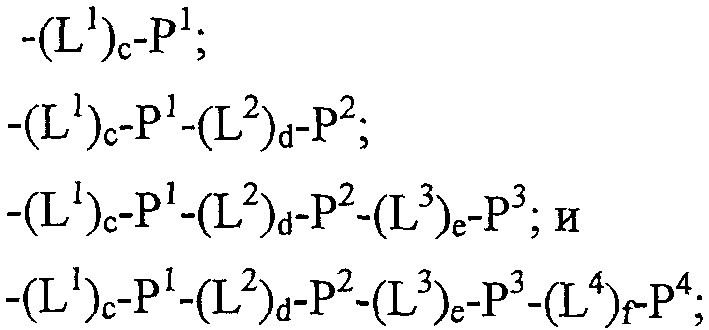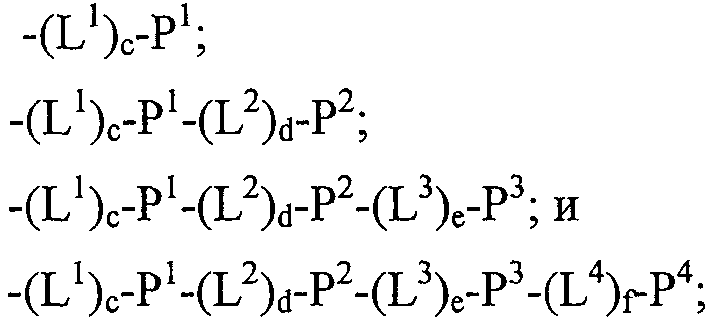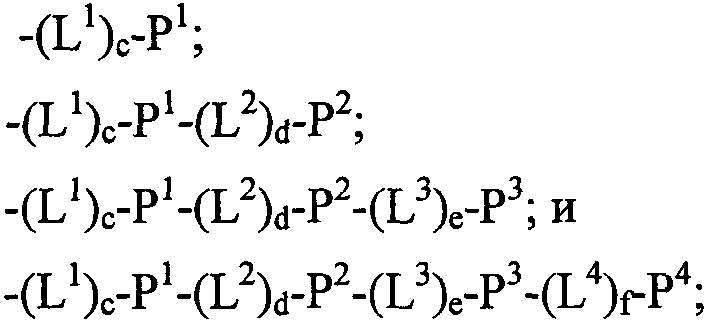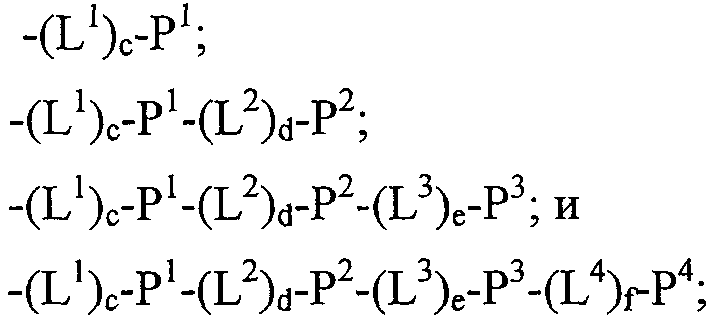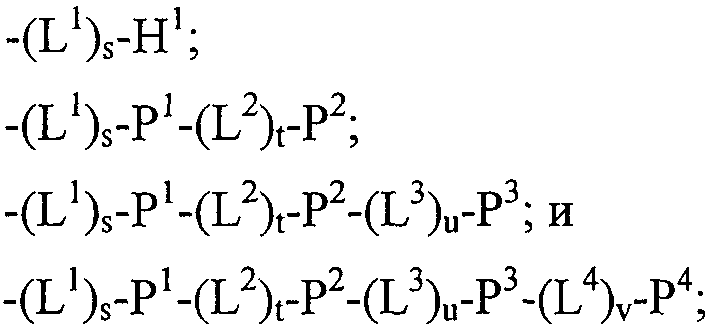|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000008248b*\id |
|
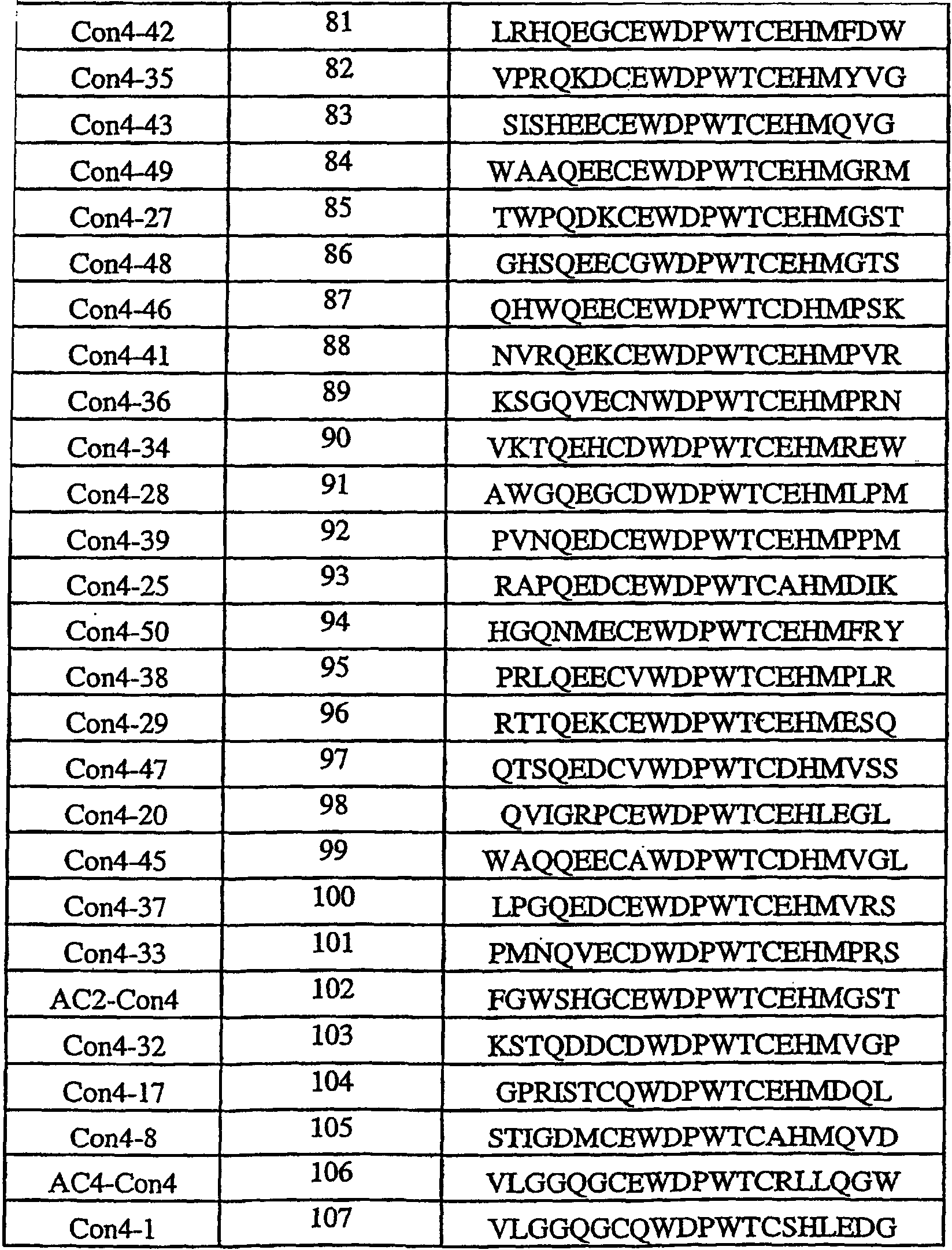
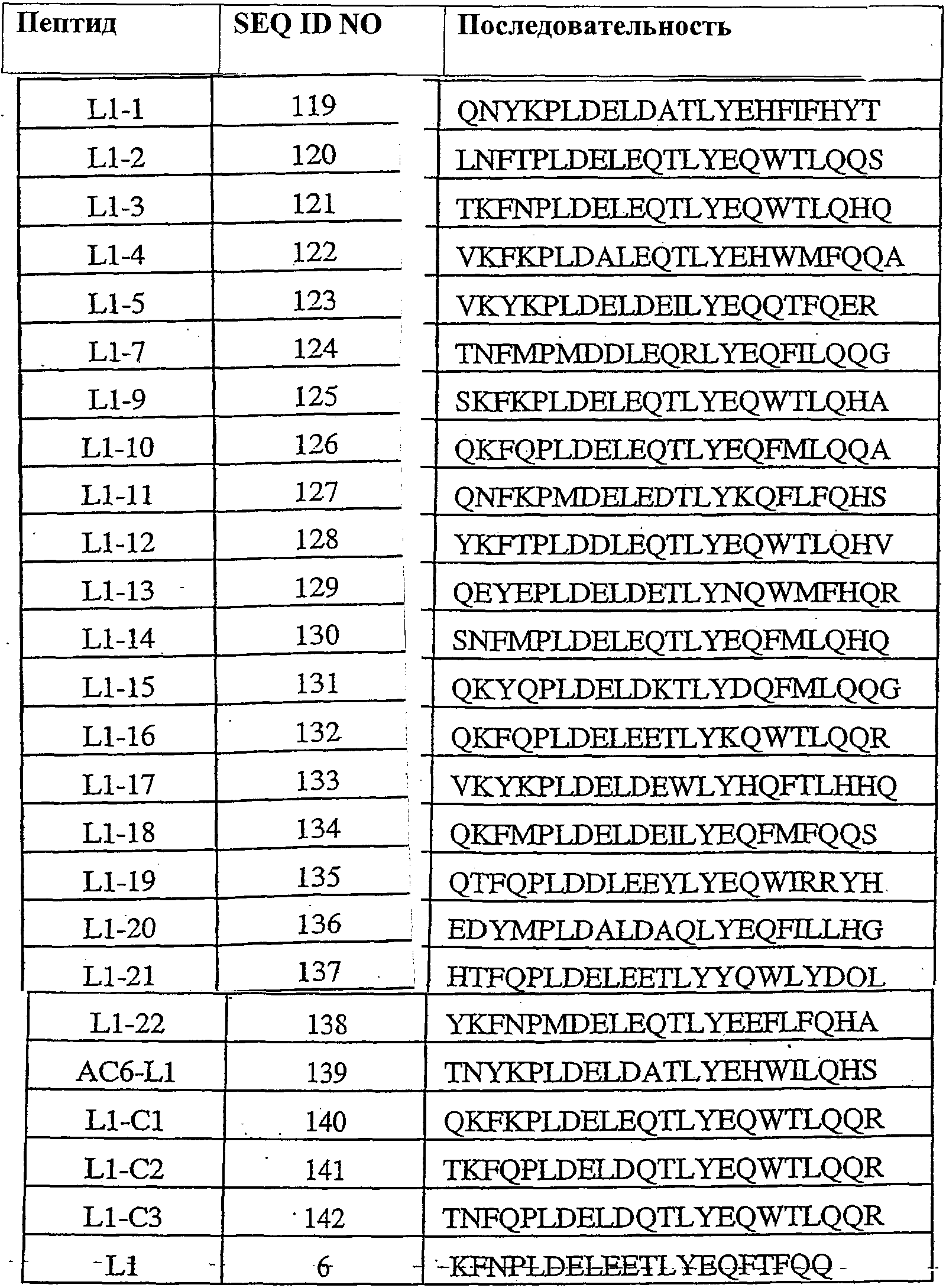
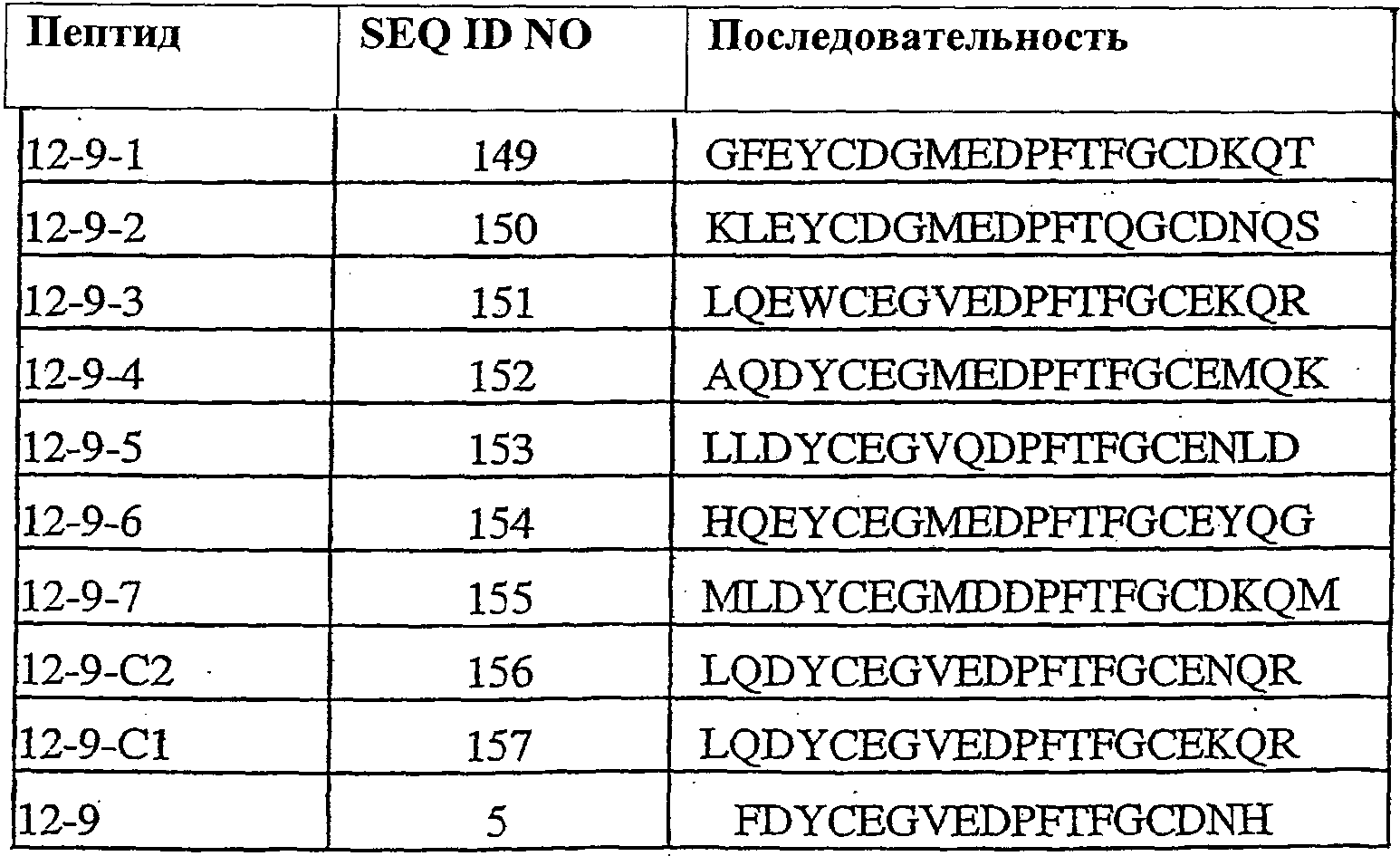
Термины запроса в документе Реферат 1. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность WDPWT (SEQ ID NO: 65) длиной в 5-50 аминокислот, и его физиологически приемлемые соли. 2. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность WDPWTC (SEQ ID NO: 66) и его физиологически приемлемые соли. 3. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность Cz 2 WDPWT (SEQ ID NO: 67) где z 2 представляет собой кислый или нейтральный полярный аминокислотный остаток, и его физиологически приемлемые соли. 4. Полипептид по п.3, содержащий аминокислотную последовательность Cz 2 WDPWTC (SEQ ID NO: 68) где z 2 обозначает кислый или нейтральный полярный аминокислотный остаток, и его физиологически приемлемые соли. 5. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы a 1 a 2 a 3 Ca 5 WDPWTCa 12 a 13 a 14 (SEQ ID NO: 69) где а 1 , а 2 и а 3 , каждый независимо, обозначают аминокислотные остатки; а 5 обозначает аминокислотный остаток; а 12 отсутствует или обозначает аминокислотный остаток; а 13 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; а 14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; и его физиологически приемлемые соли. 6. Полипептид по п.5, отличающийся тем, что а 1 обозначает V, I, P, W, G, S, Q, N, Е, K, R или Н; а 2 обозначает V, Р, М, G, S, Q, D, Е, K, R или Н; а 3 обозначает А, V, Р, М, F, T, G, D, Е, K или Н; а 8 обозначает А, V, G, Q, N, D или Е; а 12 обозначает S, Q, N, D, Е, K или R; а 13 обозначает L, Т или Н; а 14 обозначает V, L, I, W или М; и его физиологически приемлемые соли. 7. Полипептид по п.5, отличающийся тем, что а 1 обозначает Q; а 2 обозначает Е; а 3 обозначает Е; а 5 обозначает D или Е; а 12 обозначает D или Е; а 13 обозначает Н и а 14 обозначает М, и его физиологически приемлемые соли. 8. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы b 1 b 2 b 3 b 4 b 5 b 6 Cb 8 WDPWTCb 15 b 16 b 17 b 18 b 19 b 20 (SEQ ID NO: 70) где b 1 отсутствует или обозначает аминокислотный остаток; b 2 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; b 3 , b 4 , b 5 и b 6 , каждый независимо, отсутствуют или обозначают аминокислотные остатки; b 8 обозначает аминокислотный остаток; b 15 отсутствует или обозначает аминокислотный остаток; b 16 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; b 17 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; b 18 , b 19 и b 20 , каждый независимо, отсутствуют или обозначают аминокислотные остатки; и его физиологически приемлемые соли. 9. Полипептид по п.8, отличающийся тем, что b 1 отсутствует или обозначает А, V, L, P, W, F, T, G, S, Q, N, K, R или Н; b 2 отсутствует или обозначает А, V, L, I, P, W, M, T, G, S, Y, N, K, R или Н; b 3 отсутствует или обозначает A, L, I, P, W, M, T, G, S, Q, N, E, R или Н; b 4 обозначает V, I, P, W, G, S, Q, N, Е, K, R или Н; b 5 обозначает V, Р, М, G, S, Q, D, Е, K, R или Н; b 6 обозначает А, V, Р, М, F, T, G, D, Е, K или Н; b 8 обозначает А, V, G, Q, N, D или Е; b 15 обозначает S, Q, N, D, Е, K или R; b 16 обозначает L, Т или Н; b 17 обозначает V, L, I, W или М; b 18 отсутствует или обозначает А, V, L, P, W, F, T, G, Y, Q, D, Е или R; b 19 отсутствует или обозначает V, L, I, P, T, G, S, Y, Q, N, D, Е или R и b 20 отсутствует или обозначает V, L, P, W, M, T, G, S, Y, Q, N, D, K или R; и его физиологически приемлемые соли. 10. Полипептид по п.8, отличающийся тем, что b 1 отсутствует или обозначает Р или Т; b 2 отсутствует или обозначает I или N; b 3 отсутствует или обозначает R или I; b 4 обозначает Q; b 5 обозначает Е; b 6 обозначает Е; b 8 обозначает D или Е; b 15 обозначает D или Е; b 16 обозначает Н; b 17 обозначает М; b 18 отсутствует или обозначает W или Р; b 19 отсутствует или обозначает G или Е и b 20 отсутствует или обозначает V или K; и его физиологически приемлемые соли. 11. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбираемую из группы, приведенной в табл. 1, состоящей из SEQ ID NO: 4 и с SEQ ID NO: 76 по SEQ ID NO: 118 включительно, и его физиологически приемлемые соли. Таблица 1 12. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пептид по пп.1, 5, 8 или 11 и носитель, и его физиологически приемлемые соли. 13. Слитый полипептид по п.12, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 14. Полипептид по пп.1, 5, 8 или 11, который является циклическим. 15. Димер или мультимер полипептидов по пп.1, 5, 8 или 11. 16. Состав полипептида, имеющий формулу (X 1 ) a -F 1 -(X 2 ) b , и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид, выбираемый из группы, состоящей из: (a) аминокислотной последовательности WDPWT (SEQ ID NO: 65), отличающейся тем, что указанный полипептид имеет протяжённость 5-50 аминокислот; (b) аминокислотной последовательности WDPWTC (SEQ ID NO: 66), (c) аминокислотной последовательности Cz 2 WDPWT (SEQ ID NO: 67) и (d) аминокислотной последовательности Cz 2 WDPWTC (SEQ ID NO: 68), где z 2 обозначает кислый или нейтральный полярный аминокислотный остаток; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначает линкеры и а, b, с, d, e и f, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 17. Состав полипептида по п.16, отличающийся тем, что z 2 обозначает А, V, G, Q, N, D или Е. 18. Состав полипептида по п.16, отличающийся тем, что один или более из Р 1 , Р 2 , Р 3 или Р 4 , каждый независимо, представляет собой полипептид, выбираемый из группы, состоящей из SEQ ID NO: 4 и с SEQ ID NO: 76 по SEQ ID NO: 118 включительно. 19. Состав полипептида по п.16, соответствующий формулам X 1 -F 1 или F 1 -X 2 и его физиологически приемлемые соли. 20. Состав полипептида по п.16 формулы F 1 -(L 1 ) c -P 1 и его физиологически приемлемые соли. 21. Состав полипептида по п.16, соответствующий формуле F 1 -(L 1 ) c -P 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 22. Состав полипептида по п.16, соответствующий формуле P 1 -(L 1 ) c -F 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 23. Состав полипептида по п.16, отличающийся тем, что F 1 обозначает домен Fc или его фрагмент. 24. Состав полипептида по п.16, отличающийся тем, что F 1 содержит аминокислотную последовательность SEQ ID NO: 60. 25. Полинуклеотид, кодирующий состав полипептида по пп.1, 5, 8 или 11. 26. Вектор экспрессии, содержащий полинуклеотид по п.25. 27. Клетка-хозяин, содержащая вектор экспрессии по п.26. 28. Клетка-хозяин по п.27, отличающаяся тем, что является клеткой прокариот. 29. Клетка-хозяин по п.28, отличающаяся тем что является клеткой Е. coli. 30. Клетка-хозяин по п.27, отличающаяся тем, что является клеткой эукариот. 31. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы Pc 2 Dc 4 Lc 6 c 7 c 8 LY (SEQ ID NO: 71) где с 2 обозначает нейтральный гидрофобный аминокислотный остаток; с 4 обозначает A, D или Е; с 6 обозначает кислый аминокислотный остаток; с 7 обозначает аминокислотный остаток и с 8 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; и его физиологически приемлемые соли. 32. Полипептид по п.31, отличающийся тем, что с 2 обозначает L или М. 33. Полипептид по п.31, отличающийся тем, что с 6 обозначает D или Е. 34. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы d 1 d 2 d 3 d 4 Pd 6 Dd 8 Ld 10 d 11 d 12 LYd 15 d 16 d 17 d 18 d 19 d 20 d 21 d 22 (SEQ ID NO: 72) где d 1 отсутствует или обозначает аминокислотный остаток; d 2 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d 3 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d 4 отсутствует или обозначает аминокислотный остаток; d 6 обозначает нейтральный гидрофобный аминокислотный остаток; d 8 обозначает A, D или Е; d 10 обозначает кислый аминокислотный остаток; d 11 обозначает аминокислотный остаток; d 12 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; d 15 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d 16 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d 17 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d 18 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d 19 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; d 20 отсутствует или обозначает аминокислотный остаток; d 21 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d 22 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; и его физиологически приемлемые соли. 35. Полипептид по п.34, отличающийся тем, что: d 1 обозначает Т, S, Q, R или Н; d 2 обозначает Т, Q, N или K; d 3 обозначает F; d 4 обозначает М, Q, Е или K; d 6 обозначает L или М; d 8 обозначает D или Е; d 10 обозначает Е; d 11 обозначает Q или Е; d 12 обозначает Т или R; d 15 обозначает Y, D, Е или K; d 16 обозначает Q; d 17 обозначает W или F; d 18 обозначает L, I, М или Т; d 19 обозначает L, F или Y; d 20 обозначает Q, D или Е; d 21 отсутствует или обозначает Q или Н; d 22 отсутствует или обозначает A, L, G, S или R; и его физиологически приемлемые соли. 36. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 2, состоящей из SEQ ID NO: 6 и с SEQ ID NO: 119 по SEQ ID NO: 142 включительно, и его физиологически приемлемые соли. Таблица 2 37. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пептид по пп.31, 34 или 36 и носитель, и его физиологически приемлемые соли. 38. Слитый полипептид по п.37, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 39. Полипептид по пп.31, 34 или 36, который является циклическим. 40. Димер или мультимер полипептидов по пп.31, 34 или 36. 41. Состав полипептида, имеющий формулу (Х 1 ) a -F 1 -(X 2 ) b и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид по пп.31, 34 или 36; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначают линкеры и а, b, с, d, e и f, каждый независимо, обозначают 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 42. Состав полипептида по п.41, отличающийся тем, что один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 6 и с SEQ ID NO: 119 по SEQ ID NO: 142 включительно. 43. Состав полипептида по п.41, соответствующий формулам X 1 -F 1 или F 1 -X 2 и его физиологически приемлемые соли. 44. Состав полипептида по п.41 формулы F 1 -(L 1 ) c -P 1 и его физиологически приемлемые соли. 45. Состав полипептида по п.41, соответствующий формуле F 1 -(L 1 ) c -P 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 46. Состав полипептида по п.41, соответствующий формуле P 1 -(L 1 ) c -F 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 47. Состав полипептида по п.41, отличающийся тем, что F 1 обозначает домен Fc или его фрагмент. 48. Состав полипептида по п.41, отличающийся тем, что F 1 содержит аминокислотную последовательность SEQ ID NO: 60. 49. Полинуклеотид, кодирующий полипептид по любому из пп.31, 34 или 36. 50. Вектор экспрессии, содержащий полинуклеотид по п.49. 51. Клетка-хозяин, содержащая вектор экспрессии по п.50. 52. Клетка-хозяин по п.51, отличающаяся тем, что является клеткой прокариот. 53. Клетка-хозяин по п.52, отличающаяся тем что является клеткой Е. coli. 54. Клетка-хозяин по п.51, отличающаяся тем, что является клеткой эукариот. 55. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы RPe 3 e 4 e 5 e 6 e 7 G (SEQ ID NO: 73) где е 3 обозначает нейтральный полярный аминокислотный остаток; е 4 обозначает кислый аминокислотный остаток; е 5 обозначает нейтральный полярный или кислый аминокислотный остаток; е 6 обозначает нейтральный гидрофобный аминокислотный остаток; е 7 обозначает нейтральный гидрофобный аминокислотный остаток; и его физиологически приемлемые соли. 56. Полипептид по п.55, отличающийся тем, что е 3 обозначает Y или С. 57. Полипептид по п.55, отличающийся тем, что е 4 обозначает D или Е. 58. Полипептид по п.55, отличающийся тем, что е 6 обозначает I или М. 59. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы f 1 f 2 f 3 f 4 RPf 7 f 8 f 9 f 10 f 11 Gf 13 f 14 f 15 f 16 f 17 f 18 f 19 f 20 (SEQ ID NO: 74) где f 1 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 2 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 3 обозначает нейтральный полярный или кислый аминокислотный остаток; f 4 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 7 обозначает нейтральный полярный аминокислотный остаток; f 8 обозначает кислый аминокислотный остаток; f 9 обозначает нейтральный полярный или кислый аминокислотный остаток; f 10 обозначает нейтральный гидрофобный аминокислотный остаток; f 11 обозначает нейтральный гидрофобный аминокислотный остаток; f 13 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 15 обозначает нейтральный полярный аминокислотный остаток; f 16 обозначает нейтральный полярный аминокислотный остаток; f 17 обозначает нейтральный полярный или кислый аминокислотный остаток; f 18 обозначает нейтральный гидрофобный или основной аминокислотный остаток; f 19 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и f 20 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; и его физиологически приемлемые соли. 60. Полипептид по п.59, отличающийся тем, что f 1 обозначает S, А или G; f 2 обозначает G, Q или Р; f 3 обозначает Q, G или D; f 4 обозначает L, М или Q; f 7 обозначает С или Y; f 8 обозначает Е или D; f 9 обозначает Е, G или D; f 10 обозначает I или М; f 11 обозначает F или L; f 13 обозначает С или W; f 14 обозначает G или Р; f 15 обозначает Т или N; f 16 обозначает Q, Y или K; f 17 обозначает N, D или Q; f 18 обозначает L, V, W или R; f 19 обозначает A, Q, Y или I и f 20 обозначает L, A, G или V; и его физиологически приемлемые соли. 61. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 3, состоящей из SEQ ID NO: 3 и с SEQ ID NO: 143 по SEQ ID NO: 148 включительно, и его физиологически приемлемые соли Таблица 3 62. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один полипептид по пп.55, 59 или 61 и носитель и его физиологически приемлемые соли. 63. Слитый полипептид по п.62, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 64. Полипептид по пп.55, 59 или 61, который является циклическим. 65. Димер или мультимер полипептидов по пп.55, 59 или 61. 66. Состав полипептида, имеющий формулу (X 1 ) a -F 1 -(X 2 ) b и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид по пп.55, 59 или 61; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначают линкеры и а, b, с, d, е и f, каждый независимо, обозначают 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 67. Состав полипептида по п.66, отличающийся тем, что один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 3 и с SEQ ID NO: 143 по SEQ ID NO: 148 включительно. 68. Состав полипептида по п.66, соответствующий формулам X 1 -F 1 или F 1 -X 2 и его физиологически приемлемые соли. 69. Состав полипептида по п.66 формулы F 1 -(L 1 ) c -P 1 и его физиологически приемлемые соли. 70. Состав полипептида по п.66, соответствующий формуле F 1 -(L 1 ) c -P 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 71. Состав полипептида по п.66, соответствующий формуле P 1 -(L 1 ) c -F 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 72. Состав полипептида по п.66, отличающийся тем, что F 1 обозначает домен Fc или его фрагмент. 73. Состав полипептида по п.66, отличающийся тем, что F 1 содержит аминокислотную последовательность SEQ ID NO: 60. 74. Полинуклеотид, кодирующий полипептид по любому из пп.55, 59 или 61. 75. Вектор экспрессии, содержащий полинуклеотид по п.74. 76. Клетка-хозяин, содержащая вектор экспрессии по п.75. 77. Клетка-хозяин по п.76, отличающаяся тем, что является клеткой прокариот. 78. Клетка-хозяин по п.77, отличающаяся тем что является клеткой Е. coli. 79. Клетка-хозяин по п.76, отличающаяся тем, что является клеткой эукариот. 80. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы Cg 2 Gg 4 g 5 DPFTg 10 GCg 13 (SEQ ID NO: 75) где g 2 обозначает кислый аминокислотный остаток; g 4 обозначает нейтральный гидрофобный аминокислотный остаток; g 5 обозначает Е, D или Q; g 10 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; g 13 обозначает кислый остаток; и его физиологически приемлемые соли. 81. Полипептид по п.80, отличающийся тем, что g 2 обозначает Е или D. 82. Полипептид по п.80, отличающийся тем, что g 4 обозначает V или М. 83. Полипептид по п.80, отличающийся тем, что g 10 обозначает F или Q. 84. Полипептид по п.80, отличающийся тем, что g 13 обозначает D или Е. 85. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы h 1 h 2 h 3 h 4 Ch 6 Gh 8 h 9 DPFTh 14 GCh 17 h 18 h 19 h 20 (SEQ ID NO:158) где h 1 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; h 2 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h 3 обозначает кислый аминокислотный остаток; h 4 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h 6 обозначает кислый аминокислотный остаток; h 8 обозначает нейтральный гидрофобный аминокислотный остаток; h 9 обозначает Е, D или Q; h 14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h 17 обозначает кислый аминокислотный остаток; h 18 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; h 19 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и h 20 отсутствует или обозначает аминокислотный остаток; и его физиологически приемлемые соли. 86. Полипептид по п.85, отличающийся тем, что h 1 отсутствует или обозначает A, L, M, G, K или Н; h 2 обозначает L, F или Q; h 3 обозначает D или Е; h 4 обозначает W или Y; h 6 обозначает D или Е; h 8 обозначает V или М; h 14 обозначает F или Q; h 17 обозначает D или Е; h 18 обозначает М, Y, N или K; h 19 обозначает L или Q и h 20 отсутствует или обозначает М, Т, G, S, D, K или R; и его физиологически приемлемые соли. 87. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 4, состоящей из SEQ ID NO: 5 или с SEQ ID NO: 149 по SEQ ID NO: 157 включительно, и его физиологически приемлемые соли.
Таблица 4 88. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пептид по пп.80, 85 или 87 и носитель, его физиологически приемлемые соли. 89. Слитый полипептид по п.88, отличающийся тем, что носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 90. Полипептид по пп.80, 85 или 87, который является циклическим. 91. Димер или мультимер полипептидов по пп.80, 85 или 87. 92. Состав полипептида, имеющий формулу (X 1 ) a -F 1 -(X 2 ) b и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид по пп.80, 85 или 87; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначает линкеры и а, b, с, d, e и f, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 93. Состав полипептида по п.92, отличающийся тем, что один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 5 и с SEQ ID NO: 149 по SEQ ID NO: 157 включительно. 94. Состав полипептида по п.92, соответствующий формулам X 1 -F 1 или F 1 -X 2 и его физиологически приемлемые соли. 95. Состав полипептида по п.92 формулы F 1 -(L 1 ) c -P 1 и его физиологически приемлемые соли. 96. Состав полипептида по п.92, соответствующий формуле F 1 -(L 1 ) c -P 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 97. Состав полипептида по п.92, соответствующий формуле P 1 -(L 1 ) c -F 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 98. Состав полипептида по п.92, отличающийся тем, что F 1 обозначает домен Fc или его фрагмент. 99. Состав полипептида по п.92, отличающийся тем, что F 1 содержит аминокислотную последовательность SEQ ID NO: 60. 100. Полинуклеотид, кодирующий полипептид по любому из пп.80, 85 или 87. 101. Вектор экспрессии, содержащий полинуклеотид по п.100. 102. Клетка-хозяин, содержащая вектор экспрессии по п.101. 103. Клетка-хозяин по п.102, отличающаяся тем, что является клеткой прокариот. 104. Клетка-хозяин по п.103, отличающаяся тем что является клеткой Е. coli. 105. Клетка-хозяин по п.102, отличающаяся тем, что является клеткой эукариот. 106. Полипептид в соответствии с любой из последовательностей SEQ ID NO: l, SEQ ID NO: 2, SEQ ID NO: 3 или SEQ ID NO: 4 или его вариант. 107. Полипептид в соответствии с любой из последовательностей SEQ ID NO: 5, SEQ ID NO: 6 или SEQ ID NO: 7 или его вариант. 108. Состав полипептида формулы (X 1 ) q -F 1 -(X 2 ) r и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид, выбранный из группы, состоящей из: (a) аминокислотной последовательности WDPWT (SEQ ID NO: 65), причём указанный полипептид имеет в длину 5-50 аминокислот; (b) аминокислотной последовательности WDPWTC (SEQ ID NO: 66); (c) аминокислотной последовательности Cz 2 WDPWT (SEQ ID NO: 67), где z 2 обозначает кислый или нейтральный полярный аминокислотный остаток; (d) аминокислотной последовательности Cz 2 WDPWTC (SEQ ID NO: 68), где z 2 обозначает кислый или нейтральный полярный аминокислотный остаток; (e) аминокислотной последовательности Рс 2 Dc 4 Lc 6 с 7 с 8 LY (SEQ ID NO: 71), где с 2 обозначает нейтральный гидрофобный аминокислотный остаток; с 4 обозначает A, D или Е; с 6 обозначает кислый аминокислотный остаток; с 7 обозначает аминокислотный остаток и с 8 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; (f) аминокислотной последовательности RPe 3 e 4 e 5 e 6 e 7 G (SEQ ID NO: 73), где е 3 обозначает нейтральный полярный аминокислотный остаток; е 4 обозначает кислый аминокислотный остаток; е 5 обозначает нейтральный полярный или кислый аминокислотный остаток; е 6 обозначает нейтральный гидрофобный аминокислотный остаток и е 7 обозначает нейтральный гидрофобный аминокислотный остаток; (g) аминокислотной последовательности Cg 2 Cg 4 g 5 DPFTg 10 GCg 13 (SEQ ID NO: 75), где g 2 обозначает кислый аминокислотный остаток; g 4 обозначает нейтральный гидрофобный аминокислотный остаток; g 5 обозначает нейтральный полярный или кислый аминокислотный остаток; g 10 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и g 13 обозначает кислый остаток; (h) полипептида SEQ ID NO: 1; (i) полипептида SEQ ID NO: 2 и (j) полипептида SEQ ID NO: 7; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначает линкеры и q, r, s, t, u и v, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из q и r обозначает 1; и его физиологически приемлемые соли. 109. Фармацевтическая композиция, содержащая эффективное количество состава полипептида по пп.1, 31, 55 или 80 в смеси с фармацевтически приемлемым носителем. 110. Способ ингибирования нежелательного ангиогенеза у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 111. Способ лечения ангиогенеза у субъекта, заключающийся во введении эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 112. Способ модулирования ангиогенеза у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 113. Способ подавления роста опухоли, характеризующейся нежелательным ангиогенезом, у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 114. Способ лечения рака у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92 и химиотерапевтического агента. 115. Способ по п.114, отличающийся тем, что химиотерапевтический агент является по меньшей мере одним из 5-FU, СРТ-11 и Таксотер. 116. Способ модулирования по меньшей мере одного нарушения у млекопитающего: сосудистой проницаемости или истечения плазмы, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 117. Способ лечения по меньшей мере одного из заболеваний: реваскулярного заболевания глаза, ожирения, ангиоретикулёмы, гемангиомы, артериосклероза, воспалительного заболевания, воспалительных нарушений, атеросклероза, эндометриоза, опухолевого заболевания, костного заболевания или псориаза, у млекопитающего, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92.
Полный текст патента (57) Реферат / Формула: WDPWT (SEQ ID NO: 65) длиной в 5-50 аминокислот, и его физиологически приемлемые соли. 2. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность WDPWTC (SEQ ID NO: 66) и его физиологически приемлемые соли. 3. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность Cz 2 WDPWT (SEQ ID NO: 67) где z 2 представляет собой кислый или нейтральный полярный аминокислотный остаток, и его физиологически приемлемые соли. 4. Полипептид по п.3, содержащий аминокислотную последовательность Cz 2 WDPWTC (SEQ ID NO: 68) где z 2 обозначает кислый или нейтральный полярный аминокислотный остаток, и его физиологически приемлемые соли. 5. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы a 1 a 2 a 3 Ca 5 WDPWTCa 12 a 13 a 14 (SEQ ID NO: 69) где а 1 , а 2 и а 3 , каждый независимо, обозначают аминокислотные остатки; а 5 обозначает аминокислотный остаток; а 12 отсутствует или обозначает аминокислотный остаток; а 13 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; а 14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; и его физиологически приемлемые соли. 6. Полипептид по п.5, отличающийся тем, что а 1 обозначает V, I, P, W, G, S, Q, N, Е, K, R или Н; а 2 обозначает V, Р, М, G, S, Q, D, Е, K, R или Н; а 3 обозначает А, V, Р, М, F, T, G, D, Е, K или Н; а 8 обозначает А, V, G, Q, N, D или Е; а 12 обозначает S, Q, N, D, Е, K или R; а 13 обозначает L, Т или Н; а 14 обозначает V, L, I, W или М; и его физиологически приемлемые соли. 7. Полипептид по п.5, отличающийся тем, что а 1 обозначает Q; а 2 обозначает Е; а 3 обозначает Е; а 5 обозначает D или Е; а 12 обозначает D или Е; а 13 обозначает Н и а 14 обозначает М, и его физиологически приемлемые соли. 8. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы b 1 b 2 b 3 b 4 b 5 b 6 Cb 8 WDPWTCb 15 b 16 b 17 b 18 b 19 b 20 (SEQ ID NO: 70) где b 1 отсутствует или обозначает аминокислотный остаток; b 2 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; b 3 , b 4 , b 5 и b 6 , каждый независимо, отсутствуют или обозначают аминокислотные остатки; b 8 обозначает аминокислотный остаток; b 15 отсутствует или обозначает аминокислотный остаток; b 16 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; b 17 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; b 18 , b 19 и b 20 , каждый независимо, отсутствуют или обозначают аминокислотные остатки; и его физиологически приемлемые соли. 9. Полипептид по п.8, отличающийся тем, что b 1 отсутствует или обозначает А, V, L, P, W, F, T, G, S, Q, N, K, R или Н; b 2 отсутствует или обозначает А, V, L, I, P, W, M, T, G, S, Y, N, K, R или Н; b 3 отсутствует или обозначает A, L, I, P, W, M, T, G, S, Q, N, E, R или Н; b 4 обозначает V, I, P, W, G, S, Q, N, Е, K, R или Н; b 5 обозначает V, Р, М, G, S, Q, D, Е, K, R или Н; b 6 обозначает А, V, Р, М, F, T, G, D, Е, K или Н; b 8 обозначает А, V, G, Q, N, D или Е; b 15 обозначает S, Q, N, D, Е, K или R; b 16 обозначает L, Т или Н; b 17 обозначает V, L, I, W или М; b 18 отсутствует или обозначает А, V, L, P, W, F, T, G, Y, Q, D, Е или R; b 19 отсутствует или обозначает V, L, I, P, T, G, S, Y, Q, N, D, Е или R и b 20 отсутствует или обозначает V, L, P, W, M, T, G, S, Y, Q, N, D, K или R; и его физиологически приемлемые соли. 10. Полипептид по п.8, отличающийся тем, что b 1 отсутствует или обозначает Р или Т; b 2 отсутствует или обозначает I или N; b 3 отсутствует или обозначает R или I; b 4 обозначает Q; b 5 обозначает Е; b 6 обозначает Е; b 8 обозначает D или Е; b 15 обозначает D или Е; b 16 обозначает Н; b 17 обозначает М; b 18 отсутствует или обозначает W или Р; b 19 отсутствует или обозначает G или Е и b 20 отсутствует или обозначает V или K; и его физиологически приемлемые соли. 11. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбираемую из группы, приведенной в табл. 1, состоящей из SEQ ID NO: 4 и с SEQ ID NO: 76 по SEQ ID NO: 118 включительно, и его физиологически приемлемые соли. Таблица 1 12. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пептид по пп.1, 5, 8 или 11 и носитель, и его физиологически приемлемые соли. 13. Слитый полипептид по п.12, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 14. Полипептид по пп.1, 5, 8 или 11, который является циклическим. 15. Димер или мультимер полипептидов по пп.1, 5, 8 или 11. 16. Состав полипептида, имеющий формулу (X 1 ) a -F 1 -(X 2 ) b , и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид, выбираемый из группы, состоящей из: (a) аминокислотной последовательности WDPWT (SEQ ID NO: 65), отличающейся тем, что указанный полипептид имеет протяжённость 5-50 аминокислот; (b) аминокислотной последовательности WDPWTC (SEQ ID NO: 66), (c) аминокислотной последовательности Cz 2 WDPWT (SEQ ID NO: 67) и (d) аминокислотной последовательности Cz 2 WDPWTC (SEQ ID NO: 68), где z 2 обозначает кислый или нейтральный полярный аминокислотный остаток; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначает линкеры и а, b, с, d, e и f, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 17. Состав полипептида по п.16, отличающийся тем, что z 2 обозначает А, V, G, Q, N, D или Е. 18. Состав полипептида по п.16, отличающийся тем, что один или более из Р 1 , Р 2 , Р 3 или Р 4 , каждый независимо, представляет собой полипептид, выбираемый из группы, состоящей из SEQ ID NO: 4 и с SEQ ID NO: 76 по SEQ ID NO: 118 включительно. 19. Состав полипептида по п.16, соответствующий формулам X 1 -F 1 или F 1 -X 2 и его физиологически приемлемые соли. 20. Состав полипептида по п.16 формулы F 1 -(L 1 ) c -P 1 и его физиологически приемлемые соли. 21. Состав полипептида по п.16, соответствующий формуле F 1 -(L 1 ) c -P 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 22. Состав полипептида по п.16, соответствующий формуле P 1 -(L 1 ) c -F 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 23. Состав полипептида по п.16, отличающийся тем, что F 1 обозначает домен Fc или его фрагмент. 24. Состав полипептида по п.16, отличающийся тем, что F 1 содержит аминокислотную последовательность SEQ ID NO: 60. 25. Полинуклеотид, кодирующий состав полипептида по пп.1, 5, 8 или 11. 26. Вектор экспрессии, содержащий полинуклеотид по п.25. 27. Клетка-хозяин, содержащая вектор экспрессии по п.26. 28. Клетка-хозяин по п.27, отличающаяся тем, что является клеткой прокариот. 29. Клетка-хозяин по п.28, отличающаяся тем что является клеткой Е. coli. 30. Клетка-хозяин по п.27, отличающаяся тем, что является клеткой эукариот. 31. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы Pc 2 Dc 4 Lc 6 c 7 c 8 LY (SEQ ID NO: 71) где с 2 обозначает нейтральный гидрофобный аминокислотный остаток; с 4 обозначает A, D или Е; с 6 обозначает кислый аминокислотный остаток; с 7 обозначает аминокислотный остаток и с 8 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; и его физиологически приемлемые соли. 32. Полипептид по п.31, отличающийся тем, что с 2 обозначает L или М. 33. Полипептид по п.31, отличающийся тем, что с 6 обозначает D или Е. 34. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы d 1 d 2 d 3 d 4 Pd 6 Dd 8 Ld 10 d 11 d 12 LYd 15 d 16 d 17 d 18 d 19 d 20 d 21 d 22 (SEQ ID NO: 72) где d 1 отсутствует или обозначает аминокислотный остаток; d 2 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d 3 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d 4 отсутствует или обозначает аминокислотный остаток; d 6 обозначает нейтральный гидрофобный аминокислотный остаток; d 8 обозначает A, D или Е; d 10 обозначает кислый аминокислотный остаток; d 11 обозначает аминокислотный остаток; d 12 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; d 15 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d 16 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d 17 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d 18 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d 19 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; d 20 отсутствует или обозначает аминокислотный остаток; d 21 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d 22 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; и его физиологически приемлемые соли. 35. Полипептид по п.34, отличающийся тем, что: d 1 обозначает Т, S, Q, R или Н; d 2 обозначает Т, Q, N или K; d 3 обозначает F; d 4 обозначает М, Q, Е или K; d 6 обозначает L или М; d 8 обозначает D или Е; d 10 обозначает Е; d 11 обозначает Q или Е; d 12 обозначает Т или R; d 15 обозначает Y, D, Е или K; d 16 обозначает Q; d 17 обозначает W или F; d 18 обозначает L, I, М или Т; d 19 обозначает L, F или Y; d 20 обозначает Q, D или Е; d 21 отсутствует или обозначает Q или Н; d 22 отсутствует или обозначает A, L, G, S или R; и его физиологически приемлемые соли. 36. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 2, состоящей из SEQ ID NO: 6 и с SEQ ID NO: 119 по SEQ ID NO: 142 включительно, и его физиологически приемлемые соли. Таблица 2 37. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пептид по пп.31, 34 или 36 и носитель, и его физиологически приемлемые соли. 38. Слитый полипептид по п.37, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 39. Полипептид по пп.31, 34 или 36, который является циклическим. 40. Димер или мультимер полипептидов по пп.31, 34 или 36. 41. Состав полипептида, имеющий формулу (Х 1 ) a -F 1 -(X 2 ) b и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид по пп.31, 34 или 36; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначают линкеры и а, b, с, d, e и f, каждый независимо, обозначают 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 42. Состав полипептида по п.41, отличающийся тем, что один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 6 и с SEQ ID NO: 119 по SEQ ID NO: 142 включительно. 43. Состав полипептида по п.41, соответствующий формулам X 1 -F 1 или F 1 -X 2 и его физиологически приемлемые соли. 44. Состав полипептида по п.41 формулы F 1 -(L 1 ) c -P 1 и его физиологически приемлемые соли. 45. Состав полипептида по п.41, соответствующий формуле F 1 -(L 1 ) c -P 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 46. Состав полипептида по п.41, соответствующий формуле P 1 -(L 1 ) c -F 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 47. Состав полипептида по п.41, отличающийся тем, что F 1 обозначает домен Fc или его фрагмент. 48. Состав полипептида по п.41, отличающийся тем, что F 1 содержит аминокислотную последовательность SEQ ID NO: 60. 49. Полинуклеотид, кодирующий полипептид по любому из пп.31, 34 или 36. 50. Вектор экспрессии, содержащий полинуклеотид по п.49. 51. Клетка-хозяин, содержащая вектор экспрессии по п.50. 52. Клетка-хозяин по п.51, отличающаяся тем, что является клеткой прокариот. 53. Клетка-хозяин по п.52, отличающаяся тем что является клеткой Е. coli. 54. Клетка-хозяин по п.51, отличающаяся тем, что является клеткой эукариот. 55. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы RPe 3 e 4 e 5 e 6 e 7 G (SEQ ID NO: 73) где е 3 обозначает нейтральный полярный аминокислотный остаток; е 4 обозначает кислый аминокислотный остаток; е 5 обозначает нейтральный полярный или кислый аминокислотный остаток; е 6 обозначает нейтральный гидрофобный аминокислотный остаток; е 7 обозначает нейтральный гидрофобный аминокислотный остаток; и его физиологически приемлемые соли. 56. Полипептид по п.55, отличающийся тем, что е 3 обозначает Y или С. 57. Полипептид по п.55, отличающийся тем, что е 4 обозначает D или Е. 58. Полипептид по п.55, отличающийся тем, что е 6 обозначает I или М. 59. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы f 1 f 2 f 3 f 4 RPf 7 f 8 f 9 f 10 f 11 Gf 13 f 14 f 15 f 16 f 17 f 18 f 19 f 20 (SEQ ID NO: 74) где f 1 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 2 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 3 обозначает нейтральный полярный или кислый аминокислотный остаток; f 4 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 7 обозначает нейтральный полярный аминокислотный остаток; f 8 обозначает кислый аминокислотный остаток; f 9 обозначает нейтральный полярный или кислый аминокислотный остаток; f 10 обозначает нейтральный гидрофобный аминокислотный остаток; f 11 обозначает нейтральный гидрофобный аминокислотный остаток; f 13 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f 15 обозначает нейтральный полярный аминокислотный остаток; f 16 обозначает нейтральный полярный аминокислотный остаток; f 17 обозначает нейтральный полярный или кислый аминокислотный остаток; f 18 обозначает нейтральный гидрофобный или основной аминокислотный остаток; f 19 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и f 20 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; и его физиологически приемлемые соли. 60. Полипептид по п.59, отличающийся тем, что f 1 обозначает S, А или G; f 2 обозначает G, Q или Р; f 3 обозначает Q, G или D; f 4 обозначает L, М или Q; f 7 обозначает С или Y; f 8 обозначает Е или D; f 9 обозначает Е, G или D; f 10 обозначает I или М; f 11 обозначает F или L; f 13 обозначает С или W; f 14 обозначает G или Р; f 15 обозначает Т или N; f 16 обозначает Q, Y или K; f 17 обозначает N, D или Q; f 18 обозначает L, V, W или R; f 19 обозначает A, Q, Y или I и f 20 обозначает L, A, G или V; и его физиологически приемлемые соли. 61. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 3, состоящей из SEQ ID NO: 3 и с SEQ ID NO: 143 по SEQ ID NO: 148 включительно, и его физиологически приемлемые соли Таблица 3 62. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один полипептид по пп.55, 59 или 61 и носитель и его физиологически приемлемые соли. 63. Слитый полипептид по п.62, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 64. Полипептид по пп.55, 59 или 61, который является циклическим. 65. Димер или мультимер полипептидов по пп.55, 59 или 61. 66. Состав полипептида, имеющий формулу (X 1 ) a -F 1 -(X 2 ) b и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид по пп.55, 59 или 61; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначают линкеры и а, b, с, d, е и f, каждый независимо, обозначают 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 67. Состав полипептида по п.66, отличающийся тем, что один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 3 и с SEQ ID NO: 143 по SEQ ID NO: 148 включительно. 68. Состав полипептида по п.66, соответствующий формулам X 1 -F 1 или F 1 -X 2 и его физиологически приемлемые соли. 69. Состав полипептида по п.66 формулы F 1 -(L 1 ) c -P 1 и его физиологически приемлемые соли. 70. Состав полипептида по п.66, соответствующий формуле F 1 -(L 1 ) c -P 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 71. Состав полипептида по п.66, соответствующий формуле P 1 -(L 1 ) c -F 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 72. Состав полипептида по п.66, отличающийся тем, что F 1 обозначает домен Fc или его фрагмент. 73. Состав полипептида по п.66, отличающийся тем, что F 1 содержит аминокислотную последовательность SEQ ID NO: 60. 74. Полинуклеотид, кодирующий полипептид по любому из пп.55, 59 или 61. 75. Вектор экспрессии, содержащий полинуклеотид по п.74. 76. Клетка-хозяин, содержащая вектор экспрессии по п.75. 77. Клетка-хозяин по п.76, отличающаяся тем, что является клеткой прокариот. 78. Клетка-хозяин по п.77, отличающаяся тем что является клеткой Е. coli. 79. Клетка-хозяин по п.76, отличающаяся тем, что является клеткой эукариот. 80. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы Cg 2 Gg 4 g 5 DPFTg 10 GCg 13 (SEQ ID NO: 75) где g 2 обозначает кислый аминокислотный остаток; g 4 обозначает нейтральный гидрофобный аминокислотный остаток; g 5 обозначает Е, D или Q; g 10 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; g 13 обозначает кислый остаток; и его физиологически приемлемые соли. 81. Полипептид по п.80, отличающийся тем, что g 2 обозначает Е или D. 82. Полипептид по п.80, отличающийся тем, что g 4 обозначает V или М. 83. Полипептид по п.80, отличающийся тем, что g 10 обозначает F или Q. 84. Полипептид по п.80, отличающийся тем, что g 13 обозначает D или Е. 85. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы h 1 h 2 h 3 h 4 Ch 6 Gh 8 h 9 DPFTh 14 GCh 17 h 18 h 19 h 20 (SEQ ID NO:158) где h 1 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; h 2 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h 3 обозначает кислый аминокислотный остаток; h 4 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h 6 обозначает кислый аминокислотный остаток; h 8 обозначает нейтральный гидрофобный аминокислотный остаток; h 9 обозначает Е, D или Q; h 14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h 17 обозначает кислый аминокислотный остаток; h 18 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; h 19 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и h 20 отсутствует или обозначает аминокислотный остаток; и его физиологически приемлемые соли. 86. Полипептид по п.85, отличающийся тем, что h 1 отсутствует или обозначает A, L, M, G, K или Н; h 2 обозначает L, F или Q; h 3 обозначает D или Е; h 4 обозначает W или Y; h 6 обозначает D или Е; h 8 обозначает V или М; h 14 обозначает F или Q; h 17 обозначает D или Е; h 18 обозначает М, Y, N или K; h 19 обозначает L или Q и h 20 отсутствует или обозначает М, Т, G, S, D, K или R; и его физиологически приемлемые соли. 87. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 4, состоящей из SEQ ID NO: 5 или с SEQ ID NO: 149 по SEQ ID NO: 157 включительно, и его физиологически приемлемые соли.
