca gatcatagag ggtatggcct taggcaattg ggtaagtctt atgcatgact gagagagaat tccaacacaa aatttgtcaa tatctgtata gcagagatac tttacttaca attgctccta tcacacaaag ataggaccac aaaatatatg agttgagtac - 152 - 008252 gatccaaaaa tacaaaactt acataagaca gaagtttatt tatgcaggga aagcaggtgg ctgtgccccc catggtcacc gacctgtcgt attctatgca agccttctat ttctggtcca tccatttcca ggcaagagaa aggtatttta aaaaggattc caccaaaact tgtacaaatc atttcctctc acattccgta gagacagcta gctgcaaggg agactggaat gtgtaggcta ctaggttaga attcggaaag ttctgttttt taaaggagga ctgataatct caataagaag taaagattct tggcaaaagt ttgtgatttt ctaataatac ctaggaaaat attttatgag aggttgagta tctgtatccc aaatgcttgg gaacagaagt tcagattttg gaatatttgt atgtatgtgc ataatgagat gtctaaacac aaaaattgtt tatgttttat ttgtatgcaa tattcttaat agttttttta catcataaag caaaggtgtc aggtgtagaa aaaaacttta aatttttgag cattttatgt attaaaggtt tccttgcata tttgaacgtt gtactagtga atatcatttt gtataatcat tcatttggga tttattgagc tagaccaatg tattcaaata gagaggtatg attttcataa atatttaaga ttcagtgtaa ataaatgatc tgataggttc tccgagtagt tcggctgatt tacaagatat ttggtcctgg aaaaaagctt ttgattgtta tgtcagcaat tagtttgcag tttactacgt ttccgagggt aagagtttta attttagtaa taatgattaa catcaaagta ctatttttga taacttggga taattagatt tgcaggtgtt tgagaccatt ttcagcattg aaataatcta atagagttag atgctgaaac ttaaaataca gcagaatttt aaaataaagc atttttaatg ttttataatt tatatattgt tgattttcaa taaaataaaa ttcataggct tctctgcagc agttggtaag cagcctgtcg ggttggacag aaggggctac agagaaagtt ggtctgaacc tagccccagg aagagaaaat ttgcttctct acagtgaata actatttgtt taacttaatg cagtccactg caaccacttg ggatactggg ttccatttcc taactaatac gttttggctt tcgatgtttg atcctgggaa tgggacccaa catgtaagag gtacagctct taaccagaag gaaacataaa aaaatacaat gaccagacaa atagtaaggc ctattatatg gtgtattagt cgatgaaact gggtaattta taaagaaaaa ctggagaggc ctcacagtca tggtggaagg agagagaatg agagccaagc taaaggggga gacttattca ctaccacaag agcagcaggg acacctggtc ccttccacaa cacatgggaa gggtggggac acagccaaac catatcatat tatattttta aaagttaagc tttgacagca aaacacctca aatacattcc ctgaaggtag tgcatgaaac aaagtgtgta tacattgaac tttttcactt atggcctcat atcagcactc tttaaattag gaatgttcaa cctgtattga agaaaaatat taattttatt tcaacaatac ataatatgtt aaaacactta gttcatccat gttattctat tgttaatttt agtcactaga agcactataa aataatattt tagtatgaat atatgtgtaa ataaatgttt ctctctccct aagatttcac ctgcattaga agactttgtg gggagtttgg ttgtatttac tgccagcctc ttgtygaaga actggacaga aagttaaatt cattgttctt agtcattcag aagtattatc aaattgtctc ttctgcaggg tatcaattct aaaattctgg gggttcttag cacctataaa gtgttcaaga gagttaacat agaatctaga aggaagcttt taaatgtctc tcagctttct -"*"."¦¦".""-• +• /-" /-Ч +- au^^Q-v^uaL aaov^Baaavji. gaccttgaaa tttaactcta tgtggactta ataatgtata tatttttacc ctaagttgaa ccattcttac actgatgata aagacatact gaggtttaat ggactcacag ttccacgtgg tgaaaggcac atcttacatg gtggcaggca aacctcttat aaaaccatca gatctcgtga gggaaaccgc ccccatgatt caattatctc ttatgggagc tacaattcaa gatgacattt gggaagaata atattggggt aaaaacaaca aaagggtctg tacttctaat ggatagctta atgttctgct aaaatgcagt atttcacttt ttaatattta tcagattata ctgaatgcta ttgcattttt tatgttagtc cttcagattt +- 4- -4 - gctttatttt agggattttt ttaaaaatcc tatgctaaat ttattatttt agctaccagt gttggaattc atccttttat aatgcagatt agtgccttta aagcatctgg tcatttaggt cttcattcat gtgccgccaa ggatttgtcg aatcatgggg acataagtcc tcaagaaatt ctcagttgag gggaggaacc agtaggtaag cacagggcac cacaggcctt gatgaaggca gacaccaggt gctctgggga cacagaggca aggccgattc ccaggcatgg aaaagtccag aaaggcttat gggaacaggt gactccaatg gctggatgta gccaaaggaa gtgcaaggga acggattccc atcacaggaa cagcatgtac taaatgtctg aggtgagcaa atccttcttc cagaacttag caagaaccat caatggtaag aggaggacaa aaggccaagg aactctgaag tcggttgaaa gtgtgatcgt atctgtcatg ccaaactcct atcaactcta gtagggaagg caccaggttc aagaggctga agaagagacc cagagccagc agatgagaca tggggtttca ttgagggctt acatacaggg gagggagtcc cctgcttgca aaatgcatgc aatttacata cctccacatg gaaatcttaa ttcaccccag agtgttccac gggaggagat ggggactcag gggttggcca ctcctagatt ccctagctca gggtcattct cggggtatgc ttgagttatt agtgctgatg agagagggca agtagctagt gatggcaaag gttcctgcag aggtgggaga tggccttaga caggtgttga gctgacaggt gctgaaagtt acaatcaaga ctttgttccc gtaggcaccg ctctccagct ctccagcgtc aatgctggct ggctggtcag gagaacaaga gcatttttac ttagcacact cccccagcaa acttggggcc tcaattccct gtgtatagcc atgttcctca tcaacaagga gtcagtctct gaacacacat tcaggtgtgt ctggcaagaa gctttcaggt gcacttacat acgatgttta gaggttgcag atgggtgaga taaggaggat aggcagtatc cacagcaaga ggacaaagat gtcgaggtga tcctcaggac agcaagccaa tcactgcaac agtacaggca ggaggcaaag cggaggtgtt ggttaagggt caggtggatc - 153 - 008252 agcaggcatg ggggagggag ggctcttttc cttgctaagc agtcctgaac aaaaggttcc 109200 agccattcct gcacctaggg aggggaagca taaggaagga agggatagga gaagaaagga 109260 acggagccag catggaggaa agtgcctcat ctgggccctg atcaggaggc ctctgcacag 109320 aggcacctgg tctgggactg cagctatggc catgatccca gctggcctgg cagggcagag 109380 ctgagtcata aattgcaagt ccctccagaa atggcagtgc cctggtcatg caccatgtct 109440 agggtgggga tggcagtggt ccccgggagg tctcccaggc aaggccttgc catcaggtct 109500 gaatggatca cacataggat tttggccagg agcaggactc ctggaacgga ggctggaggg 109560 aggcaagcca aaatgatagc cacatcatta ggcccacctt ctctgggtct gccccttcac 109620 tgggtaaata aaaagatcag ctggccatct tttccatggg aacatgggga tggttattgt 109680 aagagttaaa gaggtaagaa acacaaaaag cagctcaaca gtaaaagaca gatttatttt 109740 ggagaataaa ctttagaggg gcttctggcc aattttgctc aggagcattc tctcttacag 109800 actaagggta tttaagggtg taggaagtgg ggagcttatc gcaggttcgg aatgtttcta 109860 tgtgagggaa agtttattgt ggggttggga ttgggatgtc tggttggagg ggaggctgtc 109920 tcagggttgg cgtgtttctg gtcagagggg gtttatctta gggttggaat gtttctggtg 109980 atgctgacat taggcattag gctgatgttt gggngcttga tttgggtgtt tttttaatca 110040 aagggaactt aaaatggtgg tgtttctcca agacgacgat ctcctgctct gtcaatccag 110100 tccctacagt tatagaaagg aagaggggtg gtatattctt tctggctact tcctgctgag 110160 tggagggtca gagggttctt tggtcttggg ttgattgtgg gagtaatgcc atctgtagat 110220 gtttttaggt aattgtgaga tgtccatgat cctgtcagtt aaaaaatctt tgaaaaaggt 110280 gaattaagcc aggtaagcac attagtccta ggcatattat taggagaggg cccaagaatg 110340 ggatggtcca tgttaggatt ttgtttctaa accaagaatg tatttggttg ttttggtatt 110400 cctttcacgt tttagccctt tcttcaagtt tttcagcagc atctcttact aggcctgatt 1104 60 ggttgagata gaagcaacat tcctcactca gtgagaggca gagaccccct ttttcagcca 110520 ttgtaagatc taatcccagt ctattttgga ggactgctcc aggcaaggag tctaattggt 110580 cttcgactct tataaggctt tgggctatat cttttaataa atcctgtagt tctgttgaaa 110640 gaactttaaa gtatgttaag gagttggcca gtctgcctgc tcccaatcca agcccggagg 110700 ttatacataa ggcagccatt aaaggaatga cttggatgct cctccttttt ctaacatact 110760 ggatgaatgg aacaggtgaa gattgattag gaggaactag tccaacggag ggagaaagat 110820 aaactagggt acaggttctg gtctagttgg tggggagaaa aagatatgtg ttggtaccgc 110880 ataaaaagaa cgagcctttg tcataatgca ggcggagata gggaaagaag ataagcaaat 110940 taaagatggg gtgttttttc tttcctgcgg ttttttactc caggtggaca aggaggaggc 111000 cagggagaca cccactatag tagagatata ggaagagtta ttttttacac atttaatgca 111060 atcggatgtt gcaccattga tattgagcca tggaaaaagt tgggcaccag gggcagcgca 111120 agttaggcag gaggcattgg ttgacggaga ggctgtaaac tagctggttg cagaaatgct 111180 ccatttgttt caggtgatac ttggtagtca acgtggctac tttgatggtc agaggggtgt 111240 ttaacaatga tagtgtgatt gcattggtta gatgttaagg accctacagg gaaatttccc 111300 tccgaggctg aaaagcaaag tgaggcttgt tgtaaaaggg gggtgtgtgt agttatgggc 111360 ccttcaatgg gagggtcttg gtgcttaagg .cattgtagag agttgtaata ggtttgaaat 111420 attttgttgg cccgattggc cctggaagtg aaataatctc tgaacagggt gtcggctctc 111480 tcaaaaaagg aagctccttt tgggagttta taagttaggg ttatatttcc tgttcaaagg 111540 tcatgaagag gtgtaggaag ggctgtgtaa gctgaggaag acagcgataa gcatatccag 111600 cactctggag caaaggaaga gttagctttg cagcaagagg gattgtgtaa ggtttataga 111660 gcattctagt ttgagagcag tgaggagtgg ttttattggt gaggttgata tgattaggga 111720 atttatattg aaagaaacaa tcaaaaagag aaaaaagtta tttgggtagt agtaaagtcc 111780 tggaaagttc cttgaagcca taggtaccat agaacagtca gaaactgatt agcatgtata 111840 atgggagctt tgtaagggta ccactgaagg tctgtagcaa ggttggttag gaagcagtga 111900 aagtttggga gagaaagaga cataagcagc ttatgtggat ttctcttcct tttcctccag 111960 gatttaggtg aggcgaaggg aagtaggtcc tgtggagaca caagagaagg ctgaaggggt 112020 tttagatttg gttgtttgtg tatgtggtga agggaagtct gttttcttga gagaggtata 112080 atgaaaccag ttggggagtc cctggagttt tgctgccatg ggtgtagtaa ggatgatctg 112140 gtaaggtcct ttccatttag gtgtaaggag ggaattgggg ctaggactgg gatcttttat 112200 cagaacccag tctcctggct gtaggaaggg attagaggag ttggtgacaa gttgtggcag 1122 60 gtgtttgtca gcatattccc aaatgaaatg gcagatggta tgtacaagag gggaaatgag 112320 aggggttggc agaggtgggg cttgaccctg aggtggacca aaaggggcaa gtggtctccc 112380 atatgtgagt tcaaaggggc tgagcattaa aggtttatgt gagagcacct gaatttttag 112440 aagggctaaa ggcaaaagtg taaccgagtc tttatgtgtc tggagtgagt acctggtgag 112500 ggtgtttttt agaatgccat tcattttttc aaccttttct gaagattgag gtcgataggg 112560 gatgtgtagc ttccaggtaa tttgtagggc ttgtgaaagt gtttgagtaa tctgagaaat 112620 gaattcaggg ccattatcag attgaaaaga aagaggcacc ccaaacctgg ggatgatttc 112680 tgttattaat ttggaggtga cattagaggc tcgtttgttg gttgtgggaa aagcctcaag 112740 ccattccaaa aaggtatcaa ccagaactaa aagaaatcaa acccttttta ctgggggtat 112800 atgggtaaaa tcaatttgcc aatcctgtcc tggaaggtgt cccctggctt gatggattgg 1128 60 gaaagaagag tgtctagtgt tggaatgggg tgaagctttc tggcaaatag agcattgatg 112920 - 154 - 008252 ggaaatggct tttaactgtt cctttatatc tggggttatg tgtttatggg aatttaagaa 112980 gtgttgtaga ggtgaatggc tagcatggaa gaggttgtga atgtcccgta aaagagttgt 113040 ttttttcagg gtcaggtagg actaatttgt tttgtatgaa tcagtttggg gggtttgaat 113100 tgtgcccctg ccatgattag ttgttgtgtt tggtgttctg gataaaagaa ggggatatgt 113160 tgtatgaagg gaaataagta ttggggaatt gggtgattgg ttgagacatg ttttgcccag 113220 tagtcagcct catggttccc taaagaaatg tggcttttgt ctgatgttct ttgccatgaa 113280 taactgcaac cttttctgga agtagaagtg cctttaatag aggatgaatg agctttccat 113340 taatgatagg ggttcctata actgtgagat agccccgctc cctccaaatt taggcattgg 113400 aatggatgat gttataagca tatttagaat tggtgtatta ttaacttgtg tgttttttgg 1134 60 tagggttagt gctcttatta gggcaactaa ttctgcttgt tgggcggatg tgcccaaagg 113520 caagggggca gcctctatca ctcttctaga tgggagagag tgggtatcat aacgctatcc 113580 ctcaatgatg gcatatcctg cttggagagg agggtttttt gatgcactgc catctataaa 113640 ccagtctggg gctcccttta tgtgagtgga agtaagatgg tgaaacatgg taggaggact 113700 ttcaattaga tcagagcatg agtattggtc aggatctaaa atcggtattg agggtaaaag 113760 agaggcggga ttaggtgggg agcatctatg aagagagata gcgggttgaa ggagggttaa 113820 atgtaaggct tgcatatgag aggatgagat gggggtgagc gcctatggct gaccatatct 113880 tgtagactgt gagaagaaaa tacctgaagg ggtttgtaga acatgagttt ttgtgtctca 113940 gagataatta gagaggctgt ggccaaaatt tttaagaaaa gggaccagat tttataaatg 114000 gggtctagtt gttttgaaaa atatgccact gttgagaagg agtctcccat aagttgggct 114060 aatagtccaa gggcctgatt atgagaactg tgtaaatata gatgaaaagg tcttaggagg 114120 tttggaatgt ctaaagcatg ggcctgcaat aaggcacgtt ttagatgaga aaaagcatga 114180 taaaggtctg gggtgggagt gagtggttgg tcaagatttc cttttgtgtg ttcataaagg 114240 ggtttagcaa taatggcaaa atttgctacc cataatcgga aatatcccac taatccaagg 114300 aaggaaagta agtccctttt cgttttagga aatgggattt gttgaacacc ttgctttcat 114360 gccaacagca tttctcgggt atttggagtt atgatcagtc ctaggtatga aactttcggt 114 420 gaggttaatt gggccttcct tttggacacc aggtacccac attcagccaa aacatttaaa 1144 80 atcttggtgg tgtgttgaat acaatggtca agaaaggggc tgcaaaggag taaactgtcc 114 540 acatattgca gcaaaatcct gggttgtaga ggtagttggg agaggtctaa ttgaagtgac 114 600 tgaaatagtg atggctgtct ctaaaccccc aggggaggac agtccagatg agttgttggg 114660 agtagcctgt gtcagggttg gtccaagtga aagcataaag gttttgagag gagggatgta 114720 gagggatagt aaaaaaggtg tctttggcca ggtgcagggc tcatgcctgt aatcccagca 114780 ctttgagagg ctgaggtggg cggatcatga ggtcaggaga tggagaccat cctggccaac 114840 atggtggaac cctgtctcta ctaaaaacac aaaaattagc tgggtgtggt ggcgtgtgcc 114 900 tgtaatccca gctactcagg aggctgaggc aggagaatca tttgaaccag ggagtcagag 114 960 gttgcagtag attttgctac taaatgtact ataagaaaaa aaaaaaattc aaaaggacct 115020 tcaagtattt tcagaaattt ttaaattttg gagttgtgaa taaggactgt gggcctctat 115080 ttttaaaaaa tgcatttaag agtctgtact tctgtcccta caagtccgat tgtgcatcaa 115140 tgccccattg ctttttagct atgtgatact .gggcaaatca ttctttacat ttttattctt 115200 cacaaccaag atgtaagccg aaatgtgttt ttttttcttc tgcattttgt gtattaaata 115260 atatacaaag atgacacaac tgggcttaga aagtatttct taaaagacta gattacagcc 115320 tatgtaatat atttgaaatt gataatatta tctgtataaa tggcaaggct gctttttaga 115380 tccattcctg acttagcccc tctatacttg ctctattttc cagctataat agtaatcaag 115440 tagtattaca ttttcagatc tccaattact caattgcacc attttgcctg attattttga 115500 ccatcataat gaaagcagct cggcctctcc caggccccct aatcagttat tcagcagctg 115560 acaatggtga cagcctgaac gtgtgaagtg actagaagga gtatattttt cctttcacat 115620 aaatgttact aaaggtgaaa tccatgaaga gtaataaaat tttacaagga aatgttaaaa 115680 aatattacaa ttacaaaagg ggtcattatc atccctaatc aaataaagca tcaccaattg 115740 gtcttacttg ccagatagaa aatcattcag gagagatgat taaaaacata ttttactcaa 115800 ggctagattt gtcgtcagac gcacatccct gggtttcttt tccctccctt ctctagacat 1158 60 tatccagtgg ttgagtgctt aaggtaggtt ggttaattac agtattagcc aaggcacagg 115920 acatgaccag aacaaatttt ctgcaagaaa aatcacatga tgtatcgcaa agtgaaatac 115980 agactttaaa aatcagcaat tgtctatgtt atcttgactt gggtccaggc cgtcttggag 116040 gctgcctccc agcaatgttt atagggctgc agaatggtgt gggaaaataa agtgtggaag 116100 cacatgggag gcttggggag ggcactcacc ctgaatctgc ttcagcacta tttaccaggt 116160 gtgtagtctt aggtgacact taatttgtct gagcttcatt catttttaaa atggtaatga 116220 tgtctatcaa acaggcatga taactaagag ccctacttac acatagtaag actttgaagc 116280 ttggagagct tgcataaccc acacaaagtt acacagctgg acagctggat ttgaagttgt 116340 ctactgctcc taccaactat tatccattag gtcacttcta gccaagcaaa cacactagaa 116400 agtgttcagt taggaaatac ttattgagtg ccaagaatat aaatattttg tcaatctgta 1164 60 cctggaagga aaggcctctg gaatagttca tcacaaactt cccagaggac ttcaggaaat 116520 caagaaaggt atagaggtat taacattttg gggcacagcc tgtgagtgtt aggaatgatc 116580 atgtcaggat cccaaaccag cacctgatag tctctgtgcc acagcagaaa cagcatttgt 116640 ctaagaagat cccacacatg agacaggtga aaggatgttc caagtgacag cttgtgaaga 116700 - 155 - 008252 ggtgtttgaa atactgggtg tcctcattac gttgcaaaca ggaaatttat tttcactgaa tgatatccca taaaatgaag aaagaacaga gttctcacat attgctcccg aaaccacaag ggctggagca ttcatgatga ctgagaaacc ttacaattga tggccatggg ccagaagccg gctaacacct ttttaagcca ccttctccat gatgacatct aaccttttct tgagtaacag agtagatgtt agctctgtct ggtagaaaga cccaagatgc tagatcagtt taccattgac gagagcatgt П /"• г^> Гж/-"-Ь -4 agagatggag tagaatttaa caaacagcta aacagttcct attctggaac ctttctgtat cagcttcagt aaattactga tattccttct aatggggata ttttagtatg ttgctctatt aacaaatgct gactagtcgt ctccttaatt ttcaaggttt tttttgttct atatgaataa tagacctgaa atcacttttc atatggacat ctcattctgt gttacctaaa actacatatt ttcattttcc gtccataccc ctgtcattcc tgtcatgtgt aagggtcttc tgcagcacac taccctagaa tggagaatga aggctcagca cctccatcat ctttggctgc tccataaaat attctcttaa actcccagaa taatcagggt cacaaatatg tgctataaag gttctgcagg aaacttacaa ggaggaaaag ctcactcagt acccctataa acatgagatt actggaatgg taagatacaa taattttaga ccaggtttgt gggttgtctt agaggaatat atcatccctt ggtcattcct gtgaagacaa ccaaagctgc aagctggtgt ccaaataggt atggattcct attacacatg gcattcctta ag uagagtica aagggatgct taaaacaact cactgctaaa gttttgttgg agcctgtaca caaaaacaaa aagattttac aataaatgca gatttcccac gtttttattt atttatttga gagttttcac ataaaacaca gcagttagtt ttttaaatta attttaatat gtctttcatg taataaaaca gtccctgggt tgattttgat caactgaaaa actatttctc tgagtacatg atattgcata tgtgcagtag tgcatcatcc tgtgaatagg gggcctgtcc tgttggcttg caacgtggca cttaaagtat gcaacaatgc tgccggattc tccccttacc aagtaacaga tcaagagatc ttgttgtttg cctcaaaata ggaccctaat cataagagga aaatacctga ttgtatagga tcacagcaga aaagagagag gtcatgagaa tctaatcact tgggtgggga catctacaag gggaagattt cttctggcct ggagcttttt cctatgttag gtgggtcctc tccttccagc agaattaaaa ctacttgggt tcctatattc ttaattcctt aggtacctgc gctctccctg ggtcttactg cacctccaag gaaaggaaga cgaattctac ttgtcatctc tagaaataac gatgaatata ggtcctaaga ttgcatgttt . tcagtttttc gtaaacattc taaaataata ctaagtaata tataggtttt ttataaaata taattaaaaa gtattaaagg tttgtcagat ttaaaactat tatttttaat tactcctctg gcctcttcct gtgtgttttg tcatatatct ctgctaattg gagctgtatt tataatacat gcagattcta cctacccttc ttgctaagca taatcagagg cattaggaga catgtataca aataaattaa tgaaccaaca cttttcccag ctatccaatc aaacttgatt aggtagcttt aattgtcagt taactatatt ccagtatgcc agatgatata gactgggtaa agcatggcag aggcaaaggg gaggtgctac cagcactgag cccatcaggc cacagatcta ccaaggaatg ttttccctag ccatacctgt taccacagcc caaaatggat tgttcccagg aaacttgctg ttactggctt cagacccaaa cccatgcaaa tcaccagaaa aaagtagcta tttttatggt cctttgctag agcacttaac caaacatctg tattgttcat agaagcgtct ccagttgact caggaagtca gttcccccaa tgaaatacag tgatgctttt gacccacagt ttacaaagaa catctgcttg gaagcaggca acacttttaa aagatggtgc cccacctcca aactgtatga tcaaagaatg agctgtcaga gagacaatac ctagcaaact gtgacatttc cctcctcctt tcatctctca tggttaatca acctggacaa caaggtttgt catctggacc gcatagggct actggtcagt tctacaggta actgtgcagt tcatccttac tcctccagga gcataattcc ttaagaataa ggactggatc aaatgtatgt gtaatcaagt atgaaaaggg atattagcct gagaggttta gctcctgggg agtcttatat gcaacctgat tcaaccattc atattgggga cacggggata ctggcaaaca gcacagccct atttctgttg catacagtga caggcttcct aggaccagga ttggcaggat catgggaagg gatttctctc aaatagacgt ccagatcctt ggacacagag ttctatcaca ccttgaggtt gggatctgta 40V> Ц-wb tcgatggaaa aagccaataa aagatgaaat ttgtatgagt cagattgcca .ttccataatt ctccatttat actgtccatc aacagacctc tattatcctc cttaattttc tgtccaaaac ttaactttcc aattaccaaa gtagtgagag tagttcaatt tttatcatga tggccatact gatctctgtc ttaataaatt tttgtgtaat cttttattgc ttatttattg tctttataca agctgtcccc gatactggta tttaatgttg tactggcctt tatacctaat tatgtaacaa taaataaatt aacaaagaat ctttctgttc cctgatgtaa acaatttaga taagcaagaa gcctaacaaa tatagaaagc tttaaccagc atatgtttag tataacacaa attctctaaa agttattgca tttcctcttg aaatttgaag tgtgtgacat aatattgctg aatatttgat taacaaacac ctctcactct tctttatact ttaaacttgc tcaatattat tcattactgt gacagaacac catgatatct ggatttgtga agtcagtcat ttcttagatt gtaaatgatg acctgcacgt aataaatagg gctatattga tagtttggac ggcaatatgc tgaactattc caaccacagt agctcataag aactgcctct tctgtatctt cggttgtata agtacttaaa tgttaattac cgatagaagc ccaccaaaat aaacaatttg ttttaataga aaatctttaa gtgcaaaaag taaattgcaa cccatggaga tttaatgcat cttatatggt gtttatgaga ataaaaagca aatttatttt ccgcacgagt atatgatggg aaatagagat taaagcaaga agttaacggg tgtgcacatg aaaaaaaaag - 156 - 008252 taagcctcca gcaggactct tagaagaaga ttttgagact aaaactgtga caatagaaaa actgctctga atgtatctgg attgataaac tgagagtatt tcjaaacactt acaattttat catgccccct gcccaagtca tggatctgga gtatctcagc cattaaaaac ttcaagctac cctttaaaag tttccataca ctccatggac atgtgcataa ccattaactc accccatgac tcaattccca gctcagaatg ttttatggct atcattgttg aacatacgtg agtaatggga actgtcttcc atttctctat ctggtgtgag atgagcattt tcatatcctt tttgtagatt ttctgtaggt cttaattaga catgaagtcc ttttatggtt aggtgtaagg ccatttatta cagatggttg atctctgttt gaagtcaggt gcgggctctt agtcattggt cattttcatg taaaatgagc acagtctttt agctttaccc atacccattt caacattctt cctctttaac tagaacacat tcagttctaa tgtgtgccta gaaggaggtg gtagagattt aaattagaga caatgggaaa tgaagatttt ttttgaggaa tgttgtgaag cagtcctgca ccgcaagctc ctgagctgag agtaacaaat gtaatgcatc acatttttgt gaatagagtt tttcattcca ctctacttta tgctgtgtgg accctaagga cagatgagac agcacagtta gagtgttaag caggtgttgg acattgattt gctgggtctt aagtagtttt ccactcagga itagectgaa ataaacaatt catatgccag gctcccttgt ctctgaggtt tttcaaaggt cagagatcgg ttggtccctg gcattttatc ctgaggttca atagactttt tgttacatat ttaggtatat gtgtgatatt agaacatgca gcttcatcca ccatggtgta ttggttccaa tttatagcag aaatggtatt aactagttta cacctgttgt attgtggttt cttttggctg ttgaaggagt gccctttgtc ctctgatggt ctattttggc ctatatcctg catttaagtt gtttcagctt cctttcccca gtattatttc accatgctgt ctccagcttt atatgaactt cgatggcatt ttcctatcag gcaggccatc gcattgagga tgccacacca taaataaatc agctaggaag tcaagaaccc tcaagtaagc aagacatttg atatcagctc tataaagatt attagatgaa cacaggcatg agggtcttca tttttaattt gaagaaatgt ccaagaccct acaaccctct caaagcccag tattacacca ctgcttcgtt atgtttctaa gtgcacatgt agaataaaca accaatttat acacttggtc ttgatttttg gccacagtac ctgtgtcata catccccttt taaaaccaat tcatgacttt aaattctcta gagtttttaa cagaatttcc ctaataaatt ttaaaaatta gtatacatgt ctcctaatgc ccccaccctg gtgtttggtt tgtccctaca tacatgtgcc gtctttgcta catgatttat tctagttcta cagtcccacc ttcctgactt tgatttgcat cataaatgtc tgtttgattt agatgggtag agtttctttt .tcctgttgcc aatggtattg tttaatccat tctacatatg tttcttgttt tgagggctct tttggttact gttcttttgg taaagtagtt gaatctataa atggataaga ccagtgtttg tattgtgtcc ctgaaaatca atgttcttgg catactgaca aaggaaggac caaaggtgac ggagagtatt atttaatgag tgtctttagg ggcattccat gagggacata gtggttggaa tttttttctt tatttctatg gggcaacagc gataataacc ccagcatctt tgcccacatt tgtggtaatt atattgtaaa gaaataattc tatcctcatt cctaagctat tggtcaggct tggctgctgt catttattga acgtggttcc tgccattgaa aactatatag gtaccttatc gagaaactga ttattttctt tggggttttt tttgagaaga tactttaagt gccatgttgg tatccctccc tgtccaaatg ttctgtcctt aaggacatga acattttctt ttgtgaatag aatcctttgg gatccttgag aacagtgtaa tttaatgatc ttctctgatg ttcttttgag tttcttgtaa attgtaaaaa gctgtgcaga attgcgttgg cctagatttt cttgaattaa gctagccagt ttgtcaggtt gttctattcc gtagccttat cttaggattg ttttccaatt attaccttgg cttcagatgg cctttactgt tctttgaatg tctttcacaa tttttcattc tgaacaaatg tgaaacaact atatgctaca ttaaaattct agaaaaatat ggtgaatata ataagaaaaa tgaccaaagg ctgtgacaca ggagaatagc aggaggtatt catcaagaaa caaatgaagt gatttccacc ctgaccttca tgttacacat tccaaacagt tttttagggt tctcattggt cagcagtata ttattttgct c_g__c_tcat ttccatttgg agttctgcat agaggcacag tcaacctcct agtctcctcc caaagtctag ttcttagctt gatgcttttt ggatgagaaa tctcagatgg cgtgcaggtt atcatttaca aggccctgga cctatcagtg atggtttcca gcatagtatt gacacttggg tgcatgtgtc gggctgggtc acaatgattg atgcctccag atggcatctc tttcatgtgt cgcccacttt ctggatatta tgcctgttca tcccatttgt ttgcccatgc ttaggtctaa aagggatcca aatagggaat tagatgtgtg tggtaccagt agcgtgatgc ttttggttcc agcttgatgg gtattgatcc atgatctagg gcctttgttt tccctcagtc atgatgaaga tatgcaacac tatagcattg aagaaagatc agtaatagca gcacttccaa tgcagagctt gggtcatggc catggagaat ccagacatgg tactttattt agttacactg у ciyciyciyy yet ccaatctatt atatgtgagt ttttttttac ctttgctcaa tctgacacaa cttttcttga ggactattca gaatcatttt agaaacagta tctagggtac tgtgctgcac acctcccccc ttctcattgt gcgatagttt actcatccat aatccagtct tgccacaata gtatataccc gaatcgccac aagtgttcct gccattctaa gccagtgatg aagtgtctgt atttaagttc ttttctctca ggctctttag gtgttttact cttctagggt tttttgtata tttcccagca tgtcaaagat attggtctat agtatagttt tcttggcaat ctgtgaagaa gcagtatggc atagactttt ccgcagcctc tcatagcctc aatatctgat ataattctat tactcactgc catcccatag atagatgtac gggaatttac tgcttccaga atttcaacag ccaaatgagt atggcagtag agggagcttt agaagagcta ttcattgaac - 157 - 008252 aaaattcatt ctttggagat ttcataaaat actctgcaat tatatatata tcattatttc atttttgtgg ttccttttag atcattggtc gtttcctacc tcttattttg catcttacat aatgaaagtc gttttcagtg aagaggacag cggtggaatt acagtctgta aaaacaggat tgcaaagtag gtccttctta actgaggtag cagttttctg ggagattatt atcatattac tcattaacta cattctttct agactaaaat tgaaatctta catgcatctg ccctaagcaa ggaaaagcag ggtctgcagt tcagaattcc aaatgtactt aatatgaacc ctcttgggat tgcaaaggta tcctgtatta caataaataa cggcaataat aacaaattaa cactcatgtt ttgatagaca agagaagcat ttaatggtta aaatggagag acatatatct aaagaagcag ctacaataac ttgctataaa accattctgc caagcttcca gaaacaaaag agcttagagc gccccgtgac acatgggatt atgtaacaaa actaaagccc aatgaagccg ttgttccttc cgctggcctc gcacatctgg tggtcttgct tttattgatg attttatatt tgagtctcat agaaactaac tagtttattg ctttctacta tgactcatct aggatgattc cactgaactg cctaacccta gtagtttttc taatattatt attaatgaaa atgaaaacat ttcatacatt ttgaacaaca atgagactct aataatacat ttagcactgc aatgcatgga cccagcatcc tgttgtatag tttaatcagt agaaaaataa ttacatatac cttgggtata gcctacttct atttccagga gctgagcaca atggaagcta agcaggtggg attgctgctg ggagaaaaag gctaaattcc accaaaaagt aaaccaagtc cagcaaatga ctcactagac aaagaaatgt tatatccagg gatggaaaca tcccagaatc aacatgttta aattattcct ctatatgtga atggatgggt aatataattc tgaggaacag tagaagtgca gaaatacctg aggctgtaca gtcatggcag agagaggagg gagagctcac ctaaacacct tggaaaggac gtgtcacaat tctcatactg cagacccttg tgatatttgg aggagtgaag agttgtttgt ggcttcagga aaagacaaga tttttctgtg ctgaagcttt aatttggctt tcattgtctc taacattagg gtggcataat acataaaatt caatgatcag gaagagtact atttatgatc aggctgttac aggccagatt attttaaagt cccccccaaa gttttttttt gatgaaatga ggccatcttt aattatgact ctttcaggga ttatttgcaa gactgttcca cattccatta ttaaatattt tgccttttag atgatttatg tctcatggct attggtagga taaacaggca cttagcatgg gactggagaa actttaaaaa tttccagaca caagttattt agagaccatt atgtaacacc gtctctaaca gataaagcat gatgggggtg tcctataatc gaaagaacet tgttgcttct aaagataaaa ctgcaaagtt aacattctag gagtaaaatg aacactctct atcttcatct tggccttgtt ttaatcaaaa ctccaaaacc cgtgactaat acagtgatta agaaagacat tgtgtgagag agagagaggt ttcattaagg tgatgagaac ttcgcagaca atttctaaag caccagcgtc gggagcaagc ttcaaacaac agacagcacc cccacctcca accatatcac aagacatatt tgttgggttc tacagttctt agtttctgcc ctgtgatgag gggagttctt agaccttcgt agttgataaa tgtgtcaaat gttatccatg gataccatga aaaaactgta ccttgctcct taaaataata tcctgctgtt ttccttgaat gaacctattt tttcaggcat ctttcacagt gttgagcatg gatatatata aaattaattt gttggaattt tcctgtgtac gtatattaat aaagcatcct ttgcagctca atttaattgt tttaaaaata cctcaagtat tgcctacaaa taaagaatgt tacaaaaaat tcctatcaat aagtaacagg agtagcagtt tcctagcagc ggtgggcact ttccagttta gtaaaaaaaa aaatagctta tatgtatatt gtgtgccaga gaatcaatgt cgtgtgtgag gagggacccc caatacaaaa tttagatgga attttagttt aatccaacag aatgcatatt catttgctgt agtgggatta gggagaactt gccaattcaa ataatgtgtc ttaaaaataa atcatgagga aatatacact actagttcct gttattaatc ccttgaagaa atagatatcc ; acgttataaa acatataagg caaacacgca taattggctc gaggcctcag ggcgagaaca gtgaatttag tgagacagct tcacatttca ctcaaacaac agttgggaat tgacttcaag tgtctagagt ttcctggtct cataaaggtg ggtgggttcg acagttcata dUUd-LLdLL ttaataatca agtattgtct gaccgtgtta tgtgtctgta aaacaggcac attgtcagtc aacttagagg gcctgccatg aagcccatgt aagggtaaat aaagtttaaa cttttgcgac gacaaaccga ggaactaaac cccttcttgg gcagaggact agcatgatgg tgtgcatgca atgggctgca accagatgca tggcttttgc gcaacaggca tgtgagaact gaattctgtt atttcaagtt aaggaaaggc aaagattatt ttattatatt tcaaataaat aaaaatggca ccggaccaag tacaagttct gatcaagtcc ttgcttttta atacacagag tcaaaacgac caccagaaat tgacttaggg agactgagca ggaagtgtag aaggtgaaag cgccaactct tcgttatcat cccagcaggc acacatgcaa gaatgattaa tgtccggaat cggtgagtgt acgtgtccgg ctgcagacct tcctcccgtc gtgaagctgc L> L. ULULddU L. ttgttcaaat taatgagctt tgtggattct ctctactgca tagtcttgcc acccacaatt agattaaagg cttaacaagt catgctgaat ttaatggaat tagtccccaa aatgcaaaag ccagacgcgg cacccaatga cagagtctta ttttttctag aatgtgtgta gggaaggaga ttaaattgtg tagtagctag ttgtgttgtt atcctgcctc ggcaataaag acaaatgtca aagtaaagcc tgaagagcga tatagttgct ctaacagaat tttaaagtaa taaaaggctc ttttctagca tgaggtacca ttttctggat tttacaagag gcacacacac aaaattatgg caacaagaaa gatgaatggg caaagaggtt tgctccttgt ggtttcacat cagatctcct aagccatttg acactgggga catatattca aaacaagtta ttggtcttgc aaagatggtg ttctggtggg tgttacagtt tgtccctcct ggtgagtgtt - 158 - 008252 aaggtggcgc atctggagtt gttcgttcct cctgttggga gttgtttgtc cctcctggtg 128100 ggttcgttgt cttgctggct tcaggagtga cgctgcagac cttcgtggtg agtgttacag 128160 ttcataaagg cagcgcctct ggagttgttc attccttccg gtgggttcgt ggtctcgctg 128220 gcttcaggag tgaagccgca gacctttgcg gtgagtgttg cagctcataa agacagtgcg 128280 gacccaaaga gtgagcagca gcaagatgta ttgcaaagag caaaagaaca aagcttccac 128340 agcacggaag gggacccaag caggttgctg ctgctggctt gggtggtctg cttttattcc 128400 cttatctggc cccacccaca tcctgctgat tggtccattt tacagagcgc tgattggtcc 128460 attttacaga gtgctgattg gtgtgtttac aaatctttag ctagacacag agtgctgatt 128520 ggtgcattta caatcctttg gctagacaga aaagttctcc aagtccccac tcaacccaga 128580 agcccagcca gcttcatgtc tcaatactat gtatcaaaac attttcttga tgaattagag 128640 caaggatatt taaagaatga aaaatgtaga gaacaaaaag aaaataaaat cttaatcttg 128700 gatagtcaaa ggccacctaa gcctaacaga ataaaatcaa ttacaaagaa gaatgaattt 128760 gaatttttaa acaagtcaaa aataaccacc aaaaatattt tctaaacaaa atatgaaata 128820 caaccttaac ttaccatttc agtaagaaaa tgtcaaatgg acaagattta tttgctatat 128880 ttggtttttg ttatcaacaa tgcacactgc tggcaaggaa gcacagatac tgtgatatat 128940 tgcatgtggc agtgaaaatt tgtgtaactt ttctgagaat tggtttttta ttatataaaa 129000 tgacactaaa aagcatctta tcctttgatc ttctaattgc atttataaaa atctctccta 129060 nagaanagta acctgagatg catacaaaga ttgatcttca aaaaatgttt tttgttgcat 129120 tatttactgt tttgaaaatt ttgaaacagt ttttctgtcc aaccatagta gaatgactaa 129180 gtaaattttg aaatatagtg gtttatcatg caattatcaa aactatattt ttatagacta 129240 attagtgatg taagtcatga gtgagagaaa tggtgatgat aaaatatttt taaaaatttt 129300 accaaactat atttgcagct gaattcaatt ttgattcaag tacatttgta tgcctagaaa 129360 caaactgata gaatgtttaa attacaaaac aaaatatata cattttttgt tatatgtgta 129420 tatatatata tatatattta aaaaacctgt tgtttgaatt attttatgct tcatatattt 129480 tggtattttg agatttttta ctatgaaaat cttcaaaaat ggaaacaggt ggactgataa 129540 acaaatagat ataaaacctt actttatagt taaacatcag tgagaaaaaa actgtttttt 12 9600 aaatggcaag tacatgaaca ggtaaagttc taaaagaaag aaaatacaca cacacacgga 129660 catgctagtt atatttacaa taaaactaat ttaggttttt gacttgtaaa atattttatt 129720 aaagtaaaat aatttcaatt aagtgtttat tcataaacag ctatcaagtt tatttttaag 129780 gtattaaagt actaaataag cactattaga agtgtggctt tgaaatttat atctgcaaaa 129840 gaccacatca tttttgataa agctctttat ttgtgagctt agtgaagaga aatgagaaat 129900 caaagttgat tacacaggtt actttttcac atgataagag aaagatacag ggagggagga 129960 aggcaggaat tgnntgagat tttagaaaat ctcagattta aaacttttaa aaattaagta 130020 ttgggcatga ggaaatgagc agagaaaata gtagtcatat ttccacattt gtaaagagaa 130080 gaacaatagc caggcaaact ccatgtctga agtgtggttt attttaatat gcaaaacata 130140 agcatttctc tgaccaaagc tgctttctat ttaaagctaa atataaaaat attatttttg 130200 agcatagtat tgcgagaaca tagtgaaatt tcctatgtag attctgtgct gttttagtta 130260 cggaaatatt tttaattaat cctccattta .acaatggggc atacatctgt gtttttcgaa 130320 gcactgtttt gtatatatta gtaattatca ttgcactgaa atgaaaatat caatttcatg 130380 tacaatttta tagcacatat