Таблица 4 88. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пептид по пп.80, 85 или 87 и носитель, его физиологически приемлемые соли. 89. Слитый полипептид по п.88, отличающийся тем, что носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 90. Полипептид по пп.80, 85 или 87, который является циклическим. 91. Димер или мультимер полипептидов по пп.80, 85 или 87. 92. Состав полипептида, имеющий формулу (X 1 ) a -F 1 -(X 2 ) b и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид по пп.80, 85 или 87; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначает линкеры и а, b, с, d, e и f, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 93. Состав полипептида по п.92, отличающийся тем, что один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 5 и с SEQ ID NO: 149 по SEQ ID NO: 157 включительно. 94. Состав полипептида по п.92, соответствующий формулам X 1 -F 1 или F 1 -X 2 и его физиологически приемлемые соли. 95. Состав полипептида по п.92 формулы F 1 -(L 1 ) c -P 1 и его физиологически приемлемые соли. 96. Состав полипептида по п.92, соответствующий формуле F 1 -(L 1 ) c -P 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 97. Состав полипептида по п.92, соответствующий формуле P 1 -(L 1 ) c -F 1 -(L 2 ) d -P 2 и его физиологически приемлемые соли. 98. Состав полипептида по п.92, отличающийся тем, что F 1 обозначает домен Fc или его фрагмент. 99. Состав полипептида по п.92, отличающийся тем, что F 1 содержит аминокислотную последовательность SEQ ID NO: 60. 100. Полинуклеотид, кодирующий полипептид по любому из пп.80, 85 или 87. 101. Вектор экспрессии, содержащий полинуклеотид по п.100. 102. Клетка-хозяин, содержащая вектор экспрессии по п.101. 103. Клетка-хозяин по п.102, отличающаяся тем, что является клеткой прокариот. 104. Клетка-хозяин по п.103, отличающаяся тем что является клеткой Е. coli. 105. Клетка-хозяин по п.102, отличающаяся тем, что является клеткой эукариот. 106. Полипептид в соответствии с любой из последовательностей SEQ ID NO: l, SEQ ID NO: 2, SEQ ID NO: 3 или SEQ ID NO: 4 или его вариант. 107. Полипептид в соответствии с любой из последовательностей SEQ ID NO: 5, SEQ ID NO: 6 или SEQ ID NO: 7 или его вариант. 108. Состав полипептида формулы (X 1 ) q -F 1 -(X 2 ) r и его мультимеры, где F 1 обозначает носитель; X 1 и X 2 , каждый независимо, выбирают из где один или более из Р 1 , Р 2 , Р 3 и Р 4 , каждый независимо, представляют собой полипептид, выбранный из группы, состоящей из: (a) аминокислотной последовательности WDPWT (SEQ ID NO: 65), причём указанный полипептид имеет в длину 5-50 аминокислот; (b) аминокислотной последовательности WDPWTC (SEQ ID NO: 66); (c) аминокислотной последовательности Cz 2 WDPWT (SEQ ID NO: 67), где z 2 обозначает кислый или нейтральный полярный аминокислотный остаток; (d) аминокислотной последовательности Cz 2 WDPWTC (SEQ ID NO: 68), где z 2 обозначает кислый или нейтральный полярный аминокислотный остаток; (e) аминокислотной последовательности Рс 2 Dc 4 Lc 6 с 7 с 8 LY (SEQ ID NO: 71), где с 2 обозначает нейтральный гидрофобный аминокислотный остаток; с 4 обозначает A, D или Е; с 6 обозначает кислый аминокислотный остаток; с 7 обозначает аминокислотный остаток и с 8 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; (f) аминокислотной последовательности RPe 3 e 4 e 5 e 6 e 7 G (SEQ ID NO: 73), где е 3 обозначает нейтральный полярный аминокислотный остаток; е 4 обозначает кислый аминокислотный остаток; е 5 обозначает нейтральный полярный или кислый аминокислотный остаток; е 6 обозначает нейтральный гидрофобный аминокислотный остаток и е 7 обозначает нейтральный гидрофобный аминокислотный остаток; (g) аминокислотной последовательности Cg 2 Cg 4 g 5 DPFTg 10 GCg 13 (SEQ ID NO: 75), где g 2 обозначает кислый аминокислотный остаток; g 4 обозначает нейтральный гидрофобный аминокислотный остаток; g 5 обозначает нейтральный полярный или кислый аминокислотный остаток; g 10 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и g 13 обозначает кислый остаток; (h) полипептида SEQ ID NO: 1; (i) полипептида SEQ ID NO: 2 и (j) полипептида SEQ ID NO: 7; L 1 , L 2 , L 3 и L 4 , каждый независимо, обозначает линкеры и q, r, s, t, u и v, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из q и r обозначает 1; и его физиологически приемлемые соли. 109. Фармацевтическая композиция, содержащая эффективное количество состава полипептида по пп.1, 31, 55 или 80 в смеси с фармацевтически приемлемым носителем. 110. Способ ингибирования нежелательного ангиогенеза у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 111. Способ лечения ангиогенеза у субъекта, заключающийся во введении эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 112. Способ модулирования ангиогенеза у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 113. Способ подавления роста опухоли, характеризующейся нежелательным ангиогенезом, у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 114. Способ лечения рака у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92 и химиотерапевтического агента. 115. Способ по п.114, отличающийся тем, что химиотерапевтический агент является по меньшей мере одним из 5-FU, СРТ-11 и Таксотер. 116. Способ модулирования по меньшей мере одного нарушения у млекопитающего: сосудистой проницаемости или истечения плазмы, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 117. Способ лечения по меньшей мере одного из заболеваний: реваскулярного заболевания глаза, ожирения, ангиоретикулёмы, гемангиомы, артериосклероза, воспалительного заболевания, воспалительных нарушений, атеросклероза, эндометриоза, опухолевого заболевания, костного заболевания или псориаза, у млекопитающего, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92.
008248 Область, к которой относится изобретение Настоящее изобретение относится к агентам специфического связывания, которые распознают ан-гиопоэтин-2 (Ang-2) и связываются с ним. Более конкретно, изобретение относится к получению, диагностическому и терапевтическому применению специфически связывающихся агентов и их фрагментов, которые специфически связываются с Ang-2. Предпосылки создания изобретения Ангиогенез, образование новых кровеносных сосудов из уже имеющихся, является существенно важным для многих физиологических и патологических процессов. Обычно ангиогенез непосредственно регулируется про- и антиангиогенными факторами, но в случае таких болезней как рак, глазные реваскулярные заболевания, артрит и псориаз, процесс может идти "неправильно". Folkman, J. Nat. Med., 1:27-31 (1995). Известно, что многие заболевания ассоциируются с разрегулированным или нежелательным ангио-генезом. Такие заболевания включают, но без ограничения, глазную реваскуляризацию, такую как ретинопатии (включая диабетическую ретинопатию), пятнистую дегенерацию, связанную со старением, псориаз, ангиоретикулёму, гемангиому, артериосклероз, воспалительное заболевание, такое как ревматоидное или ревматическое воспалительное заболевание, в особенности артрит (включая ревматоидный артрит), или другие хронические воспалительные заболевания, такие как хроническая астма, артериальный или пост-трансплантационный атеросклероз, эндометриоз и опухолевые заболевания, такие, например, как так называемые солидные опухоли и "жидкие" (или гемопоэтические) опухоли (такие как лейкемии и лимфомы). Другие заболевания, связанные с нежелательным ангиогенезом, очевидны для специалистов в данной области техники. Хотя в регуляции ангиогенеза участвуют многие системы сигнальной трансдукции, одна из лучше всего охарактеризованных и наиболее селективных в отношении эндотелиальных клеток систем включает Tie-2 рецепторную тирозинкиназу (называемую как "Tie-2" или "Tie-2R" (также называемую "ORK")); мышиную Tie-2, называемую также "tec") и её лиганды, ангиопоэтины (Gale, N.W. and Yancopoulos, ОЛ., Genes Dev. 13:1055-1066 [1999]). Существуют 4 известных ангиопоэтина; от ангиопоэтина-1 ("Ang-1") до ангиопоэтина-4 ("Ang-4"). Эти ангиопоэтины можно также называть "Tie-2 лиганды" (Davis, S., et al., Cell, 87:1161-1169 [1996]; Grosios, K., et al., Cytogenet. Cell Genet., 84:118-120 [1999]; Holash, J., et al., Investigative Ophthalmology & Visual Science, 42(40?):1617-1625 [1999]; Koblizek, T.I., et al., Current Biology, 8:529-532 [1998]; Lin, P., et al., Proc. Natl. Acad. Sci. USA, 95:8829-8834 [1998]; Maisonpierre, P.C., et al., Science, 277:55-60 [1997]; Papapetropoulos, A., et al., Lab. Invest., 79:213-223 [1999]; Sato, T.N., et al., Nature, 375:70-74 [1998]; Shyu, K.G., et al., Circulation, 98:2081-2087 [1998]; Suri, C., et al., Cell, 87:1171-1180 [1996]; Suri, C., et al., Science, 282:468-471 [1998]; Valenzuela, D.M., et al., Proceedings of the National Academy of Sciences USA, 98:1904-1909 [1999]; Witzenbichler, В., et al., J. Biol. Chem., 273:18514-18521 [1998]). Наблюдали, что в то время как связывание Ang-1 с Tie-2 стимулирует рецепторное фосфорили-рование в культивированных эндотелиальных клетках, Ang-2 является как агонистом, так и антагонистом Tie-2 рецепторного фосфорилирования (Davis, S., et al., [1996], см. выше; Maisonpierre, Р.С., et al., [1997], см. выше; Kim, I., J.H. Kim, et al., Oncogene 19(39): 4549-4552 (2000); Teichert-Kuliszewska, K., Р.С. Maisonpierre, et al., Cardiovascular Research 49(3): 659-70 (2001)). Фенотипы нокаутов мышей Tie-2 и Ang-1 сходны, и полагают, что стимулированное Ang-1 фосфо-рилирование Tie-2 опосредует реструктурирование и стабилизацию образующихся сосудов в матке за счёт сохранения клеточной адгезии к эндотелиальным клеткам в качестве подложки (Dumont, D.J., et al., Genes & Development, 8:1897-1909 [1994]; Sato, T.N., et al., Nature, 376:70-74 [1995]; Suri, C., et al., [1996], см. выше). Полагают, что роль Ang-1 в стабилизации сосудов сохраняется у взрослых, где он экспресси-рует широко и конститутивно (Hanahan, О., Science, 277:48-50 [1997]; Zagzag, D., et al., Experimental Neurology, 159:391-400 [1999]). Напротив, экспрессия Ang-2, прежде всего, ограничена сайтами васкулярно-го (сосудистого) рестуктурирования, где, как полагают, блокируется функция Ang-1, тем самым вызывается состояние сосудистой пластичности, ведущее к ангиогенезу (Hanahan, D., [1997], см. выше; Holash, J., et al., Science, 284:1994-1998 [1999]; Maisonpierre, P.C., et al., [1997], см. выше). Многочисленные опубликованные исследования как будто демонстрируют экспрессию Ang-2 при болезненных состояниях, ассоциированных с ангиогенезом. (Bunone, G., et al., American Journal of Pathology, 155:1967-1976 [1999]; Etoh, Т., et al., Cancer Research, 61:2145-2153 [2001]; Hangai, M., et al., Investigative Ophthalmology & Visual Science, 42:1617-1625 [2001]; Holash, J., et al., [1999] см. выше; Kuroda, K., et al., Journal of Investigative Dermatology, 116:713-720 [2001]; Otani, A., et al., Investigative Ophthalmology & Visual Science, 40:1912-1920 [1999]; Stratmann, A., et al., American Journal of Pathology, 153:1459-1466 [1998]; Tanaka, S., et al., J. Clin. Invest., 103:34-345 [1999]; Yoshida, Y., et al., International Journal of Oncology, 15:1221-1225 [1999]; Yuan, K., et al., Journal of Periodontal Research, 35:165-171 [2000]; Zagzag, D., et al., [1999] см. выше). Основная часть этих исследований сосредоточена на раке, при этом многие типы опухолей выявляют сосудистую экспрессию Ang-2. В отличие от его экспрессии при патологическом ангио-генезе экспрессия Ang-2 в нормальных тканях чрезвычайно ограничена (Maisonpierre, Р.С., et al., [1997], см. выше; Mezquita, J., et al., Biochemical and Biophysical Research Communications, 260:492-498 [1999]). У здорового взрослого человека существуют три основных сайта ангиогенеза, это яичник, плацента и матка; эти ткани являются первичными в нормальных (не злокачественных) тканях, где обнаружена мРНК Ang-2. - 1 - 008248 В некоторых функциональных исследованиях высказывается предположение, что Ang-2 может принимать участие в ангиогенезе опухолей. Ahmad et al. (Cancer Res., 61:1255-1259 [2001]) описывает сверхэкспрессию Ang-2 и показывает на мышиной модели ксенотрансплантата, что она, по-видимому (якобы), ассоциирована с ускоренным (повышенным) ростом опухоли. См. также выше Etoh et al., и Tanaka et al., данные в которых предполагают связь сверхэкспрессии Ang-2 с гиперваскулярностью опухолей. Напротив, данные, сообщаемые Yu et al. (Am. J. Path., 158:563-570 [2001]), показывают, что сверхэкспрессия Ang-2 в раковых клетках при раке лёгкого Льюиса и в раковых клетках ТА3 молочной железы, по-видимому, продлевает продолжительность жизни мышей, которым инъецированы соответствующие трансфектанты. В последние несколько лет в различных публикациях предлагаются Ang-1, Ang-2 и/или Tie-2 в качестве возможной мишени для противораковой терапии. Например, в каждом из патентов США 6166185, 5650490 и 5814464 описываются представления об антителах против лиганда Tie-2 ligand и рецепторных "тельцах". Lin et al. (Proc. Natl. Acad. Sci. USA, 95:8829-8834 [1998]) инъецировали аденовирус, экспрес-сирующий растворимый Tie-2, мышам; растворимый Tie-2, по-видимому (якобы), уменьшал число и размер опухолей, образовавшихся у мышей. Lin et al. (J. Clin. Invest., 100:2072-2078 [1997]), инъецировали растворимую форму Tie-2 крысам; как пишут, это соединение уменьшало размер опухолей у крыс. Siemeister et al. (Cancer Res., 59:3185-3189 [1999]) генерировали клеточные линии человеческой мелано-мы, экспрессирующие внеклеточную область Tie-2, инъецировали эти клеточные линии "голым" мышам и сделали вывод, что Tie-2 (как полагают) вызывает "заметное ингибирование" роста опухоли и ангиоге-неза опухоли. В свете этой информации и при условии, что Ang-1 и Ang-2 связываются с Tie-2, из этих исследований не ясно, являются ли Ang-1, Ang-2 или Tie-2 привлекательными в качестве мишени для противораковой терапии. Слияние некоторых пептидов с устойчивым плазменным белком, таким как константная область Ig, с целью увеличения периода полужизни этих молекул описано, например, в международной заявке РСТ WO 00/24782, опубликованной 4 мая 2000 г. Слияние белка или его фрагмента с устойчивым белком плазмы, таким как константная область Ig, с целью увеличения периода полужизни этих молекул многократно описано (см., например, патент США 5480981; Zheng et al., J. Immunol., 154:5590-5600, (1995); Fisher et al., N. Engl. J. Med., 334:1697-1702, (1996); Van Zee, K. et al., J. Immunol., 156:2221-2230, (1996); патент США 5808029, вьвданньш 15 сентября 1998 г.; Capon et al., Nature, 337:525-531, (1989); Harvill et al., Immunotech., 1:95-105, (1995); международная заявка WO 97/23614, опубликованная 3 июля 1997 г.; заявка PCT/US 97/23183, поданная 11 декабря 1997 г.; Linsley, J. Exp. Med., 174:561-569, (1991); международная заявка WO 95/21258, опубликованная 10 августа 1995 г.). Эффективная affra-Ang-2 может принести пользу многочисленной группе больных раком, так как для роста твёрдых опухолей более 1-2 мм в диаметре требуется реваскуляризация. Такая терапия могла бы найти более широкое применение при других ассоциированных с ангиогенезом заболеваниях, таких как ретинопатии, артрит и псориаз. Существует настоятельная необходимость в идентификации новых агентов, которые специфически распознают Ang-2 и связываются с ним. Такие агенты были бы полезны для диагностического поиска и терапевтического вмешательства в болезненные состояния, ассоциированные с активностью Ang-2. Таким образом, целью настоящего изобретения является создание специфических агентов для связывания Ang-2, которые модулируют активность Ang-2. Такие агенты по данному изобретению имеют форму "пептител" (peptibodies), т.е. пептидов, слитых с другими молекулами, такими как Fc домен антитела, при этом пептидный фрагмент специфически связывается с Ang-2. Сущность изобретения Настоящее изобретение в одном своём варианте относится к пептидам (также называемым в данном описании "полипептиды"), которые связываются с Ang-2. Также данное изобретение охватывает варианты и производные таких пептидов. В другом варианте изобретения пептиды и их варианты и производные по данному изобретению связаны с носителями. В другом варианте изобретения пептиды могут быть слиты с Fc доменами, тем самым создавая "пептитела" (peptibodies). "Пептитела", необязательно, содержат по меньшей мере один пептид, например, из SEQ ID NO: 3-SEQ ID NO: 6 или SEQ ID NO: 76-SEQ ID NO: 157, а также их варианты и производные. Кроме того, пептиды могут содержать по меньшей мере один пептид, соответствующий формулам, представленным SEQ ID NO: 65-SEQ ID NO: 75 и SEQ ID NO: 158. Ещё в одном варианте изобретение охватывает молекулы нуклеиновых кислот, кодирующие специфически связывающиеся агенты и их варианты и производные. Ещё в одном варианте изобретение включает молекулы нуклеиновых кислот, кодирующих "пепти-тела", а также их варианты и производные. Необязательно, такие молекулы нуклеиновых кислот включают SEQ ID NO: 33-SEQ ID NO: 53. Ещё в одном варианте изобретение охватывает способ уменьшения опухоли путём введения эффективного количества специфически связывающихся агентов по настоящему изобретению нуждающемуся в этом субъекту. Изобретение также охватывает способ ингибирования ангиогенеза у субъекта, заключающийся во введении эффективного количества специфически связывающихся агентов по данному - 2 - 008248 изобретению нуждающемуся в нём субъекту. Кроме того, изобретение включает способ лечения рака субъекту, заключающийся во введении эффективного количества специфически связывающихся агентов по данному изобретению субъекту, нуждающемуся в этом. Изобретение также относится к полипептиду, способному связывать Ang-2, причём этот полипептид содержит аминокислотную последовательность WDPWT (SEQ ID NO: 65) и длина этого полипептида 5-50 аминокислот, а также к его физиологически приемлемым солям. Изобретение может также включать полипептид с аминокислотной последовательностью WDPWTC (SEQ ID NO: 66) и его физиологически приемлемые соли. Кроме того, изобретение охватывает полипептид, который может содержать аминокислотную последовательность Cz2WDPWT (SEQ ID NO: 67) где z2 представляет собой кислый или нейтральный полярный аминокислотный остаток, и его физиологически приемлемые соли. Кроме того, изобретение относится к полипептиду, который может содержать аминокислотную последовательность Cz2WDPWTC (SEQ ID NO: 68) где z2 обозначает кислый или нейтральный полярный аминокислотный остаток, и его физиологически приемлемым солям. В другом варианте изобретение относится к полипептиду, способному связываться с Ang-2, содержащим аминокислотную последовательность формулы ala2a3Ca5WDPWTCa12a13a14 (SEQ ID NO: 69) где а1, а2 и а3, каждый независимо, обозначают аминокислотные остатки; а5 обозначает аминокислотный остаток; а12 отсутствует или обозначает аминокислотный остаток; а13 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; а14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; и его физиологически приемлемым солям. В предпочтительном варианте изобретения а1 обозначает V, I, P, W, G, S, Q, N, Е, K, R или Н; а2 обозначает V, Р, М, G, S, Q, D, Е, K, R или Н; а3 обозначает А, V, Р, М, F, T, G, D, Е, K или Н; а8 обозначает А, V, G, Q, N, D или Е; а12 обозначает S, Q, N, D, Е, K или R; а13 обозначает L, Т или Н и а14 обозначает V, L, I, W или М. В более предпочтительном варианте изобретения а1 обозначает Q; а2 обозначает Е; а3 обозначает Е; а5 обозначает D или Е; а12 обозначает D или Е; а13 обозначает Н и а14 обозначает М. Понятно, что строчные буквы (нижние литеры) с верхними индексами по данному описанию (например, а1 и b1) определяют положения аминокислот, но не являются однобуквенными сокращениями для данной аминокислоты. Далее изобретение относится к полипептиду, способному связываться с Ang-2, содержащему аминокислотную последовательность формулы bVbWbWWDPWTC^VVVVV0 (SEQ ID NO: 70) где b1 отсутствует или обозначает аминокислотный остаток; b2 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; b3, b4, b5 и b6, каждый независимо, отсутствуют или обозначают аминокислотные остатки; b8 обозначает аминокислотный остаток; b15 отсутствует или обозначает аминокислотный остаток; b16 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; - 3 - 008248 b17 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; b18, b19 и b20, каждый независимо, отсутствуют или обозначают аминокислотные остатки; и их физиологически приемлемые соли. В предпочтительном варианте b1 отсутствует или обозначает А, V, L, P, W, F, T, G, S, Q, N, K, R или Н; b2 отсутствует или обозначает А, V, L, I, P, W, M, T, G, S, Y, N, K, R или Н; b3 отсутствует или обозначает A, L, I, P, W, M, T, G, S, Q, N, E, R или Н; b4 обозначает V, I, P, W, G, S, Q, N, Е, K, R или Н; b5 обозначает V, Р, М, G, S, Q, D, Е, K, R или Н; b6 обозначает А, V, Р, М, F, T, G, D, Е, K или Н; b8 обозначает А, V, G, Q, N, D или Е; b15 обозначает S, Q, N, D, Е, K или R; b16 обозначает L, Т или Н; b17 обозначает V, L, I, W или М; b18 отсутствует или обозначает А, V, L, P, W, F, T, G, Y, Q, D, Е или R; b19 отсутствует или обозначает V, L, I, P, T, G, S, Y, Q, N, D, Е или R и b20 отсутствует или обозначает V, L, P, W, M, T, G, S, Y, Q, N, D, K или R. В более предпочтительном варианте изобретения b1 отсутствует или обозначает Р или Т; b2 отсутствует или обозначает I или N; b3 отсутствует или обозначает R или I; b4 обозначает Q; b5 обозначает Е; b6 обозначает Е; b8 обозначает D или Е; b15 обозначает D или Е; b16 обозначает Н; b17 обозначает М; b18 отсутствует или обозначает W или Р; b19 отсутствует или обозначает G или Е; и b20 отсутствует или обозначает V или K. Также будет оценено, что изобретение предпочтительно относится к полипептиду, содержащему по меньшей мере одну аминокислотную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 4 и SEQ ID NO: 76-SEQ ID NO: 118 включительно, причём полипептид способен связываться с Ang-2, a также его физиологически приемлемые соли. Ниже представлены пептидные последовательности. Таблица 1 ПЕПТИД SEQ ID NO ПЕПТИДНАЯ ПОСЛЕДОВА -ТЕЛЬНОСТЬ Соп4-44 PIRQEECDWDPWTCEHMWEV Соп4-40 TNIQEECEWDPWTCDHMPGK Соп4-4 WYEQDACEWDPWTCEHMAEV Соп4-31 NRLQEVCEWDPWTCEHMENV Соп4-С5 AATQEECEWDPWTCEHMPRS Соп4-42 LRHQEGCEWDPWTCEHMFDW Соп4-35 VPRQKDCEWDPWTCEHMYVG Соп4-43 SISHEECEWDPWTCEHMQVG Соп4-49 WAAQEECEWDPWTCEHMGRM Соп4-27 TWPQDKCEWDPWTCEHMGST Соп4-48 GHSQEECGWDPWTCEHMGTS Соп4-46 QHWQEECEWDPWTCDHMPSK Соп4-41 NVRQEKCEWDPWTCEHMPVR Соп4-36 KSGQVECNWDPWTCEHMPRN Соп4-34 VKTQmCDWDPWTCEHMREW Соп4-28 AWGQEGCDWDPWTCEHMLPM Соп4-39 PVNQEDCEWDPWTCEHMPPM Соп4-25 RAPQEDCEWDPWTCAHMDЖ Соп4-50 HGQNMECEWDPWTCEHMFRY Соп4-38 PRLQEECVWDPWTCEHMPLR Соп4-29 RTTQEKCEWDPWTeEHMES Q Соп4-47 QTSQEDCVWDPWTCDHMVSS Соп4-20 QVIGRPCEWDPWTCEHLEGL Соп4-45 WAQQEECAWDPWTCDHMVGL Соп4-37 100 LPGQEDCEWDPWTCEHMVRS Соп4-33 101 PMNQVECDWDPWTCEHMPRS АС2-Соп4 102 FGWSHGCEWDPWTCEHMGST Соп4-32 103 KSTQDDCDWDPWTCEHMVGP Соп4-17 104 GPRISTCOWDPWTCEHMDQL Соп4-8 105 STIGDMCEWDPWTCAHMQVD АС4-Соп4 106 VLGGQGCEWDPWTCRJLLQGW Соп4-1 107 VLGGQGCQWDPWTCSHLEDG - 4 - 008248 Con4-Cl 108 TTIGSMCEWDPWTCAHMQGG Con4-21 109 TKGKS VCQWDPWTCSHMQS G Соп4-С2 TTIGSMCQWDPWTCAHMQGG Соп4-18 111 WV№W(^WDPWTCNHWDTP Соп4-19 112 WQVGMCQWDPWTCKHMRLQ Соп4-16 113 AVGSQTCEWDPWTCAHLVEV Соп4-11 114 QGMKMFCEWDPWTCAHTVYR Соп4-С4 115 TTIGSMCQWDPWTCEHMQGG Соп4-23 116 TS QRVGCEWDPWTCQHLTYT Соп4-15 117 QWSWPPCEWDPWTCQTVWPS Соп4-9 118 GTSPSFCQWDPWTCSHMVQG TNS-Con4* QEECEWDPWTCEHM *Следует понимать, что названия некоторых пептидов и/или "пептител" могут содержать префикс "TN", "TN8" или "TN12" и что этот префикс может присутствовать или может отсутствовать в названии (обозначении) данного "пептитела". Так например, в данном описании чередуются взаимозаменяемые термины "TN8-Con4" и "СОП4". В другом варианте предметом изобретения является состав полипептида, отвечающий формуле (X1)a-F1-(X2)b и его мультимеры, где F1 обозначает носитель; X1 и X2, каждый независимо, выбирают из -(L')c-P1; -(LVP'-CLVP2; -(L^c-P1 -(L2)d-P2-(L3)E-P3; -4LVP4LVP4L3)E-P3-(L4)RP4; где один или более из Р1, Р2, Р3 и Р4, каждый независимо, представляет собой полипептид по данному описанию. Например, в предпочтительном варианте изобретения Р1, Р2, Р3 и Р4, каждый независимо, представляет собой полипептид SEQ ID NO: 3 - SEQ ID NO: 6 и/или SEQ ID NO: 76 - SEQ ID NO: 157. В другом варианте предметом изобретения является состав полипептида, отвечающий формулам X1-F1 или F1-X2 и его физиологически приемлемые соли, где X1, F1 и X2 имеют значение по данному описанию. В другом варианте предметом изобретения является состав полипептида, соответствующий формуле F1-(L1)c-P1 и его физиологически приемлемые соли, где L1, F1 и Р1 имеют значение по определению в данном описании. Ещё в одном варианте предметом изобретения является состав полипептида, соответствующий формуле F1-(1L1)1c-P11-(L22)d-P2 и его физиологически приемлемые соли, где L1, F1, Р1, Р2 и с и d имеют значение по определению в данном описании. Ещё в одном варианте предметом изобретения является состав полипептида, соответствующий формуле P1-(L1)c-F1-(L2)d-P2 1 и его физиологически приемлемые соли. В предпочтительном варианте изобретения F1 обозначает домен Fc или его фрагмент. Кроме того, изобретение относится к полипептиду, способному связывать Ang-2, содержащему аминокислотную последовательность формулы Pc2Dc4Lc6c7c8LY (SEQ ID NO: 71) где с2 обозначает нейтральный гидрофобный аминокислотный остаток; с4 обозначает A, D или Е; с6 обозначает кислый аминокислотный остаток; с7 обозначает аминокислотный остаток и с8 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток, и к его физиологически приемлемым солям. В предпочтительном варианте изобретения с2 обозначает L или М. В другом предпочтительном варианте с6 обозначает D или Е. Кроме того, изобретение относится к полипептиду, способному связываться с Ang-2, содержащему аминокислотную последовательность формулы d1d2d3d4Pd6Dd8Ld10d11d12LYd15d16d17d18d19d20d21d22 1 (SEQ ID NO: 72) где d1 отсутствует или обозначает аминокислотный остаток; - 5 - 008248 d2 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d3 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d4 отсутствует или обозначает аминокислотный остаток; d6 обозначает нейтральный гидрофобный аминокислотный остаток; d8 обозначает A, D или Е; d10 обозначает кислый аминокислотный остаток; d11 обозначает аминокислотный остаток; d12 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; d15 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d16 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d17 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d18 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d19 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; d20 отсутствует или обозначает аминокислотный остаток; d21 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d22 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; и к его физиологически приемлемым солям. В предпочтительном варианте изобретения d1 обозначает Т, S, Q, R или Н; d2 обозначает Т, Q, N или K; d3 обозначает F; d4 обозначает М, Q, Е или K; d6 обозначает L или М; d8 обозначает D или Е; d10 обозначает Е; d11 обозначает Q или Е; d12 обозначает Т или R; d15 обозначает Y, D, Е или K; d16 обозначает Q; d17 обозначает W или F; d18 обозначает L, I, М или Т; d19 обозначает L, F или Y; d20 обозначает Q, D или Е; d21 отсутствует или обозначает Q или Н; d22 отсутствует или обозначает A, L, G, S или R. В предпочтительном варианте изобретения полипептид содержит по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6 и SEQ ID NO: 119 - SEQ ID NO: 142 включительно, причём полипептид способен связываться с Ang-2. SEQ ID NO: 6 и SEQ ID NOS: 119-142 представлены ниже. - 6 - 008248 Ll-l 119 QN\^mELDATXYEHFIFrIYT Ll-2 120 li^FTPLDELEQTLYEQWTLQQS Ll-3 121 TKENPLDELEQTLYEQWTLQHQ Ы-4- ~ - - 122 VE^KPmAXEQTLYEHWMFQQA Ll-5 123 VKYKPLDELDEILYEQQTFQER Ll-7 124 TNFMPMDDLEQRLYEQFELQQG Ll-9 125 SKFKPLDELEQTLYEQWTLQHA Ll-10 126 QKFQPLDELEQTLYEQFMLQQA Ll-11 127 QNFKPMDELEDTLYKQFLFQHS Ll-12 128 YKFTPLDDLEQTLYEQWTLQHY Ll-13 129 QEYEPLDELDETLYNQWMFHQR Ll-14 130 SNFMPLDELEQTLYEQFMLQHQ Ll-15 131 QKYQPLDELDKTLYDQFMLQQG Ll-16 132 QKFQPLDELEETLYKQWTLQQR Ll-17 133 VKYKPLDELDEWLYHQFTLrfflQ Ll-18 134 QKT^LDELDEILYEQFMFQQS Ll-19 135 QTFQPLDDLEEYLYEQWTRRYH Ll-20 136 EDYMPLDALDAQLYEQFILLHG Ll-21 137 HTFQPLDELEETLYYQWLYDQL Ll-22 138 YKFNPMDELEQTLYEEFLFQHA AC6-L1 139 TN\TCPmELDATLYEHWILQHS Ll-Cl 140 QEDRKPLDELEQTLYEQWTLQQR L1-C2 141 TKFQPLDELDQTLYEQWTLQQR L1-C3 142 TNFQPLDELDQTLYEQWTLQQR KFNPLDELEETLYEQFTFQQ Изобретение также относится к полипептиду, способному связываться с Ang-2, содержащему аминокислотную последовательность формулы RPe3e4e5e6e7G (SEQ ID NO: 73) где е3 обозначает нейтральный полярный аминокислотный остаток; е4 обозначает кислый аминокислотный остаток; е5 обозначает нейтральный полярный или кислый аминокислотный остаток; е6 обозначает нейтральный гидрофобный аминокислотный остаток; е7 обозначает нейтральный гидрофобный аминокислотный остаток; и к его физиологически приемлемым солям. В предпочтительном варианте изобретения е3 обозначает Y или С. В другом предпочтительном варианте изобретения е4 обозначает D или Е. Ещё в одном предпочтительном варианте изобретения е6 обозначает I или М. Далее изобретение относится к полипептиду, способному связываться с Ang-2, содержащему аминокислотную последовательность формулы f f2 f3 f4RPf7 f8 f9 f10 f11Gf13 f14 f15 f16 f17 f18f19 f20 (SEQ ID NO: 74) где f1 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f2 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f3 обозначает нейтральный полярный или кислый аминокислотный остаток; f4 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f7 обозначает нейтральный полярный аминокислотный остаток; f8 обозначает кислый аминокислотный остаток; f9 обозначает нейтральный полярный или кислый аминокислотный остаток; f10 обозначает нейтральный гидрофобный аминокислотный остаток; f11 обозначает нейтральный гидрофобный аминокислотный остаток; f13 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f15 обозначает нейтральный полярный аминокислотный остаток; f16 обозначает нейтральный полярный аминокислотный остаток; f17 обозначает нейтральный полярный или кислый аминокислотный остаток; f18 обозначает нейтральный гидрофобный или основной аминокислотный остаток; f19 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и f20 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; - 7 - 008248 и его физиологически приемлемым солям. В предпочтительном варианте изобретения f1 обозначает S, А или G; f2 обозначает G, Q или Р; f3 обозначает Q, G или D; f4 обозначает L, М или Q; f7 обозначает С или Y; f8 обозначает Е или D; f9 обозначает Е, G или D; f10 обозначает I или М; f11 обозначает F или L; f13 обозначает С или W; f14 обозначает G или Р; f15 обозначает Т или N; f16 обозначает Q, Y или K; f17 обозначает N, D или Q; f18 обозначает L, V, W или R; f19 обозначает A, Q, Y или I и f20 обозначает L, A, G или V. В более предпочтительном варианте изобретение относится к полипептиду, содержащему по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 3 и SEQ ID NO: 143 - SEQ ID NO: 148 включительно, причём полипептид способен связываться с Ang-2, и к его физиологически приемлемым солям. SEQ ID NO: 3 и SEQ ID NO: 143 - SEQ ID NO: 148 суть следующие. Conl-l 143' AGGMRPYDGMLGWPNYDVQA Conl-2 144 QTTODPCMrTILGPVTWRRCI Con 1-3 145 APGQRPYDGMLGWE.TYQRTV Conl-4 146 SGQLPvPCEEIFGCGTQNLAL Conl-5 147 FGDKRPLECMFGGPIQLCPR Conl-6 148 GQDLRPCEDMFGCGTKDWYG Conl KRPCEEIFGGCTYQ Ещё в одном аспекте изобретение относится к полипептиду, способному связываться с Ang-2, содержащему аминокислотную последовательность формулы Cg2Gg4g5DPFTg10GCg13 (SEQ ID NO:75) где g2 обозначает кислый аминокислотный остаток; g4 обозначает нейтральный гидрофобный аминокислотный остаток; g5 обозначает Е, D или Q; g10 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; g13 обозначает кислый остаток; и к его физиологически приемлемым солям. В предпочтительном варианте изобретения g2 обозначает Е или D. В другом предпочтительном варианте изобретения g4 обозначает V или М. Ещё в одном варианте изобретения g10 обозначает F или Q. И еще в одном варианте изобретения g13 обозначает D или Е. Далее изобретение относится к полипептиду, способному связываться с Ang-2, содержащему аминокислотную последовательность формулы h1h2h3h4Ch6Gh8h9DPFTh14GCh17h18h19h20 1 (SEQ ID NO: 158) где h1 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; h2 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h3 обозначает кислый аминокислотный остаток; h4 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h6 обозначает кислый аминокислотный остаток; h8 обозначает нейтральный гидрофобный аминокислотный остаток; h9 обозначает Е, D или Q; h14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h17 обозначает кислый аминокислотный остаток; h18 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный - 8 - 008248 остаток; h19 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и h20 отсутствует или обозначает аминокислотный остаток; и к его физиологически приемлемым солям. В предпочтительном варианте изобретения h1 отсутствует или обозначает A, L, M, G, K или Н; h2 обозначает L, F или Q; h3 обозначает D или Е; h4 обозначает W или Y; h6 обозначает D или Е; h8 обозначает V или М; h14 обозначает F или Q; h17 обозначает D или Е; h18 обозначает М, Y, N или K; h19 обозначает L или Q и h20 отсутствует или обозначает М, Т, G, S, D, K или R. В более предпочтительном варианте изобретение относится к полипептиду, содержащему по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5 или SEQ ID NO: 149 - SEQ ID NO: 157 включительно, причём указанный полипептид способен связываться с Ang-2, и к его физиологически приемлемым солям. SEQ ID NO: 5 и SEQ ID NO: 149 - SEQ ID N0: 157 представлены ниже. Пептид SEQ ID NO Последовательность 12-9-1 149 GEEYCDGMEDPFTFGCDKQT 12-9-2 150 KLEYCDGMEDPFTQGCDNQS 12-9-3 151 LQEWGEGVEDPFTFGCEKQR 12-9-4 152 AQDYCEGMEDPFTFGCEMQK - 12-9-5 153 - - LmYCEGVQDPFTEGCENLI) 12-9-6 154 HQEYCEGMEDPFTFGCEYQG 12-9-7 155 MLDYCEGMDDPFTFGCDKQM 12-9-С2 156 LQDYCEGVEDPFTFGCENQR 12-9-С1 157 LQDYCEGVEDPFTFGCEKQR 12-9 FDYCEGVEDPFTFGCDNH В наиболее предпочтительном варианте изобретение относится к составу полипептида, имеющему формулу и его мультимерам, где F1 обозначает носитель; каждый независимо, выбирают из -(L')s-H1; -(Ь'^-Р'-СД-Р2; -(L'VP'-^rP'-CL^u-P3; и 1 2 3 4 где один или более из Р1, Р2, Р3 и Р4, каждый независимо, представляют собой полипептид, выбранный из группы, состоящей из: (a) аминокислотной последовательности WDPWT (SEQ ID NO: 65), причём указанный пептид имеет в длину 5-50 аминокислот; (b) аминокислотной последовательности WDPWTC (SEQ ID NO: 66); (c) аминокислотной последовательности Cz2WDPWT (SEQ ID NO: 67), где z2 обозначает кислый или нейтральный полярный аминокислотный остаток; (d) аминокислотной последовательности Cz2WDPWTC (SEQ ID NO: 68), где z2 обозначает кислый или нейтральный полярный аминокислотный остаток; (e) аминокислотной последовательности Pc2Dc4Lc6c7c8LY (SEQ ID NO: 71), где с2 обозначает нейтральный гидрофобный аминокислотный остаток; с4 обозначает A, D или Е; с6 обозначает кислый аминокислотный остаток; с7 обозначает аминокислотный остаток и с8 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; (f) аминокислотной последовательности RPe3e4e5e6e7G (SEQ ID NO: 73), где е3 обозначает нейтральный полярный аминокислотный остаток; е4 обозначает кислый аминокислотный остаток; е5 обозначает нейтральный полярный или кислый аминокислотный остаток; е6 обозначает нейтральный гидрофобный аминокислотный остаток и е7 обозначает нейтральный гидрофобный аминокислотный остаток; - 9 - 008248 (g) аминокислотной последовательности Cg2Cg4g5DPFTg10GCg13 (SEQ ID NO: 75), где g2 обозначает кислый аминокислотный остаток; g4 обозначает нейтральный гидрофобный аминокислотный остаток; g5 обозначает нейтральный полярный или кислый аминокислотный остаток; g10 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и g13 обозначает кислый остаток; (h) полипептида SEQ ID NO: 1; (i) полипептида SEQ ID NO: 2 и (j) полипептида SEQ ID NO: 7; где L1, L2, L3 и L4, каждый независимо, обозначают линкеры; и q, r, s, t, u и v, каждый независимо, обозначают 0 или 1, при условии, что по меньшей мере один из q и r обозначает 1; и к его физиологически приемлемым солям. Будет оценено, что изобретение, кроме того, относится к слитому полипептиду, содержащему по меньшей мере один пептид по данному описанию и носитель, причём слитый полипептид способен связываться с Ang-2, и к его физиологически приемлемым солям. В слитом полипептиде носителем является по меньшей мере один из ряда: Fc домен, полиэтиленгликоль, липид, группа холестерина, углевод, оли-госахарид. Другие подходящие носители, такие как альбумин и т.п., будут оценены специалистами в данной области техники и входят в объём изобретения. Специалист в данной области техники понимает, что в структуру специфически связывающегося агента могут быть встроены различные молекулы. Так, данную молекулу можно ввести, например, между участками пептида и носителя специфически связывающихся агентов или внутри участка самого пептида, при сохранении заданной активности специфически связывающегося агента. Можно легко встроить, например, такие молекулы, как Fc домен или его фрагмент, полиэтиленгликоль или другие родственные молекулы, такие как декстран, жирная кислота, липид, группа холестерина, низкомолекулярный углевод, пептид, цитотоксический агент, химиотерапевтический агент, обнаруживаемый фрагмент, такой как представленный в данном описании (включая флуоресцентные агенты, радиоактивные метки, такие как радиоизотопы), олигосахарид, олигонуклеотид, полинуклеотид, "интерференционная" (или другая) РНК, ферменты, гормоны и т.п. Другие молекулы, пригодные для внедрения таким образом, будут понятны специалисту в данной области и входят в объём данного изобретения. Сюда входит, например, инсерция заданной молекулы между двумя последовательными аминокислотами, необязательно соединёнными соответствующим линкером. В качестве примера приведена последовательность СОП4(С) "пеп-титела" M-Fc-GGGGGAQQEECEWDPWTCEHMLE (SEQ ID NO: 23) Cпециалист в данной области техники может легко встроить заданную молекулу, например, между двумя остатками глутамина ("QQ") для получения заданной структуры и/или функции при сохранении способности пептида связывать Ang-2. Так, эту последовательность можно модифицировать следующим образом: M-Fc-GGGGGAQ-[мoлeкyлa]-QEECEWDPWTCEHMLE Если требуется, можно добавить линкерные молекулы. Кроме того, понятно, что молекулу можно встроить в различные местоположения в молекуле, включая подходящие боковые цепи, между носителем и пептидной последовательностью, как показано ниже М-Fс-[молекула]-GGGGGАQQЕЕСЕWDРWTCEHMLE или в любом другом месте по желанию специалиста в данной области техники. Другие подходящие варианты изобретения будут понятны специалистам в данной области техники. Ещё в одном варианте изобретение относится к полинуклеотиду, кодирующему специфические связывающие агенты (включая, но без ограничения, пептиды и/или "пептитела") по изобретению по данному описанию. Специалист в данной области техники понимает, что если известна аминокислотная последовательность, то с помощью известных методов можно легко определить соответствующую(ие) нук-леотидную(ые) последовательность(и). См., например, Suzuki, D., An Introduction to Genetic Analysis, W.H. Freeman Pub. Co. (1986). Примеры нуклеотидных последовательностей, кодирующих пептиды по изобретению, представлены ниже. Специалист в данной области техники знает, что данную аминокислоту могут кодировать более одного кодона, и, следовательно, изобретение относится к любым нуклеотид-ным последовательностям, которые кодируют пептиды и/или "пептитела" по изобретению. - 10 - 008248 Пептид Seq Пептидная последовательность Примеры ДНК-последовательностей Con4-44 PIRQEECDWDPWTCEHMWEV ccgatccgtcaggaagaatgcga ctgggacccgtggacctgcgaac acatgtgggaagtt (SEQ ГО NO: 159) Соп4-40 TNIQEECEWDPWTCDHMPGK accaacatccaggaagaatgcga atgggacccgtggacctgcgacc acatgccgggtaaa (SEQ ID NO: 160) Соп4-4 WYEQDACEWDPWTCEBDVLAEV tggtacgaacaggacgcttgcga _ atgggacccgtggacctgcgaac acatggctgaagtt (SEQ ГО N0:161) Соп4-31 NRLQEVCEWDPWTCEHMENV aaccgtctgcaggaagtttgcgaa tgggacccgtggacctgcgaaca cateeaaaacett CSEO ГО No7l62) ~ Соп4-С5 AATQEECEWDPWTCEHMPRS gctgctacccaggaagaatgcga atgggacccgtggacctgcgaac acatgccgcgttcc (SEQ ID NO: 163) Соп4-42 LRHQEGCEWDPWTCEHMFDW ctgcgtcaccaggaaggttgcga atgggacccgtggacctgcgaac acatgttcgactgg (SEQ ID NO: 164) Соп4-35 VPRQKDCEWDPWTCErJMYVG ^ttccgcgtcagaaagactgcga itgggacccgtggacctgcgaac icatgtacgttggt (SEQ ID 40:165) Соп4-43 t t 5ISHEECEWDPWTCEHMQVG i ccatctcccacgaagaatgcgaa gggacccgtggacctgcgaaca ;atgcaggttggt (SEQ ID 40: 360) Соп4-49 WAAQEECEWDPWTCEHMGRM tgggctgeteaggaagaatgcga atgggatccgtggacttgcgaaca catgggtcgtatg(SEQID . NO: 166) Соп4-27 TWPQDKCEWDPWTCEHMGST acttggccgcaggacaaatgcga atgggatccgtggacttgcgaaca catgggttetact (SEQ ГО NO: 167) - 11 - 008248 Con4-4S GHSQEECGWBPWTCEHMGTS ggtcactcccaggaagaatgcgg ttgggacccgtggacctgcgaac acatgggtacgtcc (SEQ Ш NO: 168) Con4-46 QHWQEECEWDPWTCDHMPSK cagcactggcaggaagaatgcga atgggacccgtggacctgcgacc acatgccgtccaaa (SEQ ГО NO: 169) Con4-41 NVRQEKCEWDPWTCEHMPVR aacgttcgtcaggaaaaatgcgaa tgggacccgtggacctgcgaaca catgccggttcgt (SEQ ID NO: 170) Con4-36 KSGQVECNWDPWTCEHMPRN aaatccggtcaggttgaatgcaac tgggacccgtggacctgcgaaca catgccgcgtaac (SEQ ID NO: 171) Соп4-34 VKTQEHCDWDPWTCEHMREW gttaaaacccaggaacactgcga ctgggacccgtggacctgcgaac acatgcgtgaatgg (SEQ ID NO: 172) Соп4-28 AWGQEGCDWDPWTCEHMLPM gcttggggtcaggaaggttgcga ctgggacccgtggacctgcgaac acatgctgccgatg (SEQ ГО NO: 173) Соп4-39 PVNQEDCEWDPWTCEHMPPM ccggttaaccaggaagactgcga atgggaccc gtggacctgcgaac acatgccgccgatg (SEQ ID NO: 174) Соп4-25 RAPQEDCEWDPWTCAHMDIK cgtgctccgcaggaagactgcga atgggacccgtggacctgcgctc acatggacatcaaa (SEQ ID NO: 175) Соп4-50 iGQNMECEWDPWTCEEMFRY cacggtcagaacatggaatgcga atgggacccgtggacctgcgaac acatgttccgttac (SEQ ID NO: 176) scgcgtctgcaggaagaatgcgtt -tgggacccgtggacctgcgaaca ;atgccgctgcgt (SEQ Ш ЧО: 177) Соп4-38 3RLQEECVWDPWTCEHMPLR Con4-2S RTTQEKCEWDPWTCEHMESQ cgtaccacccaggaaaaatgcga atgggacccgtggacctgcgaac acatggaatcccag (SEQ ED NO: 178) Соп4-4Ч QTSQEDCVWDPWTCDHMVSS cagacctcccaggaagactgcgtt tgggacccgtggacctgcgacca catggtttcctcc (SEQ ED NO: 179) Соп4-20 QVIGRPCEWDPWTCEHLEGL caggttatcggtcgtccgtgcgaa tgggacccgtggacctgcgaaca cctggaaggtctg(SEQID NO: 180) Соп4-45 WAQQEECAWDPWTCDHMVGL tgggctcagcaggaagaatgcgc ttgggacccgtggacctgcgacc acatggttggtctg(SEQro NO: 181) Соп4-37 100 LPGQEDCEWDPWTCEHMVRS ctgccgggtcaggaagactgcga atgggacccgtggacctgcgaac acatggttcgttcc (SEQ ED NO: 182) Соп4-33 101 PMNQVECDWDPWTCEHMPRS ccgatgaaccaggttgaatgcga ctgggacccgtggacctgcgaac acatgccgcgttcc (SEQ ED NO: 183) АС2-Соп4 102 FGWSHGCEWDPWTCEHMGST ttcggttggtctcacggttgcgaat gggatccgtggacttgcgaacac atgggttctacc (SEQ ГО NO: 184) Соп4-32 103 KSTQDDCDWDPWTCEHMVGP aaatccacccaggacgactgcga ctgggacccgtggacctgcgaac acatggttggtccg (SEQ ID NO: 185) Соп4-17 104 GPRISTCQWDPWTCEHMDQL ggtccgcgtatctccacctgccag tgggacccgtggacctgcgaaca catggaccagctg (SEQ ID NO: 186) Соп4-8 105 STIGDMCEWDPWTCAHMQVD tccaccatcggtgacatgtgcgaa tgggacccgtggacctgcgctca satgcaggttgac (SEQ ID NO: 187) АС4- . Соп4 106 VbGGQGCEWDPWTCRLLQGW gttctgggtggtcagggttgcgaa -gggacccgtggacctgccgtctg ;tgcagggttgg (SEQ ID 40: 188) Соп4-1 107 VLGGQGCQWDPWTCSHLEDG i ^ttctgggtggtcagggttgccag gggacccgtggacctgctccca xtggaagacggt (SEQ ГО 40: 189) - 12 - 008248 Con4-Cl 108 TTIGSMCEWDPWTCAHMQGG accaccatcggttccatgtgcgaa tgggacccgtggacctgcgctca catgcagggtggt (SEQ Ю NO: 190) Con4-21 109 TKGKSVCQWDPWTCSHMQSG accaaaggtaaatccgtttgccag tgggacccgtggacctgctccca catgcagtccggt (SEQ ID NO: 191) : Con4-С2 TTIGSMCQWDPWTCAHMQGG accaccatcggttccatgtgccag tgggacccgtggacctgcgctca catgcagggtggt (SEQ ID NO: 192) Соп4-18 111 WVNEWCEWDPWTCNHWDTP tgggttaacgaagttgtttgcgaat gggacccgtggacctgcaaccac tgggacaccccg (SEQ ID NO: 193) Соп4-19 112 W QVGMCQWDP WTCKHMRLQ gttgttcaggttggtatgtgccagt gggacccgtggacctgcaaacac atgcgtctgcag (SEQ ID NO: 194) Соп4-16 113 A VGS QTCEWDPWTCAEDLVE V gctgttggttcccagacctgcgaat gggacccgtggacctgcgctcac ctggttgaagtt(SEQID NO: 195) Соп4-11 114 QGMKMFCEWDPWTCAHTVYR cagggtatgaaaatgttctgcgaat gggacccgtggacctgcgctcac atcgtttaccgt (SEQ Ш ЬЛГ\. 