ttgattgaca atgaaatgca tttaacttat aaaaatgtaa 130440 tatggctgac attagcaaaa tctcttatta agaaataggt cataagtact ccaagaaact 130500 tgacagtctg ttctatgaag agcattaatt caaaagtccc ctatttgcat aaatctccag 130560 gtctcatttt cacagctgtt tgttgtcagc tagacggttg cagttcaaca ttattttaac 130620 ataatcggtg ggaagtatac ctttcaacta aaaataccat gctctgcaag aaagattgtc 130680 aggggtaaaa tataagttca ttcagttgat cttgatattt ttccttaaaa taaatatttc 130740 taatacagga actttaagtc tcaaattact gaaatatttt tattattaaa atatattttg 130800 taattttata ttttcataaa aataaaaagg ttatgtgttc atacataaaa atgttaatca 130860 gtagagctta ttttagcaaa gagtaaaaaa tactgaggat atctattttt cattaatcag 130920 ccaagcatta tatggagtgc aatataggtg tgctggaaaa aagaggatta aaatttttag 130980 atattgctga gaatatgcat gtatgttctt tgttttatta attttaataa tggaaaggga 131040 tcagcaatta ttagatctta aatgtctttt acctgtaaca cccaagagca cagaaggaat 131100 caaagtcaag agatctcagc ctggatactg agcaacacca tttgctacct gtaagctgga 131160 gttcttgatt taaaccctat gtacttcagc tctccatctg cagtaggggc cattatatct 131220 acagtaatca cttcacaggg tcattgctaa gattatatta aataatggac aattaactac 131280 tgtgggaatt gcaaagcacc atacaaatgc tagtcctggt tcagtgcctg ttctcatatc 131340 atttcatctt agctgcaatg aaatcctagt acaaattcat gaccttgtcc agctgtcatt 131400 tatggtataa tacataaaag ttaacccaag catagccaag gaaagagaca attggacata 1314 60 cctacatata ttttcataac tagcaaaaag attcgctggt ttttgccaat gtgcttctgt 131520 tacttttttt ttcttttact cttgaaatta tatttaagtg tagactccac ttaaagtaat 131580 ttgctgctgg aataggcata atgttctgat gtctgctttg cttctaatta tgttcaagct 131640 cattttgaca tttaaggcca attacaaaag tctagattca gaagactttt ccttagtcat 131700 ctaaaatttg aacaataaac agtttttttc tcctctctct ttattcattt aagtggtatg 131760 cttggcaaca agcatatttt gggaattgta aaagcaaaat aaataatgtg ggttcatttt 131820 - 159 - 008252 gaaaaaagaa ctacttggtc tttcttgcca acataaaagg cagtcacaga attaactgaa 131880 accaccagtt tcaactttat ttcctcctaa atcacaagga ataaggaatt attttacaga 131940 tacagtgata ttttaaacgc ctaagttcta ttaacataaa ggtaattgac ttcaacagat 132000 ctctaattcc aaaagagttc atacatagag agatatgtat aaaaatagct acaaaatctt 132060 tatcatttaa aagtcactca taattaaact ccccagacta ggaccctaca ccaaaattca 132120 gttagtgttc cattgcgttt taacatccag tcatgtgatg acttacgaca tagagcaaag 132180 cacttaattt acctgcctcc ttattttcag tcaatttgcc aactgcaatt tttactgttg 132240 attcagtaga agaccctgat gagttgacag caccaaaagg caattcatag catctattag 132300 taagaagtca ctgtagcttc tttgtttaac ttttataggt aagatctggc ttggaaaaat 132360 agactggaaa gtatattgta aaaaaaacaa aattttccaa ttgactgtca ttatcagtta 132420 atagattata gaatagtaga cctttatagc tggaagatat gtatcgtgta cttcaaagat 132480 gaaaggaaat aatgtctttg aaaacactgt caaattttca aaattatgta tctaaaagaa 132540 attccttaaa aaatagttta tcctttataa tttttgaact ttataaaaat ggaatcacac 132600 catatatgac caaacatttt aagtcataaa gaatacacag gaattctggg ccgggcacag 132660 tggctcacac ctgtaatccc agcactttgg gaagctgagg tgggtggatc acctgaggtc 132720 aggagttcaa gaccagcctg gccaacatgg tgaaaccccg tctctactaa aaaatacaaa 132780 agtcagccag gtgtggtggt aggcacctgt aatcccagct acttaggaga ctgaggcagg 132840 agaatcgttt gaacctggaa ggtggaggtt gcagtgagcc aagatcgtgc cattgcactc 132900 cagcctgggc gacaagagcg aaactctgtc tcaaaaaaaa gaaaacatag gaattccgga 132960 atttaaccac tgtgatagaa gttgcttttc cttgggacaa tatttttaag gcagatttca 133020 aaattattat ttcaactatt ctttatataa tgctttatta tttttaaagt attcacatat 133080 atgttatttt gattttcaaa acaatcttcg agtgacaatg ggattgcata tttgagaaag 133140 gctggtctca ggacggctga atgtttttcc caaagccgtt aagtaagtcg gtgacttaga 133200 ctcagacgct gtgaaagcag aggctgagta cttggttctt gacctcactg atctgctggc 133260 ttacagagca tcctggtctg catactcagc gattcaatct ctatcttaaa cgcagtccct 133320 cagccattcc ttctagaaag gtgatctctt tagagtgaga gcagtgatgg aggtggctga 133380 agttacacca gggggagcag ctttgaaaag ctcctggctg ataggcatgt actaatggga 133440 gcaggagctt tgttggcagt gtcgaagtcc ctttggaggc tgtgtgagga gggaacccat 133500 ttccctggga tatagctctc tccacagggc agctggatgg ctaggggcag ccatggagta 133560 tcagattcac ctaggaatcg atatttttcc tgagggtttc acaaaaaaag acagagagat 133620 ggaatttgaa gaactgggaa gagtgtgatt gaattggagg gatctgaaat aaagaaatag 133680 ctatggtcaa tggctctaac caacagtact atatttatac tccaaattcc aaatccacac 133740 tatgtgtatt taggtacaat ttagtttagc catagtaagt tgtggcttct ggagctttta 133800 tttaccatag gattttaaat acatttcatg tcatcttcat tggttttcct taacccctaa 133860 ttctccacaa gatcttcatc ccgtcagtta tgactttgcc aaccaatata ctttttagaa 133920 atgtaacccc aacattaggt acgaccagca tttactgtat atcaatatta tcctaaatat 133980 tgctgtagct ttgaatagct gaattaagtt ttaaataaaa gcaaaggaat tataaaacaa 134040 aatattaacg gattatgagg tcagtgggtt .tttctgaaga taaaactatt ctactaatta 134100 ttgctgtttg ggaaaaatag aaacaaggag actgagtaaa tgaatcatta aggcaattaa 134160 aaacagaaga gaagtcactt ttgcactcag tgaataccat caaggcaagt cttctatttt 134220 agggctgtct tatagtaaga tataacaggg actctctcaa taaaaaggtt gagctactat 134280 ataaatcatc aagtagattt tattttgaaa gcaacttttt ttgtagagaa agagttcttg 134340 aaaccagtta aagatttaaa aagttgatac aagttatgtt gtaaaatcaa agttatatgt 134400 ggatgatacg agttgtattc cccaccacat tttaacattt ttagtgtata tatttttaaa 134460 tgaaaaaatg tacattttac acgtatttta gtacaaacag taaaaaaaaa gtcagaagta 134520 atttatcaaa acatatgttt acacaaaaac ttgtatataa atgttcataa cagcattatt 134580 cataatagcc aaaaggtgga aacaacccaa atatcaactg ataaataggt aaaatgtggt 134640 atatccatta ataaaatatt atttggcaac aaaagggaat gaagttccga tgcatactac 134700 aacatggatg acccctggcc acatattggg tgattccatt tatagaaaat atccagagta 134760 caaaaagcca tagagacaga aagtagatgt ctttgatttc caaaggctga gggagggagg 134820 aatggagaat gacagctaac aagtatgtag cttcttttgg ggataaagag aatgttctta 134880 aattagatag tggtaatggc tgcacaacca tgtgaataaa ctaaaaccca cttaatcgta 134940 cactttaaaa ggtgaacttt atggtatgca aattgtatct caaaaaccta tttcttttaa 135000 aagtcaaaaa aaatagtata gaccccttcc atagttcctt agtcctttcc attctagtcc 135060 ttagagttac aggtttattg tatacacttt caagcattta tatgcacaca aaagcatatc 135120 tatgtgcttg ggatatagca aagggatggt actatatatt ctacttactc tactttgacc 135180 aaaacccaag atgtttaacc attctacttt ttcttaatat ttgactatgt ttgaacttaa 135240 attgatatcc acacatacta aaataccaaa atatgcattg agtcatcaaa tttcaaaggt 135300 catccatgat attttttaag ggcttaaaac tccttngcat gatcnttcag cctgtttant 135360 tcagcatagg aaaataccct tgctttgcct tcacctaact ctaaggaagt ctattagaga 135420 tatttagggt cagaaatggg ttcttggggc ttgagagaga aaaggatctt ctgctattga 135480 tgatctggct ctgccatgac cctatggcat gggtcctcag tggccctgtg ggaagcctag 135540 tgtaggcaac atggcctagc ccacctatgt tgatgtttaa ggggtgctag gaacaaaaga 135600 - 160 - 008252 gaagctgtgg aaacttttgc ttatccccct ttcttccctc tttggtggtc ttatttttat 135660 ccgacgctct taatctcaac ttataaccgg aaaatacagc accaaagcgt gagggcattt 135720 aggggaaaaa tgttatctgt tttggaagta atatcaagga agatcaggga caaatgcaga 135780 actggaagca aggactaagg ttggatttag caggatttcc tgaaagattg gatgtaggat 135840 gtgaggaaat gagatgaacc aaagatgtct tcagggtcat tggcctgaat aacttgaaga 135900 atagatttgc cattttcttg gctgggaagg agtgcagaaa gaacaggttt gaaagtgggg 135960 catgaggagt ttcatatgac acatcaaata cttgagattc ctgttatcta acatctgtcc 136020 aagtaaaaat gtggagttga ttctggaggt ggggctggaa atataactgt ggaagtcatc 136080 agaacataca agggatgtga aggcattaca atggatgaga tcaagcaggt agtgagtatt 136140 gagatataag aaaagatctg aggtcaagta ctggagctct gtaacattta gacattagga 136200 aattgaagag aaaccagcaa aggagcatgc aaagggctgg cattggattt tgtggctctg 1362 60 tagtgcagtg ctctggctct tttggtgact gaaagatgct tgttcactgc ttttctctgt 136320 gttttcaccc ggagcatccc tgtgacattt ctatatatat atatttattt atactctaag 136380 ttctagggta catgtgcaca acgtgcaggt ttgtacatat gtatacatgt gccatgttag 136440 tgtgctgcac ccattaactc gtcatttaca ttagctatat ctcctaatgc tatccctccc 136500 ccctcccccc accccacaac aggccccagt gtgtgatgtt ccccttcctg tgtccaagtg 136560 ttctcattgt tcaattccca cctatgagtg agaacatgcg gtgtttggtt ttttgtcctt 136620 gcgatagttt gctgagaatg atggtttcca gcttcatcca tgtccctaca aaggacatga 136680 actcatcatt ttttatggct gcatagtatt ccattgtgta tatgtgccac attttcttaa 136740 tccagtctat aattgttgga catttgggtt ggttccaagt ctttgctatt gtgaatagtg 136800 ccacagtaaa catatgtgtg catgtgtctt tatagcagca tgatttattt atttattttt 136860 tccaactctt ttttttatta tactttaaat tttagggtac atgtgcacaa tgtgcaggtt 136920 tgttacatat gtatatatat gccatgttgg tgtgctgtac ccattaactt gtcatttaca 136980 ttagctatat ctcctaatgc tatacctccc acctcccccc accccacaac aggccccagt 137040 gtgtgatgtt ccccttcctg tgtccatgtg ttctcattat tcagttccca cctatgagtg 137100 agaaagtgca gtgtttgact tttgtccttg tgatagtttg ctgagaatga tagtttccag 137160 cttcatccat gtccctacaa gggacatgaa ctcatcaatt ttatggctgc atagtattct 137220 atggtgtata tgtgccacat tttcttaatc cagtctatca ttgttggaca tttgggttgg 137280 ttccaagtct ttgctattgt gaatagtgcc acaataaaca tacatatgca tgtgtcttta 137340 tagcagaatg atttataatc ctgtgagtat atacccagta atgggatggc tgggtcaaat 137400 ggtaattcta gttctagatc cctgaggaat tgccacactg acttccacaa tggttgaact 137460 agtttgcagt cccaccaaca gtgtaaaagt gttcgtattt ctccacatcc tctccagcac 137520 ctgttgtttc ctgacttttt aatgatcgcc attctaactg gtgtgagata gtatctcatt 137580 atggttttga tttgcatttc tctgatggcc agtgatgatg agcatttttt catgtgtctg 137640 ttggctgcat aaatgtcttc ttttgaaaag tgtctgttca aatcctttgc ccactttttg 137700 atggggttgt ttgttttctt cttataattt gtttgagttc attgtagatt ctggatatta 137760 gccctttgtc agatgagtag attgcaaaaa ttttctccca ttctgtaagt tgcctgttca 137820 ctctgatggt agtttctttt gctgtgcaga agctctttag tttaattaga tcccatttgt 137880 caattttggg ttttgttgcc attccttttg gtgttttaga catgaagtct ttgcccatgc 137940 ctatgtcctg aatggtattg cctaggtttt cttctagggt tttcatggtt ttatgtctga 138000 catttaagtc tttattccat cttgaattaa tttttgcata agattcctca tacattcctg 138060 ttgtgctttc accctctact ctctggttct caccctgttc tcatatatcc aaggaagatg 138120 atacactcta attagaaaca acaactgaac tcagctcatt attttcaaaa tctttcttct 138180 ccattaggta aaaatcattg cagtctgttt attcaatatg ttttcttttc cacttaaatt 138240 ctgttcatgc agagactgat tcctgggaac atcaatgtgc tggcaacttt gactttgtaa 138300 tttccctttt gttgtcccac tcaactgcat ggctgatctc cattcatttg ttcacttcca 138360 gagcctactg ttggcaatct gatttttctt catagtttcc cattaaaatt cactagagat 138420 gcttttgatt tacttatttc ttccctggca gggtagagtt gcttgttcca gatcacctca 138480 tagcaaactg gaccattctc aagtaaactg ctcatccctg gtgcagttat cagcagggag 138540 gcatctgcag cattaacagt gattgacccg tatagtgatg tttaatatgt gataatgtta 138600 ctagcagcaa acaaaatagc acttgtaaac acagttgttt actcatactt acatgtatag 138660 gtttggactt gaggggtaaa atagcaaaat tacccaactc ccctgctaat aaaacctaag 138720 gaaatgaata aaatgacctg agaaaattat accagcataa ctacagtaag gaagagctac 138780 aatttcatgc cattttctcc gaatagcagc taggaggaaa gaaaagaaga gaaagatgaa 138840 ggaggaggaa aaggaaaggg tcgcggagga ggagaagaga atgagagaga acatgaatat 138900 tctaggctca tcccttaaga ctgtcttctg gaagaagtta agaagaaata aacttggaaa 138960 aacaacgaaa ccgtatactc aaattcccca atctgtgaag tctgtggcca aagggctatg 139020 cagagccctg gaaagtgata agaaccactc atgtatagaa gccacttgtt ccagaggcaa 139080 aataatccac tagatctcac catttttatt tttttcagct gaggaggctt gagagtgaag 139140 gcaaaatgtc ataccttctg aaaatgaatc atctcccatt ctgctacaaa aatgtagcat 139200 aatgaaaata tgaagagaca aaaacagaag cttcaagagt acaggaagct caggataaag 139260 aggggtgtcg aaagccccag gagttgtcct attcgcccat acagagaacc aaaaccaaca 139320 ggaggacatt gacttctccc tcacctggag tcagtggagt gggaatgatg gctggagaga 139380 - 161 - 008252 ggccagtgaa tttggggaag tgaacactta ggactgatta gttctggctg caataaagat ttcctgaacc tacctgaatt gaagaaatat ctgaagcctg atatttcaag cataattgcc catacacagc ctcagctaat acttctctcc ctgtacccct ctcctcatac tgaaacagta aaacacagag aagaaaacct cgttgataaa ttgaaaaagg atgctaatga gtgggatgag atatctggaa taattctctg cccctgtatt agaaagaatg aaagatctag ataataatgt taaaagtgga aattttggca actgtggaac cagtctagaa agaaattagt tcaaattaaa acaggtagtg agaggacccc aagaataata tctatgagag taagaatcaa atggtttctg ggcaattgat ggtttaaaga aagaacatct gcaggatagt aagaagggtt tattctaaat gataagtgta atgtgtgaga atctggaagc tacttgagat agaagaaaga aagaatatat ttanttttcc caatgagaca tgtgagaaat agtcagaaaa tctcagcaaa aacaaaacca aaccaaagat tgtattaaaa aaaaatggct caaataacgt gagttaaatt ggcccagatt ctgagcaatt aatggaaagt aaggaaggtt ccatttaacc ttgaccattt actgtctaat agagtcttgg agggggaagt ataattttag aacattacct ggcccaagag tgagcaatgt tttcatagga aacattgact atcaacatat tgattatcaa cctgctgaga agcacggatg acccaattat gatagaaaaa caaaatgtaa atattgtcaa acttgaggat ataaatgtat acatgacacg agatggaaaa tgaaagcaga gagaagaggt gacctgacaa gtggcaatgg taataccttc atctcacact ggaaggggtc aagtctttct agtttgatgg aaaaagaaat cgagatttac atacaaattt gggagtggtt aagggaagtg gggagtgatg ttgtgcctga atgaaatctt cacatatcat agcagaagtc tatgtctaaa gtggataaat caagaaatcc taaataggga taggcataac gccatcagaa gtgaaaacaa aaaattcctg tctcattact tataccaaaa taaatttcag atttaaactg atttaaacag ccaattcagg ttcaacatgt ttaaaataat aaagcaagga aagtgtcaaa ctactgcata aaaccatggg taaatttatg tctaatcatg cagtgagaga agtcttttta agcaaaataa aaacacttaa aaggaatgac aaattagacc acactgaaat ttaaaactga tctttgcaaa aatataagaa atgtataggg gaaaatgcac tgaacataca tggcagataa aagatgggtt ttcttatttg taaagagttg gtggaaacca ataaagaaaa taaaatggtc aacagggcag aaaaatggaa taacatgcga ataggtagtt cagagaaaca acttcctctc tgccttcttt ccactgaaac tctactcctg tgcattactg aggtcagttg cttcattttc ccaaaactgt tcttgattca cccaagagga ctaaattggc tcttctctgt tgcccctcag tactttaacc atattttgct tacacaccaa agttactatt cttccctttc tttttatttt cttttttctt ttgagacagg atctcgctct gttgtccagg ctggagtgca gtggcacaat ctcagctcac tgaaacctcc gcctcccagg ttcaagcaat tcttcttcct cagcctcccg agtagctggg attacaggtg accatcacca caactggcta atttttgtat ttttagtaga gacggggttt caccatgttg gccaggctgg tctggaactc ctgacctcaa gtgatctgcc cgccttggcc tcccaaagtg ctgggattac aggcgtgagc catcatgccc agccagtatt cttccttttt gagaattgat cctgttcatc tttttcttcc cctctcaatt atacaattct taagggaaga aactatgtct tactccactc aatccccaat gcatcaaacc tttagtaacc actaataatc cttgtttgaa taaataaaga gcacaggccc tacggacaat gtaagtcagt aagtgcactt gagtcactat ctcaagcaca ctaccatttg aatttaaatt tctggataaa aatctccaca aaactgataa ggcacccttc aagcaagaca caagttaatt ttcagcctct aatagaagta tttgttggca ctacctttca ttggtttatt catttcagca gccataaaca cacagatccc attgtcacag aaagccttgt tcgcaaatat aatttagtgc atctcacaat atgagctttc aggtggttga gctgtgtgga aattcgattt ttttttctcc cgagactctg aaatgaaaag aggataaccc cagtgctctt gcccacatcg cttatctcac aacaacagat tacaatccca taactccacg ttcccaagca gctactgtgt tgatcaaatg taatgcaaca gatgagaaaa catcagctgg ttcatgccca agacagctga aaggtgcact tacattttta gtctccagat accaaaataa atctgttggg cttaatcatg agttgtcctg ctacttttgg ggaaagaaag ccatgtccag tgaaatttct tttatactgg caggcatttt gactcttcac ctattgctgt tgaatctcca ggatatgtac atgttttcac accgatacct tcattgaaga aaatacagca atctgatatt tttatgtgaa tcatctttgg aattcgtgtt cctcaccttt ggatacaata aatattcaag agaaaaaaaa tacaggaaaa aaataattgc cctcaatgat gagttctgaa atacttataa ggataagtga tgtgttgaaa aattctcaaa agcaatttta gaaggaataa tttttgcaga gagtggctgt ctcttgggca ccataacaag aaaaatagta tattgccctt ggtgatggtg cggtcaagta tcaataacaa cacgcttcat attttcatga acaaaacact tcttcataag gtaatgagat tatagaaaaa tcaactcaag gtaaaaataa ttactttgca catggacaaa ggataagtgg aagaacttat cttctagttc gtggttcatt taacaattaa ttgctccttt ttcaaatagg tccactgtgt tctgggcctt gaacagtgta cctctaagaa agtttgttat ccttactgaa ggaaattaca gttaggtcgc ttctcagcct tttggctaag atcaagtgtg aaaattatac ttagattact atgtaattag gcctggtaga tcctgggtag acttcactga atgcaggatg caatatttgg aaaagtactt actttggagt cctaatgcct gggtgctact tctattagtg ccagaaatta atagtgtgac tttgggataa tcacttaaca tctttgagcc tcattttcct tatcttcaaa aaaaaaaaaa aaagaaggaa tcagatgaaa tcatctttac cagattaggt ctatctttag gtcccccagt gtaactgggt ttctgtcttg gtctcagtcc atccttgcat - 162 - 008252 tatcctggac tactgaagtc cagatatcat tgagttggag agccaaagat gctctatttt tcttgaccct gtccagtgtg atgatagcta tgccttagac aaaagtaatc accaaagatc ttttaattca agatctttat ataaaaatca atgcaagcct atgttccaga gtgggacata actctggtga atcaacagat taatctggtt tgatttgtta gacagaacaa acctagatga cactgtcttc atagacctca gactggaaaa aagaaaactg tcttcaatat actaaggaat gcaaacctaa v. wu.ba^^ae agttacaaaa cccagttaac atggaatgta tcttgcagct gtctttgtca ctctaaggtc tataacttag caccaaaaag gaactacata gaaagatagg gtgctggtgt taagcctgtg agtttattaa attgtgggtt taattatttt ttcctcactg aaaatgaaag caagtgaaaa agccttcttg acttaaagtt caagatgaaa aaacatctgt aaattgaaat ttcagaagtc aaattggagg ttctgaagac ataattcaca tctgaggatg gcatttaaag tatgcaagta tcttctttct tttatttatt tataaagatg catagtctca actcacagat attggcagaa ttattttttt gtgccttgaa gcaatatcac aaataatatg aagtgtaatt tgttgaaaat ttgaatttgt gggtaattat tcttatatct ataatattga aaagctagtg tcctcaccca tgtgggcacc gttcagcatt atttggccta ggatgagatc agttagatat aatgaaaatc tctcaggatc agtttcactg gggtgcagtc aaacacatgg aataccaaag acttccatta gtaataccaa gttctaacat +- i r> т" /т _"-_> "¦ tatcttacaa agatgataaa aatatgacca aattattatt aatcaaactt agttttaatt aagttgtgtt ttaatgagaa gacatagttt aacattgtgc ttgcacaaca ccatttattc gctacttgtg aaatgcagga atagtaatca aaaatgtttc ctggaagaaa ttctactggg ggcaatctaa catccaagga accaatctct tctaactttt aacttgttct tacatgttaa gcaaaaatcc tcacataact atacacccag ttttagagca ccatatatta tcatttatta ctccttccct ggaatcccaa tttttaaaga ttaaaaagta acacatatat agtgtgtgaa gtttgtttgt acttttttta ttagaaatat atagaaaggt cactataaaa gttagatagc ttctaatttg tttcttattc ttcctatgtg gatcaaagtc tgtaatacct ggaaggatgg cgtggttgcc tttttttcaa ttgtcaattt cctccttcca tttctccatc tcacatttac aaaccggaac catcctcttc agttaattag aaaaaaataa tccagtaaga aaaacaaaga atgctcatct cacgttagaa aaatacaact tattattgct actgatactc gagtgttccc gcttccctgg ctgccaaaga ctaaatatct tgctatagac ttcattgtta tggccttttt aataggaatg cttagccctg attcatccat tttaagttac tagatttcat acctaatcat acactcagat aaactagcct gtcactaata atcttcttta ctcctggggt aattctactc ttattgcaat tttattcaca cgagaacagt agattcaggt tccctgatct tttccaaggt ccagcactga ataaatgtta gggtatcatt atacctcttc tatctgagaa gtcactagac tttattgact ttaattagaa atttttagct ttgttttttt aatcatgaaa ttctttttta accataaaaa ataagtttct ttgctgaaag ctcaacagca cacatcaaat ttgaatcaag ttcaaacatc ttgttctttt gtggacgtaa atctatttca ttatatcttg caaaatggga ttctaagttt aggtaccagt ataccaattt tccattatta acagctgtaa aaatcagcac aacacattgc accgacttct aaaaatgtta ctatgtaata gttccaagtt -"^-(--l Г-*-- aatctttgga agaagagttt aggtctgtct ataatgaagt atcactcaca atgaatgaaa agacagaggc catggtgtga atacatggag gaagcctggc gggtctttgc tcatttacac tctaaaacta tgaaatcatc tttcctaata atatgtttga attacattat taataaaata tgggctaaaa tcacctgagg tttccactta tttcaaacaa ctttttaaaa ttggtaaatg tctagctctg ctctgttgtt tgtcatgtat gcccctttaa aatttatatt tatgcccctt tcccaggcaa ttcaatttat tccagtgggg actaatgtgt aaatatgtaa tggctggcct ggtcttggct aatgtgtttg aagagtccca taaaattata cttagcagca gaaaaaaatt tttactgcga agttaaatat actgttcttg acatgctaat cctcttaggc tggaccaaac ttctctatca gaatttataa gccaaaggac ctacacatca gtttgtaaga aagtatcccc ttttttttct ttaaactgaa agtggttttg tgcaagtatt gtcacacaag tgaaatgcag aatttgttca gaggactcaa +¦¦-+¦+• 4- nnr* -i -i acacccgatt agtaatttat gatctagatt ttaaagttgt tcaaacaaat taagtccttg tttcaaaact agtcatgggc ttgactttta ttgcaaagct atcttccctt atcatttatg tggaattgct tatgtcaggg aagcaacaaa ttcttaaatc ttagtaaagt aaatattaaa tatggcattg ctttaggaat ctgatttaag cagttataag gtctgtaagg tggtgtcgta ctggtcactc tgtatcggaa ctgcgagtta atgtagataa atgttcagat cctgaattaa tgactctgat gtctttatcg caacttgctc ataggataca gaatgaaact ttgttggaat tcttccatga tcttcaagct tgtttttaat tgcctttata ggaacaaaga aacggcaaaa acaaaactaa cacattttta ccctgatcag ttgaagttgt atttatgtcc tctaaatgct tctttttgcc tgggatgaac taatgcagac aagaactgct gggaaaacaa aagcattcac catgtagaga gacagaagct tgctatgtgc gtaaaagcac cagatttttc cctgtgactt tatttaagaa atgtttccct agatagtatt ctgatgtcac tggtccttag ctgaaaaaat tacatataat ataaactaga aagacatgtc tcaaacagta aaagttacgc gaatctggct ccaggacctt cgttcaacta cagactgttg aatatcaatc atggtttact agggacacaa aacatttttt actggtatat gttatagaac ccacagagca tatctaccca ttaaaactga caattgagaa caagactacg gatatctgat agtgaggata accctgtatt gacggtttgt atttttaatt tttccattct aattttaata - 163 - 008252 taaattcatt ttccaatttt tagaacatat ttcactaaat ttaacattct gtcattcagt 14 7000 agaagactca gtttgtgtaa cactaaggag aaaaacaatt aaactataca catagaggat 147060 aaggtgcatc cctacttcag agtttttaaa aggtgaaagt aaatgtgcat cttaaaatca 147120 atgaaatgca gaggtttccc acacatatat gcatccatta aaaatatagt atttagggcc 147180 agcacggtgg ctcacgcctg taatcccagc actttgggag gctgaggcag gtggatcacc 147240 tgaggtcagg agttaaagat cagcctggcc aacatggtga aaccctatct ctactaaaaa 147300 tacaaaatat tagccaggca tggtggtgag cacctgtaag cccagctact caggatactg 14 7360 aggcaggnna gaatcacttg aatctgggag gtgggggttg cagtgagccg agatcatgct 147420 gttgtacccc agcctgggca acaacagtga aactccatct caaaaaaant atatatatat 147480 acacacacac agacacacac acacacnnnn gtgtgtgtgt atatatgtgt atatatgtat 147540 atatgtgtgt atatatgtat atatatgtgt gtgtatatat gtatatatat gtgtgtgtat 147600 atatatgtat acatatagta ttttaaactt aaggttacca tctaaataca ttttatgccc 147660 aaaattctga agattaatat taagttttct tagtttgcaa agcactgctc cagtcaactg 147720 atggccatcc ctaggggagt ggtctggtaa tacatacttc tctcccntat aagtccacgc 147780 tggggccaag ttatacatgg tcatgtgttt ccctccttat caattatttg tagcagcaat 147840 ttttagaaca aatggtatca aactcaacat atattctgca gttaaatttt tcactggcta 147900 tttttttctg agatcttttt atcttactcc acatgaagct aatttggtca ttgttataga 147960 gagccgtgtg gtactccgcg gactgtgaat atgccatatt tgtttatcag ttccctttct 148020 gatggctatt taggctgttt tccaaagttg tacttttata atgctgcaat acatttatgt 148080 atcgtgccca cttatgtgaa tttctctagg atatatgtcc aaaagtagaa tgcttagatc 148140 ttaggtactt cccaaattgc tctataccag ggacctgcaa gcccagatac caaatccagc 148200 ccaccacctg tattatacaa cctgtgagct aaaaatggtt tctataattt agtggttata 148260 caggtaccca cagaatatcc tcagtattgc ctcatgaccc tcaacgtcta aaatgtttac 148320 tgtctggacc tttaagaaaa cactgtggac tcctgctcta tagataaatt gtgctaattt 148380 accctcccaa gtgcaatagt tgagaattcc tgtgttctta cactctcact tagacgtcac 148440 attgtttgac atgtgaggtt ttggcaatct tatgagtgaa cactggtatc aattgggctt 148500 gcatttttgc atttccataa ttattaataa gggagcacct cttactatgt ttattgacca 148560 tcttttttgt gactcaattg tttgctttac ctattttcct aaaaacaaat ttgtcttaac 148620 tcatttattg gatatttgtt ggatattaat ctcttgacta ttacatgtgt tgcgaatatc 148680 ctctcttagg cagtattttg tcttccagtt tgtctgtgat gtcttttgac attctatata 148740 tttttatttc atttttagtt aacaaataat aattgtatat atagtagtcc tcctttgtcc 148800 ccaagggaaa ctttccaaga ttcccagtag atacctgaaa ccatggatag tacagaaccc 148860 tatatatact atgtgtttta ttatacatac ataactttga-taaagtttaa tttgtaaatg 148920 aggcatagta agaaattaac aacaataata acaaaataga acagttatga caatatactg 148980 taataaaagt tatgtgaata cgcattctct ctctgtctcc atctctcctt ttcaaaatgt 149040 cttaatattt tcaggctgtg gctgaccaca gggaactgaa actacagaaa gtgagacagt 14 9100 ggataagggg ggggcctact gtatttatgg ggtacaacgt gatgttttga tacatgcata 149160 cattgtggaa tgatcaaatt aggtcaaaac .tatcggtttt tcactgatgg tctgtattta 149220 ttttgacctc atgcttaagg aattctttcc cacctgaggt aatacaaata ttctcctata 14 9280 tcttcttcta attgtttaca atttttgctt ttcaccttta agtctttagc tcatctctac 149340 ttactagtta gatatagcat aagctagatg aaatatactg tttaaataaa actcaaatca 14 9400 atttttctca gatttatatg agaaactgca aggtttagaa tatttatttg atttatctga 149460 ttatgaagtc taacacaaaa aaggtaaact gttcaaactt ttaaatttct aaattatttt 149520 ttaaatgata tataatttaa taaaaggaaa cactatataa ctaaattaaa acaaagatta 14 9580 tcacttctat actacccaaa aaaagtctaa tactgaagtc acttccccag ttcacaaaaa 149640 attttgctta aaacatggtc agcagttgtt tctcttttcc tttgtaccct tagactaaca 14 9700 gttactgtaa tgggctttat gaagtgagtt caatttgatt tgatacagtt ttactgaacc 149760 tctactacca cgtgcaggta ctaacctacg tcctggccaa agtgggaaag agcttaattc 14 9820 atgtcctctt aggggctaca aataattgga aattagcaat aacgaacaca gacaggctaa 149880 ccaacaaaca aataaaacca gtggttccaa ggcctaaaga tgcaaatggt aagagaggca 149940 tgtgtacatt gctacttgct atgtgatcaa taaacatctg cttcccagtt gttcccaatc 150000 tgccttatga gaaatatcaa aagtagtggc agtgaaacag ctggagatag tctgtgggct 150060 ааС-а + яУяая r*=ir"af" rtf-nt* n япппт-пягягт nt-r*t-rfr-rr*-+- д няга^а^яИ1 rrpa f rrrrrrt 1 ЦП"! 70 ---------------U — Z7 - ZJ - -э ~Э TJ ~----"Э -)-----~ ~ - -----. - ~ ~j-----^ ~ ~ ~ '-- aaaaactaaa cttcacaagt taaaatattc cataagcctg tcttcagagc tgaacttacc 150180 ttaaaatgag catttttgtt taattgtatg tttcctttta caaaatgcta caacgcattt 150240 taacccaaat gggtgagaag ggaaaaaaaa agtagtctgt gtaaacatac aagctctgat 150300 gtgtcttttg ttcacaccga acagtggtgc catacaactt gatgagttcc tgatttagtc 150360 ttcccaggga caggcatgtg ccaagcagcg aaaaataaaa tgtgtggttg cagcttgttc 150420 tggaataaag aacagcaggc tatttcatca cccctggggg caccaatttt cagcccacaa 1504 80 tgaaaatcaa gctgaagtat atagcaaatg catattcata acctccatgt attgtccctt 150540 tcaaacatct ctctcttccg ccaggtatgt attctgtatt tctgtcctga ctgattgcag 150600 tttattgaat gcttgttaaa ttcatctttt tccccatctt tgccaatgat ttggggatag 150660 ggactatgga agacctaaca agttgaaaga ctccattttt cctgcaatgt ttttctgtct 150720 - 164 - 008252 attactgttt agcgttcatt tcttgatttt tgggcattcc ctgtgttgaa tataaaaagt taaatggtaa ctagtgtata acgaaaataa gaaaaaaatc tagggtaagg cagggagaga aaataaatca aatgagaact attatacttg gttaaataga ataactacca gaggaggtat agaccctggt tgaaaggtca cagcaggcct ttagcaatgg attctccact aaaaagtgga ttaatatgat tagatatttg atacatgtac ctgaaattat aaatatcggg gggtggatca gtctactaaa caaaggaaag gtgggtagtc gcctcggtag aaagtgtaaa cgtatcccat gaaattgttt acaggaaaag tttcaaaata gattaaaggt ataaaattaa tcatggctat aattttcaac tacttttcag acattatgtt caaatgcaac aaaaaaataa cagncattaa tagagaaaga ccaggtacca aaatcctatt gataatgtgg gcattataat ttttcattga tacatacatc caaacgctat gctttctttt ttagcagggt atcattaaaa ccaggcgtgg cctgaggtcg cacacaaaaa gatcgcacca caatcactca aattagttgc tgacactcct gttttaatat acatgtggta ttctgattac cagagaactg tttccatacc atccacattg gatggacacc accttattag tacagactca cttgggtcag cctacatgcc tcagcggaaa agtttgtgct atttgctgaa atttttaaat gttgtgaagt cttctcctat tgccgtggct agtatatacc tttttttttt tctcggctca gagtaactgg agaaacgggg cggccttggc acattatata agtgagccag ctgccatcca yaavjuavj^avj ctgcactcca atagaaatat ataatttaga gtgatttaat gaacctgtaa gtgtaggtaa taatgagcat aaaaatggtt atgtgttggc cactgggttt aagctgccat agtctgtgag ccctgcacct acttcagtca agggacagtt cccaagcccc tttgtcatgc taaaattagg ttgttgcctc cacgcagcgt ccacagcaaa ctctctgtcc agtttcttgt tttgagatgg ctgcaagctc gactacaggc tttcaccatg ctcccaaagt tctaataaca tttatttttt ggcctagagt ccctgtgatg tttttgttac tggcagcttg ttaaataact tttacactgc cataccacag aaaaacacaa atttaaagat gaaatttcag acttcataaa ctgcagagtc cagaaaatcc ctgtttgaat ggacaaaatg tatttcagta atatatctgt ctgaggtgaa ggagcgtcat gtttgaacct atatcattca cattttccac ctgagggaaa catcactgat agctgtatat agacctgtta ttacagcatt tcaagaggag aaaaataggg tggctccccc ggaattccag ttagcctggt aaccacw ~ga gcctggggga cacataaatc acagtcaaag gtggcaagtc atcacattta acattagctg tacctcagct cttcctttca aagtgtctga gtaggaaaat tcttccttca aaaagaaccg cctgaatctt tctctcaccc tgatgaggat ctaccacaca cactgtcaaa tttatgctac cccaagagtt tttcccagag tatcaaatgc tgctctgagg ttaaccaatg agtctcgctc cgcctcccgg acccgccacc ttagccagga gctgggatta ttttgtcaaa tgttttgttt gcagtggcac tgaatactga ctttgaagtg ctgagaatag tgattgtata gctgattaca gcggcaagac gagtgtggga aagaaattat ctaaagggac aagaaagaaa agagagaatg attgtgaatg catgctaatc atctgcatca atttattgaa ggtcacatgt tctgggctgt aaagaatatt ggaaaattac taaatattct actgaatgca taaagaatta tcataatgta tacacagtag aaagtgccaa gtacaaacta atgaaaggtg acatagtata tataatccca accagcctga gtggtggcag aaagaaagaa ctgaaattaa tcatattttg gaattcattt tttgactttt agcttaggat cattgattcc taataaagtt atctttaagc aatgaaaagg taaaatagga acaaggctgc caaaagtctt caacaaaatt tttgactgaa ataaaactta ttcagaggna agagaaatca gcttttgaaa gagagagact ttgtcaatgc gatatctgaa tcaatatcgg gacaaatgct atagtaaatg aacaccactg gtgtcattag tcctgctgca gcactttggg tcaacatggt gcacctaaaa aatgtaatta cacagcaatc ctgacacaaa aaaagtgcat tgttgaccca agggtgatta tgctgtgttt cacagcaaag tatataaaag gaaagctata tatgttgctg cctcttgcaa attccgaggg ggaagaaata atttattgta aacagagcaa gaaaaaggaa aacgcctgtg tccactaagc tcccagactt taaacatacc atacattgtt tcataaaatt gtgttgcata catgaaaaaa tgttgaaagc cctttttccc gtataaattg aggccgaggc aaaaccctgt tcccagctac LUUUL cagagagaga taaatattaa cacaaaggta cctgtaaaat gtggctatag aaggtaatat ttctgtaaaa gaatgagggc cacccaggtc actgtatcat cttctaccct caaagtcatt ctgtttccat ggagtgccag caatctgaaa catacatctt ttaattgaaa tttcaagcaa tgttctaatt aagccaaagc atctgccttg gtggcttttc taagaaaaca tgttgcccag gttcacgcca acgcccggct tggtctcgat caggcgtgag ctgcacttaa gtttttagtt attcacagct ctctgcctca gaaataaata aaagtgtttt tcctttatta aaaaaccttt gcagacatcg acagaataaa ttgtcaccca gacctgctct tgataacata ttatcgcagt ctcacctcct tgcaaagaca ttgccacctt gtccacattc gaaccagttt aatgatgcct acttcaaact ctttaagttt tcttccttta aggagtcagt tctgtttatc ttacatatct gctggagtgc ttctcctgcc aatttttttg ctcctgacct ccactgcacc ccttgtctag tttggagaca cactgcagcc atcaatcaat atgcaaagca atacatatga catcaatacc tggcacagca ctgtaatggg tgagaacact ctcccatgac ttcagtccaa aattaaatat gcttattgtc ggcaatcagc cagccaaagc atcacagtgc aaatagggct ctgttttgta aaacaaagat ttattatatt gggccttcct ggagtcttgc gcagctggca ccaaggaaaa tttttttttt agtggcacga tcagcctcct tacttttagt cgtgatctgc cggctagaaa ttactatttc gggacttgct tcgacctccc - 165 - 008252 aggctcaaga aatcctccca cctcagcctc ctcccaagta gctggggcca caggcatgcg 154560 ccaccatgcc tgactaattt ttgttttttt ttttttttga gagacagggt tgcccaggct 154620 ggtctcaaat tcctggggtc aagcaatcct gcctcagctt cccaaactgc taggattaca 154 680 cgcatgagtc accatgccca gctgagccag tttaaacttg ctgcattaca tgcattaaaa 154740 acaaaaacac acacacaaaa agccccagaa cttttaaaag gaaaaattta gcagaaagaa 154800 actgatgtca tcactttgtg attattccag cctccctctc ttatccgtga cctctgtggc 154860 actcaagacg cctgtccatc ctgccttcct ctccagcctc caaaccctct gcctggaagc 154 920 cagactctgg ccagacttac tcaccaggcc ctcaccacac cacctggaat gcgttcctca 154 980 cagctctttt ccaaaatcag tcttctcctt ggcatctgcg tggcttggga gagcatcctt 155040 ggctctctgt cattgtctca caatcttcga taagtagtgt tcggaaagtg cccccctggc 155100 ccatgtgccc agatgggctc tccaagtcta aatgcttcaa aatggtacag aactaaatgt 155160 gtgaatttat gatggggctt catttctggt cctccttctt cacacttgcg tggccgccct 155220 tcccatccgc tctccaattt tctttcacgt ttctgctttc tgagaagaaa taagactgat 155280 ttctgggatg gagcatgaga gagacagatg cagagagaga gagagacaaa gcgtgtttcc 155340 gtgggagatg cccacacacc tcacttggga gcatcggcag tgttggtacc cacacaggtg 155400 tgttttgaca tccaagactt gaccctaatt cttcttggaa cctgatgggg agagaacctt 1554 60 gctgggggcc ccacatgggc caggggcagc cagggcagag ttgggcagct gcctgagagt 155520 tctcggtgga gaggcccaga atatcccaca agaacaagtc tccgttagtt ccaagtggaa 155580 tgacagccct ttctgagttt ccctgagttg agagctaata catgtgttgt gggaacaaca 155640 gcccaagcca aaggccacgc ggaggccagg cgtggtgact cacacctgta atcctagcac 155700 tttgggaggc caagatccac ggatctcatg agctcaggag ttcaagacca gcctggtcaa 155760 ccttgtctct acaaaaaata caaaaaaaaa aaaaagtagc caggcatggt ggtaggtgcc 155820 tgtagtccca gctacttggg aggctgaggt gggaggatcg cttgagccca tgaggtggag 155880 gctgcatgag ttgagatcac accactgcac tccagcctgg gtgacaatgt gaggccctgt 155940 ctcaaaaaaa aaaaaaaaaa aaagaccagg tggagagtat agtggattaa atgggcccct 156000 acaaaagata ggaccatggc ttaacccaca gaacctatga acacgacctt attggaaaaa 156060 gagtctttgc agatgtaatt aaattaagga tcttgtgatg agatcatcct ggattatctg 156120 ggtgggccct aagtccaatg acaagcgtcc ttgtaagaca caaaagagca caagacacag 156180 agagaaagcc ttgtgactgg tgttagataa aggccaagtg actggtgtta acaaagccac 156240 aagccgaggg acagaggagg ccaccagcag ctggaagaga caagggagta ttctcccccg 156300 tggcctttgg agagagtgtg gtcttgccga caactcaatt ttggactctg gcctccagaa 156360 ttgtgatcaa ataaatgtct gttgtttgaa gctaccaact ccgtgcacgg tgttacagta 156420 ctctaggaag cgcatatgga ggacatcact gggaatcaga gctcttagct ttgggagctg 156480 agatggtggt caagtcagca gaaagaaagc cccaaacccc ctgtacagcc tcatcagttc 156540 ccctgatggg ggatggagga catattaccc agcttgtagt gggtaacctt gaccttaaac 156600 taccctaaaa agaaaaggag catttattca aatgtaactg agcttgaaag gactgaatgg 156660 aactgatgta gcaagataat gtttccagcc cccaacagaa atggagggat tgtgcaccaa 156720 attcaattca tttgtagaaa aaccaagaaa tttattttag ctttgcataa ctaattttat 156780 gtctgtatat tttcagacat tgtatagcta tactacatgt cagggttgct cggggagaat 156840 taaatgaaaa aaaagctgca atttgttgag ctattattac atttgtaata ttttacaaac 156900 gagagcacca aaactcactg tgggctggca gttgccacgt ggctcatagc tggcacagcc 156960 tgtttgctac ctgagctctc tgactccaag cacagtgctc tcttctatgt ctgcttaatt 157020 cagtgataaa catcactgca gtgagtgcaa tattgtattt atgtatcaga ccaggcagtg 157080 cctcacacat ccatggagtc cagtcaactg tttgtcaaat ttaatagaat aacatagcct 157140 tgctgtcctt aagtagcctg ccctctgtcc agtttggggg atataattaa gaataaaatc 157200 tcctgccaac ccacgaaacc tctccttaaa ggtagaggag agagagccca gttttattat 157260 tgaaaaagct ctaacgcagg ctgcaatgca catcactggc aatctgctaa agagatcacg 157320 gaggcagaaa atacctccct cttttatatc accgctttcc atacgtgttc tcaagataga 157380 aataactagt cctcaagtaa gaaggcttga cagcaccact tgttaaatat agtttatcct 157440 aaattcacct ggtaagtggg gtggccatct gtgggagctg attgccttta tccaaaggaa 157500 aattaaaact tctcatattt ccgtgacaag taggtaggtt tacaacttgg agccagatgt 157560 ctaaactccc accctctacg gagaagggga gacggaggct cttgtcttcc ttgatgttta 157620 cattacttca aggagacagc cctcaaggaa aacattcttt taggttgtag aactggtgag 157680 aagcttattt ggctcttaaa agtatttata tacatctcaa acagaaaggg aaagaattta 157740 caattccaag tcttataaag gaaatgctct aagaaaaggg aggcgaacag gatcccttcc 157800 gacccccctt tttttagatt catgtttgta ctttcactag agagagagga aacatgatta 157860 aattaggaca cagaacaatg cctgctccag tgcccaggtg tgagagaagg ggtataagat 157920 atcccctcct gcagaaggaa accagcaggt gaacatctca gaattcgggg tgagaaatca 157980 agacagggga aggtagactc tgcctctgat attactttct gtcgtcgagg tattgggaaa 158040 tgggctgaac atggagcaaa ctttaaaatg tccagtttgg aatgatgtct ggttcagaat 158100 aagcctgatg cccagttacc agtggtgtac cagtcatggt gatgttaata aaaaccctct 158160 ggtctagatc attctgtatc ttcttcaagt tatggtaaga ctccaaaggg catggctgtt 158220 gaaaagaagg gtgctggcat ttgtcccaag ccaagcatat cccatcataa tttaatcatg 158280 - 166 - 008252 ttgatatact tagcactggc aatgggaata gatgatcaag aatattttta acatttttta ccaatacaca tattactccg cgtatttact tgttgctgtg tctcaattag aaagtgtcat ctagggctgg cagttcctca ctgcttccac tgcctagcct catattcagt gaggccatct acataaatct tccttattac tcagatcact tttcttaatt tgaacgtatg attcatcact gctatattat caccatttta tgatgaagac ggacaaaaaa agtaaaeata tatctaatat ttatttttga aattatatgt ctagacacta tgatcaaatg ataccccaga acagctatcg ttggctctat tgggtggaac accaaactct gttaacagaa cagaatttcc ttaatttttt ttaaagccta acaaaaaaaa tttaccatcc tcttccccgt attcctttat taataggtga tctgctgctg ttccttcatg aatgtgtatt aggttccata ctgctggaaa acctaaggat gtatatgcat tttcatatct gacattcttc gtcattattt tggaaattct tactctgggt gtacttttta tctattaatt cattaaaatt tgaattatct attcattaag tgaatatggg aaaattatac aaaactgaga aaagcaagta ttatatgcat gttataatgt taataaatta gaaaaagagc caactttctg aagaaaactg acttaaaaaa tattttgttt aaatcataac ccattaatat agttgagctt gttttcaaca cccacacatt ttgggaatct ttgagatgtt tccaagatgg ctctccagag atctaggaga acacaattat actacacaaa ctaccacatg attgactatt ctaataagca attatcacac gtaagttggt catgtatttg tttgcattta ttttgatact tgagtttgtt ttaaacaagt accatttgct aggctatcga aactttctat aatgggcaga /-r+- f nf f f пла atcatcatca tagtctgggt tatgtatgaa agaaataggt ctatagcttc agaccctttg caccctcccc tagaatgtta gaaactttcc tatttttaac tcatgatcag agagggtatc agattcctac caggcaaatg tgtggcagca aaaatacttt gaaagtgacc ttagtccatg tatgcacaga aatttaaaat taaccatata aaacttgtat gttcattttt aattttttgt gaagaagcaa aagttttaat ttttaatcac actggttatc tgaaaactcc tattcatggt aactcgttga tagccattaa accttcatgc aaaagaaaaa attcaaagtt tattttattt attttttaaa ttctaaagta ctaccatata ccataaaaat agaagcctac gagacaggct gtctacagct gtcactccag ggctgcttga gaccaagggt ttctgcaacc ggcatgaacc ggaatttgta catgcaaagt cctgtgtata aagtgggatt ttcataaagc tgataaatac atgaattata tatggcatgt tgttgctgtt aagtagtgtg tataggcaat aggtggtaaa tactctataa gtatagttaa taaaaatact gcccatagtg aacagacggg gaatgctgtg tgttgcatgg agaagaagag aaaactgaga taatgttgac atttggaatt aaagcaaaaa gttaggcatt aacaaggaaa ttggagcata ggcatcttcc gtgatagatt gtgtggcttc agccatataa ctttacaaat tatgtactat atccaatttt attccagccc gtagcaattt ttcaacattt catctttata caataaaaag aaactatatg ttgctagata caattgcact aatcataacg tggactgaaa acctgaaatt cttctgcata ttaaccacct tatgagttag gaagagggac acctgttcta agaggaaaaa cctccttcaa tatatatata tcaccagtgt caggaaatag tagctgagtg gcagtcacct tggctgttgg gcatcctcac aaagacacag tgctgatgac aggagtctag atggctgcat tggatgttat ttaagctttt gttgagttca aagttaacat cttgaaatgc tactcaatat gggttttttc attttggaca tgggggattt attaaaaatt atggtgttaa atatcctagg ttggatttgg CtCuuCfCjCCu acaatcaacc agtataagaa atgtcatcat ttcttaatgg aggatatttg ctcgcaagag gaatagttga agaaccacac ataagcaggc gtataaccta tcattttatt aaaaaaaaca ccaagaaagg cggctctgct cttcaaatga tagttgcaga ttagacatga ctggtattga cctctgagac ttttagctag tcatttgtaa tttcaggatt taacttattt tctagtggtc tcaggacttt gcccttcact ttagtgaagc agcctacaat cagcaattcc ctatgtatct cattatagca ataacttctc aatattaaca aaatgaaaaa tacagcgtaa aaagaagcat ttgtattact gcattttctc tatatatact attaggtatc atgtttatta tttctagttc tgaagtttta caggaggcct aacatagcac tgtattttat acaggccaga aacaacgctg gtatcttatc ttctaatatt catcaatagt aaaaaacctt attatttcaa ctatgcaatg attttaggta attgttattt acttagaact tttttaaaaa tgagcttcaa gtttaacttt aatagtttta agactgttct "-" W l_ _> V.vjwWv. tgcctggtct acagaataaa ggtgaaagtt caggaggatt gcttgaatat agtgggacaa gatgttaact ctgttataat cctccatatt tttaaatttt actcttacct gaacaaacaa ttttgagttc agagatgatt gagttgggac aaaaaatatc tcaataaaaa gctgtcactt agtatgttac tgctaccgcc aataattttt gtgaattcaa atatatacac acaataggca aattggcatt ttgggtaaat tttgtgtttc ttggaccata atttttgtta attaaattct aatattatta LLi.wLwwwyy gaggttgcag agtgcctctt ccacatggac agagcaggtg cataagtcac gtggaaggga tgaagagtag ctataactgt aaacaaccct tttttagaat tcctttttaa cattttattt gcatgtttat acatataata tagatcctcc ctaaagaagc ccaggggcaa aaatgtaata taacaaggga ataaaagtaa 4- "- a a = +¦ r"-i гыч i- LaaaLvjoovj aatagtcttt aaggacaatt tggacccagt gaatgaaact tcaatcattc caaagagtac gttcttgtat acccaactgt ccatggctct caaagctctc ttaatctatc tgcttttaac attgttatga tacagctacc cttttcactc gccatcaatg attcaagatt tagaaccact ccatggttat ctatagataa tgcttttaaa gcaacaaaat actttgcagt agcctactta gacccaaagt ctgggctata ataaaatggc gtatatcaga atttgggcat cactattctt tcatacttga - 167 - 008252 aactaagtta aagtgttcct ggttaacttt ctctcatttt tgtatgatta tgttttgttt 162120 tcagtagttt ttaacaacgg tgtcattttt tatgcccctg atttttctta ttttcagtga 162180 tcttggcctt taaggtaggg ctcatttgca tatttttaaa aacacacaca ttctcatgca 162240 ttcactttca tatacacatg tggtcatgtg gatacctgaa actttagact gtcttttaca 162300 gattctttta aaaaacaatg aaattggtac cctaaatatt tgtttcaatt gtccatgtac 162360 aaatccagtc cttaagattt aaacttatgt actctatttt tactactccc agatagagag 162420 tagagtggag gaaatgttca aaaaaagtat ttgattttct taatgtttca agtttcagaa 162480 agcgctatgc attccaaatt ccatctttgc tccgttcaaa ctttttgctt cttttaccac 162540 atctgcttgt atgtagattc tcatagctat ggtgacattg gttgaattct tgctcagtat 162600 caagtgtttt atgttttcac ttatttttca caacgaccca ttcaatgcag gtgttattat 162660 tatgtccatc ttacagatga ggaaactaag acatagagag gttaagtaac ttgctcaagg 162720 ttacacagcc aacaaatgtc ggagctagaa tttgaaccta ggcaatttgg accgaaagcc 162780 cacactctta acctctaaac tgtattgatt caaactaaag ctatataaag gccaaacatg 162840 agtcatgcat actggggagg gcactcatag gactatgaat tatttgttta taaataatga 162900 tgttgtagat aacaaaggca tcctcatatt caagtaactg ttagaatgaa ggacagtttc 162960 tgagaatagc ggtgaatgca gcaacccaat cgttaaaact attcacatta gttctttgtt 163020 tcctctcaaa actcagagga tctaatcaga gccactggag ttacagtctt gctgcgcgat 163080 ggcaaacgcc actcaaggtg ccactttgca natgatcttc ttggagtgat gatcctcctc 163140 agacgcctac ataacaggct tgcagcatag cagcatttta gtgactttaa tgtcttttat 163200 tatcgatatg tttttaactg tgttaatatg taaaagttgg ttcatagatt tttttcaaga 163260 gtaataaaaa ccagggcctg ttcgctctaa aatgggggtg ttggccaggc gcggtggctc 163320 acacctgtaa tcccagcact tggggaggct gaggtgggcg gatcacgagg tcaggagatc 163380 gagaccatcc tggctaacac ggtgaaatcc cgtctctact aaaaatacga aaaacaaaaa 163440 aattagctgc gcgtggtggc gggcacctgt agtcccagct acttgggagg ctgaggcagg 163500 agaatggcgt aaacccggga ggcggagctt gcagtgagcc aagatggcgc cactgtactc 163560 cagcctgggt gacagagcga gactccgtct caaataataa taacaataaa tgggggtgtt 163620 aacatgcaac actcagtagg gctcaacaga ggaagaggaa gcagccctca tacaggggag 163680 aggagaggcc gcccttaagc taagggtgtg agatggtggt agaaagtgaa aaggtactag 163740 tgaagaaatt ataatggaaa agaaggaaga agtgaagtag gaaaacacca gatatgaaat 163800 aaagagtatg actataactc agggataaag gtgcaggaat gtggccaaat ccagagttag 163860 gaatgtgagc aagtcagtgg ccaaaagtca aagcccacca gggcgttccc ctcagaagct 163920 -.-.-,4-__-,____4-____4--__4-__4--_ .1...41.-Ы.. -.4-4- "4- -,-.4- 1 CWOn Ч"1_-1~ L.y CIVjyy V- CIVJVJwCl tyyyU UVJVyOV.Vjai.VjVj Ct V- L,y L.V~VJVJ L.VJ UVJUV.UVJLVJLU V~ V, V~.V~ L.Vjy СД CLVJ J.VIJ3UU agaagcatgg cgctgtatgt cttttcccat tatttttcta tgcaacctcc gtgtgcatca 164040 gaggtggttt ctagccaagg actgctgagc tctaggcgtc ctcatgagaa gctatggtgt 164100 catttttaat acttcaggaa gctggaagag aattgtgcat tcacttaata taagcccgca 164160 cctgctaaat aaaatgtcaa gtttaatact tttagctgga aacctgaaaa ttctaagtgg 164220 cagcactgtt tccttaatac tatagttctg aagtgttttc atatatagtt tcctttaagc 164280 aaatttgttt aaagctacaa tgaatttctt attgcttaat cgaggcattt tggcaggaaa 164 340 gaagtttggg ttttagaatc cagaggagaa agtaaaagga ctttttgcac agaaattttg 164400 agaggctctg gtgtgtttat tcccatcacc gcctctccaa gacatcacca gtgtgtgccg 164 4 60 tgatggtcac ctctatggcn ttttcctttg tcacttccca caatcatgtt ccaaaaactc 164520 acctcaatca tataaccata tgaaattcct ctattgcctt ccctgtgtgt tatatatatg 164580 acacagctga ttgctaatag caatagggca gcgtaatctt ttcatagctg ccatcattta 164640 catgaccccc atattgaagt acgtagaaac catatttctc ttttcagcat ccccaaaagc 164700 caatacagag acctcggaaa aaaatatata tgtgtgtgtg tgtatatcta ttattacata 164 760 tatgtaatat atgtaacata tatgtatatg tcatacatta aatacaaaaa tatattataa 164820 ttttgtatta catatatgta atatctatac atatatgtat atattaaata taaagattat 164880 atttataaat atatgtaata catgacataa attatacata atgtgtgtaa tatatgtaac 164 940 agtatataaa aattatattt tttatattta gtatttatac acacatatat ataaaacatt 165000 gtttctgcat gacatttaaa aattggctaa aaatgagatt gtggtgagct gaggagattc 165060 ctgataaatc agttggtata gaggctacaa caaaaccctt gacctaaaac ttctctgaac 165120 agaagagttg atgactgtga agctcaagtg atgcaggaat gacactgggg aatgtacaca 165180 gagttttgca aggtttgaat atgaagagaa ttcttgcagc tttaatgatt tttgcaaatt 165240 tggtgaaaga cagatataaa gggttttctt ttaagtggta taaaattcat taagctttga 165300 gggaagaaaa aacccacaga aaagcctgag gggaggaatc atcgaaaagg gaagtatagg 165360 tagtctgttt agattaaatt cagagctgtg aaattgaaaa ctgtttaccc agccctgtat 1654 20 ccagtctcac ttgaatcctg gttccaattt aatgatgttt ggtctttgct agagtccact 165480 gcatagtgtt ggatagatat tatatattaa gaagctttcc tttcttattg actgtgactt 165540 ttttaaacaa aaaccataaa tatcttatat tctcttttgt taaagtatga gctctaggtg 165600 aatgtttttc tttagcctta agctgcatga tagttgattt tatgtgtcaa cttgactggg 165660 ctaagggatg cccagatagc tggtaaaaca ttgtttctgg gtgcatctgt cgggtggttc 165720 tggaaagatt tgcatttgaa ttggtagcct gagtaaagca gacctccttc cccaatgtcg 165780 ttaaaccact tccaaagctt tgaggtccca gatagaacaa aaaggtggag gaaagctgaa 165840 - 168 - 008252 tttgctgtct gtctcattgc ctgcaccatg taaacaacct tctcctgccc tgaggaatcc 165900 tggttcttag gccttcaaaa tgttgccctg gcttccaaag gtctccagct tgaagagagc 165960 agatagtggg acttatcatc agcctccata attacatgag ccaatacctt aaaataaata 166020 ctgatgtata ttttattggt cctgtttctc tggagaatct tgcctaacac acctgaagct 166080 ctcattkgca tgtccatttt gtcgcctata ttgagttcat tccagattga gggcggtagc 166140 aaggtggcag ggatactgct ggaagatgcc attcggctga gcactggttg catctcctga 166200 tgttagttga atcttcaatc ttctcagcat tttgtcagtc tctcatgaat gaactcaaac 166260 taatgtaatt ttaaaagact gatataactg ctgcaggagc aaatgctatc tattgagaac 166320 taccttgtgt aggctrcttt atattaaatt atcacattta atcctaaata ttaaagacgt 166380 ggcaactaac attcagggga gtgaatttct cacccaagtc cacacaattc atatgtgcct 166440 gggtcagagc cacaaaggct gtctgaccct aaaccaatgt ecttgctgaa aaaggccatg 166500 ctgcctgcag agaaccagct ccttcattta taatgaaaag catatctagg tgtggattgg 166560 atgagatgct tatccatttt ttagatcagc atattttctc cctgtgggta ttcaaagaag 166620 aagccaaatt ataatcagac tatagcttag tgagccagga atcaggatgg agagggaggc 166680 gagaaaggtt gtgtgattta aatacataga tgatggtttg atagatgcag caaaccacca 166740 tggcacatgt aagcctgtgt aacaaacctg cacattctgc acatgtatcc cagtatttaa 166800 agtaaaatct aaaaataaat aaaaatattt ttaaaaatac ataatggcat aggctaagat 166860 aggctgagat caactgagtt atctctttta cagttgagat attagtgacc tagaaactaa 166920 atctactcat tctgtgaaac tggggtctgg acactcatct actatgctat agatattgtg 166980 ttggggctag ggacatgtga tggtcaggac actgtttcta ccctcaaaaa taactcctga 167040 gccagtggga gaaaaagaca agtaaacagg cagtttgggt tcagtggtga tgattataag 167100 gataatgttt cggaagcaca cagaaaacat ctaaagcaga ctgtgaaatc agggacgttt 167160 tctcacaagc aggtgcagac tgaaatctgg aggttgagta gtgctaatcc aggtgaagag 167220 ggttggagga agatggcatg aaggagggta gtgaagacca agggaactgc agagtctttt 167280 ttaggtggga aaaaaacaga gtggctttct cgacactaag agtttgctgt ggctaatgtg 167340 tagagcaaga gagaaaattg ttaactggaa aaataaggag ataccagcac aggaaatgct 167400 tgagcagcac taagcatttt gatattattc tgatggtatt gatgccacag ggtgtcataa 167460 tcatacctac attttggtgt cagtgtgaaa tgcactggag ggcacccaga ctggagatgg 167520 aagaatccgt tcttgaggca agtggtgatg gcagcctgaa ccaaaggaaa taggaatgag 167580 agaatatcga aagcggtgga gtacagagaa gaggatttag tgacaaattg gatgtaggaa 167 640 gtgagaaaaa ccaggacaca gggatgtcac ccagatttct agatttcttt aagtgcgctg 167700 atgcagaaga aatacaagag aaagaagaaa tgagagtttt gtttatttgg gttttttttc 167760 gggggaagaa gtttggtcat attttccctg agatgcctgg gaaatatcta tgagacaatg 167820 tctatggaga gatacaaagt ttaaagctta ggtggaatat ctaggctgaa atagatactg 167880 aaaaagagca aaatcaacat gcagatggta atttaagcct ttagagagca ggaacttacc 167940 aagggaagga tggttcccta aggggtaata aataaatggt atgaaccagt gaggaggagc 168000 caaaatgtgt atgagaagtc acagcacaag aggtaagagg aaagcctaga atgtgtttgt 168060 caccaagatc aaaaggtgaa agagtttcaa gagagatcct tgttcaacga tctcaaagac 168120 ttcagagagg tcaaaaaaat agtcaccatg gaaagcaatc ccatttacag tagcaccaaa 168180 aagaataaaa tacttggaat aaacttagcc aaggaggcaa agacacatac cctgaaaact 168240 ataaaatact gaggaaagaa cttaagccac aaataaagat gaattctgac ttacgatgat 168300 tcgacttaac agtttttgtt tttgtttttt tttgacagag tctcaacctt ttccttaggc 168360 tggagtgcag tagcatgatc tttggctcac tgcaacctct gcctcccagg ttcaagtgat 168420 tctcctgcct cagtctacca agtagctggg attacaggca tgcaccacca aatccaacta 168480 atttttgtat tttcagtaga gacagggttt caccatgttg gccagggctg gtctcgaact 168540 cctgacctca agtgactcac ctgcctcagc ctcccaaagt gctgggatta caggcgtgaa 168600 ccacctaacc tggcaactta caattttctg attttatgat ggtgcaaaac agatacacgc 168660 tcagtatgct cctcaactta tgatggaatt atatccagaa acactcatag taacttgaaa 168720 atatcataag ttaaaaaatg cacttttaca gtattttcaa cttataatga atttataaac 168780 cctattgtaa gttgaggaac atctgcaagt gaaaagacat tctgtgttga agaactggaa 168840 ggcttaatat tgttaaaatg ttcataccat ccaaagcaat ctacagattc aatgcaatct 168900 ttcgtaaaat tccaatgacg ctttttgcag atatagaaaa gaaatcttaa aattcatacg 168960 aaatcttaag ggataccaaa tagccaaaac aattctgaaa aagaacaaac ttgaagatgt 169020 catacttcct gatttcaaaa catattacaa agataaaata atcaaaacag tgtgatactg 169080 gcataaaggc acacatatag accagtggaa cagaatagag aggccacaag taaaccctca 169140 catatatagt cacatgatct tcaacagtgg tgtcaagacc acactatggg gaaaggacag 169200 tctcttcaac aaatgatgtt gaaaaattgg ataccacaca caaaagaatg aagtttggcc 169260 cttaccttat atcatatata aaaattagct caaaatggat taaggacctt aacaaaagac 169320 ctaaaaccat aaagctccta gaagaaaaca aggcaaaagc ttcatgacat tggaattgtc 169380 aatgatttat tggatatggc acagacagca aaagcaaaaa taggagactg cattaaactt 169440 aaaaacactt gcacgtcaaa ggaaacaatc aatagagtga aaagccaacc tatgggattg 169500 tagaaaatat ttgcaaatcg tgtgtctgat gaggagttaa tattgagaat atgaaagagc 169560 tcctatagtc aacaagaaaa atacaaataa tctgttttaa aaataggcaa aatacttaag 169620 - 169 - 008252 ctgacattta actaatcatt aatggccact tactttgttg tcaaaaaatt aaagaaatga ttcacaataa aaatgtggta cacatgctac aaaagacaaa gaaacagaag tcaatgggta caatgtgagt gcgtggtggc gagcccagga acaaaaaatt gccaaggtga ccattacatt aaaatgatca tttaaaactg agggccttcg agtgaaggtg acctggttgc tacccaacct tcaataggta agggaacagc gctgagagtt gaagccattc ccataacttt ttctgaaata tttataataa tctaaaaaag agagaaatgc atcaaaaaaa atgagaatgt aaaagtagca aaatagaatc cagaaggtgg tctacacgca acatggatga ttctgtatgg acagaatgag cagagtttcc atgtttaaca tcacgcctgt atttgagatc agcctagccg aaggattgct tcggcctggg agatggcaaa atcatattaa atgatacaga gatcaccaac tggctgattc tttgccattg ggaagcaaaa tgcccaggag gaaaaactct tttcaatctt ccatgatgta gctacttact attctctgtc gtatacaaat aaatagaaac aaancagttt aaaatggtgc tgatcatatg tcaaagacat aagcaaccct aggaaatatc accttgagga tttctcttat agttgccagg actttgaaag ctgctgaact aatcccaaaa agcctgcgca gacatggtgg tgagcccaag caagagtgag atttatgttg caataaggag gctgggttgc cttttcctca ttaattttgt tggtatgcat gcagctggaa cgagaggacc gcatttcact ttggtcagag aatctcaaaa tttataataa acactaagac ggccaagtag cacaatgaga ggcaagaatg tgctgctgcg ataaaatata atttgcacac agtgtccacc attcagcctt catcatgcta gtgagtttat ggctagggag atgaaaaagt gtacacttaa ctttaggagg acgtggcaac cgcatgcctg agatcaaggc accctgtctc tatgtttttt gtggttagtg agaggatagg aacgcctcat tagatatgtt agagaatgat atatcagata gtaccctgaa caaatctgtt atgactcttt gatcagctta attcttactt ataaacagca atgaaaagat taccacttta tgaaaaaatt gaaaacagta cttctggata tcatgtteat agtggatgaa aaacatgaat agtgaaataa ctaaagtagt gggagtaaac tctggagatc aatggttaag ccgaggtaag accctgtctc tagtcacagc ttcagtgagc aaataacaac accacaatta aaggcaataa gaatcaatga actctgtttc aacaccattt atttgtacaa tcaaggattc gaacctataa gatgetaect tgaggtcagt tagagcattt ttataataaa ataaaattgg gttcaacatc tacctattag gcaacatttg tggcagttcc tgtatatcca tgcagtattt tagataaaca aaaactctga gccagtcaca caaattcaga ccattgttgt tgttgtacaa atggggctag cacattgttt tactaaaaat tacgtaggtg tgtgatcatg aacaacgaca atttctttta gagcaagctc egggtaagga cagatagacc gctacagtgc caegctgaag ctgatgctcc atcattagtg aactccactt ttattattgc gatctgetat tgttagactt tttgaagaaa ctcagaattt attcaaaaag agaccaacct tgtggtggtc agectgggag acagagaaag ggatttttac tacctgttag tggaagcect atgcaatcag aaagagaaga gaccagagtg aaagtcccat ttgatctcag cagttttttg tctctggact tggggcagca gagcagcttt aaccccatca gcccttccaa gaccttttct accatggaac gttgtcggtt acagcactct gactcacacc ggagtttgag attagecagg gaatcacttg catctcaaca ttggagggaa cacccactgc gggaatatca tagaaatatg gggcaatata catacctgtg gtcaaggctg accctgtccc agtcaaataa aattgatgtt acgcctcagt gttaagatga gacacagaca atgtggtcca ggagtagacc acttccagcc gaactttgtg caagaagaaa gggaacaatg ctcagcagtt tagtaaaatt aatttcatgt ttgcctcatc aaacatgaaa gtatcattct gcaaggtaac tgtaatccca actagectgg cgtagtggcg aacccaggag agagtgaaac gatgtgccca ttaactgtaa attatcaaca ggaggccaag gtgagaccct gtcccagctg cagtgagctg cccaaccccc gtataggtgg ctcctgatat ccctgtgaat ggtcattagg cagacacaca caaggaacac cccctcagaa cccagaaata gcagcggccc cgatcccttc gacgtaccct ggagttattt ttatagactt catgtgtgat catgggtact ctacacacct ttaattgtgc ccttcaatta gcactttggg ccaatatggt ggcgcctgta gcagaggttg tccatctcaa aacaggagag ggatgetatt taaaaacaac gtgggaggat catctctaca cttgtgaggc tgattgtgcc caagaaagat gagattcaaa gagtggaatt gtaacttgat gtggactcaa gagagagcac ecaggattge cctccagaag tgagacaata tagacaacta taaagctggc tggaatccta tcttctgcct gagactttct taactcaaac cgggactgtt aagattgtcc taattcagaa aaatatatat aagecgagge gaaaccctat gtcctagcta cagtgagctg aaaaataaat aagctcacac ttattttgtt agectaggaa cgcttgagcc aaaaataaaa tgaggtagga actgeattet aaagatagaa aagtcagttc ctgtccctca tggaaatagg tatgactgtt tgtgtgttga tggcatcaca gagcttgect catttctgtt acacacctcc aagtagcctg cgccagcctg ttctttgatt tagaggaaag cagaaggaca ttgttagcat actcgagtct ttattttcat atatataggc aggtggatca ctctactaaa cttgggagac agatcgegtc aaaataaaat ttcaactcca tcctttaatn tctagaagtc cagtagttgg attategcag gaatcacttg agectgggea acagccaact aaaacagaag aataggtacg gtctttgtag gtcctcataa tgaagecaca agcagctgga tgetgacate ttcagccacc agagecttgt gctgagagca cgccaccctg ataggaatca gtttctgtaa ttgttttaga gatctctctt acaatggtgg tctagtttcc caggcatggt cctgtggtca aatacaaaaa tgagacagga tgegaatetc atagtgggat actgaacacn ncaagagnnn - 170 - 008252 nacaaanatt gttnaagana gcatgnagct tactgcactg taccattgaa aacagcaagg 1734 60 atgataagga agagcaacct cgcagcactt tttctttcag atcctgtcac aggcagtggg 173520 aaactagatt gcaatggttc taagagaaaa gcatgacagt tttcagtttt atgcggtgcc 173580 tggcagttgt tgaattacaa aaaggcactc ttatagttgt gttaagattt gttgtgagga 173640 gttagaatag tgaggtttac cctgtgtgga cctacacatt aaataggaac atgctggagc 173700 tggagtcatt gccctgattc cttttggact caactttcac acagtccgtt tggatctttt 173760 ttcaagtaat tcatgtactt ttccgttctt ctggaagcat cctcacaatc tgctttatca 173820 tccattcttc aaatatttga gtacctacca tgtgtacaac gctaggccag gtacaataat 173880 aagtaagttc tttgctccca tggagccttc tggtctaata ggggagttgg atgtaaatca 173940 agtaattaaa ataattccta actgtgtaaa gattgagatg agctttcttc agaaaataac 174000 atcatttaag gagagacact cgtagaagac cccgacacaa atggtggtca ggggtgagga 174060 tgaagaaagt tacaggagag ctaagttttg caagatgagc cttgtggact tcacggaggt 174120 gtggtgggtc aggggagggg agcacaactc cagggagaag ggaaagcctc tgtgcaccca 174180 gaatgcagga agaccagggt ggagtggtat agagtaagtg tggaaaagtc agcagggcat 174240 gaaccatgca gtgccttgag aaacacggta aggtcattta ccatttgttt tttgtttgtt 174300 tgtttgtttt gagacagtct cactctgtca cccaggctgg agtgcagtgg tacagtcttg 174360 gctcactgca acctctacct ccaaggttca agcgattctc ctgcctcagc ctcccgagta 174420 gctgggatta caggcgtgcg ccaccactcc tggctaattt ttatattttt agaagagacg 174480 gggtttcacc atgttggcca ggctggtctt gaactcctga cctcaggtga tccgcctgcc 174540 ttggcctccc aaagtgctgg gattacaggc atgagccacc ataccagcca gaacatttac 1746O0 ctttaatgtt tacctttaac ctgagccact gaaagcttat aagatgggag gcagtggtga 174660 accagagagc tgtttggaaa ataaaatcaa gagggacttg gtgataggtt ggatttaaag 174720 acatgagtga gagagtctca aagatgattc ctggtttcag gtttctacag ttgcatgaat 174780 gaaatgtcat taactgagtn aagaaacact agcatacaac cgtatttggg ggaaacgttc 174840 atcaactcag ccttgggcat gttgagttgg tgaatctttc aagatgtctg aacngagatg 174900 cctaatagtt gtgtataggt ctggagttca ggggagaagg atggctgcct agagatataa 174960 atgtggacat tcaatgtaat tctcaattat ttattcaggt gctgttctag acatgtggcg 175020 taggtcaatg aacagaacag ncaaaaatga ctgcctcctt tgagcttcag tcctagtaaa 175080 gttctccatg ttttcataat actggaaggt gtaaatctgg atgagattgt atagggtggg 175140 aagaaaaaat atgctggagc cttgaggaag agcaacattt aataactgct ggtgtttgcc 175200 tttatggtaa ctcaacctgt ctctgttttc ctccttgtgt attttcactt tcagttccat 175260 tgcaaatagt caacattttc tgtatctcag gttcaaaact tgatcaataa gaacagattt 175320 ggtcgatcca gctaatcaat atgattcctt ttaggcagga atttcatctc tgattgcctc 175380 gcaatgctgg ccagtcttat agaaagtccc tgtctttggc ccggtttctg gctcaaccag 175440 ctttgacaag ggtgatgagt tactttgtac cataaggcaa tttatnagga agaaacacct 175500 tcataaaaaa aaacaaaaac aaaacaaaac aaaaaacaga acaggcagac atttggagaa 175560 ctcacctgca tattacagag agatttaata tgtttatcta ttgcatgcca ctgtaattta 175620 acagtctagg caaaggtaac agaaaagagg agctgagata atccagctcc tccccaacca 175680 ctcattcagg aggaagcaga tgtgagtgcc attgtagggt cctgagggag tattggttcc 175740 catttcctag cctgcccctc ctccccacgc ttggtcagtt acaccttcat atggctcctg 175800 ttctcacttt acctctctgc tttatgataa gcgcagataa aaggcaagag acacagacaa 175860 agcgaaaaag aaggaagagg gaaggaaatt tagaagggac cttgcaaaca ttttccttcc 175920 actgagtggt ccacaggagg aaacaatgaa aacttctctc tcctttggga ggccaaggcg 175980 ggcagatcac aaggtcagga gatcgagacc atcctggcta acatggtgaa accctgtctc 176040 tactaaaaat acaaaaaatt agtcaggcat ggtggtgtgt acctgtagtc ccagctactc 176100 aggaggctga ggcagtagaa tcgcttgaac ccaggaggca gaggttgcag tgagccgaga 176160 tcgcaccgct gcacttcagc ctgggcgaca gaatgagact ccgtctcaaa ataaaataaa 176220 ataaaaataa caaaaaataa caaaaaacaa aaaaanacct tctctctcac taacaaagag 176280 atgtgatttt ggagaaatgt cttccctctc tttcagtttt accaaatcta acttgggtat 176340 ctttagatcc ggtgtgccct cttttctttg atgacacgag tcctaactcc tatatattgt 176400 cttatatgat tgttttatgt catccaagct ggcacttata tcatctccct ggtctcatag 1764 60 gcatttaaat ctgggcccct gcactagacc ctgacaagat ctcttctaga tctaaaattc 176520 tcatccggga aagagtcgag agctgtcatt actgggacac ctgtagctgt atacttcatt 176580 atgacagatc tgttgtttat gtccagtgtc tcttctgatt ttggggtgtt catgctgcat 176640 ggttttctaa tatacgtagt tctaatatgt tctctgttca ttactgcgaa tttctgctcc 176700 gacccctggc cacctacaca aaccgcaatc cccagcatag gcaacatgtt agtaaccacc 176760 cggagggggc gactgcaagc catgatgaag agctgggtct accatgtacc tgacctgtgc 17 6820 tgcaagctct ggcactagag ctggggtcac agggccctag ataagtaatc tcctctgaga 176880 atagtcagat tttgtcaata catacaattt tctcttagca gaaatcttgg ttgtattaat 176940 aaatgctgtc attgacagca ttgctttgta tttgtacatg gatcaaatga aagaataaaa 177000 gtcaggaaag tgatgatatt cattccttgc catttactta atatgccatc aaactttccc 177060 actttcctat tattttttca ctctgggagt ttaggaaagg gaattatatg actacctgga 177120 ttctcatatg ccttcaacat atttttaact ggatcatttc atcttactct ggatttgtgc 177180 - 171 - 008252 ctaaaatctc agtgctaagt gctctatctt actggtggtc ctcttattcc tgattatctt gagagtgctg tgggaggaaa gttgtggtag attattcctc ctttgccaaa gcgcaagaga ggagtaccag tcaaacagaa agagactggc ctctactcag ttcagccact agattacagc atcttccttc tgaagaatgg tgcagatggg gacagttctg aaagtcagag gccagccaga actgtcaaga agaatatttt ctaagattaa ccttctctgc cacctcagac agtgcagagg gaggaaggtt ggggaggcag agggctgcac caggagagga ttaaaggaag gacctaagga ttggctgcgg aagaccagag taaaactctc atgttaaccc tatcaacaaa ggacataagt gaagggaggt cagtggtttc ctaagataat ccaactttgg aggagctaca cccaaaagaa ttggttgctc accctgtaat ctcagcactt caagagtttg agaccagcct ggccaacgtg aattagctgg gtgtggtggc gcctgcctat agaattgctt gaacccagga ggcagaggtc cagctctggg caacagagca agactccatc ccctccctag ctcaaaacca tatgtcatac gtttgaatca gatatgagac tagactgcac caaaaagtta gggaattaac tagggataac actttcctat aattgaggat gactgatttc agcattactc agtgagaggt agggcatccc cattctgtca ccgtgaaatt tccagtcctc gccagccatt tcttcaaaga gtgctgattt aaancctagc actaggtttc tcattgtttc caagaagagc agggaaatac atgtatgtgt agctagtttt gtatatattt acctatctat +* /"?/"•/••+- ¦¦-(-# fi 4~ __ -i /"•¦• "-"^ "-.¦¦- #-> ^" гчгч' \, v-J V. V. t-j U " C-\-.U> U^i-CIV-> -.(-> gacatgagtg cagtatatac atgttatttt ttaataaaat atgtggatta atgtgaattt acataggaga catttaaaaa ctattcattt gaaatgcatg attgtggcca gaaaagaaat caagacccca atgttaagac tgcattccaa gtgtcatatt tatttgatag taaataccat gcaacaaata agggaatttt ctatagaaaa gaagttataa atgagcagag gggagatgcc tatcagtaag aactcagctt attttaagag tatatatata cctgcataca tatggttgga atatacaaaa ctaggtaaat gtgagagagt atttccccgc tctttctgct gagggagcct aacctagtgc tgggattttg gtccctaaat tgaagaaatg gctggcttta gagctgaatt ttgtggtgcc agaaaataaa gaagacctaa acaaacaaaa tagatagaag tatgtctgaa gtggccaaag caggaagact ttgagcaatg ccaaagaata caataaatag tcacaaatac aaataaagga ggagggacaa ctgattctct aatagaaata atagaataat aaatgaagta +* rrt- a at* *¦ n r*r* алапалаагга rrr"+- ла rra at +¦ ctaaattgag cccagctcac aatcatcagt ggacttccag tcttctatct aagctgagtt ggtagagtga gtacagctag tgctgggctg ggagctcagc attccacctt actcccccag agcagtgttt agaatcactt ggaagcttgg ggttgtgaaa acctgccctt cagtagcttg ggggaacccc ggtgcccatg oyywtayciyyv- agacaagatt cttatacaaa gctgaatggg accttctgct gaagatgtgg tgggaggctg gtgaaaccct aatcccagct gtagtgagct tcaggaaaaa tgtcatgtaa tgaagtaaat aaccagtctt actgttggcc caggcacagt atcttgtgtt gttttattgg cagtatatca actaactcaa gcatgtgtct ctttaatgtt tcatagtgta gaccacagac tacttaaggg agtacgtatt .tactgataca tgagagccag acaggagaac agaggatagg gggttatgta gtgtgtacat agaagtaagg aacattctcc aaagagaaca aaaggaaaaa gactagagaa aaatgaataa atatttatac cagaataatt aacagaaaat ar"r"aaaa+*4-a waayL,4.aaya ctctttctct aaagaaatca tgtagatcca cctaactctc agcaatgaaa aggtgggtgg gtctctacta actcaggaag gagatcacgc aaaaaaaaaa tggggtgagg ttttaataac ttatcctcat ttcccttaca gagaggtgag ttctgtaatt agaatggtat gtgtttctag gtacacacat tacataacca *- 4- +- 4- -"4- 4- atccctattt tctaataagc atacacaaaa aataggacac ttccatgcct acgttacatg agttcatacc ctgtggggct taggtagtta agatacatac gagttagtat atatactaga aataaaacaa caagatgagg acaaacaagc gccgactgaa ggaaagtact agatacatga ccaagtaaga tgccattagg ccaaagatgn atgaaataag acggagagag gtaggagtga aaacccgtag ggaagctggc gcgtgacata tcaagaaagg acccctttct acagatgtta catgctcatt taggacgtct cacaaattat aaccaaacta cactctgacc gaaaagacag atcacaaggt aaaatacaaa ctgaggcagg cattgcactc agaccatgtc aagaagggag caaagttgac ttataatcca ttccacttgg acattcctgg cagctctaaa ttagggacca gctctctcag acatgtattt tagaaccatc V 4- -.