1 п <сч N\J. ±y <)j -------- Соп4-С4 115 TTIGSMCQWDPWTCEHMQGG accaccatcggttccatgtgccag tgggacccgtggacctgcgaaca catgcagggtggt (SEQ ID NO: 197) Соп4-23 116 TSQRVGCEWDPWTCQHLTYT acctcccagcgtgttggttgcgaat gggacccgtggacctgccagcac ctgacctacacc (SEQ ID NO: 198) Соп4-15 117 QWSWPPCEWDPWTCQTVWPS cagtggtcctggccgccgtgcga atgggacccgtggacctgccaga ccgtttggccgtcc (SEQ ID NO: 199) Срп4-9 118 GTSPSFCQWDPWTCSEMVQG ggtacctccccgtccttctgccagt gggacccgtggacctgctcccac atggttcagggt(SEQID NO: 200) TN8- • Соп4 3EECEWDPWTCEHM caggaagaatgcgaatgggaccc atggacttgcgaacacatg (SEQ ID NO: 201) - 13 - 008248 Ll-l 119 QNYETIJDELDATLYEHHFHYT cagaactacaaaccgctggacga actggacgctaccctgtac gaaca cttcatcttccactacacc (SEQ ED NO: 202) Ll-2 120 LNFTPLDELEQTLYEQWTLQQS ctgaacttcaccccgctggacgaa ctggaacagaccctgtacgaaca gtggaccctgcagcagtcc (SEQ ГО NO: 203) Ll-3 121 TKFNPIJDELEQTL^QWTLQHQ accaaattcaacccgctggacga actggaacagaccctgtacgaac agtggaccctgcagcaccag (SEQ ГО NO: 204) Ll-4 122 VKPICPLDALEQTLYEHWMFQQA gttaaattcaaaccgctggacgct ctggaacagaccctgtacgaaca ctggatgttccagcaggct (SEQ ID NO: 205) Ll-5 123 VKYKPLX)ELDErLYEQQTFQER gttaaatacaaaccgctggacgaa ctggacgaaatcctgtacgaacag cagaccttccaggaacgt (SEQ ED NO: 206) - - - - - Ll-7 124 TNFMPMDDLEQRLYEQFILQQG accaacttcatgccgatggacgac ctggaacagcgtctgtacgaaca gttcatcctgcagcagggt (SEQ ID NO: 207) Ll-9 125 SKFKPLDELEQTLYEQWTLQHA tccaaattcaaaccgctggacgaa ctggaacagaccctgtacgaaca gtggaccctgcagcacgct (SEQ ED NO: 208) Ll-10 126 QKEQPLDELEQTLYEQFMLQQA cagaaattccagccgctggacga actggaacagaccctgtacgaac agttcatgctgcagcaggct (SEQ Ш NO: 209) Ll-11 127 QNFKPMDELEDTLYKQFLFQHS cagaacttcaaaccgatggacga attggaagacaccctgtacaaaca gttcctgttccagcactcc (SEQ ED NO: 210) Ll-12 128 YKFTPLDDLEQTLYEQWTLQHV tacaaattcaccccgctggacgac ctggaacagaccctgtacgaaca gtggaccctgcagcacgtt (SEQ Ш NO: 211) Ll-13 129 3EYEPLDELDETLYNQWMFHQR caggaatacgaaccgctggacga actggacgaaaccctgtacaacc agtggatgttccaccagcgt (SEQ ID NO: 212) - 14 - 008248 Ll-14 130 SNFMPLDELEQTLYEQFMLQHQ tccaacttcatgccgctggacgaa ctggaacagaccctgtacgaaca gttcatgctgcagcaccag (SEQ ID NO: 213) Ll-15 131 QKYQPmELDKTLYDQFJSdbQQG cagaaataccagccgctggacga actggacaaaaccctgtacgatca gttcatgctgcagcagggt (SEQ ID NO: 214) Ll-16 132 QKFQPLDELEETLYKQWTLQQR cagaaattccagccgctggacga actggaagaaaccctgtacaaac agtggaccctgcagcagcgt (SEQ ID NO: 215) Ll-17 133 VKYKPLDELDEWLYHQFTLHHQ gttaaatacaaaccgctggacgaa ctggacgaatggctgtaccacca gttcaccctgcaccaccag (SEQ ID NO: 216) Ll-18 134 QKEMPLDELDEILYEQFMPQQS cagaaattcatgccgctggacgaa ctggacgaaatcctgtacgaacag ttcatgttccagcagtccc (SEQ Ю NO: 217) Ll-19 135 QTFQPШDLEEYLYEQWIRRYH cagaccttccagccgctggacga cctggaagaatacttgtacgaaca gtggatccgtcgttaccac (SEQ ID NO: 218) Ll-20 136 EDYMPLDALDAQLYEQFILLHG gaagactacatgccgctggacgc tctggacgctcagctgtacgaaca gttcatcctgctgcacggt (SEQ ID NO: 219) Ll-21 137 HTFQPLDELEETLYYQWLYDQL cac accttccagccgctggacga actggaagaaaccctgtactacca gtggctgtacgaccagctg (SEQ ID NO: 220) Ll-22 138 YEENPJVTOELEQTLYEEELFQHA tacaaattcaacccgatggacgaa ctggaac agaccctgtacgaaga attcctgttccagcacgct (SEQ ID NO: 221) AC6-L1 139 INYEPIX> ELDATLYEITWILQHS accaactacaaaccgctggacga actggacgctaccctgtacgaaca ctggatcctgcagcactcc (SEQ ID NO: 222) Ll-Cl 140 3KFKPLDELEQTLYEQWTLQQR cagaaattcaaaccgctggacga actggaacagaccctgtacgaac agtggaccctgcagcagcgt (SEQ ID NO: 223) L1-C2 141 TKFQPLDELDQTLYEQWTLQQR accaaattccagccgctggacga actggaccagaccctgtacgaac agtggaccctgcagcagcgt (SEOIDNO:224) - 15 - 008248 Ll-СЗ 142 TNFQPLDELDQTLYEQWTLQQR accaacttccagccgctggacga actggaccagaccctgtacgaac agtggaccctgcagcagcgt (SEQ ID NO: 225) EI^NPLDELEETLyEQFTFQQ aaattcaacccgctggacgagctg gaagagactctgtac gaacagttt acttttcaacag (SEQ ID NO: 226) Conl-1 143 AGGMRPYDGMLGWPNYDVQA gctggtggtatgcgtccgtacgac ggtatgctgggttggccgaactac gacgttcaggct (SEQ Ю NO: 227) Con 1-2 144 QTWDDPCMHILGPVTWRRCI cagacttgggacgatccgtgcatg cac attctgggtccggttacttggc gtcgttgcatc (SEQ ID NO: 228) Conl-3 145 APGQRPYDGlsdLGWPTYQRIV gctccgggtcagcgtccgtacga cggtatgctgggttggccgaccta ccagcgtatcgtt (SEQ ID NO: 229) Conl-4 146 SGQLRPCEEIFGCGTQNLAL tccggtcagctgcgtccgtgcgaa gaaatcttcggttgcggtacccag aacctggctctg (SEQ ID NO: 230) Conl-5 147 FGDKRPLECMFGGPIQLCPR ttcggtgacaaac gtccgctggaa tgcatgttcggtggtccgatccag ctgtgcccgcgt (SEQ ID NO: 231) Conl-6 148 ( 3QDLRPCEDMFGCGTKDWYG ggtcaggacctgcgtccgtgcga agacatgttcggttgcggtaccaa agactggtacggt(SEQID NO: 232) 12-9-1 149 ( 3FEYCDGMFJDPFTFGCDKQT ggtttcgaatactgcgacggtatg gaagacccgttcaccttcggttgc gacaaacagacc (SEQ ID N0:233) 12-9-2 150 KLEYCDGMEDPFTQGCDNQS aaactggaatactgcgacggtatg gaagaccc gttcacccagggttg cgacaaccagtcc (SEQ ID NO: 234) 12-9-3 151 LQEWCEGVEDPFTFGCEKQR ctgcaggaatggtgcgaaggtgtt gaagaccc gttcaccttcggttgc gaaaaacagcgt (SEQ Ш NO: 235) 12-9-4 152 AQDYCEGMEDPFTFGCEMQK gctcaggactactgcgaaggtatg gaagaccc gttcaccttcggttgc gaaatgcagaaa (SEQ ID NO: 236) 12-9-5 153 LLDYCEGVQDPFTFGCENLD ctgctggactactgcgaaggtgtt caggacccgttcaccttcggttgc gaaaacctggac (SEQ Ю NO: 237) 12-9-6 154 HQEYCEGMEDPFTFGCEYQG caccaggaatactgcgaaggtat ggaagacccgttcaccttcggttg cgaataccagggt (SEQ ID NO: 238) 12-9-7 155 MLDYCEGMDDPFTFGCDKQM atgctggactactgcgaaggtatg gacgacccgttcaccttcggttgc gacaaacagatg (SEQ ГО NO: 239) 12-9-C2 156 LQDYCEGVEDPFTFGCENQR ctgcaggactactgcgaaggtgtt gaagacccgttcaccttcggttgc gaaaaccagcgt (SEQ ID NO: 240) 12-9-C1 157 LQDYCbGVEDPFTFGCEKQR ctgcaggactactgcgaaggtgtt gaagacccgttcaccttcggttgc gaaaaacagcgt (SEQ ID NO: 241) 12-9 FDYCEGVEDPFTFGCDNH ttcgactactgcgaaggtgttgaa gacccgttcactttcggctgtgata accac (SEQ ID NO: 242) Ещё в одном варианте изобретение относится к экспрессирующим векторам, содержащим по меньшей мере один нуклеотид по изобретению. В другом варианте изобретение относится к клеткам-хозяевам, содержащим вектор экспрессии. Следует понимать, что клетки-хозяева представляют собой - 16 - 008248 предпочтительно прокариотные клетки (такие как клетки Е. coli) или эукариотные клетки. Изобретение также относится к фармацевтической композиции, содержащей эффективное количество состава полипептида по данному описанию в смеси с фармацевтически приемлемым носителем. Изобретение также относится к способу ингибирования нежелательного ангиогенеза у млекопитающих, заключающемуся во введении терапевтически эффективного количества полипептида или состава полипептида по данному описанию. Изобретение также относится к способу модулирования ангио-генеза у млекопитающих, заключающемуся во введении терапевтически эффективного количества полипептида или состава полипептида по данному описанию. Кроме того, изобретение относится к способу ингибирования роста опухоли, характеризующегося нежелательным ангиогенезом, у млекопитающих, заключающемуся во введении терапевтически эффективного количества полипептида или состава полипептида по данному описанию. Помимо этого, изобретение относится к способу лечения рака у млекопитающих, заключающемуся во введении терапевтически эффективного количества полипептида или состава полипептида по данному описанию и химиотерапевтического агента. В предпочтительном варианте изобретения химиотерапевтический агент представляет собой по меньшей мере один из 5-Fu, СРТ-11 и Taxotere (таксотер). Однако следует отдавать себе отчёт, что можно применять другие химиотерапев-тические агенты и другие методы терапии рака. Изобретение также относится к способу модулирования по меньшей мере одного из свойств: проницаемости сосудов или просачивания плазмы у млекопитающих, заключающемуся во введении эффективного количества полипептида или состава полипептида по данному описанию. Кроме того, изобретение относится к способу лечения по меньшей мере одного из следующих заболеваний (нарушений): глазного реваскулярного заболевания, ожирения, ангиоретикулёмы, гемангиомы, артериосклероза, воспалительного заболевания, воспалительных нарушений, атеросклероза, эндометриоза, опухолей, костного заболевания или псориаза у млекопитающих, заключающемуся во введении терапевтически эффективного количества полипептида или состава полипептида по данному описанию. Понятно, что специфически связывающиеся агенты по изобретению можно применять для лечения ряда заболеваний, ассоциированных с нарушенным или нежелательным ангиогенезом. Такие заболевания включают, но без ограничения, глазную реваскуляризацию, такую как ретинопатии (включая диабетическую ретинопатию и пятнистую дегенерацию, связанную со старением), псориаз, ангиоретикулёму, гемангиому, артериосклероз, воспалительное заболевание, такое как ревматоидное или ревматическое воспалительное заболевание, в особенности артрит (включая ревматоидный артрит), или другие хронические воспалительные заболевания, такие как хроническая астма, артериальный или посттрансплантационный атеросклероз, эндометриоз и опухолевые заболевания, такие, например, как так называемые солидные опухоли и "жидкие" (или гемопоэтические) опухоли (такие как лейкемии). Дополнительные заболевания, которые можно лечить введением специфически связывающихся агентов, будут очевидны для специалистов в данной области техники. Такие дополнительные заболевания включают, но без ограничения, ожирение, проницаемость сосудов, просачивание плазмы и костные нарушения, включая остеопороз. Таким образом, изобретение далее относится к способам лечения этих заболеваний, ассоциированных с нарушенным или нежелательным ангиогенезом. Другие варианты данного изобретения будут совершенно очевидны из прилагаемого описания. Краткое описание чертежей На фиг. 1 дан график изменения объёма опухоли (ось у) во времени (ось х) у мышей с опухолью линии А-431, пролеченных либо "пептителом" TN8-Con4-C по данному изобретению, либо фосфатно-солевым буферным раствором (PBS). Подробности описаны в примерах. На фиг. 2 изображён график зависимости концентрации "пептитела" (ось у) от времени после введения дозы (ось х) у мышей дикого типа, пролеченных дозой 50 мкг "пептитела" 2xCon4-C, L1-7-N или L1-21-N. Подробности описаны в примерах. На фиг. 3 дан график изменения объёма опухоли (ось у) во времени (ось х) у мышей с опухолью А-431, пролеченных либо "пептителом" TN8-Con4-C по данному изобретению, либо фосфатно-солевым буферным раствором (PBS), либо контрольным "пептителом". Подробности описаны в примерах. На фиг. 4 изображён график, представляющий in vitro рост культивированных клеток А-431, обработанных "пептителом" СОП4-С по настоящему изобретению, контрольным "пептителом" или необработанных. Подробности описаны в примерах. На фиг. 5 изображён график изменения объёма опухоли (ось у) во времени (ось х) в опухолевых клетках Сок> 205 у мышей, пролеченных "пептителом" СОП4-С, "пептителом" L1-7-N, "пептителом" L1-21-N или "пептителом" 2ХСОП4-С по данному изобретению, либо фосфатно-солевым буферным раствором (PBS), антителом против Ang-2 (Ab536) или Fc. Подробности описаны в примерах. На фиг. 6 дан график зависимости объёма опухоли (ось у) от времени (ось х) у мышей с Сок> 205 ксе-нотрансплантатом опухоли, пролеченных варьируемыми дозами "пептитела" 2ХСОП4-С по данному изобретению, или фосфатно-солевым буферным раствором (PBS), или Fc. Подробности описаны в примерах. На фиг. 7 дан график зависимости объёма опухоли (ось у) от времени (ось х) у мышей с Сок> 205 ксенотрансплантатом опухоли, пролеченных "пептителом" 2ХСОП4-С по данному изобретению или контрольными "пептителами". На фиг. 7 также изображён график - область, окрашенная CD31/общая пло - 17 - 008248 щадь опухоли для этих "пептител". Подробности описаны в примерах. На фиг. 8 изображён график зависимости объёма опухоли (ось у) от времени (ось х) у мышей с Со1о205 ксенотрансплантатом опухоли, пролеченных "пептителом" 2ХСОП4-С по данному изобретению или фосфатно-солевым буферным раствором (PBS) или контрольным "пептителом". Подробности описаны в примерах. Этот график показывает, что affra-Ang-2 "пептитела" способны ингибировать рост опухоли Со1 о205 независимо от того, когда начинают вводить дозу лекарства. На фиг. 9 дана сводка скоростей полной реакции (CR) у самок "голых" мышей, полученных при использовании антитела АЬ536 или "пептитела" 2ХСОП4-С как на модели ксенотрансплантата А431, так и на модели ксенотрансплантата Colo-205. Подробности описаны в примерах. На фиг. 10А дан график изменения объёма опухоли (ось у) во времени (ось х) у мышей с ксе-нотрансплантатом Colo-205 опухоли, пролеченных либо "пептителом" 2ХСОП4-С по данному изобретению, либо комбинацией 2ХСОП4-С и таксотера, либо фосфатно-солевым буферным раствором (PBS), либо PBS плюс таксотер. Подробности описаны в примерах. На фиг. 10В дан график изменения объёма опухоли (ось у) во времени (ось х) у мышей с ксе-нотрансплантатом Colo-205 опухоли, пролеченных либо "пептителом" 2ХСОП4-С по данному изобретению, либо комбинацией 2ХСОП4-С и 5-FU, либо фосфатно-солевым буферным раствором (PBS), либо PBS плюс 5-FU. Подробности описаны в примерах. На фиг. 11 А дан график уровня опухания лапы (AUC±SE) у индуцированной адъювантом модели артрита у крыс, пролеченных "пептителом" 2ХСОП4-С по данному изобретению, либо фосфатно-солевым буферным раствором (PBS), либо контрольным "пептителом", либо нормальным контролем или контролем на артрит. Подробности описаны в примерах. На фиг. 11 В дан график минеральной плотности костей лапы (BMD) у индуцированной адьювантом модели артрита у крыс, пролеченных "пептителом" 2ХСОП4-С по данному изобретению, либо фосфатно-солевым буферным раствором (PBS), либо контрольным "пептителом", либо нормальным контролем или контролем на артрит. Подробности описаны в примерах. На фиг. 11 С дан график изменения веса тела у индуцированной адьювантом модели артрита у крыс, пролеченных "пептителом" 2ХСОП4-С по данному изобретению, либо фосфатно-солевым буферным раствором (PBS), либо контрольным "пептителом", либо нормальным контролем или контролем на артрит. Подробности описаны в примерах. На фиг. 12 даны 2 графика, изображающих ингибирование индуцированного VEGF корнеального ангиогенеза у крыс. На первом графике изображено число кровеносных сосудов у крыс, обработанных альбумином бычьей сыворотки (BSA), VEGF плюс фосфатно-солевой буферный раствор (PBS) или VEGF плюс "пептитело" СОП4-С по изобретению. Второй график изображает площадь кровеносных сосудов (мм2) у крыс, получавших BSA, VEGF плюс фосфатно-солевой раствор (PBS) или "пептитело" СОП4-С по изобретению. Подробности описаны в примерах. На фиг. 13А, 13В и 13С изображены данные эпитопного картирования (O.D) для полноразмерного Ang-2 (hAng-2), присоединения по N-концу hAng-2 и по С-концу hAng-2, соответственно, для "пептител" TN8-Con4-C, L1-7-N и 12-9-3-С по изобретению, а также для контрольного "пептитела" Tie2-Fc, C2B8 или 5В12. Подробности описаны в примерах. На фиг. 14 изображает аффинность связывания (Кц) "пептитела" по изобретению, определяемую методом Sapidane KinExA. Подробности описаны в примерах. Подробное описание изобретения Оглавления разделов даны лишь в целях упорядочения материала, но никоим образом не для ограничения описываемого предмета изобретения. Для получения молекул рекомбинантных ДНК, белков и антител, а также для получения тканевой культуры и трансформации клеток можно применять стандартные методы. Ферментативные реакции и очистку, как правило, осуществляют в соответствии с рекомендациями производителя или обычно принятыми в данной области техники методами, такими, как описанные в Sambrook et al. (Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY [1989], или методами по данному описанию. Если не указано иначе, применяемые в данном описании номенклатура, а также лабораторные методики и методы аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, хорошо известны и применяются обычно в технике. Для химических синтезов, химических анализов, приготовления и доставки фармацевтических препаратов и лечения больных можно применять стандартные методы. Определения Ниже приводятся определения терминов, употребляемых по всему данному описанию (если не указано иначе в конкретных примерах). Термин "Ang-2" относится к полипептиду, представленному на фиг. 6 в патенте США 6166185 ("Tie-2 лиганд-2"), или к его фрагментам, а также к родственным полипептидам, которые включают ал-лельные варианты, "сплайсинговые" варианты, производные, варианты с заменами, делециями и/или ин-серциями, слитые пептиды и полипептиды и межвидовые гомологи. Полипептид Ang-2 может включать или не включать дополнительные концевые остатки, например лидерные последовательности, целевые - 18 - 008248 последовательности, метионин по аминоконцу, остатки метионина и лизина по аминоконцу и/или последовательности-метки или последовательности слитых белков, в зависимости от способа его получения. Термин "биологически активный" в применении к Ang-2 или к агенту, специфически связывающемуся с Ang-2, относится к полипептиду, имеющему по меньшей мере одну активность, характерную для Ang-2 или к агенту, специфически связывающемуся с Ang-2. Агент, специфически связывающийся с Ang-2, может иметь агонистическую, антагонистическую или нейтрализующую или блокирующую активность в отношении по меньшей мере одной биологической активности Ang-2. Термин "специфически связывающийся агент" относится к молекуле, предпочтительно белковой молекуле, которая специфически связывается с Ang-2 и его вариантами и производными по данному описанию. Специфически связывающийся агент может представлять собой белок, пептид, нуклеиновую кислоту, углевод, липид или низкомолекулярное соединение, которое связывается преимущественно с Ang-2. В предпочтительном варианте специфически связывающийся агент по данному изобретению представляет собой пептид или "пептитело", а также его фрагменты, варианты или производные, одни или в сочетании с другими аминокислотными последовательностями, получаемые известными методами. Такие методы включают, но без ограничения, ферментативное расщепление, химическое расщепление, пептидный синтез или методы рекомбинантной ДНК. Affra-Ang-2 специфически связывающиеся агенты по данному изобретению способны связываться с участками Ang-2, которые модулируют, например ингибируют или промотируют, биологическую активность Ang-2 и/или другие типы ассоциированной с Ang-2 активности. Термин "варианты" по данному описанию включает такие пептиды и полипептиды, в которых аминокислотные остатки встроены в природную (или, по меньшей мере, в известную) аминокислотную последовательность, делетированы из неё и/или заменены в ней на связывающийся агент. Варианты по изобретению включают слитые белки, описанные ниже. " Производные" включают такие связывающиеся агенты, которые были химически модифицированы каким-либо способом, отличным от вариантов инсерции, делеции или замены. " Специфически связывающийся с Ang-2" относится к способности специфически связывающегося агента (такого как "пептитело" или его пептидный участок) по данному изобретению распознавать и связывать зрелый, полноразмерный или неполноразмерный человеческий Ang-2 полипептид, или его орто-лог, так что его аффинность (определяемая, например, методами анализа Affinity ELISA или Biacore) или его нейтрализующая способность (определяемая, например, методом анализа Neutralization ELISA по данному описанию или аналогичными методами анализа) по меньшей мере в 10 раз выше, но, необязательно, в 50 раз выше, в 100, 250 или в 500 раз выше или даже по меньшей мере в 1000 раз выше, чем аффинность или нейтрализующая способность того же агента в отношении любого другого ангиопоэтина или другого пептида или полипептида, в котором пептидный участок "пептитела" сначала слит с человеческим Fc фрагментом для оценки с помощью такого метода анализа. Термин "эпитоп" относится к участку любой молекулы, способному распознаваться и связываться специфически связывающимся агентом, например "пептителом", по одному или более антиген-связывающих участков связывающегося агента. Эпитопы, как правило, состоят из химически активных поверхностных молекулярных группировок, таких, например, как боковые цепи аминокислот или углеводов, и имеют специфические признаки трёхмерной структуры, так же как специфические характеристики заряда. Эпитопы по данному описанию могут быть смежными и несмежными. Термин "ингибирующий и/или нейтрализующий эпитоп" представляет собой эпитоп, который, будучи связан специфически связывающимся агентом, таким как "пептитело", приводит к утрате (или, по меньшей мере, к снижению) биологической активности молекулы, клетки или организма, содержащих такой эпитоп, in vivo, in vitro или in situ. В контексте настоящего изобретения нейтрализующий эпитоп локализуется на биологически активной области Ang-2 или ассоциируется с этой областью. Термин же " активирующий эпитоп" представляет собой эпитоп, который, будучи связан специфически связывающимся агентом по изобретению, таким как антитело, вызывает в результате активацию или, по меньшей мере, сохранение биологически активной конформации Ang-2. Термин "фрагмент "пептитела" относится к пептиду или полипептиду, который содержит область, меньшую, чем полное интактное "пептитело". Термин "природный" в применении к биологическим материалам, таким как нуклеиновые кислоты, полипептиды, клетки-хозяева и т. п., относятся к тем из них, которые обнаружены в природе, а не модифицированы человеком. Термин "изолированный, выделенный" в применении к Ang-2 или к агенту, специфически связывающемуся с Ang-2, относится к соединению, свободному по меньшей мере от одного загрязняющего полипептида или соединения, в смеси с которым он обнаруживается в природе, и предпочтительно практически не содержит любые другие загрязняющие полипептиды, обнаруживаемые в организме млекопитающих, которые могли бы помешать его (Ang-2 или агента, специфически связывающегося с Ang-2) применению в терапевтических или диагностических целях. Термин "зрелый" в применении к Ang-2 "пептителу", или его фрагменту, или к любому другому белковому агенту, специфически связывающемуся с Ang-2, относится к пептиду или полипептиду, у которого отсутствует лидерная или сигнальная последовательность. Если связывающийся агент по изобре - 19 - 008248 тению экспрессирует, например, в прокариотной клетке-хозяине, "зрелый" пептид по изобретению может также включать дополнительные аминокислотные остатки (но по-прежнему не содержит лидерной последовательности), такие как концевой метионин или один или более остатков метионина и лизина. Пептид или полипептид, полученный таким образом, можно употреблять, удаляя или не удаляя эти дополнительные аминокислотные остатки. Термин "эффективное количество" и "терапевтически эффективное количество" в применении к агенту, специфически связывающемуся с Ang-2, относится к количеству специфически связывающегося агента, которое пригодно или необходимо для поддержания наблюдаемого изменения уровня одного или более типов биологической активности Ang-2. Изменение может представлять собой либо повышение, либо понижение уровня Ang-2 активности. Предпочтительно изменение представляет собой уменьшение Ang-2 активности. Термин "пептитело" относится к молекуле, содержащей Fc область (домен) антитела, связанный по меньшей мере с одним пептидом. Получение пептител в целом описано в международной заявке РСТ WO 00/24782, опубликованной 4 мая 2000 г. Термин "варианты" по данному описанию включает молекулы таких соединений, как пептиды или комбинации пептид-носитель, такие как "пептитела" по данному изобретению, в которых аминокислотные остатки встроены в аминокислотные последовательности таких молекул, делетированы из них и/или заменены на них. Варианты, содержащие одну или более встроенных аминокислот, включают слитые белки, описанные далее. "Производные" включают такие пептиды и/или комбинации пептид-носитель, такие как "пептитела", которые были химически модифицированы иным способом, нежели варианты с инсерцией, делецией или с заменой. Термин "фрагмент" относится к пептиду или к комбинации пептид-носитель, который содержит аминокислотную последовательность меньшей длины, нежели полноразмерная аминокислотная последовательность таких пептидов и/или таких комбинаций пептид-носитель. Такой фрагмент может получаться в результате усечения по аминоконцу, усечения по карбоксиконцу и/или внутренней делеции ос-татка(ов) аминокислотной последовательности пептида или комбинации пептид-носитель. Фрагменты могут получаться в результате альтернативного сплайсинга РНК или вследствие in vivo или in vitro про-теазной активности. Такие фрагменты можно также получить методами химического пептидного синтеза или модификацией полинуклеотида, кодирующего пептид, комбинацию пептид-носитель или Fc фрагмент и/или пептидный фрагмент "пептитела". Термин "Fc" относится к одному типу носителя по данному изобретению и содержит последовательность неантигенсвязывающего фрагмента антитела, образующегося в результате протеолитического расщепления целого антитела, в мономерной или мультимерной форме. Источником Fc по данному изобретению является предпочтительно полностью человеческий Fc и может быть любой из иммуноглобулинов, хотя предпочтительными являются IgG1 и IgG2. Однако также по данному описанию охватываются молекулы Fc частично человеческие или получаемые из нечеловеческих видов. Fc составлены из мономерных полипептидов, которые могут быть связаны в димерные или мультимерные формы кова-лентной связью (т.е. дисульфидными связями) и с помощью нековалентной ассоциации. Число межмолекулярных дисульфидных связей между мономерными субъединицами молекул нативного Fc составляет от 1 до 4 в зависимости от класса (например, IgG, IgA, IgE) или подкласса (например, IgG1, IgG2, IgG3, IgA1, IgA2). Одним примером нативного Fc является связанный дисульфидной связью димер, получающийся при расщеплении IgG под действием папаина [см. Ellison et al. (1982), Nucl. Acids Res. 10: 4071-9]. Термин "нативный Fc", применяемый в данном описании, является родовым по отношению к мономерным, димерным и мультимерным формам. Термин "Fc область (домен)" охватывает молекулы нативного Fc и Fc вариантов и последовательности по определению выше. Как в случае Fc вариантов и нативных Fc, термин "Fc область (домен) охватывает молекулы в мономерной и мультимерной форме, полученные либо при расщеплении целого антитела, либо другими способами. Термин "мультимер" в применении к Fc областям или молекулам, содержащим Fc области, относится к молекулам, имеющим две полипептидные цепи или более, ассоциированные за счёт ковалентных взаимодействий, нековалентных взаимодействий, или как за счёт ковалентных, так и за счёт нековалент-ных взаимодействий. Молекулы IgG, как правило, образуют димеры; IgM-пентамеры; IgD-димеры; и IgA-мономеры, димеры, тримеры и тетрамеры. Мультимеры могут образовываться за счёт использования последовательности и результирующей активности нативного Ig-источника Fc или за счёт дериватизации (по определению см. ниже) такого нативного Fc. Термин "димер" в применении к Fc областям или молекулам, содержащим Fc области, относится к молекулам, имеющим две полипептидные цепи, ассоциированные за счёт ковалентных или нековалентных связей. Термин "носитель" относится к молекуле, которая предотвращает расщепление и/или повышает период полужизни, снижает токсичность, уменьшает иммуногенности или повышает биологическую активность терапевтического белка. Примеры носителей включают Fc область, а также линейный полимер (например, полиэтиленгликоль (ПЭГ), полилизин, декстран и т.п.); разветвлённый полимер (см., например, патент США 4289872, принадлежащий Denkenwalter et al., выданный 15 сентября 1981 г.; патент - 20 - 008248 США 5229490, принадлежащий Tam, выданный 20 июля 1993 г.; международная заявка WO 93/21259 авторов Frechet et al., опубликованная 28 октября 1993 г.); липид, группа холестерина (например, стероид); углевод или олигосахарид; или любой природный или синтетический белок, полипептид или пептид, который связывается с рецептором-"утилизатором". Носители дополнительно описаны ниже. Термины "дериватизация" и "производное" или "дериватизированный" обозначают процессы и полученные в результате соединения соответственно, в которых (1) соединение содержит циклический фрагмент; например перекрёстное связывание между цистеинильными остатками в соединении; (2) соединение перекрёстно связано или содержит сайт перекрёстного связывания; например, соединение содержит цистеинильный остаток и, следовательно, образует димер за счёт перекрёстного связывания в культуре in vivo; (3) одна или более пептидильных связей замещаются непептидильной связью; (4) N- конец заменяется на -NRR1, NRC(O)R1, -NRC(O)ORl, -NRS(O)2R1, -NHC(O)NHR, сукцинимидную группу, или замещённый или незамещённый бензоилоксикарбонил-NH-, где R и R1 и заместители в цикле определены ниже; (5) С-конец на -C(O)R2 или -NR3R4, где определение R2, R3 и R4 дано ниже; и (6) соединения, в которых индивидуальные аминокислотные фрагменты модифицированы обработкой агентами, способными реагировать с выбранными боковыми цепями или концевыми остатками. Производные дополнительно описаны ниже. Термин "пептид" относится к молекулам, содержащим, примерно, 3-75 аминокислот, при этом предпочтительны молекулы, содержащие около 5-50 аминокислот, более предпочтительны молекулы, содержащие около 8-40, и наиболее предпочтительны молекулы, содержащие 10-25 аминокислот. Пептиды могут иметь природные или искусственные (т. е. не встречающиеся в природе) аминокислотные последовательности. Примеры пептидов могут получаться любыми методами, представленными в данном описании, такими как использование пептидной библиотеки (например, метод фагового дисплея), химический синтез, расщепление пептидов или метод рекомбинантной ДНК. Термин "фармакологически активное" обозначает, что найдено, что описанное таким образом вещество обладает активностью, которая влияет на медицинские показатели (например, кровяное давление, число гемо-цитов, уровень холестерина) или на болезненное состояние (например, рак, аутоиммунные расстройства и т. д.). Термины "пептид-антагонист, пептид антагонистического действия" или "ингибирующий пептид" относятся к пептиду, который блокирует биологическую активность, или каким-либо образом препятствует биологической активности ассоциированного представляющего интерес белка, или обладает биологической активностью, сравнимой с известным антагонистом или ингибитором ассоциированного представляющего интерес белка. Так, термин "пептид-антагонист Ang-2" охватывает пептиды, которые, как можно идентифицировать или установить, обладают антагонистическими в отношении Ang-2 признаками. Кроме того, в данное описание включены физиологически приемлемые соли соединений по изобретению. Под "физиологически приемлемыми солями" подразумеваются любые соли, известные или станут известными позже как фармацевтически приемлемые. Некоторые конкретные примеры: ацетат; трифторацетат; галогенводороды (галоидводороды), такие как хлоргидрат и бромгидрат; сульфат; цитрат; тартрат; гликолят; и оксалат, мезилат и фосфат. " Пептитела" Один аспект настоящего изобретения связан с созданием Ang-2 "пептител". Взаимодействие белка-лиганда со своим рецептором часто происходит на относительно большой поверхности раздела. Однако, как показано для соматотропина и его рецептора, только несколько ключевых (важных) остатков на поверхности раздела вносят вклад в энергию связывания. Clackson et al., Science 267; 383-6 (1995). Основной объём белка-лиганда просто выявляет связывающие эпитопы в правильной топологии или служит в качестве функций, не связанных со связыванием. Так, молекулы только на длине "пептида" (как правило, 2-40 аминокислот) могут связываться с рецепторным белком данного большого белка-лиганда. Такие пептиды могут имитировать биоактивность большого белка-лиганда ("пептидные агонисты", "пептиды-агонисты") или за счёт конкурентного связывания ингибировать биоактивность большого белка-лиганда ("пептидные антагонисты", "пептиды-антагонисты"). Фаговый дисплей является эффективным методом для идентификации таких пептидных агонистов и антагонистов. См, например, Scott et al., Science 249: 386 (1990); Devlin et al., Science 249: 404 (1990); патент США 5223409, выданный 29 июня 1993 г.; патент США 5733731, выданный 31 марта 1998 г.; патент США 5498530, выданный 12 марта 1996 г.; патент США 5432018, выданный 11 июля 1995 г.; патент США 5338665, выданный 16 августа 1994 г.; патент США 5922545, выданный 13 июля 1999 г.; международная заявка WO 96/40987, опубликованная 19 декабря 1996 г. и международная заявка WO 98/15833, опубликованная 16 апреля 1998 г. (каждый из этих источников вводится в данное описание в качестве ссылки). В пептидных библиотеках для выявления пептидов методом фагового дисплея случайные пептидные последовательности можно выявлять слиянием с оболочечными белками нитчатого фага. Если требуется, выявленные пептиды можно, используя метод аффинной хроматографии, элюировать против связанного с антителом внеклеточного домена рецептора. Удержанный фаг может быть обогащен за счёт последовательных циклов аффинной очистки и повторного культивирования. См., например, Cwirla et al., Science 276: 1696-9 (1997), где были идентифицированы два различных семейства. Пептидные последовательности могут подсказать, какие остатки можно безопасно заменить на аланин ("alanine-scanning") - 21 - 008248 или мутагенезом на уровне ДНК. Можно создать библиотеки, провести мутагенез, а затем скрининг для дополнительной оптимизации последовательностей лучших связывающихся агентов. Lowman, Ann. Rev. Biophys. Biomol. Struct. 26:401-24(1997). Для определения пептидов, которые имитируют связывающую активность больших белков-лигандов можно также использовать структурный анализ белок-белкового взаимодействия. В таких анализах структура может подсказать идентичность и относительную ориентацию существенно важных (критических), остатков большого белка-лиганда, из которого можно построить (создать) пептид. См., например, Takasaki et al., Nature Biotech 15: 1266-70 (1997). Эти аналитические методы можно также применять для изучения взаимодействия между рецепторным белком и пептидами, выбираемыми методом фагового дисплея, который может подсказать дальнейшую модификацию пептидов с целью повышения аффинности связывания. Другие методы конкурируют с методом фагового дисплея в исследовании пептидов. Пептидную библиотеку можно сливать (присоединять) по карбоксильному концу белка-репрессора lac и экспресси-ровать в Е. coli. Другой метод с использованием Е. coli делает возможным дисплей на внешней клеточной мембране за счёт слияния с липопротеидом, ассоциированным с пептидогликаном (PAL). В дальнейшем эти и родственные методы в данном описании объединены общим названием "Е. coli дисплей". В другом методе перед выделением рибосом прекращают трансляцию произвольной РНК, в результате получают библиотеку полипептидов со всё ещё присоединённой ассоциированной с ними РНК. В дальнейшем в данном описании этот и родственные методы употребляются под общим названием "рибосом-ный дисплей". Другие методы используют химическое связывание пептидов с РНК. См., например, Roberts and Szostak, Proc. Natl. Acad. Sci. USA 94: 12297-303 (1997). В дальнейшем в данном описании этот и родственные методы в целом называются "РНК-пептидный скрининг". Создаются получаемые химическим синтезом пептидные библиотеки, в которых пептиды иммобилизуют на стабильных материалах небиологического происхождения, таких как полиэтиленовые стержни или полимеры, проницаемые для растворителя. В других образованных химическим способом пептидных библиотеках используют фотолитографию для сканирования пептидов, иммобилизованных на предметных стеклах. В дальнейшем в данном описании эти и родственные методы имеют общее название " химический пептидный скрининг" (скрининг пептидов, полученных химическим путём). "Химический пептидный скрининг" может иметь преимущества, так как позволяет применять D-аминокислоты и другие неприродные аналоги, а также непептидные элементы. Обзор как биологических, так и химических методов дан в Wells and Lowman, Curr. Opin. Biotechnol., 3: 355-62 (1992). Умозрительно понятно, что можно обнаружить пептиды-миметики любого белка, используя метод фагового дисплея и другие приведённые выше методы. Эти методы были использованы для эпитопного картирования, для идентификации важных (критических) для белок-белкового взаимодействия аминокислот, и в качестве ключевых методов для открытия новых терапевтических агентов. См., например, Cortese et al., Curr. Opin. Biotech. 7: 616-21 (1996). В настоящее время пептидные библиотеки очень часто используются в иммунологических исследованиях, таких как эпитопное картирование. См. Kreeger, The Scientist 10(13): 19-20 (1996). Пептиды, определённые скринингом библиотек методом фагового дисплея, скорее рассматриваются как "ключевые" в создании терапевтических агентов, нежели как сами терапевтические агенты. Аналогично другим белкам и пептидам, они, по-видимому, быстро удаляются in vivo либо почечной фильтрацией, либо по механизмам клеточного клиренса в ретикулоэндотелиальной системе, либо путём проте-олитического распада [Francis, (см. выше)]. В результате в настоящее время в технике пептиды используют для подтверждения целевых препаратов (препаратов-мишеней), либо в качестве "скелета," основы для дизайна органических соединений, которые , которые невозможно так легко или так быстро обнаружить скринингом химических библиотек [Lowman, см. выше; Кау et al., (см. выше)]. Способ, позволяющий, используя такие пептиды, более просто получить терапевтические агенты против ангиогенеза, дал бы большие преимущества. Структура " пептител" В составах полипептида, получаемых в соответствии с данным изобретением, пептид может быть связан с носителем по N- или по С-концу пептида. Таким образом, молекулы носитель-пептид по данному изобретению можно описать в виде следующих пяти формул и в виде из мультимеров: (Xi),-F,-(X2)b (ФОРМУЛА I) Xi-Fi (ФОРМУЛА И) Xi-F2 (ФОРМУЛА III) FHLi)c-Pi (ФОРМУЛА IV) FKLi)c-PKL2)d-P2 (ФОРМУЛА V) где F1 обозначает носитель (предпочтительно, Fc область); X1 и Х2, каждый независимо, выбирают из -(L1)c-P1, -(L1)c-P1-(L2)d-P2, -(L1)c-P1-(L2)d-P2-(L3)e-P3 и - 22 - 008248 -(Ll)с-Pl-(L2)d-P2-(Lз)e-Pз-(L4)f-P4; Р1, Р2, Р3 и Р4, каждый независимо, обозначают последовательности или фармакологически активные пептиды по данному описанию; L1, L2, L3 и L4, каждый независимо, обозначают линкеры и "а", b", "с", "d", "e" и "f", каждый независимо, обозначают 0 и 1, при условии, что по меньшей мере один из "а" и "b" обозначает 1. Пептиды В настоящем изобретении рассматриваются пептиды, которые селективно связываются с Ang-2. По данному изобретению может использоваться любое количество таких пептидов. Метод фагового дисплея, в частности, применим для получения пептидов, используемых по данному изобретению, так как было показано, что аффинная селекция из библиотек случайных пептидов может быть использована для идентификации пептидных лигандов для любого сайта любого генного продукта. Dedman et al., J. Biol. Chem. 268: 23025-30 (1993). Пептиды по данному изобретению могут быть получены любыми методами, описанными в уровне техники. Используются однобуквенные аббревиатуры аминокислот. Термин "X" в любой последовательности (и по всему описанию, если в конкретном случае не указано иначе) означает, что может присутствовать любой из 20 природных аминокислотных остатков или любая из синтетических (неприродных) аминокислот (описанных далее в разделе "Варианты"). Любые из этих пептидов могут быть связаны парами, тандемом (т. е. последовательно), с линкером или без него, и примеры пептидов, связанные тандемом, представлены в таблице. Линкеры обозначены "L" и могут представлять собой любые из линкеров по данному описанию. Показаны тандемные повторы и линкеры, разделённые для ясности тире. Любой пептид, содержащий цистеинильный остаток, может быть перекрёстно связан с другим Cys-содержащим пептидом, каждый из которых или они оба могут быть связаны с носителем. Любой пептид, содержащий более одного Cys остатка, может также образовывать внутрипептидную дисульфидную связь. Любой из этих пептидов может быть дериватизирован по данному описанию. В производных, у которых карбоксильный конец может заканчиваться аминогруппой, эта завершающая аминогруппа представляет собой NH2. В производных, у которых аминокислотные остатки заменены фрагментами, отличными от аминокислотных остатков, замены обозначаются S, что обозначает любой фрагмент, описанный в Bhatnagar et al., J. Med. Chem. 39: 3814-9 (1996) и Cuthbertson et al., J. Med. Chem. 40: 2876-82 (1997), которые вводятся в данное описание в качестве ссылки. Если не указано иначе, все пептиды связаны пептидной связью. Носители В одном варианте настоящее изобретение включает по меньшей мере один пептид для присоединения по меньшей мере к одному носителю (F1, F2) по N-концу, С-концу или по боковым цепям одного из аминокислотных остатков пептида(ов). Можно также использовать разные носители; например, Fc на каждом конце или на одном конце Fc, а на другом конце или в боковой цепи - группу ПЭГ. Область (домен) Fc является одним предпочтительным носителем. Область Fc может быть слита с N- или С-концом пептидов или как с N-, так и с С-концом. Как отмечалось выше, Fc варианты являются подходящими носителями в объёме данного изобретения. Нативный Fc может быть значительно модифицирован для образования Fc варианте по данному изобретению при условии, что связывание с рецептором-утилизатором сохраняется. См., например, международные заявки WO 97/34631 и WO 96/32478. В таких Fc вариантах можно удалять один или более сайтов нативного Fc, которые придают структурные особенности или функциональную активность, не требующиеся в слитых молекулах по данному изобретению. Можно удалить эти сайты, например, заменой или делецией остатков, инсерцией остатков в сайт или отсекая участки, содержащие сайт. Встроенные или замещённые остатки могут также быть изменёнными аминокислотами, такими как пептидоми-метики или D-аминокислоты. Fc варианты могут быть желательны по ряду причин, некоторые из этих причин описаны ниже. Примеры Fc вариантов включают молекулы и последовательности, в которых 1. Сайты, участвующие в образовании дисульфидной связи, удаляют. Такое удаление позволяет избежать реакции с другими цистеинсодержащими белками, находящимися в клетке-хозяине, используемой для продуцирования молекул по изобретению. Для этой цели цистеинсодержащий сегмент на N-конце можно отсечь или можно удалить цистеиновые остатки или заменить их на остатки других аминокислот (например, аланил, серил). Даже если цистеиновые остатки удалены, одноцепочечные Fc области всё ещё образуют димерный Fc домен, который удерживается вместе за счёт нековалентных связей. 2. Нативный Fc домен модифицируют, чтобы сделать его более совместимым с выбранной клеткой-хозяином. Например, можно удалить РА последовательность близ N-конца типичного нативного Fc домена (нативной Fc области), который может распознаваться расщепляющим ферментом в Е. coli, таким как пролиниминопептидаза. Также можно добавить N-концевой метионильный остаток, особенно, когда молекулу экспрессируют методом рекомбинантной ДНК в бактериальной клетке, такой как Е. coli. 3. Часть N-конца нативного Fc удаляют с целью предотвратить N-концевую гетерогенность при экспрессии в выбранной клетке-хозяине. Для этой цели можно удалить любые из первых 20 аминокислотных остатков на N-конце, в частности, в положениях 1, 2, 3, 4 и 5. 