+¦ 4- -" -.4-vV> yW> k."wwyy ccacttgttt ccttttatca atgataattg tgctggggtc ctaatttaaa cacactaagt atcagagaaa agattaggag catgcatata gtaggtaaat acatacatgt aacaatgaga gtcggcactc ataggacatt aaacaaacaa agtgccccca gggttataac ttgaataaat aatgtagaga aaactattgt rrtr at +" rr +" rra a a r*? +- а а гтпа^т aaataaaaca cagctttaac aattcttcta agaaacactg aggacagtta atggtatttg caaaattggg ttaacaacaa gaaatgcaat acctgggaat tttgcatggc atccacccac agtgtcagtt aatgtaactt acctcaatgt tcccaaaaat ggacactctg gaagaattag gtggtttcca tcccaataac ctcaaagtgt aaattctttg ggagtcagtg tataatgcag catatacaca gtataagctc caaattacct aggaattgtc ggatgagctc ttctttagtt cctctccaag atactcctcc acttgcttca aaacctggca ctgatatgac agtctaatca gactgatact acagccaaga ctggatcaga aaaagtattt tcttttttta ttcctaagag agccaatgga gacatcacct gagaagggcg tgagaaaaga cctcgaaacc gaaacttcag cgaaagtcat gaccaaggtt attattgtag ataaagcttt gtaggaaaag tacccaagta tatcatctcc tcagacaacc atcaaagtca atgtgacaat tcgtgcaaaa aattttgtag - 172 - 008252 ttttgataaa actatctttg ggatgagaat ccttgttcna tttatacttt gccaaatgct cctctttcat ataatataga atattacaaa ctcatctcat tgttctatgg caactctcat tttctatggt aagnatatgt attattcttt ttttgtgatt ctttgccagt tgattccata tgatcctctt caccattgga tatataacat gtgagtttta aagctgtaat acatctgtgt tttgatcttn ttgtttncta aatataactc ttgtaaaggc ctgcctgttg ttatcattgt gctgggtcaa aattacttca aatgcaacat agctttctgc aaaaagctgg atagaaaatc atctgaaatc tctaagaaaa atgggactcc attgtgtgca a-ca ugc uag aggtgcaaca tggtttagag aaacgagaga gtaccaattg agctctggaa ttcaggcagc ggagctaatt aaagtgttca tgagaaatat tgcaggcatg caggttagag agaatttctt aaggtatttc aagccagtta attggattca ggggggacac taagcatctc tttaactgcc caacatagca ccagagggaa ggtatacagg aaatagtgtt agaaattagc catgaatacg gaacacacac tgatgtaact ggcagacttg caataataga gttagtgttg caatttgagg aactctctgt tttgtaaaca tcctgtgttg ttgtatcaag tttatcaaga tgtcattact cttgtgctgg atgaagagaa ccttcttgtt ccacagggcc gcacacguaa tgactaaatg gaagatgtgt tataagagga ttgttttggt tgttgtcaaa tccatggacc catcagggag cagcacgcgt gatggttttg tcttcatgtg aaagcactgg ttgatagtct gttctaaaag aggagtctca aaatacgctg agaggctgga caaccttaaa ttcattttta aattggggaa cacacatatg Я Г""!" rrt- П rtn Я t----1 - - - ---. caggcatatt ttttttattt tttaaatcca tttataaatt gtaagtgtgt acgaagaatc ttcctctgaa atgactcagt tataatataa attcatcacc aatatgagca ttttctttta atgttgttgc gtgataacag aatgcaacag cacttaattg agggacatgc gctaatgtta ataggctaac aataattgaa ttagaaagtt caaggtgcac ctaaaaatga ataacaggca gtggcagtat ttccagttct tgtttcctca ctaattttta gtttactgct gaaggattta acatccctgt aagacccgct ttgcattact attggtttaa acgagatgaa actttgccaa agaggggtgg tagcaaagga caaccaagag agagggagag * п*г*япг*г*пя --,------- ggtgatagct gctatctcac gagaacactg tgtcttctta aaaaaaaaga gtgtgtgtat atctcacaat gtggagcaaa aaattctctg aatagagatg actgtctcta aaatgtgctc ctaattcatt acaaagtaac catgaaagta tctgttataa cccagtgcat tgcttggacg tgaacatttt caagaaatga aaccccactt attagtgact tctatgcact acctgaagaa tattaaggtt ccagatgccc gcaagagaaa ttgctatgtc agagacttca aagataatgt actcacaaat atagtgcaaa agaggaagat tgcataaaat caagattcac acaacagttt gccacttcta tcgatgacat gggttaaata tcacctacca taagagttaa gtgagggggg tatgccatgc ttaaggacca agggaaaaaa atttggatga gagcagcccc gaagcgatat ggcatgatca tcttgagaaa gttgagttgt cccctgagca gtgcctaatt tttcttagga aagtaaaagt tagaagctca gctatgtgac tgcataatat atattagcat gttgttattc ataagtgtat cacagaccta taagaagatt gagggacata ctaagacaga ccaattgacc actgaggcag aatggtgctg ggagaaatgg acaatccaat ctcagattag L avjyyyaaaa gagggagatg gcaggtcctg aaatatccca cttgaaagtg tcaggaaaat taattgaagc atggtaccga gcatcatatc tcctggagaa gggagcagta catagagggc ctgtacataa ttttctagat ggaaggtttc gcaaaatggg ctgagacagg tttgaagaag tattttcgtt aacgagaaga gggactcaag ttggtttgta ttacacgtct aaatccaaag ctttgttttt gaaaaattac cctgaattcc aagaaccaag aaagatatat atggaaacaa ttntgggacc agaaattatt attcaacata agagttgttg cttctgaagg aacattttat gtgctctgct ccgcccctcc aatgagtaac ggatgtcagc gccctccaat t* rra r-r-t-t-rrrra taaacatgtt aatttgactt attctgtatt ttataattac catctgtttt ctcccttgaa atatatatag gccttaattt ggttattttt ctatgcacag tagcacattt ggcatttata gggtgaagag tactgttttt atgggagtgt ctttggtgtg tatctagttg actttttttt ctgtgtctca tagaagaata aattacatta attaggtatt gttagcttag acatttaaag gcacacttca gagctcccaa gaaaacagag cagccaggat tatcacataa ttctatctgt tatarfrinnf _.M- - - w ttttgatgaa ggttgccttt ggatttcaca acagaggtat gactgtgcga aaataatagt atacagaaaa attcacataa cctatattga ttttcaaact cccttgagta tgaaagacta attataaaac acttatgcta atgtgcgtgt acatttttcc acatgaaact atcacactga gtggagtaac aacctacaaa atgggatatt ctggatttga ccttgctgca cttgtaagat gggatatcat accaacaaac cttctcttca acagcacatg aaatgtagct ggcttcagtc attcaaatta aagatcctca tgaacaaaag tctctgatgt ttgtgtgtgt ctagcactta ctaatataat gcatatttct aaaggacaga atcaatcttt aaaggacaat atagtttttt aatattatct cttaagggtt aagtggcagg acaaaacagc tacttactta cagttaatca tttatggccc gatgcgtcca ni-attaaat-a t-r t--г t-rrt* я a a acgtttgaaa tgtccacagc ggccagtgag tattaaaaac tttgggttgt ttacgcctta aatagctaaa caaagaggag acaaccaaaa aacacggact actgtacagt agaaaagaca ggatattctt caacatgtgt aaaatcacat gcaagactct gccactgtgt ggactccttt aatattctac agatttcaaa atcgctgtct gtgcccaccc tgtatttgca gatcattatt atacaaatta atgttctgaa aaagaagcaa cttctgattg agttacatca atgagaatct - 173 - 008252 cccaagtctg aaatttttat tgaaactgaa tttaagcgat tgagaagtca gcagatgctg gtttgaagtt gtgaatgtgt tgaggccatt taaaatccta aatgtttttt ttaatgtaat taatgtttga gaggcagaac gggaaagcag agccgtagaa tgtttctttg gacaaagcta ataaacttcc ggattctcta tggacattta atgatcttaa agtaaattta aaaggtagaa tgtaatactt tcccatctta gaaagacgtc aactgcatca aggtatagta ctgtgcctta gaccactctg acgtggaatg caatgttatg tttaaataaa attttgggac tgtgcacgca cctgtgctca actgctgtcc gagattcaaa taatgcacaa ctccaaaaag tgagttctaa gaagtaaatt tatctctaat actgttggta tgccatttct tgtgtccccc gggttcacgc ccacgcccag atggtctcga acaggcgtga ttcattagtt attctcttct caatagatat atttcccagt ccattattcc ctgcccgtgg tcagggaaaa agtagggggt catctttgga cagtgtttgg cgcaggccca agggaactcc cgcctggagc tccagaatgc tgacaggagt atgttcaact tgagcttgta tgaaatctag aacccagcag cagagggtgc gagctagctg aagattgaga aaattgtttt tcttcaattc atattttaac acagtatgtg aggatttcag tagcactgag attctaattt tcatttgatc tttgtactgt cactgtgttc cttttgggaa atcataccaa agttattcct cacagcaatc agaagaaagc acaggctgaa tcgtgcagag caattacacc ccgcccgatg tgtaatgata cttagacgac ttctagtttt cattcttgca aatgtgcttt tctcggttgc agccagactt tctttacatt agatttttca ttttccttca agaatcatga tactttatgt taaaagatct ccacttctta taatatatat atctttgctt aaatgcaaaa ttacatttta aagaaactta ggtcacctca tcccccacca ctttttctgt gtctgtagct tggctgatag tgccgaagta tgaaaaactt ttaactgaac taaactgaac acacccagca tccatactcc ctcccgctcc gcagcacacg tcagtgccac aacagtatgt actctgttgt cacatttctt acactgagat ttttcccact caactgctag tgacactgct gtggccaagg gcacgcacct aggagaggat atgctgcttc tctcttaata gtccttgctt ttaggaggtt ttcaagcagc cgctcatcct atgatgccag gtttgcacaa ctgttctttg tagtttcaca tacaaagagt tctatttnct tcttgttcct aaggagcgta atgcgggtga aacccatttt gttaaacaat atattcaagg cctttataat tttttttttt gcagtggcac aatcttggct cctcagcctc cctagtagct gtatttttag cagagacggg tcgtgatccg cccacctcgg ccagcctaca aaatttttat agacatcagt tcaggttaaa ggacacgcat actccacact tctctctacc cttcaggata agagtttagc ttagagttga atttgctcag ctcaccttgt agagccctgc atggttcctg gtgcaagggg tcacacaata cccccaaaat ttttgtccag ttgcagaggt aaccagttaa gaaaaggaga gggggattca tggtgacaga agcagggatg gagaatcatg agaaacaaga agcatggccc tgctggcgcc aatttctatt gtgttaagct taatgtattg tggattcaat acacaaacag caatattata tttgtcagag aaagcttctg gaatttgatg gagttcaaga gagggccggc agaggtgggg aagcaactga aggatgtgac ggcagggagg ggccaaaaaa gcatggcagc tcgcacctgt aaaattacaa agaaaggaac atagggaaga cgaagtctct tctcagccag aaagctcaag tctttaccat catgctcagc atatgagaaa tcttaaaaac a <_u ll. L. Laaa gtatagtata tttgcatcaa aatttggata caaaataaca ccctgtctac catgggttgg aattttgctt atatagaaag gaaatattga cacccaggac aacttcctcc tcccttcact cattcaaata gacagttcac gccttgtggg gctggattta gtgcatggga cctcagccat aactggccct ccttctgagt caagtngtgc atgtactata ggagtgtaaa aaacaaggtg aaatataact acagtctcgc tccacctccc gcgcctgcca gttagccagg tactgggatt tgtctaaaca gtctaatttt ttcaatcaat cttttctttt ctgatcaata atgccagcca gtttacagtc cgacagtacg agaatgagac ctggatggtc cacagaaaca gctacaagcc gaaggactct acttctagcc tggtaatgtg gaaattgcag taccatgtta ctagtggaac aagaaagggg ggttctgagg tgtggggata gcctagattg LciaL LclUclau aaattcataa atcttgcaaa taccatttca tgtgtatcat ctctttcttc aaagttttgg tcttcccttt tacatttttg gaagctcctt cagacacccg gagtacatac ttgtgaaatc cccaaatgta ttttaacaac gttcttgcag tttttagttg agaagaacaa atactcaccc ctgtagggca agcttcccgt ttcatttatt taaaatggta accaattaga tcctacatat ccaacatgtc aggctggagt cattctcctg ctaatttttt tctccttacc gccactgcgc tgtcaacgac gtgaatctaa tttatgctta tgtctgctgg ttcactgaat aataatgagc gagaactgtt ttaaggcatt aattgggtct catctataag gaaggccatg gagcatggct cttcagagag tgagagagta cctaggaaac gctttgagcc agaggaaaac aaaacaagta agggccagca agatgtttcc aggctggaca ttttgaccag L.wL.L.Uwwl.L.U' tattcattca aaggggaaag atagtggctt ccaacacaca tggtcaaaat agaatggaat tctaatatac ccctgctcag ccgccacccc ctgctggttt tgcgtctggc gtagttcact tgggcatttg gagggcttgt ccacagggct acagcaaaga tgattcagtg tctctgtgtt tcgtgaagaa gcgcaatgac actccctaga caagctttgg taaaatgtgt atattcgata tttttttgag cactgtaagc gggactacag gtttcaccgt cctcccaaag atctgtactt cgacacagga aacaaattta tatctaaaat gttccgccac gtccagtgcc tcctcaccaa tagtgtcaaa ctggatcctc gttaattgtt gattcagaga gggtgttaca gaaaggcaag ttggtttttg aatgatctag tgacccgtaa ggattcatcc tgaggaattt gaaaaagagg gaggcacaga tggaacccgg aggcctctga agtcccggct - 174 - 008252 actcaggagg ctgaggcaag aggatccctt gagcccagga gtttgaggcc agcctgggca 188580 acattgcaag atcccatatc tttaaaaaag aaaagtttat atttttgttt ttgtttgtta 188640 aaacaaaagt ttttttgttt gtttgttaaa agcagtgggt agccaggtgt ggtggtgcat 188700 gcttataatt ccagcgactc aggaaactga ggcttgaacc agaaggcaga ggttgcggtg 188760 agccaagatc gcaccactgc actccagcct gggagacaga gcaagactcc atctcaaaaa 188820 aaaaaaaaaa aaagagagag agaggaagaa aaaggcagtg gaatgccatt gaaaagtttt 188880 aagacagagc gtgagaaatg gcaataaccg cagaatgaag agcagattag aaagagctga 188940 agcagatgga gaaagcactg caggaaatgc aggaacccag tggtgcattg gccagggagg 189000 cactggtggg gatgcaggca gtgggcaggc ctagaggctt ttaggaaaga actctgtgtt 189060 ctcatttcaa tgatatgtcc caattaatat accatttgcc cttgtgtctg tttgaagaca 189120 ttgaattctc ataaaatatg cataccagtc ctgatacatc tagagccccc aagtcaactc 189180 ttctgaccac gtgtgtctat tctgctgccc gtgaagtctc gtggagtaga agtagtgcct 189240 tcattttttt tcttatttct ctctttgctc tataccttgt gtttccgagc actctgttcc 189300 tatatctgcc agagtggcca ttctatttta ttatcattgt ctctttatag aactatctac 189360 ccagttatac tgtgggctcc gtgagggcga gtgcagtgac ttatttcttt ttatatcccc 189420 tgagcttagc acagaacctg acctgtaaga tttgatttat aagtgacagt atcaatgtat 189480 aaataaatga gtcagcaaat gcatgaatta acaatacttt atttctgctc tgtcttatca 189540 cctttggcct gatatacctg ccttacactc tgatctgatc taatagatta actgcttttt 189600 ctacttctgc tctatttacc actgacaagt atttttctac tagttgtatc aacttttaga 189660 aaacaagact cttcctggta aatctcttta ctcgtgtgtg gaatcatttt tctaactgtg 189720 tacctctcac agagatgcta ctactttttg ttttgtgtct ttgggagcta attccttacc 189780 attgtcatca tcataaccat ggaactgttg agaacgtgcc attttgcaca cattactgcg 189840 ttgagcactc cagttaggaa gggaggaagg caacgaacag cagtgactcc ttctccaaat 189900 aagattacca tctaagacag aaaaaccaaa ggctacccac aaaatcaaat cagacttaca 189960 aatggctaag gtaaatgttt atgttacaca aaacaaaaac ataagagcga tttctgcagc 190020 ctaataagaa aaccagaaat caggatgtat aatctcatca tgagctcaca accaatagga 190080 tagttgagtc atccatcata ttgaaatggg aatgtttccc caaaaagaag ccctcctgtt 190140 ctccttagcc aatttgtatt tccacatata ggagggatgc attgccatct ctcttcctgt 190200 tttaaaagct ctcttggcct ctaattctta tctttccata attcctaaaa caaattatag 190260 agtaaaacca tacccaacaa ggaggtaaat gagtggatga tttttgaaac ccacccccca 190320 aatccctcca aaaaggcagc ctgaattaca tgtactcttt gaaaggcatc agtgttaaaa 190380 gatgcagttt gggaagccgg gacagaagac agggcagacg tggggaaggg tgttccactc 19044 0 tgcagaaggt cctggagaag gggagaggta aacagaagta aagagggact ccaagagacg 190500 tcggcactgg gagagaaaaa gagagcatag gaaagtatag catgaattct ggaagccaca 190560 gtccaccttt gtagtatctt agttcagcct tgttccagtt tatgatgtta aacctgagct 190620 tccctgaaac tcaatttgta gaaatataaa tatacactct taaataaatc catttgattg 190680 tcattcctca caaagatatc atatggcatt taagttcgtc gtgtccaagc atgtttgcag 190740 ccaaatggaa aactcgctgc atggaccaaa .tgtggtattt gtgttttgag cttttctgtt 190800 ttaatctgct aggcgacata accaattttc ctgccattgt ctctctttac aaatgctact 190860 acttagtatg attctgtagt taattttttt taaatttttc atggttttta aaaattttta 190920 attctagcgt taggcatgtg taaacaaatt ttattgttta aatatgcaag taatagtaaa 190980 ccattaaaaa tgtctgagat gaaaagagac ttgtctttgg gaataaaatg acataaagtt 191040 ttccttgaat ttttttctgg tttanttttt ttgttgttaa gttttgtttt tgttttttct 191100 gttaactcaa aagttttagc tcatagagca atttatggcg ggcaagagag gtatacttgg 191160 taggcctttg ctgccatcta ctggcatttc atgtaattac ttattgcttc cattctaaaa 191220 ctaattttgt gtaacgtata taatagtatt ataggttccc ccttttnttc ttctttccaa 191280 tttttcttta gttaaagcaa agtgaagcaa atgcggacac caaagaaaag ctccctttac 191340 gcctgcgaat ctttgaaaaa tttccaaaca gacctcaaat ggtgaaaatc tcaaagcttc 191400 cttcagattt tacagttcct aaaatcaggt accaaaatac ttctttaatt gcatatttca 191460 catatattgt aaaagattca tcatcttaaa tctgatttaa cccttctttt aagtatatgt 191520 gggttgggtt tctttctatt ttattgtcat ctttctacta cctctacctc tatttaaaat 191580 aaataaataa ataaacaact ttgactcttt taagcaatgt attcacagaa gagtcacttt 191640 д 13 д гтттгэ n tartr-haanr a a ~a з ~rrr*f n rhfrpz-af-rrt ft* ¦-/*•¦* r-t" r"t*rr ггпл +¦ /Т ap f" r*r" IQITDH -------~> --w, WW.- - - w--.-- - - ----- WW- - WW---- W-- - W.W - -- -- - - ---- - - - W-- -.-.WW tcgggaatgg gacatcccag gaaactggag aagctgctca gtgttgcagg gatggcaggt 191760 agacatcaat ctcagacccc aaccgtatga gactaaaatt gcctctgtgg cccttttatt 191820 ttcaagcaaa cagggaaacc acagctggcc cagtttgtgt actactacag tgtttatgga 191880 gcctaccatg tcaggttacc atatataatt tgccttccac tttttaaaag aagaaccgtg 191940 gcgtttttcc actcatactg atttgatttt ctaccacgtt tgccctccag ttaagcaaat 192000 gcataaaaga tgctttaaac acctattttt aataataaaa ggaaattctt ctagttatag 192060 ctcaagaaat tacaaacaga attacagtaa tatgttacta gctttaaaaa taattctgtg 192120 gggggttata aaagtatgag actatatatg gtgccataat atttaaaatt cgggtagaaa 192180 tatatttgca tttctaattt taaaaaacaa aattaattga atttggtaat ttgaagagac 192240 gttacttgaa ttttcgttat gctttgaaga tcagatgaac gcacttgctt ttgagtctta 192300 - 175 - 008252 aaatgtaaaa ttctctctta gtcattctta tgataaatgg acatttagag agagcaacta 192360 attcttccaa gtcttagagg gggaaaaaaa gtgttttatt accacaataa gattcaaaaa 192420 actgggctgg gtgcggtggc tcacacctgt aatcccagca ctttgggagg ccgaaccggg 192480 cagatcacct gaggtcggga gttcgagacc agcctgacca acatggagaa atcccatctc 192540 tagtaaaaat acaaaattag ctgggcgtgg tggcgcatgc ctgtaatccc agctactctg 192600 gaggctgagg caggagaatt gcttaaactc gggaggcgga ggttgtggtg agctgagatt 192660 gcaccattgc actccagcct gggcaacaag agtgaaactc cgtctcaaaa aaaaaaaaga 192720 ttcaaaaaac taccatcggt tactgagtaa tattgcttga aagttatttc aaggatattg 192780 ctttttaagc aggagtcact gagtttcagg aagctgttgc taatgaaata ttttagtcat 192840 gatctagttt tctatgatct tttatcactg ctttatcttt gctcaagcaa actcatcatt 192900 gcaaaatgct gtcacttgct cccttccact aataatgcct gatgcacata tacttcattt 192960 gcaaagggag agttttatga agcagtttat tgaccgccag caacaggaca catgttgcct 193020 cctgagaagc ctcccgacca cctcttcctc ctcctgcgac cactccaaac gctcagcaat 193080 tgaggacaac aaatacatcg accaaaaagt aagatcngct gtttcgaata accaggcttc 193140 taaaaagcat ctatgtttga gaaaagaacc gcagagttct ttagggtcct ggaatccaat 193200 ttnctcaaga gaataatgaa aanaaaatga tttcatttgt aaaagatatc atgaaaccta 193260 acatttttta ctctatgcat tgaaattttg catttgcccg tgtaagcttt tgagctgcgc 193320 cacgtggcag ccatgcgact ttgtaaacat caagagcctc ccaaatctgc attttctcan 193380 gtgcggaata aagggctggg atctccaagg ctcagtccag gtgtaatgac ctgccatctc 193440 ctgcaagtac aactgcttat gtctacttat tacatcagag aggatacgag agcatgtttc 193500 tctaaaatcg gtcgaaagaa ttgccattta ttcaatgctt agtatgtggc aggaacttag 193560 gtgcaagagc ttattcatgt tctagtttac ctatttcttt atcatttgaa aagtagaagg 193620 gaaaattttt ttcagattcc acagcaaagt atcagtaggg aaccacatat aaataaagca 193680 aatttaggta ataattttaa agaaatgtac tttttcattt attcactttt aaaaaaaata 193740 atataagggc tgtgtgccgt ggctcacccc tgtaatccca gcactttggg aggccgaggc 193800 gggtggatca cttgaggtca gaagttcgag accagcttgg ccaacatggt gtaaccccat 193860 ctctactaaa aatacaaaag tcagccgggc gtggtggtgg atgcctgtaa tcccagctac 193920 tcgggaggct gaggcaggag aatcactcga acctaggagg cagaggttgc agtgagctga 193980 gattgtgcca ctgtactcca gcctgggcaa cagagtgaga ctccatctca aaaaaaaaaa 194040 aaaaaaaaaa nangacntag aatgtattat ctaggtataa taagtaaaca tatttatata 194100 tacacacata gacatataaa tgtatatgtg tgtatatatg tatgcatagt atcaaactat 194160 atatatcagc ttggtcacat aatcaatgtc agttcagtta gaaaatgaaa gattaataat 194220 tgtcctttaa tattaaagag attaaacagt agagtttgca aaaacatcta gttcacttgc 194280 aagtattcca ctttggcatt cctagagtgt tttttattcc tttacattgt tactgccctt 194340 tttcttttgc ttatagtgaa atccatattt ctctatgatc atgtgacaat aaaatgccaa 194400 ctgtttcatc ctccaggatc ttctttctct ccgctttctt tcccttccca cagggatgaa 194460 ccctttggaa ttccctcacc tttctcttta tctcgcctcc cctttttctc ctctttcttg 194520 cctccccttt ttcctccttt ttcacctccc .ccttttctcc tcctctttct cacctcccct 194580 cccctaatgt gatactactc ccgtttacct ctcccccagc tcccaggttc tctgctgtag 194640 ttcagtcatg ttgctcttgc tgaaaatgat gatgattttc tttgcaggat gtctcttctt 194700 tttcaaggat ttccatgtgt tgattttcct tcagccatct agagatgcta ttctactatc 194760 tttgcagaat ccaagattgt aaatgacaag tctgatgcca atgtcatttt ttttttaagt 194820 aattagcctt ttggttggta gacttggttt gcttgtttct tcaggaaaac ttgtaagatt 194880 ttctgtgtat ccttggagtt cagaagcttt cttagaatat accttatatg tcttttttca 194 940 tcagaactgc tccaactaga gtcttttcag cttgccaact aagatatttg ctcagggaaa 195000 tttccattta ttttgcgttc aattgttact cttccatctg cttctctctc ccctgggatt 195060 gccatctctc tatttttgtt ctgtgcttgg tcttccagga cactaattag atttcaactg 195120 aggtcatcct ttgcttctaa atttctcgtg aaaaattatg atatggacat atgaccctta 195180 actatcctta aaggcttgca tattaccctt agctgtcctt agaggcttta cttttatctt 195240 ttatttttat ttttttagag atgagatctt gctgtgttgc ccaggctggg gtgcagtggc 195300 tattcacagg cacggccata gtgcactata gccttgaact cctgagctca agtgatcctc 195360 ctgcctcagc ctcccgagta gccgggacta taggcatggc caccatgcct ggcttttttg 195420 atttttttaa attaaactta tcacctctct tcttcacagt tataatccac tcaacactgt 195480 gcatggggca tggaagcaga aggtcatatg tcagtggata aacaagtggg aagatagtgc 195540 cttggcttca gccagtcagt ctgaaattgc agtttgcgtg gactagaaag caggttgttt 195600 tgttttgttt tgtttccaaa gcagattgta agccagaagt ctgggctgtc cttgtgacca 195660 cttatctcca ggaagaggat agagagctga tttccattat ggccaaagtc acaaaaccat 195720 gacatgcttt taaaaatcac tggtaacact accccactgg aaacacaaaa ttgaaatatg 195780 agtttatttt ctctctaaaa ggaaatgtga ttgtttctca catttcctga acacagtatt 195840 ttgatttggt gaggacaaag ataaaaagag tcttgagagg catgtgccgg gctgaggagg 195900 agtctaaaga gaatgtggcc cgcccctctg tgcgcccgcc cgaagcgaac cctccccagg 195960 tacagttgga ataacacgtt ttatcccata caacatgtct tattgctccc acttggaaag 196020 tggaatgtca aaaacatact ttttcttcat cagaggaccc tattatggga gtggcaagac 196080 - 176 - 008252 aactgctcct aagtagactt gtgttcctca tctaaagcag attgcggaga ttaagaaacc tcacagaccc tttccattaa ttcttcactc gctgtgtttc gattctcata gtaccaggag aaaatagtga gacagcaacc attaggcaag tggcaacctc tatacagttt aatcttgggg tcgtttgaga gctacctacc ttgacagggg tttttcctcc aagtgagatc ccgaagtgag actgccctgt tgtcagaacc agggctgtcc acataataat gtcagcctcc ctaacttatg tttgatttta gttattcatc accctttaga gtattactgg attgataaag catcaggtgc caaccctcat taatgtaata caaaccagac agatttctat cagtctagtt atttttttag gtcttcgaat ttattgccat cttgtaactc tgtttgttca acagcctgnt ggcttagctt atgatgctac caaatatgaa tcccaattta ggccatcaga gttactatcc aattcttttc taagacctta aaaatccctc cagcattctg cgaaagccac taccaccatt aatcccaatt tacgtgnaaa ctcagtgact acatgaacaa aaacttctga agccccaggt ccagaagggg attttctccc actttgcaga aaaatacctg ctgacccaga ctccatagaa gttcctgtga cttttccagt atattattga tgaacttgac acatccagtc ggcctgagga tgcaatagcc cagcacctcc gaacagtgaa tttctgctga tacagtaaag taactcatta tgctattctc agctcatagt aaccatgaga gaagaaagca tcgatgtttg cagtaaagtg atgttaaata aagaaaggtt tcatcagtta gagtgcaata agtttgattt ctgaagaagt acagcatttt ctcacatagc ccttaaaaga ttcattttta ctctgcagta catgtaacca aactctgtac gtcacaataa gaaatgcaaa acgctttctt aggatccacc ggatttgtca ttcaaaggat gcatttgtat ttaaaaacaa agttcccctt gcctttggaa aaaaaaacca ggtatgaaag aggaacaata cttcactatg gcctgaagac gctggggttt agtaattacc aaccatgttt tccttctccc tcctaaaagg acgaccttcc gcaattgtga ttccaagcag aattaaactc tttccagtaa aaggtcaagg tggttctgtg tgggaagcat acctcgtgcc tattttctct agcatatgat aggaaaatga acattcattc tgggctgtgg ccaatgggag cagaaccccc ccccaggcag agagaaattg gagataattc cctagcattg actgttanta ggacaaaaga tattaattac ttgttgatat aaattaggtc ataatgttct aagttactca gagagagaga gngcttttgg ccattcaaga tgggttgtat acttctctct tggaaagaaa tcaaagccac acttttacac ttcctcaagg gcccaaagga cactgttcac ttaagaaaat tcatgtcctt aaggacaaaa acacttggac ggggggnnag gcacacaaac tgttggtggg atctagaact ttataaatca aatagcaaag gtggcacata tgtagggacg aaccaaatac acnaggaagg ggatagtatt atggcacatg 4^UV-*V-" 4-F U U V-J accgtaaact agaaatacat tgctgctata acctggaacc tacaccatgg tggatgaagc tgcatgttct nggnacatca aggagatata catacctatg aaagctttga attttcagca accttgttga ttcagtgata tatgaatacg gagcgcataa ataaagtatt atcctagtca cccagccctc aaaaaggaca aaataaaact ctagagggaa tgcaagttca gatgaagcat caagctttga ttaaaaatag agaaaacaac aaaagacagg caagtggtca gaggttcctt aaagaaatca tagtttcctt ccctgtaata tctcacctgt ctgctgtctt gagtgaacct cctgtacttc gacagtaggg agttctttaa tcattggaat agtcattcaa gaattcaaca gaaatgcaca taagttcccc agggggacat ggtagaagag tcattttcaa .cacctgggct aactgtacca cttttacata tgatcaattt tttagttgct attctggtaa atatgattat gcctgatcta gagagagaat tcttttgaaa tgtatctcca ctcaaaggga agcttaaaaa taaaaagatg aatgagatac agttcaacca ttgacccagc aagacacatg aacccaaatg aatactatgc tggaaaccat cactcatagg cacactggga cctaatgtaa taacaaacct ttcgatctag atagttccca gtgcccctgg aggatttaat gcaagccttt acacagaaca tttcagcgag cagtaaagtg tgcaaattgt ttagaggctg gttctcttaa tcacctttct gggagaaaaa tgcagaaaac acagggatga ccctggtgga cagcaaacaa aaagagtgtt tttccccatc ttcttcctga agcaacaatt aagcagagtg aaagcagccc ggttgagtct ggctgatgca cctggagcca ccagcataga caatatctgc taagccttca gactatgttt caagcactta gtgagcaaga tacatgcatg ttacagaaga atatgcaaag tgattaagtg ggtttgttgt agcatatgtt tgaagtcaga aatttaacct ccttatggta atccctattg gatttactct agagtgcttg taatttccag gaacaaanga gaattttttt ggaaaagagt agaattggtg tgctttagta cgtaagattt catctcacac gttgtcagaa ccagttcttg gccagggaaa ggaagcacaa ttcattttta aaaatccccg cacttagaaa acaggcaggg gcaaattgag gtttatcggt ".-¦> -_•--.".-.- <. *" uy у L. i_a L. w l. u ctttccaggg aacaacaact acaaaagcac gtaattccca tcacctttat aaatctttaa ttcctgcctg cctttatcct ttcaaggatt tctacttgta gtgattaata gctctgttct acagcgtctc ccatttctta tgctaggatg atcagagccc ccaccctctt caatgaacag tgtttaccca tagagcaccc tgcgaactgt aaagtaaaaa agtgtgattg gtatgcaaag gtgcaggttc gagaatcaaa ctgtgcctct ctcagcaaac tcagatgtaa cctaagtctg tctacacggt tccgnttttt ataatttgtt gatgaatgaa acgaacgaac tttaaataat gacagatgac ttcctcatgt gataatggat catcatcact cagttagaat yy_-.y _yy_.y ttgtggaaga catcccatta cacatgtatg tccatcaatg agtcataaaa cattctcagc tgggaanctg cctgtcatgg atgatgagtt gcacgttgtg cagcgtgcta ctgggtacat tttattgcag atagactgga aaggatgagt aaactatcac aacaatgaga ggcaggggga aatgggtgca cacatgcacc - 177 - 008252 ctagaaccta aagtataata taaataaata aataaataaa taaataaata aataaataaa 199920 taaaacgatg cgtactataa aatcaaacca ttttttcttc ttactggtct ttgcagtaaa 199980 ttctcaagct aaaaccaaat atctgtgtgt gtgagcactc accctttcat ccaacaatca 200040 tttttgagga ccattatatg caaacagttt gctgggccct gagtaaacaa acacatcttt 200100 ctatattcaa ctaacattag agtaggaatt ccctctcaat gattattttt gcagaatatt 200160 taattgtgta accattttta ggttggaact atgtatcagc tgtttgtatt ccttagtaaa 200220 tgcgcattga gagtcaaaag taaatcagtc taggattaag aattttttat aggtaattta 200280 aaagattttt aatcgcattt tctgtatcat gcacaaaaga ggtatcaagg agttaaggcc 200340 gctgctaaat ctatcttgga aatttacatt ttgctcttct ttttaaactt tttcctcttc 200400 tgactacagt tttacttata aattgtctct tctatcaagt gtcatgctag tctcagtgtg 200460 ttgtatttgt tgctgatgat ggtgttgagt ccaagtcctg cttccttgtg ttgccacaga 200520 ccttaaattt cgaatttcta cacccataat cccagcactt tgggaggcca aggctggcgg 200580 atcacctgag ctcaggagtt cgagaccatc ctgggcaaca tggtgaaacc ccatctctac 200640 taaaaaaaag tagctgagca tggtggcaca caactatagt cccagctact cgggaggctg 200700 aggcatgaga attgcttgag cccagggggc ggaggttgca gtgagccaag atcatgccac 200760 tgcactccag cttgggcgat acagcgagac tccgtctcaa aacaaaaaaa aattttggaa 200820 tttctaaatt cacaaacagg ttaaaagcat ctgacattta ctctagaatc acaagcccac 200880 tcaccttctt ttgtctctgc agatttccaa tctttgcctt cctgtttata ttagatataa 200940 tcataaagtc attttgtatt ttacttttgt aattaaaatc tatctagcgt atatttatag 201000 ttttgtttgt ttgtttgttt gagacggagt ctcgctctgt cacccagcct ggggtgcagc 201060 ggcacgatct cggctcactg caagctccgc ctcccgggtt cacgccattc tcctgcntca 201120 gcctcctgag tagctgggac tacaggcgcc cgcaaccacg cccggctaat ttttttgtat 201180 ttttagtaga gacagggttt caccgtgtta gccaggatgg tctncnattt cctgacctcg 201240 tgatccaccc gcctcggcct cccaaagtgc tgggactaca ggcgtgagtc tccgcacccg 201300 ggcatttata gttncttatc attaagtact gtatattaaa cctatctaaa tttgtttcag 201360 tactcgaaga tattgctttt ccctcgattt ttgcacagtt tcattatttn atgtgtaaga 201420 tgttggagat ttgaccaagt gaccccaggc tactgtgtta gtggccgaaa cattgtcata 2014 80 acattcacgt gcaaaaaaaa atcagatcca ttaagtttta aattgtttag tttctaatat 201540 tacaagctgt tcctatatat caagttagtt gattcatttc aaaataagca gtccctcttt 201600 gtaattttta cgtcattcat atttcttatt ttcaaggact aactacatta aaatatataa 201660 gatagaaaaa gtaatacttt aatattatga aattaattca attttcaaat atatttcaag 201720 caccaacaat gcacagtgct agacaatgaa agaaaatgta ataaacaaag aatacattat 201780 acatatccta aggagtttac aatctattgg aatttacaga aagataaaaa gatgtttcac 201840 acagggcagg gatcaacttg tgctgggtgc ctaggagtcc aaaggaagag cgaagatttg 201900 gtgtatgcat ggcatgcaca cacgtgcaca tgcatgttta tatgaatatt ttaactccta 201960 cacatagtgt ttttacaggg ttcagaacac atgatattga aataagttac aataggaaat 202020 cttttttttt gtttgtttgt ttttttgaga cagagtctcg ctctgtggcc catgctggag 202080 tgcagtggca cgatctcagc tcactgcaag .ctctgcctcc caggttcacg ccattctcct 202140 gcctcagcct cccgagcagc tgggactaca ggcgcccgcc accatgcccg gcttattttt 202200 tctatttttt agtagagacg gggtttcacc atgttagcca ggatggtctc gatctcctga 202260 cctcgtgatc cacccacctc ggcctcccaa agtgctggga ttacaggcgt gagccaccgc 202320 acctggccta caataagaaa tcttgaccca tcatctaaaa ttctcttcac aaaattcaca 202380 ttttaagata ctgttcaaag cttcataaca tgatattata atggtattca cctttatata 202440 catccataat caacatttta atcttgctga tgctcacatt aatcaccttt taaaattttt 202500 tttctaagca taatgtcaat tccnagagaa agtttatttt atagaaacat cttgccacac 202560 acatgatctg tcgtatgcac tttgggatta attatgaatt cacgggtgca tgaaaattca 202620 aatacagtca aatgagatac caccttcgac ccatgcaatt ttaagtaatt gcttattccc 202680 aattactata ttttatttgg gagacttatt caataaagtg gctcgttaca ataattacac 202740 tacaactttc tagcagaaat gacattcatc aagccatatt cattggaccg tggttttgtg 202800 tcttatggtc atatgttttt ataaccactt tcccatttat tattaacctg ttaatatttg 202860 taaagtgaga aaagaaaaat ggagccactc aaactttttc ccagttagaa acagaaatgt 202920 attgttaaca ttgtaaatca aaaaagctat tgaatattta taatacttgt ggtgtgcttt 202980 gtcaaataaa tattgatggc ttcattctga aggtggtttc tagggtcaag ctatggttct 203040 ttaggtctaa gcattgccgc atcctaaagg aatctatatt caagataggt ctgtggttac 203100 ttctgggtga ttgaattaat caaattaata ttctgattga ttgagagttt tgatggatcc 203160 cagattttaa ggaatcagta agtgtaaaat ataaaaacta tgatcatact agctttaatg 203220 agttaggaaa tgtattgatc tctttcagtg cttcagagtc attattctga aaatcaatta 203280 tgcatatgta gaacaaacaa aaagttggtt agtttcgatt ttgcattagg atgtgagaat 203340 agcaagagtt tataatatgt gtaaacaaaa tatttcaagt caagcagata atattttgtt 203400 atgatatagt ttagaaaatg cttttttata ctaagcctag tgacatttta catagacata 2034 60 atgcagtgaa aagatttgaa cactgaagct gcatgatcca gagttctagt aataattttg 203520 caatttatta tctgagtgaa cttagacaac ttaattttta tgagccttag atctttaatc 203580 tggaaaaaag agataataat accttaccta cctatgagga ttagtttgaa gattaagttt 203640 - 178 - 008252 aaaatatgta tatatcacct cagcacagat caaggagcca agaaatacag atgtagaagt 203700 tataaaatgt acatttttaa atgtgcaatg ggcatctata aaattcttca ttcaatgact 203760 agagaataac attctaattc aagaacacat gtatgatttt aaatgtaacc atttaccaca 203820 gacttctaaa tatatacaca tcagtgtcct agaaaagata ttatttgacc aaaatgtaac 203880 aaaatcngaa acctaataag aagaaaataa ccacaaaaaa acctacacat tcagaaattt 203940 aaaactattt tcaaataagc acagaagaaa caccaatgaa aataagaaaa tatctacaat 204000 taaaaaataa tgaacaaaag atgcatcaaa cttgtgggaa gtagctaatg gcatttttag 204060 aggaaaaaat ttagccctga atgaatatct agtaaagaaa taaaggctta aattaatcct 204120 ccaagcattc atttcaagaa tttagaaaaa ttatcaacca aggaaacact aagatagcag 204180 aaagaaagaa ataataaaga gaaaagcagt tattaatcaa atggggggaa agaggtgata 204240 aataatatca acataattga aattttgttc tttggaagca gtaataaaat tgataaatct 204300 actttaaaat tcaacaagaa aaaattaagg agccttcttc tataaagatg acagaggcat 204360 tgacattgtt ttttaacctt gaattcccac atgaanaaca aaaagaatac caagatatca 204420 aaaccccaaa tccatagtta atatttacca taaaactcgg taatccacaa accctaaata 204480 aaagcaagtg