4. Один или более сайтов гликозилирования удаляют. Остатки, которые, как правило, гликозили - 23 - 008248 руются (например, аспарагин), могут придать цитолитическую активность. Такие остатки можно удалить или заменить на негликозилированные остатки (например, аланин). 5. Сайты, участвующие во взаимодействии с комплементом, такие как Clq-связьтающий сайт, удаляют. Например, можно удалить или заменить ЕКК последовательность человеческого IgG1. Восстановление комплемента может быть неблагоприятным для молекул по данному изобретению и поэтому его можно избежать с помощью такого Fc варианта. 6. Сайты, которые влияют на связывание с Fc рецепторами, иными, нежели рецептор-утилизатор, удаляют. Нативный Fc может содержать сайты для взаимодействия с некоторыми лейкоцитами, которые не требуются для слитых молекул по данному изобретению и поэтому их можно удалить. 7. Сайт ADCC удаляют. Сайты ADCC известны в технике. См., например, Molec. Immunol. 29 (5): 633-9 (1992) относительно сайтов ADCC в IgG1. Эти сайты также не требуются для слитых молекул по данному изобретению и поэтому их можно удалить. 8. Если нативный Fc получают из нечеловеческого антитела, нативный Fc может быть "гуманизирован". Как правило, для гуманизации нативного Fc можно заменить выбранные остатки в нечеловеческом нативном Fc на остатки, которые обычно обнаруживают в человеческом нативном Fc. Методы гуманизации антител хорошо известны в технике. Альтернативным носителем может быть белок, полипептид, пептид, антитело, фрагмент антитела, малая молекула (например, пептидомиметик), способные связываться с рецептором-утилизатором. Например, можно в качестве носителя использовать полипептид, описанный в патенте США 5739277, выданный 14 апреля 1998 г. Presta et al. Пептиды можно выбирать методом фагового дисплея по признаку связывания с рецептором-"утилизатором" FcRn. Такие связывающие рецептор-"утилизатор" соединения также охватываются понятием "носитель" и входят в объём данного изобретения. Такие носители следует выбирать вследствие увеличенного периода полужизни (например, за счёт отсутствия последовательностей, распознаваемых протеазами) и пониженной иммуногенности (например, за счёт того, что предпочитаются неиммуногенные последовательности, описанные при гуманизации антител). Как отмечалось выше, полимерные носители также можно использовать в качестве F1 и F2. Различные значения для связанных химических фрагментов, пригодных в качестве носителей, доступны в настоящее время, см., например, международную заявку РСТ (Договор о патентной кооперации) WO 96/11953, озаглавленную "N-Terminally Chemically Modified Protein Compositions and Methods", которая ссылкой вводится в данное описание во всей полноте. В этой публикации РСТ, среди прочего, описывается селективное связывание водорастворимых полимеров с N-концом белков. Предпочтительным полимерным носителем является полиэтиленгликоль ПЭГ. Группа ПЭГ может быть любого подходящего молекулярного веса и может быть линейной или разветвлённой. Средний молекулярный вес ПЭГ предпочтительно составляет примерно от 2 до 100 килодальтон (кДа), более предпочтительно около 5-50 кДа, наиболее предпочтительно около 5-10 кДа. Группы ПЭГ, как правило, присоединяются к соединениям по изобретению с помощью ацилирования или восстановительного алкили-рования за счёт взаимодействия реакционноспособной группы в ПЭГ-фрагменте (например, альдегидной, амино, тиольной или сложноэфирной группы) с реакционноспособной группой соединения по изобретению (например, альдегидной, тиольной или сложноэфирной группы). Применяемая стратегия ПЭГирования синтетических пептидов состоит в присоединении за счёт конъюгирования в растворе пептида и фрагмента ПЭГ, причём каждый несёт особую функциональность, так что они являются реакционноспособными (реактивными) по отношению друг к другу. Пептиды можно легко получать обычным твердофазным синтезом, известным в технике. Пептиды "предварительно активируют" с помощью соответствующей функциональной группы в конкретном сайте. Предшественники очищают и полностью характеризуют перед реакцией с фрагментом ПЭГ. Лигирование пептида с ПЭГ обычно происходит в водной фазе и его легко контролировать (мониторинг) обращённо-фазовой аналитической ВЭЖХ. ПЭГированные пептиды можно легко очистить препаративной ВЭЖХ и охарактеризовать аналитической ВЭЖХ, аминокислотным анализом и лазерной десорбционной масс-спектрометрией. Полисахаридные полимеры представляют собой другой тип водорастворимых полимеров, которые можно использовать для модификации белков. Декстраны являются полисахаридными полимерами, состоящими из отдельных субъединиц глюкозы, преимущественно связанных связями а1-6. Сам декстран выпускается с различными интервалами молекулярной массы, легко доступен декстран с молекулярной массой 1-70 кДа. Декстран представляет собой водорастворимый полимер для применения по данному изобретению в качестве носителя как сам по себе, так и в комбинации с другим носителем (например, Fc). См., например, международные заявки WO 96/11953 и WO 96/05309. Сообщалось о применении дек-страна, конъюгированного с терапевтическими или диагностическими иммуноглобулинами: см., например, европейскую патентную заявку 0315456, которая вводится в данное описание в качестве ссылки. Предпочтительно, чтобы декстран, применяемый в качестве носителя по данному изобретению, имел молекулярную массу 1-20 кДа. Линкеры Любая "линкерная " группа является необязательной. Если эта группа присутствует, её химическое строение не важно, так как прежде всего она служит в качестве спейсера. Линкер предпочтительно со - 24 - 008248 стоит из аминокислот, связанных вместе пептидными связями. Так, в предпочтительных вариантах изобретения линкер состоит из 1-20 аминокислот, связанных пептидными связями, причём аминокислоты выбирают из 20 природных аминокислот. Одна или более из этих аминокислот могут быть гликозилиро-ванными, что хорошо понимают специалисты в данной области техники. В более предпочтительных вариантах изобретения аминокислоты от 1 до 20 выбирают из глицина, аланина, пролина, аспарагина, глу-тамина и лизина. Ещё более предпочтительно, когда линкер состоит из большинства стерически незатруднённых аминокислот, таких как глицин и аланин. Так, предпочтительными линкерами являются полиглицины (в частности, (Gly)5, (Gly)8), m^^G^-A^) и полиаланины. Комбинации Gly и Ala также являются предпочтительными в качестве линкера, называемого в данном описании К1 и имеющего аминокислотную последовательность, представленную далее в примерах. Возможны также непептидные линкеры. Например, алкильные линкеры, такие как -NH-(CH2)s-C(O)-, где можно использовать значение s=2-20. Эти алкильные линкеры могут дополнительно иметь в качестве заместителя любую пространственно незатруднённую группу, такую как низший алкил (например, низший ацил, галоген (например, Cl, Br), CN, NH2, фенил и т.д. Примером непептидного линкера является линкер ПЭГ с молекулярной массой, равной от 100 до 5000 кДа, предпочтительно 100500 кДа. Пептидные линкеры можно варьировать, образуя производные описанным выше способом. Варианты и производные В объём настоящего изобретения входят варианты и производные специфических связывающих агентов. Понятно, что конкретный специфический связывающий агент по настоящему изобретению может содержать один, два или все три типа вариантов. Варианты с инсерцией или заменой могут содержать природные аминокислоты, нетрадиционные (неприродные) аминокислоты или и те, и другие. В одном примере включены варианты с инсерцией, в которых один или более аминокислотных остатков, либо природных, либо нетрадиционных аминокислот дополняют аминокислотную последовательность пептида или "пептитела". Инсерции могут быть локализованы на любом конце белка или на обоих его концах или могут быть позиционированы во внутренних областях аминокислотной последовательности "пептитела". Инсерционные варианты с дополнительными остатками по любому концу или по обоим концам могут включать, например, слитые белки или белки, содержащие аминокислотные "хвосты" или метки. Инсерционные варианты включают пептиды и "пептитела", в которых один или более аминокислотных остатков добавлено к аминокислотной последовательности пептида или "пептитела" или ее фрагменту. Варианты продуктов по изобретению включают также зрелые пептиды и "пептитела", у которых удалены лидерная или сигнальная последовательности, и образующиеся в результате белки, содержащие дополнительные аминокислотные концевые остатки, причём аминокислоты могут быть природными и неприродными. Рассматриваются специфически связывающиеся агенты (такие как "пептитела") с дополнительным метионильным остатком в аминокислотном положении -1 (Met-1-"пептитело"), так же как специфически связывающиеся агенты с дополнительными остатками метионина и лизина в положениях -2 и -1 (Met-2-Lys-1-). Варианты, содержащие дополнительные остатки Met, Met-Lys, Lys, особенно пригодны для повышенной продукции рекомбинантных белков ив бактериальных клетках-хозяевах. Изобретение также охватывает варианты специфически связывающихся агентов, содержащие дополнительные аминокислотные остатки, образующиеся при использовании специфических систем экспрессии. Например, применение промышленных векторов, которые экспрессируют заданный полипептид как часть продукта слияния с глутатион^-трансферазой (GST), даёт заданный полипептид, содержащий дополнительный остаток глицина в аминокислотном положении -1, после отщепления компонента GST от заданного полипептида. Рассматриваются также варианты, получающиеся в результате экспрессии в другой векторной системе, включая варианты, в которых в аминокислотную последовательность, как правило, на карбокси- и/или аминоконце последовательности, встроен полигистидиновый "хвост". Инсерционные варианты также включают слитые белки, в которых амино- и/или карбоксиконцы пептида или "пептитела" слиты с другим полипептидом, его фрагментом или аминокислотами, которые, как правило, не распознаются как часть последовательности любого конкретного белка. Примерами таких слитых белков являются иммуногенные полипептиды, белки с продолжительным периодом полужизни в кровотоке, такие как константные области иммуноглобулина, маркерные белки, белки или полипептиды, которые облегчают очистку заданного пептида или "пептитела", и полипептидные последовательности, которые промотируют образование мультимерных белков (такие как лейциновая застёжка" молния", которая пригодна для образования/стабильности димера). Такой тип инсерционного варианта, как правило, содержит всю нативную молекулу или её значительную часть, связанную по N- или по С-концу с целым вторым полипептидом или с его частью. Например, слитые белки, как правило, используют лидерные последовательности от других видов, чтобы благоприятствовать рекомбинантной экспрессии белка в гетерологичном хозяине. Другой применяемый белок включает добавление иммунологически активной области, такой как эпитоп антитела, с целью облегчить очистку слитого белка. Включение сайта расщепления в место слияния или рядом с ним облегчает удаление " чужого" полипептида после очистки. Другие применимые слияния включают связывание функциональных областей, таких как активные сайты ферментов, области гликозилирования, сигналы - 25 - 008248 нацеливания на клетки или трансмембранные области. Существуют различные промышленные системы экспрессии слитых белков, которые могут использоваться по данному изобретению. Особенно пригодные системы включают, но без ограничения, систему глутатион^-трансферазы (GST) (Pharmacia), систему белка связывания мальтозы (NEB, Beverley, MA), систему FLAG (IBI, New Haven, CT) и систему 6xHis (Qiagen, Chatsworth, CA). Эти системы способны продуцировать рекомбинантные пептиды и/или "пептитела", несущие только небольшое количество дополнительных аминокислот, которые, по-видимому, незначительно влияют на активность пептида или "пептитела". Например, как система FLAG, так и система 6xHis добавляют только короткие последовательности, о которых известно, что они обе являются слабо антигенными и не оказывают вредного влияния на укладку полипептида в нативную конформацию. Другое слияние по N-концу, которое рассматривается как пригодное, представляет собой слияние дипептида Met-Lys по N-концевой области белка или пептидов. Такое слияние может дать благоприятное повышение экспрессии или активности белка. Другие слитые системы продуцируют полипептидные гибриды, в которых следует вырезать партнёр слияния от заданного пептида или "пептитела". В одном варианте связан с рекомбинантным "пепти-телом" с помощью пептидной последовательности, содержащей специфическую последовательность распознавания протеазой. Примерами соответствующих последовательностей являются последовательности, которые распознаются протеазой вируса гравировки табака (TEV) (Life Technologies, Gaithersburg, MD) или фактором Ха (New England Biolabs, Beverley, MA). Изобретение также охватывает слитые полипептиды, которые содержат целый пептид или целое " пептитело" по данному изобретению или их часть в комбинации с усечённым тканевым фактором (tTF). tTF представляет собой агент, нацеленный на сосуды, состоящий из усечённой формы человеческого белка, вызывающего коагуляцию, который ведёт себя как агент, коагулирующий кровь в кровеносных сосудах опухоли, как описано в патентах США 5877289; 6004555; 6132729; 6132730; 6156321 и в европейском патенте ЕР 0988056. Слияние tTF с anra-Ang-2 "пептителом" или пептидом или его фрагментом облегчает доставку anra-Ang-2 к клеткам-мишеням. В другом аспекте изобретение включает варианты с делецией, в которых удалены один или более аминокислотных остатков в пептиде или "пептителе". Делеции можно получать на одном или обоих концах пептитела или удалением одного или более остатков внутри аминокислотной последовательности "пептитела". Варианты с делецией обязательно включают все фрагменты пептида или "пептитела". Ещё в одном аспекте настоящее изобретение включает варианты пептидов или пептител по изобретению с аминокислотными заменами. Варианты с заменами включают такие пептиды и "пептитела", в которых удалены один или более аминокислотных остатков и заменены на одну или более альтернативных аминокислот, причём эти аминокислоты могут быть природными и неприродными. "Субституционные" (с заменами) варианты приводят к пептидам или "пептителам", которые "аналогичны" исходному пептиду или "пептителу", так как две молекулы содержат определённый процент идентичных аминокислот. Варианты с заменами включают замены 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30 аминокислот пептида или "пептитела", при этом число замен может составлять вплоть до 10% или более аминокислот пептида или "пепти-тела". В одном аспекте замены являются консервативными по природе, однако, изобретение также охватывает замены, которые являются неконсервативными, и также включает нетрадиционные аминокислоты. Идентичность и подобие родственных пептидов и "пептител" можно легко рассчитать известными методами. Такие методы включают, но без ограничения, методы, описанные в Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York (1988); Biocomputing: Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York (1993); Computer Analysis of Sequence Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey (1994); Sequence Analysis in Molecular Biology, von Heinje, O., Academic Press (1987); Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M. Stockton Press, New York (1991); и Carillo et al., SIAM J. Applied Math., 48:1073 (1988). Предпочтительные методы определения родства или идентичности в процентах двух пептидов или полипептидов, или полипептида и пептида предполагают дизайн, дающий наибольшее количество совпадений между проверяемыми последовательностями. Методы определения идентичности описаны в общедоступных компьютерных программах. Предпочтительные компьютерные программы для определения идентичности двух последовательностей включают, но без ограничения, пакет программ GCG, включающий GAP (Devereux et al., Nucl. Acid. Res., 12:387 (1984); Genetics Computer Group, University of Wisconsin, Madison, WI, BLASTP, BLASTN и FASTA (Altschul et al., J. Mol. Biol., 215:403-10 (1990)). Программа BLASTX является общедоступной из National Center for Biotechnology Information (NCBI) и других источников (BLAST Manual, Altschul et al. NCB/NLM/NIH Bethesda, MD 20894; Altschul et al., см. выше (1990)). Для определения идентичности можно применять также общеизвестный алгоритм Смита-Уотермана (Smith Waterman). Некоторые схемы совмещения (выравнивания) для сравнительного анализа первичной структуры двух аминокислотных последовательностей могут дать совмещение только короткого участка двух последовательностей, и эта малая область совмещения может иметь очень высокую степень идентичности последовательностей, даже если между двумя полноразмерными последовательностями нет заметного родства. Соответственно, в некоторых вариантах изобретения выбранный метод совмещения (программа - 26 - 008248 GAP) даёт в результате совмещение, которое перекрывает по меньшей мере десять процентов полной длины сравниваемого целевого полипептида, т. е. по меньшей мере 40 последовательных аминокислот, если сравниваются последовательности, содержащие по меньшей мере 400 аминокислот; 30 последовательных аминокислот, если сравниваются последовательности, содержащие по меньшей мере около 300400 аминокислот; по меньшей мере 20 последовательных аминокислот, если сравниваются последовательности, содержащие около 200-300 аминокислот; и по меньшей мере 10 последовательных аминокислот, если сравниваются последовательности, содержащие примерно 100-200 аминокислот. Например, с использованием компьютерного алгоритма GAP (Genetics Computer Group, University of Wisconsin, Madison, WI) два полипептида, для которых следует определить процент идентичности последовательностей, выравниваются с оптимальным совмещением их соответствующих аминокислот ("общее перекрывание", как определено с помощью алгоритма). В некоторых вариантах изобретения в сочетании с алгоритмом применяют штрафную функцию за раскрытие щели (гэпа) (которое обычно рассчитывается как средняя диагональ 3Х; "средняя диагональ" обозначает среднее значение диагонали применяемой матрицы сравнения; "диагональ" означает количественный показатель или число, отнесённое к каждой идеальной паре аминокислот при использовании конкретной матрице сравнения) и штрафную функцию за расширение щели (гэпа) (значение которого обычно составляет 1/10 от величины штрафной функции за раскрытие щели), а также матрицу сравнения, такую как РАМ 250 или BLOSUM 62. В некоторых вариантах изобретения стандартную матрицу сравнения (см. Dayhoff et al., Atlas of Protein Sequence and Structure, 5(3) (1978) относительно матрицы сравнения РАМ 250; Henicoff et al., Proc. Natl. Acad. Sci. USA, 89:10915-10919 (1992) относительно матрицы сравнения BLOSUM 62) также применяют с помощью алгоритма. В некоторых вариантах изобретения параметры полипептидной последовательности сравнения включают следующие: Алгоритм: Needleman et al., J. Mol. Biol., 48:443-453 (1970); Матрица сравнения: BLOSUM 62 из Henikoff et al., см. выше (1992); Штрафная функция за щель: 12; Штрафная функция за протяжённость щели: 4; Порог подобия: 0. Программу GAP можно применять с приведёнными выше параметрами. В некоторых вариантах изобретения вышеуказанные параметры представляют собой параметры по умолчанию для полипептидов сравнения (наряду с отсутствием " штрафная функция" за концевые щели) при использовании алгоритма GAP. В некоторых вариантах изобретения параметры последовательности сравнения полинуклеотидной молекулы (в противоположность аминокислотной последовательности) включают следующее: Алгоритм: Needleman et al., см. выше (1970); Матрица сравнения: совпадения (пары)=+10, несовпадение=0; Штрафная функция за щель: 50; Штрафная функция за протяжённость щели: 3. Программа GAP может также применяться с приведёнными выше параметрами. Вышеуказанные параметры являются параметрами по умолчанию для полинуклеотидньгх молекул сравнения. Можно применять другие примеры алгоритмов, штрафной функции за раскрытие щелей, штрафной функции за расширение щелей, матриц сравнения, порогов подобия и т. д., включая значения этих показателей, представленные в Program Manual, Wisconsin Package, Version 9, September, 1997. Практический выбор, который следует сделать, очевиден для специалиста в данной области техники и зависит от конкретного сравнения, которое следует сделать, такого как ДНК-с-ДНК, белок-с-белком, белок-с-ДНК; и, кроме того, проводится ли сравнение между данными парами последовательностей (в этом случае, как правило, предпочтительными являются GAP или BestFit) или между одной последовательностью и большой базой данных последовательностей (в этом случае предпочтительными являются FASTA или BLASTA). В данном описании придерживаются общепринятых правил при употреблении двадцати традиционных аминокислот и их сокращений. См. монографию Immunology-A Synthesis (2nd Edition, E.S. Golub and D.R. Gren, Eds., Sinauer Associates, Sunderland, Mass. (1991), которая вводится в данное описание ссылкой для любых целей. Аминокислоты могут иметь либо L, либо D стереохимию (за исключением Gly, который не является ни L, ни D), а полипептиды и составы полипептидов по настоящему изобретению могут содержать комбинацию стереоизомеров. Однако предпочтительными являются L-стереоизомеры. Изобретение также охватывает молекулы с обратной последовательностью, в которых последовательность аминокислот от амино-конца до карбокси-конца является обратной. Например, если молекула имеет нормальную последовательность Х1-Х2-Х3, то молекула с обратной последовательностью будет Х3-X2-X1. Изобретение также охватывает "ретро-обратные" молекулы, в которых, как указано выше, последовательность аминокислот от амино-конца до карбокси-конца является обратной и остатки, которые обычно являются "L" энантиомерами, находятся в "D" стереоизомерной форме. Стереоизомеры (например, D-аминокислоты) двадцати традиционных аминокислот, неприродные аминокислоты, такие как а-, а-дизамещённые аминокислоты, N-алкиламинокислоты, молочная кислота - 27 - 008248 и другие нетрадиционные аминокислоты могут также быть подходящими компонентами для полипептидов по данному изобретению. Примеры нетрадиционных аминокислот включают, без ограничения, ами-ноадипиновую кислоту, бета-аланин, бета-аминопропионовую кислоту, аминомасляную кислоту, пипе-ридиновую кислоту, аминокапроновую кислоту, аминогептановую кислоту, аминоизомасляную кислоту, аминопимелиновую кислоту, диаминомасляную кислоту, десмозин, диаминопимелиновая кислота, диа-минопропионовая кислота, N-этилглицин, N-этиласпарагин, гидроксилизин, алло-гидроксилизин, гидро-ксипролин, изодесмозин, алло-изолейцин, N-метилглицин, саркозин, N-метилизолейцин, N-метилвалин, норвалин, норлейцин, оритин, 4-гидроксипролин, у-карбоксиглутамат, e-N,N,N-триметиллизин, e-N-аце-тиллизин, О-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, o-N-метиларгинин и другие аналогичные аминокислоты и аминооксикислоты (например, 4-гидроксипролин). Аналогично, если не указано иначе, левый конец однонитевых полинуклеотидных последовательностей обозначается как 5' конец; направление двухнитевых полинуклеотидных последовательностей влево называется 5' направление. Направление от 5' конца к 3' концу образующихся РНК транскриптов называется направление транскрипции; участки последовательности на нити ДНК, имеющие ту же последовательность, что и РНК, и которые расположены 5' к 5' концу РНК транскрипта, называются "последовательности в обратном направлении, upstream"; участки последовательности на нити ДНК, имеющие ту же последовательность, что и РНК, и которые расположены 3' к 3' концу РНК транскрипта, называются "последовательности в прямом направлении, downstream". Понятно, что аминокислотные остатки можно разделить на классы, исходя из свойств общих боковых цепей: 1. Нейтральные гидрофобные: Аланин (Ala; А), Валин (Val; V), Лейцин (Leu; L), Изолейцин (Не; I), Пролин (Pro; Р), Триптофан (Trp; W), Фенилаланин (Phe; F) и Метионин (Met, M). 2. Нейтральные полярные: Глицин (Gly; G); Серин (Ser; S), Треонин (Thr; T), Тирозин (ТУГ; Y), Цис-теин (Cys; С), Глутамин (Glu; Q), Аспарагин (Asn; N) и Норлейцин. 3. Кислые: Аспарагиновая кислота (Asp; D), Глутаминовая кислота (Glu; E). 4) Основные: Лизин (Lys; K), Аргинин (Arg; R), Гистидин (His; H). См. Lewin, В., Genes V., Oxford University Press (1994), p.11. Консервативные аминокислотные замены могут включать нетрадиционные аминокислотные остатки, которые, как правило, встраиваются скорее химическим пептидным синтезом, нежели синтезом в биологических системах. Эти замены включают, без ограничения, пептидомиметики и другие обратные или инверсные формы аминокислотных фрагментов. Неконсервативные замены могут включать замену представителя одного из этих классов на представителя другого класса. При осуществлении этих замен (изменений), согласно некоторым вариантам изобретения, можно учитывать гидропатический индекс. Каждой аминокислоте присвоен гидропатический индекс, исходя из показателей её гидрофобности и заряда. Эти индексы суть следующие: изолейцин (+4.5); валин (+4.2); лейцин (+3.8); фенилаланин (+2.8); цистеин/цистин (+2.5); метионин (+1.9); аланин (+1.8); глицин (-0.4); треонин (-0.7); серин (-0.8); триптофан (-0.9); тирозин (-1.3); пролин (-1.6); гистидин (-3.2); глутамат (-3.5); глутамин (-3.5); аспартат (-3.5); аспарагин (-3.5); лизин (-3.9) и аргинин (-4.5). Из уровня техники понятно, как важен гидропатический индекс аминокислоты для приобретения белком интерактивной биологической функции. Kyte et al., J. Mol. Biol., 157: 105-131 (1982). Известно, что некоторые аминокислоты можно заменить на другие аминокислоты, имеющие сходный гидропатический индекс или показатель и всё ещё сохраняет аналогичную биологическую активность. При изменениях на основе гидропатического индекса некоторые варианты изобретения включают замены аминокислот, гидропатические индексы которых находятся в пределах ±2. Некоторые варианты изобретения включают замены аминокислот, гидропатические индексы которых находятся в пределах ±1, а некоторые варианты изобретения включают замены аминокислот, гидропатические индексы которых находятся в пределах ±0.5. Также из уровня техники понятно, что замену подобной аминокислоты можно эффективно провести с учётом гидрофильности, когда полученное таким образом биологически функциональное "пептите-ло" или полученный таким образом биологически функциональный пептид предполагается использовать в иммунологических вариантах изобретения, как в настоящем случае. В некоторых вариантах изобретения наибольшая локальная средняя гидрофильность белка, обусловленная гидрофильностью его смежных аминокислот, коррелируется с его иммуногенностью, т. е. с биологическим свойством белка. Следующие значения гидрофильности относятся к этим аминокислотным остаткам; аргинин (+3.0); лизин (+3.0); аспартат (+3.0 ±1); глутамат (+3.0 ±1); серин (+0.3); аспарагин (+0.2); глутамин (+0.2); глицин (0); треонин (- 0.4); пролин (-0.5 ±1); аланин (-0.5); гистидин (-0.5); цистеин (-1.0); метионин (-1.3); валин (-1.5); лейцин (-1.8); изолейцин (-1.8); тирозин (-2.3); фенилаланин (-2.5) и триптофан (-3.4). При изменениях на основе сходных значений гидрофильности некоторые варианты изобретения включают замены аминокислот, величины гидрофильности которых находятся в пределах ±2. Некоторые варианты изобретения включают замены аминокислот, величины гидрофильности которых находятся в пределах ±1, а некоторые варианты изобретения включают замены аминокислот, величины гидрофильности которых - 28 - 008248 находятся в пределах ±0.5. Можно также идентифицировать эпитопы первичных аминокислотных последовательностей исходя из гидрофильности. Эти области также называются "эпитопные коровые области". Примеры аминокислотных замен представлены ниже, в табл. 2. Таблица 2 Аминокислотные замены Исходный остаток Примеры замен Предпочтительные замены Ala Val, Leu, He Val Arg Lys, Gin, Asn Lys Asn Gin, Glu, Asp Gin Asp Glu, Glu, Asp Glu Cys Ser, Ala Ser Gin Asn, Glu, Asp Asn Glu Asp, Glu, Asp Asp Gly Pro, Ala Ala His Asn, Gin, ys, Arg Arg Leu, Val, Met, Ala, Phe, Норлейцин Leu Leu Норлейцин, He, Val, Met, Ala, Phe lie Lys Arg, 1,4-Диамино-масляная кислота, Gin, Asn Arg Met Leu, Phe, He Leu Phe Leu, Val, He, Ala, Tyr Leu Pro Ala Gly Ser Thr, Ala, Cys Thr Thr Ser Ser Trp Tyr, Phe Tyr Tyr Trp, Phe, Thr, Ser Phe Val He, Met, Leu, Phe, Ala, Норлейцин Leu Опытные специалисты в данной области техники способны определить подходящие варианты полипептида, представленные в данном описании, используя широко известные методы. В некоторых вариантах изобретения специалист в данной области техники может идентифицировать подходящие области молекулы, которые можно изменить, не нарушая активность, нацеливаясь на участки, которые, как полагают, не являются важными для активности. В некоторых вариантах изобретения можно идентифицировать остатки и фрагменты молекул, являющиеся консервативными в аналогичных пептидах или полипептидах. В некоторых вариантах изобретения даже области, которые могут быть важны для биологической активности или структуры, можно подвергнуть консервативным аминокислотным заменам, не нарушая биологическую активность или не оказывая вредного влияния на структуру полипептида. Кроме того, специалист в данной области техники может изучить работы по теме структура-функция, в которых идентифицируют остатки в сходных пептидах, которые важны для активности или структуры. Сравнение позволяет прогнозировать важность аминокислотных остатков в белке, которые соответствуют аминокислотным остаткам, важным для активности или структуры в аналогичных белках. Специалисты в данной области техники могут выбрать химически аналогичные замены на такие прогнозированные важные аминокислотные остатки. Специалисты в данной области техники могут также проанализировать трёхмерную структуру и первичную структуру аминокислотной последовательности в сравнении с такой структурой в аналогичных полипептидах. С учётом такой информации специалист в данной области техники может прогнозировать совмещение аминокислотных остатков антитела относительно его трёхмерной структуры. В некоторых вариантах изобретения специалист в данной области техники может предпочесть не делать радикальных изменений с аминокислотными остатками, которые, как прогнозировалось, расположены на - 29 - 008248 поверхности белка, так как такие остатки могут участвовать в важных взаимодействиях с другими молекулами. Кроме того, специалист в данной области техники может создать пробные варианты, содержащие единственную аминокислотную замену при каждом заданном аминокислотном остатке. Затем можно провести скрининг вариантов, используя методы анализа активности, известные специалистам в данной области техники. Такие данные можно использовать для сбора информации о подходящих вариантах. Например, если обнаруживается, что изменение в конкретном аминокислотном остатке приводит к нарушенной, нежелательно пониженной или неподходящей активности, вариантов с такими изменениями можно избежать. Другими словами, с учётом информации, собранной в таких рутинных экспериментах, специалист в данной области техники может легко определить аминокислоты, дальнейших замен которых, одних или в комбинации с другими мутациями, следует избегать. Ряд научных публикаций посвящен прогнозированию вторичной структуры. См. Moult J., Curr. Op. in Biotech., 7(4):422-427 (1996), Chou et al., Biochemistry, 13(2):222-245 (1974); Chou et al., Biochemistry, 113(2):211-222 (1974); Chou et al., Adv. Enzymol. Relat. Areas Mol. Biol., 47:45-148 (1978); Chou et al., Ann. Rev. Biochem., 47:251-276 и Chou et al., Biophys. J., 26:367-384 (1979). Кроме того, в настоящее время компьютерные программы могут помочь при прогнозировании вторичной структуры. Один метод прогнозирования вторичной структуры основан на моделировании по гомологии. Например, два полипептида или белка, последовательности которых идентичны более чем на 30%, или подобие которых выше 40%, часто имеют подобную структурную топологию. Увеличение за последнее время структурной базы данных белков (PDB) обеспечило повышенную предсказуемость вторичной структуры, включая потенциальное число складок в структуре полипептида или белка. См. Holm et al., Nucl. Acid. Res., 27(1):244-247 (1999). Было высказано предположение (Brenner et al., Curr. Op. Struct. Biol., 7(3):369-376 (1997)), что в данном полипептиде или белке существует ограниченное число складок и что, если решено критическое число структур, точность прогнозирования структур резко возрастает. Дополнительные методы прогнозирования вторичной структуры включают "вытягивание нити" (threading) (Jones, D., Curr. Op. Struct. Biol., 7(3):377-87 (1997); Sippl et al., Structure, 4(1):15-19 (1996)); "профильный анализ" (Bowie et al., Science, 253:164-170 (1991); Gribskov et al., Meth. Enzym., 183:146-159 (1990); Gribskov et al., Proc. Nat. Acad. Sci., 84(13):4355-4358 (1987)), и "эволюционное связывание" (Holm, см. выше (1999), и Brenner, см. выше (1997)). В некоторых вариантах изобретения варианты "пептитела" включают варианты гликозилирования, в которых один или более сайтов гликозилирования, таких как N-связанный сайт гликозилирования, добавлен к "пептителу". N-Связанный сайт гликозилирования характеризуется последовательностью: Asn-X-Ser или Asn-X-Thr, в которой аминокислотный остаток, обозначенный как X, может быть любым аминокислотным остатком, кроме пролина. Замена или введение (добавление) аминокислотных остатков с целью создания этой последовательности даёт потенциально новый сайт для введения N-связанной углеводной цепи. Или же замены, которые элиминируют эту последовательность, удаляют имеющуюся N-связанную углеводную цепь. Также охватывается перегруппировка N-связанных углеводных цепей, когда один или более N-связанных сайтов гликозилирования (как правило, тех, которые являются природными) элиминируются, а один или более новых N-связанных сайтов создаются. Изобретение также охватывает "производные", которые включают "пептитела", несущие модификации, отличные от инсерций, делеций или замен аминокислотных остатков или в дополнение к этим инсерциям, делециям или заменам. Предпочтительно, модификации являются ковалентными по природе и включают, например, химическую связь с полимерами, липидами, другими органическими и неорганическими фрагментами. Производные по изобретению можно получать с целью увеличения периода полужизни "пептитела" в кровотоке или для повышения целевой способности "пептитела" по отношению к заданным клеткам, тканям или органам. Примеры производных включают фрагменты, в которых сделаны одна или более из следующих модификаций: одна или более пептидильных [-C(O)NR-] связей (связей с пептидильным фрагментом) заменены на непептидильную связь, такую как -СН2-карбаматную связь (связь с карбаматной группой) [-CH2-O(CO)NR-]; фосфонатную связь; -СН2-сульфонамидную [-CH2-S(O)2NR-] связь; уреиленовую (мочевина) [-NHC(O)NH-] связь; -связь -СН2-вторичный амин; или связь с алкилированной пептидильной группой [-C(O)NR6-, где R6 обозначает низший алкил]; пептиды, в которых N-конец превращен в -NRR1 группу; в -NRC(O)R группу; в -NRC(O)OR группу; в -NRS(O)2R группу; в -NHC(O)NHR группу, где R и R1 обозначают водород или низший алкил, при условии, что R и R1, оба вместе не обозначают водород; в сукцинимидную группу; в группу бензилокси-карбонил-NH-(CBZ-NH-); или в группу бензилоксикарбонил-NH-, содержащую от 1 до 3 заместителей в фенильном кольце, выбираемых из группы, состоящей из низшего алкила, низшего алкокси, хлор и бром; и пептиды, в которых свободный С-конец дериватизирован в -C(O)R2, где R2 выбирают из группы, состоящей из низшей алкоксигруппы и -NR3R4, где R3 и R4 независимо выбираются из группы, состоящей из водорода и низшего алкила. Под "низшей" понимается группа, содержащая 1-6 углеродных атомов. - 30 - 008248 Кроме того, модификации отдельных аминокислот можно вводить в полипептиды или составы полипептидов по изобретению с помощью реакций нацеленных аминокислотных остатков пептида с органическим дериватизирующим агентом, способным реагировать с выбранными боковыми цепями концевых остатков. Ниже следуют примеры. Лизинильные аминокислотные остатки могут реагировать с ангидридами янтарной или других ди-карбоновых кислот. Дериватизация этими агентами влияет на реверсию заряда лизинильных остатков. Другие реагенты, подходящие для дериватизации альфа-аминосодержащих остатков, включают имидо-эфиры, такие как метилпиколинимидат; пиридоксальфосфат; пиридоксаль; хлорборогидрид; тринитро-бензолсульфоновая кислота; О-метилизомочевина; 2,4-пентандион и глиоксилат, реакция с которым катализируется трансаминазой. Аргинильные остатки могут быть модифицированы реакцией с одним или несколькими обычными реагентами, среди которых фенилглиоксаль, 2,3-бутандион, 1,2-циклогександион и нингидрин. При де-риватизации аргининовых остатков необходимо, чтобы реакция проводилась в щелочной среде из-за высокого рКа гуанидиновой функциональной группы. Кроме того, эти реагенты могут реагировать с функциональной группой гуанидина остатков лизина, а также аргинина. Специфическую модификацию тирозильных остатков изучали очень тщательно, особый интерес вызывало введение спектральных меток в тирозильный остаток реакцией с ароматическими диазоние-выми соединениями или тетранитрометаном. Обычно можно применять N-ацетилимидазол и тетранит-рометан с образованием О-ацетилтирозильных фрагментов и 3 нитропроизводных соответственно. Карбоксильные группы боковых цепей (аспарагил или глутамил) можно селективно модифицировать реакцией с карбодиимидами (R'-N=C=N-R'), такими как 1-циклогексил-3-(2-морфолинил-(4-этил)карбодиимид или 1-этил-3-(4-азониа-4,4-диметилпентил)карбодиимид. Кроме того, остатки аспар-тил и глутамил можно превратить в аспарагинил и глутаминил по реакции ионами аммония. Остатки глутаминил и аспарагинил часто дезаминируют в соответствующие остатки глутамил и ас-парагил. Или же эти остатки можно дезаминировать в слабокислой среде. Любые формы этих остатков входят в объём данного изобретения. Дериватизация с помощью бифункциональных агентов применима для перекрёстного связывания (сшивания) пептидов или их функциональных производных с нерастворимой в воде подложкой (мат-рикс) или с другими макромолекулярными носителями. Общеупотребительные агенты для сшивания включают, например, 1,1-бис(диазоацетил)-2-фенилэтан, глутаральдегид, N-гидроксисукцинимидоэфиры, например эфиры 4-азидосалициловой кислоты, гомобифункциональные имидоэфиры, включая дисукцини-мидильные эфиры, такие как 3,3'-дитиобис(сукцинимидил пропионат), и бифункциональные малеинимиды, такие как бис-^малеинимидо-1,8-октан. Дериватизирующие агенты, такие как метил-3-[(п-азидофенил) дитио]пропиоимидат, дают фотоактивируемые интермедиаты, которые способны сшиваться на свету. Или же для иммобилизации белков можно использовать реакционноспособные нерастворимые в воде матрицы, такие как активируемые цианогенбромидом углеводы и реакционноспособные субстраты, описанные в патентах США 3969287; 3691016; 4195128; 4247642; 4229537 и 4330440. Другие возможные модификации включают гидроксилирование пролина и лизина, фосфорилирова-ние гидроксильных групп серильного или треонильного остатков, окисление атома серы в Cys, метилирование альфа-аминогрупп боковых цепей лизина, аргинина и гистидина. Creighton, T.E., Proteins: Structure and Molecule Properties (W.H. Freeman & Co., San Francisco), pp. 79-86 (1983). Такие дериватизированные фрагменты предпочтительно улучшают одну или более характеристик соединений, включая антиангиогенную активность, растворимость, абсорбцию, биологический период полужизни и т.п. Или же дериватизацией фрагментов можно получить соединения, которые имеют такие же или практически такие же характеристики и/или свойства, что и у недериватизированного соединения. Фрагменты можно либо элиминировать, либо можно ослаблять любой нежелательный побочный эффект соединения и т. п. Соединения по данному изобретению можно также изменять на уровне ДНК. Последовательность ДНК любого участка соединения можно изменять на кодоны, более совместимые с выбранной клеткой-хозяином. Для Е. coli, которая является предпочтительной клеткой-хозяином, оптимизированные кодоны известны из уровня техники. Кодоны могут быть замещены таким образом, чтобы элиминировать сайты рестрикции или ввести молчащие сайты рестрикции, которые могут способствовать процессированию ДНК в выбранной клетке-хозяине. Последовательности ДНК носителя, линкера и пептида могут быть модифицированы таким образом, чтобы включить любые из вышеуказанных изменений в последовательностях. Таким образом, все модификации, замены, дериватизации и т.д., обсуждаемые в данном описании, равно применимы ко всем аспектам настоящего изобретения, включая, но без ограничения, пептиды, димеры и мультимеры пептидов, линкеры и носители. Кроме того, специалист в данной области техники может просмотреть работы по теме "структура-свойство", идентифицирующие остатки в аналогичных пептидах, важные для активности или структуры. С учётом такого сравнения можно предсказать важную роль аминокислотных остатков в пептиде, которые соответствуют аминокислотным остаткам, имеющим важное значение для активности или структуры аналогичных пептидов. Специалист в данной области техники может выбрать химически аналогич - 31 - 008248 ные замены на такие прогнозированные важные аминокислотные остатки. Специалист в данной области техники может также проанализировать трёхмерную структуру и аминокислотную последовательность в сравнении с такой структурой в аналогичных полипептидах. С учётом такой информации специалист в данной области техники может прогнозировать совмещение аминокислотных остатков пептида относительно его трёхмерной структуры. Специалист в данной области техники может предпочесть не делать радикальных изменений аминокислотных остатков, которые, как прогнозировалось, расположены на поверхности белка, так как такие остатки могут участвовать в важных взаимодействиях с другими молекулами. Кроме того, специалист в данной области техники может создать пробные варианты, содержащие единственную аминокислотную замену при каждом заданном аминокислотном остатке. Затем можно провести скрининг вариантов, используя методы анализа активности, известные специалистам в данной области техники. Такие данные можно использовать для сбора информации о подходящих вариантах. Например, если обнаруживается, что изменение в конкретном аминокислотном остатке приводит к нарушенной, нежелательно пониженной или неподходящей активности, вариантов с такими изменениями можно избежать. Другими словами, с учётом информации, собранной в таких рутинных экспериментах, специалист в данной области техники может легко определить аминокислоты, дальнейших замен которых, одних или в комбинации с другими мутациями, следует избегать. Ряд научных публикаций посвящен прогнозированию вторичной структуры. См. Moult J., Curr. Op. in Biotech., 7(4)-422-427 (1996), Chou et al., Biochemistry, 13(2):222-245 (1974); Chou et al., Biochemistry, 113(2):211-222 (1974); Chou et al., Adv. Enzymol. Relat. Areas Mol. Biol., 47:45-148 (1978); Chou et al., Ann. Rev. Biochem., 47:251-276 и Chou et al., Biophys. J., 26:367-384 (1979). Кроме того, в настоящее время компьютерные программы могут помочь при прогнозировании вторичной структуры. Один метод прогнозирования вторичной структуры основан на моделировании по гомологии. Например, два полипептида или белка, последовательности которых идентичны более чем на 30%, или подобие которых выше 40%, часто имеют подобную структурную топологию. Увеличение за последнее время структурной базы данных белков (PDB) обеспечило повышенную прогнозируемость (предсказуемость) вторичной структуры, включая потенциальное число складок в структуре полипептида или белка. См. Holm et al., Nucl. Acid. Res., 27(l):244-247 (1999). Было высказано предположение (Brenner et al., Curr. Op. Struct. Biol., 7(3):369-376 (1997)), что в данном полипептиде или белке существует ограниченное число складок и что, если решено критическое число структур, точность предсказания структур резко возрастает. Дополнительные методы предсказания вторичной структуры включают "вытягивание (нити)" (threading) (Jones, D., Curr. Op. Struct. Biol., 7(3):377-87 (1997); Sippl et al., Structure, 4(1):15-19 (1996)); "профильный анализ"" (Bowie et al., Science, 253:164-170 (1991); Gribskov et al., Meth. Enzym., 183:146159 (1990); Gribskov et al., Proc. Nat. Acad. Sci., 84(13):4355-4358 (1987)), и "эволюционное связывание" (Holm, см. выше (1999), и Brenner, см. выше (1997)). Кроме того, изобретение охватывает производные специфически связывающихся агентов, например, " пептитела", ковалентно модифицированные таким образом, чтобы включить присоединение одного или более водорастворимых полимеров, таких как полиэтиленгликоль, полиоксиэтиленгликоль или по-липропиленгликоль, описанные в патентах США: 4640835; 4496689; 4301144; 4670417; 4791192 и 4179337. Другие известные в технике подходящие полимеры включают: монометокси-полиэтилен-гликоль, декстран, целлюлозу или другие полимеры на основе углеводов, поли-^-винилпирролидон)-полиэтиленгликоль, гомополимеры полипропиленгликоля, сополимер полипропиленоксид/этиленоксид, полиоксиэтилированные полиолы (например, глицерин) и поливиниловый спирт, а также смеси этих полимеров. Особенно предпочтительными являются "пептитела", ковалентно модифицированные субъединицами полиэтиленгликоля (ПЭГ). Водорастворимые полимеры присоединяться по конкретным положениям, например, по аминоконцам "пептител", или произвольно присоединяться к одной или более боковых цепей полипептида. Применение ПЭГ для улучшения терапевтических свойств специфически связывающихся агентов, например, "пептител", и для "гуманизации" антител в частности, описано в патенте США 6133426, принадлежащем Gonzales et al., выданном 17 октября 2000 г. В изобретении также рассматривается дериватизация пептидной области соединений и/или области соединений, являющуюся носителем. Такие производные позволяют повысить растворимость, абсорбцию, биологический период полужизни и т.