ttgacaaaca gccacatgat ttgcatgata ttagtgtctg tgggaggaaa 204540 ccaagcttag ttacagagcg tccaatgttc atggaatggt agaacatctt ctttgctcca 204600 ttctgaaata aaaagtccag gagtggaatc aaaattgaac aggccaggaa caagagtgat 204660 aaggataaaa gcagatccag acaaaatttg tggggggagg aggattggga aacagagctt 204720 ggaaattcca gaaagaaagc taccatattt gatcgatata ttaaaacaac agaagaggaa 2047BO actctgaagt tagaagagaa atactaaact gtttgggaaa actaattttg tataaaaatg 204840 agtaaggaaa attatcaaag ccaaattccc tgcaaacgtt attataagaa aaaaacagca 204 900 gaatactgtc tctcactgta gacaaggaaa gcaacctana aaaacgtgct ctaaccagct 204 960 caaactgtag catactattg aaaaattagt taaaagacat taaaaattac ataaggatga 205020 aatagcaaca taaataaaag ttagaaaaac tcagaaatga ggtgccagaa ctgagaaaag 205080 ttataaatta aagcaatgaa aactaaacga gaaggaacat aaagagacct cagatgattc 205140 aaaagtgaga aatagaaaat ggagaagagg gcatttttta aaatcaaaga gaaatgaaga 205200 aagagatgaa aatgatttgc aagaaaaatg acaaattgaa gaaagtcaaa aatccgacat 205260 ctgcattata ggtgtcccta aggaagaaaa accacagcaa gagaataaaa cgtatactaa 205320 acttttctga aacataaaaa gatatgaaac tatatattga cagcacacac ctaaaaatat 205380 tggagtaaat gattaaaaac aatacataca ccagacaaat gacaggtctt taaatttttt 205440 aaaaatatat tatctagccc aaataagaac aagtgattta taaaggaaag aacgttagat 205500 tatcatgaga tttttcagca gcaatgcttt atgtcagaag aaaatagagt aacatatata 205560 ggatgcccag gcaaagaata tctgagccag atttttttcc atcctgaaaa ctgactttca 205620 actttcaagg tcagagaaac tgctttcaag atctaagaat atgagtaatg taagatacaa 205680 gtaatcttgt ttcaatgaac ctgtaatcta ctaaaaaaca tgcctcagat aacctaaatg 205740 aactccaaaa gggactctta aaccatttgg cataaaaatt cacattcata tgtaacaact 205800 aactacataa ggacaacatg aagagtgaaa tatgtaataa ttatttgttc taatatggat 205860 agagtacaga tatacaaatg aggaggtgaa ngggaagaag atatgcaaac ccttaagata 205920 ttcagtagtn ttcttggttg tgtattttca ttctgagact attgtctatg aaatgtgaaa 205980 taaaactgag tcactgtnga gacaatctaa ttctggcatc cctggtgctc ttgataatca 206040 agagtgctct ttgtggagaa aaaaggagat atagatatca tatagaaaag cttaggtaaa 206100 aataaaaatc ctaaagaaaa cgtggcatgt atacaccatg gaatactatg cagccatata 206160 aaagaatgag ctcatgtcct ttgcagggac acggatgaag ctggaaacca tcattctcag 206220 caaactaaca caggaacaga aaaccaaaca cagcatgttc tcagtcataa gtaggaattg 206280 aacaatgaga acatatgggc acagggaggg gaacatcaca cactgggtgc tatcggggtg 206340 gggggcaagg ggagggatag cattaggaga aatacctaat gtagatgacg ggttgatggg 206400 tgcagccaac cactgtggga catgcatacc tacgtaacaa acctgcacgt tctgctcatg 206460 taccccagaa cttaaagtat aactaaaaaa aaaaaaaaaa aatcctgggt catgaattta 206520 aattgaaaac atcagaataa acccaggcaa aatatttgaa acagtgacca actccatagc 206580 agtgagcatc tatgggctct agatatgttc ttgaaacacc atttttccaa ttttccaata 206640 agtaaaccag ggcttcttga agaaaatggt tgattttgaa taggaagtgt acaagctgac 206700 cttggagcat cttgtcatac taggaacaag gaagctttgt caaagactac gagggtcata 206760 taaaaagaac ctacagactg accaacttga aaaactccca atgagcacag tagagataat 206820 ttgactaaca gtaagaaatg gtaattgaca tggactgaaa tccattaaac attaaataag 206880 tttaaatctt gagttcataa tgacattntt ttaaatagta attggttacc ttgagcgatg 206940 atagaaaatc aaccatgtaa aattaataag taaaagtcga taatcaagca tttatcttgg 207000 cttttccata ccagttttgc tactgggtaa ccaaataata caggagggaa agcacctcat 207060 tataaaacta tcccaactaa taaatgaaaa agaaatgaca gttttgcaaa cggaatgaag 207120 tcaagggatc taagaaatga acatcaagac ccgtaacatc acaaaaagaa acacacacac 207180 tctgttcctt tcgatggaag aatctgacac cacctatggc cttgccaaag ggatgaaacc 207240 agagtctgta tgatgaaacc tctggatcca gctgccgatt ttcagaaaat agaagacaga 207300 agaacgtgtt gaactgtacc aagagtgtgc tttcagcaaa atccagactc tctcttggct 207360 ggnngtgcgg tggcttactc ctacaatccc agcatattgg gaggccacgg caggaggatc 207420 - 179 - 008252 acttgagccc aggcatttca agaccagcct gtgcaacatg aagaaacccc atctcttaaa 207480 aaaaaaaaaa tgttacttag ctatatgtga tggcacacgc ctgtagtccc agctactcag 207540 aaggctgaga caggaggatc gattcagccc aggaggtcaa gactgcagtg agtcatgata 207600 attccactgc atgcactcca gcctgggtga cagaacgaaa ccctgtcaaa aaaagaaaaa 207660 aaaaaatcca gactcaaaaa ctataggtca aatggcccag gtttctctat tttaagggga 207720 aaaaagggta acatgtagat taattagaag acatcgccaa aaaatcagca agattcatac 207780 attggaatgt ctttcaggtc caaattattc aatcacaaat aactctatgg gaattgagtt 207840 gaagcttttg aattatgtgt taaagtatgt aattctggaa acgtgtttta acttagaagt 207 900 tcaagactta aaatatctct atattgttga taatttaaat ttaatttact tgttttgagt 207960 tattaattct gtcaaagaat tgttatctga ggcagttgga tgttttcagt ttccaaacaa 208020 aaccgaagaa aatcttcttc aacgaatctt tgactttttt caaagtaaac tccttgtaat 208080 tccatcattc atccccttcc tttttattcc cacatctctg atatgtttac atccagaatg 208140 agcaattact ggctcctttc atagctgtaa tttacaattt cagatctgaa attactcatc 208200 atgaaatgga tctctgtatt ctttgttttg ctaattattt caggtgattg tcagtatctt 208260 ctagagccag aagagggcac tgcagagcag tgtagctctg agctctttat gtttagcaca 208320 gtctaggaat gccctttgcc aaaaaaaatt cgcaaagctt tactatagaa gtaagaaaaa 208380 agaagggaat aactagataa tcactatagg ccttctttat ttagtagaaa aaagaaagta 208440 tataacgatc taaatacata tctaggacat caaatgtaat atcatatata tatgtataag 208500 tagatgaaag aagacatata tatgtgtgtg tgagtccttt ccttctttta agaacatccc 208560 aaaggccagg tgtggaggct cacacctgta atcccagcac tttgggaggc caaggtcggt 208620 ggatcacttg aggtcaggag tttgaggcca gcctggccaa catggtgaaa cctcgtctct 208680 actaaaaata caaaaattag ccggtgtggt agtgcatgcc tgtaatccca gctactaggg 208740 aggctgaggc aggagtattg cttgaaccca gggggcggag gttgcagtga gccgggatca 208800 caccactgcc ctccaacctg ggcaacagag tgagactctt tctcaaaaaa caaaacaaaa 208860 caaaagaaca tcccaaagct aaatctgatg tttttctatt ataatattga ttctcctgct 208920 acctctggca ttagttataa aaacagttct caagtactct cctgtatgaa ttaatactgt 208980 attttgtctt agttcaaatt caggacaaag gccattcagt agccttttag caatcttgtg 209040 cagggcagaa atcatttagg caatagtgta ccagatatta tcccaaaagt ttttcctaac 209100 gtgaaattgc aggggcagta gataagtgat tggattcaga ctctcatctt ccactgaagg 209160 aaagacaact aacacatgtc attgtccttt caagatgaaa taagagttgt ggaatctgga 209220 gtcctctctt aaaactcact gtcacaggac ctctgtgcct gggtctgtct caggactatg 209280 tgtgttctta ggtgtctaaa gagcagatgc tccacgatga tggggccacc agctgagtct 209340 tccatctcca aaagcggctc caagcagctt tccttaggag ctgagctgtc aactcaagag 209400 aggttaaatc catgtcaaaa gtcaactttt ggccgggtgt ggtggctcat gcctgtaatc 2094 60 tcagcacttt gggaggctga ggcaggtgga tcacaaggtc aggagttcaa gaccagcctg 209520 gccaatatgg tgaaacccca tctctactaa aaatacaaaa atcagtcggg catggtggcg 209580 ggcacctgta gtcccagcta ctcgggtggc tgaggcagga gaatcgcttg aacccaggag 209640 gtggaagttg cggtgagctg agattacgcc actgcactcc agcctggatg gcagagcaag 209700 actatcttaa aaaaaaaaaa aaaattatgc caaaagtcaa ctctttcagg aggggagaag 209760 taggaaagtt tcaagattta tcattggaaa tagcatgtat gtgctaccaa gaaaatggaa 209820 gttccaaata gttggtatcc actcttgtct ccttgtccat acattaattt ttctcctaag 209880 agttctagtg aataatgaaa ccaattccgg tttttcattt ttgagcagaa aaatgaacaa 209940 accacctttc ctcatgaggg ggaaatcacc agtatgttca gtgatgctga tctttactac 210000 atatctgctg taaaatatga aaagtggatt gctagttttg atgttatcta ggttatcatt 210060 tgtgactaaa aagacctgtt tgttactctg gttcttctcc actatggctt tatggacaca 210120 ctgactgaca ttatagcatt tgttcatagc tttgtttgta gaatggggtg ctgtgagttt 210180 gtacctcaac atacctaacc cttgttgaaa agaaactaac taggtgatct atttttactg 210240 aatacagagt tttctgtctt ctgagggttt tctttccccc ctcaactttg tttgacacac 210300 agtatgccct gtagggcttc ttgcctggtt tcaagatctg gctagcacag cacttataca 210360 ggtagtttgt accaaattaa cttgccctgt gcacacatgt ccacttcagt ccttcaccca 210420 gtaccgcctt tgcctctagg aatcttagcc ttccttcaag gcccagtata acctccactt 210480 tttcattgaa gcctttgccg actagtccag acttgctgtt ccaccgcaga gagctccagc 210540 ccctctgatc ttgtgttttc tcctttacgt atcatctagc actgtgccat ctactgcact 210600 gtacttgtca ctgatgatct catgtgtcct acccttgtct tcctatctag gctattcact 210660 tctagaaacc aagaactcca aattctattt accctaaatc tcctatggtc actaatatca 210720 tgctaagtgc tcagtaaatc taatctattg atcaaaatga acaggtctca attactatct 210780 tcagtctgac tactggagtt acactttcct gttatttatt atttattcta gttatttatt 210840 attgtgaatc tagttattta tttattatta tgagaacata gtggcccatt tgcctgtttt 210900 ctctttgccc ttcttctgcc tgctttaact ccttgtanac cttccaccaa tctaaaatct 210960 gcaactagca agccacacag aactgagctt ggggagtata tctttgaatt tacagtcagc 211020 atccaggatg gttttctcct tagttctctt ctctggagta attttatcat cgccaactag 211080 gaagagatct tgaatgccca tttcattcat tccccacacc tgcaagcaaa acaactccta 211140 aactcattgt tcagattagt gttaggttac aggccaagaa cctgcagcag acataatgtg 211200 - 180 - 008252 gattagagaa cattgcgcta atcaaagaca aataaggagt ccagtatgag ggtcaacgtt 211260 aaagtctttg gaaactgaaa attttattca gacgaaggcc attatcagag tgaggcacag 211320 ccaatgcacc atccgtggct gggacgtggg gccccacaag tttgttcatg gtgctgccca 211380 aataaaagta aaaatagaca agaattgtca attgtacgat gatgctttag cgaaacatat 211440 tcatgtttca tgttttaaca atgtcatcta ttgaatgatt tgacagtata ttatttctta 211500 acattccaaa actaaaagtc atactcttat cattttcagc ttcgcaagtc tgttttcagc 211560 atcagggcaa ggaaccttct ggaaaaggag accgcagtca ctaaaatctt aaggtaacac 211620 caattaatgt tgacacatat ttaaacacaa gtgggattat attaatcgaa aaatagcttt 211680 ttgaaaaatg aattaatgtt ctgggcatca gcatatactt tctgccagaa agtagctttg 211740 agtatctatg tacttataaa caatgtaaaa gacatattga tgtcatttct aaaaaaaaat 211800 tttagtagct cagtaggcct ttctttcaag tatttaaaca taaatgataa gtataaacat 211860 tgaccaaaag gtgaaaataa cttactgtta tttttaacag ttttatatct tttgtgatat 211920 gtaaaggaga aattgcaatc taaaggataa atggtaaaag ggtaaacttt ttcattatag 211980 ttcattacac tattaagagc acatttatta aaaatccnga aataataaaa agtataccaa 212040 tcttagaatg cctatgaaaa catatgcttt taatgctact ttctagaaga tgcgctgaaa 212100 tgaagtatca aggaggaaaa ctaagcaacc gacttgagtg ttttataaga ctatatgtta 212160 tattatggag taacatgatg agtataaaaa cacataggca tttagaatat catttattgg 212220 tacatatatt acatagacat atgtagatga cacaactcaa gattgggagc tggaggtagg 212280 atcgctcgca aggcgagtgg acagattcat gccaggaagc cccctgcaaa cctgcggtga 212340 gtgataagga gttctagctc ccttctacta gatttattac taggaataga cattaatgca 212400 ttctctccag aatgcatatt actataactt ggaagactgt gtgagaaaat aaagcttagt 2124 60 atatatgatc tttaaaacta ttcttacttt ataatactac ctaaaatttg tgtgcaaaaa 212520 aaatttccta actcatcaag acagtatttc tctcaatatt ggctgacgtt catgcataga 212580 gttaagtatg tgttggagta gctgtcatag gatgaaaata agcatttcta gggcttgtat 212640 attatcacca caaaatgtga tgagataaac agacaaggtc tcattgaaat agacaacctg 212700 cgccccaact taggcttact cccgactggc tggtaactcc atcctggtat taccatcaga 212760 tcatgcagtg aggctggcct tgaaccacac atgttacatc aggatcatga ttccagctgc 212820 acctacatca gacaggttca tggtagggtc ctacagcaag gctgatccac tcactaaaaa 212880 gagaaattca ccctcaagcc agaaactacc tacaaggaca tcataattct cagtctgcat 212940 cgagtatctg cctgccaagt caggaactac agggacctta ctcacctgtc cagagccact 213000 gccttgcagt gtgaatgggc cctctgtgtg aatgggttcc cagagacttt ccaaggcaga 213060 tgcagaaaca gatactggca aggaagagat ttccagacct cacttctcta ggaactcttc 213120 cttaaaactg atctatcact ttaccttttt caccctcctg tccttattta cccctctctc 213180 agtttacaaa agaaaggtga aactctcact catccggaaa ctatggcgtg tattaatctg 213240 ggatgtaaaa tcctctgggt tgccaagcaa aaggaccgtt cccacctgga tttctccaaa 213300 aagagggtcc catggggccg tgccttatta tttatccagg tgacagtttt atggataact 213360 tccttcccaa agctatccac cttagattag gccttagaaa taacatgatt gtcacaaggg 213420 catgtggtta catgttattt gaattgaaaa .ataaagccag actttttctg acagattcag 213480 ctgattggtt caacaatgaa aactgattta ataagtgccc aatgtcataa aattgaatag 213540 attggatggc gaagagtctc caatattttc ttaaaattat tttcaagcac aaacacaata 213600 atgctagaaa ttacattctt ctatttaacg tggtgcaaga gggatatagt tgctgatttt 213660 taaatataca gaagtgtttt tgttttttca attatataat gtacatgagc aatattttta 213720 agactatcag ggaatataga cacctctaac ttgcctaaca atggtgtagt ttaggggata 213780 cccacagcca ataaataata ttttactgca gcccagcata ctacagaatg agaggcagca 213840 ccttaaaact acctgagcta tatcaccttt tgaaactcag gaaataaaaa gccataattt 213900 gaaaaaataa aaaaccttag aattttttaa aatacaaatt tgatcttaga aagccatttt 213960 gttttttctt ctgtgcgaat ggtttcttca ttgcttctat gctagtttct gcatgaatta 214020 agggcaagtg caaaaaatca aactgatatt ttgggaatac ttatcttttc accactaatc 214 080 gtagtaatag taatggcaat gataccatac ctgcattata atgtgatcca tcagttaaaa 214140 agtcaagaat agattgggtc catgtcaatg agtgtttgta ttgttttata ctgtttttca 214200 aggaacaata acagctaaaa ggaaaaaaag agggagaaaa agaatttttt ttnnaaaaaa 214260 aaaaaaggaa aacatttgca gtaacctcag gacaattgat tcgaataacc ttccacaaaa 214320 ttgggaagga gaaaaagctc tccaccactt tttcagtgtc tttttactgc ctctagggac 214380 tccgctctcc taggctccaa ccttgctgtc gattaaactc tgccaaggtt cctggataac 214440 ttagcagtgg ggtctcccag gaccatttgc attttgaaag aaggctttta aaaaatgaaa 214500 gggcttttaa gacaaagagg atgcagattc tgtcttttaa tagatgggtc tatcttgata 214560 taaataaaag aaatctggaa gagactgccg ttttagtgtt ctattcctct taactttcta 214620 ggtgttctta atttcataga attattgttt ttagtcttat ccaagaggtt cctcgccagg 214 680 gaggtaagta acagcagtat agggaggctt gaaaaagaga cctgagctgg atcattccgg 214740 caaatatcac aggttggcat tcgtcctaat ttgtgtgcag atagagataa atcacaaaaa 214800 ccttaaatcc agagtgatat aagaatagga agtttttatg aaatattagc gtcttgagat 214860 aactttccaa aaaataacaa gtatttggga aaaaataaag caaccccttg acttaaggtt 214 920 tctggaggcc tggctgtgta aacagacaat taatttcaat tctcagctct cttgtctatt 214 980 - 181 - 008252 ccatgggagc taggaggagg cgatcttcag ggcccctgaa gatctaatgt aggatactgg 215040 acttggtgtt aaatcccccc tggctgggat caggctctgc ttctctgtaa cattgaacac 215100 ctcactaatc tgaccctcag ttttcttatc taccttaaaa agattgtctg aataatctat 215160 tgaccaagca aaatttaata cttattccag taagggaaac tgccactttg ttttcataat 215220 gtctcagaag ggataagtga gaggtggata tatgtaaggt tttgggtttg ggtttcagtg 215280 tttgtaggat ggaggtctgt ttagactggg cagaattctt gatatgagtt gaggagtatt 215340 gaagatgcct ggtatggaca agtaatagtt actgttctga agacggcagt gctgtggttg 215400 aggaccagaa agagaaagcg acctgcctgg tgcctctcca ctgctttcca tgttctttcc 215460 cccggctttg gttcaacatt tgagtaatcc attctttaca gcgagtttgc ccttctattt 215520 tcattctcct tcttgacttg ggtgatctaa gagaagaaaa ataaatggtg accttatgtt 215580 gtcctcttca cacatgaaat ttattttttc ttttgtgcct ttcagatttt taaccagaca 215640 catgtattac ctttaacaaa caaatacaac tgacttttat agtggttttt tttttttttt 215700 aagtttggca ttgatcccca ttgggtagag tcttcacaaa aaatcctaat gaataccaac 215760 tatttgtagg tgactagctt gaccagttga gtgacctact gtctggagaa aggggttggc 215820 ccagcagtct atagttcaga taaatgaatt tgtgggaagt gcctgaaatc aacggtgaag 215880 ttgctgactg tttaacagct ttaccttcct ggacaagtca agtcatattg acgcagaggc 215940 tcagctctca gccctggaaa ttaagctgga gagctgccag gggtcttagt tctcaggggg 216000 gacccacatt tccaaatatc catcttccat ttgaaacagg atccacatta ttctagcaga 216060 aagtatgtta tatacattac tatgaacagg aaattccagc ccacagagga agaagtcttt 216120 cagagaggaa gggccctaga gcctgaggta tgtaactcac aggtagaatc acccatttct 216180 atgtataatc atctttaatt catgttcttg tacttttgaa aattaattga tgttttgaga 216240 aatataaatg aattttttga tatatttgag aactctttga tgagaaaaaa aaaagtgggc 216300 catttttaca gaacttgagg ctactggccc atgctaaaat gcagatgatt cacttggcac 216360 aaacaggcat ctgagttgtt gttttttaag agttatccga agaggatgtt ccactacctg 216420 aattataaaa tccaggattg ctattaaatt tattgttgtt ctgacttaga ctgtatcgtt 216480 cagctccact aaataaagct tcttttcaaa aggcagtgac cttttctctc atcaggcgaa 216540 ataaaaaaca aatagcaatg tactttcaaa agcatcacta cctttgttga gatttcctct 216600 aattcaaagc cctgaaatcg agcaaaaaat gcattatcgt tcctgcaagg ctgaatgaat 216660 agagaaatgt taaaataaaa caattaaaca ttttacatta gtttagaaag ttccatcctt 216720 gtgtggtttg ccatgagtac ctccttagtg aggacaatat atttataacc tttcgaatgc 216780 tcagtctcgt atgatgtttg cattctgaca tttggtttta taacgtaact ttctttgcca 216840 tgcacttata ttaataactg tttaagtctt aaggaatagt ctttagttta gtgctactta 216900 ctacagaaaa aaattaaacg gatttaataa aatacataga gatagttttt ctcttatcat 216960 tttaaaagaa tcatttaatg actataaata caacttatat actacttaca tatatatata 217020 ancactatgt atttattaat ctgcaattat gtaaatgaca ttagctggat tttttttcct 217080 gaaagaaagt gcccctaaga attcacgcat tcattcgaga aatatctctg tagtgtctca 217140 catgccaggc atttcacacc atttgctttt tcccatttgg aaagttctcc ttttataatt 217200 attgttccat aggttagcca gctattcagc .aactttggat aaggtcatat tatttctctg 217260 cagctccgca ttctgggcag aactagcaag acaccctgta cactgctggg agggcctgag 217320 taccttacag cgntcccctg gacctttgag ctcagctcct cagtggaaag ggaaggtgct 217380 ggactgaccc tacagatgag agaaagaaac gacagagacc cttgcagcct agggtttgtg 217440 ccaacccctg agatagcagc aatttttttc ttagcatttg gtatgaataa ccccaagttt 217500 taagaaaata gagtatgaga atgaatgtca taaatattta tctttgcaat tctatgaagt 217560 ctgcctggtt attgttggct ttttcttaag attttcctga cacttaaaaa ctctgataag 217620 ttagttgaaa gactatttgg aaataagcta agtagataga tgaaaattaa ctgtattcag 217680 acaaccataa actaaggaga caagatgtgt agatatatgt ttttgcataa aacataaaat 217740 agtaaagatt taaaagtggc aaaatataac taaaacaaaa ataatatatt ctaatcacat 217800 ttttagatag tacctgattt gcggtaatca aaatatgtta tattttattg agatagcata 217860 tgatttatag attgttttta gttaagccca aaagtgatta taatagtgtt atggattaga 217920 agggctacta ttgagtttta ccaaacagat gtgggaagtt ttttttaata gactattttt 217980 tgagcagttt taggtttgcg gcaaaatgga gcaaaggtac agagatttcc catgtacctt 218040 ctgcccccac acaggcacag tctccccaac tatcaacatc cccatcagag tggtacattt 218100 gttacaattg atgaacctac actgacacat cactatcacg ccaagtctgt agctgacact 218160 agggttcact cctagagtac tatagtctat gggtttgaac aaatatgtga tgatgtgtat 218220 ctatcattat aacatcatac agagtaattt gagtgctcta aaaatcctct gtgctccacc 218280 tattcattcc tccctcccca caaaccctgg caaccaatga tctttttact gtttccacag 218340 acatagaaat gcacatagaa atacatagat atgtgtctcc atagacatgc attgatgttg 218400 atggcagctt tattcctaat tgccaaaact tggaagcaac caagatgtcc ttcagtagga 2184 60 gaatggataa gcaaactgtg gtacatccag acagtggaat aattattcag catggaaagg 218520 aaattagata tcatcatgga aagacatggt gtaaacataa atgcctgtta ctgagtaaaa 218580 gaagtcagtc tgaaaagtct acatactgta tgattctaac tatatggcat tctgggaaag 218640 atagaaccat ggagatggaa agatatctta acaaggtctt tcttttgtta ttttgtcact 218700 ggagagtaaa gtagtaaaga aaaatggaat acatatctgt tgaaaattgg ccaatgcagt 218760 - 182 - 008252 tcctgcagca tttacaaaat ttcactttta ttgtagtaaa aagttcagcc tttgcattgt 218820 aatttgcatg tttcggagtc agtagttggg ttactaaatt tgaagcccag tgaatgatgc 218880 ggggttgcag acatttttta tgctgaaatt taatagatgc acgaaaggca tggatttctc 218940 atgagttaat ttttcttttt tgtttatagt atgggtataa gtgcgtaaga taaagacatt 219000 caaggtttat tttattttct gcctacnatg aagttctggt cgcccatgta tttacatgcc 219060 tttctatgaa aacattttta tgaataacac aagattggga agaactaact ggcagttgct 219120 atgaaagagc cagatccatt tactcaggat agggctgact cacaaaggag tggggtatcc 219180 actaggataa aaatgcagct ttttcctcat ctcagcaaga ggccattacc gacgagctca 219240 cccattagca tccgctgttg tctctacttg ggaagatcac attgtcactc tagtggagtc 219300 aggtaagagc catttccagg aaatactcat tatctattct cttagttata cagtcatagt 219360 tttacataat taccactgaa agactat Cut ectgttgtet gctaatgtga ttacat.aca _ 1 .-4 2u ttaaccaacc ttggataata cttttaaatg tattacttgt ggagttcaag atttgagcaa 219480 aatccctcaa atgtaatgct ctgcaaagtt atactgcagt tcctttagat ggtaattacg 219540 tttaatacca gacggtattg tgtaatctaa cttttctata atgagtgaaa ataatttgaa 219600 gactcatagg cctctccctt tcattacaga atgtataggc tttttccaca cccttgttct 219660 gggacattaa gaaagaagac atggagacaa agaagaaaag aaccggggac cccctgctac 219720 tgtgtctata aacaactagt ggtgtttaaa ggctaataaa gctccaaaga tctctcttat 219780 aaccataatc tttgggccaa ttttaaaata ttttagatgg agaaatgtca agaaaaatag 219840 agacaatttc aagacatgat tctctttccc caacgtccat aaaagattat atatatatat 219900 atatatatnn nnggctgtca tataaggagc aattgaaatt ccacagcaca aaggcaatgc 219960 agggggaaaa aaaaggatca atctttcata ttattttttc tcaaaggctt ggcaagaaac 220020 caggggtaat atttttgcaa ctgggaatgt gtgtttgctg aagaatattc ccacatttga 220080 atcatcctca ggggtcctca tcttgtttta ctttcaaagg tagacagggc ataacattaa 220140 ccccaactgg gaaaaggaaa atgcctcact gtaattctcc tccagggaac aggaataaag 220200 ggattacata tgcctgcata atgaagaatt tcctcaccca gacaagacaa aagacttcat 220260 tatttttcac cagacacttg aagtcttgtg cctcccagtt gcagaagctg aggaggcagg 220320 ccagtttttt atatgaagcc actagatgga gttcctaaga aactggcctt ctagtacttg 220380 ccattgtgaa aagatctagg gaagatgtta aaaaccttgg tacctgacac tacccagtct 220440 ttcctctaag aaaagtgaga ctgtccctga agatcacagt attcttccag taaaattagc 220500 cttgccttta ctctgggaag caaataaatg agcattttct ttctgaaaat gcacctcctt 220560 tgtccttaca tgagaccaaa aggcatctca atactaatga aaaatgtaaa gtggcaaaat 220620 ataacaaaat ggaatattca agttcttctc cttctgggta ggtctgttac ttcgtaactt 220680 taaggatgta tgcatctcat ttcttatata aaatattgga agtctcaaat tttgtagtta 220740 ctcactattt cggcttgcaa gtattagatt cctgtaagag gaggtatatg gagactatgt 220800 tcccagcaac agatctgatc tccagttaaa gaaattgaac taagttcacc tgctacagcc 220860 tgctctgctc cctcttggct ttcaggcttc cctaatagct ttcacataca acatagagaa 220920 agangtatga agtttttagt cgtaagaagc aatgtctttt ttttttcatt actgtctctt 220980 ctagtgtagg tatgcagccc attattcttg agtggcatca tattctaact tgagataaaa 221040 ttaagattat ttaacttgcc cctttaaaag tatattttaa acacatgctg ttttgtccgt 221100 gattattttc atgcttcaaa agcagggagt gatcagaaaa gaaaatattt gatcttcacg 221160 gaatttcaag gttcattcat tccacaaata tttattgagc acctactgtg gataagagct 221220 gagaaataat catgaccaat tttaaagaaa accctactct cttagagctc tttggagcat 221280 ggcatatgtg acgaactaaa agaacaaaga aaatggcatg gagagaacga ggttcctcct 221340 ggtaaagatg agaccagaaa gggaggttag agggacacta aacggggctg tgcaagccat 221400 gctgctcgtt aggcttagtc tttttaaggc agtgataata accgcattcg tgatctattg 2214 60 ctgcaggaca aattactcca aaacttaagt gatttaaaca acacgttttt catctcacag 221520 ttttagtcag gaatctgggc atcccttagt catgtctctc tccaagctgc atgcatttgg 221580 acttgaatgg gaagagcccc actcaatcgc atggttgtta gtggattcag ttcctcgtga 221640 gctgtgatat gaaggcctca gttccatccg gtttgctgga ttcctcccca agtggaactc 221700 tccgtagagc acctcacaat atgcagcttg cttcaccagg gcaagcaagc aagatgggaa 221760 gagcaagtca gcgagagaga gagagagaga gagagagaga atgaatatgc tatgtagagt 221820 gttataacat ccaccatgac gcgggaaaag atacaacagg aaataatagc aactaaaatt 221880 caaagaaagc gttgatgttt ccaaagcact gtcacattca taatcatctt tacttcttcc 221940 agctccctgt gaagtcagaa ggaaaggcat tattgttctc tgcattttac atatgggaaa 222000 tggaggctta gagaaagcac atcacctgtc taaggtccca cagcttataa aagctgaagt 222060 cagaagtgtg tgcttttaaa gcttatgttt tctcaactca tcttctccat taggcttcca 222120 gacaaaggca gaactttcca tcagcagctg cactgcaggg tgcacttctt tgaaccatga 222180 gaggctagca agttagggat ggggtcacac agtccccacg cacctgccgc taccctccct 22224 0 actcacccct ccccaccttt tcacttactc attctgttca tcaggcagca tgagggccag 222300 aaactgaact tcctaggaat gggaggctga gtgataaagt ggtccctaat caactcacag 222360 cctagtaagg ggaataacta atgataacaa aatgccatga gcgttaaaca agcttggtta 222420 ataacatact gtaggaacat ggagtataaa acaagtcttt ttttgtgctt tcacataggt 222480 gaaatattca tttctcattt catgttactt ttcagagccc atttatatca gtctgtcatt 222540 - 183 - 008252 atactgtgca tgttaaaaca ctatttggta tttatttcat ggtttgcctt actgtatatg 222600 cttatctata gtaacattgn tattcccttt aaaaaataat aaagcaaact tttaccatgc 222660 ttttgtatga acctgggcac attggattag agtatatgta acctcttagc aggtctgatt 222720 gtgtccaact gaagttcata gctttctgcc aacactgtta ttttccaaaa tagcaaaatt 222780 tcttacatga ccaacaaatg tgacaatggc ttaggctgta caaggctgtt tcattatgca 222840 gaaacgtcct atcatgggaa tctttctata cctgtctcaa tntccaggtg agctctaggc 222900 caaagggtca taaaacttcc ttcccattgc cttgctcctg catccaccgt ccaacctcag 222960 gtttataact tccatctgcg gtccttccct cattcaccag gaacctgtgt gtggccagtg 223020 gtctccttgt ataatggaat aagatttttc agaatgttct cgttagacat atttatatat 223080 ctgtagaaaa gttgcttcat gctgtttagt aaatgaaaaa tagtgattat ttagggtttg 223140 tttccaagaa tacatttaca ttcagtgaat ttgtatgatt tttcatagat attcaccata 223200 attatcttca tccacattaa aagcattttt aaggacttaa atatgacata tttagtaagt 223260 tcttatataa tgtattacta tataataagt tcttatatga tgtattatat aagttcttct 223320 gtaatatatg tgttatattt aaatacttaa aaatgcttct attatacatg tatcataaga 223380 gctgctattt taagtgcgct aagcacatac aaatcttttt ctcaaagatc ttaattctca 223440 gacagatggt aatcatattc aaagccattc aagaaatgca ttttaagaaa cataaatgtg 223500 aaaataatat gtaaaaaatg gcagaattgt gtgttcaggt gttgagcaaa tagagtatca 223560 ttagatttgc atatcataaa atcattgcat tttcaacctg tgagctatct ttcagagcaa 223620 ggctttctct tcattttacc aatgaggaag ttaggcccaa agatcactca actacttcat 223680 aacagtgtaa cttctggcac acaggtcttt taatcccttg tgttattttc ttgccactta 223740 tcaactatct caagggaatt cctgtcattc ccagttggag aaaattttct ggagaataat 223800 ttttttttct caccaaaatc attcacacca caggtgttag gccacccaat tgtcttcact 223860 cattagcaan atcagaaatt aacacaagtc cagttattgt ccaaaaagaa catgttttta 223920 aaagtcttcg tggtttaaac aagtggtaac tggttaccag taagctgcaa gcttcagatt 223980 gaggtttctg taggtaataa ttccaaaaac tgtgtgcttt tttttttttt ttgaactgtg 224040 gttttaaaaa caacacataa catttaccat ctaaaccatt tctaagtgta cagttcagtc 224100 gtgttaagta tgtttacatt gttgtgcata attccagctt tagttacttc tctttatgac 224160 tcatttttgc ttgtaagctt atgttttcct atcttcctaa agttctacaa aatctccttg 224220 gaacatatat ttaatcattc caggagcctg agtggacaaa tccttcccaa agggggttta 224280 ttagaaacct tgtttaaatt caaaaccgac atcatgattg cctgtgtgta ttaatctgtt 224340 ttcatgctgc tagtaaagac atacctgaga ctgggcaatt tacgaaagaa agaggtttag 224400 *.____---I.*.-.____J_______________л________---_4-__------. Ш Л Л СГ\ LyaaL-i. Laua yLL-uauyuy yu-uyyciyc-yy uuivduocii.. didyuayauyj у иус-ас-у y\~c- --ччиу cgtcttatat ggcagcagac aagagaagag agcttgtaca gggaaactcc cctttttaaa 224520 accatcagat cttatgagac taattcacaa tcacgagaaa agcacaagaa agacctgccc 224580 ccatgattca attacctccc accgggtccc tcccacgaca catgggaatt caagatgaga 224640 tttgggtggg gacaaagcca aaccatatca ctgtgggtag gctggccttc tgcttatggt 224 700 gtcctgatag aagctctacc attatttttg gaaaggaggg aggaggcaga aggaggaggg 224760 gcaagagggg agctttctgt ttattttcca .gcccttcttc atagctgtga tacttctgct 224820 ttgtaagtgt tcttgacgca gagcgtaaag ttcattttcc tttataatta tacgttattt 224880 gctgtatttt gtgttttcat tnnggtgttg nccntttaaa ataataaaag taatgagcac 224 940 ttatgtgtta ttcatctcaa gtgtacaaac catgcattta tcttgggctc cttttttttt 225000 cctagagctt gcacccgaca gcgcatgctg agcggatcat ttgaggggca gcccgcaaag 225060 gagaggtcaa tcctcagcgt gcagcatcat atccgccaag agcgcaggta aatacttggg 225120 aagacagggc ttcagagact ctaaaaaagg ggggttttac aggaaatggt cgctgtacac 225180 agtcacggct gtcagccctg gctgccagac aggcatggct gaggaatcgc gttctatttg 225240 ctgtttgtta caataatgaa acacctaatc agatccaaag attcaaaaaa atctctactt 225300 cacaatcctt ttccttagag atggccaaat tctcaccttt aagatcagaa ggaaaaattc 225360 tgaattttgt tagatgggat caaagacatt tggagagagt ataagcttct gctatgaaaa 225420 gaaatcatag gcatttgtgg cgttgtagca gaaaaagtgt tttattttga ttttatcatg 2254 80 gtcactgctt gaaaggccag tttgatcttg agatgtatta accagtgcct gtggttgttt 225540 taggtcacta agacatggat caaacagcca gaggatcagc aggggaaaat ctcttgaaac 225600 tttgactcaa gatgtaagtt ttaagcaatg ctttggattt cagaaacgtg attatgaaaa 225660 tgtatttaaa tgactgctta gacaatgccc agtgtctttc tgctgcattc ccatgtctcc 225720 ttaagacttc ataccagagg gaatcaggaa actcaggata tgaattatgt agccaagtat 225780 cagaagtaac tgttaacagc agaaaactcc agtcatccca cacaaagcac atgtcttagt 225840 taaaatagta acacaaaagt gtcattatca gttggatttg acttgaaatt aaagacattt 225900 taagcattgt atataggatt gatttgcatc attgactttg aatattcact gagaatcgaa 225960 tattctagaa tctagatcaa gggtctataa tttcagggct ttatatatta ggaagtgaaa 226020 ttttatttct agaagaattt tcctgtcaag gactacctag tatttcccag gaaagctttt 226080 tgcttttaga ctctaagtca tggatttatt tactggctct ataaatagtt atcaagtgtt 226140 tgctctgtgc tgggttctgt tttaaggaac gaggagcagt ggggtaataa gatagactca 226200 gtgccctcct ggagcttaaa ttttagaata ggagacagat aataagttca taaaaatata 226260 atggaatttc tggagtgatt tctttttatt ttccatcatc tcttcattac tgtgacagat 226320 - 184 - 008252 ttgctttata atgtaaagta aaatgaagtc atgtaaataa atcaaaggtg gggatttggg 226380 ggcagggaat tttagatacg atggttatcg tagacccatc tgcattatcc cggagaccca 226440 aatgaaatga aggagacatg aaaaaaatgc agagaagcca agcgttatat ggcccagcag 226500 ctataccatg caaagattac aatgaaaggt ttgggaaatg catattaaat aaccgtaatt 226560 atgaagccct ggtaaaaaga caggcgttag gttgaatctg gatgaactgg cttaaaaagt 226620 aatcttgaat gttcagagcc aaggatggca cttgagaaag gagggctagc aagagaaagt 226680 tagcaaagcc ttggggttat ttcgtttttc tctttttcac cacttatctc ttatttcttg 226740 agcttttgga gtgcttgcat actacatctg ctttcattct gtagaaactg ctgttttagt 226800 ccaagtacat ggtttttatt aagtgttccg ggcaaaatca agagcgagaa gttggcaatg 226860 agaacacctg atatcaccaa taccctatag acatggttnc tagttggcac agagnctctg 226920 tttaccacca tgatttttaa tatccatagc tctatctgat tggctaccat attgttgaca 226980 tagaattgcc atcaaatcca acatatggcc tgcgcaactt agaactgcag agtcagccat 227040 attgcacgga gaacattttc aggtgaaaag tcaaagaggc aaccctcatt caaagcgcct 227100 tcaatatgtt tcaaccgaaa tgtacaacct acttaagaat caaatcagaa catgtctttg 227160 aaaataatgt gtttaaacat agacagcaaa ggccctattt gaatagtctc agtgccggag 227220 cttacttctt cnnagagact tcctacccta tttttcatgt agaagtatag taaaatccat 227280 ctcccacctt ctccatagca aaatgccatg gggttctgat catttccaat aaattcttat 227340 aatgtctttt ctcccacaaa ggggagattt aaattagaga attttaactt tttcacttat 227400 tcggaatgat agcangacac agataattaa gtctaaggag tgctttctgt aaaatctact 227460 gagttactgg aactctggcc ttgcaccttc catggggtcc atctaaaggg ctgtgaaggg 227520 cctgcatggc ctgctcacct ctcttagctt tacccacact ggcatcctgg ctattccaca 227580 tgtagacacg ccaggcctgc tcctagctct ggaagaacac atttctgtag tcctaccaca 227640 gcctgcaccc tcacctcctt ctagtctttc ctcaaatagc accttcatat tgaggtctac 227700 cctgacctgt tcatttaaaa tcataatctt ctagcctgtc tgtgtctctc tatgcctttc 227760 acccattgtg attttttaaa aaatcggatc actaagcatc ctcttacata cttattttga 227820 attatttttt atctgtctcc cactgactag aacatcctat gagaataagg atgttttgtt 227880 tccacttgcc atatcctgca cactagaata gtgcctggca cataatagga tttcaatcaa 227940 tatgtgctga atgaattact ttttagagga acaaatactc ttgctatgaa gactgtggcc 228000 attaaatatg tgcatttaat cgtatacata ttttatatac tgtaaattta tatatttaat 228060 gtaagtatat acatttactt taaattacat atttaatgta aatatatacc tttagtataa 228120 atatacataa ttataaatat atacatttca tatagatata tatttattgt