п. соединений. Фрагменты можно либо элиминировать, либо можно ослаблять любой нежелательный побочный эффект соединения и т. п. Примеры производных включают соединения, в которых 1. Соединение или некий его участок является циклическим. Например, пептидный участок можно модифицировать таким образом, чтобы он содержал два или более Cys остатков (например, в линкере), которые могут циклизоваться за счёт образования дисульфидной связи. 2. Соединение является сшитым или его молекулы способны образовывать перекрёстные связи (сшивание). Например, пептидный участок можно модифицировать таким образом, чтобы он содержал Cys остаток и поэтому мог образовывать межмолекулярную дисульфидную связь с аналогичной молекулой. Соединение может также сшиваться по С-концу. 3. Одна или более пептидильных [-C(O)NR-] связей (фрагментов) замещается на непептидильную - 32 - 008248 связь. Примерами непептидильных связей являются -СН2-карбамат [-CH2-OC(O)NR-], фосфонат, -СН2-сульфонамид [-CH2-S(O)2NR-], уреилен (мочевина) [-NHC(O)NH-], -СН2-вторичный амин и алкилиро-ванный пептид [-C(O)NR6-, где R6 обозначает низший алкил]. 4. N-конец дериватизирован. Как правило, N-конец может быть ацилирован или модифицирован превращением в соответствующий амин. Примеры производных N -концевых групп включают -NRR1 (иной, нежели -NH2), -NRC(O)R1, -NRC(O)OR1, -NHS(O)2R1, -NHC(O)NHR1, сукцинимид или бензилок-сикарбонил -NH- (CBZ-NH-), в которых R и R1, каждый независимо, обозначают водород или низший алкил и фенильное кольцо может иметь от 1 до 3 заместителей, выбираемых из группы, состоящей из С1-С4алкила, С1-С4алкокси, хлора и брома. 5. Свободный С-конец дериватизирован. Как правило, С-конец этерифицируют или амидируют (превращают в амид). Например, можно использовать описанные в технике методы присоединения (NH-CH2-CH2-NH2)2 по С-концу соединений по изобретению. Аналогично можно использовать описанные в технике методы для присоединения -NH2 по С-концу соединений по изобретению. Примеры производных С-концевых групп включают -C(O)R2, где R2 обозначает низший алкокси, или -NR3R4, где R3 и R4 независимо обозначают водород или ^-^алкил (предпочтительно Q^^ICM). 6. Дисульфидную связь (мостик) заменяют другим, предпочтительно более устойчивым сшивающим фрагментом (например, алкиленовым мостиком). См., например, Bhatnagar см. выше; Alberts et al. Thirteenth Am. Pep. Symp., 357-9 (1993). 7. Один или более отдельных аминокислотных остатков модифицирован. Известны различные де-риватизирующие агенты, которые специфически реагируют с выбранными остатками боковых цепей или концевыми остатками, как подробнее описано ниже. Лизинильные остатки или аминоконцевые остатки могут реагировать с ангидридами янтарной или других карбоновых кислот, которые меняют на обратный (реверсия) заряд лизинильных остатков. Другие подходящие реагенты для дериватизации альфа-аминосодержащих остатков включают имидоэфиры, такие как метилпиколинимидат; пиридоксальфосфат; пиридоксаль; хлорборогидрид; тринитробензол-сульфокислоту; О-метилизомочевину; 2,4-пентандион; и глиоксилат, реакцию с которым катализирует трансаминаза. Аргинильные остатки могут быть модифицированы реакцией с любым или с комбинацией нескольких обычных реагентов, включая фенилглиоксаль, 2,3-бутандион, 1,2-циклогександион и нингидрин. Дериватизация аргинильных остатков требует, чтобы реакцию проводили в щелочной среде вследствие высокого значения рКа функциональной группы гуанидина. Кроме того, эти реагенты могут реагировать с группами лизина, а также с эпсилон-аминогруппой аргинина. Очень тщательно изучалась специфическая модификация тирозильных остатков, при этом особый интерес вызвало введение спектральных меток в тирозильные остатки по реакции с ароматическими диа-зониевыми соединениями или тетранитрометаном. Чаще всего используют N-ацетилимидазол и тетра-нитрометан для образования О-ацетильтирозильных фрагментов и 3-нитропроизводных, соответственно. Карбоксильные группы боковых цепей (аспарагил или глутамил) можно селективно модифицировать реакцией с карбодиимидами (R'-N=C=N-R'), например с 1-циклогексил-3-(2-морфолинил-(4-этил)карбоди-имидом или с 1-этил-3-(4-азониа-4,4-диметилпентил)карбодиимидом. Кроме того, остатки аспарагил и глу-тамил могут быть превращены в аспарагинильный и глутаминильный остатки реакцией с ионами аммония. Глутаминильный и аспарагинильный остатки можно дезаминировать до соответствующих глута-мильного и аспарагильного остатков. Или же можно дезаминировать в слабокислой среде. Любая форма этих остатков входит в объём данного изобретения. Цистеинильные остатки могут быть замещены аминокислотными остатками или другими фрагментами либо с целью элиминировать дисульфидный мостик, либо, наоборот, стабилизировать сшивание (молекул). См., например, Bhatnagar, см. выше. Дериватизация с помощью бифункциональных агентов применима для перекрёстного связывания (сшивания) пептидов или их функциональных производных с нерастворимой в воде подложкой (матрикс) или с другими макромолекулярными носителями. Общеупотребительные агенты для сшивания включают, например, 1,1-бис(диазоацетил)-2-фенилэтан, глутаральдегид, N-гидроксисукцинимидоэфиры, например эфиры 4-азидосалициловой кислоты, гомобифункциональные имидоэфиры, включая дисукцинимидиль-ные эфиры, такие как 3,3'-дитиобис(сукцинимидил пропионат), и бифункциональные малеинимиды, такие как бис-N-малеинимидо-1,8-октан. Дериватизирующие агенты, такие как метил-3-[(п-азидофенил)ди-тио]пропиоимидат, дают фотоактивируемые интермедиаты, которые способны сшиваться на свету. Или же для иммобилизации белков используют реакционноспособные нерастворимые в воде матрицы, такие как активируемые цианогенбромидом углеводы и реакционноспособные субстраты, описанные в патентах США 3969287; 3691016; 4195128; 4247642; 4229537 и 4330440. Углеводные (олигосахаридные) группы удобно присоединять к сайтам, известным как сайты глико-зилирования в белках. Как правило, О-связанные олигосахариды присоединяются к остаткам серина (Ser) или треонина (Thr), тогда как N-связанные олигосахариды присоединяются к остаткам аспарагина (Asn), когда они являются частью последовательности Asn-X-Ser/Thr, где X может быть любой аминокислотой, кроме пролина. X, предпочтительно, является одной из 19 природных аминокислот, кроме пролина. - 33 - 008248 Структура N-связанных и О-связанных олигосахаридов и остатков cахаров, обнаруживаемых в каждом типе, различна. Один тип cахаров, который обычно находят в обоих случаях, представляет собой N-ацетилнейраминовую кислоту (называемую сиаловой кислотой). Сиаловая кислота является обычно концевым остатком как N-связанных, так и О-связанных олигосахаридов и, вследствие своего отрицательного заряда , может придавать кислые свойства гликозилированному соединению. Такой(ие) сайт(ы) можно включать в линкер в линкер соединений по данному изобретению и предпочтительно гликозилировать при участии клетки в процессе рекомбинантного продуцирования полипептидных соединений (например, в клетках млекопитающих, таких как СНО, ВН^ COS). Однако такие сайты можно дополнительно гликозилировать синтетическими или полусинтетическими методами, известными в технике. Другие возможные модификации включают гидроксилирование пролина и лизина, фосфорилирова-ние гидроксильных групп серильного или треонильного остатков, окисление атома серы в Cys, метилирование альфа-аминогрупп боковых цепей лизина, аргинина и гистидина. [Creighton, Proteins: Structure and Molecule Properties (W.H. Freeman & Co., San Francisco), pp. 79-86 (1983). Соединения по данному изобретению можно также изменять на уровне ДНК. Последовательность ДНК любого участка соединения можно заменять на кодоны, более совместимые с выбранной клеткой-хозяином. Для Е. coli, которая является предпочтительной клеткой-хозяином, оптимизированные кодоны известны из уровня техники. Кодоны могут быть замещены таким образом, чтобы элиминировать сайты рестрикции или ввести молчащие сайты рестрикции, которые могут способствовать процессированию ДНК в выбранной клетке-хозяине. Последовательности ДНК носителя, линкера и пептида могут быть модифицированы таким образом, чтобы включить любые из вышеуказанных изменений в последовательностях. Созревание аффинности Один вариант настоящего изобретения включает "зрелые с точки зрения аффинности" ("зрелые по аффинности") пептиды и "пептитела". Этот метод рассматривает повышение аффинности или биоактивность пептидов и "пептител" по данному изобретению с использованием фагового дисплея или других методов отбора. На основе консенсусной последовательности (которую создают для коллекции родственных пептидов) могут быть созданы направленные вторичные библиотеки пептидов методом фагового дисплея, в которых "коровые" аминокислоты (определяемые из консенсусной последовательности) остаются постоянными или смещёнными по их встречаемости. Или же для создания смещённой направленной библиотеки пептидов по методу фагового дисплея можно использовать отдельную пептидную последовательность. Пэннингом таких библиотек можно получить пептиды (которые можно превратить в "пептитела") с повышенным связыванием с Ang-2 или с повышенной биоактивностью. Непептидные аналоги/белки-миметики Далее рассматриваются также непептидные аналоги пептидов, которые обеспечивают стабилизированную структуру или пониженное биологическое разложение. Аналоги-пептидомиметики можно получать на основе выбранного ингибирующего пептида заменой одного или более остатков на непептидные фрагменты. Предпочтительно непептидные фрагменты дают возможность пептиду сохранять его естественную конформацию или стабилизировать предпочтительную, например биоактивную конформацию, которая сохраняет способность распознавать Ang-2. В одном аспекте полученный аналог/миметик проявляет повышенную аффинность связывания с Ang-2. Один пример способов получения непептидных миметических (имитирующих) аналогов с участием пептидов описан в Nachman et al., Regul. Pept. 57: 359-370 (1995). При желании пептиды по изобретению можно модифицировать, например, гликозилиро-ванием, амидированием, карбоксилированием или фосфорилированием или получать из них соли присоединения, амиды, эфиры, в частности, С-концевые эфиры, N-ацильные производные пептидов по изобретению. "Пептитела" также можно модифицировать, получая пептидные производные за счёт образования ковалентных или нековалентных комплексов с другими фрагментами. Ковалентно связанные комплексы можно получать, соединяя химические фрагменты с функциональными группами в боковых цепях аминокислот, составляющих "пептитела", или на N- или С-конце. В частности, предполагается, что пептиды могут конъюгировать с контрольной группой, включая, но без ограничения, радиоактивную метку, флуоресцентную метку, фермент (например, катализирующий колориметрическую или флуорометрическую реакцию), субстрат, твёрдая матрица (подложка) или носитель (например, биотин или авидин). Таким образом, изобретение включает молекулу, содержащую молекулу "пептитела", причём эта молекула предпочтительно дополнительно содержит репортёрную (контрольную) группу, выбираемую из группы, состоящей из радиоактивной метки, флуоресцентной метки, фермента, субстрата, твёрдой матрицы (подложки) и носителя. Такие метки хорошо известны специалистам в данной области техники, например, специально рассматриваются биотиновые метки. Применение таких меток хорошо известно специалистам в данной области техники и описано, например, в патентах США 3817837; 3850752; 3996345 и 4277437. Другие подходящие метки включают, но без ограничения, радиоактивные метки, флуоресцентные метки и хемилюминесцентные метки. В патентах США 3817837, 3850752; 3939350 и 3996345 рассматривается применение таких меток. Любые пептитела по данному изобретению могут содержать одну, две или более любых из этих меток. - 34 - 008248 Методы получения пептидов Пептиды по данному изобретению можно получать, используя широкий ряд методов, известных в технике. Например, такие пептиды можно синтезировать в растворе или на твёрдой подложке (твёрдом носителе) обычными методами. Промышленностью выпускаются различные автоматические синтезаторы и их можно применять в соответствии с известными протоколами. См., например, Stewart and Young (см. выше); Tam et al., J. Am. Chem. Soc., 105:6442, (1983); Merrifield, Science 232:341-347 (1986); Barany and Merrifield, The Pentides, Gross and Meienhofer, eds, Academic Press, New York, 1-284; Barany et al., Int. J. Pep. Protein Res., 30:705-739 (1987); и патент США 5424398, каждый из источников вводится в данное описание в качестве ссылки. В методах твердофазного пептидного синтеза применяют сополимер стирола с дивинилбензолом, содержащим 0,1-1,0 мМ аминов/г полимера. В этих методах пептидного синтеза используют бутилокси-карбонильную (трет-ВОС) или 9-фторметилоксикарбонильную (FMOC) группы для защиты альфа-аминогрупп. Оба метода включают постадийный синтез, при котором на каждой стадии добавляют одну аминокислоту, начиная с С-конца пептида (См. Coligan et al., Curr. Prot. Immunol., Wiley Interscience, 1991, Unit 9). По завершении химического синтеза можно провести депротекцию синтетического пептида с удалением блокирующих аминокислоты групп трет-ВОС или FMOC и отщеплением полимера обработкой кислотой при пониженной температуре (например, жидкий HF-10% анизол в течение, примерно, 0,25-1 ч при 0°С). После упаривания реагентов пептиды экстрагируют из полимера 1% раствором уксусной кислоты, который затем лиофилизируют, получая сырой продукт. Этот продукт можно обычно очищать такими методами, как гель-фильтрация на Sephadex G-15, используя в качестве растворителя 5% уксусную кислоту. Лиофилизацией соответствующих фракций после элюции с колонки получают гомогенные пептиды или производные пептидов, которые можно затем охарактеризовать такими стандартными методами, как аминокислотный анализ, тонкослойная хроматография, высокоэффективная жидкостная хроматография, УФ спектроскопия, молекулярное вращение, растворимость и количественно рассчитывают методом расщепления по Эдману в твердой фазе. Также применяются другие методы, такие как селекция пептидов из библиотеки на основе метода фагового дисплея. Библиотеки можно получать из наборов аминокислот по данному описанию. Метод фагового дисплея может быть особенно эффективен при идентификации пептидов, применимых по данному изобретению. Коротко говоря, получают "фаговую библиотеку" (используя, примерно, мл 13, fd или лямбда фага), выявляющую инсерты от 4, примерно, до 80 аминокислотных остатков. Инсерты могут представлять собой, например, полностью выраженный или смещённый ранжированный ряд. Затем можно выбрать несущие фаг инсерты, которые связываются с нужным антигеном. Этот процесс можно повторять с помощью нескольких циклов повторной селекции фага, который связывается с антигеном. Повторные циклы обогащают несущих фаг конкретных последовательностей. Можно проводить анализ последовательности ДНК для идентификации последовательностей экспрессируемых пептидов. Можно определить минимальный линейный участок последовательности, который связывается с заданным антигеном. Можно повторить процедуру, используя "смещённую" библиотеку, включающую инсерты, содержащие часть целого или минимальный линейный участок плюс один или более дополнительных вырожденных остатков в обратном (3'-5') или прямом (5'-3') направлении от него. Эти методы позволяют идентифицировать пептиды, которые ещё сильнее связываются с Ang-2, чем агенты, уже идентифицированные по данному описанию. Вне зависимости от метода получения пептидов, молекулу нуклеотида, кодирующего каждый такой пептид и каждое такое "пептитело", можно получать стандартными методами рекомбинантной ДНК. Нуклеотидной последовательностью такой молекулы ДНК можно манипулировать соответствующим образом, не изменяя аминокислотной последовательности, которую они кодируют, чтобы получить вырожденный нуклеотидный код, а также предпочтение по кодону в конкретных клетках-хозяевах. Методы рекомбинантной ДНК являются удобными методами получения полноразмерных полипептидов и других больших белковых специфически связывающихся агентов по данному изобретению или их фрагментов. Молекулу ДНК, кодирующую "пептитело" или его фрагмент, можно встроить в экспрес-сирующий вектор, который, в свою очередь, вводят в клетку-хозяина для продуцирования антитела или фрагмента. Как правило, молекулу ДНК, кодирующую пептид или "пептитело", можно получить по методикам, представленным в разделе "Примеры" данного описания. Зонды и типичные условия гибридизации описаны в Ausubel et al. (Current Protocols in Molecular Biology, Current Protocols Press [1994]). После гибридизации блот с зондом отмывают в условиях с подходящей жёсткостью в зависимости от таких факторов, как размер зонда, ожидаемая степень гомология зонда с клоном, тип библиотеки, скрининг которой проводится, и число клонов, которое подвергается скринингу. Примеры скрининга в очень жёстких условиях суть 0.1 X SSC и 01% SDS при температуре около 50-65°С. Для идентификации пептидов по изобретению, связывающихся с Ang-2, можно также применять метод двухгибридного скрининга в дрожжах. Так, антиген или его фрагмент можно использовать для скрининга пептидных библиотек, включая библиотеки "фагового дисплея", чтобы идентифицировать и выбрать агенты, связывающиеся с Ang-2, например "пептитела", по данному изобретению. - 35 - 008248 Или же для содержания и экспрессии пептидов по изобретению можно применять системы экспрес-сирующий вектор/хозяин. Эти системы включают, но без ограничения, микроорганизмы, такие как бактерии, трансформированные рекомбинантным бактериофагом, векторы экспрессии плазмидных или кос-мидных ДНК, дрожжи, трансформированные с использованием дрожжевых векторов экспрессии; системы клеток насекомых, инфицированных вирусными векторами экспрессии (например, вирусом мозаики цветной капусты, CaMV; вирусом табачной мозаики; ТМВ) или системами животных клеток. Клетки млекопитающих, применимые для получения рекомбинантных белков, включают, но без ограничения, клетки VERO, клетки HeLa, клетки яичников китайского хомячка (СНО), клетки COS (такие как COS-7), клетки W138, ВШС, HepG2, 3Т3, RIN, MDCK, А549, РС12, K562 и 293. Примеры протоколов рекомби-нантной экспрессии пептидов представлены ниже в данном описании. Термин "вектор экспрессии", "экспрессирующий вектор" относится к плазмиде, фагу, вирусу или вектору для экспрессии полипептида при использовании последовательности ДНК (РНК). Экспресси-рующий вектор может содержать транскрипционную единицу, представляющую собой сборку (1) генетического элемента или генетических элементов, играющих роль регуляторных элементов в генной экспрессии, например, промоторов или энхансеров, (2) структуры или последовательности, кодирующей связывающийся агент, которая транскрибируется в мРНК и транслируется в белок, и (3) соответствующих последовательностей инициации и терминации транскрипции. Структурные единицы для предполагаемого применения в дрожжевых и эукариотньгх системах экспрессии, предпочтительно, включают лидерную последовательность, способствующую внеклеточной секреции транслированного белка клеткой-хозяином. Или же, если рекомбинантный белок экспрессируется без лидерной или транспортной последовательности, он может включать аминоконцевой метионильный остаток. Этот остаток может затем отщепляться или не отщепляться от экспрессированного рекомбинантного белка с целью получения конечного пептидного продукта. Например, пептиды можно рекомбинантно экспрессировать в дрожжах, используя промышленную систему экспрессии, например, Pichia Expression System (Invitrogen, San Diego, CA), следуя инструкциям производителя. Основную роль в непосредственной экспрессии в этой системе также играет пре-про-альфа последовательность, но транскрипция инсерта управляется промотором алкогольоксидазы (АОХ1) при индукции метанолом. Секретируемый пептид очищают от питательной среды для дрожжей, например, методами, применяемыми для очистки пептида от супернатанта бактериальных клеток и клеток млекопитающих. Или же кДНК, кодирующую пептид, можно клонировать в бакуловирусный вектор экспрессии pVL1393 (PharMingen, SanDiego, CA). Этот вектор можно использовать в соответствии с рекомендациями производителя (PharMingen) для инфицирования клеток Spodoptera frugiperda в безбелковых средах sF9 и для продуцирования рекомбинантного белка. Рекомбинантный белок можно очистить от среды и концентрировать на колонке с гепарин-сефарозой (Pharmacia). Или же пептид можно экспрессировать в системе экспрессии в клетках насекомых. Системы экспрессии белка в клетках насекомых хорошо известны специалистам в данной области техники. В одной такой системе вирус ядерного полиэдроза Autographa californica (AcNPV) можно использовать в качестве вектора для экспрессии чужеродных генов в клетках Spodoptera frugiperda или Trichoplusia larvae. Последовательность, кодирующую пептид, можно клонировать в не основной участок вируса, такого как ген полиэдрина, и поместить под контроль промотора полиэдрина. Успешная инсерция пептида сделает ген полиэдрина неактивным, в результате продуцирует рекомбинантный вирус без оболочки оболочечного белка. Рекомбинантные вирусы можно использовать для инфицирования клеток S. frugiperda или Trichoplusia larvae, в которых экспрессируется пептид. Smith et al., J. Virol. 46: 584 (1983); Engelhardt et al., Proc. Nat. Acad. Sci. (USA) 91: 3224-7 (1994). В другом примере последовательность ДНК, кодирующую пептид, можно амплифицировать при использовании ПЦР и клонировать в соответствующий вектор, например, pGEX-3X (Pharmacia). Вектор pGEX-3X создан для продуцирования слитого белка, содержащего глутатион^-трансферазу (GST), кодируемую вектором, и белок, кодируемый фрагментом ДНК, встроенным в сайт клонирования вектора. Можно получать праймеры для ПЦР, включающие, например, соответствующий сайт расщепления. Когда фрагмент слияния используют только для облегчения экспрессии или иным образом, нежелательным с точки зрения присоединения к пептиду, представляющему интерес, рекомбинантный слитый белок можно затем отщепить от участка GST слитого белка. Пептидную конструкцию pGEX-3Х/связывающийся агент трансформируют в клетки Е. coli XL-1 Blue (Stratagene, La Jolla CA), и отдельные трансформанты выделяют и выращивают. Плазмидную ДНК индивидуальных трансформантов можно очистить и частично секвенировать, используя автоматический секвенатор, с целью подтвердить присутствие заданного специфически связывающегося агента, кодирующего нуклеотидный инсерт в подходящей ориентации. Некоторые составы пептидов по данному изобретению представляют собой такие составы, в которых "пептитело" конъюгировано с любым противоопухолевым пептидом, таким как фактор некроза опухолевых клеток (TNF). В особенно предпочтительном методе химерные пептиды TNF-специфически связывающийся агент получают в виде рекомбинантных слияний с кодирующими пептид последователь - 36 - 008248 ностями, слитыми в рамке считывания с последовательностями, кодирующими TNF (Novagen, Madison, WI). кДНК пептид-TNF можно клонировать в вектор pET-11b (Novagen) и экспрессию TNF-пептидов в BL21 Е. coli можно индуцировать в соответствии с инструкциями производителя рЕТ1Ш. Растворимые TNF-пептиды можно очистить от бактериальных лизатов препаратом сульфата аммония, хроматографией с гидрофобным взаимодействием на Фенил-Сефарозе Phenil-Sepharose 6 Fast Flow, ионообменной хроматографией на DEAE-Sepharose Fast Flow и гель фильтрацией Sephacryl-S-300 HR. Слитый белок, который можно продуцировать в виде нерастворимых телец включения в бактериях, можно очистить следующим образом. Клетки-хозяева можно разрушить центрифугированием: отмыть 0,15М NaCl, 10 мМ Tris, pH 8, 1 мМ EDTA и обработать 0,1 мг/мл лизоцима (Sigma, St. Louis, МО) в течение 15 мин при комнатной температуре. Лизат можно осветляют ультразвуком и клеточный дебрис высаждают центрифугированием в течение 10 мин при 12000 X g. Пеллеты, содержащие слитый белок, снова суспендируют в 50 мМ Tris, pH 8 и 10 мМ EDTA, расслаивают с 50% глицерином и центрифугируют в течение 30 мин при 6000 X g. Дебрис можно снова суспендировать в стандартном фосфатно-солевом буферном растворе (PBS), не содержащем Mg++ и Са++. Слитый белок можно дополнительно очистить, фракционируя повторно суспендированный дебрис в денатурирующем SDS-PAGE (Sambrook et al., см. выше). Гель можно пропитать 0,4М KQ для визуализации белка, вырезать и выделить методом электроэлюции, элюируя буфером для промывания геля, не содержащим SDS. Если GST/слитый белок получают в клетках бактерий в виде растворимого белка, его можно очистить с применением GST Purification Module (Pharmacia). Слитый белок можно подвергнуть расщеплению, чтобы отщепить GST от пептида по изобретению. Реакция гидролиза (20-40 мг слитого белка, 20-30 единиц человеческого тромбина (4000 Ед/мг, Sigma) в 0,5 мл PBS можно инкубировать в течение 16-48 ч при комнатной температуре и загружают на денатурирующем SDS-PACE геле с целью фракционировать продукты реакции. Гель можно пропитать 0,4М KO для визуализации полос белка. Идентичность полос белка, соответствующих ожидаемой молекулярной массе пептида, можно подтвердить анализом аминокислотной последовательности (Applied Biosystems Model 473A, Foster City CA). Или же идентичность можно подтвердить методами ВЭЖХ и/или масс-спектрометрии пептидов. Или же последовательность ДНК, кодирующую пептид, можно клонировать в плазмиду, содержащую заданный промотор и, необязательно, лидерную последовательность [Better et al., Science 240; 1041-43 (1988)]. Последовательность этой конструкции можно подтвердить автоматическим секвенированием. Затем плазмиду можно стандартными методами трансформировать в штамм МС1061 Е. coli, подвергая бактерии инкубации с Са02 и тепловому шоку (Sambrook et al., см. выше). Трансформированные бактерии можно выращивать в среде LB, дополненной карбенициллином, и продуцирование экспрессированного белка можно индуцировать выращиванием в соответствующей среде. Если присутствует лидерная последовательность, она может осуществлять секрецию пептида и её можно отщеплять в процессе секреции. Секретированный рекомбинантный белок можно очищать от бактериальной культуральной среды, представленными далее в настоящем описании. Системы хозяев-млекопитающих для экспрессии рекомбинантного белка хорошо известны специалистам в данной области техники. Штаммы клеток-хозяев можно выбирать в соответствии с конкретной способности процессировать экспрессируемый белок или продуцировать некоторые посттрансляционные модификации, которые могут быть полезны для придания белку активности. Такие модификации белка включают, но без ограничения, ацетилирование, карбоксилирование, гликозилирование, фосфори-лирование, обработку липидом и ацилирование. У различных клеток-хозяев, таких как СНО, HeLa, MDCK, 293, W138 и т.п., различные клеточные механизмы, и можно выбрать характеристические механизмы для такой посттрансляционной активности, чтобы гарантировать корректную модификацию и процессирование введённого чужеродного белка. Предпочтительно, чтобы трансформированные клетка можно было применять для получения белка с высоким выходом в течение длительного времени и, следовательно, желательна стабильная экспрессия. После того как такие клетки трансформированы при использовании вектора, который содержит селективные маркёры наряду с заданной кассетой экспрессии, клетки можно выращивать в течение 1-2 дней в обогащенной среде перед тем, как поменять её на селективную среду. Создают селективный маркёр для придания резистентности к селекции, и его присутствие даёт возможность выращивать и очищать клетки, которые последовательно экспрессируют введённые последовательности. Резистентные группы стабильно трансформированных клеток можно пролиферировать, используя методы работы с тканевыми культурами, подходящие для клетки. Для очистки клеток, трансформированных с целью продуцирования рекомбинантных белков, можно использовать ряд систем селекции. Такие системы селекции включают, но без ограничения, гены HSV тимидинкиназы, гипоксантин-гуанин фосфорибозилтрансферазы в tk-, hgpt- или aprt-клетках соответственно. А резистентность к антиметаболитам можно использовать как основу для селекции на (ген) DHFR, который придаёт устойчивость (резистентность) к метотрексату; (ген) gpt, который придаёт устойчивость к микофенольной кислоте; (ген) шо, который придаёт устойчивость к аминогликозиду G418 к хлорсульфурону; и (ген) hygro, который придаёт устойчивость к гигромицину. Дополнительные селек - 37 - 008248 тивные гены, которые можно применять, включают trpB, который делает возможным использовать в клетках индол вместо триптофана, или hisD, позволяющий использовать в клетках гистинол вместо гис-тидина. Маркёры, наличие которых даёт визуальную индикацию для идентификации трансформантов, включают антоцианины, в-глюкуронидазу и её субстрат, GUS, люциферазу и её субстрат, люциферин. Очистка и переукладка специфически связывающихся агентов В некоторых случаях может потребоваться "переукладка" (повторная укладка) и окисление специфически связывающихся агентов, таких как пептиды и/или "пептитела" в соответствующие третичные структуры, и образование дисульфидных связей для достижения биологической активности. "Переукладку" можно выполнить многими методами, хорошо известными в технике. Такие методы включают, например, экспонирование стабилизированного полипептидного агента при рН, как правило, выше 7 в присутствии хаотропного агента. Селекция хаотропного агента подобна отбору, используемому для со-любилизации телец включения, однако, хаотропный агент, как правило, используют в более низкой концентрации. Примером хаотропного агента является гуанидин. В большинстве случаев раствор для переукладки/окисления содержит также восстанавливающий агент плюс его окисленную форму в специфическом соотношении с целью достижения конкретного окислительно-восстановительного потенциала, который делает возможным "перетасовку" дисульфидных групп с образованием цистеиновых мостиков. Некоторые общеупотребительные пары окислитель-восстановитель включают цистеин/цистамин, глутати-он/дитиобисGSН, монохлорид меди, дитиотреитол DТТ/дитианDТТ и 2-меркаптоэтанол (ЬМЕ)/ДИТИО-ЬМЕ. Во многих случаях для повышения эффективности переукладки можно использовать сораствори-тель. Обычно в качестве сорастворителей используют глицерин, полиэтиленгликоль с различной молекулярной массой и аргинин. Может потребоваться очистка пептидов и "пептител" по данному изобретению. Методы очистки белков хорошо известны специалистам в данной области техники. Эти методы включают, на первой стадии, грубое разделение белковых и небелковых фракций. После отделения пептида и/или "пептитела" от других белков представляющие интерес пептид или "пептитело" можно далее очищать хроматографиче-скими и электрофоретическими методами, достигая частичной или полной очистки (или очистки до гомогенности). Аналитическими методами, особенно пригодными для получения "пептител" и пептидов по данному изобретению, являются ионообменная хроматография, эксклюзионная хроматография; электрофорез в акриламидном геле; изоэлектрофокусировка. Особенно эффективным методом очистки пептидов является скоростная жидкостная хроматография белков или даже ВЭЖХ. Некоторые аспекты настоящего изобретения относятся к очистке и, в конкретных вариантах, к практически полной (существенной) очистке, "пептитела" или пептида по данному изобретению. Понятие "очищенное "пептитело" или "очищенный пептид" по данному описанию предполагает состав полипептида, способный отделяться от других компонентов, при этом "пептитело" или пептид очищают до некоторой степени, сравнимой с его естественньм состоянием. Следовательно, понятие "очищенный пептид" или "очищенное "пептитело" также относятся к "пептителу" или к пептиду, не содержащему примесей среды, в которой он может встречается в природе. Как правило, понятие "очищенный " относится к составу пептида или "пептитела", который подвергся фракционированию с целью удаления различных других компонентов, при этом составы пептида или "пептитела" практически сохраняют свою экспрессируемую биологическую активность. Если употребляется понятие "практически чистый", это обозначение относится к составу пептида или "пептитела", в котором "пептитело" или пептид образуют основной компонент состава, например, составляющий около 50, около 60, около 70, около 80, около 90, около 95% или более белков в составе. Различные методы количественной оценки степени очистки пептида или "пептитела" станут понятны специалистам в данной области техники в свете настоящего описания. Эти методы включают, например, определение активности специфического связывания активной фракции или оценку количества пептида или "пептитела" во фракции методом SDS/PAGE. Предпочтительный метод оценки чистоты пептидной фракции или фракции "пептитела" представляет собой расчёт активности связывания фракции, сравнение с активностью связывания исходного экстракта и следующий из этого расчёт степени очистки, в данном описании оцениваемой как "-кратный показатель очистки". Действительные числа, применяемые для того, чтобы представить величину активности связывания, естественно, будут зависеть от конкретного метода анализа, выбранного для очистки, и от того, проявляют или нет "пептитело" или пептид обнаруживаемую активность связывания. Различные методы, пригодные для очистки, хорошо известны специалистам в данной области техники. Эти методы включают, например, осаждение сульфатом аммония, ПЭГ, антителами (иммунопре-ципитация) и т. п. или тепловой денатурацией с последующим центрифугированием; стадии хроматографии, такие как аффинная хроматография (например, на Протеин-А-Сефарозе), ионообменная, гель фильтрация, обращённофазовая, хроматография на гидроксилапатите и аффинная хроматография; изо-электрофокусировка; электрофорез в геле; и комбинация этих и других методов. Как общеизвестно в технике, порядок проведения различных стадий очистки можно изменять или некоторые стадии можно опустить и всё равно получить практически чистый специфически связывающийся агент. Не всегда требуется, чтобы пептид или "пептитело" по настоящему изобретению были абсолютно - 38 - 008248 чистыми. На самом деле, рассматривается, что в некоторых вариантах изобретения применяются менее чистые продукты специфически связывающегося агента. Частичную очистку можно осуществлять, применяя сочетание меньшего количества стадий или используя иные формы той же самой общей схемы очистки. Например, понятно, что катионообменная хроматография с применением ВЭЖХ хроматографа, как правило, даёт большую "кратность" очистки, чем тот же метод с применением хроматографической системы при низком давлении. Методы, дающие более низкую степень относительной очистки, могут иметь преимущества для тотальной очистки пептида или "пептитела" или для сохранения активности связывания пептида или " пептитела". Известно, что миграция пептида или "пептитела" может изменяться, иногда значительно, с изменением условий SDS/PAGE [Capaldi et al., Biochem. Biophys. Res. Comm., 76: 425 (1977)]. Поэтому понятно, что в других условиях электрофореза явная молекулярная масса продуктов экспрессии очищенного или частично очищенного специфически связывающегося агента может изменяться. Анализы связывания В иммуноанализах связывания, как правило, используют агент для захвата (улавливающий, захватывающий агент) с целью специфического связывания с аналитом, антигеном-мишенью, и часто его иммобилизуют. Захватывающий агент представляет собой частицу, которая специфически связывается с аналитом. В одном варианте настоящего изобретения захватывающий агент представляет собой пептид или "пептитело" или его фрагмент, который специфически связывается c Ang-2. Эти методы иммуноана-лиза хорошо известны в технике [Assai, ed., Methods in Cell Biology, Vol. 37, Antibodies in Cell Biology, Academic Press, Inc., New York (1993)]. В иммуноанализах связывания часто применяют метку, которая сигнализирует, сообщая о существовании связанного комплекса, образованного захватывающим агентом и антигеном. Меткой может быть одна из молекул, составляющих связанный комплекс; т. е. это может быть меченый специфически связывающийся агент или меченое антитело против специфически связывающегося агента. Или же, меткой может быть третья молекула, обычно другое антитело, которое связывается со связанным комплексом. Метка может являться, например, антителом против специфически связывающегося агента, несущим метку. У второго антитела, специфического в отношении связанного комплекса, может отсутствовать метка, но оно может быть связано с четвёртой молекулой, специфической в отношении вида антител, представителем которого является второе антитело. Например, второе антитело можно модифицировать, используя детектируемую частицу, такую как биотин, который можно затем связать с четвёртой молекулой, такой как меченый ферментом стрептавидин. Другие белки, способные специфически связываться с константными областями иммуноглобулина, такие как протеин А или протеин G, также можно применять в качестве метки. Эти связывающиеся белки являются нормальными составляющими клеточных стенок стрептококковых бактерий и проявляют сильную неиммуногенную реакционную способность в отношении различных видов константных областей иммуноглобулинов. Akerstrom, J. Immunol., 135: 2589-2542 (1985); Chaubert, Mod. Pathol., 10: 585-591 (1997). Во всех анализах после каждой комбинации реагентов могут требоваться стадии инкубации и/или отмывания. Время стадий инкубации может варьироваться от 5 с до нескольких часов, предпочтительно примерно 5 мин - 24 ч. Однако время инкубации зависит от формата анализа, аналита, объёма раствора, концентраций и т.п. Как правило, анализы проводят при комнатной температуре, хотя их можно проводить в интервале температур. A. Анализы методом бесконкурентного связывания. Иммуноанализы связывания может быть или может бесконкурентного типа. В этих методах анализа непосредственно измеряется количество захваченного аналита. Например, в одном предпочтительном "сэндвич" анализе захватывающий агент (антитело или "пептитело") может непосредственно связываться с твёрдым субстратом, на котором он иммобилизуется. Эти иммобилизованные захватывающие агенты затем захватывают антиген (связываются с ним), присутствующий в испытуемом образце. Иммунобили-зованный таким образом белок затем связывается с меткой, такой как второе антитело, содержащее метку. В другом предпочтительном "сэндвич" анализе у второго антитела отсутствует метка, но оно может быть связано с меченым антителом, специфическим в отношении антител вида, к которому принадлежит второе антитело. Второе антитело также может быть модифицировано детектируемой частицей, такой как биотин, с которым может специфически связываться третья меченая молекула, такая как стрептави-дин. См. Harlow and Lane, Antibodies, A Laboratory Manual, Ch 14, Cold Spring Harbor, NY (1988), вводится в данное описание в качестве ссылки. B. Анализ методом конкурентного связывания. Иммуноанализы связывания могут быть конкурентного типа. Количество присутствующего в образце аналита определяют опосредованно, измеряя количество добавленного аналита, замещённого или конкурентно вытесненного из захватывающего агента (антитела или "пептитела") аналитом, присутствующим в образце. В одном предпочтительном варианте анализа методом конкурентного связывания известное количество аналита, как правило, меченого, добавляют к образцу, а затем образец контактирует с захватывающим агентом. Количество меченого аналита, связанного с антителом, обратно пропорцио - 39 - 008248 нально концентрации аналита, присутствующего в образце (См. Harlow and Lane, Antibodies, A Laboratory Manual, Ch 14, pp. 579-583, см. выше). В другом предпочтительном варианте анализа методом конкурентного связывания захватывающий агент иммобилизуют на твёрдом субстрате (подложке, носителе). Количество белка, связанного с захватывающим агентом, можно определить либо измеряя количество белка, присутствующего в комплексе белок/антитело, либо измеряя количество остающегося незакомплексованного белка. Количество белка можно обнаружить, делая белок меченым. Harlow and Lane, см. выше. Ещё в одном предпочтительном варианте анализа методом конкурентного связывания используют ингибирование гаптена. В данном случае известный аналит иммобилизуют на твёрдом носителе. Известное количество антитела добавляют к образцу и образец контактирует с иммобилизованным аналитом. Количество антитела, связанного с иммобилизованным аналитом, обратно пропорционально количеству аналита, присутствующего в образце. Количество иммобилизованного антитела можно определить, детектируя либо иммобилизованную фракцию антитела, либо фракцию, которая остаётся в растворе. Обнаружение может быть прямым (непосредственным), когда антитело является меченым, или опосредованным, при использовании последующего добавления меченой частицы, которая специфически связывается с антителом, как описано выше. C. Применение анализов методом конкурентного связывания. Анализы методом конкурентного связывания можно применять для определения перекрёстной реактивности, что позволит специалисту в данной области определить, является ли комплекс белка или фермента, который распознаётся "пептителом" по изобретению, заданным белком, а не перекрёстно-реактивной молекулой, или определить, является ли "пептитело" специфическим в отношении данного антигена и не связывает ли неродственные антигены. В анализах этого типа антиген может быть иммобилизован на твёрдом носителе и в аналитическую смесь добавляют смесь неизвестного белка, который конкурирует со связыванием "пептител" с иммобилизованным белком. Конкурентная молекула также связывается с одним или более антигенов, не родственных данному антигену. Способность белков конкурировать со связыванием "пептител" с иммобилизованным антигеном сравнивают со связыванием тем же самым белком, который был иммобилизован на твёрдом носителе, с целью определить перекрёстную реактивность смеси белков. D. Другие анализы связывания. Настоящее изобретение также охватывает методы Вестерн-блоттинга для обнаружения или количественного определения присутствия в образце Ang-2. Метод, как правило, включает разделение белков в образце по молекулярной массе электрофорезом в геле и перенос белков на соответствующую твёрдую подложку, такую как нитроцеллюлозный фильтр или дериватизированный найлоновый фильтр. Образец инкубируют с "пептителами" или их фрагментами, которые специфически связываются с Ang-2, и обнаруживают полученный комплекс. Эти "пептитела" могут быть мечены непосредственно или их обнаруживают потом, используя меченые антитела, которые специфически связываются с "пептителом". Диагностические методы Производные связывающие агенты, такие как пептиды и "пептитела" или их фрагменты по данному изобретению применимы для диагностики состояний или заболеваний, характеризующихся экспрессией Ang-2 или субъединиц, или при мониторинге больных, проходящих лечение с помощью индукторов Ang-2, его фрагментов, агонистов или ингибиторов Ang-2 активности. Диагностика Ang-2 включает методы, использущие "пептитело" и метку для обнаружения Ang-2 в жидкостях или клеточных экстрактах и тканях из организма человека. "Пептитела" по данному изобретению можно использовать с модификацией или без неё. В предпочтительном диагностическом анализе "пептитела" метят, присоединяя, например, метку или репортёрную молекулу. Известен большой ряд меток и репортёрных молекул, некоторые из них уже представлены в данном описании. В частности, соединения и методы по настоящему изобретению применимы для диагностики заболеваний человека. В технике известно множество протоколов для определения белков Ang-2 с использованием "пеп-тител", специфических в отношении соответствующего белка. Примеры включают твердофазный имму-ноферментный анализ (ELISA), радиоиммуноанализ (RIA, РИА) и метод, использующий лазерный анализатор (сортер, "сортировщик") клеток по интенсивности флуоресценции (FACS). Предпочтительным является двухстадийный моноклональный иммуноанализ, использующий моноклональные антитела, реактивные в отношении двух "не интерферирующих" эпитопов на Ang-2, но может использоваться конкурентное связывание. Эти методы анализа описаны, например, в Maddox et al., J. Exp. Med., 158: 1211 (1983). Для получения исходных данных ("основа") для диагностики обычно определяют нормальные или стандартные значения для экспрессии человеческого Ang-2. Это определение можно выполнять, объединяя жидкости из организма или клеточные экстракты здоровых субъектов, предпочтительно человека, с " пептителом" к Ang-2 в условиях, пригодных для образования комплекса, хорошо известных в технике. Количество образованного стандартного комплекса можно определить, сравнивая связывание "пептител" с известными количествами белка Ang-2 с контрольным образцом и образцом больного. Затем стандартные значения, получаемые из нормальных образцов, можно сравнить со значениями, полученными в образцах от субъектов, возможно, подвергнувшихся заболеванию. Отклонение от показателя стандартного - 40 - 008248 и больного субъектов наводит на мысль о том, что Ang-2 играет роль в болезненном состоянии. Для применения в диагностике в некоторых вариантах "пептитела" или пептиды по настоящему изобретению, как правило, метят детектируемой частицей. Детектируемая частица может быть любой частицей, способной давать, прямо или опосредованно, детектируемый (обнаруживаемый) сигнал. На- 3 14 32 35 125 пример, детектируемая частица может быть радиоактивным изотопом, таким как Н, С, Р, S или I, флуоресцентным или хемилюминесцентным соединением, таким как флуоресцеинизоцианат, родамин или люциферин; или ферментом, таким как щелочная фосфатаза, в-галактозидаза или пероксидаза хрена. Bayer et al., Meth. Enz., 184: 138-163 (1990). Заболевания Настоящее изобретение охватывает связывающийся агент, такой как пептид, "пептитело", или его фрагмент, вариант или производное, которые связываются с Ang-2, применимые для лечения заболеваний и патологических состояний человека. Агенты, которые модулируют активность связывания Ang-2 или другую клеточную активность, можно использовать в комбинации с другими терапевтическими агентами с целью повышения их терапевтического эффекта или уменьшения возможных побочных эффектов. В одном аспекте настоящее изобретение охватывает реагенты и методы, применимые для лечения заболеваний и состояний, характеризующихся нежелательными или аберрантными уровнями Ang-2 активности в клетке. Эти заболевания включают рак и другие гиперпролиферативные состояния, такие как гиперплазия, псориаз, контактный дерматит, иммунологические нарушения и бесплодие. Настоящее изобретение также охватывает методы лечения рака у животного, включая людей, заключающиеся во введении животному эффективного количества специфически связывающегося агента, такого как "пептитело", который ингибирует или понижает Ang-2 активность. Помимо этого, изобретение относится к методам ингибирования роста раковых клеток, включая процесс клеточной пролиферации, инвазивность и метастазы в биологической системе. Методы включают применение соединение по изобретению в качестве ингибитора роста раковых клеток. Предпочтительно методы применяются для ингибирования или уменьшения роста раковых клеток, инвазивности, метастазов или заболеваемости опухолевыми заболеваниями у животных, таких как млекопитающие. Способы по изобретению также легко адаптировать для применения в аналитических системах, например, для анализа роста раковых клеток и их свойств, а также для идентификации соединений, которые влияют на рост раковых клеток. Типы рака, которые поддаются лечению способами по данному изобретению, предпочтительно встречаются у человека. Млекопитающие включают, например, человека и других приматов, а также домашние животные, такие как собаки и кошки, подопытные животные, такие как крысы, мыши и кролики, и сельскохозяйственные животные, такие как лошади, свиньи, овцы и крупный рогатый скот. Опухоли или новообразования включают рост тканевых клеток, при котором размножение клеток является неконтролируемым и прогрессирующим. Некоторые из этих опухолей являются доброкачественными, но другие называются термином "злокачественные" и могут привести к гибели организма. Злокачественные новообразования отличаются от доброкачественных опухолей тем, что помимо проявления агрессивной клеточной пролиферации, они могут поражать окружающие ткани и метастазировать. Кроме того, злокачественные новообразования характеризуются тем, что они у них наблюдается большая утрата дифференцировки (большая дедифференцировка) и их организации относительно друг друга и окружающих их тканей. Это свойство также называется "анаплазия". Новообразования, которые поддаются лечению по данному изобретению, также включают солидные опухоли, т.е. карциномы и саркомы. Карциномы включают злокачественные новообразования из эпителиальных клеток, которые проникают в (поражают) окружающие ткани и приводят к метастазам. Аденокарциномы представляют собой такие карциномы (рак), которые образуются из гландулярных (железистых) тканей или которые образуют распознаваемые гландулярные структуры. Другую большую группу раковых заболеваний составляют саркомы, представляющие собой опухоли, клетки которых расположены ("вкраплены") в фибриллярное или гомогенное вещество, подобное эмбриональной соединительной ткани. Изобретение также делает возможным лечение рака миелоидной или лимфоидной систем, включая лейкозы, лимфомы и другие разновидности рака, которые, как правило, присутствуют не в виде опухолевой массы, но распространяются в сосудистой или лимфоретикулярной системах. Типами раковых или опухолевых клеток, поддающихся лечению по данному изобретению, являются, например, АСТН-продуцирующая опухоль, острый нелимфотический лейкоз, рак коры надпочечника, рак мочевого пузыря, рак мозга, рак молочной железы, цервикальный рак, хронический лимфолейкоз, коло-ректальный рак, кожная Т-клеточная лимфома, эндометриальный рак, эзофагеальный рак, саркома Юин-га, рак желчного пузыря, лейкемический ретикулёз, лимфома Ходжкина, саркома Калоши, рак почки, рак печени, рак лёгкого (мелкоклеточный и немелкоклеточный), плевральный выпот при злокачественном новообразовании, меланома, миеломная болезнь, нейробластома, глиома, лимфома не-Ходжкина, остео-саркома, рак яичника, рак яичника (половых клеток), рак поджелудочной железы, рак простаты, рети-нобластома, рак кожи, саркома мягких тканей, плоскоклеточный рак, тестикулярный рак, рак щитовидной железы, трофобластные новообразования, рак матки, рак влагалища, рак вульвы и аденомиосаркома. Изобретение в данном описании иллюстрируется, в частности, ссылкой на лечение некоторых ти - 41 - 008248 пов экспериментально определённого рака. В этих иллюстрирующих лечение примерах используют реальные стандартные in vitro и in vivo модели. Эти методы можно использовать для идентификации агентов, которые, предположительно, эффективны в схемах лечения in vivo. Понятно, однако, что метод по изобретению не ограничивается лечением этих типов опухолей, но распространяется на любые солидные опухоли любых органов. Рак, инвазивность и метастазирование которого обусловлено экспрессией или активностью Ang-2, особенно восприимчив к ингибированию или даже индукции регресса по способу изобретения. Изобретение можно осуществлять на практике, включая соединение по изобретению, такое как " пептитело", в комбинацию с другим противораковым химиотерапевтическим агентом, таким как обычный химиотерапевтический агент. Комбинация специфически связывающегося агента с таким другим агентом может усилить химиотерапевтическое действие (химиотерапевтический протокол). Опытный практик знает множество химиотерапевтических протоколов, которые можно включить в способ по изобретению. Можно использовать любой химиотерапевтический агент, включая алкилирующие агенты, антиметаболиты, гормоны и антагонисты, радиоактивные изотопы, а также природные продукты. Например, соединение по изобретению можно вводить с антибиотиками, такими как доксорубицин и другие аналоги антрациклина, мустины, такие как циклофосфамид, аналоги пиримидина, такие как 5-флуорацил, цисплатин, гидроксимочевина, таксол и его натуральные и синтетические производные и т.п. Другой пример: в случае смешанных опухолей, таких как аденокарцинома молочной железы, когда опухоли содержат гонадотропинзависимые и гонадотропиннезависимые клетки, соединение можно вводить в сочетании с лейпролидом или гозерелином (синтетические пептидные аналоги LH-RH). Другие анти-неопластические протоколы включают применение тетрациклинового соединения с другими возможными методами лечения, например, хирургическим вмешательством, облучением и т. д., также называемыми в данном описании "дополнительные антинеопластические методы". Таким образом, метод по изобретению можно применять с такими обычными схемами, что благоприятствует уменьшению побочных эффектов и повышению эффективности. Таким образом, изобретение охватывает составы и методы, применимые для лечения большого ряда разновидностей рака, включая солидные опухоли и лейкозы. Типы рака, которые можно лечить, включают, но без ограничения, аденокарциному молочной железы, простаты и ободочной кишки; все формы бронхогенного рака лёгкого; миелоидный рак; меланому; гепатому; нейробластому; папиллому; апудо-му; хористому; бранхиому; злокачественный карциноидный синдром; карциноидную болезнь сердца; карциному (например, карциному Уолкера, базалиому, базосквамозный рак, карциному Брауна-Пирса, дуктальную карциному, опухоль Эрлиха, опухоль Кребса-2; рак клеток Меркеля, слизистый рак, немел-коклеточный рак лёгкого, овсяно-клеточный рак, папаллярный рак, скиррозный рак, бронхиолярный рак, бронхогенный рак, плоскоклеточный и переходно-клеточный рак); гистиоцитарные нарушения; лейкоз; злокачественный гистиоцитоз; болезнь Ходжкина; иммунопролиферативный мелкоклеточный рак лёгкого; неходжкинскую лимфому; плазмацитому; ретикулёз; меланому; хондробластому; хондрому; хондро-саркому; фибросаркому; гигантоклеточные опухоли; гистиоцитому; липому; липосаркому; мезотелиому; миксому; остеому; остеосаркому; хордому; краниофарингиому; дисгерминому; гемартому; мезенхимому; мезонефрому; миосаркому; амелобластому; цементому; одонтому; тератому; тимому; "тофобластную" (tophoblastic) опухоль. Кроме того, можно лечить следующие типы рака; аденому; холангиому; холестеа-тому; циклиндрому; цистаденокарциному; цистаденому; зернистоклеточная опухоль (яичника); андроб-ластому; гепатому; гидраденому; островковоклеточную опухоль; опухоль клеток Лейдига; папиллому; опухоль клеток Сертоли; опухоль клеток слизистой оболочки желудка; лейомиому; лейомиосаркому; миобластому; миому; миосаркому; рабдомиому; рабдомиосаркому; эпендимому; ганглионейрому; глиому; медуллобластому; менингиому; нейрилеммому; нейробластому; нейроэпителиому; нейрофиброму; нейрому; параганглиому; хемадектому; ангиокератому; ангиолимфоидную гиперплазию с эозинофилией; склерозирующую ангиому; ангиоматоз; гломангиому; гемангиоэндотелиому; гемангиому; гемангиопе-рицитому; гемангиосаркому; лимфангиому; лимфангиомиому; лимфангиосаркому; пинеалому; карцино-саркому; хондрасаркому; филоидную цистосаркому; фибросаркому; гемангиосаркому; лейомиосаркому; лейкосаркому; липосаркому; лимфангиосаркому; миосаркому; миксосаркому; карциному яичников; раб-домиосаркому; саркому; новообразования; нейрофиброматоз и цервикальную дисплазию. В другом аспекте настоящее изобретение охватывает применение материалов и способов по данному изобретению для предупреждения и/или лечения гиперпролиферативного состояния кожи, включая псориаз и контактный дерматит или другие гиперпролиферативные заболевания. Было показано, что у больных псориазом и контактным дерматитом наблюдается повышается Ang-2 активность в местах этих поражений. [Ogoshi et al., J. Inv. Dermatol., 110: 818-23 (1998]). Предпочтительно, чтобы специфически связывающиеся агенты, специфичные в отношении Ang-2, применялись в комбинации с другими фармацевтическими агентами для лечения людей с подобными клиническими симптомами. Специфически связывающиеся агенты можно доставлять с помощью любого носителя, вводя различными способами по данному описанию и другими способами, хорошо известными специалистам в данной области техники. Другие аспекты настоящего изобретения включают лечение различных ретинопатий (включая диабетическую ретинопатию и старческую дегенерацию жёлтого пятна), которые протекают с участием ан - 42 - 008248 гиогенеза, а также нарушений и/или заболеваний женских половых путей, таких как эндометриоз, фиброз матки и другие подобные состояния, обусловленные нарушением сосудистой пролиферации (включая микроваскулярный эндометриальный рост) во время репродуктивного цикла у женщин. Ещё один аспект настоящего изобретения относится к лечению аномального микроваскулярного роста, включая артериовенозные пороки развития (AVM), поражение и заживление слизистой желудочно-кишечного тракта; изъязвление слизистой желудочно-кишечного тракта с анамнезом язвенной болезни желудка, включая ишемию вследствие удара; широкий спектр сосудистых нарушений в лёгких при заболевании печени и портальную гипертензию у больных с непечёночной портальной гипертензией. Другим аспектом настоящего изобретения является предупреждение рака с применением составов и способов, охватываемых данным изобретением. Такие реагенты включают специфически связывающиеся агенты, такие как "пептитела" против Ang-2. Фармацевтические композиции Фармацевтические композиции агентов, специфически связывающихся с Ang-2, таких как "пептитела", входят в объём данного изобретения. Фармацевтические композиции, содержащие антитела, подробно описаны, например, в патенте США 6171568, принадлежащем Lam et al., выданном 9 января 2001 г. Такие композиции содержат терапевтически или профилактически эффективное количество специфически связывающегося агента, такого как антитело или его фрагмент, вариант, производное или слияние по данному описанию, в смеси с фармацевтически приемлемым агентом. В предпочтительном варианте изобретения фармацевтические композиции содержат антагонистические специфически связывающиеся агенты, которые модулируют, частично или полностью, по меньшей мере одну биологическую активность Ang-2, в смеси с фармацевтически приемлемым агентом. Как правило, специфически связывающиеся агенты являются в достаточной степени очищенными, чтобы их можно было вводить животному. Фармацевтическая композиция может содержать входящие в рецептуру материалы для модификации, поддержания или сохранения, например, рН, осмомолярности, вязкости, прозрачности, цвета, изо-тоничности, запаха, стерильности, стабильности, скорости диффузии или выделения, адсорбции или проникновения композиции. Подходящие вещества для рецептуры включают, но без ограничения, аминокислоты (такие как глицин, глутамин, аспарагин, аргинин или лизин); антимикробные вещества; анти-оксиданты (такие как аскорбиновая кислота, сульфит натрия или гидросульфит натрия); буферные вещества (такие как борат, бикарбонат, Tris-HCl, фосфаты, цитраты, другие органические кислоты); наполнители (такие как маннит или глицин), хелатирующие агенты [такие как этилендиаминтетрауксусная кислота (EDTA)]; комплексообразующие агенты (такие как кофеин, поливинилпирролидон, бета-циклодекстрин или гидроксипропил-бета-циклодекстрин); моносахариды; дисахариды и другие углеводы (такие как глюкоза, манноза или декстрины); белки (такие как сывороточный альбумин, желатин или иммуноглобулины); красители; ароматизаторы и разбавители; эмульгаторы; гидрофильные полимеры (такие как поливинилпирролидон); низкомолекулярные полипептиды; солеобразующие противоионы (такие как натрий); консерванты (такие как бензалкония хлорид, бензойная кислота, салициловая кислота, тимерозаль, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексин, сорбиновая кислота или перекись водорода); растворители (такие как глицерин, пропиленгликоль или полиэтиленгликоль); спирты сахаров (такие как маннит или сорбит); суспендирующие агенты; поверхностно-активные вещества или увлажняющие агенты (такие как плуроники, ПЭГ, эфиры сорбитана, полисорбаты, такие как поли-сорбат 20, полисорбат 80, тритон, трометамин, лецитин, холестерин, тилоксапаль); агенты, повышающие стабильность (сахароза или сорбит); тонизирующие агенты (такие как галогениды щелочных металлов (предпочтительно хлорид натрия или калия), маннит, сорбит); носители для доставки; разбавители; экс-ципиенты и/или фармацевтические адъюванты (Remington's Pharmaceutical Sciences, 18th edition, A.R. Gennaro, ed., Mack Publishing Company, 1990). Оптимальную фармацевтическую композицию определяет специалист в данной области техники, например, в зависимости от предполагаемого способа введения, формата доставки и желательной дозировки. См., например, выше: Remington's Pharmaceutical Sciences. Такие композиции могут влиять на физическое состояние, стабильность, скорость in vivo выделения и скорость in vivo клиренса специфически связывающегося агента. Основной наполнитель или носитель в фармацевтической композиции может быть либо водным, либо неводным по природе. Например, подходящий наполнитель или носитель может представлять собой воду для инъекции, физиологический раствор или искусственную спинномозговую жидкость, возможно, дополненную другими веществами, общепринятыми в композициях для парентерального введения. Физиологический раствор с нейтральным буфером или физиологический раствор в смеси с сывороточным альбумином является ещё одним примером носителя. Другие примеры фармацевтических композиций включают буфер Tris с примерным рН 7,0-8,5, или ацетатный буфер рН около 4,0-5,5, который может дополнительно включать сорбит или его соответствующий заместитель. В одном варианте настоящего изобретения можно получать композиции со связывающимся агентом для хранения, смешивая выбранную композицию с заданной степенью чистоты с необязательными агентами, входящими в рецептуру (Remington's Pharmaceutical Sciences, см. выше), в форме лиофилизированной лепёшки или водного раствора. Кроме того, продукт со связывающимся агентом можно готовить в виде лиофилизата, исполь - 43 - 008248 зуя подходящие эксципиенты, такие как сахароза. Фармацевтические композиции можно выбирать для парентеральной доставки. Или же композиции можно выбирать для ингаляции или для энтерального способа доставки, такого как оральный; для аураль-ного, офтальмического, ректального или вагинального способа доставки. Приготовление таких фармацевтически приемлемых композиций находится в компетенции специалистов в данной области техники. Компоненты рецептуры присутствуют в концентрациях, которые приемлемы для данного места введения. Например, буферы применяют для поддержания композиции при физиологических значениях рН или при немного более низких значениях рН, как правило, в примерном интервале 5-8. Если предполагается парентеральное введение, терапевтические композиции для применения по данному изобретению может быть в форме не содержащего пироген парентерально приемлемого водного раствора, содержащего заданный специфически связывающийся агент в фармацевтически приемлемом носителе. Особенно предпочтительным носителем для парентеральной инъекции является стерильная дистиллированная вода, в которой связывающийся агент приготовлен в виде стерильного изотонического раствора, соответствующим образом сохраняемого. Ещё один препарат может включать рецептуру заданной молекулы с агентом, таким как инъецируемые микросферы, биоразрушаемые частицы, полимерные соединения (полимолочная кислота, полигликолевая кислота), бусины или липосомы, обеспечивающие контролированное или пролонгированное выделение продукта, который можно затем доставлять с помощью депо-инъекции. Можно также использовать гиалуроновую кислоту, и это может повлиять на промотирование пролонгированного пребывания в кровотоке. Другие подходящие способы введения заданной молекулы включают имплантированные устройства для доставки лекарственного препарата. В другом аспекте фармацевтические препараты, пригодные для парентерального введения, можно готовить в водных растворах предпочтительно в физиологически совместимых буферах, таких как раствор Хэнкса, Рингеровский раствор или забуференный физиологический раствор. Водные суспензии для инъекций могут содержать вещества, которые повышают вязкость суспензии, например, натрий-карбоксиметилцеллюлозу, сорбит или декстран. Или же суспензии активных соединений можно получать в виде соответствующих масляных суспензий для инъекций. Подходящие липофильные растворители или носители включают жирные масла, такие как кунжутное масло, или синтетические сложные эфи-ры жирных кислот, такие как этилолеат, триглицериды, или липосомы. Для доставки можно также применять нелипидные поликатионные полимерные амины. Необязательно, суспензия может также содержать подходящие стабилизаторы или агенты для повышения растворимости соединений, что позволяет готовить препараты высокой концентрацией этих веществ в растворах. В другом варианте изобретения фармацевтическую композицию можно готовить для доставки методом ингаляции. Например, связывающийся агент можно приготовить в виде сухого порошка для ингаляции. Раствор полипептида или нуклеиновой кислоты можно также готовить с диспергатором для доставки в виде аэрозолей. Ещё в одном варианте изобретения растворы можно для распыления. Кроме того, доставка в лёгкие химически модифицированных белков описана в международной заявке PCT/US 94/001875. Рассматривается также, что некоторые препараты можно вводить перорально. В одном варианте настоящего изобретения молекулы связывающегося агента, которые вводятся таким способом, можно приготовить с такими носителями (или без них), которые обычно применяют для приготовления твёрдых лекарственных форм, таких как таблетки и капсулы. Например, можно создать капсулу для высвобождения активного компонента препарата в точке желудочно-кишечного тракта, в которой биодоступность является максимальной, а предсистемное расщепление минимально. Для облегчения всасывания связывающегося агента можно включать дополнительные агенты. Можно применять также разбавители, вещества, придающие запах и вкус, низкоплавкие воски, растительные масла, смазывающие вещества, суспендирующие агенты, вещества, способствующие измельчению, и связующие. Можно готовить также фармацевтические композиции для (пер)орального введения, используя фармацевтически приемлемые носители, хорошо известные в технике, в дозах, подходящих для орального введения. Такие носители способствуют тому, что фармацевтические композиции можно готовить в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, эмульсий, суспензий и т.п., проглатываемых больным. Фармацевтические препараты для орального применения можно получать, объединяя активные соединения с твёрдым эксципиентом и готовя из полученной смеси гранул (необязательно, после измельчения) таблетки или ядра драже. Если требуется, можно вводить подходящие добавки. Применимые эксципиенты включают углеводные или белковые наполнители, такие как сахара, включая лактозу, сахарозу, маннит и сорбит; маисовый крахмал, пшеничный крахмал, рисовый крахмал, картофельный крахмал или другой растительный крахмал; целлюлозу, такую как метилцеллюлоза, гидрокси-пропилметилцеллюлоза или натрий-карбоксиметилцеллюлоза; камедь, включая аравийскую камедь и камедь трагаканта; и белки, такие как желатин и коллаген. При желании можно добавлять вещества, способствующие измельчению, или солюбилизирующие вещества, такие как сшитый поливинилпирроли-дон, агар-агар и альгиновая кислота или её соль, такая как альгинат натрия. Ядра драже можно применять в сочетании с подходящими покрытиями (оболочками), такими как концентрированные растворы сахаров, которые могут содержать также аравийскую камедь, тальк, поли - 44 - 008248 винилпирролидон, карбопол гель, полиэтиленгликоль и/или диоксид титана, растворы лаков и подходящие органические растворители или смеси растворителей. Для идентификации продукта или для характеристики количества активного соединения, например, дозировки, в покрытие таблеток или драже можно добавлять красители или пигменты. Фармацевтические препараты, которые можно применять перорально, также включают солидные желатиновые капсулы, а также мягкие герметичные капсулы из желатина, и покрытия, такого как глицерин или сорбит. Твёрдые капсулы могут содержать активные ингредиенты, смешанные с наполнителями или связующими, такими как лактоза или крахмал, смазки, такие как тальк или стеарат магния, и, необязательно, стабилизаторы. В мягких капсулах активные соединения можно растворить или суспендировать в подходящих жидкостях, таких как жирные масла, жидкость или жидкий полиэтиленгликоль со стабилизаторами или без них. Другая фармацевтическая композиция может включать эффективное количество связывающегося агента в смеси с нетоксическими эксципиентами, пригодными для производства таблеток. Стандартные дозы растворов можно приготовить, растворяя таблетки в стерильной воде или в другом подходящем носителе. Соответствующие эксципиенты включают, но без ограничения, инертные разбавители, такие как карбонат кальция, карбонат или бикарбонат натрия, лактоза или фосфат кальция; или связующие, такие как крахмал, желатин или аравийскую камедь; или смазки, такие как стеарат магния, стеариновая кислота или тальк. Дополнительные фармацевтические композиции очевидны для специалистов в данной области техники, включая рецептуры, содержащие связывающиеся агенты в препаратах пролонгированного или регулируемого (контролируемого) действия. Способы приготовления составов для доставки многих других препаратов пролонгированного или контролируемого действия, таких как липосомные носители, биораз-рушаемые микрочастицы или пористые бусины и депо-инъекции, хорошо известны специалистам в данной области техники. См., например, международную заявку PCT/US 93/00829, в которой описано контролируемое выделение из пористых полимерных микрочастиц для доставки фармацевтических композиций. Дополнительные примеры препаратов пролонгированного действия включают полупроницаемые полимерные матрицы в форме имеющих определённые очертания изделий, например, плёнок или микрокапсул. Матрицы (носители) пролонгированного действия могут включать полиэфиры, гидрогели, по-лилактиды (патент США 3773919, европейский патент 58481), сополимеры L-глутаминовой кислоты и гамма этил^-глутамат [Sidman et al., Biopolymers, 22: 547-556 (1983)], поли(2-гидроксиэтилметакрилат) [Langer et al., J. Biomed. Mater. Res., 15:167-277, (1981)] и [Langer et al., Chem. Tech., 12: 98-105(1982)], этиленвинил ацетат (Langer et al., см. выше) или поли-ф)(-)-3-гидроксимасляную кислоту (европейский патент 133988). Композиции пролонгированного действия также включают липосомы, которые можно получать любым из методов, известных в технике. См., например, Eppstein et al., Proc. Natl. Acad. Sci. (USA), 82: 3688-3692 (1985); европейские патенты ЕР 36676; ЕР 88046; ЕР 143949. Фармацевтическая композиция, применяемая для in vivo введения, как правило, должна быть стерильной. Этого можно достичь фильтрованием через стерильные фильтрующие мембраны. Если композицию лиофилизируют, стерилизацию с применением этого методы можно осуществлять либо до, либо после лиофилизации и восстановления. Композицию для парентерального введения можно хранить в лиофилизированной форме или в виде раствора. Кроме того, парентеральные формы композиции, как правило, помещают в контейнер, имеющий стерильное входное отверстие, например, контейнер или флакон с пробкой, через которую может проникнуть гиподермическая игла для инъекций. После приготовления фармацевтической композиции её можно хранить в стерильных ампулах (флаконах) в виде раствора, суспензии, геля, эмульсии, твёрдого вещества или в виде дегидратированного или лиофилизированного порошка. Такие препараты могут храниться либо в виде готовой формы, либо в виде формы (например, лиофилизированной), требующей восстановления перед употреблением. В конкретном варианте настоящее изобретение относится к наборам для приготовления дозы для одноразового введения (одноразовой дозы). Каждый набор может включать как первый контейнер, содержащий сухой белок, и второй контейнер, содержащий водный препарат (препарат в воде). В объём настоящего изобретения также входят наборы, содержащие многокамерные предварительно заполненные шприцы (например, шприцы с жидкостями и "лиошприцы" (шприцы с лиофилизатом)). Эффективное количество фармацевтической композиции для терапевтического применения зависит, например, от терапевтического контекста и от задач. Специалист в данной области техники понимает, что, следовательно, соответствующие уровни доз для лечения варьируют в зависимости, частично, от доставляемой молекулы, показания, в соответствии с которым применяется связывающийся агент, способа применения и размеров (веса тела, площади поверхности тела и величины органа) и состояния (возраст и общее состояние здоровья) пациента. Соответственно, клиницист может определить (рассчитать) дозу и модифицировать способ применения, чтобы получить оптимальный терапевтический эффект. Типичная доза обычно находится в пределах, примерно, от 0,1 вплоть до 100 мг/кг или выше в зависимости от вышеуказанных факторов. В других вариантах изобретения интервалы доз могут составлять, примерно, 0,1-100 мг/кг; или 1-100 мг/кг; или 5-100 мг/кг. - 45 - 008248 Для любого соединения терапевтически эффективную дозу можно сначала определить либо анализом клеточных культур, либо на животных моделях, таких как мыши, крысы, кролики, собаки, свиньи или мартышки. Животные модели можно также использовать для определения подходящего интервала и способа применения. Эту информацию можно использовать для определения применяемых доз и способов применения для лечения людей. Точную дозировку определяют, принимая во внимание факторы, связанные с субъектом, подлежащим лечению. Дозы и путь введения корректируют, чтобы обеспечить достаточные уровни активного соединения или для поддержания заданного эффекта. Факторы, которые можно принимать во внимание, включают тяжесть болезненного состояния, общее состояние здоровья субъекта, возраст, вес и пол субъекта, время и частоту введения, комбинацию(-и) лекарственных веществ, чувствительность и реакция на лечение. Фармацевтические композиции пролонгированного действия можно вводить каждые 3-4 дня, раз в неделю или раз в 2 недели, в зависимости от периода полужизни и скорости клиренса конкретного препарата. Частота введения дозы зависит от фармакокинетических показателей связывающегося агента в применяемой композиции. Как правило, композицию вводят до тех пор, пока не достигнут дозы, дающей нужный эффект. Следовательно, композицию можно вводить в виде однократной (разовой) дозы или в виде многократных доз (в тех же самых или различных концентрациях/ дозу) во времени, или в виде непрерывных вливаний. Дальнейшее уточнение подходящей дозы делается обычными методами. В том, что доза является подходящей, можно удостовериться с помощью данных об эффекте дозы. Способ применения фармацевтической композиции представляет собой известный метод, например, оральный, внутривенная инъекция, интраперитонеальный, интрацеребральный (интрапаренхима-тозный), интрацеребровентрикулярный, внутримышечный, внутриглазной, внутриартериальный, интра-портальный, интралезиональный (в место поражения), интрамедуллярный, внутриоболочечный, интра-вентрикулярный, трансдермальный, подкожный, интраперитонеальный, интраназальный, энтеральный, местный, сублингвальный, уретральный, вагинальный или ректальный способы с помощью систем пролонгированного действия или имплантированных устройств. Если требуется, композиции можно вводить с помощью болюсной инъекции или непрерывно, с помощью вливания или с помощью имплантированного устройства. Или же, или в дополнение к сказанному, композицию по изобретению можно вводить местно, имплантируя мембрану, тампон или другой подходящий материал, на котором абсорбировано или в который инкапсулировано нужное вещество. Если используется устройство для имплантации, прибор можно имплантировать в любую подходящую ткань или в любой подходящий орган, и доставка молекул заданного вещества может осуществляться диффузией, болюсом пролонгированного действия (с выделением во времени) или непрерывным введением. Иногда может быть желательным применять фармацевтические композиции ех vivo. В таких случаях клетки, ткани или органы удаляют из организма больного и экспонируют с фармацевтическими композициями, после чего клетки, ткани и/или органы последовательно имплантируют обратно, в организм пациента. В других случаях связывающийся агент по данному изобретению, такой как "пептитело", можно доставлять, имплантируя определённые клетки, полученные с помощью генной инженерии (методом рекомбинантной ДНК), такими, которые представленные в данном описании, для экспрессии и секреции полипептида. Такие клетки могут быть животными или человеческими клетками и могут быть аутоло-гичными, гетерологичными или ксеногенными. Необязательно, клетки могут быть иммортализованными. Чтобы понизить вероятность иммунного ответа, клетки можно инкапсулировать, чтобы избежать инфильтрации окружающих тканей. Материалы для инкапсулирования, как правило, представляют собой биосовместимые, полупроницаемые полимерные капсулы или мембраны, которые делают возможным выделение протеинового(-ых) продукта (-ов), но предотвращают разрушение клеток под действием иммунной системы больного или других вредных факторов из окружающих тканей. Комбинированная терапия Специфически связывающиеся агенты по изобретению, такие как "пептитела", можно применять в комбинации с другими терапевтическими средствами лечения болезней, обусловленных экспрессией Ang-2. Эти другие терапевтические методы включают, например, облучение, химиотерапию и нацеленные терапевтические средства, такие как Herceptin(tm), Rituxan(tm), Gleevec(tm) и т.п. Дополнительные варианты комбинированной терапии, не указываемые конкретно, также входят в объём настоящего изобретения. В химиотерапевтическом лечении можно использовать противоопухолевые агенты, в том числе, например, алкилирующие агенты, включающие: мустины, такие как мехлорэтамин, циклофосфамид, ифосфамид, мелфалан и хлорамбуцил; нитрозомочевины, такие как кармустин (BCNU), ломустин (CCNU) и семустин (метил-CCNU); этиленимины/метилмеламин, такой как триэтиленмеламин (ТЕМ), триэтилен, трифосфорамид (тиотепа), гексаметилмеламин (НММ, альтретамин); алкилсульфонаты, такие как бусульфан; триазины, такие как дакарбазин (DTIC); антиметаболиты, включая аналоги фолиевой кислоты, такие как метотрексат и триметрексат, аналоги пиримидина, такие как 5-флуорурацил, флуорде-зоксирубицин, гемцитабин, цитозин арабинозид (АгаС, цитарабин), 5-азатидин, 2,2'-дифлуордезокси - 46 - 008248 цитидин, аналоги пурина, такие как 6-меркаптопурин, 6-тиогуанидин, азатиоприн, 2'-дезоксикоформи-цин (пентостатин), эритрогидроксинониладенин (EHNA), флударабин фосфат и 2-хлордезоксиаденозин (кладрибин, 2-CdA); природные продукты, включая антимитотические лекарственные средства, такие как паклитаксель, алкалоиды винка, включая винбластин (VLB), винкристин и винорелбин, таксотере, эстрамустин и эстрамустин фосфат; пиподофилотоксины, такие как этопозид и тенипозид; антибиотики, такие как актимомицин D, дауномицин (рубидомицин), доксорубицин, митоксантрон, идарубицин, блео-мицины, пликамицин (митрамицин), митомицин С и актиномицин; ферменты, такие как L-аспарагиназа; модификаторы биологического ответа, такие как интерферон-альфа, IL-2, G-CSF и GM-CSF; различные агенты, включая координационные платиновые комплексы, такие как цисплатин и карбоплатин, антра-цендионы, такие как митоксантрон, замещённые мочевины, такие как гидроксимочевина, производные метилгидразина, включая N-метилгидразин (МН) и прокарбазин, адренокортикальные супрессоры, такие как митотан (o,p'-DDD) и аминоглютэтимид; гормоны и антагонисты, включая адренокортикостероидные антагонисты, такие как преднизон и эквиваленты, дексаметазон и аминоглютэтимид; прогестин, такой как гидропрогестерона капронат, метилпрогестерон ацетат и мегестрол ацетат; эстроген, такой как ди-этилстильбестрол и эквиваленты этинилэстрадиола; антиэстроген, такой как тамоксифен; андрогены, включая тестостерона пропионат и флуоксиместерон/эквиваленты; антиандрогены, такие как флутамид, аналоги гонадолиберина и лейпролид; и нестероидные антиандрогены, такие как флутамид. Комбинированная терапия с применением факторов роста может включать цитокины, лимфокины, факторы роста или другие гемопоэтические факторы, такие как M-CSF, GM-CSF, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, IFN, TNF0, TNF1, TNF2, G-CSF, Meg-CSF, GM-CSF, тромбопоэтин, фактор стволовых клеток и эритропоэтин. Другие композиции могут включать известные ангиопоэтины, например, Ang-1, -2, -4, -Y и/или человеческий Ang-подобный полипептид и/или сосудистый эндотелиальный фактор роста (VEGF). Факторы роста включают ангиогенин, костный морфогенный белок-1, костный морфогенный белок-2, костный морфогенный белок-3, костный морфогенный белок-4, костный морфогенный белок-5, костный морфогенный белок-6, костный морфогенный белок-7, костный морфогенный белок-8, костный морфогенный белок-9, костньш морфогенный белок-10, костный морфогенный белок-11, костный морфогенный белок-12, костный мор-фогенный белок-13, костный морфогенный белок-14, костный морфогенный белок-15, рецептор IA костного морфогенного белка, рецептор IB костного морфогенного белка, нейротрофический фактор мозга, рецептор цилиарного нейротрофического фактора, индуцируемый цитокинами хемотактический фактор 1 нейтрофилов, индуцируемый цитокинами хемотактический фактор 2 нейтрофилов, фактор роста эндо-телиальных клеток, эндотелин 1, эпидермальный фактор роста, эпителиальный аттрактант нейтрофилов, фактор роста фибробластов 4, фактор роста фибробластов 5, фактор роста фибробластов 6, фактор роста фибробластов 7, фактор роста фибробластов 8, фактор роста фибробластов 8b, фактор роста фибробла-стов 8с, фактор роста фибробластов 9, фактор роста фибробластов 10, кислый фактор роста фибробла-стов, щелочной фактор роста фибробластов; рецептор-1 нейротрофического фактора глиальной клеточной линии; рецептор-2 нейротрофического фактора глиальной клеточной линии; ростовой белок, ростовой белок-1, ростовой белок-2, ростовой белок-3, гепарин-связывающий эпидермальный фактор роста, фактор роста гепатоцитов, рецептор фактора роста гепатоцитов, инсулиноподобный фактор роста I, рецептор инсулиноподобного фактора роста, инсулиноподобный фактор роста II; белок, связывающий ин-сулиноподобный фактор роста, фактор роста кератиноцитов; фактор, ингибирующий лейкоз; рецептор-1 фактора, ингибирующего лейкоз; фактор роста нервных клеток, рецептор фактора роста нервных клеток, нейротрофин-3, нейротрофин-4, плацентарный фактор роста, плацентарный фактор роста 2; полученный из тромбоцитов фактор роста эндотелиальных клеток; полученный из тромбоцитов фактор роста цепи А; полученный из тромбоцитов фактор роста АВ; полученный из тромбоцитов фактор роста цепи В; полученный из тромбоцитов фактор роста ВВ, рецептор-1 полученного из тромбоцитов фактора роста; рецеп-тор-2 полученного из тромбоцитов фактора роста; фактор стимуляции (роста) пре-В клеток; фактор стволовых клеток; рецептор фактора стволовых клеток; трансформирующий фактор роста-1, трансформирующий фактор роста-2, трансформирующий фактор роста-1.2, трансформирующий фактор роста-3, трансформирующий фактор роста-5, латентный трансформирующий фактор роста-1; белок I, связывающий трансформирующий фактор роста-1; белок II, связывающий трансформирующий фактор роста-1; белок III, связывающий трансформирующий фактор роста-1; рецептор типа I фактора некроза опухоли; рецептор типа II фактора некроза опухоли; рецептор плазминогенного активатора урокиназного типа; васкулярный эндотелиальный фактор роста и химерные белки и их биологически и иммунологически активные фрагменты. Иммунотерапевтические средства Иммунотерапевтические методы лечения, как правило, основаны на использовании иммунных эф-фекторных клеток и молекул для нацеливания на раковые клетки и их разрушение. Иммунные эффекторы могут представлять собой, например, "пептитело" по данному изобретению, которое распознаёт некий маркёр на поверхности клетки-мишени. "Пептитело" само по себе может служить в качестве эффектора в терапии или оно может рекрутировать другие клетки для того, чтобы фактически осуществить лизис клеток. "Пептитело" может быть также конъюгировано с лекарственным веществом или токсином - 47 - 008248 (химиотерапевтическим соединением, радионуклидом, А цепью рицина, холерным энтеротоксином, токсином