aaatatatac 228180 ttagtataaa tatacatatt taatgtaaat atatacattt agtgtaaata tacatatttg 228240 ttgtaaatat atacatttag tataaatata cattgtaaaa tatacattta atataaatat 228300 ataaatatat gtttaacagt aaatatatgt aattaatatt tatactttat attgagggag 228360 atattagttt taagtttttt aactgataat atattttgca ttcaaaaccc tcctgtttta 228420 acacaccccc tgttaattta ntttcntagc attccaatac agtgagatat agaaatgcac 228480 aaagagaaga cagtgaaata aagatgattc aggaaaaaaa ggagcaagca gagatgaaaa 228540 ggtacaggtt gctgcagtgt cccatggctc tcattcggga atggcaaaat ttcccttagc 228600 ctcattctcc catgaatttc tccttcaagg aaagtgcaag aagaggaact cagagagaac 228660 cacccatact tcgataagcc actgttcatt gtcgggcgag aacacaggtt cagaaacttt 228720 tgccgggtgg tggtccgagc acgcttcaac gcgtaagtac acttcatttc agccaggtgt 228780 ttggcctgag ccatgcaaac agcaggggtg ttgtcatatg aagaagaagg aacgtgtaat 228840 cctaatgggg tcacaaaata aactgcggac taattaatca tgctgctcct ctttgctgaa 228900 cacccgacca ggtggaaaac aaaggtcgca agtagttaac atttatagaa aactcgcatt 228960 gataaaacac aaaacagcca ttttcgtttg tgcattttaa cagtttcctg atacacttat 229020 atcggcatta ttaactccat attttatttt ggaagtactg agacaaaagg agactaaagg 229080 acatggtgaa aggaaggtca ttgtccatgt cttccatgag gcttgtagtt cctttgtggt 229140 aaaaatacct caatttgttt tttggagaat aacgtaaagt tgattagtgc tttcattacc 229200 aactgttata agaaggaaac ccactgttca ctgtctctac cctaccaaca tggaagaaat 229260 gtgtaagaag ccgtaagtag agtggacatt gaagacacta gttgtggaca cctaagccaa 229320 gtttgacagg caacaaatgg atgaaatgga tttttgtcaa gcatacctgt atnagtccat 229380 tttcatgcca ctgataaaga catacctgag actgggtaat ttataaagaa aaagagattt 229440 attggattca cagttccaag tggctgggga ngcctttcaa tcatggtgga aggcaaaagg 229500 cacatcttac atggcagcag acaagagagg gaagtgaaag cagaaacccc tcataaaatc 229560 atcagatctc atgagactta ttcaccacca caagaacagt atgggggaac cgcccccatg 229620 attcaattat ctctgactgg gtccctccta caacatgtgg gaattatggg agctaccatt 229680 caagatgaga tttgggtagg gacgcagagc caaaccctat cagtaccttt aagtaaatat 229740 tctgaagaca ttaagagtca tggaggaaat gagaaaactt tccatgttcc cattatgtag 229800 ttggagtgtc cctgatgcca tcgtttaagt aaattgattt ctttgttatt aattagtagc 229860 atatgataat cactatctgt tatggactga attgttcccc ctccacaaaa tccatacggt 229920 agagtccaaa ccctcagaac ctcagaatgt gactgtattt ggaaatagga attttaaaca 229980 ggttaattaa aatgagatca ttagggtggg ccctaatcta atatgattgg tttccttaag 230040 agaacatttg gacacagaca tatacagaga gaagaccttg cgaagacacg tagggagaag 230100 - 185 - 008252 acggctctcc acaagtcaag gagacagccc tggaacaggg ttttccctca cagccctcag 230160 aaagaaccaa ctcagctggc atcttgatct tggatttcca gcctccaaaa ctatgagaaa 230220 ataaacttat gttgcttgag ccatccagtc ttaagcacca gtggtacttt gttacagcag 230280 ctcgaggaaa ctaatacaca ctattataac atttcaggga tggaagagac ctcagggacc 230340 actcggtgtc atttgtgtgc cccctccttc aacagaggag gacatttaag cgtgcagaag 230400 ttgagggctt tgctcaggga cccagctact acctggctct ctcttgattg ccagactatt 2304 60 gttctttctg ctgttctata aagactttca caatttaaag caactgagtc acaattatta 230520 aaatcttcca tgagttttca aagtaaagga agaaaatgcc attttggaga ttgatttgat 230580 cagttcttga agtacgtttt ttttatcttg aattatcctg ttggctgcct gtaccttaac 230640 atctcatggt taattctcca cttcacttag tgtgatgtaa tttccctttt atcagcaagg 230700 cagctgatgt tgaatacaat tgatgaagac tctgttgtaa ttggggacac atacacaaag 230760 actgtaagca atctcattga cagtgctcac aggcttcctg aaagtagaaa caacgactct 230820 cccttccaag aaattctaaa taaatcattc taccacatca cagttaaatg cagggggaag 230880 aagtctattt aaattatgga gtatttaatt aaataatctt gaataactaa cattcttgaa 230940 ctattaaggc aaagatttca aatgagtgtt tttaaatgaa agtcaaaact taattttgta 231000 ctacccatgt cagaaaattc tgaacaaagt cacaaagtct aaaatcaagt attttgagaa 231060 atataaaact ttattttcat aagcatcaac tgcaaattct ttggattttt tctttcatca 231120 atgttctctg acctccagag ttacctacag ttaacatatt ttataatgga cattgtagag 231180 aggctgtaaa agagatttta ggtagatgag atgtttgcaa atatgatttt tagcaagtta 231240 aaacatgctt gctttgttgc agaattttat atatgtgttc tcttacagat ctaaaacaga 231300 ccctgtcaca ggagctgtga aaaatacaaa gtaccatcaa ctttagtaag catcaccaac 231360 ttcattttcc atgcacattt aaagttttca tgtactattt tatgctgcat acaacgatta 231420 aaattctctt ttattttctt gctatatagt gctatgaaat agatgattgg ggaatgatgt 231480 ttttaaaata ccagtcatgt tgatagagcc acagaagcca tcatctctaa ctcttagaag 231540 cctttcctgg aacctttatg tttctcaatt ttgtttggtc tttggcagtg atttgctggg 231600 attggtcact tacctggact gggtcatgat catcgtaacc atctgctctt gcatttccat 231660 gatgtttgag tccccgtttc gaagagtcat gcatgcacct actttgcagg taccgcttac 231720 cgctgcaaaa tctcagctta caactaggag gtaaaatact gacctgctgt ttttccgttc 231780 attgtaacca aactgccttt tgcccgtttc agattgctga gtatgtgttt gtgatattca 231840 tgagcattga gcttaatctg aagattatgg cagatggctt atttttcact ccaactgctg 231900 tcatcaggga cttcggtgga gtaatggaca tatttatata tcttgtaagt ctttgcttat 231960 tgcctaaatg aaaaatgtaa tggttttgta aaatctcctg catttcttgg tataagcgca 232020 agaggtatga gttacagagg cttagattcc ctaaccgaat tcttaaatgg actctagagt 232080 agttcattat atgcaaataa gggccatgca tgtaaaatct aaattaagag catcataaat 232140 atctatggat agttatattg ttcaaataca ctaatcacaa ttttatgtat gaatgagaca 232200 tacattaata aggtctaaca tggcaaatct agtttgcatc ccacaaacct tatggcttct 232260 atactaaaaa aaaaaagttt gcctcatata aaccatgtta ttgattatga gccccttatt 232320 gctgaaagca aactaaagtg attgtgattg .tatctctctc ctgattccat tgcataaaat 232380 acctggctat agcagttgcc cattcaactg tctagtaaaa ctcgctggaa aaccatagaa 232440 atgggctcag agaatttttt atttttcctg tatgtccaag gaggcatgaa atcaatgtct 232500 ttgcagagac aacaaggaga attgtgatca ttaacaagta gcatgttgca ggtcattaat 232560 aaaatagcag taaccttgat aaaaggaatt ccagccagat cgacatcagt ggagcctcca 232620 gagtagtaag ttaaaagaag aggcagaacg cactggtgta cacacacaca cacacacggt 232680 gtggacaccc ccatttcttt cctgcataca catgatgact ttcctttgag gtacctagca 232740 agtctagatg aggtttgttg tttaaagtta acagatggca cgtacaacca gtgagtggaa 232800 tctctgctaa atgaaaagtt gtggccagtc ggctgtcaga aaatgtaaat agtaggggat 232860 gaattgtata aattccagag atcatttgtt cccctttagg tgagcttgat atttctttgt 232920 tggatgcctc aaaatgtacc tgctgaatcg ggagctcagc ttctaatggt ccttcggtgc 232980 ctgagacctc tgcgcatatt caaactggtg ccccagatga ggaaagttgt tcgagaactt 233040 ttcagcggct tcaaggaaat ttttttggta tgtgagaatt atcctttgat tccagttcac 233100 agtagtccac ctctaatgca gctaatgagg gaaaagtgag tcttgcttat tgaggtcaat 233160 tcaatagttt tggttattgc cagagggtaa tggaggcact aacaaaacac aatgcacctt 233220 гяяягя PT'T't' ar-*- -•=¦ t- лп/т i-" a -*t- *¦-"¦• rtrtr*(tm) f- a f- frarth t- +" a *-¦*+¦ a =_¦•¦* pa г" р|*ааалапая 9 ~ < 4 9 Я П -----------.- -.-.-.WW---. _w_ WW-- - W 4. v.--.-. ---¦- - " gcactctgag atttgaaaga gttcntatct gctgactctt tcggtactaa cctcttgact 233340 ctaattccta ctctgggttt tagtttcctc ccatctgaaa atgaaggata agaagattca 233400 acacngtgta ctccacaaat gggattagga attcatgctt ttgggctcct aatctcattc 2334 60 tgtgtgttcc tgaaataata tagcacagaa tttgtcttta gatctcagga gntgttaaca 233520 taaaacaact ttaaaaatgc aacatatagg tataacccac ttcaattaac ttcagcatta 233580 cagtaatgag tgattacaaa ngatgattga gggggcaaaa cgtactttnt agctgggcaa 233640 acattgattt agggtcttct gaaaacaagg tcagagataa aacaacaaga aagaggctca 233700 ttgttcacaa tctgttacag aaagatcttt gccttacaaa acagtgatca aaagtttttt 233760 ttattattga tgaggtctgc ttgtgcatga gaagcatgaa ttaatttaca taggcttgtt 233820 ttgtatcttg ctttctagag acttacctgt gatgagaaaa cagctgtagt tgtagttgac 233880 - 186 - 008252 acaaatgtat tttaaagata gattgtgatg ttactagagg tatatatatc attactgact 233940 ttttttttct ttttttttga gacagaatct tgctctgtcg cccaggctgg agtgcaatgg 234000 cgcaatctca gctcactgca acttccacct actgggttca agagattctc atgcctcagc 234060 ctcccaagta gctgagacta cacgcatgtg ccaccactcc cagttaattt tttgtatttt 234120 agtagacacg gggctgcacc atgttaccca tgctgccctt gaactcctga cctcaggaga 234180 tccacccacc tcagcctccc agttgctgaa tttctttttt aagatgggtt taaaaattct 234240 tgttttcttt atgcaaagcc ataaaacagt gactcttcac acagctggaa ttaaaagaaa 234 300 atatcacctg actgctaatg tatatgaatc ttttgatcaa taataaattt tagaggagaa 234360 tttgtgcctt tccctaatgg tggtcagtgt gattttagta cacttaggta tatgtttagg 234420 ttattcaacc agtgcagttc acttgttcca tattttgccc tctgccactc gtctctgctg 234480 agaccaaaga agtcagccag cagagggagc atgagttaat taacagattt gctcttcagg 234540 actgaattct ctcctctcct ctatgcatag aacattctcc taacgcacac cttcacagtt 234 600 acaagacagc taagccactg tcccgctcat ttgtgctctg ctaataaaaa tatgaatcca 234660 ggccgggcac agtggctcac gcctgtaatc ccagcacttt gagaggccca ggcgggtgga 234720 ttgcctgagg tcaggagttc aagaccagcc tgaccaatat gatgaaaccc tgtctctact 234780 aaaaatacaa aaattagctg ggcttggtgg cgtgtgcctg tagtcccagc tactcgggag 234840 gctgaggcag tagaattgct tgaacccagg aggtggaggt tgcagtgagc tgagattgca 234 900 ccattgcact ccaccctggg ggacagagtg gcactccatc tcaaaaaaaa aaaaaaaaaa 234960 agaaaaaaaa ataaatatat atatgtccat ttgggaaaaa atagacaaaa tataattact 235020 taaacatttt tttcctttgt tccattaaat aaaaaagatc aaggtgttga ttttaaagtc 235080 agaaaagttc aagccatatc aaaaatacct tattccattt tcttacatga aattcatgtt 235140 tccatttaaa aaattcaaag cttgttaatt ttctcctttt gaccttccag taaggctaat 235200 tgtgtgacct atggtggaga agttaaaata tatcatcaaa aacccatgat ttttatacaa 235260 tgttatctta gaagtcctat ttatgtcact gtttaaacca aacctaatat ttagtcttta 235320 acaagagatt taacatttca tcaatttctt tcaggtctcc attcttttgc tgacattaat 235380 gctcgttttt gcaagctttg gagttcagct ttttgctgga aaactggcca agtgcaatga 235440 tcccaacatt attagaaggg taagatgtat ctttctgtct tttgaaataa aaattaaggg 235500 ccaagtgtgg tggcatgtgc ctacaatcct agcactttgg aatgccaagg agggaagatt 235560 acttgaactc aggaggtcaa ggttgcagtg agctatgatc tcaccactgt actccaacct 235620 gggcaacaga gcgagagccc tgtccctaaa aagaaaacaa aataataata aggtgtttac 235680 actcccttgg caagcatgtg gtaccagtgg ctgacaagcc tagtttaaga actccctata 235740 а а a r*r*r*+ a a n -i-,*-*--.*--.*-*-^-. а. а -"/ч-* -v -i-.-.-.*-.-**----* +• a *¦ -r+- -•+¦ ОТНЯЛА ______ __._.y __v-.v-i.____ __________ v.b.v-.-.-.yv.-- __y--.-.у-у- ---у-_-_-¦- -_--__ ctggctagaa aggttaataa gatgaaaagc acacctcaga gaatgtcact cttccctcct 235860 attccagtgt caacccactg ttgacaactt gagygtggtc acccaagatt ccagctgttt 235920 acagctgtaa gctttggata ctttcccttg aggtcttagc taacttgatc cttcctttgt 235980 tgtcacataa gactcatatc agcaaacagc cccatgcaaa aatgtccaag aagcagcgtt 236040 tggttctcac cactttttat tgcttcccca ctctcagctt tctccccatt agaaangcca 236100 gtgcctttac attagctcta gctgcccaga .atctccctct ttatgacaac caacaaggac 236160 ctccacaaag acccaaactt gaatacttgc actacggcgt ttctttctct ttcttcagtc 236220 attggttata agggaaggct agctgctgta acaagcccaa tatttcggta gcttcacata 236280 atgaaatgta tatattgcat cttggatcag ttgcaatata tacatttgca atatatacac 236340 tatatacagc tctcctccaa atgaccactc agggactcac atgccttcta tcctatgccg 236400 ccaccctttg ccatgtcttc agagccttct tcaatgagct ggagataaca gtggagagcc 236460 atctccaact gagtgggctg tgggagaatc tgctccaccc acattcctct gtaaagagca 236520 gagatactgc aaggggggct ggaatttagt tgtgtgctcc agaaaaataa aagggggaca 236580 cagtttggtt aataattaga caatttttgt aatatcataa agtgtcccat tctcttagga 236640 ttcttatgtt acaagtaaat tcataaagta aaccaaccac tgctgatgta taaactagaa 236700 aagcattttc tctttgatcc caataaaatg atgcctcagt tcctaaatgt atatcttcaa 236760 aaggcctact agacctgaac tacattattt atttctagca aatatggcta tctagtatag 236820 attttaaatg atacaaattg ttctctaatt gtgacaaaat acaggtaaca tctttgttca 236880 gccaaaatac attataaatt atattaataa tgagaatttg aaacaaaggc cagactggga 236940 gagacattcc aagagctttg atacacctga gaagacagaa caaggaatgt ggcctgacgg 237000 tcatgccgta tcaaattagc tgataaagaa agtgacagag acctgacgca ttcatccaca 237060 agcttgtgtg tggaaagaag agtgattttt gatccacact gaacagaggc cagaaagata 237120 ataacacttc aggatgaagc ctgctaggaa tcttggatga ctcagtatat taacagtggc 237180 ctggccgggc gcggtggctc acgcctgtaa tcccagcact ttgggaggcc gaggcgggcg 237240 gatcacgagg tcaggagatc gagaccatcc tggctaacaa ggtgaaaccc cgtctctact 237300 aaaaatacaa aaaaaaatta gccgggcgta gtggcgggcg cctgtagtcc cagctacttg 237360 ggaggctgag gcaagagaat ggcgtgaacc cgggaggcgg agcttgcagt gagccgagat 237420 cccgccactg cactccagcc tgggcgacag agcgagactc cgtctcanaa nnaaaaannn 237480 nnaaanannn anaannanna aaannrmana aaaccggggg ccctaaaaac gggggggttt 237540 tactttttcc ccttatcccc ccccaaagtt aaaaattggg ggggaccgcc ccccccaaaa 237600 aaccctaacc cccattttta ggggcctttt ttcctttttc caaaaacagg ggggttttgc 237 660 - 187 - 008252 caaggtttaa atggaaaccc taagggaacc tccccttttt aaaagccccc ccccccaacc 237720 aaaaccttta aaaaaaattt ggcccgggaa aacggggttt aaattttttt gtttttcaaa 237780 aaccaaaaaa aaaaggccgg ggccggggcc aaaaacccct taaagaaagg ggacccccgt 237840 taaatttttt ttgaaaaact tccccccttt tttctaaaaa aaaaaataat tccaaaaaag 237900 gggggaaagg taatcccccc acttttaaaa cccccccctt tcctttaaaa aaaaaaaaaa 237960 t 237961 <210> 3 <211> 47841 <212> ДНК <213> Homo sapiens <220> <221> экзон <222> 3895..4001 <223> экзон 28 <220> <221> экзон <222> 9611..9731 <223> экзон 29 <220> <221> экзон <222> 9816..9914 <223> экзон 30 <220> <221> экзон <222> 15776..15869 <223> экзон 31 <220> <221> экзон <222> 16382..16488 <223> экзон 32 <220> <221> экзон <222> 16697..16771 <223> экзон 33 <220> <221> экзон <222> 17934..18053 <223> экзон 34 <220> <221> экзон <222> 23644..23712 <223> экзон 35 <220> <221> экзон <222> 24928..25076 <223> экзон 36 <220> <221> экзон <222> 25913..26006 <223> экзон 37 <220> - 188 - 008252 <221> экзон <222> 30767..30899 <223> экзон 38 <220> <221> экзон <222> 31561..31676 <223> экзон 39 <220> <221> экзон <222> 34044..34201 <223> экзон 40 <220> <221> экзон <222> 37493..37643 <223> экзон 41 <220> <221> экзон <222> 39652..39801 <223> экзон 42 <220> <221> экзон <222> 41563..41680 <223> экзон 43 <220> <221> экзон ylllv Л Л 1 -1 1 1 J.JJ. <223> экзон <220> <221> признак ошибки копирования <222> 45842..47841 <223> 3'регуляторная область <220> <221> аллель <222> 31646 <223> 99-79310-29 : полиморфное основание А или С <220> <221> аллель <222> 42373 <223> 99-79311-50 : полиморфное основание G или С <220> <221> связанный праймер <222> 31618..31635 <223> 99-79310.ри <220> <221> связанный праймер <222> 32080..32100 <223> 99-79310.гр комплемент <220> <221> связанный праймер <222> 42324..42344 <223> 99-79311.ри - 189 - 008252 <220> <221> связанный праймер <222> 42704..42723 <223> 99-79311.гр комплемент <220> <221> связанный праймер <222> 31627..31645 <223> 99-79310-29.mis <220> <221> связанный праймер <222> 31647..31665 <223> 99-79310-29.mis комплемент <220> <221> связанный праймер <222> 42354..42372 <223> 99-79311-50.mis <220> <221> связанный праймер <222> 42374..42392 <223> 99-79311-50.mis комплемент <220> <221> ошибочное связывание <222> 31634..31658 <223> 99-79310-29.зонд <220> <221> ошибочное связывание <222> 42361..42385 <223> 99-79311-50.зонд <220> <221> признак ошибки копирования <222> 26,350..351,353,356.-357,366..367,371,383,389,392,396,399..400 404,411..412,416,421..422,435..438,440,442,44 6..447,450,455 457..459,461,463..4 65,1805,2175,2328,24 02,2561,4048,4 661,6659 6799,7509, 7534, 828 9..8290,8294,10663,10789,11052,12083,12904 13017,13916,14116..14117,14680,14 971..14974,17093,18448,18669 18767,18808..18811, 18838,18971,18985, 20714,20903,21355..21367 22901,24093,24112..24115,24171..24173,24868..24869,25356,25797 25879,26062,26234,26824,27130,27254,27437,28745,28781,29278,29475 29526..29528,29617,31152,31790..31791, 31796,32011,32262..32273 32296,32415,33656,33719,39140,39142,39343, 3994 4,4094 5.-40947 41948,42166..42169,43371..43372,46709..46720 <223> n-a, д, с или- <400> 3 gtttaaaaaa aaggtgaagg cctctnggct agattttttt ttcccccctc cacaagggcc gcttgggaat tttggggggc tccgggaatt ccccccttta ttcccccccc ttttgggggc taaacccttt ttggttaaaa aggggaaacc aacccggggg gtgggggggg gccccttttt aagggnntaa ncccgggggg ggnggtttnc nncggggggg aaaannnnan ancccnntcn aaaaaaaaaa aaaaaaaaac agtggcctaa acccagccaa agctgaagat tgatggtgac ttcatgatgc atttatcctg tttcctagag ttttttcccc aaattttttt gggaaaaaaa 60 caaaaaataa aaacccttcg ggagaaaccc 120 taccaagggc ccggcccggg ggggggggtt 180 caaggggggg ggtttccagg gttgggggat 240 cccttttttt ttaaaaaaca aaaaaaaaat 300 cccccttcct tgggggggtn ngncannaaa 360 antaanccnn aatncccccc nngccnccca 420 cccanannna nannnaaaaa aaaaaaaaaa 480 gagcatggtg attttacatt atccactcat 540 agcagccagc agatgagcat agacacagat 600 cagtgaggtc tgtgcgaggc tcagatggaa 660 - 190 - 008252 agcctaagga aattggcacg ccgaggccga aacttcagcc accaacattt catagtttcc ctcctttgtt tggagcctag aatagtcatt gggaaacatt tttcatgttc aaataacatg actgggaggg agagcaagca gaccgaacaa acttcaggag ttgaatgctt ctaacctgat gaacaaggcc ggggntttgg atcagacaca cccatatcct aatttgtcca ctggcttttc caggaagaca cactacaaca taatcgcaat ctcagataca gtctgtggca gnatgtgtta gaagtaacgt алаллпа at sirt gggacagaaa gattaaaatg ccttgaaccc aataaccaaa aaccttttta gtgttcaatt agaacatttg ttagcctcca ctgttctgga aattagcata tcctatggct caaaaatgga cattttatac agtggagttg ttttccaaag gattgcgatt tttctcagtg agatggcaca tggagaagtg aactgcctgt ttagataaag tctgagttca tgcaatggca cctggagaga atattctgaa taactaaaag ctagattata ttctgtatag aatatgtatc taaaactaaa aaccctaaat gactcagcca gtgagacggg ggccaaaaac catcttacta agatcacaca agttgggaag tgttgtctta gactttgtca aaatgtcagt aggaggtcct aagcaacagt taatctatga cttcagggag gtgaagaagg ggaggaagga aggtgatggc ctcacacaat ctcaacagtg agaaatgtaa aatttcgttg tcctagtgcc aatgaggata aggtcaccca accaagatac aattggaaaa cagcnctttc gtagttttag tataggaaaa attgtgggta tttgcattaa tggcattact __ гчгчп •_"¦_-/*¦> ggaggaggcc gaatttttgt agaaaagaga gggagaatag acagctttat caatggtttt tatctccttg cctcaagacc cttctatatg atattttcaa gaataatatt catctggatt aagtttctat ctggctcata tggttgcacc cctccacatc ggtgtgaggt tgatactgag tctactcagg ctccaaagtc ggcttttggg ttttctgacc tattcagaat aaaaacctgg gaatggtgtg tagttttgct ttggaagata cttcatgaaa tggattcaaa cttatcctga cataaagcta tcatagagag gcttgagtct gccatgagtg aaagaagaga cagttcctgt atgtgttttc aaactcaaca ccaaaaccct tgtgtccaag tgcttctgtt aatataaagt tgtcagcatg gaggtgggat acacggagca gcatgggctt ctggtctggt gatattatga agtgttagtt gttagggaga atgagaagtg cacaaagtac tccattccta cctaataagt cagactgctt ggcagtgagg caaataatgt ctgaaaaaag aagactggaa agttttcttc aatattctta aaactcatcg aagaggagac tgaaatattc tttaggttca gaaatgagtc tgagatgtaa ttagcgtatt aaaagaaacc taaacaacaa aataaaatca ggatcctcca cacttgtatg gtttccacct agggacatat tggtaactct attctacatc ctcgccaatg gatatatcat catcttgtcg tcctttgctc tttgttacta attccgtgaa aggtttcccc taatgtcagt attttgggtg tttgtcanta agagatgtta gaaataaggt ctagagaaat tgccttaaaa gtttatatct tttccatttt aagcagaacg attgaaaaga tcaaatccaa acataagagg cggctgaagt tctgttggag gaaaggggga aagtagatac gaagcttcag ttgctgcgcc ctgagcactt cctccctctg gatcaatata gatatagata gctgaggata ccttttgaca aggtgaaatg aaatgtggaa ttctgtccaa tcagaacgtg caaaagccta tgagaagcag cttccaactg ggagtactgt caggaaagga ccttccaagg ggtcacagtg tagaggcagg gcttctgaga ttggaatcct ttctggaacc tatttctgcc accttgttta tgcgaagtac ccatgtagga aaaaactgaa tgcctgaaat gcctggctaa aaaggttggt aagaagagga tcctagtgca gtttttgtta caggtgtaga aagattaaat ggaactagtg tccaggcacc aaggtgggct caggtgggtt caagggacct ttatacaata aaggtatggt tatggaaaga aagaatataa actttatgag caaaacgntt cattgtgttt tccaggcttt ataaattaaa tcatgctggt aatccagaaa ggagacttgg gccaagaagg т*ч rtr> i- firm a +* rrfr- /-"/"-"¦-r* r^r-a Ji\v CVJ W - <-)l-|--lU W Y-J > -) l_ "_••_.v* gtcaaagagg acgtgaagca caagaatttt accatctaca atcatagcca aagaaacaaa tatagatgtt taatttttaa tccacatacc acacaataca cccacttcaa catagagttg tatacacgac agccactttt ctatgtctct taactattac ccccatccct ctaatattct ttttgtctct atagagttca taacatatgg tcttttgtga cttttttctc tgttgtagca cgtatcagga cttcatccct gttggaccac attttgttta tctgtttgtt tttggctatt gtgaataatg cttatataaa attttcatct ctcttgggta gatacctagg gtgttagtgg tttgaggacc tgctagaatg cccaccagta tgtaagggtt ccagtatgag cttgttgtta tctgactttt tggttatggc tatggttttg atttgcattt tccggatggc tgtgcttttt ggccatttgc atatcttcct atttaagtaa tattctcagc atcgattttt tttgctgggt agctgtccac atcactgttg agaagaaaga gtttcctatc ctttatcttt tctcctcttg tttttctact ttaggaagat gtgtcaaaga acttaaattt aaaattgagg ccccgtgttt ggtaagtata tcataaaaaa taaggcttta aataagtatc atatggcatg aaatgatctt aagtcataca gggcgaagcg aaaaaataag agagcaagta gtgcaatgag attatctcag tatttttcta atacttcaga aattaggctt aacaggattt tctgtcaaat ttaaaggagg atctgatgtc ctcataatag aaaacccatt tacactgaag gacttccatt caccagcacc ttttgtcttc aaggtgacgt taaatccaaa taaagacaac ttgtatgagg gatgtcatga gtaaacaaac gcactgtgat ttgtggcatt cagcagtttt ggaaattgtg ttgagatgag tacaaagaag cgaaatcaca cacttaatcc agattagatt tcattcccgc gatcctcaag agaaaccaca aaagcaccat atgaggaaaa ggagaagcag catgcaatca ccaaatatgt tctacataaa tgggcgatgg ggatatactc cttttatttg ttttaaagaa tagtaagaat -..Vj <_ L_. \_> V-) Х-} v. atggcttagg aaacagtgat aaatatgacc aaaaacttcc - 191 - 008252 attgaaacac tagagaggaa taatcagtga cacagttgat tgcttctaac tttcccttta atattccttt ttctttaaat tgagttttca aagcacttag atcaaagcta gaaataccta gtaccttctt tcttatttta ttaatgacca tgagtgaatc tctcagtctg tgtcattggt gaactgagtg ttctgtcacc ttcatttgtc ctttaaaatt nttatctgtt ttttaagttt tcttttagaa aatgtgtaag aataacaact gagtcacagc acctctgctt ttgtcacaat tgatgttctt ttctaaatgt caaaactctt taaaaggaac agcatttcac ccagctgtac cctagcactc actgttacct ctctttggca tttttctcat atgaaattcg cttttatacg taatccaata gaatgcatat tgggttttta ttttttaaaa tatccaagaa ctgagaaaaa ggattcattt ggggtatgtg gcttcagctt ggagagagag gaatgtgaga catgagaata tcctggcccc actgctgact agaaaagccg tccaatttcc attgaaataa aaattaaagg agcataaaaa ttttcttcca tgagataaaa attaattata gcataaagtt atttatgtta taatttatta atttttatta attcaaagat aatatcatgg aagtaaatgt agtcaggaat agagaacact ggggcatttc atttccttca ctatagagaa aggtagaggg tgcctcccac ctctgccaag tgaggtaccc cagggggatc acttaacgtc agagatgggc agggcaagaa gagagactct tgtggccagg ctgctcatca ccatctggag agcagggtta tctggtgttc ctggagagcc cagtagtcac tgcctagggc cacatcatct gagtcctctt gagccttgcg gcaaagggct ttgtgctggg aaagggtaac cccctccaaa gggtccgtag gtgtgtttcc aggagccaga cttgagaatc acccagagac tgcacactta caggagaaaa ggtgcaatga ggcactcctg ggctggagag ggggttcaaa ctcttgaaca cctatggcca gaatccaatt gtatcagatg cacaagactt aaactccatt cctccattcc tttcaacgac atcccacctc agtcatgtga gaatctcgtg gtggcctgaa gaggatgagt ggaagtgaag aaagtggcca acttttcttc ctttcctcaa accggggtcc ttgagctgct aaggatggga ttgttatatg agttatagga tcagttggat gtaaaactga attactgaaa ataaccaaaa agctctggga cctccccaag aattcagaca gggagctgga acacacttga aggtagtgtt tggagaaaag taaacctgcc agttgtgccc tatagcactc aacctagtcc ataaaaccag ctacatggga tttttaaaat caaagttaat ttaaatgcaa tattttatgg aaggtattat taggggtttc aaaggcaatg ccattctaga agcataagca aatagccagg cattccttga acatcatcac aagtgctctt gtccatttct ccaggataat gtgacttgga ttgtagtgaa atagacacct tcgatctaat aaatgtcatc tgtaaccatg ataacttttt acactttcct tcttaagtaa gctaatcaag agttggaagc tgaaattttg taattttata ttggctaaat tatgattaag aaaataacat atctaggaat acactagagg aatgctttgg taattataat taggacataa a -I- a +• rm a /¦"•¦• a aff^a гчгч -"/-"•-. _¦¦"" <-"¦• -J, *- +-/-•- actttcttct gaatcactca gacttgatga gtcacctcag taaaaaggat taaacatatc atggattata aatacagtgt gatttatttt aagaggaggt ctggaatata ggaaaatggc atgcatggga gaaaaactaa caaagtattc aggcttataa taaatgtatt tattgagtgc ttgctcttca ccagctagta tgcttgtcac tgtggtaagg ccctgaataa gcgatggccc ctacccctta ggagcttaaa gtctagtggg aaacagaaaa taaaattaca aataatttna tataatatgc taagggcttt aatgctgcac agagaagccc ctaagcttac ctctggctca cccgaaactc atctccatgt aatgttaagc tacactgtga tatgttctag gaagaaggtg atcaggtttt atagggtcna aggcccaaac aatttggggt accaacatct cagaagggcc tgtacaaaag aggggtccta ccccttaggt ttctttggct tcatcatata cctgcctctg attcctacag aaaggctaag ggtacaatgg acacacagga gaagcaccta accccaacta tgcagggcca aagagaagga ccagatgaga agacacactg gttacctttt gaaggacaac caataggaac tagccaggtg aagggggcag ggtcagaggc aggtgaggag gaaagtctag gcataggttt ggtattacaa gttaaggcaa ctctatgccc aaagctcaag gcaaaaagcg ttggggtgct gtaggaagta cgacgtttct gatttctttt aggagagatg gttgtgtaag atacagcatg ccgtttttgg acaaagtctt tttctgcatt tccacatcca agggcagacg ctcagtcacg tcttcagggt gcccccatgt tccccaaggg aagagcagct ctccctacct tgatcttggg gggccttgga gcacctcctc tgggaggtaa aaaactgaca aaattatgtg gtcagatcta atgtcttagt tttctgaatt ggacaggcaa gattacaata tttcacaaat taaccagaaa aaatgacaac cttttcaatt caaggaaggg tagcaggtaa aaatgacaat aatagatgnc atttgggatt tattgaagga ctcngtagtt acaggtaaag tttcaaactt tgcaactttg ctagcatgaa gaaagcataa gaaaacactt tacatgatta aaactgaaaa agacttattt ttttaatgtg ataaattctt ccaactattg aagtctttga tgctacattt tgttttactt ggtttagtta gaatattttt cacatattct aacttctaga agatctagaa cataacagta gtttcagtgg ctgcaaattc agctaagaaa gcacttctac aaaagaaaat gagcaggtaa ggaagtccaa caccggctct gagaaccggt tcacttgctt gtaaaaaatc cttattcaag caaactgtgg catctgtaga atccttataa cttctgtcta ctgtaaaata tatgaaattt agtggaaaca tcccaaagga aagattgttg gaagcatcca aaagaatata tttgtacttt catacataaa aaaatataaa gcagaattcc aaaatatgca tttctgagtg gggcttttgc actatattaa agaccgtctc cactaatcag ccctaagccc ttaagaacaa atcagtttca gtttaatgtt ttcttcttat atcctcatct atttaatatt caaaaaactt atgaaggcac atcttgtgat tgtctctata gtttaaaaaa cataaaagat ggtagaaaag - 192 - 008252 tacagaacca gattttccct agagcgttgc ctgtcctgag ggttaaattt tgagaaatga gctagcagnn gggnaaaaaa tgatcaaaat ataatttaaa taggctccaa ggggcctact ctagtcagtg gtccggagct cagttcttca ttaaattcaa gaaatatagt ccatgatttt ccagtgtttg gttatcaaaa tcttaattat ttaaaattag cctctaataa gaaggaaaga gggggaaaag ccactgaaag cttcttctat taacaacaaa aatgttagct aagaaccata acaattaaag aagatgcaat ctagtaaaaa ggatcacttt tataattggg aagagaaaag tataaccatg ttaaaagaac aagaaatgag acatccctgg cccccacaga agtgggagct aaacagtggg tacacctcga cataaagttg gaaactctgg acagcgggga cttcaaatgg gaggaggaag gaaggaggtc ataggttgga aaaattacct gttggattct gcgtgcacta tctgagtaat gggttcatta gaagctcaaa ccccagcatt atgcaataca cccatgtaac aagcctgcac gtgtaccccc caaatctata agaaaacaga aaatgaataa gccaatgagt ttcaggaatc aggaatcagc ggtggaggta gaaagcatac cacccataaa ggggaccaga ctaagagtcc actgagaaga ctcagagtta ataagaaaaa atccaactgg aagagagtaa tttggaaaaa tacatactac aagcaaacac ctcttgccgt ctcagtgacc tctaaatgca tcactacttt ccagccctgt gttcaccatg atgacacaaa gtgaaataaa gtgactaaat tctactctac actttctata ctaacacagg caattgtaat ttatgccttt cgaaagattt tcagttacaa aactatttca aaatccacat ccaaaatcca atgcttcttt gtgcaaatcg ataaaatcat ctgtccagtc ctcagggtaa tcaggacaag gtagaaatgt tgcagaacct ctcaagctag aactagagct gttttacatt gagatcaccc catcattacc taatgaacac aatcaattat aacacattga tgcattgtta tgatttagca aacgaatgcc atatagcgta aaaatgtgtg tgtatacata tacacacata tgtatgtgtg tgtgcacgca cacgtgtgtg tgtatatatg tgtgtgtata tacacacaca cacacatatc ccacactcat gactggaaag ggctacagat atttgccttc gggtgctgct tattggaatt tcacagaaca ttcctgtatt ggaatcgcag ggcgaatcct cggaacttta atttcgacaa tgtgggaaac gctatgctgg cgttgtttga agttctctcc ttgaaaggct gggtggaagt gagagatgtt attattcatc gtgtggggcc ggtaagcgag cacatcgaat cctttgaaca cttcaagagc agctttcatt tgttttgatg aataaccata ttttccttta tgcagatcca tggaatctat attcatgttt ttgtattcct gggttgcatg attggactga ccctttttgt tggagtagtt attgctaatt tcaatgaaaa caaggtaagg atttttggtt tggataccac agggtttagc cttttctagc cactcacttt gagctatttg tttcttttca cacgcggcgt gaagccgctt taaactgtaa tgtgactggt gtaaataatc tctataactg ttttacaagc tccctcattg ctgcttttca gacacattat ttaagagtac agttctcttg ctaaatttac ccttttggta acagtgtcat taattcaccg acaactgaga ggcgcgtctc ctattgttca aattgaaaag aaagatgggc agcaagttat taaaatacaa tagatgaaag tgattcgcta attacaggta atattacttt cactaaatgt aaattgtatg cttctgtgta actcagtagg tgaactcagg attttgatgg gggtgagggg agtctggtag aaatttcaaa atcctaattc acagaaaaaa agattttgtg gcagggcatg gtggctcacg cctgtaatcc cagcactttg ggaggccaag gcgggtggat cacttgaggt caggagttca agaccagcct .ggccaacata gtgaaaccct gtctctacta aaaatacaaa aattagccgg gcgtggtggc gcatgcctgt aatcccagct actgggttgg ctgaggcatg agaatcgttt gaacctagaa ggcagaggtt gcagtgagcc gagatcacac cactgcatcc agcctgagca acagagcgag actcaaaaaa aanaaagaaa aaaaagaaaa aaagattttc taatttcata tgaacttgac atactttgtg aactcgggaa gcctgttgag tttgtaacag ctctaatgat tagaacaaag gctcacaaat gggaaaatna caatgtagtt tcctatcaaa gtgtgagttt ggggaagaac attctattag cactccttgg agaggggctt gctcatctag gcacttgagc tatgccctct attgtttgag acgcagtctt tggaatcata gtaagggaag tgcatgcagt ttttgagcac acactgtgta ctaagatttt acatattcaa tcaattctca caactctcaa atgcacggtt gcagaaaaag caatgactag tctggggctt ataaggtgtg anaaacaggg ccagaattca gaattggtgc ctattaccgc aggctctgtg attcttcgaa ggcatgagta gaataaagtc aaccttcact ctgcattaag ttgaagctag atctgcctgt aaagcaccgt gctgctcgat gttattcatg gctaggactg attcccatgc gttcacatca gagctcttca gtttgtgttg ctacacatgg gactgtatct gtcattcact gtcctcaggc tcaacgtgag tggtaccagt tagccagtca attgacacct ttgatgtgtg gctaatgagc atgtcatttc attcaggctt tcaaagggtg agatttaaaa attcctcttt tgggccccga ctctacttgg accctcagaa acatgggatg ttagagacag aaagaacagt agtgatcatg tcacccagcc ccttatttta ctcctatcac attcattaac gagatgcttt gagccctaca gtgttgagtt taataccaag cagaaagagg aagaaggatc tttttagatg cctttgagcc atgtaatgca gcagacactg agaattctag aatataagca gaagctaatc agtcaattaa caggagagtt taggcactgc ttgcccagaa caccgttaag aagccgtctg agcagacacc tgatttcagg aagattagaa tgtatcacag ctcacaattc agcaacatac atagaacaac ataaaatgtt cagtaactgg aaataaatgt ataattaatt gcaacacttg agcttagaga agtggctgag gttagcagaa taaccaggga ggtctccatc cagcactgta tttatgttct gcaggtagga aagagtaacg tggcctgttt gggggatctg gggcgtatgt tctatcagct caaatgggca cttaaagaaa ggagaaattg acaagaaatt atattaccaa - 193 - 008252 gctttacaaa gaaaagtttc atctaacagc tgccagaggc aaagttattt aatttccaaa gaatcatgca tatattagta tattaaccta atatctccat ttatgtataa ataattagtg acactattat tagaacttcc agaaaatcca gagnagaata ggaaaactct cccagaaggc aaacaaaaac atgagttgaa cgtcatatac tttaagtgta tcaataaatt aggacacaaa aataacaaaa gaacaaaaat ttgttatagg atgtatgtag ggagtggtgt cactgcatac ttaaaccatt cttgatttgt tttctctgca atttatacat tgctcttgac gccctcatgg tgtagagaca aaaatatgtc atatagattt acttgataat gcctattatt caaagtaatt agaaaagtgg tgggaataaa aaatcccaaa aaatcatagt aaaggaaata aattacaaca cataattacc aaaaaaaact gaaattataa aaaaactaat tatattaaag ataataaagt ttaatgatac gctgggtttt aaaaacatga tatagaaaat aaaacagaag tgggagtatt tcaaaaatga tttaaagcat taattggaga agnttttttt tattcaatca actgaggacc agcacacatc acagagatag ttcttttgaa ttttctgatt aataaacttt cttttagatc acattagtgt tttagcaatg tgatgtacta atgtacaaag gaaagatatt tgagatttga atttgagaat caagaattga ctacaatttg tagaaaatca gaggtatagg aagaatcaca aaaatcaaat agaaagtaag ggtttttttt aacaagaaag tagacttaca cagctcaaca gattgtattg attccagatg gtcagcattt tatcaattgc ttttttttag ttcaacagat tagtgaacta ttaatagtca aaaatgaggg aatggaaacc tctctactat cagaggtttt ctttcagagt ctatggtggc tgattatcag aagaatgaca caaaaattaa aacatatatc tacaataaac gtgcattatt taatgtaaga gaaatattaa caaaatattt taaatcagta agttaaatat aaagtagggg gaaacttgta aaatcaagtc taagctacaa gaaaaaacta aattagtgca atgtaaagag aattaaagac attttttatt taaattatgt aaagcagctc gtatttgagc acctaaacac tacccaaagg aatctttagg tctttaaaga gtaagacaat tatatattaa tcaactttcc cagagaacaa aaaagaacaa aaataatcac acaagttgta tcaaaaaatc acaatcaagt ttcaagcaca atgttttctg gacaatacta acagacaaaa attacaaata taatatgtca ggaagaaagt tagcttctat aataaacctc tcattaaaag gaaagaccca gagggtggtg ctatgctgag ttcaatatga tgaatttcag 4- ~. 4- ---. 4- 4- Luyuaua.aL tcaaatcagg aacattcata attcaccata catatccatt gcatttgttt ctataaagga aagacactct cataattgat gtaaatatca acaagcatca agatacattt gcaagaatat ttgctctttg accccatcaa gagacagtac acttcggata atctagttgt acatgttgga ttcatgcgta tggttagctc L L vayu.u v L L tcatatgcat atcacagcac ttgcaattgc acagcattga tgagtatcct tgaatattac tggtcatcat tcaccaccac cggcacccca cttttgtctg gagcctttta atgagctgcc atctactagg taaacacaga cacataggta gtacaattca gttccaagtc gtgtttgatt tgatcagatg aaaatttttt gtgtttgctt ttttaaacaa cagaaagata aaaagaagct tcataattca gaatgtataa aaatgtntct accataatga taaataattt cattttatag aatgtatcat aatatccaaa aacaaagatt aatattcaag taggaaggtt agtgtcagta gcagggaccc agaccaacaa atctctacct aaagcaaaat ataagcagca u. La иСпд <-р -.а ataattagca atcctctctt tattaccatg aatcaacatt tttgccctgg aaaaaattat aagcaaaata aaaagattag aaatacaaaa ccatttacac tcacacctat taggatatgg gaaagcaagt ttcttctcct atggatgctt ttcgtacagt atggaaagta gtccatttct agcacaaaag tgggtaagga ". L^o out- tttaatcaag tactttctta acatattaga caagctataa tttttgaaaa acatctccaa tttgggaact ggttggaaga tctgccatgt ttaaggggac aBtaai.yuLL cagtgtgata tatctatcac ctaggtattt gaacactaaa tgactatccc agtagttatg aaactcgatt aaaatacaag tatccagcat atgagcaaag acatgcaaat caaaatggtc taacacctgt tggcaacatc agagatcaac attgcagcat ggatggtgtt tactgttcaa ataaatatta tctgaaaaca a actnfrtartfarft -"--_-¦-"•-*•-> - ataatgcagt tactctatta aaatgcaaac cattttttaa ttttgctgct tgtaattatg agatacaagt cctcccaaga acatggtcag tcttggtgca -yLwLLLQU LCI L L L L L CICI L tttcaattca ctcaaacatt aaaaataaac atttannttc tattctagtt gatttcatcc atatttaaat ccaagactaa gtataaagaa gatatgaatg gtttcatctc agattacaat gtattgctgg tggtaaatgt aatggagaaa tggttatagt tgtgagcagt tgcagatata aaaaattcag tagtaaggag -#-•-"--+• *- +¦ *- +- ¦-".'-( У LL.LLLL- gcagtgcaat tacagaatat tctatgactt agaaagacaa atgaaatgaa tagaaactcc cgaaggctgc ccaactttga tgcaagccct agcacatgac tgtattataa tatcactgtt aattaattgt ctccattcta cccctcccca cattagccac taaaattttc gagaaaatat tctctacaaa ggcaattcac gttaatatta gtctggcaat taggaaattt aaagatgctt ccctcataga ccataaatca taaacaaatt aagaaacagc aacaaattct gatgcacgcc +- -" -"-4- +- 4- +• L tgtgtaatga ttgtgttggg tgtcaattag gctgtatttt cccctagtaa .caagtgaaat tgaatgacaa gtacaactta ccattaaaaa ataggaagat aaacaacatt atcaagtatt gttttggttn aaattcctag tatttactgt aaaatgaata agatttagta nnnnataatc tacgtattga tgaagggtag aaaatlt-v-CciC ataattttaa ttaggcagtt aaaaacagtt actggaagcc ctactattag ttcccaaacc ctgaaacacc catgtttctc ctgtttgtct ttcctgcttt - 194 - 008252 ggctttgctg acagcctctt gggcactagt ggccgctggg cataaggcta taatgaaaaa gaaatgtatt catgcactgt acggaactgt cgacaagaaa gataatgatg acaatcgcat accctaagac gctatatgca aagtgggcag gctgtttctg agttgttttc gaatctgtgc tctgtgttcc ttaactcgaa gtcgattgtt aatggtgcaa gcccagacaa ctgaaataca gccttttgct aagagtgtag tggtaataaa ataaccgagc gatcacctga ctaaaaatac aggctgaggc tgccactgca taaatagaca ggcattaaag taaaattttc aaatatgtac tgaagatcat tcctggtgac tagaagtcaa ttccagtctc gcccagcttt cactcttcac gaaggacctt gagagaggaa aagactatag ttataatcag acatttagcc tattttacct ctatgggtnt acagatatca aactattaaa aatatatttt accgtcgatc catcttccgc gtgcacgatt aggttgtgca ttaaaatcaa atgaattttt aaactgacag cgttttgaat ttggggagaa gtaaatgtga gttttagagc tactcgtcct tccgggaatg ttcacggtta aagaggggca ttgcagtggg accttcatct acagttttaa attgcctttg caaagacatc gtgtctgaac gccactataa ggttatttta agcagattct taaggaagaa agaacacaga ctcttatttt atgctggctc ggccaggagt aaaaattagc aggagaattg ctccagcctg gacagacaga tgtttgtctc ttcatttgaa taaacactct agcaatgtcg gtcgcttggc gttgatttca attttactcc gactgtgatg acaggcaaaa caagatcttc gactgcaaca tcctgttttc tatttcacaa tccacttctc aaagtgcctt actgaactgt cttaacattt ttgttaataa attatttaaa agagaagatg ctcgcccggg ccccaggagt taaggtaata ccacccaaag aaataaaata aaattgaatg attcagagcc aaatgtaatg cctgtgtcta taaaatgtat ggcccagtcg aattttgtat tgttcttggg agcctgtatg acgtcgagga ttgttctgga agtaaatcag aagtttgccc actttatcca attggcctgt ttcattgact acatgtgatt tattttgccc aaatgtccca tttgtaagct tagttactac actcctgtaa ttgagaccag caggagtggt cttgaacccg ggtgacagag tagatagtgt attttcaatg aaaaatacat ctctgtttat cctgctggct gttgtatggg gctgattatt acatcaacct ggctctcaga cctggcccct tggtttagcc gagactgaga ctgttttnct tgtgattctc taaagcatat caccctccat aatctagggc tctagaagcc ttattaannn atatgtatta ggaagacctg tacccaaaat ctcagattta cagaaacagg gctgtgcttc tatttgctct atgcacttct agccattgct ggttttaaga aaatgccctt gacataaccc gtgttgctct tcactgctac tgtaaaaatt tgtccgatgg cccggtgacc ggtactgtaa cctccctctt ttaggatatt tcagattttt ctagttatgc caaattgcaa ctcataatat tacactggtt tgatcagaac acttagcttc atgtttacaa tcccagaact cctggccaac tgcacgtgcc ggaggcagag cgagactcag aggcacgtgg gaaaagatac acacataatg tttcttcttt tctggcaaag .tggtgcttca tcagtttggt gctctgaggt catgcttgtg caaagaccta cctcatttta actgtccaaa cagctgaaat aagtgcttcc gattctccat tcagtctgag catattgtcc acaataaaaa nataaataat attaaaagat aagagccgac gctctgattt aattaaaata ctccaaacca agtgctctca tgccttttaa ccttccaata gcaaaaacac taagcattta gcccaaacct agcatccatt ctgtcaaggt atccgagagt aagattcact acgtgggcaa gtacctttgg tgagtcagct tgatgccact tcctacttaa ctaactacag gagttctgat tagagacgca tgctttccag gatgtttccc tagtcataaa aaaaataaaa ttattcttta ttgggaagcc gtggtgaaac tataatctca gctacagtga tctcaaaaaa cctaaaagga tgtgtccaca catctcaagt gctttttctc cagaagaaac ctttgccctc tcagttctgc gttagatgga tctacatgta gacgatgcaa ccaaagggga ttcacacaag atagcatttg actgtttcat ctctatatct atcccactca aatgaaaata agtagtngtt aattaattat gcattatgtg tgaagatcgc tattcatgtt ataatcgtga cagatctctc gactaagcct aaatataagt tagacggcat ctcacaatag agaaaataaa ttcttttcta ttttaagagg acttcgtttc atcacaagct gtcaatggca cgactcgtgt caacaatgtc agctctgggg tactctcctc gcggttctgc aaatgctgct atttagaggg aancaagttg ggcagtcttc aaaatctatg acgaagagga tcatatatct aatagatagt gacgtgggtg cccatctcta gctactcggg gccaagatca taataataaa aacctcaaag tcaaataatt taatacattt caggttacca cgatacgatc ctggtaagca tcagtcctgt cctccctaaa cacaccccca gctagaaaga aaacaagaaa ctagtatcag ttccactttg tttctataag gtgttctcac aaatctgcag aaatgtggcc tagaaaatat ttaattangt tatgtgtatg t- a +¦ a t- a t-t- t-t- f.H-aftfat af-a-ihtatat a ft- a *- a f a-t-f aatat-aH-fa a(-faaahaat taatttaatt aattatttat atgtaattat actatatcca gtatccttgt tcttcattta ctggttacca attttccagc gccaatataa tatatttgag nattatataa gtataattat aattatttat aagtattatt atctgtgtgt aattgccaca tgttggatag cttgactaat cttcataatc gctttcttga ttaanttaat tatttatata ataatttaat attttcatgt gaagtctttg ttttaagtgc cacagctctg ttatttgcct cacctattcc actagaaaga aattatttaa cttataacta tattaacaat gtaagcaata gctgagtatg tcagtaacca gaagatatgg tcaatgatcc ctctggcatt ttatacaagc ttatataatt ttatgtatgt ttaataatta taaaattatt tatcttacat catgtggttc gtcctagtct tttgtttcct atgaagatca ctgtaggcat atttggttat aattatactt tattttctaa gagataagta ttacagcata catgtggctg aggttccacc gctctgaatg agtgagataa tattataatt - 195 - 008252 accttctttt tgttagtcat acttgatctc atcccaagga atattatcgg tttttccctg tttccttcta tccattctcc catcccctta gtgttagaga catgctggga aagactcatt tacacatcct aaagggcttg ccttaatacg agacgctgtg ctcatgaaca tagtgagcgt ccttcttaaa ccagtggtta ccttcagcat caggggctgt atgaagccta ggtgagctct cagtcatcct gaagtatttt ctcagcaata aatggttctc aattcctagt acaagacaca gtactagtaa ctgagaagaa tgtaataagc aaaacaaata tagttgctgc tcttgtgaac cttatatcct agaaaagtag atagatagtc atcaattgct cacaaatcta tgagtataat gactatgatg agggagatga aggagagcct gtgttcctgt gacaacatag ataatagtgg gagatggctt catcagagag gtcttggaag acatccccaa gggagtgatg gctccatctg taggataagg ggagggcatt tcaggtagtt gtccctgtga tggaaagatg acagagctgg aagaaggata gatacatagt actagcatag tacaagattt ggatgggtaa gaaggtatca gacctcttag gaccttgcag tccaggtgaa gaattcagat tctcatccta aaaactattg gttgtcatta cttccaaaac ataaagaggg aaggtatctg caatagatct ttagctaaca tgggaagaaa aggcagcaag gcccatgttt ttcctttctc cagataaaat ttgtattagt cccttctcac actgttatga agaaataccc aaaactgggt aatttataaa ggaaagatat ttaattgact caaagttcca catggctggg aaggccccag gcaacttaca atcatggtgg aaagggaagc aaacacatcc ttcttcacaa gggaacagga gagagaagaa tgagtgccca gcaaaggggg aagtccctta taaaaccatc agatcttgtg agaactaact caatatcatg agaacaggat gggggaaact gcctccctta attcaattat ctccacctgg tccctcccac aacatgtgga gatnatggga actacaattc aagatgagat ttgggtgggg acacagttaa accatatcat tctgcctgga cccctcccaa atctcatgtc ttcacaattc aaaatacaat cataccctcc caacagtccc cgaaagtctt aattcattcc agcattaact taaaagtcca agcccaaagt cttatctgag aanaggaaag tttctgctgc ctatgagcct gtaaaatcaa aagcaagtta gttacttcct agatacaatg ggggtacagg cattgggtaa atatacccat tccaaatggg agaaattggc caaaacaaag gggctatagg caccatgcaa gtccaaaatc caatagggca gtcattaaac attaaagttc caaaataatc tcctttgact ctgtcttata tatatccagg gcacactgat gcaagaggtg ggctcccatg gccttgggca gctccaccat tgtggctttg cagggtacag accccctcct ggactggtat tgagtgtctg tggcttttcc aggcacacag tgcaagctgt cagtggatct accattctgg agtctggagg atggtggccc tcttctcaca gctccactag gcagtgctcc agtggggact ctgtgtgtgg actcnnnnnn nnnrmnncaa ccacacattt tccttccgca ctgccccagc agaggttctc catgagggct ctacccccac agcaaacttc tgcctggaca tccagttgtt tctatacatc ctgtgaaatc taggcagagg ttcccaaacc tcagttcttg acttctgttc acccacaggc ccaaaaccat gtgtaagcca ccaagtcgtg gggcttgcac tctttgaagc atcagcctga gctgtatatt ggcacctttt agccatggct ggagcagaag ctgctggaag gcagggcacc atgtcccaaa gctgcataga gcagagggag cctggggccc acccacaaaa ccattgttcc cttctaggct tccaggcctg tgatggaagg ggctgctgtg aaggtctctg gcatgccctt ggggacattt tccccattgt cctggtgatt agcattcagt ccctcgttac ttatgtaaat ttctgcaact ggcttcaatt tctccctagt aaataaggtt ttcttttcta taaaaatcat cagactgcac attttccaaa tatttatact ctgcttcctc ttgaacactg tgtcacttag aaatttattc taccagatac cctaaatcat ctctcttaaa ttcaaagttc cacagatctc tagggcaggg gcaaaatgct gccagtctct ttgctaaagc ataacaagag tcacctttgc cctagttccc aataagttcc tcatctccac ctgagaccac ctcagcctgg acttcactgt ccatatcact atcagtattt tggtcaaagc tatccaacaa gtaggaagtt ccaaactttc ccatatcttc ctgtcttctt ctgagtcctc caaattgttc caacctctgc ctgttaccca gttccaaagt cgcttccaca ttttcaggta tccttatagt agccccccac tctaagtacg aatttactgt gttagtcctt tttcatgctg ctatgaagaa atacccaaaa ctgggtaatt tataaaggaa cgaggtttaa ttgactcaca gttctgcatg gctgggaagg cctcagaaaa cttatgatca tggtggaagg ggaagcaaac acatctttct ccacatggtg gcaggaagaa gtatgagtga ccagtgaaag gggaagcccc ttataaaact atcagacctg gtgagactta atcatcacaa gaacaggatg agggaaacca ccccagtgat tcaattatct ccacctggtc cctcccagga tacatgggga ttatgggaac tacaatccaa aataaggttt gggtgggaac atagccaaac catatcgaaa tgcaacctaa agctactgag tccaagcaaa agtcaagtag actctctaat cccagataac ctcagagagg aaatcttaaa cccaaagacc aacttccctt ttctgcctaa ttgcctttgt tcttctccaa gaacctcttt ggtcttggaa ggatgctcat tctccaccgt attttgcagt ggactattgg gaaggaaagt tccccatcca nggatgctat gtgcacacat tcatgtgctt ggacctcacc aaggatctca ctggaccccc ttcgcggggc ccatctgtga ctgtcatcat ttggctaggc aagcctcctt gagagtaatc acactctggc atcggctgaa tgtcaaggtt gttattgcag ttatagctgg tgggatcttt gtttctcaaa tgaagctttc ttattcataa agtgcagtca tctcttcctg ctaatcacac ccttgaaaaa agagctggat gtgttcctga gcagggagga cctctctgaa atcagcttag aggaatgagt gtatgtttgt tcttcagtgg tgtctagaat ctgacttctg gcaatctttt ccaccatcag ccctttctct ttgccaggat tcctttacct cctatgtcag ttgaccttcc tatgttgcac tgttaccctg - 196 - 008252 tgtttttcac accctgtatg caaaagagct ggtcaccatt ttatgtaaaa taaatttcaa aatacagaat ttagctgagc aaaacagaac ccgaagtttc atttgagact gaattttaaa tctaatattt attcatgagt ttttgcagtg accttttcag cacctttttt agatcacttt cttataataa tttacacata ttaagctaat tattcagtat acttcacaga aggcttaaaa aatggaactg taacaagatc ttttgtgcaa attatgagaa gctgtcttct gtttaaattt cagaatgcat atacttacat gatgggcgct tgtgtgattg tatttaggtt tttctccatc tgtggaaaac atgtaagtac attgttaaga atcttatctt aaaggtcata tatgtgttaa tgaaagaatg gtgattaatt gattacagcc ttacctcggt tatttaggga tgtaatcaca aattactaga catcataagt caagattaac aaatattaaa cattgttaaa tgaatcagta gcttagttat cattcgatgt gtttcctgcc atttatgttg cttattcttt atattcaata +¦ +--•.••-¦*-+¦ -> ¦-. i- ¦.- v-aa u i. l. aacuaouLaa j eta. a. av. aaaaaccaat attctttctt aatgcatttt tatttttatt ttcaataaag ttaagtaact tcaaagaaaa ataatcttta atgaattttt atngtttcca tgattccaga tnnnnttttt tttttttttt tttgagatgg agtcttgctc tgtcaccagg ctggagtgca nnngtggcag gatctcggct cactgttttc tccacctccc aggttcaagt gattctcctg cctcagcctc ccgagtagct gagattacag gcatgtgcca ccacacccag ctaatttttg tatttttagt agagacaggg tttcaccacg ttagccagga tgagattcca gattttttaa tattatttgt taatgtccat gggtatattc cacatatatt tttcattttt tacaatgctg tttccttttt tattacaaac gttatttcat atatacattc catatataca cataaccaat atacttaata tttgatagtt atttagtatt ataccattgc ttaggtttgt ttttgtgaaa gtctgtttgt ggaagaaacc tgagtttact tgctgtcagt caaaaatgtg tagcccttat catagagaat tgaggagcag aagagaaaat ggaaggtgaa aatgcttagt gttactctta ttaatttgtg tgaagaggct gaaatgtttg aagtaaaaat aaaattcgcc caacgcctgg tgggttttga agaatatttt gatgtaatac cttatagaaa gcaactttca aatgatacac ctaagagtga ttttgagcca atctgcttaa agattagttg ctacccaggt cttccatagc ttaatagatt gaatattctc tctctctctc tctctctnng tctctgtctc tctctctctc tctctctgtc tccccgcttc ccacccctgg tccccaggta acgctaaaga tgctcctctt gacagtggtc gtcagcatgt acaagagctt ctttatcata gtaggcatgt ttctcttgct gctgtgttac gcttttgctg gagttgtttt atttggtact gtgaaatatg gggagaatat taacaggttg gtatatactt atctttctat tcatttatca agcaaaccct ttggtaataa aattttaaat aaacaaatgt tttttgtcta ttaaagtcta ggagagggga aagggaagta atcagttgga aattaaaata aaatagccaa gttttaaatg atatttttcc agttgaaaaa ggttattcaa ctgaaaaggt ctactttcaa aaccttagaa aaattagaaa taaaagaaaa caaaaagaat ctataattcc accaacattc agtaatcatc actccnaatt ccagcacttt ttctatctac atattgccaa aaaatgatat attattttta gatggaacaa ttttaagata actctgggtt tctttctcag caaaatatgg gaatttagag ccccccaaat ggttgtaata tatgaaaagc agttttcaac gatttgccta agttttatag atgcatcatt ttttttcaag tcctgataca acaaagtata taccgatatg tcaagttgta .ggaattagat acatggctgc cccacctttt atagcatcgt ccaagtgaaa gaaggaatca cttagttgcc aaagcctgcc tacttagtca ttcaagatga atggtaataa attagtggcc atgctagagt tgggtgttca gacggctata cttggaggca agtataaatg acaaattaat aaaaaggtat aacctttgga aaaggtnact tagcaaccaa atttgctaca acctccactt gttttggtgt tggcaaacca tgctatttca tgacaaaccc ttctataant cacattctgt taattttttt ttcttaaatt aggcatgcaa atttttcttc ggctggaaaa gctattaccg tactgttccg aattgtcaca ggtgaagact ggaacaagat tatgcatgac tgtatggtaa atatctctca tcatgaacaa ggcttagact tagaatcagc atgaaactct tncaacacaa tgttggcagg atcatccttg ttagcgttaa gactttatgt atgaaatata aatcctgaca cttcagtcca taaaattgca ttattataca ttccaatgtg tgaatattta gggcaaaata ctcaattaac ccaacttgag ctattgtagt tccattcaac aaantatgca ttttgtgagt gttgttcttt ccctcccatc tttaatattt tggtaatgaa agtcaaacag gaagcccttt aatttaatgt gttcagtgga tagttcactg cccctcagct atatgctagg ccctaggtct tcaacaactc caaggcacac tcaaggagct tacctcaagg gctctgagtt actcttcctt tatactataa agtatacttt atacttcctt cttactatat cctgtcttcc ccaagcatca tccttctggt ctcagtttaa acaggtcttt tgccttaagc taagacagcc agctacatgt tcttatgctg ctctatagtt ctcctttgta ataaggagct gtagtgaatt actggcttac attcatttat tttatctctt tctcaccgta ggacagacca atgctgtgtt aaaaaatgtc tgcagtgatg gaaatctgca ctgtccaatt cgggagccac tagccatatg tggttatgga gcatttaaaa tgtggctaac acaactgagg aactgaattt tagttttaat tttatattat tacattttaa ttaacataaa tttaaattta catngctgca tggagctagt ggtcaccata ttggactgca cagcactcaa ctgtcagtag gaagaagaga tagggccgcc tatgtctttc ttaccagtct ctccccaaac ttggcacaaa cctagcacac agtaggagtg ctgggacttc atgcatgaat taacaaggag aatttgccaa atcaaataga atacaaggca aaatttaatg aatcttaaat aaaaatacaa gcagaatgct gggattatag aagagagagc tagtttatct ggctgagaaa aaccagaaag ttctctcaga - 197 - 008252 ggtgtcggcn tgctattcca tgaatgtgaa ttgcagacat atcagggcca taattctcac tcttttgatg tggagaattt tccaactctt taggccgatt atcagtcact agctgagtta aaaggaaaag aatagcctta gccacaacat ttatgaaggg tggttcatta gctgatataa acactaaaca ctagaacttt gacttactct tttaccattc catcttgaat ttgaatcatg tagccgtcat cttgctgcca tcctgaggcc ttacncagtc agaattatgc cagaatagct ctcagaaaaa cacaacatcc cagaaataga ggcaaagtat ttcatggcca gtggatcacg actaaagata ggaggctgag tgcaccactg aaaaannnaa tttggaagaa aggaggtcgc cctgaagacg tccttatgga gaacatagtc agcagcaggg gctagcaaac atgataaact agccaccacc agaagattac ctttttatta gtatggtata tgcaatattc ctgtagagtc tgtcttttta tggtggtggc tggcaataac atttctgcta attatatgct attctctact ttgtactcca tatgtatttc ttttatagag tagcatccca tttnctatgt tgcattttta tttttccaat tatttcngac tttcttttgt aattgataaa ctctcttttc cacaacccta tcaaatcaaa gcccaaaagc ttcttgaagg tggctgatat gcccttaagc tgagagaggt tgtttaagga aagctgtggc atggattttc catagctaga gttgttaggg caaaaatcct tgtagttccc ggggtggttt gagatctgat ccatgtgaag tcccaatccc tcaggtatgt taaatctacc gtttgcagca aaagattgct cctgtaaaca taacagtgtt gcaggcacat caagttgaag agcatgtttg tttttcatta agtttttttt tgtcatatgt ttgggcctcc caatggccta agctagaaat taggtccctt gaattaaaat ggagaaaaaa taaagccaaa gaggacgctg aagaaagaag aagttatcta aatgtagaca aaggagaagt actaatgcag aggaccctgc ttaatctcca cttccatcct ggttttataa aagaacatat tgctgaactg ctttattagc ctgcctgtgc tggtttgctg ttcaaagtag atgagagagt ccaagaatgg cttgttttac gtttatggca ctcactttct ttatctgttt gaggggtgcc cttctgattg ctatgtcctg taagtgttca gattaagctt gcaccaaata gctactccag ttttaccggt tgcagaagaa tagaaaaaaa ccatctccat gaagatctag aaacagcctt caatgcctgg ctggtgactt aatggtttga tgtgtcgtgg gttcttgtga gaggtttttc ttgcttcccc tgagtcaatt agtgtgagaa tctataaatg gatatttaag tactgctcat ttttgatgac ggcttactcc tgtgcttccc atcttgtgtg ggttctgtat tggtgatctg atgaaccatg ctgcaccgaa agacacaaca agtgaataga atggaggaag gttactcaag tgaacacatg ctcagtagaa ggcccttact tgtttgacgc aatatgaaag ctcaaataat ctattggaag attcaaagct tcagtggaag ctgtgtcccc gagggaccag tactgagtga cccccatttt ttccgccgtg aaatctcttt nggactaata gaacaacaaa cccactcttg taacaatgca iyy aLjaLyuai.aa LjyjaLjaLLaBL aLLyLLLLL." attctgtcca tttcataagg attgaaaacc ggcgcagtgg aggtcaggag caaaaaatta gcaggagaat cactncagcc cattcatgat gttgattcca tgcacatgtg cgactgaatt tgatcagagt ataatgacaa tttgagaggg agcatcacat tccatgttga ctgctcagtc aactcactga aggtctgtac aacatgagtt actttatcac tatattccca tttgattttc agaaagaata cttgggattc cagatgatgt atatgtttag cttatcctct gatgaattta tgttcatttt tggatcaaga ctaatagctg ttctggaaag ctcacgcctg atcgagacca gccgggcatg ggcgtaagcc tgggcgacag tcatggaagg gccattatgg gttggtagaa gctgcagtct ggtttcttga caaaggattt ttgactccaa gctgcagaaa cttgttttaa agcagctgtc aggctcacat ctttttggac gcatgtgcac agcagtctgg aatcttaacc tgcagtttga tctagtggag tacaagataa acccaatgta gaatctattc ccttcttcca catactgggc atgtcatcat agtcatttca ccatagatag gattcactat taatcctagc tcctggctaa .gtggtgggcg caggaggcgg agtgagactc aggtcaaaat atgactntga atagcaagag cctgatcaaa aatggaatct ataatgttac attcgaaaga aatctttcat gaaatcgcca aacattgagg gattgttagc ataatgccat tgggaaacca aactgaaact cattacttca tcacagtggc atcagttcat cagcatagca tgtggctttt attaactata ccccaatgcc aacagactgt tgcctacatc actttcaagt tgattgctct tctacatgcc actttgggag catggtgaaa cctgtagact agcttgcagt cgtctcaaaa atcatcaaca ggggttcaag aactagaatt cttgaaggaa actcttgcag ataaaattac agttctattg gaaaggaaga cagccacccc caagaccctc ttttcttagc tgtgcactta aaacatgcat gcaatgtctc gtcagagaaa tacttatcac tttcattgtg taagtgaaat gaaagcttac gtcagctaaa ccaaaggttc ggaaattatg atgctaaatc ttaagtcttg tcacacaatt ttattgcctt gagcaggtct cacattttgg tgatcagtga ctcaataaga catctgtttc atattgacat agattcacac gcatgtcaaa ttgtaaattc actgataaga gatcagacta ctcttcaaat tagcaaaggt tgtaagtgct taaggtggct aagatgccct tcaaataaca ccaatgctca acccaaatct gcagagataa gttctcacta cttggcactt attgtaaatc cctttataaa cagtctctta gcctggatga agacctactg tctacctggt +--"-.-.+- ГЧ a LVJL-L.Lywl.Cia cttattattt gatggatctg attaagaaca gtcaaggngg ccttgtctgt cagctactcg gagcggagat aaaaaaaaaa ttaacaggaa acttcagtga agaaatggaa tgaagattgc aagatgctat tcgataaacc tgggtaaaat gtcaattgat aaccttcagc caccaccagc aataaaagta ataagccaca aggactttat cgaggtatac aacttgcaaa ttggggctgg cttttaaaaa attcttcttt tttgaagtta agaaatttgt agcctccgtt ctggggcact• tgcttgtagg - 198 - 008252 taagtgatgt ttattttcaa attgttgtct actgcaattt atggattgag tatgtttttg acctttaaaa tctatacaca gagaaatagc aaaaatttgg tagttgaata gcaattaccc ctcaatccag taagagttgt tngaacaaat aaaggtacac atggccctgg gaatgtgtgt ttagtccaag cttatgatat ctttttactc tggatttctt aagctgaaaa agcatttatt atcagagagg cttatttacc aactctcaac ttgtgcttac tgtaaacatg aggtgataga gca L,aa -.gg t tccttgtttt caaatmatat tgcctcacct caaatgacac tcagcagaat gaacatcctc gtaggaatca cagtatatta aaaggagtta gaatgtttta ctgctcttcc tcctcctcct tccccctcct cctcctcctc cctcctgttc agttctaggc cttacagtct atttccagaa ccatgaagga gccagggtat gaccaagaag ttgctccctc aaggtagtga cctgtctctc cccactgccc cgcccagcac ggaaggtagc gcagagctga aaggagttag aggctggaat cctcccaaat tagtagagac atccacccgc cttttaatgg gaagaggagg actcagtgca gagagcaaga ctgcaatgtg tatcctctgc cttcacatct tgagaaaccc cagggggtga gaggtggacc ctccacaatg agggctgaat aacaagtgtg gtcccaggcg tcgcagggga gtgcaccccg tgatgcggga agctgggact tgctctgtga tttaaactca ggggtgggtg gacctttgaa atgatcatta actatctttc cagatgtggg taagagttaa gaccctcatg ccaatttgga atccattatt ggtacttaga tttcatggca aggcacgtaa atttcaaaag tagccttgaa cccacccaga agccagctgg ttttggaaag aaccatacta aataatagat taattatgta ttaattataa atgacagttg ggctcttctc atatcacatt actcaatttg catgactatc gcaggagctg aaagcacttt gttttactct L L L.wa.ciaa - a ccataattgt acaatgatct gggagtgggt ttaaatggtt agaaagccat cttacatgat aaggaagcca cactgaaact gcaatcccaa ttatttttga ttagccaaag cctccttccc cctcctcttc ccacctcatc cacctcctcc cctagcaaac cagctcctgt ataataataa ggctgcgccg tgaacaacaa gagttgatga ggcagtgcac gtcttgaggg tctcagctcc atgcaataca agtcatggtc catggtggtt agttctgacc aatcctgaag tggagtccca aagtgattcc ccgggtaatt cgaactcctg catgatccaa tcctagcatc gtgtgagtac tatgcaagca cccatgcctc aaatttatca ccaaggaggt gcaccttttt acttcttgtg ggctactgcg tgtgctacga ggtacaaggc aaggcctctg gtcgtgaagt aacatttcca agggacccct acagtcccac cggttacagg tgatgagaat .ya-avja u La ggagaatttc tcgccacttt tgttgtttca ttcaggttgg catttacagt cttcaaccat aagggactct accaaaggga ttaaaagtac gttactactt tagctgcagt ttcctcctac cccctnctct ttccccccta ttctncactt actcaggcac actcacagag atagtgcagt gcaggaatga ggaaagttct agggcacaaa tcaccccaat ctttcctttc tcaggatcca cacatgaaga gccagtgttc tgtgttgcat ccagaagagg aagacatgaa ctctgtcccc ccagcctcag tttttatttt acctcaggtg cacgagttat ccgattgagt attatggccc tcagcctggt actccaaatg ctcagttttc gaccagcttt ttttttcttt ctctgccccg tgggaggctg aatggagagg aggctttcgc cagtacaatt ggggtcacat agtaggtgcc gcccactgag cccttccctg gctctggcaa ttgcctggcc LltOaLL ULLVJ attccactga ggaacatggt caaagagact agattgaatt cctctaggaa ttcttcagac gccttgggct ncaaattcat tttaagtggc accataaagt tcttcttgtc ctaccccctc cttccccctc ctctacccct ttcctccctc actgaggata aataggtgga gttaaaacta ggcctgaggg ttggggcgag ccagcctggg ttcttggtac gagccctgcg tgtaactgtc tgcacaccga tttcattcaa aacacatctt aggcctaatt ttagctcttt gcagtggtgc agctgggact tgggcttcat ctcggcctcc tagaaaaaca attacacgtg tgtgttggag gtagggtcct gtgacatccc tcttcttagt ggtgctaatt aagtgaatca tccccacgtt tggacaagga gcggcgacgt gggggccaga tttccaaccc ctaagtgccc gggctgggcg aggggagagg actgcccaga aacatgcaga tggttgcccc aa LLLLLLLL ggaggaccag ggatgataaa tatcaacatg tttaaatggn gggagacagc tttccgaagg caggccagaa gatcacctca atctgaaggt ttacccaacg attgtcgtca tnnnnnnnnn ctcctcgtcc tcctctacct tcctcttctc tagcagtgaa gacaaataat tagacagtag gtgctgtggg atctggaaat catcataaat ccctgcccgc tgcagggcat acccacggga gatgtcagta agaaagatac agattctcag ctcttgtata tttttttttt aacctctgct acaggcatgt catgctggcc caaagtgctg tgtaataaac tgtgtcttgt gtggcgtctg ggggggctga agaaagagaa gcacttaatt atgctagctt tcgaggcctg ccgcgtcaag caagctcctg caccttccat aaccacagct aaaatgccac tttcctgccg gctgtgccct tggaagggat agagctgagt gcacctcact gttggctaaa LLLLuaaaay cttttaagtt agagaggtat tctcaaacac ntgaanttgc atcgttcctc ccagcaggga aaagaggtga aactcaagta gtgtattgga tttcttaaaa tcaacctcca nnntcttccc acctcctctt tctcctcccc tcttcctcat caagatggaa aaaaagtgaa ttggggttgg acctcatgtc agttcaagaa gaggggctcc ccaggcttca ctgagctcac acctgctttc actggcaggc agccaggggg tgtctggatc tagtttttct ttttttgaga tcctgggttc accactatgt aggctggtct agattacagg attgagacgt ccattncctt tgggacagnc aggggagctt gcaccagcag ctaccaggct ttgagaaata atgatttgct ttcctgctgc tttaagcaca gatgtcctga gtcagccttc atctgctgtg tcctccagtg gatgtcccgt ctgggagagc gacttgcttg tgcatgggtc acaccagatt - 199 - 008252 ttccagacat cttcttcctg ctgagggatt atggaggtat ctcccctggc cagcttggtc tgtgtgtgtg aggagtccat tgttctattt tttttttggg acagaggccc tttgaaatgt aatatccatg gccttgactg catttgagct ttactttatc cctttgtaat ataagctcca gtagctattt tcaaatttca aaaaggatca cctcccacta aatgcatgtg agtggcaaat atattcagcc gtgatggggc aacagtgaag gaagagaaaa aatgtaccag ctcacagaag aatcctgtgg gg~ggac-gc tgtgttggct gatattcaac tggctcacac aggagtttaa ctagctgggc aaatgcttga gcctggtcaa atacagaagg tgacgcacag gaggtgcggt ctttacaggc agtcattaaa gggagacaca aaaacactag gtaaatctca cataccggtc agctggagta agtgcctgaa ccgccccgcc ttcatgagcg ggagacttct taaacaacag tgcggggctg tgctggaggt tcccttcatc tatctatgtt tctactccac ccaaataacc aggaaacatt atggacttgc gtcattctct tctttgttac acatcttaaa ctgctagatg actgccgttt acggtgtatt ctgccacagg taatcacata ggagaactca gatcagcaca tccagttctt aatcacaaag ttttttggtt tctttggaac tgtctctccc ataaaagcaa ttcatttttt tgctctcaag tgggtagctc ctacacctgg •b "*-* -a /*r+* у*"/-" пыт i_ w"_-i^ h. v- we* у у ttacctacaa cctgggtttg ctgtaatccc gaccagtttg atggtggcac acccaggagg caagagtgaa atgaatgaga ccgagcatat cttccttctg acttagatgg taaaggagtt gagaggtgac acttattaga ggtgaatcag cgtggacatc caccatagag gcgcatcaga ctgccccgca cccagacagg gtatttgttt aaaggtatgg attccttctg gcgctgccaa ttcacatggc caaattcctc tgtgatctca ttacgttctg taacccataa aagggggaaa tccatgatga caggagaaat atatcattta gtattattta taaagtaaat tcattccgtg ggtccatgcc gcagtaccta ggtgtacaga ggaagaatct ccaccaacaa gaggtcatct gacatgaggc acaccgatgt cttcctattt tgaagctttg ttccctctgt agcctccagg acaggtgcac gtgcatcaca Г* f_ -t" <^ /-"¦¦" *_ I-• -- t- > _- I", ^ Ы ctgcatgggc tttggtagcc agctctttgg accaacatgg acacctgtaa cggaggttgc gctccatctc gaaacagaga taaccatcac cagagaaagt cacttactac accaggacta aggaccagcc ctgagtgatg agctgcgatt cggaagagct gaggaggtgg gctgtgagtg ggaggatgaa acagagggtc ccttgggcta tctcgaagtc aggcctgtga tgttggtcat attctgtttg cttttataag ttttaactta aggtactaga cagcttctta caattacatc acgtaggcaa catagatatt tgctcaccag atggaataat ttgagaattg catttaaagc acagaaaaaa caaggagtgt aatatgcttg gggaaaagtt gctaaaaagc ctgtcagaat agataggcat atctaataaa attcccctag atgggattct ggtgggaaga ctgtgatttg cagggtgcat ctgccaagag /"--Г/-"+-/-"Гл-"/-"+* +• -5 +- -•^W-.^WWWt-.C-.'.-. ctgctgtgtg tctgaaatta gaggctgagg tgaaaactcg tcccagctac agtgagctga aaaaaaagaa gggaagtggg ctcggggggc cgcgagcctt tataagagcc tctatcatct caagacatgc tcctctctgg gcaggtgtgt tgcagctgga ccaagcagac aactgcacac agaaaagagc aaatggagtc ctgtaactta ctggatgctg gggagaatct tccttggctt tgtgtgtgtg aacatctgtc attactaatt ggttaggact agcaggagtc tttattttca caaaccacag ttcagatgac tgcttctgag aaaggaagca tattttaata attatttcaa ataagagcga gtgggatatt tggccagaag ctctcaagtt agaaaaacat gtttctgagt actgttgtca aatagaccca cccagtatat tgatgagtca gatttagcaa ggctctagga gggtctgagc gggctgtctt <- у wwww" v у fc* tcaagggtta ataaggaaga caggtggatc tctctactaa ttgggaggct cagcgcgcca aaaattaatt atgcagggag taagtgtgaa ttcctagcct tcacagatat ttgttttaca gggtggcgag attttgacgt ttctcccacc ggaactcctg catccgcatg atgcaggttg ttcgtgaaca tctgcctttt gcaccttaag gaagcccgag agcctagcct gtagaagcat tgtgtgtgtg gtattacatt acatctgcaa ccaacatatc catggataaa ctaatcactc tcgcattagc ataactgact ttatggtagt cttacttatg taattggttt gaaatgtccc ctgactagca gtgttgtgtc ctgaaaaaaa atcaatttca gggacggaga ctttcctcct ttaataatta ggcaaaatct gataaaaaca gggaagagag agagaagccc tggaactgag cataacctgg ctcctctggt /-¦•> гтч- a a rt r* caw *¦" w *"*b*--jw aatgggagtg ccaggcatgg atttgaggtt aaatacaaaa gaggcagaag ttacactcca aataaggaag tagggatgtg gaaagtatgg ggatcttttg taactcactt gacgagggaa gggtggcgtc atctaagggt agcatgcttt gcgagggagc tggctcaaga ggtccccgcc tgggattagc tccttctgtg atcagccttc tatcccttta aactatgtcc actacctcca cctacagtgg +¦ r*a rra t" ra t" rr **it-rrr"arTar--t"t- trart--tanr* nr*a na rrl- r-э rr r-r"r-t- nnrr-t r- tccattagga gtttattggc gggagctcaa catctctact actcaggagg gagatcatgc aaaaaagttt ctagaatttt caaactgtga tgcttcctga gtttccatgg caggtgtggt ctcacttgag aaaaatacaa ctgaggcatg cactgcactc attgatatta ccacatattg taatgtcatt cttataagcc aatcccattt ggctcatgcc gccaggagtt aaattaacca agaatcactt cagcctgggt aaatatgatg aatttgcaaa tattctgaaa tataagattc ttataatatg tgtaatccca ggagaccagc ggcgtggcac gaacctggga gacacagtga tccatttatt actactttaa agaccacctc tctaactcat acatgaagac gcactatgag ctgccaacat acatgcctgt ggtagaagtt gaatctgtct aaactcaact attgctgcta tcaagattaa aaattaatga ttaaaaagca aggtcaaggg ggcaaaaccc agtcccagct gcagtgagct caaaaaaaaa gttcaaaatt aaaatttcaa tgatgtttcc atgtatactg - 200 - 008252 cattgacata ttttggcatg acagagattg tctattggag cagctgccca aatctgtcct acagactggt ctcatccacc attgtattgg tagtaatttt ttgggaaact agagtgagac agtcatgcac ctaggagttt acatagtgag aaaaagaaaa cacaaatgaa cctaccttaa cacccgtcat caggcaggat cagtcgtgca ctgaacccgc gacaacagca gggcttcttt gtgacaatat tgatccagat gtctgactca tatttttgtc catggagtat tagaaatttg atcagactca tctatgtatg gctgtgagaa aggttaacaa catggtgtgc cccagcaggg gttttgggct ttagtcaacc gatggttggc gacagggcta aagcctaaga gaccaaggct tgtgggaagt cctccaccag gtgtggatct accttccttc aaacaatgac tgtcccagga gtgacccttt cttcccactt gggttaaaaa ctcctgagcc aaaccccaga ctccagcgat ttcaaaatgc tcttgtcaaa attttccggg gattcagnaa atgggcaaaa cgaagttctc ggtcactggc acttacgtag tactgtattt tttcacagac tttcaatcct gagaaaaggt gtgatgtccc cctctggctc ctcctggatt caaggctctt cttaatgagc aaataagaca gaggcgggag ctcctctctn acttttagtc aaggttacag atcctatcta acaaataaat aatggactgg ttgaacattg gcaggtctta gctgggctta gtatcatcca ccagcatcga tgcaacctga cttttaataa ttctaatagg tatgtctatg tatgaatgcc taaagagtta ggtttacagg cagtaaaatc ttgaagatat **•/¦-+• *• гч^гчгч^г* tgtctatata gcagctggct cgtggagctg agcaacgcag tgctccagga ccccagcctt aaggggcaag aatgtctcgc atgaggttgg aagaaagagg tttgttagga cttggcatct ggcccatcat ccaggctggt ctgcagttcc attctcaggc agagtggtgt atacccattt gtcctcctgc gacggtaact ccacgctgtc ggaaatttgg cgaggcattt aagtctgcat aacattgtcc tacatttcag gagggagaaa gtattcaact aaaaaaggga tttaggcaag tactcactgt acagtgaatg agggaaaaaa cttcctcaga tcccagagaa atcactgctg tgagagggca gacctgaaaa tgcctcttac ctagaataaa taagctgggc gatcgcttaa gnaaaaaaaa tcagcttctc tgagctataa aaaaaaaaaa aaaatacaag gaagtcaatg aaattagttc ggagagggga ttttactgaa cagcctgaga gaccacccag ggtatgggca aaataaataa aatattttca tctttgccat aaaaaaagtg acatcagctc aagctctgcc agtgaaaatg atcctttcct -.+¦ -> +-¦*-¦• aaaaagtaca gggaggacct atcccgtgct ccaggggcca ggcccactgg tagaaatcat gggaggcatc atgggagagg agaaagtgat tgacnnnaaa aagggagagg ctctttcttt acagctttgc gacagtagac ccttccttac atggagggga ctccttcagc tcacttttcc catgtgtttt gtgtgcttct ggatcgagga gcagtggcgt taaaggtatt ctacagttgt cccaaaataa gctatcatgg tttccaaggg gaagagggag caggcccccg ctgctttacc atgccagtca tttctatgag cccaaagcaa gcaacaacgt aagatacaac gccactgtag caaggagtcc cttcagaacc aacttgcagg ataagaggag acagtgtctc gaccagaagt aaaaaaaaaa aggaggctga taatcgcacc aaaaaaaaaa catcaatgac catacctctc aggatcctta gaagaaaata caaacaatcg gagagtcagc cccagtgagg cgaggcgtcc aatgggtgct ttaaaaaaaa cttggatgca aatgccctga taagggctgc ctaaatattt tttgtaatta ggttgggtat +¦ n •_" /-T-b r* _" •" 4, V-" V-W_A aggagttatc gcccctttat ctgtgcagag gagtaccttt tgccatagct ctgccaccga tgcagtggcc gaggcaggct gatttcagag aaaaaaaaaa tgacaggtga cactgtagct ctgcagagaa tgcccccgcc cccctggttt acgaagcagt agggcgtgca ccattgtgga cctttcttat cctcccatgc ggaagtcggc ctgccctcag tactctgttt ttacagacag cttctctaaa ttgagaagcc ctttctgtgt tgggcagatg atttctctca tccatacaat ctcttctaag ataattactt tcactaaaca caattgcaga tcaacctcct ctatgttagc tggctttagt aagacaagca aaagtgggaa aaaaaataga atgcctgtaa ttgaaactag aaaattaatt ggcaggagca actgaactct aancagaaaa cgtcactatt taagcccacc cttggaaatt agatttgtgt catttccata agcaagagct acacgaatgc tctgtcccag ggtcagatca aaanggcaac gttgaggtta acttcataat atttgaatct tcctcccttc gctttcaacc cagaagaaca goat at- tti t с agcgtcctgg gaggcagcaa cccccagcag ggaagcctaa cagttttcag ctgcagggac ttgccactgg gaagtcagga ccctctgttt ggatactaaa acctgagaac tagattgtta ctctttccct cagccggtcc gccagcgaag gggggttacc gtggtggctc cactctgagt agtaaaagga agacaagcag aagatgcagc gtaggtccat gtgtgttttg caacataatg tatgacttac agcacctatg gcctcaaggc caatcattca gcacactttg aacaannnnc tgcattactt ttatgaatgc tttatatata aacatgtatt atagcaaggc tccagggatc atatgccgag caaaggtttt aggggttggg atagccacat tcccagcaca cttgggcaac agccagccac tagcttgagc agtctacggg caccaacaac ctgattgtta tttgcatcag cctaagaggt ggcggtgggg gaaacagcag gagccggttt caacagtcag gacactctgt ttcaaacaag ggaaatccat tttaaaaaga tctgtagttt gggtatatta tctaaagtca tccttaaata tatggaattc SttttC233C agagtcctgt ttacgatttg caccaggctc ggctccacgg tagatctcat ttctgcggag tcaccctgaa cagacagtga ttagatgaaa gagatgtgaa aggtgaaatg tcagctccat gaagctctaa ccactgcccc atatgaaggg caggacaaga tttttcttct ccatgttgtt ggagagcgca ccagcagcag cgacgcaggg ccaggcattt ggtcttgcct aaaaatgtag attagccccc aaaaagacaa atgctcaaac gaaaatgcca cagctgagcc aacaacaaac gtgttctctc - 201 - 008252 attcaatcct ttttcttcta gcactcagga tttctataga cccaacttca gacagcacag ccaaggaaag aggttcctcc tgccaggtca tagcaaagat gcagcaaagg ttaccgtggt agggacagat gacaaaagaa tgtaatccca accagcctgg ggtagtggtg agcctgggaa agagagtgag actggtccat ggggtcaggc aacaatgtca atcctgacaa tgggttcaga caacaccatg cccctcgcct taatgaatgc ttttctagtt atgacatcat atcaagtcct atcactgatg ciy t CJCJQ uCat cagtcaactt ccatgactga ctgtggagag atgaatttct ctgtaattga tcctgccaca aaaatttttg atcacttaaa ctaatgcagt gggtacaggg tgagagaatt gatagtagtt tatattttta ttatgttaat gaaatggact catagttgcg aagaataaaa ttggcttcat ttctgggtga gaatcttcta ttaataaaca tgagcttctt atsgagggag taactaagtt gcccaattga cttacatttt tatccccttt tgattcaaac taaattacaa ttctgagcag agggaattgt gtgagcacaa aggagaggtg atgatgctgt gcactttggg tcaacatggt tgcacctgta gcagaggttg aacctgtctc gcctaggtgg tgagtttgag caggtgtcat tagtctaatt gacgtgaagt actgtgaggc ttactgtggg acaggtttct tttttaatta tagttgtcca gaaaaggatg ggaatatcat -.-.4-+-UVJLJU V L.U L L acggtttgga cgcggcttcc cgacgaaagt agtggtggaa tctataactg gaaaggcata tataatgaca aatgtcatat attatcaccg gttatagctt tattatttat ttacaaacta aatatacaca gaaactgttg caggatgctt caaagatgtt ctatatagct ttcttaccca tcctaacaca tggaaataat 4" +- =1 Г-*Г 4- a --+¦ -> gtcccattat ctctataggg agcaatgctc catgctaaag aaggagaaga gaggctgacg catatccagg acagcagaga gctgagtcgg tatgtgactt ctatgacaga ggcaacaaag acctggggag caaaagtact aggccgaggc gaaaccctgt atcccagctg tggtgaacca ccaaaaaaaa tccccaccaa gaggcagagg gctatgcact aggacatttt tgcccatagc cccagaacca tcacactctc gtgacactta tttcaaaatg tgagcacaat tttacttagc gggtttttag tttccatatt ttttggaaag cccacagccc caggagcttt tgtgtagcat atgttcagtg aacttatttt atgacagcat ttcaatttta ttggaatctg gcctggaggc gcatggacag aatgcacagt tccattgggc aggtggatca ctctactaaa cttgggaggc agattgtgcc aaaaaaaaaa aaacggtgat agaagaaaaa ttcctaagtc cattgtcttc tacatatcca cacctttcct cttccttttt atagttcccg aaaggggtga taaaaaaaaa atctgtggct gagtgattac aaui> 4._a-avj wwwwaaczG-ww ggaaggacaa tgcggttctg ggggatgacc accgttttct aattccagct agctgcatgt gtactgataa attctgtctg ccccgcaaaa tagttcacat gattgatctg cagattctga agacatttgg cctccttcta agcggtttaa aagtttctta caatttattt aggtatgtct ccattgctat ggtgccctgc rtl- 4- -H--T*- +• <-+¦ -n- ttactgattc ccaggataat agtgtccttg ataatgctct atgtgttcca ggtcttcact tcagagatca caagacaacg tcctgtaggc tgctttagga aggttacaag agctgggggc ccttggtatt caggcgcagt tttgaggcct aatacaaaaa tgaggcagga gctgcactcc aaaaaaaaaa cgtgaaatag tagtattttg atctcatgta atcttacaat caggttgcag cccataccct gtttttttta aattggaatt gcagcacgtt atctatcgtg aggacacgtt aatagggtga autuuCjCCut tgtcgtgtgc gtggtggacc ttaggtggat ttgattgtcc tgtttataaa actatcaatc gataccgcaa ggtatatatt cccaaagtgg tccactagaa cactgaactt gtatcttctt aagttgattt gtgtgcaagc tagataaatc taataactga ttgcatgtgt aaatcagcag aaatgacatc tctgaactca tr-t- i~i~? *-=¦*- 31 4- agtaagagaa cccattaaga tacatgtttt ttaactgtgc caaagcagat gggactacca gagaagatct tagatggtgg aattaggagc agttgacttg ctggaggaag cagatactgc catttaaatg ggctcacccc ggagttcaag ttagctgacc gaatcacttg tgtctgggtg nngtacttcc acatgctctt agggggcagc acctgtttta ggaggaaact aggtgggtcc cactgcatcc ttgcatatgc tttttagtga tttttcccaa aacataaaga agccaaaagt agcaatacca L-V-U'wj t- \j WWW.. aaaatgaacc cggcagctga gtcaatgtag agtgagcaat ttgattttca attgctcaaa gcaaatgtat catatgtgct ctgatattta atatttctct atgctaaaat tttctcgtgt ctgtcctcag accaggcaaa acttgagtga gtttagcaaa gatgttatca agtcatttca tctgttaaaa caggttaggg r,tc3atrir*cc gatttcttgt attacagatt tgtgacctcc ggcaaatagg atatgattca tttgatttga atttgtaaaa cactcggtag aagctgtgtc cttgtgctga catgtcctcc gctgggccag ggacatttgc aggttgtttt acattccttc ttgtaactat tgttcacttg taatttactc cagcacattg cctgatactc gggaaaatta ccaagtagaa tgttcagatg ataatctatt tgtgtcacag gagataaaga gagtttactt tgagttgacc caaacctgga atttcaactc gaaaagcaaa gtggacttga ttttttaaca tttctctatt aagtatcgtg acaaccagtc tttgtacata atttttaaac ttgaatattc aattctctga atgattacaa cttggacctt gagggattat taacattcct gtagtggcag atgtggccaa caagataaaa ccttttcttg agtccataat gccagaatgt agtgctgtta taactatttt ctcagaatcc cagatagata tctacagttt aaggattaga taaatattgg gtggggcagg tagttggcaa tcctttcaca - 202 - 008252 taaggctgag ggtccagggc acttgagtaa ttgatgttca aatgtattta tgtaggactc taagcgccac acacagatgg tactaatcct ggcaccagaa ccacaca ug. aatggaaggg cggagtcttg ctccacctcc ggtgcccgcc atgttagcca agtgctggga ccagaccaac aagggtacat gtggaatgat tcagtatatc accagccaaa gtggtctctt gtcctaaccc aattacttat tcctctgaag ggagctatgc aaggcgcctc cttctagcca ctgctttttt ccctttccaa tgaataccct atgggtctgt tacagcttca gcagaacatc aagccatagc ctctctacga ccccatttag gaaaacttgg gtcatcacgc agcatatggg gtgccctcta actagtgctg ttctcacgaa attgttgttc accctatatt ttttcctgaa attttacccc gtcacagtgt cacccactgc gtggaatact agcctacaat cataaaaaca actgctgagc attctgcagt ttcaatgaag acaggtagag tagatatgta tccctggtcc ctttaattag tacttgactc aggagactca nnnnnnnnnn tgcagtggca gcctcagcct ttgtattttt cctcgtgatc gcccggccca tggctgtgcc aaagccttac aggaagtggc ctaaggcctt agttgaggtg gaagtgtgac cttattagga gggtaatccc tggccatgag ctgggctacc atggccccct atttctgttg ttttagttta gaacagaaaa agatgttttt cctttctaaa gaagccaggg cgaaacagtc catatgcaag aacatatgaa gtagggaagc agggtgctgt agagcccctg aagggaacaa tttttttttt cgatctcggc cccaattagc agtagagacg cacccgcctc ctctattctt ttggccaagc aggaaagcag attctataca caacagatcc aagatgaaaa ccccagaagg aatagggtct ttactccaac acggtgaagg agaagctgca gtcaacacct ttttaagctg gtctctgtct ggcacttaca ggctcattac acgtctggaa gcagaatacc acacacttcc ccagagaaat aaatcccatt cagacactga cagttccaaa agagagcagg ggatggacag ttttttgaga tcactgcaag tgggactaca gggtttcacc ggcctcccaa aatggtactt tgaatgagat ctttaagcca tgggtggttt caaaatttta cggcactcat ataggttgaa tcgcagatat atacctggta cagaggcatc agaggcaagg tgattctgga cccagtttgt tctactaact ccaatgctat ctctgtcacc cgggttcaca accatgccca ggatggtctc ttacaggcgt atacatcagc agaagaaaaa gtgtttgcct agtaagcagt agccaggcag ctcacccact ttagtacctg gaggaggtca acacaggcat tgcccagagc ccagaggctt gaagaactgt atcattttat ttctacttta tcactctann caggctggag ccattctcct gctaattttt gatctcctga gagccaccgt agtgtctggc ccagtgttat gtttccttgt ggaactgttt ccctcgttga attatgggtt tgaatatgac tactagagga gtggggagga caagggacac ctaagggaga gagaacatac cattttagtt <210> 4 <211> 6799 <212> ДНК <213> Homo sapiens <220> <221> 5'UTR <222> 1..65 <220> <221> CDS <222> 66..5282 <220> <221> 3'UTR <222> 5283..6799 <220> <221> полиА-сигнал <222> 6081..6086 <220> <221> полиА-сигнал <222> 6777..6782 <220> <221> аллель <222> 1658 <223> 99-79316-158 ¦ <220> полиморфное основание С или Т - 203 - 008252 - 204 - 008252 - 205 - 008252 aga cct саа Arg Pro Gin 640 cct aaa ate Pro Lys lie сад дас аса Gin Asp Thr tec tgc gac Ser Cys Asp 690 gac caa aaa Asp Gin Lys 705 ctg gaa aag Leu Glu Lys 720 сад cgc atg Gin Arg Met tea ate etc Ser lie Leu aga Arg act Thr aga Arg 800 gag Glu cat gga His Gly 770 ttg act Leu Thr 785 gaa gac agt gaa atg gtg Met Val agg gag Arg Glu 660 tgt tgc Cys Cys 675 cac tec His Ser ctt cgc Leu Arg gag acc Glu Thr ctg age Leu Ser 740 age gtg Ser Val 755 tea aac Ser Asn caa gat Gin Asp atg aaa Met Lys tac ttc gat Tyr Phe Asp aac ttt tgc Asn Phe Cys 850 gac cct gtc Asp Pro Val 865 gat ttg ctg Asp Leu Leu 880 acc ate tgc lie Cys Thr agg aaa Arg Lys 820 aag cca Lys Pro 835 egg gtg Arg Val аса gga Thr Gly gga ttg Gly Leu tct tgc Ser Cys 900 aaa ate Lys lie 645 agt ttt Ser Phe etc ctg Leu Leu aaa cgc Lys Arg aag tct Lys Ser 710 gca gtc Ala Val 725 gga tea Gly Ser сад cat Gin His age сад Ser Gin cat tec His Ser 790 ata aag т i _ т___ 805 gtg caa Val Gin ctg ttc Leu Phe gtg gtc Val Val get gtg Ala Val 870 gtc act Val Thr 885 att tec lie Ser tea aag Ser Lys atg aag Met Lys aga Arg tea Ser 695 gtt Val age Ser 680 gca ttc Phe act aaa Thr Lys ttt gag Phe Glu cat ate His He 760 agg ate Arg He 775 aat аса Asn Thr ctt cct Leu Pro 650 сад ttt Gin Phe 665 etc ccg Leu Pro att gag age ate Ser He ate tta He Leu 730 ggg сад Gly Gin 745 cgc caa Arg Gin age agg Ser Arg gtg aga Val Arg tea gat ttt Ser Asp Phe att gac cgc He Asp Arg acc Thr gac Asp agg Arg 715 aga Arg acc Thr aac Asn 700 gca Ala tct Ser 685 aaa Lys agg Arg get tgc Ala Cys ccc gca aag Pro Ala Lys atg att сад gaa gag cgc Glu Arg gga aaa Gly Lys 780 tat aga Tyr Arg 795 aaa aag Lys Lys agg Arg 765 tct Ser aat Asn аса gtt Thr Val 655 сад caa Gin Gin 670 tec tec Ser Ser tac ate aac ctt Asn Leu acc cga Thr Arg 735 gag agg Glu Arg 750 tea eta Ser Leu ctt gaa Leu Glu gca caa Ala Gin gag caa gca gaa gag Glu Glu att gtc He Val 840 cga gca Arg Ala 855 aaa aat Lys Asn tac ctg Tyr Leu atg atg Met Met gaa Glu 825 ggg Gly 810 etc Leu cga Arg aga gag aac Arg Glu Asn gaa cac Glu His cgc ttc Arg Phe аса aag Thr Lys gtc atg cat gca cct act ttg сад ttc atg age Phe Met Ser 930 ttc act cca Phe Thr Pro 945 ttt ata tat Phe He Tyr 960 915 att gag He Glu act get Thr Ala ctt gtg Leu Val ctt aat Leu Asn gtc ate Val He 950 age ttg Ser Leu 965 ctg Leu 935 agg Arg att T 1 -. 4 1C 920 aag Lys gac Asp gac Asp ttt Phe 905 get tgg Trp 890 gag Glu aac Asn tac Tyr 875 gtc Val gca Ala 860 cat His atg Met agg Arg 845 tct Ser caa Gin ate He cac His 830 ttc Phe 815 cca Pro aga Arg tec ccg ttt Ser Pro Phe gag tat gtg ttt ata ttt He Phe att atg He Met ttc ggt Phe Gly ctt tgt Leu Cys 970 925 gca gat ggc Ala Asp Gly 940 gga gta atg Gly Val Met 955 tgg atg cct Trp Met Pro aaa аса Lys Thr ctt tat Leu Tyr ate gta He Val 895 cga aga Arg Arg 910 gtg ata tta ttt Leu Phe gac ata Asp He caa aat Gin Asn 975 - 206 - 008252 - 207 - 008252 ttc tec ate tgt gga aaa cat gta acg eta Phe Ser He Cys Gly Ьуз His Val Thr Leu 1315 1320 gtg gtc gtc age atg tac aag age ttc ttt Val Val Val Ser Met Tyr Lys Ser Phe Phe 1330 1335 etc ttg ctg ctg tgt tac get ttt get gga Leu Leu Leu Leu Cys Tyr Ala Phe Ala Gly 1345 1350 gtg aaa tat ggg gag aat att aac agg cat Val Lys Tyr Gly Glu Asn He Asn Arg His 1360 1365 gga aaa get att acc gta ctg ttc cga att Gly Lys Ala He Thr Val Leu Phe Arg He 1380 1385 aac aag att atg cat gac tgt atg gtt cag Asn Lys He Met His Asp Cys Met Val Gin 1395 1400 gat gaa ttt аса tac tgg gca аса gac tgt Asp Glu Phe Thr Tyr Trp Ala Thr Asp Cys 1410 1415 ctt atg tat ttc tgt tea ttt tat gtc ate Leu Met Tyr Phe Cys Ser Phe Tyr Val He 1425 1430 aat ctg ctt gta gec ata att gtg gag aat Asn Leu Leu Val Ala He He Val Glu Asn 1440 1445 act gag gag gac cag ctt tta agt tac aat Thr Glu Glu Asp Gin Leu Leu Ser Tyr Asn 1460 1465 atm ata tgg aac atg gtg gat gat aaa aga aag atg etc etc ttg аса Lys Met Leu Leu Leu Thr 1325 ate ata gta ggc atg ttt He He Val Gly Met Phe 1340 gtt gtt tta ttt ggt act Val Val Leu Phe Gly Thr 1355 gca aat ttt tct teg get Ala Asn Phe Ser Ser Ala 1370 1375 gtc аса ggt gaa gac tgg Val Thr Gly Glu Asp Trp 1390 cct ccg ttt tgt act cca Pro Pro Phe Cys Thr Pro 1405 gga aat tat get ggg gca Gly Asn Tyr Ala Gly Ala 1420 att gec tac ate atg eta He Ala Tyr He Met Leu 1435 ttc tec ttg ttt tat tec Phe Ser Leu Phe Tyr Ser 1450 1455 gat ctt cgc cac ttt caa Asp Leu Arg His Phe Gin 1470 gag ggg gtg ate ccc acg 1475 1480 1485 ttc cgc gtc aag ttc ctg ctg egg eta ctg cgt ggg agg ctg gag gtg Phe Arg Val Lys Phe Leu Leu Arg Leu Leu Arg Gly Arg Leu Glu Val 1490 1495 1500 gac ctg gac aag gac aag etc ctg ttt aag cac atg tgc tac gaa atg Asp Leu Asp Lys Asp Lys Leu Leu Phe Lys His Met Cys Tyr Glu Met 1505 1510 1515 gag agg etc cac aat ggc ggc gac gtc acc ttc cat gat gtc ctg age Glu Arg Leu His Asn Gly Gly Asp Val Thr Phe His Asp Val Leu Ser 1520 1525 1530 1535 atg ctt tea tac egg tec gtg gac ate egg aag age ttg cag ctg gag Met Leu Ser Tyr Arg Ser Val Asp He Arg Lys Ser Leu Gin Leu Glu 1540 1545 1550 gaa etc ctg gcg agg gag cag ctg gag tac acc ata gag gag gag gtg Glu Leu Leu Ala Arg Glu Gin Leu Glu Tyr Thr He Glu Glu Glu Val 1555 1560 1565 gec aag cag acc ate cgc atg tgg etc aag aag tgc ctg aag cgc ate Ala Lys Gin Thr He Arg Met Trp Leu Lys Lys Cys Leu Lys Arg He 1570 1575 1580 aga get aaa cag cag cag teg tgc agt ate ate cac age ctg aga gag Arg Ala Lys Gin Gin Gin Ser Cys Ser He He His Ser Leu Arg Glu 1585 1590 1595 agt cag cag caa gag ctg age egg ttt ctg aac ccg ccc age ate gag Ser Gin Gin Gin Glu Leu Ser Arg Phe Leu Asn Pro Pro Ser He Glu 1600 1605 1610 1615 acc acc cag ccc agt gag gac acg aat gee aac agt cag gac aac age Thr Thr Gin Pro Ser Glu Asp Thr Asn Ala Asn Ser Gin Asp Asn Ser 1620 1625 1630 atg caa cct gag аса age age cag cag cag etc ctg age ccc acg ctg Met Gin Pro Glu Thr Ser Ser Gin Gin Gin Leu Leu Ser Pro Thr Leu 1635 1640 1645 - 208 - 008252 cga gga gga Arg Gly Gly 1650 aaa ttt ggg Lys Phe Gly cag agg Gin Arg 1665 age cat Ser His 1680 atg aaa tct gtc gtg tea gtg tec Ser Val Ser agt egg Ser Arg cag tgg Gin Trp 167( tea gtc Ser Val 1685 tgc aaa caa gat Gin Asp 1655 cgt ctg Arg Leu gca gee gac Ala Ala Asp gca ggg Ala Gly 1660 cca aaa Pro Lys ccc tea gee Pro Ser Ala I 1675 aac tta egg ttt gga gga agg Asn Leu Arg Phe Gly Gly Arg 1690 atg aac ccc atg act gac gcg aaa ccc Lys Pro cca ata Pro He аса acc Thr Thr 1695 get tec ncu uya oci voiJ. vetJ. i_.ys J-iys hkl паи r.u net 1700 1705 tgc ggt tct gaa gtt aag aag tgg tgg acc egg Cys Gly Ser Glu Val Lys Lys Trp Trp Thr Arg 1715 1720 age gac gaa agt ggg gat gac ctt ctg gat att Ser Asp Glu Ser Gly Asp Asp Leu Leu Asp He nil. i-u.a rtj-a _> -=x 1710 cag ctg act gtg gag Gin Leu Thr Val Glu 1725 tag gtggatgtca 1730 1735 atgtagatga atttctagtg gtggaaaccg ttttctaata agcaatctgt aattgatcta taactgaatt ccagcttgtc ttttcatcct gecacagaaa ggcataagct gcatgtatga ctcaaaaaaa tttttgtata atgacagtac tgataatatt atgtatatca cttaaaaatg tcatatattc tgtctgcgta tgtgctctaa tgcagtatta tcaccgcccc gcaaaagagt tatttagggt acaggggtta tagctttagt tcacatcttt ttctcttgag agaatttatt atttatgatt gatctgaaaa taaaatgata gtagttttac aaactacaga ttctgaattt tcgtgttata tttttaaata tacacaagac atttggtgac cctcagttat gttaatgaaa ctgttgcctc cttctaagaa ggcaaagaaa tggactcagg atgcttagcg gtttaaaaca gagtgacata gttgcgcaaa gatgttaagt ttcttaagaa agcaaaaaga ataaaactat atagctcaat ttatttaaaa ttatcattgg cttcatttct tacccaaggt atgtctgttt atttcattct gggtgatcct aacacaccat tgctatgtta ttaaaagaat cttctatgga aataatggtg ccctgcaaaa ttagggatca cacaacttac ttaatcgttt tttgtttttg atggcccatg tcctccggga aaattagaaa agcaaaatga tcttgtgctg ggccagccaa gtagaagtgg .acttgacttg cagattggac atttgctgtt cagatgtttt ttaacagagg acctccaggt tgttttataa tctatttttc tctatttaac aataggacat tccttctgtg tcacagaagt atcgtggtag gattcattgt aactatgaga taaagaacaa ccagtcatgt atttgatgtt cacttggagt ttactttttg tacatacaag gtaaaat <210> 5 <211> 1738 <212> PRT <213> Homo sapiens atgtccttga acaagatgtt tgggttacta agaaatgata aactaaggta gctaagccca cccatttcca ggtcagcact taaaaagtat cagaacaagt aattgtgtgt aacctgtaga accttttaat aaatctttgc tgccataaat gatttgaaat tcttcctctg ttttttttcc ttacaaagtg gacctttaac gattatctca attcctcaga tggcagtcta ggecaaaagg ataaaataaa ttgtccagtg tataaattga tcaatcattg ccgcaagcaa tatattcata aagtggctga ctagaaatat gaacttatgc cttctttttc tgatttctgt gcaagcacca taaatcactt aactgagttt atgtgtgatg cagcagagtc gacatctctg aactcacagg ttatatgtca ctgttagatt tattttatta gaatcctgtg tagataggca cagtttatat attagatttg tattggattt <400> 5 Met Leu Lys Arg Lys Gin Ser Ser Arg Val Glu Ala Gin Pro Val Thr 15 10 15 Asp Phe Gly Pro Asp Glu Ser Leu Ser Asp Asn Ala Asp He Leu Trp 20 25 30 He Asn Lys Pro Trp Val His Ser Leu Leu Arg He Cys Ala He He 35 40 45 Ser Val He Ser Val Cys Met Asn Thr Pro Met Thr Phe Glu His Tyr 50 55 60 Pro Pro Leu Gin Tyr Val Thr Phe Thr Leu Asp Thr Leu Leu Met Phe 65 70 75 80 Leu Tyr Thr Ala Glu Met He Ala Lys Met His He Arg Gly He Val 85 90 95 - 209 - 008252 - 210 - 008252 - 211 - 008252 Met Leu Ala Leu Phe Glu Val Leu Ser Leu Lys Gly Trp Val Glu Val 1105 1110 1115 1120 Arg Asp Val He He His Arg Val Gly Pro He His Gly He Tyr He 1125 1130 1135 His Val Phe Val Phe Leu Gly Cys Met He Gly Leu Thr Leu Phe Val 1140 1145 1150 Gly Val Val He Ala Asn Phe Asn Glu Asn Lys Gly Thr Ala Leu Leu 1155 1160 1165 Thr Val Asp Gin Arg Arg Trp Glu Asp Leu Lys Ser Arg Leu Lys He 1170 1175 1180 Ala Gin Pro Leu His Leu Pro Pro Arg Pro Asp Asn Asp Gly Phe Arg 1185 1190 1195 1200 Ala Lys Met Tyr Asp He Thr Gin His Pro Phe Phe Lys Arg Thr He 1205 1210 1215 Ala Leu Leu Val Leu Ala Gin Ser Val Leu Leu Ser Val Lys Trp Asp 1220 1225 1230 Val Glu Asp Pro Val Thr Val Pro Leu Ala Thr Met Ser Val Val Phe 1235 1240 1245 Thr Phe He Phe Val Leu Glu Val Thr Met Lys He He Ala Met Ser 1250 1255 1260 Pro Ala Gly Phe Trp Gin Ser Arg Arg Asn Arg Tyr Asp Leu Leu Val 1265 1270 1275 1280 Thr Ser Leu Gly Val Val Trp Val Val Leu His Phe Ala Leu Leu Asn 1285 1290 1295 Ala Tyr Thr Tyr Met Met Gly Ala Cys Val He Val Phe Arg Phe Phe 1300 1305 1310 Ser He Cys Gly Lys His Val Thr Leu Lys Met Leu Leu Leu Thr Val 1315 1320 1325 Val Val Ser Met Tyr Lys Ser Phe Phe He He Val Gly Met Phe Leu 1330 1335 1340 Leu Leu Leu Cys Tyr Ala Phe Ala Gly Val Val Leu Phe Gly Thr Val 1345 1350 1355 1360 Lys Tyr Gly Glu Asn He Asn Arg His Ala Asn Phe Ser Ser Ala Gly 1365 1370 1375 Lys Ala He Thr Val Leu Phe Arg He Val Thr Gly Glu Asp Trp Asn 1380 1385 1390 Lys He Met His Asp Cys Met Val Gin Pro Pro Phe Cys Thr Pro Asp 1395 1400 . 1405 Glu Phe Thr Tyr Trp Ala Thr Asp Cys Gly Asn Tyr Ala Gly Ala Leu 1410 1415 1420 Met Tyr Phe Cys Ser Phe Tyr Val He He Ala Tyr He Met Leu Asn 1425 1430 1435 1440 Leu Leu Val Ala He He Val Glu Asn Phe Ser Leu Phe Tyr Ser Thr 1445 1450 1455 Glu Glu Asp Gin Leu Leu Ser Tyr Asn Asp Leu Arg His Phe Gin He 1460 1465 1470 He Trp Asn Met Val Asp Asp Lys Arg Glu Gly Val He Pro Thr Phe 1475 1480 1485 Arg Val Lys Phe Leu Leu Arg Leu Leu Arg Gly Arg Leu Glu Val Asp 1490 1495 1500 T л.- 7V л*-. T • ¦ л 7\ T ¦> ¦ n T . ч T л i •" ПкЛ 7 *.~ U-i e. U"f ,r- T1 ¦ r-r- ПЛ ii Мгъ.+- Г1 n JuiwU ПО^ ±jy ГЛ О Jj-1 JjyO -UCU UCU C1IC Jul у _> _-__.-> UCL V^-> J. у J. \_> J- -- IJC v, v_> Jv _Л 1505 1510 1515 1520 Arg Leu His Asn Gly Gly Asp Val Thr Phe His Asp Val Leu Ser Met 1525 1530 1535 Leu Ser Tyr Arg Ser Val Asp He Arg Lys Ser Leu Gin Leu Glu Glu 1540 1545 1550 Leu Leu Ala Arg Glu Gin Leu Glu Tyr Thr He Glu Glu Glu Val Ala 1555 1560 1565 Lys Gin Thr He Arg Met Trp Leu Lys Lys Cys Leu Lys Arg He Arg 1570 1575 1580 Ala Lys Gin Gin Gin Ser Cys Ser He He His Ser Leu Arg Glu Ser 1585 1590 1595 1600 Gin Gin Gin Glu Leu Ser Arg Phe Leu Asn Pro Pro Ser He Glu Thr - 212 - 008252 1605 1610 1615 Thr Gin Pro Ser Glu Asp Thr Asn Ala Asn Ser Gin Asp Asn Ser Met 1620 1625 1630 Gin Pro Glu Thr Ser Ser Gin Gin Gin Leu Leu Ser Pro Thr Leu Ser 1635 1640 1645 Asp Arg Gly Gly Ser Arg Gin Asp Ala Ala Asp Ala Gly Lys Pro Gin 1650 1655 1660 Arg Lys Phe Gly Gin Trp Arg Leu Pro Ser Ala Pro Lys Pro He Ser 1665 1670 1675 1680 His Ser Val Ser Ser Val Asn Leu Arg Phe Gly Gly Arg Thr Thr Met 1685 1690 1695 Lys Ser Val Val Cys Lys Met Asn Pro Met Thr Asp Ala Ala Ser Cys 1700 1705 1710 Gly Ser Glu Val Lys Lys Trp Trp Thr Arg Gin Leu Thr Val Glu Ser 1715 1720 1725 Asp Glu Ser Gly Asp Asp Leu Leu Asp He 1730 1735 <210> 6 <211> 453 <212> ДНК <213> Homo sapiens <220> <221> аллель <222> 105 <223> 99-62617-105 : полиморфное основание G или С <220> <221> ошибочное связывание <222> 93..117 <223> 99-62617-105.зонд <220> <221> связанный праймер <222> 86..104 <223> 99-62617-105.mis <220> <221> связанный праймер <222> 106..124 <223> 99-62617-105.mis комплемент <220> <221> связанный праймер <222> 1..20 <223> 99-62617.pu <220> <221> связанный праймер <222> 435..453 <223> 99-62617.гр комплемент <400> 6 tgaggatgag gggcaaacac agaggaagaa tctgggcccc atgaggtcaa cagtgagctg 60 ctaaaattaa cctgaaattt ccctgctggg agacattgtg ttgtsaaata gttttatacc 120 atcttttact aggttttcaa ttaccagcag ttggatgcac cctgtgatcc aatggataca 180 aatgagattt agaagtgcac tttaacttaa ataaaataaa tagccttgtc taatgtatta 240 acgagaaaga catttttgtc ttatttatat gtcttatgtt ctttgaaatg aaataaatga 300 ccatatatgc ttgtaaagag agggagtcta cagatatttg gagaaaggac attctgggtg 360 gagagaacac agagtgcaaa ggacttgaag ttaaagtgca tccagcagtc tcaaggaaga 420 gtaaaggggt ccatgtgagt ggaataaggt gag 4 53 <210> 7 <211> 18 <212> ДНК <213> синтетическая последовательность <220> <223> секвенирующий олигонуклеотидный праймер PU <400> 7 tgtaaaacga cggccagt 18 <210> 8 <211> 18 <212> ДНК <213> синтетическая последовательность <220> <223> секвенирутощий олигонуклеотидный праймер RP <400> 8 caggaaacag ctatgacc 18 - 213 - 008252 ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Антитело или антигенсвязывающий домен, специфически связывающийся с полипептидом с последовательностью SEQ ID NO: 5. 2. Антитело или антигенсвязывающий домен по п.1, который является химерным, гуманизированным антителом, или антигенсвязывающим доменом, или моноклональным или поликлональным антителом, или антигенсвязывающим доменом человека. 3. Антитело или антигенсвязывающий домен по п.1 или 2, которое является антителом или анти-генсвязывающим доменом с одной цепью. 4. Антитело или антигенсвязывающий домен по любому из пп.1-3, который является моноспецифичным, биспецифичным, триспецифичным или мультиспецифичным антителом или антигенсвязываю-щим доменом. 5. Применение антитела или антигенсвязывающего домена по любому из пп.1-3 для диагностики шизофрении или биполярного расстройства. 6. Набор антисмысловых последовательностей, ингибирующих экспрессию полипептида ионного канала, кодируемого полинуклеотидом, показанным в виде SEQ ID NO: 1, 2, 3 или 4, содержащий фрагмент полинуклеотида с последовательностью SEQ ID NO: 1, 2, 3 или 4, для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 7. Набор антисмысловых последовательностей по п.6, которые являются рибозимами. 8. Применение антитела или антигенсвязывающего фрагмента по пп.1-4 для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 9. Применение набора антисмысловых последовательностей по п.6 или 7 для получения лекарственного средства для лечения шизофрении или биполярного расстройства. 10. Фармацевтическая композиция, содержащая антитело или антигенсвязывающий фрагмент по пп.1-4. 11. Фармацевтическая композиция, содержащая набор антисмысловых последовательностей по п.6 или 7. 12. Применение по крайней мере одного двухаллельного маркера (ДМ), связанного с ионным каналом, выбранного из группы двуаллельных маркеров, показанных ниже, для определения подвержен ли индивидуум риску или страдает ли больной шизофренией или биполярным расстройством. Название маркер Полиморфизм Положение ДМ в SEQ ID Аллель 1 Аллель 2 N0:1 NO : 2 NO : 3 N0:4 NO:06 99-62626-168 12642 99-62632-275 14088 99-62633-409 24981 99-62611-51 69248 99-62605-56 73428 99-62635-443 80250 99-79335-60 51090 99-79336-369 61293 99-79338-332 80602 А10 99-79314-201 100485 99-79314-225 100509 А12 99-79316-158 106725 1658 А13 99-79322-224 166087 А14 99-79322-473 166336 А15 99-79306-182 235894 А16 99-79310-29 31646 4481 А17 99-79311-50 42373 А18 99-62617-105 105 - 214 - 008252 13. Способ генотипирования, включающий стадии: a) выделения нуклеиновой кислоты из биологического образца; b) обнаружения наличия нуклеотида в одном или нескольких зависимых двухаллельных маркерах, связанных с ионным каналом, А1-А18, перечисленных в п.12. 14. Способ по п.13, где способ осуществляют с использованием анализа микросеквенирования. 15. Диагностический набор для диагностики шизофрении или биполярного расстройства, содержащий полинуклеотид, который является фрагментом полинуклеотида с последовательностью SEQ ID NO: 1, 2, 3 или 4, содержащего один или несколько биаллельных маркеров А1-А18, как описано в п.12. 16. Диагностический набор для диагностики шизофрении или биполярного расстройства, содержащий: a) поликлональное или моноклональное антитело, которое специфически связывается с полипептидом ионного канала, содержащим аминокислотную последовательность SEQ ID No 5, или с фрагментом пептида или его вариантом, необязательно меченным; b) реагент, с помощью которого осуществляется определение образовавшегося комплекса антиген-антитела, указанный реагент либо необязательно содержит метку, либо который может быть распознан меченным реагентом. область LE 8000000 85О0ОО0 9000000 9500000 10000000 10500000 110О0ООО 115ОООО0 120О0ООО 125СКХ С0001А06 О0001А10 Фиг. 1 данные поискового устройства 120 Фиг. 2 - 215 - 008252 200 Q запуск ^)-201 сохранить новую послед овательность в память открыть базу данных последовательностей прочтение первой последовательности в базе осуществление сравнения новой послед, и сохраненной последовательности 202 204 206 210 212 224 перейти к след последовател-Ti/ в базе данных одинаковые 2.14 дисплей сохраняет название последовательности J для пользователя ' 218 •да нет 220 (^окончание Фиг. 3 250 С за"Уск 252 сохранение первой последовательности в память 254 256 сохранение второй последовательности в память прочтение первого символа в первой последовател-ти 260 прочтение первого символа во второй последов-ти 262 264 одинаковые? 268 прочтение следующего символа в первой и второй последовательностях_ 270 одинаковые? 274 1несколькТ символов дляда-ючтен! нет 276 уровень дисплейной гомологии между первой и второй последовательностями у окончание у/ Фиг. 4 278 - 216 - 008252 300 ( запуск J-302 сохранение первой последовательности в память открытие базы данных элементов последовательное,-! 304 308 прочтение первого элемента в базе данных 310 сравнение определяющих элементов с первой J последовательностью У 316 нахождение? 318 дисплейное обнаружение элемента для пользователя [ 320 прочтение следующего элемента в fiaag ПЯЫЫЫУ 324 Гокочание Фиг. 5 Евразийская патентная организация, ЕАПВ Россия, 109012, Москва, Малый Черкасский пер., 2/6 - 217 - |