коклюша и т. п.) и, следовательно, может просто служить в качестве целевого агента. В соответствии с данным изобретением мутантные формы Ang-2 могут служить целью для иммунотерапии с помощью "пептител" или конъюгатов "пептител" по изобретению. В частности предполагается, что составы "пептител" по изобретению можно применять в комбинированной терапии в сочетании с Ang-2 нацеленной терапией. Оказалось, что пассивная терапия является особенно эффективной при лечении различных типов рака. См., например, международную заявку WO98/39027. Нижеприведённые примеры предназначены только для целей иллюстрации и ни в коей мере не являются ограничивающими объём изобретения. Пример 1. Экспрессия Ang-2 в патологической и нормальной ткани. Экспрессию Ang-2 изучают в нормальной и патологической ткани in situ гибридизацией. Фрагменты последовательностей человеческого (Genbank Accession Number AF004327, нуклеотиды 1274-1726) и мышиного (Genbank Accession Number AF004326, нуклеотиды 1135-1588) Ang-2 амплифицируют ПЦР с обратной транскриптазой при использовании фетальной кДНК из образцов лёгкого, клонированной в плазмиду pGEM-T, и проверяют секвенированием. Зонды меченой 33Р антисмысловой РНК транскрибируют при использовании линеаризованной плазмиды с 33-UTP и РНК полимеразой. Из блоков фиксированных в формальдегиде заключённых в парафин тканей делают срезы по 5 мкм и собирают на подготовленные предметные стёкла. Перед in situ гибридизацией с помощью 0,2М Hd получают пермеабили-зованные ткани с последующими расщеплением протеиназой K и ацетилированием триэтаноламином и уксусным ангидридом. Срезы гибридизуют с меченым радиоактивной меткой зондом в течение ночи при 55°С, затем расщепляют с помощью РНК-азы и отмывают в очень жёстких условиях, примерно, в 0.1 X SSC при 55°С. Предметные стёкла погружают в эмульсию Kodak NTB2, экспонируют при 4°С в течение 2-3 недель, проявляют и окрашивают контрастным веществом. Срезы изучают в темноте и при стандартном освещении с целью одновременной оценки морфологии ткани и гибридизационного сигнала. Результаты показывают, что у нормального новорождённого человека экспрессия Ang-2 ограничена несколькими тканями, содержащими ангиогенную сосудистую сеть, такими как яичник, плацента и матка. Никакой экспрессии Ang-2 не отмечается в сердце, мозгу, почках, печени, лёгких, поджелудочной железе, селезёнке, мышцах, миндалинах, тимусе, аппендиксе, лимфатическом узле, желчном пузыре, простате или яичках нормального взрослого человека. У пятинедельной мыши (но не у взрослой обезьяны или взрослого человека) почки выявляют заметную экспрессию Ang-2 в семенных канальцах. Чтобы определить, является ли эта экспрессия остатком развития зародыша, этот эксперимент повторяют на почках мышей в возрасте до 1 года при использовании зонда мышиного Ang-2 и в условиях, описанных выше. Выяснено, что экспрессия Ang-2 понижается в период неонатального развития, но всё ещё явственна в почках годовалых мышей. Экспрессия Ang-2 также обнаружена практически во всех изученных типах опухолей, включая первичные человеческие опухоли, такие как рак прямой (ободочной) кишки (5 случаев), рак молочной железы (10 случаев), рак лёгкого (8 случаев), глиобластома (1 случай), метастатические человеческие опухоли, такие как рак молочный железы (2 случая), рак лёгких (2 случая) и рак яичника (2 случая), который метастазировал в мозг, и модели опухолей у грызунов, такие как С6 (глиома у крыс), НТ29 (человеческий рак прямой (ободочной) кишки), Colo-205 (человеческий рак прямой (ободочной) кишки), А431 (человеческий эпидермоидный рак), А673 (человеческая рабдомиосаркома), НТ1080 (человеческая фиб-росаркома), РС-3 (человеческий рак простаты), B16F10 (меланома у мышей), MethA (саркома у мышей) и метастатический (mets) рак лёгкого Льюиса. Кроме того, экспрессия Ang-2 обнаружена в неососудах, растущих в матриксе (Матригель) в ответ на VEGF и у мышиной гипоксической модели ретинопатии у недоношенных. Пример 2. Молекулярные анализы для определения Ang-2 "пептител". Молекулярные анализы (аффинный метод ELISA, нейтрализационный метод ELISA, BIAcore) применяются для оценки непосредственного связывания пептитела с Ang-2 и представителями родственных семейств и влияния антител на взаимодействие Ang-2:Tie-2. Эти in vitro анализы описаны ниже. Аффинный метод ELISA Для начального скрининга возможных anra-Ang-2 "пептител" используют очищенный человеческий Ang-2 (R &D Systems, Inc.; номер в каталоге 623-AN; Ang-2 в виде смеси 2 усечённых вариантов) или мышиный Ang-2 полипептид (полученный, как описано выше). Для подтверждающих анализов связывания человеческий Ang-2 получают при использовании кондиционированных сред человеческих 293Т клеток, трансфицированных с помощью ДНК полноразмерного человеческого Ang-2 и культивируют в бессывороточной среде Игла, модифицированной по Дульбекко (DMEM), содержащей 50 мкг в мл альбумина бычьей сыворотки (BSA). В каждую лунку титрационного микропланшета добавляют 100 мкл Ang-2 и планшеты инкубируют в течение 2 ч, после чего планшеты 4 раза отмывают фосфатно-солевым буферным раствором (PBS), содержащим 0,1% Tween-20. Затем лунки блокируют, используя около 250 мкл на лунку 5% BSA в PBS и планшеты инкубируют при комнатной температуре, примерно, в течение 2 ч. После инкубации избыток - 48 - 008248 блокирующего раствора отбрасывают и около 100 мкл каждого возможного affra-Ang-2 "пептитела" добавляют в лунку, используя метод серийных разведений, начиная, примерно, с 40 наномолярной концентрации, а затем серийно разводят в 4 раза в PBS, содержащем 1% BSA. Планшеты затем инкубируют в течение ночи при комнатной температуре. После инкубации планшеты отмывают PBS, содержащем около 0,1% Tween-20. Отмывание повторяют ещё 4 раза, после чего добавляют около 100 мкл/лунка антитела козы к человеческому IgG(Fc)-HRP (Pierce Chemical Co., № в каталоге 31416), предварительно разведённого 1:5000 в PBS, содержащем 1% BSA. Планшеты инкубируют около 1 ч при комнатной температуре. Затем планшеты 5 раз отмывают в PBS, содержащем около 0,1% Tween-20, после чего в каждую лунку добавляют по 100 мкл субстрата ТМВ (система жидкого субстрата 3,3',5,5'-тетраметилбензидина; Sigma Chemical Company, St. Louis, MO, № в каталоге Т8665) и планшеты инкубируют около 5-15 мин до появления синего цвета. Затем оптическую плотность считывают на спектрофотометре (планшетном ри-дере), примерно, при 370 нм. Метод ELISA с нейтрализацией Микротитационные планшеты, с которыми был связан человеческий Ang-2 полипептид, готовят как описано для аффинного метода ELISA. Предполагаемые affra-Ang-2 "пептитела" титруют от 1000 нМ до 0,2 пМ в 4-кратном разведении в растворе PBS, содержащем около 1% BSA и около 1нМ Tie-2 (при условии, что молекула Tie-2-Fc представляет собой молекулу, в которой участок Tie-2 содержит только растворимую внеклеточную область молекулы; R &D Systems, № в каталоге 313-TI). После того, как в каждую лунку добавляют около 100 мкл раствора, планшеты инкубируют в течение ночи при комнатной температуре, а затем 5 раз отмывают в PBS, содержащем около 0,1% Tween-20. После отмывания в каждую лунку добавляют около 100 мкл антитела против Tie-2 (Pharmingen Inc., № в каталоге 557039) до примерной конечной концентрации 1 мкг в мл, и планшеты инкубируют в течение 1 ч при комнатной температуре. Далее добавляют около 100 мкл/лунка антитела козы против мышиного IgG-HRP (Pierce Chemical CO., № в каталоге 31432) в разведении 1:10,000 в PBS, содержащем 1% BSA. Планшеты инкубируют при комнатной температуре около 1 ч, после чего их отмывают 5 раз PBS, содержащем 0,1% Tween-20. Затем добавляют около 100 мкл/лунка ТМВ субстрата (описанного выше) и оставляют до появления синего цвета. Затем определяют оптическую плотность на спектрофотометре (планшетном ри-дере), примерно, при 370 нм. Аффинный BIAcore Аффинный анализ каждого возможного Ang-2 "пептитела" проводят на BIAcore(r)2000 (Biacore, Inc., Piscataway, NJ) с PBS и 0,005% сурфактанта Р20 (Biacore, Inc.) в качестве разделяющего буфера. Ре-комбинантный белок G (Repligen, Needham, MA) иммобилизуют на химически чистом датчике СМ5 (Biacore, Inc.) за счёт первичных аминогрупп, используя набор Amine Coupling Kit (Biacore, Inc.) в соответствии с рекомендуемым производителем протоколом. Анализы связывания проводят, сначала связывая (захватывая) по 100 Ru каждого предполагаемого affra-Ang-2 "пептитела" с иммобилизованным протеином G, после чего huAng-2 или mAng-2 в различных концентрациях (0-100 нМ) инъецируют через поверхность связанного антитела при скорости потока 50 мкл/мин в течение 3 мин. Кинетические параметры связывания, включая k (константа скорости ассоциации), kd (константа скорости диссоциации) и Ки (константа равновесной диссоциации), определяют с помощью компьютерной программы BIA evaluation 3.1 (Biacore, Inc.). Более низкие константы равновесной диссоциации указывают на более высокую аффинность "пептитела" к Ang-2. Пример 3. Идентификация пептидов, связывающих Ang-2. 1. Получение намагниченных бус с иммобилизованным Ang-2. A. Иммобилизация Ang-2 на намагниченных бусах. Для неспецифической элюции биотинилированный Ang-2 белок (Biotinylated Recombinant Human Angiopoietin-2, R &D Systems, Inc.; № в каталоге ВТ 623) иммобилизуют на Streptavidin Dynabeads (Dynal, Lake Success, NY) с концентрацией около 4 мкг биотинилированного Ang-2 белка на 100 мкл массы бус от производителя на все три раунда селекции. Для элюции антигена (Ang-2) и рецептора (Tie-2) 2 мкг биотинилированного Ang-2 белка иммобилизуют на 50 мкл Streptavidin Dynabeads для второго раунда селекции. Концентрацию покрытия уменьшают, примерно, до 1 мкг биотинилированного Ang-2 белка на 50 мкл массы бус для третьего раунда селекции. С помощью магнита бусины перемещают на одну сторону пробирки и пипеткой отбирают жидкость, бусы дважды отмывают фосфатно-солевым буферным раствором (PBS) и снова суспендируют в PBS. Биотинилированный Ang-2 белок добавляют к отмытым бусинам с вышеуказанной концентрацией и инкубируют при вращении в течение 1 ч при комнатной температуре с последующей инкубацией в течение ночи при температуре 4°C при вращении. Бусы, покрытые Ang-2, затем блокируют, добавляя BSA до конечной концентрации около 1%, и инкубируют в течение ночи при 4°С при вращении. Затем полученные в результате покрытые Ang-2 бусы 5 раз отмывают PBS перед тем, как подвергнуть их селекции. B. Приготовление бус для негативной селекции. Кроме того, готовят также бусы для негативной селекции. Для каждого условия пэннинга 500 мкл исходных бус (массы бус) от производителя подвергают вышеописанной процедуре (раздел 1 А), за исключением того, что опускают стадию инкубации с Ang-2. На последней стадии отмывания бусы делят - 49 - 008248 на пять аликвот по 100 мкл. 2. Селекция фага, связывающего TALL-1. A. Общая стратегия. Для селекции на Ang-2 связывающий фаг используют три библиотеки нитчатых фагов, обозначенных как "TN8-IX" (5х109 независимых трансформаций), "TN12-I" (1,4х109 независимых трансформаций) и "Linear" (2,2х109 независимых трансформаций) (все от Dyax Corp.). Каждую библиотеку затем подвергают либо неспецифической элюции, либо Ang-2 элюции, либо рецепторной элюции (Tie-2). Девять различных условий пэннинга применяют для Ang-2 (TN8-LX с использованием метода неспецифической элюции; TN8-IX с использованием метода Ang-2 элюции; TN8-LX с применением метода Tie-2 элюции; TN12-I с использованием метода неспецифической элюции; TN12-I с использованием метода элюции с Ang-2; TN12-I с применением метода Tie-2 элюции; Linear с использованием метода неспецифической элюции; Linear с использованием метода Ang-2 элюции; Linear с применением метода Tie-2 элюции). Для всех трёх библиотек фаг из первого раунда селекции элюируют только неспецифическим методом для других раундов селекции. Элюции с Ang-2 и с Tie-2 используют во втором и в третьем раунде селекции. В случае библиотеки Linear проводят только второй раунд для Ang-2 и Tie-2 элюции. B. Негативная селекция. Для каждого условия пэннинга из исходных библиотек TN8-IX и TN12-I отбирают аликвоты около 100 произвольных библиотечных эквивалентов (около 5х10п бое для TN8-IX и около 1,4х1011 бое для TN12-I) и около 10 произвольных библиотечных эквивалентов для библиотеки Linear (около 1 х1011 бое) и разводят с помощью PBST (PBS с 0,05% Tween-20) до 400 мкл. После последнего отмывания жидкость удаляют из первой 100 мкл аликвоты бусин, приготовленной для негативной селекции (раздел 1 В), к бусам прибавляют 400 мкл разведённого представителя исходной библиотеки. Полученную смесь инкубируют в течение 10 мин при комнатной температуре при вращении. Супернатант с фагом отбирают с помощью магнита и добавляют во вторую аликвоту 100 мкл для второй стадии негативной селекции. Таким образом осуществляют пять стадий негативной селекции. C. Селекция с применением бус с иммобилизованным белком Ang-2. Супернатант с фагом после последней стадии негативной селекции (раздел 1 В) добавляют к бусам, покрытым Ang-2 (раздел 1 А). Эту смесь инкубируют при вращении в течение 1-2 ч при комнатной температуре, что делает возможным связывание фага с нацеленным белком. После того, как супернатант отбрасывают и бусы 10 раз отмывают PBST, а затем 2 раза PBS. D. Неспецифическая элюция. После последней стадии отмывания жидкость отделяют (раздел 2С), к бусам добавляют около 1 мл раствора солей Min A (60 мМ К2НРО4, 33 мМ КН2РО4, 7,6 мМ (NH4)SO4 и 1,7 мМ цитрата натрия). Эту содержащую бусы смесь добавляют непосредственно к концентрированному образцу бактерий для инфицирования (см. ниже разделы 3А и 3В). E. "Антигенная" (Ang-2) элюция связанного фага. Для второго цикла после последней стадии отмывания (раздел 2С) связанный фаг элюируют с магнитных бус, добавляя 100 мкл 1 пМ, 0,1 нМ и 10 нМ рекомбинантного Ang-2 белка (Recombinant Human Angiopoietin-2, R &D Systems, Inc., Minneapolis, Minnesota), последовательно с 30-минутной инкубацией для каждого условия. Оставшийся фаг элюируют неспецифически (раздел 2D). Элюированный фаг после элюции с добавлением 10 нМ и неспецифической элюции объединяют и подвергают третьему раунду селекции (см. далее раздел 4). Для цикла 3 после последней стадии отмывания (раздел 2С) связанный фаг элюируют с намагниченных бус, добавляя около 1 нМ рекомбинантного Ang-2 белка и 10 нМ рекомбинантного Ang-2 белка последовательно с 30-минутной инкубацией для каждого условия. Кроме того, фаг элюируют 1 мл 100 мМ раствора триэтиламина (Sigma, St. Louis, Missuri), примерно, в течение 10 мин в ротационном шейкере. рН содержащего фаг раствора триэтиламина доводят до нейтрального (рН 7,5), добавляя 0,5 мл 1М Tris-HCl. После последней элюции 100 мМ раствором триэтиламина оставшийся фаг элюируют, добавляя бусы к бактериям (раздел 2D). F. Рецепторная (Tie-2) элюция связанного фага. Для второго цикла после последней стадии отмывания (раздел 2С) связанный фаг элюируют с намагниченных бус, добавляя 100 мкл 1 пМ, 0,1 нМ и 10 нМ рекомбинантного Tie-2 белка (Recombinant Human Tie-2-Fc Chimera, R &D Systems, Inc., Minneapolis, Minnesota) с последующей 30-минутной инкубацией для каждого условия. Оставшийся фаг элюируют неспецифически (раздел 2D). Элюированный фаг после элюции с добавлением 10 нМ и неспецифической элюции объединяют и подвергают третьему циклу селекции (см. далее раздел 4). Для цикла 3 после последней стадии отмывания (раздел 2С) связанный фаг элюируют с намагниченных бус, добавляя около 1 нМ рекомбинантного Ang-2 белка и 10 нМ рекомбинантного Ang-2 белка и 10 нМ рекомбинантного Tie-2 белка последовательно с 30-минутной инкубацией для каждого условия. Кроме того, фаг элюируют 1 мл 100 мМ раствора триэтиламина (Sigma, St. Louis, Missuri), примерно, в течение 10 мин на ротационном шейкере. Содержащий фаг раствор триэтиламина нейтрализуют (рН 7,5), - 50 - 008248 добавляя 0,5 мл 1 М Tris-HCl. После последней элюции 100 мМ раствором триэтиламина оставшийся фаг элюируют, добавляя бусы к бактериям (раздел 2D). 3. Амплификация. A. Приготовление препарата высеянных в плашки клеток. Свежую культуру Е. Coli (XL-1 Blue MRF') выращивают до ОD600 около 0,5 в среде LB, содержащей около 12,5 мкг/мл тетрациклина. Для каждого условия пэннинга около 20 мл этой культуры охлаждают льдом и центрифугируют. Осадок бактерий снова суспендируют, примерно, в 1 мл раствора солей Min A. B. Трансдукция. Каждую смесь, получаемую различными методами элюции, представленными выше (разделы 2D, 2Е и 2F), прибавляют в концентрированные образцы бактерий (раздел 3А) и инкубируют при 37°С примерно в течение 15 мин. В каждую смесь добавляют около 2 мл среды NZCYM (2XNZCYM, 50 мкг/мл ампициллина) и инкубируют примерно при 37°С в течение 15 мин, 4 мл полученного раствора помещают на большую NZCYM агаровую пластину, содержащую 50 мкл ампициллина, и инкубируют в течение ночи при 37°С. C. Сбор (харвестинг) фага. Каждую из смесей бактерии/фаг выращивают в течение ночи на большой NZCYM агаровой пластине (раздел 3В), после чего соскребают примерно в 35 мл среды LB. Агаровую пластину далее промывают ещё 35 мл среды LB. Полученную смесь бактерии/фаг в среде LB центрифугируют, чтобы высадить бактерии. Около 50 мл супернатанта с фагом переносят в свежую пробирку, добавляют около 12,5 мл раствора ПЭГ (20% ПЭГ8000, 3,5М ацетата аммония) и инкубируют на льду в течение 2 ч для преципитации фага. Осаждённый фаг (преципитат) центрифугируют и снова суспендируют в 6 мл буфера для повторного суспендирования фага (250 мМ NaCl, 100 мМ Tris pH8, 1 мМ EDTA). Этот раствор фага дополнительно очищают, центрифугированием удаляя оставшиеся бактерии, и повторно осаждают фаг, добавляя около 1,5 мл раствора ПЭГ. После стадии центрифугирования осадок фага (пеллеты) снова суспендируют в 400 мкл PBS. Этот раствор центрифугируют последний раз, чтобы отделить оставшийся бактериальный дебрис. Полученный препарат фага титруют стандартным методом бляшкообразования. 4. Дополнительная селекция и амплификация. Во втором цикле препарат амплифицированного фага (около 1010 бое) после первого цикла (раздел 3 С) используют в качестве исходного фага для проведения стадий селекции и амплификации (разделы 2 и 3). Для Ang-2 и Tie-2 элюции фаг от "10 нМ" и неспецифической элюции объединяют и амплифицируют для третьего цикла селекции. В свою очередь препарат амплифицированного фага (109 бое) после 2 цикла используют в качестве исходного фага для проведения 3 цикла селекции и амплификации (разделы 2 и 3). После стадий элюции (разделы 2D, 2Е и 2F) 3 цикла малую часть элюированного фага помещают на пластину для анализа бляшкообразования (раздел 3С). Отдельные бляшки собирают и помещают в 96-лу-ночные титрационные микропланшеты, содержащие в каждой лунке 100 мкл буфера ТЕ. Эти эталонные (master) планшеты инкубируют при 4°С в течение ночи с целью осуществить элюцию фага в ТЕ буфер. 5. Клональный анализ. Клоны фагов анализируют методами ELISA для фагов и ДНК секвенированием. Ранговое распределение последовательностей осуществляют, исходя из совместных результатов этих двух анализов. А. Анализ фага методом ELISA. Культуру XL-1 Blue MRF' выращивают до тех пор, пока OD600 не достигнет примерно 0,5. Али-квоты, около трети мкл этой культуры, помещают в каждую лунку 96-луночного титрационного микропланшета. В каждую лунку добавляют около 10 мкл элюированного фага (раздел 4) и бактерии инфицируют, примерно, в течение 15 мин при комнатной температуре. В каждую лунку добавляют около 100 мкл среды LB, содержащей около 12,5 мкг/мл тетрациклина и около 50 мкг/мл ампициллина. Затем мик-ротитрационный планшет инкубируют в течение ночи примерно при 37°С. Рекомбинантный Ang-2 белок (около 1 мг/мл в PBS) иммобилизуют на поверхности лунок 96-луночных планшетов Maxisorp (NUNC) в течение ночи при 4°С. В качестве контроля чистый стрептавидин иммобилизуют на поверхности отдельного планшета Maxisorp при концентрации около 2 мкг/мл в PBS. На следующий день жидкости в планшетах Maxisorp с иммобилизованным белком сливают и каждую лунку блокируют, примерно используя 300 мкл 5%-ого раствора молока примерно при 4°С в течение ночи (или же в течение 1 ч при комнатной температуре). Молочный раствор отбрасывают и лунки трижды отмывают раствором PBST. После последней стадии отмывания в каждую лунку планшетов Maxisorp с иммобилизованным белком добавляют около 50 мкл PBST-4% молоко. Культуры (примерно по 50 мкл) после выдерживания в течение ночи в 96-луночном микротитрационном планшете переносят в соответствующие лунки планшетов с иммобилизованным Ang-2, а также в лунки контрольных планшетов с иммобилизованным на их поверхности стрептавидином. 100 мкл смеси в двух видах планшетов инкубируют в течение примерно 1 ч при комнатной температуре. Жидкости из планшетов Maxisorp отбрасывают и лунки 3 раза отмывают PBST. HRP-конъюгированное антитело против М13 (Amersham Pharmacia Biotech) разводят примерно до 1:7500 и в каждую лунку планшетов Maxisorp добавляют примерно по 100 мкл разведённого раствора и инкубируют около 1 ч при комнатной температуре. Жидкость снова отбрасывают и лунки примерно 5 раз отмы - 51 - 008248 вают PBST. Затем в каждую лунку помещают около 100 мкл субстрата ТМВ (Sigma) и реакцию прекращают, прибавляя около 50 мкл 5N раствора H2SO4. СЮ600 считывают с помощью планшетного ридера (Molecular Devices). В. Секвенирование фаговых клонов. Для каждого фагового клона методом PCR (ПЦР) получают матрицы для секвенирования. Для амплификации примерно 500 нуклеотидных фрагментов используют следующие пары олигонуклеотидов: Праймер 1: 5'-CGGCGCAACTATCGGTATCAAGCTG-3' (SEQ ID NO: 54) Праймер 2: 5'-CATGTACCGTAACACTGAGTTTCGTC-3' (SEQ ID NO: 55) Следующие смеси готовят для каждого клона. Реагенты объем (мкл)/пробирка dH20 26.25 50% глицерин Буфер 10Х PCR (без MgCl2) 25 jwMMgCh 10 mM 0.25 100 мкМ праймер 2 0.25 Полимераза Taq 0.25 Фаг в ТЕ (раздел 4) Конечный объём реакцион- ной смеси Для проведения ПЦР с целью осуществления следующей программы: 94°С в течение 5 мин (94°С за 30 с, 55°С за 30 с, 72°С за 45 с) х 30 циклов; 72°С в течение 7 мин; охлаждение до 4°С - используют термоблок (GeneAmp PCR System 9700, Applied Biosystems). Продукт каждой ПЦР (PCR) реакции очищают с помощью набора QIAquick Multiwell PCR Purification Kit (Quiagen) в соответствии с протоколом изготовителя. Затем очищенный продукт ПЦР анализируют, пропуская около 10 мкл каждой ПЦР реакционной смеси с 1 мкл красителя (лодинг-краситель в 10 X BBXS агарозном геле) на 1%-ном агарозном геле. Сставшийся продукт затем секвенируют, используя секвенатор ABI 377 (Perkin-Elmer) в соответствии с протоколом, рекомендуемым изготовителем. 6. Ранговое распределение последовательностей и определение согласованной последовательности. A. Ранговое распределение последовательностей и анализ. Пептидные последовательности, транслируемые при использовании вариабельных нуклеотидных последовательностей (раздел 5В), коррелируют с данными ELISA. Клоны, показывающие высокое значение OD450 в лунках с иммобилизованным Ang-2 и низкое значение OD450 в лунках с иммобилизованным стрептавидином, рассматриваются как более важные (им присвоен более высокий ранг). Также важными (имеющими ранг с высоким приоритетом) считаются последовательности, встречающиеся много раз. Исходя из этих критериев, выбирают последовательности-" кандидаты" для дальнейшего анализа в качестве пептидов или "пептител". B. Спределение согласованной последовательности. Три различных класса консенсусных мотивов были получены при использовании библиотеки TN8-IX, а именно KRP С ЕЕ X WGG С X Y X (SEQIDNO:56) KRPCEEXFGGCXYX (SEQIDN0:57) XXXCXDXYWYCXXX (SEQIDN0:61) XXXCXDXYTYCXXX (SEP ID N0:62) XXX CXDXP WYC XXX (SEQ ID NO:63) XXXCXDXFTYCXXX (SEQIDNO:64) XXXCXWDPWTCEXM (SEQ ID N0:58) Один консенсусный мотив получают при использовании TN12-I библиотеки WSXCAWFXGXXXXXCRRX(SEQIDNO:59) Для всех последовательностей консенсусных мотивов подчёркнутые "основные (core, коровые) аминокислотные последовательности" из каждой согласованной последовательности получают, определяя наиболее часто встречающуюся в каждом положении аминокислоту. "X" относится к любой природной аминокислоте. Два цистеиновых остатка, прилегающие к основным (коровым) последовательностям, представляют собой фиксированные аминокислоты в библиотеках TN8-IX и TN12-I. Пептиды, идентифицированные как связывающиеся с Ang-2, представлены далее в табл. 3. - 52 - 008248 Таблица 3 Пептиды, связывающиеся с Ang-2 Пептид Seq Id No Последовательность TN8-8 KRPCEEMWGGCNYD TN8-14 HQICKWDPWNCKHW TN8-Conl KRPCEEIFGGCTYQ TN8-Con4 QEECEWDPWTCEHM TNI 2-9 FDYCEGVEDPFTFGCNH KFNPLDELEETLYEQFTQQ C17 QYGCDGFLYGCMIN Пример 4. Конструкция ДНК, кодирующей "пептитела". Модифицированные пептиды, выбираемые в качестве возможных ингибиторов связывания Ang-2:Tie-2 (см. табл. 3), используют для построения слитых белков, в которых либо мономер каждого пептида, либо тандемный димер каждого пептида (с линкером между мономерными единицами) слит в рамке считывания с ДНК, кодирующей линкер с последующей Fc областью человеческого IgG1. Каждый модифицированный пептид создан гибридизацией пар олигонуклеотидов ("олиго") с образованием полинуклеотид-ных дуплексов, кодирующих пептид вместе с линкером, состоящим, в зависимости от пептида, либо из пяти остатков глицина, либо из восьми остатков глицина, либо одного остатка лизина; эти конструкции получают в виде фрагментов от Ndel до XhoI. Эти дуплексные полинуклеотидные молекулы лигируют в вектор (pAMG21-Fc N-концевой, подробнее описан далее), содержащий человеческий Fc ген, который был предварительно гидролизован с помощью NdeI и XhoI. Смеси, полученные при лигировании, трансформируют электропорацией в клетки Е. coli штамм 2596 (GM221, подробно описанный далее) по обычным методикам. Клоны подвергают скринингу на способность продуцировать рекомбинантный белковый продукт и на содержание слияния генов с правильной нуклеотидной последовательностью. Такой единичный клон отбирают для каждого модифицированного пептида (т.е. продуктов слияния Fc-пептид). Конструкция pAMG21-Fc N-концевой вектор. pAMG21. Плазмиду экспрессии pAMG21 (ATCC No. 98113) получают при использовании pCFM1656 (ATCC No. 69576) и экспрессирующей векторной системы, описанной в патенте США 4710473, по методике, описанной в опубликованной международной патентной заявке WO 00/24782 (см. раздел в примере 2 в этой заявке со страниц 100-103, а также фиг. 17А и 17В). Fc N-концевой вектор. Fc N-концевой вектор создают, используя в качестве матрицы штамм 3788 Е. coli, pAMG21 Tpo_Gly5_Fc мономер. Информация о клонировании этого штамма обнаружена в WO 00/24782 (см. пример 2 и фиг. 10 в этой заявке). 5' ПЦР праймер (описанный ниже подробнее ) создают с целью удаления Тро пептидной последовательности в pAMG21 Тро Gly5 и замены его полилинкером, содержащим сайты АраLI и XhoI. Используя штамм 3788 в качестве темплата, осуществляют ПЦР с полимеразой для наращивания цепи, используя олигонуклеотид SEQ ID NO: 8, представленный ниже, в качестве 5' праймера и универсальный 3' праймер, SEQ ID NO: 9, изображённый далее. Полученный ПЦР продукт очищают гель-хроматографией и расщепляют рестриктазами NdeI и BsrGI. Как плазмиду, так и полинуклеотид, кодирующий представляющий интерес пептид, вместе с его линкером очищают гель-хроматографией на вращающихся (spin) колонках Qiagen (Chatsworth, CA). Затем лигируют плазмиду и инсерт обычными методами лигирования, и полученную в результате лигирования смесь трансформируют в Е. coli клетки (штамм 2596). Отбирают одиночные клоны и осуществляют секвенирование ДНК. Правильный (корректный) клон идентифицируют и используют в качестве источника вектора для модифицированных пептидов по данному описанию. 5' Праймер: ACAAACAAACATATGGGTGCACAGAAAGCGGCCGCAAAAAAACTCGAGGGTGGAGGCGGTG GGGACA (SEQ ID NO: 8) 3' Праймер: GGTCATTACTGGACCGGATC (SEQ ID NO: 9) Помимо приготовления таких модифицированных пептидов в качестве N-концевых слияний с Fc (N-концевых "пептител"), некоторые из них получают в качестве продуктов слияния по С-концу (С-концевых "пептител"). Вектор, используемый для получения С-концевых слияний, описан далее. Конструкция Fc С-концевого вектора. Fc С-концевой вектор для модифицированных пептидов создают, используя Е. coli штамм 3728, pAMG21 Fc_Gly5_Tpo мономер, в качестве темплата. Информация о клонировании этого штамма обнаружена в WO 00/24782 (см. пример 2 и фиг. 7 в описании в этой заявке). 3' ПЦР праймер (SEQ ID NO: 10) - 53 - 008248 создают с целью удаления Тро пептидной последовательности и замены его полилинкером, содержащим сайты ApaLI и XhoI. Используя штамм 3728 в качестве темплата, осуществляют ПЦР с полимеразой для наращивания цепи, используя универсальный праймер (SEQ ID NO: 11) и вышеуказанный праймер 3' праймер. Полученный ПЦР продукт очищают гель-хроматографией и расщепляют рестриктазами BsrGI и BamHI. Как плазмиду, так и полинуклеотид, кодирующий представляющие интерес пептиды, вместе с его линкером очищают гель-хроматографией на вращающихся (spin) колонках Qiagen. Затем лигируют плазмиду и инсерт обычными методами лигирования, и полученную в результате лигирования смесь трансформируют в Е. coli клетки (штамм 2596). Отбирают одиночные клоны и осуществляют секвестрование ДНК. Правильный (корректный) клон идентифицируют и используют в качестве источника вектора для модифицированных пептидов по данному описанию. 5' Праймер: CGTACAGGTTTACGCAAGAAAATGG (SEQ ID NO: 10) 3' Праймер: (SEQ ID NO: 11) GM221 (#2596). Штамм-хозяин #2596, используемый для белков слияния Fc-пептид, представляет собой Е. coli K-12 штамм, модифицированный таким образом, чтобы содержать промотор lux и как термочувствительный репрессор лямбда cI857s7 в ранней ebg области, так и lacIQ репрессор в поздней ebg области. Присутствие этих двух генов репрессоров позволяет использовать этого хозяина с различными системами экспрессии. В АТСС этот штамм имеет обозначение 202174. Пример 5. Получение "пептител". Экспрессия в Е. coli. Культуры каждой из слитых конструкций pAMG21-Fc в Е. coli GM221 выращивают при 37°С бульоне Террифика (Terrific) (См. Tartof and Hobbs, "Improved media for growing plasmid and cosmid clones", Bethesda Research Labs Focus, Volume 9, page 12, 1987, цитируется в виде ссылки в вышеупомянутой книге Sambrook et al.). Индукцию экспрессии генного продукта с использованием промотора luxPR осуществляют, добавляя в культуральную среду синтетический аутоиндуктор лактон N-(3-оксогексаноил)-DL-гомосерина до конечной концентрации 20 нг/мл. Культуры инкубируют при 37°С ещё в течение 6 ч. Затем бактериальные культуры исследуют под микроскопом на присутствие телец включения и собирают центриуги-рованием. Рефрактильные тельца включения наблюдают в индуцированных культурах; это указывает, что, вероятнее всего, Fc-слияния продуцируются в нерастворимой фракции в Е. coli. Клеточные пеллеты лизируют непосредственным ресуспендированием в буфере Лэммли для подготовки образцов, содержащем 10% в-меркаптоэтанола, и анализируют методом SDS-PAGE. В каждом случае на SDS-PAGE геле наблюдается интенсивная полоса, окрашенная Кумасси голубым, соответствующего молекулярного веса. Очистка. Клетки разрушают в воде (1/10), используя гомогенизацию под высоким давлением (два пассажа при 14000 PSI, 96.53 кПа), и тельца включения собирают центрифугированием (4000 RPM на центрифуге J-6B в течение 1 ч). Тельца включения солюбилизируют в растворе 6М гуанидина, 50 мМ Tris, 10 мМ DTT, рН 8,5, в течение 1 ч при соотношении 1/10. Для линейных пептидов, слитых с Fc, солюбилизиро-ванную смесь разводят в 25 раз в 2М мочевины, 50 мМ Tris, 160 мМ аргинина, 2 мМ цистеина, рН 8,5. Окисление проводят в течение 2 дней при 4°С, при этом образуется соединение с дисульфидной связью (т.е. гомодимер Fc-пептид). Для циклических пептидов, слитых с Fc, следуют тому же самому протоколу, добавляя три следующих условия укладки (скручивания): (1) 2 М мочевины, 50 мМ Tris, 160 мМ аргинина, 4 мМ цистеина, 1 мМ цистамина, рН 8,5; (2) 4М мочевины, 20% глицерина, 50 мМ Tris, 160 мМ аргинина, 2 мМ цистеина, рН 8.5; и (3) 4М мочевины, 20% глицерина, 50 мМ Tris, 160 мМ аргинина, 4 мМ цистеина, 1 мМ цистамина, рН 8,5. Вновь скрученный белок подвергают диализу против 1,5М мочевины, 50 мМ NaCl, 50 мМ Tris, рН 9,0. Значение рН этой смеси понижают до рН 5, добавляя уксусную кислоту. Осадок удаляют центрифугированием и рН супернатанта доводят до 5-6,5 в зависимости от изоэлектри-ческой точки каждого продукта слияния. Белок отфильтровывают и наносят при 4°С на колонку SP-Sepharose HP, уравновешенную раствором 20 мМ NaAc, 50 мМ NaCl при рН, определённом для каждой конструкции. Белок элюируют, используя линейный градиент в объёме, равном 20 объёмам колонки, в том же самом буфере от 50 мМ NaCl до 500 мМ NaCl. Фракции с максимальным объёмом удерживания (пик) собирают и фильтруют. "Пептитела", полученные вышеуказанными методами, представлены ниже, в табл. 4. - 54 - 008248 Таблица 4 Peptibody Peptibody Sequence L1(N) MGAQKFOTTJDELEETLYEQFTFQQLEGGGGG-Fc (SEQ ID NO: 12) L1(N)WT N^WLDELEETLYEQFTFQQLEGGGGG-Fc (SEQ ГО N0:13) L1(N)1KWT MKFNPLDELEETLYEQFTFQQGSGS ATGGS GSTASSGS GSATHLEGGGGG-Fc (SEQ ID NO: 14) 2xLl (N) MGAQKFNPLDELEETLYEQFIFQQGGGGGGGGKFNPL DELEETLYEQFTFQQLEGGGGG-Fc (SEQ ID N0:15) 2xLl(N)WT MKFNPLDELEETLYEQFTFQQGGGGGGGKFNPLDELEE TLYEQFTFQQLEGGGGG-Fc (SEQ ID NO: 16) Con4(N) MGAQQEECEWDPWTCEHMLEGGGGG-Fc (SEQ ID NO: 17) Con4 (N) 1K-WT MQ^OEWDPWTCEHMGSGSATGGSGSTASSGSGSATH LEGGGGG-Fc (SEQ ГО N0:18) 2xCon4 (N) IK MGAQQEECEWDPWTCEHMGSGSATGGSGSTASSGSGS ATHQEE(^WDPWTCEHMLEGGGGG-Fc (SEQ ID N0:19) LI (C) M-Fc-GGGGGAQKFNPIX)ELШTLYEQFTFQQLE (SEQ ID N0:20) LI (С) IK M-Fc- GGGGGAQGSGSATGGSGSTASSGSGSATHKFNPLDELE ETLYEQFTFQQLE (SEQ ID N0:21) 2xLl (C) M-Fc- GGGGGAQK3WLDFXEETLYEQFTFQQGGGGGGGGKF NPLDELEETLYEQFTFQQLE (SEQ ID N0:22) Con4 (C) M-Fc-GGGGGAQQEECEWDPWTCEHMLE (SEQ ID N0:23) "Cori' " XQ) iK" МПРс- GGGGGAQGSGSATGGSGSTASSGSGSATHQEECEWDP WTCEHMLE (SEQ ID N0:24) 2xCon4 (С) IK M-Fc- GGGGGAQQEECEWDPWTCEHMGSGSATGGSGSTASS GSGSATHQEECEWDPWTCEHMLE (SEQ ID N0:25) . Con4-Ll (N) MGAQEECEWDPWTCEHMGGGGGGGGKFNPLDELEET LYEQFTFQQGSGSATGGSGSTASSGSGSATHLEGGGGG-Fc (SEQ ID N0:26) Con4-Ll (C) M-Fc- GGGGGAQGSGSATGGSGSTASSGSGSATHKTM'LDELE ETLYEQFTTQQGGGGGQEECEWDPWTCEHMLE (SEQ ID N0:27) TN-12-9 (N) MGAQ-roYCEGVEDPFTFGCDNHLE-GGGGG-Fc (SEQ ЮШ:28) C17(N) MGAQ-QYGCDGFLYGCMINLE-GGGGG-Fc (SEQ ID NO:29) TN8-8 (N) MGAQ-KM'CEEMWGGCJTYDLEGGGGG-Fc (SEQ ED N0:30) TN8-14 (N) MGAQ-HQICKWDPWTCKHWLEGGGGG-Fc (SEQ ID NO:31) Conl(N) MGAQ-KRPCEEIFGGCTYQLEGGGGG-Fc (SEQ ID NO:32) В табл. 4 "Fc" относится к последовательности человеческого Fc IgG1. В столбце 2 представлены аминокислотные последовательности "пептител". Область Fc в них представляет собой меченый Fc" и представлена ниже в SEQ ID NO: 60. Ясно, что там, где в названии используется метка, например, "СОП4" или "СОП-4", это относится к пептиду СОП-4, тогда как применение суффикса "С", "(С)" или "-С"; или "N", "(N)" или "-N" указывает, что молекула представляет собой "пептитело" по данному описанию. Суффиксы "N", "(N)" или''-N" в названии "пептитела" указывают, что Ang-2-связывание пептида (или пептидов) является/являются ^концевым(-и) к Fc домену, а суффиксы "С", "(С)" или "-С" указывают, что Ang-2-связывание пептида (или пептидов) является/являются С-концевым(-и) к Fc домену. Кроме того, 2ХСОП4 (С) 1K, по определению в SEQ ID NO: 25, может в данном описании также употребляться без суффикса "1K". Аминокислотная последовательность Fc области каждого "пептитела" суть следующая (от амино-конца до карбоксильного конца): - 55 - 008248 DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDV SbiEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVL HQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPS RDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVL DSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLS LSPGK (SEQ ID NO: 60) Последовательность ДНК (SEQ ID NO: 33-53), кодирующей "пептитела", соответствующие последовательностям SEQ ID NO: 12-32, соответственно, в табл. 4 представлена ниже. SEP TP NO: 33 ATGGGTGCACAGAAATTCAACCCGCTGGACGAACTGGAAGAAACTCT GTACGAACAGTTCACTTTCCAGCAGCTCGAGGGTGGAGGCGGTGGGG ACAAAACTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTGGGGG GACCGTCAGTTTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGAT CTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGA AGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGC ATAATGCCAAGA'CAAAGCCGCGGGAGGAGCAGTACAACAGCACGTAC GGTGTGGTr! A GCGTP.r'Tr! А Г^ППТОСТГПГ. A CC A GCi A GTGGOTG A ATGGO AAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATC GAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGT GTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAG CCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGA GTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTC CCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGT GGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGAT GCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTC TCCGGGTAAATAATGGATCC SEP ID NO:34 ATGAAATTCAACCCGCTGGACGAACTGGAAGAAACTCTGTACGAACA GTTCACTTTCCAGCAGCTCGAGGGTGGAGGCGGTGGGGACAAAACTCA CACATGTCCACCTTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGT TTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACC CCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGA GGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAA GACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCA GCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACA AGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCA TCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTG CCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGC CTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGC AATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGA CTCCGACGGGTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAG CAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGC TCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAA ATAA - 56 - 008248 SEP Ш N0:35 ATGAAATTCAACCCGCTGGACGAACTGGAAGAAACTCTGTACGAACA GTTCACTTTCCAGCAGGGATCCGGTTCTGCTACTGGTGGTTCCGGCTCC ACCGCAAGCTCTGGTTCAGGCAGTGCGACTCATCTCGAGGGTGGAGGC GGTGGGGACAAAACTCACACATGTCGACCTTGCCCAGCACCTGAACTC CTGGGGGGACCGTCAGTTTTCCTCTTCCCCCCAAAACCCAAGGACACC CTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTG AGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGT GGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACA GCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGC TGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCA GCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGA ACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAA CCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACAT CGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAAeTACAAGA CCACGCCTCCCGTGCrGGACTCCGACGGCTCCTTGTTCCTCTACAGCAA GCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCAT GCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCC TCTCCCTGTCTCCGGGTAAATAA SEP Ш NO:36 ATGGGTGCACAGAAATTCAACCCGCTGGACGAACTGGAAGAAACTCT GTACGAACAGTTCACTTTCCAGCAGGGTGGTGGTGGTGGTGGCGGTGG TAAGTTCAACCCACTGGATGAGCTGGAAGAGACTCTGTATGAACAGTT CACTTTCCAGCAACTCGAGGGTGGAGGCGGTGGGGACAAAACTCACA CATGTCCACCTRGCCCAGCACCTGAACTCCRGGGGGGACCGTCAGTTT TCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCC CTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAG GTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAG ACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAG CGTCCTGACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAA GTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCAT CTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGC CCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCC TGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCA ATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGAC TCCGACGGCRCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGC AGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCT CTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA TAA SEQ Ю NO:37 ATGAAATTCAACCCGCTGGACGAACTGGAAGAAACTCTGTACGAACA GTTCACTTTCCAGCAGGGTGGTGGTGGTGGCGGTGGTAAGTTCAACCC ACTGGATGAGCTGGAAGAGACTCTGTATGAACAGTTCACTTTCCAGCA ACTCGAGGGTGGAGGCGGTGGGGACAAAACTCACACATGTCCACCTT GCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTTTTCCTCTTCCCCCC AAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATG GGTGGTGGTGG A GGTG А ОГП A GG А АГтАГПГТОАОГтТГА AGTTGA AGTG GTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGG AGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCC TGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCC AACAAAGCCCTCCCAGCGCCCATCGAGAAAACCATCTCCAAAGCCAA AGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGG ATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCT TCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGGAATGGGCAGCCG GAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCC TTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAG GGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCAC TACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATAA - 57 - 008248 SEP Ю N0:38 ATGGGTGCACAGCAGGAAGAATGCGAATGGGACCCATGGACTTGCGA ACACATGCTCGAGGGTGGAGGCGGTGGGGACAAAACTCACACATGTC CACCTTGCCCAGGACCTGAACTCCTGGGGGGACCGTCAGmTCCTCTT CCCCCGAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGT CACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTT CAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGC CGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTC ACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAA GGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAA AGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCAT CCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCA AAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGG CAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGAC GGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGG CAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCAC AACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATAA SEP ID NO:39 ATGCAGGAAGAATGCGAATGGGACCCATGGACTTGCGAACACATGGG ATCCGGTTCTGCTACTGGTGGTTCCGGCTCCACCGCAAGCTCTGGTTCA GGCAGTGCGACTCATCTCGAGGGTGGAGGCGGTGGGGACAAAACTCA CACATGTCCACCTTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGT TTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACC CCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGA GGTCAAGTTGAAGT-G^ACGTGGACGGCGTGGA GGTGC ATA ATGCCAA GACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCA GCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACA AGTGCAAGGTCTCCAACAAAGCCCTCCCAGCGCCCATCGAGAAAACCA TCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTG CCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGC CTGGTCAAAGGCTTCTATCCGAGCGACATCGCCGTGGAGTGGGAGAGC AATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGA CTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAG CAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCGGTGATGCATGAGGC TCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTGTCCGGGTAA ATAA SEP ID N0:40 ATGGGTGCACAGCAGGAAGAATGCGAATGGGACCCATGGACTTGCGA ACACATGGGATCCGGTTCTGCTACTGGTGGTTCCGGCTCCACCGCAAG CTCTGGTTCAGGCAGTGCGACTCATCAGGAAGAATGCGAATGGGACCC ATGGACTTGCGAACACATGCTCGAGGGTGGAGGCGGTGGGGACAAAA CTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTGGGGGGACCGT CAGTTTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCG GACCCCTGAdGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACC СТОАШТСААОТТСААСТООТАСОТООАСООСОТООАООТОСАТААТО CCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTG GTCAGCGTCOTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAG TACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAA ACCATCTCCAAAGCGAAAGGGCAGCCCCGAGAACCACAGGTGTACAC CCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGAC CTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGA GAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGC TGGACTCCGACGGCTCCTrCTTCCTCTACAGCAAGCTCACCGTGGACA AGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATG AGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGG GTAAATAA - 58 - 008248 SEP ID NO:41 ATGGACAAAACTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTG GGGGGACCGTCAGmTCCTCTTCCCCCCAAAACCCAAGGACACCCTC ATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGC CACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGA GGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCA CGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGA ATGGCA AGGAGTAG A AGTGCA AGGTCTCCA ACAA A GCCGTCGC A GCC CCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACC АЛА ПЛФПГП А /~1 А ^Г^Г^ФГ^ /-\Г-\ГЛГ~\ГЛ А ФЛЛЛЛ ^Л А ППП А /~1/~ч-П/~* А ЛЛ А А А А А GGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGC CGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCA CGCCTCCCGTGCTGGACTCCGACGGCTCCTrcrTCCTCTACAGCAAGCT CACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTC CGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTC CCTGTCTCCGGGTAAAGGTGGAGGTGGTGGTGCACAGAAATTCAACCC GCTGGACGAGCTGGAAGAGACrCTGTACGAACAGTTTACTTTTCAACA GCTCGAGTAA SEP ID NO:42 ATGGACAAAACTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTG GGGGGACCGTCAGTTTTCCTCTTCCCCCCAAAACCCAAGGACACCCTC ATGATGTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGC CACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGA GGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCA CGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGA ATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCC CCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACC ACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCA GGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGC CGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCA CGCCTCCCGTGCTGGACTCCGACGGCTCGTTCTTCCTCTACAGCAAGCT CACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTC CGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTC CCTGTCTCCGGGTAAAGGTGGAGGTGGTGGTGCACAGGGATCCGGTTC TGCTACTGGTGGTTCCGGCTCCACCGCAAGCTCTGGTTCAGGCAGTGC GACTCATAAATTCAACCCGCTGGACGAACTGGAAGAAACTCTGTACGA ACAGTTCACrTTCCAGCAACTCGAGTAA SEQ Ш NP:43 ATGGACAAAACTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTG GGGGGACCGTCAGTTTTCCTCTTCCCCCCAAAACCCAAGGACACCCTC ATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGC CACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGA GGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCA CGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGA AT-GGGAAGGAGTAG A AGTGGA A GGTCTCC A A G A A A GCCCTCCC A GCC CCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACC д s-\ д л лтлт к x ллптпл/^^л/ч л ПЧУ-ЧУЧУЧУ-Ч J--4 я т^*ч л лл1тл A A А /~1 А л /^/-ч л GGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGC CGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCA CGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCT CAGCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTC CGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTC CCTGTCTCCGGGTAAAGGTGGAGGTGGTGGTGCACAGAAATTCAACCC GCTGGACGAACrGGAAGAAACTCTGTACGAACAGTTCACTTTCCAGCA GGGTGGTGGTGGTGGTGGCGGTGGTAAGTTCAACCCACTGGATGAGCT GGAAGAGACTCTGTATGAACAGTTCACTTTCCAGCAACTCGAGTAA - 59 - 008248 SEP 3D NO:44 ATGGACAAAACTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTG GGGGGACCGTCAGmTGCTCTTCCCCCCAAAACCCAAGGACACCCTC ATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGC CACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGA GGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCA CGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGA ATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCC CCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACC ACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAGCAAGAACCA GGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGC CGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCA CGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCT CACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTC CGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTC CCTGTCTCCGGGTAAAGGTGGAGGTGGTGGTGCACAGGAGGAAGAAT GCGAATGGGACCCATGGACTTGCGAACACATGCTCGAGTAA SEP ГО N0:45 ATGGACAAAACTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTG GGGGGACCGTCAGTTTTCCTCTTCCCCCCAAAACCCAAGGACACCCTC ATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGAGGTGAGC CACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGA GGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCA CGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGA CCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCGCCGAGAACC ACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCA GGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGC CGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCA CGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCT CACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTC CGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTC CCTGTCTCCGGGTAAAGGTGGAGGTGGTGGTGCACAGGGATCCGGTTC TGCTACTGGTGGTTCCGGCTCCACCGCAAGCTCTGGTTCAGGCAGTGC GACTCATCAGGAAGAATGCGAATGGGACCCATGGAGTTGCGAACACA TGCTCGAGTAA SEP ГО NO:46 ATGGACAAAACTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTG GGGGGACCGTCAGTTrTCCTCTTCCCCCCAAAACCCAAGGACACCCTC ATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGC CACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGA GGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCA CGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGA ATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCC CCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACC ACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCA GGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGC CGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCA CGCCTCCCGTGCTGGACrCCGACGGCTCCTTCTTCCTCTACAGCAAGCT CACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTC CGTGATGCATGAGGCTCTGCAGAACCACTACACGCAGAAGAGCCTCTC GCTGTCTCCGGGTAAAGGTGGAGGTGGTGGTGCACAGCAGGAAGAAT GCGAATGGGACCCATGGACTTGCGAACACATGGGATCCGGTTCTGCTA CTGGTGGTTCCGGCTCCACCGCAAGCTCTGGTTCAGGCAGCGCGACTC ATCAGGAAGAATGCGAATGGGACCCATGGACTTGCGAACACATGCTC GAGTAA - 60 - 008248 SEP ID N0:47 ATGGGTGCACAGGAAGAATGCGAATGGGACCCATGGACTTGCGAACA CATGGGTGGTGGTGGTGGTGGCGGTGGTAAATTCAACCCGCTGGACGA ACTGGAAGAAACTCTGTACGAACAGTTCACTTTCCAGCAGGGATCCGG TTCTGCTACTGGTGGTTCCGGCTCCACCGCAAGCTCTGGTTCAGGCAGT GCGACTCATCTCGAGGGTGGAGGCGGTGGgGACAAAACTCACACATGT CCACCTTGCCCAGCACCTGAACrCCTGGGGGGACCGTCAGTTTTCCTCT ТССГГГГЛ A A AGGCAAGGACAGCCTCATGATCTCCCGGACCCCTGAGG TCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGT TCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAG CCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTC ACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAA GGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAA AGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCAT CCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCA AAGKJCTTCTATCCCAGCGACATGGCCGTGGAGTGGGAGAGCAATGGG CAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGAC GGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGG CAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCAC AACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATAA SEP 3D NP:48 ATGGACAAAACTCACACATGTCCACCTTGCCCAGCACCTGAACTCCTG GGGGGACGGTCAGTTTTCCTCTTCCCCCCAAAACCCAAGGACACCCTC ATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGC CACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGA GGTGGATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCA CGTACCGTGTGGTCAGCGTCCTCACCGTGCTGCACCAGGACTGGCTGA ATGGCAAGGAGTAGAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCC CCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACC ACAGGTGTACACCCTGCCCCCATCCGGGGATGAGCTGACCAAGAACCA GGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGC CGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAAeTACAAGACCA CGCCTCCCGTGCTGGACrCCGACGGCTCCTTCTTCCTCTACAGCAAGCT CACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTC CGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTC CCTGTCTCCGGGTAAAGGTGGAGGTGGTGGTGCACAGGGATCCGGTTC TGCTACTGGTGGTTCCGGCTCCACCGCAAGCTCTGGTTCAGGCAGTGC GACTCATAAATTCAACCCGCTGGACGAACTGGAAGAAACTCTGTACGA ACAGTTCACTTTCCAGCAGGGTGGTGGCGGTGGTCAGGAAGAATGCGA ATGGGACCCATGGACTTGCGAACACATGCTCGAGTAA SEP3DNP:49 ATGGGTGCACAGTTCGACTACTGCGAAGGTGTTGAAGACCCGTTCACT TTCGGTTGCGACAACCACCTCGAGGGTGGAGGCGGTGGGGACAAAAC TCACACATGTCCACCTTGCCCAGCACCTGAACTCCTGGGGGGACCGTC AGTTITCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGG ACCCCTGAGGTCACATGCGTGGTGGTGGAGGTGAGCCACGAAGACCCT GAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCC AAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGT CAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTA CAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAAC CATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCC TGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCT GCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGA GCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTG GACTCCGACGGCrCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAG AGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAG GCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGT AAATAA - 61 - 008248 SEP ГО N0:50 ATGGGTGCACAGCAGTACGGTTGCGACGGTTTTCTGTACGGTTGCATG ATCAACCTCGAGGGTGGAGGCGGTGGGGACAAAACTCACACATGTCC ACCTTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTTTTCCTCTTC CCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTC ACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTC AACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCC GCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCA CCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAG GTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAA GCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCC CGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAA AGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGC AGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGAC GGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGG CAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCAC AACCACTACACGCAGAAGAGCCTCTCCCTGTCTGCGGGTAAATAA SEOIDNO:51 ATGGGTGCACAGAAACGCCCATGCGAAGAAATGTGGGGTGGTTGCAA CTACGACCTCGAGGGTGGAGGCGGTGGGGACAAAACTCACACATGTC CACCTTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTTTTCCTCTT CCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGT CACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTT CAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGC CGCGAA A GG А ПСА GT A G А А Г А ПГ A GGT A GGGTGTGGTG A GGGTGGTG ACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAA GGTCTCGAACAAAGCGCTCCCAGCCCCCATCGAGAAAACCATCTCCAA AGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCAT CCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCA AAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGG CAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTGCGAC GGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGG CAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCAC AACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATAA SEP Ш NQ:52 ATGGGTGCACAGCACCAGATCTGCAAATGGGACCCGTGGACCTGCAA ACACTGGCTCGAGGGTGGAGGCGGTGGGGACAAAACTCACACATGTC CACCTTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTTTTCCTCTT CCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCGCTGAGGT CACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTT CAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGC CGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTC ACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAA GGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAA AGCCAAAGGGCAGCCCGGAGAACCACAGGTGTACACCCTGCCCCCAT CCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACGTGCCTGGTCA AAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGG CAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGAC GGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGG CAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCAC AACCACTACACGGAGAAGAGCCTCTCCCTGTCTCCGGGTAAATAA - 62 - 008248 SEaiDNO:53 ATGGXJTGCACAGAAACGTCCATGCGAAGAAATCTTCGGTGGTTGCACC TACCAGCTCGAGGGTGGAGGCGGTGGGGACAAAACTCACACATGTCC ACCTTGCCCAGGACCTGAACTCCTGGGGGGACCGTCAGTTTTCCTCTTC CCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTC ACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTC AACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCC QCGGGAGGAGCAGTAC A AGAGGA GGT A GGGTGTGGTP А ПГПТГГТС. A CCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAG GTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAA GCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCC CGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAA AGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGC AGCCGGAGAACAACTACAAGACCACGGCTCCCGTGCTGGACTCCGAC GGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGG CAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCAC AACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATAA Пример 6. Анализ "пептител". Четырнадцать пептител испытывают методом ELISA нейтрализацией, а три "пептитела" испытывают методом аффинного ELISA. Результаты представлены в табл. 5. Таблица 5 hAng-2 mAng-2 hANg-2 "Пептитело" IC50 (нМ) ЕС50 (нМ) IC50 (нМ) ЕС50 (нМ) IC50 (нМ) ЕС50 (нМ) 2xCon4(C) 1К 0.04 0.02 Con4-Ll (С) 0.05 0.04 Соп4 (С) 0.20 0.30 2xLl (N) 0.65 0.80 Con4(N) 0.85 0.03 0.72 0.07 Нет ингиби рования Нет связывания 2xLl (С) 0.90 1.0 Con4 (N) 1К-WT 1.9 L1(N) Нет ингиби рования C17(N) Нет ингиби рования 12-9 (N) 7.7 Нет ингиби рования Conl (N) -200 Нет ингиби рования 8-14 (N) Нет ингиби рования L1(C) 8-8 (N) -700 Нет ингиби рования Негативный контроль "Пептитело" 4883 Нетин-гибиро вания Нет свя зыва ния Нет ингиби рования Нет связывания Нет ингиби рования Нет связывания Аминокислотная последовательность негативного контрольного "пептитела" 4883 представляет собой следующую последовательность (Fc область подчёркнута, линкер "GGGGG", а пептидная область выделена жирным шрифтом): MDKTHTCPPCPAPELLGGPSVFLPPPKPKDTLMSRTPEVTCVVVD VSHEDPEVKJNWYVDGVEVHNAKTKPREEOYNSTYRVVSVLTVL HODWLNGKEYKCKVSNKALPAPIEKTISKAKGOPREPOVYTLPPS RDELTKNOVSLTCLVKGFYPSDIAVEWESNGOPENNYKTTPPVLD SDGSFFLYSKLTVDKSRWOOGNVFSCSVMHEALHNHYTOKSLSLS PGK-GGGGG-CTAGYHWNSDCECCRRN (SEP ID NG: 243) Понятно, что фраза "Нет ингибирования" в данном описании не означает, что соединение не имеет ингибирующих свойств. Скорее, выражение "Нет ингибирования", применяемое в данном описании, относится к таким соединениях, для которых в ходе испытания методом ELISA с нейтрализацией в условиях по данному описанию получено значение IC50 выше, чем 1000 нМ, что является наивысшей концен - 63 - 008248 трацией, при которой проводится скрининг этих соединений. Несмотря на то, что заметных ингибирую-щих свойств не наблюдается для молекул с пометкой "Нет ингибирования", ясно, что эти молекулы в действительности могут демонстрировать ингибирующие свойства в других условиях анализа или в других методах анализа. Ясно, что в предпочтительном варианте изобретение относится к "пептителам", проявляющим ингибирующие свойства в анализах по данному описанию. Два "пептитела" изучают аффинным анализом BIAcore (как описано в примере 2). Результаты представлены ниже, в табл. 6. Таблица 6 hAng-2 mAng-2 "Пептитело" KD (нМ) ka(l/Ms) kd(l/s) KD(HM) ka(l/Ms) kd(l/s) РЪ LI (N) 3.1 2.9 xlO5 9.1 xlO"4 0.42 5.6 x 105 2.3 x 10"4 Con4 (N) 0.67 3.3 xlO5 2.2 x КГ4 0.60 7.3 x 103 4.4 x 10"4 TN19-9 (N) 8.2 1.2 xlO5 1.0 x W3 0.32 7.2 xlO5 2.3 x 10'4 Пример 7. Изучение терапевтической эффективности при системном введении Ang-2 "пептитела". Ang-2 "пептитело", TN8-Con4-C, вводят подкожно А431 мышам, несущим опухоль, по схеме "1 раз в день" через 72 ч после введения опухоли. Дозы "пептитела" составляют 1000, 200, 40 и 8 ug/мышь/день. Всего 20 дают всем животным. Объём опухоли и вес тела записывают 3 раза в неделю. По окончании исследования животных умерщвляют и их сыворотку собирают для измерения уровней "пептитела" методом ELISA. Во всех группах собирают опухоли и панель нормальных тканей. Результаты показаны на фиг. 1. Как можно видеть, заметная разница в росте опухоли наблюдается между группой, обработанной Ang-2 "пептителом" и контролем с носителем. Все четыре дозы Ang-2 " пептитела" ингибируют рост опухоли по сравнению с контролем (носителем) (р <0,0001 по сравнению с контрольным носителем, ANOVA). Напротив, опухоли в контрольной группе продолжали расти со значительно более высокой скоростью. Обработка этим "пептителом" не оказала заметного влияния на конечный вес тела, вес органов или гематологические параметры животных, получавших указанные выше дозы. Пример 8. 1. Конструкция вторичных пептидных библиотек Ang-2. A. Электрокомпетентные E.coli клетки. Полученные электропорацией компетенетные клетки Epicurian Coli(r) XL1-Blue MRF' (Stratagene #200158) получают от фирмы Stratagene (Stratagene Cloning Systems, La Jolla, CA). B. Модификация вектора pCES1. ПЦР осуществляют, используя ПЦР темплатные системы для наращивания цепи системы (Extend Long Template PCR Systems, Roche Diagnostics Corp., Indianapolis, IN) с 1 мкг вектора pCES1 (TargetQuest Inc.) в качестве темплата. Объём ПЦР-смеси, составляющий 100 мкл, содержит 1х буфера для проведения ПЦР, 200 нМ каждого из двух праймеров: 5'-CAAACGAATGGATCCTCATTAAAGCCAGA-3' (SEQ ID NO: 244) и 5'-GGTGGTGCGGCCGCACTCGAGACTGTTGAAAGTTGTTTAGCA-3' (SEQ ID NO: 245), 200 нМ dNTP и 3 единицы (U) Tag ДНК полимеразы. ПЦР система TRIO Thermoblock (Biometra) PCR работает следующим образом: 94°С в течение 5 мин; 30 циклов 94°С по 30 с, 50°С по 30 с, 72°С по 45 с; и 72°С по 10 мин; охлаждение до 4°С. Затем продукты ПЦР наносят на 1% агарозный ген и очищают на вращающейся колонке QIAGEN Spin Column (QIAGEN Inc., Valencia, CA) в соответствии с протоколами производителя. Вторую ПЦР реакцию осуществляют с 5 мкл продуктов ПЦР и 200 нМ каждого из двух праймеров: 5'-CAAACGAATGGATCCTCATTAAAGCCAGA-3' (SEQ ID NO: 246) и 5'-AACACAAAAGTGCACAGGGTGGAGGTGGTGGTGCGGCCGCAGT-3' (SEQ ID NO: 247), в тех же условиях ПЦР, что и описанные выше. Продукты ПЦР и исходный вектор pCES1 затем расщепляют по отдельности в 100 мкл реакционной смеси, содержащей буфера 1x NEB2, 60 Ед (U) ApaLI (New England Biolabs, Beverly, MA), 60 Ед (U) BamHI (New England Biolabs) при 37°С в течение 1 ч. ДНК после отщепления очищают на колонке QIAGEN Spin Column и лигируют вместе в 40 мкл реакционной смеси, содержащей 1 х буфера для лиги-рования и 40 Ед (U) N4 ДНК лигазы (New England Biolabs) при комнатной температуре в течение ночи. Векторы трансфецируют в Е. coli и инкубируют при 37°С в течение ночи. Выделенные единичные колонии отбирают и затем очищают плазмиду на колонке QIAGEN Spin Column. Правильный инсерт подтверждают секвенированием ДНК. C. Получение векторной ДНК. 1 мкг векторной ДНК модифицированного pCES1 (из раздела 1 В, см. выше) трансформируют в 40 мкл электрокомпетентных клеток XL1-blue E. coli (из раздела 1 А, см. выше), используя Gene Pulser II (ген - 64 - 008248 ный импульсный генератор) (BIO RAD, Hercules, CA), установленный при 2500 В, 25 мкФ, и 200 ом. Затем образец трансформированных бактерий немедленно переносят в пробирку, содержащую 960 мкл SOC (2% триптона, 0,5% дрожжевого экстракта, 10 мМ NaCl, 2,5 мМ KQ, 20 мМ глюкозы, 10 мМ MgSO4, 10 мМ MgCl2), и культуры выращивают при 37°С при встряхивании в течение 1 ч. Затем клетки наносят на 2х YTAGT (2xYT с 100 ug/мл ампициллина, 12,5 ug/мл тетрациклина и 2% глюкозы) агаровую пластинку и инкубируют при 37°С в течение ночи. Одиночную колонию подтверждают секвенированием и используют для инокуляции 2 л среды 2xYTAQT при 37°С с помощью встряхивания в течение ночи. Плазмидную векторную ДНК очищают, используя набор QIAGEN Plasmid Maxi Kit в соответствии с протоколами производителя. D. Расщепление (гидролиз) векторной ДНК. Всего около 2000 мкг векторной ДНК (из раздела 1С, см. выше) расщепляют в 5000 мкл реакционной смеси, содержащей 1x NEB буфер 2, 300 Ед (U) АраLI и 300 Ед (U) XhoI, при 37°С в течение ночи. Реакцию расщепления рестриктазами инкубируют при 37°С и анализируют в предварительно приготовленном 0,8% агарозном геле (Embi Tec, San Diego, CA). Затем линеаризованный вектор ДНК вырезают из геля и экстрагируют, используя набор для экстракции в геле -QIAquick Gel Extraction Kit (QIAGEN Inc.) в соответствии с рекомендациями производителя. E. Получение олигонуклеотидов из библиотеки. Шесть олигонуклеотидов из библиотеки (1 закреплённый-фиксированный и 5 нанесённых) создают на основе последовательностей, полученных в результате описанных выше процедур. Один фиксированный олигонуклеотид из библиотеки представляет собой 5'-CACAGTGCACAGGGiWKNNK^KNNKN№CNNKNNKS ARTGGGATCCGTGG AS CNNKNNKNNKNNKNNK^^ CTCTCGAGATCА-3' (номер библиотеки 20) (SEQ ID NO: 248); и два из 70% нанесённых библиотечных олигонуклеотидов суть следующие: S'-CACAGTGCACAGGGTNr^mK^KaaKcgKccKMSTKga KgaKatKttKggKggK^KacKtaKcaK^K^KNNKCATTCTC TCGAGATCА-3' (номер библиотеки 27); (SEQ ID NO: 249); 5'-CACAGTGCACAGGGTNNKaaKttKaaKccKctKgaKgaKctKgaKga KacKctKtaKgaKcaKttKacKttKcaKcaKNNKCATTCTCTCGAGATCA- 3' (номер библиотеки 99); (SEQ П) NO: 250) Строчные буквы обозначают смесь 70% указанного основания и 10% любого из трёх других нук-леотидов). Другие три из 91% нанесённых библиотечных олигонуклеотидов суть следующие: 5,-CACAGTGCACAGGGтаNlШЧIШЧKcaKgaKgaKTGCgaKtg KgaKccKtgKacKTGCgaKcaKatl^NKNNKNNKCATTCTCTCGAGA ТС А-3' (номер библиотеки 94); (SEQ ID NO: 251); 5'-CACAGTGCACAGGGTbINKttKg^aK> fNKgaKggKgtKgaKgaKcc KttKacKttKggI^NNKgaKaaKcaKNNKCATTCTCTCGAGATCA-3, (номер библиотеки 25); (SEQ ID NO: 252); 5'-CACAGTGCACAGGGTNNKaaKttKaaKccKctKgaKgaKctKgaKga KacKctKtaKgaKcaKttKacKttKcaKcaKNNKC ATTCTCTCGAGATCA- 3' (номер библиотеки 26); (SEQ ID NO: 253) Как понимают специалисты в данной области техники, в вышеприведённых олигонуклеотидах (олиго, oligos) "N" указывает, что каждый из четырёх нуклеотидов (А, Т, С и G) в равной мере представлен в процессе синтеза "олиго", а "K" показывает, что нуклеотиды G и Т равно представлены в синтезе олиго. Строчные буквы обозначают смесь 91% указанного основания и 3% каждого из других трёх нук-леотидов. Каждый из этих олигонуклеотидов используют в качестве темплата в ПЦР. Для проведения ПЦР реакций используют набор ПЦР системы с высокоточным наращиванием цепи (Expand High Fidelity PCR System kit (Roche Diagnostics Corp.)). Каждую библиотеку олигонуклеоти-дов амплифицируют в 96-луночном планшете в 50 мкл ПЦР-реакционной смеси, которая содержит 1 нМ олигонуклеотида из библиотеки, 1X буфера для ПЦР, 300 нМ каждого из праймеров: 5'-CACAGTGCACAGGGT-3' (SEQ ID NO: 254); и 5'-TGATCTCGAGAGAATG-3', (SEQ ID NO: 255); 200 мкM dNTP, 1,5 мМ MgCl2 и 350 U полимеразы для наращивания цепи. Термоблок (GeneAmp PCR System 9700, Applied Biosystems) применяют для осуществления сле - 65 - 008248 дующей программы: 94°С за 5 мин; 25 циклов (94°С за 30 с, 52,5°С за 60 с, 72°С за 30 с); 72°С за 10 мин; охлаждение 4°С. Затем свободные нуклеотиды удаляют с помощью набора для ПЦР-очистки QIAquick PCR Purification Kit (QIAGEN Inc. Cat. #28104) в соответствии с протоколами производителя. F. Расщепление олигонуклеотидов из библиотек. Для каждой библиотеки ПНР продукты(раздел 1E) гидролизуют (расщепляют) в 1200 мкл реакционной смеси, которая содержит 1x NEB буфер 2, 750 Ед (U) АраLI и 750 Ед (U) XhoI, при 37°С в течение ночи. ДНК после расщепления отделяют на предварительно приготовленном 3% агарозном геле (Embi Tec). Нужную полосу ДНК из каждой реакции вырезают из геля и извлекают (экстрагируют), используя целлюлозно-ацетатный фильтр 0,22 мкм (Corning Inc. Cat# 8160) центрифужной пробирки COSTAR Spin-X. G. Лигирование вектора с олигонуклеотидами библиотеки. 450 мкл реакционной смеси для лигирования, содержащей линеаризованный вектор (раздел 1D) и каждый ПЦР продукт расщепляемой библиотеки (раздел 1F) в молярном соотношении 1:5, 1x NEB буфер для лигирования и 20000 Ед (U) T4 ДНК лигазы, выдерживают при 16°С в течение ночи. Продукты лигирования инкубируют при 65°С в течение 20 мин, чтобы инактивировать Т4 ДНК лигазу, и дополнительно инкубируют со100 Ед (U) NotI при 37°С в течение 2 ч, чтобы свести к минимуму самолигирова-ние вектора. Затем дотированные продукты очищают стандартной экстракцией смесью фенол/хлороформ (Molecular Cloning: A Laboratory Manual, Maniatis et al., 3rd Edition, Cold Spring Harbor Laboratory Press, 2000) и снова суспендируют в 120 мкл Н2О. H. Трансформация методом электропорации. Для каждой библиотеки проводят двенадцать реакций электропорации. Для каждой трансформации 10 мкл лигированного вектора ДНК (раздел 1G) и 300 мкл клеток XL1-BLUE MRF' (раздел 1А) смешивают в кювете 0,2 см (BIO-RAD). Полученную смесь выдерживают в пульсовом режиме в Gene Pulser II, установленном при 2500 В, 25 uФ и 200 ом. Затем трансформированные в результате двенадцати реакций электропорации бактерии объединяют и переносят в колбу, содержащую 26 мл SOC, для инкубации при 37°С в течение 1 ч. Клетки добавляют в 450 мл 2х YTAG и выращивают при 37°С при встряхивании в течение 5 ч. Клетки центрифугируют при 4000 об./мин 15 мин при 4°С. Клеточный осадок затем снова суспендируют в 12 мл 15% глицерин/2хУТ и хранят при -80°С. Это является первичным штаммом библиотек. Титры указывают размеры библиотек 5,0х109 (номер библиотеки 20), 3,3 х1010 (номер библиотеки 94), 4,7х109 (номер библиотеки 25), 5,0х109 (номер библиотеки 26), 3,0х109 (номер библиотеки 27) и 4,2х109 (номер библиотеки 99) независимых трансформантов. 2. Амплификация библиотек. A. Приготовление вторичного штамма библиотек. Из первичного штамма клеток библиотек (из раздела 1H, см. выше) достаточное количество клеток, чтобы покрыть 10Х размер каждой библиотеки, используют для инокуляции 2xYTAGT (2YT с 100 ug/мл ампициллина, 12,5 ug/мл тетрациклина и 2% глюкозы) среды, так чтобы начальная величина OD600 составляла 0,1. Культуры выращивают при 37°С при встряхивании в течение нескольких часов до достижения OD600=0,5. Аликвоту - одну десятую часть этой библиотеки отбирают и выращивают в отдельных колбах в течение ещё двух часов при 37°С. Затем эти субкультуры центрифугируют при 4000 об./мин на роторе Beckman JA-14 в течение 10 мин при 4°С и осадок бактерий снова суспендируют в 7,0 мл (для каждой библиотеки) 15% глицерин/2хYТ для хранения при -80°С. B. Индукция фага. Аликвоты М13К07 хелперного фага (Amersham Pharmacia Biotech) добавляют к оставшимся бактериальным культурам при OD600-0,5 (из раздела 2А, см. выше) до конечной концентрации 3 х 109 бое/мл. Оставляют хелперный фаг инфицировать бактерии при 37°С в течение 30 без встряхивания и 30 мин при слабом встряхивании. Инфицированные клетки центрифугируют при 5000 об./мин в течение 15 мин при 4°С. Клеточный осадок (пеллеты) снова суспендируют в том же самом объёме (из раздела 2А, см. выше) средой 2xYTAK (2YT с 100 ug/мл ампициллина и 40 ug/мл канамицина). Получение фагемид осуществляют при 30°С в течение ночи при встряхивании. C. Сбор фага. Бактериальные культуры из раздела 2В, см. выше, центрифугируют при 5000 об./мин в течение 15 мин при 4°С. Затем супернатанты переносят в новые пробирки, добавляют 0,2 объёма 20% ПЭГ/2,5М NaCl и инкубирую на льду в течение 1 ч, чтобы высадить фагемиды. Осаждённые фагемиды центрифугируют при 10000 об./мин 30 мин при 4°С и осторожно снова суспендируют со 100 мл холодного PBS. Раствор фагемид дополнительно очищают, отделяя центрифугированием оставшиеся клетки при 4000 об./мин в течение 10 мин при 4°С и осаждая фагемиды добавлением 0,2 объёма 20% ПЭГ/2,5М NaCl. Фагемиды центрифугируют при 10000 об./мин в течение 30 мин при 4°С и пеллеты фагемид снова суспендируют с 18 мл холодного PBS. 6 мл 60% раствора глицерина прибавляют к раствору фагемид для хранения при -80°С. Титры фагемид определяют по стандартной методике (Molecular Cloning, Maniatis et al., 3rd Edition). 3. Селекция Ang-2 связывающего фага. A. Иммобилизация Ang-2 на намагниченных бусах. - 66 - 008248 Биотинилированный Ang-2 (из раздела 3А, см. выше) иммобилизуют на Streptavidin M-280 Dyna-beads (DINAL, Lake Success, NY) с концентрацией около 2000 нг Ang-2 белка на 100 мкл массы бус от производителя. С помощью магнита бусины перемещают на одну сторону пробирки и пипеткой отбирают жидкость, бусы дважды отмывают фосфатно-солевым буферным раствором (PBS) и снова суспендируют в PBS. Биотинилированный Ang-2 белок добавляют к отмытым бусинам с вышеуказанной концентрацией и инкубируют при вращении в течение 1 ч при комнатной температуре. Бусы, покрытые Ang-2, затем блокируют, добавляя BSA до конечной концентрации 2%, и инкубируют в течение ночи при 4°С при вращении. Затем полученные в результате покрытые Ang-2 бусы дважды отмывают PBST (PBS с 0,05% Tween-20) перед тем, как подвергнуть их селекции. B. Селекция с применением бус, покрытых Ang-2. Около 1000-кратного библиотечного эквивалента фагемид (из раздела 2С, см. выше) блокируют в течение 1 ч с помощью 1 мл PBS, содержащего 2% BSA. Осуществляют три стадии негативной селекции блокированного образца фагемид, добавляя его к чистым бусам (те же бусы, что и в разделе 3А, но не покрытые белком Ang-2), и эту смесь инкубируют при комнатной температуре в течение 15 мин при вращении. Содержащий фагемиды супернатант отбирают с помощью магнита и переносят во вторую пробирку, содержащую чистые бусы (те же бусы, что и описанные в разделе 3А, но не покрытые белком Ang-2), и эту смесь инкубируют при комнатной температуре в течение 15 мин при вращении. Процедуру повторяют. Содержащий фагемиды супернатант отбирают с помощью магнита и переносят в новую пробирку, содержащую бусы, покрытые белком Ang-2 (из раздела 3А), и эту смесь инкубируют при комнатной температуре в течение 1 ч при вращении. После того как супернатант отбрасывают, бусы, связанные с фагемидами, 10 раз отмывают с помощью 2% молоко-РВS; 10 раз с помощью 2% BSA-PBS; 10 раз с помощью PBST и дважды с помощью PBS. Затем фагемиды элюируют в 1 мл 100 мМ раствора триэтиламина (Sigma, St. Louis, МО) в течение 10 мин на шюттль-аппарате. рН раствора, содержащего фагемиды, нейтрализуют, добавляя 0,5 мл 1М Tris-HCl (pH 7,5). Полученные фагемиды используют для инфицирования 10 мл свежевыращенных бактерий XL1-BLUE MRF' (OD600 около 0,5) при 37°С в течение 30 мин без встряхивания и 30 мин при медленном встряхивании. Все инфицированные XL1-BLUE MRF' клетки затем помещают в плашку 15х15 см 2xYTAG и инкубируют при 30°С в течение ночи. C. Индукция и сбор фага. Аликвоту 10 мл среды 2xYTAGT добавляют в плашку (из раздела 3В) для повторного суспендиро-вания XL1-BLUE MRF' клеток. Все XL1-BLUE MRF' клетки собирают в пробирку, аликвоту 250 мкл этих клеток добавляют к 25 мл 2xYTAGT и выращивают при 37°С до OD600=0,5. M13KO7 хелперный фаг добавляют до конечной концентрации 3х10 КОЕ/мл и инкубируют при 37°С в течение 30 мин без встряхивания и 30 мин при медленном встряхивании. Клетки центрифугируют при 5000 об./мин в течение 10 мин при 4°С и снова суспендируют с 25 мл 2xYTAK. Эти бактерии выращивают при 30°С в течение ночи при встряхивании. Индуцированные фагемиды собирают и очищают как в разделе 2С. D. Второй раунд селекции. Второй раунд селекции проводят, как подчёркнуто в разделе 3В-3С, за исключением следующего. Около 100-кратного библиотечного эквивалента фагемид, полученных в разделе 3С, используют в качестве исходных фагемид. Количество биотинилированного белка Ang-2 (раздел 3А), нанесённого на бусы Dynabead M-280 Streptavidin, уменьшают до 20 нг. Затем связанные с фагом бусы отмывают 10 раз с помощью 2% молоко-РВS; 10 раз - 2% BSA-PBS; 10 раз - PBST, при этом последнее отмывание включает 60-минутную инкубацию при комнатной температуре в PBST. Бусы дважды отмывают PBS. Условия элюции те же, что и в первом раунде (раздел 3В). E. Второй раунд селекции. Третий раунд селекции проводят, как подчёркнуто в разделе 3В-3С, за исключением следующего. Около 10-кратного библиотечного эквивалента фагемид, полученных в разделе 3D, используют в качестве исходных фагемид. Около 2 нг биотинилированного белка Ang-2 (из раздела 3А) используют для нанесения на бусы Dynabead M-280 Streptavidin. Связанные с фагом бусы отмывают 10 раз с помощью 2% молоко-РВS; 10 раз - 2% BSA-PBS; 10 раз - PBST, при этом последнее отмывание включает 60-минутную инкубацию при комнатной температуре в PBST. Бусы дважды отмывают PBS. Условия элюции те же, что и в первом раунде (раздел 3В). F. Четвёртый раунд селекции. Четвёртый раунд селекции проводят, как подчёркнуто в разделе 3В-3С, см. выше, за исключением следующего. Библиотечный эквивалент фагемид, полученных в разделе 3Е, используют в качестве исходных фагемид. Количество биотинилированного белка Ang-2 (раздел 3А), нанесённого на Dynabead M-280 Streptavidin, уменьшают до 0,4 нг для библиотек 25, 26 и 27. Для библиотек 20 и 94 количество нанесения сохраняется такое же, как и в третьем раунде, 2 нг. Библиотека 99 не участвует в четвёртом раунде стадии селекции. Условия элюции те же, что и в первом раунде (раздел 3В). - 67 - 008248 4. Клональный анализ. A. Приготовление эталонного планшета. Отбирают одиночные колонии из второго раунда селекции и инокулируют в 96-луночные планшеты, содержащие 120 мкл 2xYTAGT в лунке. 96-луночные планшеты инкубируют при 30°С в шюттль-аппа-рате в течение ночи. В каждую лунку прибавляют 40 мкл 60% глицерина и хранят при -80°С. B. Анализ фагемид методом ELISA. Аликвоты около 2 мкл клеток из эталонного планшета (из раздела 4А см. выше) инокулируют в свежий Costar 96-луночный планшет (Corning incorporated, Corning, NY, cat.# 69794), который содержит 100 мкл 2xYTAGT в лунке, и клетки в этом новом планшете выращивают при 37°С примерно до OD600=0,5. 13 40 мкл 2xYTAGT, содержащей М13Ю7 хелперный фаг (1,5 х 1013 КОЕ/мл), добавляют в каждую лунку и 96-луночный планшет инкубируют при 37°С в течение 30 мин без встряхивания и ещё в течение 30 мин при слабом встряхивании. Планшет центрифугируют при 2000 об./мин (настольная центрифуга Beckman CS-6R) в течение 10 мин при 4°С. Супернатанты удаляют из лунок и каждый клеточный осадок (пеллеты) снова суспендируют, используя 150 мкл 2xYTAK на лунку. Планшет инкубируют при 30°С в течение ночи для экспрессии фагемид. Человеческий Ang-2 белок наносят на 96 луночный планшет Maxisorp (NUNC) при 1 мкг/мл в 1xPBS при 4°С в течение ночи. В качестве контроля 2% BSA (Sigma) наносят на отдельный Maxisorp планшет. На следующий день стоявшие в течение ночи клеточные культуры центрифугируют при 2000 об./мин в течение 10 мин при 4°С. 10 мкл супернатанта из каждой лунки переносят в новый 96-луночный планшет, содержащий раствор BSA/PBS для разведения супернатанта 1:10. Полученные смеси инкубируют в течение 1 ч при комнатной температуре при встряхивании для блокирования фагемид. В то же время планшет, покрытый белком А^-2, блокируют 400 мкл 2% раствора BSA/PBS на лунку в течение 1 ч при комнатной температуре при встряхивании. Раствор BSA отбрасывают и каждую лунку трижды отмывают раствором PBS. После последней стадии отмывания в каждую лунку планшета, покрытого белком Ang-2, а также контрольного планшета добавляют по 100 мкл раствора блокированных фагемид и инкубируют в течение 1 ч при комнатной температуре при встряхивании. Жидкость отбрасывают, а каждую лунку трижды отмывают раствором PBST. 100 мкл HRP-конъюгированного анти-М13 mAb (мышиного антитела) (Amersham Pharmacia Biotech) при разведении 15000 добавляют в каждую лунку планшета, покрытого белком Ang-2, а также контрольного планшета, и эти планшеты инкубируют в течение 1 ч при комнатной температуре при встряхивании. Жидкость снова отбрасывают, а каждую лунку трижды отмывают раствором PBST. 100 мкл хемилюминесцентных субстратов LumiGLO (Kirkegaard & Perry Laboratories, Gaithersburg, MD) добавляют в лунки и каждую лунку прочитывают на приборе Luminoskan Ascent DLRearly machine (Labsystems, Franklin, MA). C. Секвенирование фаговых клонов. Реакцию ПЦР проводят, используя в качестве темплата 1 мкл бактерий из каждой лунки эталонного планшета (раздел 4А). Объём каждой ПЦР смеси составляет 50 мкл, и в него входит 1 х буфера для ПЦР, 300 нМ каждого из двух праймеров: 5'-GTTAGCTCACTCATTAGGCAC-3' (SEQ ID NO: 256) и 5'-GTACCGTAACACTGAGTTTCG-3', (SEQ ID NO: 257); 200 мкM dNTP, 2 мМ MgCl2 и 2,5 Ед (U) taq ДНК полимеразы (Roche Molecular Biochemicals). Систему GeneAmp PCR System 9700 (Applied Biosystems) используют для осуществления следующей программы: 94°С за 5 мин; 40 циклов (94°С за 45 с, 55°С за 45 с, 72°С за 90 с); 72°С за 10 мин; охлаждение до 4°С. ПЦР продукт очищают с помощью набора QIAquick 96 PCR Purification Kit (QIAGEN Inc.) в соответствии с рекомендациями производителя. Все очищенные ПЦР продукты секвенируют с применением праймера 5'-TTACACTTTATGCTTCCG-3' (SEQ ID NO: 258), используя секвенатор ABI 3770 Sequencer (Perkin Elmer) в соответствии с рекомендациями производителя. 5. Ранговое распределение последовательностей. Пептидные последовательности, транслированные при использовании нуклеотидных последовательностей (из раздела 4С, см. выше) коррелируют с данными ELISA. Клоны, которые показывают высокую OD при прочитывании в лунках, покрытых Ang-2, и низкую OD при прочитывании в лунках, покрытых BSA, считаются важными. 24 пептидных последовательности из библиотеки 20, 26 пептидных последовательностей из библиотеки 94, 7 пептидных последовательностей из библиотеки 25, 18 пептидных последовательностей из библиотеки 26, 6 пептидных последовательностей из библиотеки 27 и 4 пептидных последовательности из библиотеки 99 выбирают для дальнейшего анализа и образования "пепти-тел". Кроме того, 11 согласованных последовательностей из библиотек 20 и 94, 3 согласованные последовательности из библиотек 26 и 99 и 2 из библиотеки 25 выведены и использованы для образования "пептител". "Пептитела" в табл. 7 оценивают, используя протокол ELISA с нейтрализацией, описанный в примере 10 в данном описании. Результаты показаны в табл. 7. - 68 - 008248 Таблица 7 Зрелые по аффинности Pbs ("пептитела") из Соп4 hAng-2:Tie2 ICso (нМ) Последовательность "пептитела" (Seq Id No:) Соп4-44 (С) 0.09 M-Fc-GGGGGAQ-PRQmCDTOPWTCEHMWEV-LE (SEQ ID NO: 259) Соп4-40 (С) 0.10 M-Fc-GGGGGAQ-OTQmCEWDPWTC^HMPGK-LE (SEQ ID NO: 260) Соп4-4 (С) 0.12 M-Fc-GGGGGAQ- WYEQDACEWDPWTCEHMAEV-LE (SEQ 3D NO: 261) Соп4-31 (С) Э.16 M-Fc-GGGGGAQ-NT 116 M-Fc-GGGGGAQ-AATQEECEWDPWTCEHMPRS-LE (SEQ ID NO: 263) Соп4-42 (С) ( )-17 M-Fc-GGGGGAQ-LRHQEGCEWDPWTCEHIvrEDW-LE (SEQ ID NO: 264) Соп4-35 (С) 0.18 M-Fc-GGGGGAQ-WRQKDCEWDPWTCEHMYVG-LE .(SEQ ID NO: 265) Соп4-43 (С) 0.18 M-Fc-GGGGGAQ-SISHEECEWDPWTCEHMQVG-LE (SEQ ГО NO: 266) Соп4-49(С) 0.19 M-Fc-GGGGGAQ- WAAQEECEWDPWTCEHMGRM-LE (SEQ ID NO: 267) Соп4-27 (С) 0.22 M-Fc-GGGGGAQ-TWPQDKCETOPWTCEHMGST-LE (SEQ ID NO: 268) Соп4-48 (С) 0.26 M-Fc-GGGGGAQ-GHSQШCGWDPWTCEHMGTS-LE (SEQ ID NO: 269) Соп4-46 (С) 0.26 M-Fc-GGGGGAQ-QHWQEECEWDPWTOTHMPSK-LE (SEQ Ш NO: 270) Соп4-41 (С) 0.26 M-Fc-GGGGGAQ-NWQEKCEWDPWTCEHMPVR-IJE (SEQ ID NO: 271) Соп4-36 (С) 0.28 M-Fc-GGGGGAQ-KSGQVECNWDPWTCEHMPRN-LE (SEQ Ш NO: 272) Сог.4-34 (С) 0.28 M-Fc-GGGGGAQ-VKTQEHCDWDPWTCEHMREW-LE (SEQ Ш NO: 273) Соп4-28 (С) 0.30 M-Fc-GGGGGAQ- AWGQEGCDWDPWTCEHMLPM-LE (SEQ ID NO: 274) Соп4-39(С) 0.30 M-Fc-GGGGGAQ-PVNQEDCEWDPWC^HMPPM-LE (SEQ 3D NO: 275) Соп4-25 (С) 0.31 M-Fc-GGGGGAQ-RAPQEDCEWDPWTCAHMDIX-LE (SEQ ID NO: 276) - 69 - 008248 Con4-50(C) 0.38 . M-Fc-GGGGGAQ-HGQNIVffiCEWDPWTCEHMFRY-LE (SEQ Ш NO: 277) Соп4-38 (С) D.40 M-Fc-GGGGGAQ-PRLQEECVWDPWTCEHMPLR-LE (SEQ 3D NO: 278) Соп4-29 (С) Э.41 M-Fc-GGGGGAQ-(SEQ ГО NO: 279) Соп4-47 (С) 3.44 M-Fc-GGGGGAQ-QTSQEDCVWDPWTCDHMVSS-LE (SEQ 3D NO: 280) Соп4-20 (С) 3.48 M-Fc-GGGGGAQ-QVIGRPCEWDPWTCEECLEGL-LE (SEQ Ю NO: 281) Соп4^5 (С) ( ).48 M-Fc-GGGGGAQ- WAQQEECAWDPWTCD33MVGL-IJE (SEQ 3D NO: 282) Соп4-37 (С) ( ).49 M-Fc-GGGGGAQ-LPGQEDCEWDPWTCEHMVIIS-IJE (SEQ 3D NO: 283) Соп4-33(С) С ).52 M-Fc-GGGGGAQ-PMNQVECDWDPWTCEHMPRS-LE (SEQ Ю NO: 284) АС2-Соп4 (С) С ).52 M-Fc-GGGGGAQ-FGWSHGCEWDPWTCEFfMGST-LE (SEQ Ш NO: 285) Сол4-32 (С) С ).75 M-Fc-GGGGGAQ-KSTQDDCDWDPWTCEHMVGP-LE (SEQ ID NO: 286) Соп4-17(С) 0.96 M-Fc-GGGGGAQ-GPMSTCQWDPWTC33HMDQL-LE (SEQ 3D NO: 287) Соп4-8 (С) 1.20 M-Fc-GGGGGAQ-ST1GDMC33WDPWTCAHMQVD-]1E (SEQ 3D NO: 288) АС4-Соп4 (С) 1.54 M-Fc-GGGGGAQ-VLGGQGCEWDPWTCRLLQGW-LE (SEQ ID NO: 289) Соп4-1 (С) 2.47 M-Fc-GGGGGAQ-VLGGQGCQWDPWTCSHIJEDG--LE (SEQ 3D NO: 290) Соп4-С1 (С) 2.75 M-Fc-GGGGGAQ-TTIGSMCEWDPWTCAHMQGG-LE (SEQ ID NO: 291) Соп4-21 (С) 3.21 M-Fc-GGGGGAQ-TKGKSVCQWDPWTCSHMQSG-LE (SEQ 3D NO: 292) Соп4-С2 (С) 3.75 M-Fc-GGGGGAQ-TTIGSMCQWDPWTCAHMQGG-LE (SEQ ID NO: 293) Соп4-18 (С) 4.80 M-Fc-GGGGGAQ-WV^WCEWDPWCr^WDTP^LE (SEQ ID NO: 294) Соп4-19 (С) 5.76 M-Fc-GGGGGAQ- WQVGMCQWDPWTCimMRLQ-LJE (SEQ ГО NO: 295) Соп4-16 (С) 6.94 M-Fc-GGGGGAQ-AVGSQTCEWDPWTCAHLVEV-LE (SEQ 3D NO: 296) Соп4-11 (С) 9.70 M-Fc-GGGGGAQ-QGMKMFCEWDPWTCAHIVYR-LE (SEQ Ш NO: 297) Соп4-С4 (С) 9.80 M-Fc-GGGGGAO-TTTGSMCQWDPWTCEHMQGG-LE (SEQ ID NO: 298) Соп4-23 (С) 9.88 M-Fc-GGGGGAQ-TSQRVGC35WDPWTCQHLTYT-3LE (SEQ Ю NO: 299) Соп4-15 (С) 15.00 M-Fc-GGGGGAQ-QWSWPPCEWDPWTCQTVWPS-I,E (SEQ ГО NO: 300) Con4-9 (С) 20.11 M-Fc-GGGGGAQ-GTSPSFCQWDPWTCSHMVQG-IJB (SEQ 3D NO: 301) Соп4-10 (С) 86.61 M-Fc-GGGGGAQ-TQGLHQCEWDPWTCKVLWPS-LE (SEQ 3D NO: 302) Соп4-22 (С) 150.00 M-Fc-GGGGGAQ- VWRSQVCQWDPWTCNLGGDW-LE (SEQ 3D NO: 303) Соп4-3 (С) 281.50 M-Fc-GGGGGAQ-DIOLEECQWDPWTCQFFYGA-LE (SEQ 3D NO: 304) - 70 - 008248 Соп4-5 (С) No Inhibition M-Fc-GGGGGAQ-ATFARQCQWDPWTCALGGNW-LE (SEQ ID NO: 305) Соп4-30 (С) No Inhibition M-Fc-GGGGGAQ-GPAQEECEWDPWTCEPLPLM-LE (SEQ ID NO: 306) Соп4-26 (С) No Inhibition M-Fc-GGGGGAQ-(SEQIDNO:307) Соп4-7 (С) No Inhibition M-Fc-GGGGGAQ-LWQLAVCQWDPQTCDECMGAL-LE (SEQ ID NO: 308) Соп4-12 (С) No Inhibition M-Fc-GGGGGAQ-TQLVSLCEWDPWTCRLLDGW-LE (SEQ Ш NO: 309) Соп4-13 (С) No Inhibition M-Fc-GGGGGAQ- MGGAGRCEWDPWTCQLLQGW-LE (SEQ Ш NO: 310) Соп4-14 (С) No Inhibition M-Fc-GGGGGAQ- MFLPNECQWDPWTCSNLPEA-LE (SEQIDNO:3Il) Соп4-2 (С) No Inhibition M-Fc-GGGGGAQ- , FGWSHGCF^VDPWTCKLLQGW-LE (SEQ Ш NO: 312) Соп4-6 (С) No Inhibition M-Fc-GGGGGAQ-WPQTEGCQWDPWTCRIXHGW-LE (SEQ Ш NO: 313) Соп4-24 (С) No Inhibition M-Fc-GGGGGAQ-PDTRQGCQWDPWTCRLYGMW-LE (SEQ ID NO: 314) АС1-Соп4 (С) [STo Inhibition M-Fc-GGGGGAQ-TWPQDKCEWDPWTCRLLQGW-LE (SEQ ГО NO: 315) АСЗ-Соп4 (С) NFo Inhibition M-Fc-GGGGGAQ- DKTT .EECEWDPWTCRLLQGW-LE (SEQ ID NO: 316) АС5-Соп4 (С) 4o Inhibition M-Fc-GGGGGAQ-AATQEECEWDPWTCRLLQGW-LE (SEQ 3D NO: 317) Зрелые по аффинности Pbs ("пептитела") из L1 hAng-2:Tie2 IC50 (нМ) Последовательность "пептитела" (Seq Id No:) MGAQ- TNFMPMDDLEQRLYEQFILQQG- LEGGGGG-Fc (SEQ 3D NO: 318) L1-7(N) 0.03 AC6-L1 (N) 0.03 MGAQ TNiTO^LDIiLDATLYEHWILQHS LEGGGGG-Fc (SEQ ГО NO: 319) Ll-15 (N) 0.04 MGAQ- QKYQPIJDE3LDKTLYDQFMLQQG LEGGGGG-Fc (SEQ 3D NO: 320) Ll-2 (N) 0.04 MGAQ-LNFTPLDELEQTLYEQWTLQQS LEGGGGG-Fc (SEQ 3D NO: 321) Ll-10 (N) 0.05 MGAQ- QKPQP3JDELEQTLYEQFMLQQA LEGGGGG-Fc (SEQ 3D NO: 322) . Ll-13 (N) 0.05 MGAQ- QEYEPLDELDETLYNQWMFHQR LEGGGGG-Fc (SEQ 3D NO: 323) L1-5(N) 0.05 MGAQ-VKYKTIJDEIJDEELYEQQTFQER LEGGGGG-Fc (SEQ 3D NO: 324) L1-C2 (N) 0.05 MGAQ- TJCFQPLDELDQTLYEQWTLQQR LEGGGGG-Fc (SEQ ГО NO: 325) L1-C3 (N) 0.06 MGAQ- TNFQPLDELDQTLYEQWTLQQR LEGGGGG-Fc (SEQ Ш NO: 326) L1-11(N) • 0.07 MGAQ- QNFKPMDELEDTLYKQFLFQHS LEGGGGG-Fc (SEQ Ю NO: 327) - 71 - 008248 Ll-17 (N) 0.08 MGAQ- VKYISPLDELDEWLYHQFTLFfflQ LEGGGGG-Pc (SEQ 3D NO: 328) u.us MGAQ- YKT^TmDLEQTLYEQWTLQHV LEGGGGG-Fc ^SEQIBNOT329) L1-1(N) 0.08 MGAQ-QNY^mEmATLYEffiTT^TxT LEGGGGG-Fc (SEQ 3D NO: 330) L1-4(N) 0.08 MGAQ- VJCFKPLDALEQTXYEIIWMFQQA LEGGGGG-Fc (SEQ ID NO: 331) LI-20 (N) 0.09 MGAQ- EDYMPLDALDAQLYEQFTLLHG LEGGGGG-Fc (SEQ 3D NO: 332) Ll-22 (N) 0.09 MGAQ- YI^SIPMDELEQTLYEEFLFQHA LEGGGGG-Fc (SEQ 3D NO: 333) 0.1Г MGAQ- SNFMPLDELEQTLYEQFMLQHQ LEGGGGG-Fc (SEQ 3D NO: 334) Ll-16 (N) Э.11 MGAQ- Q3^QPII> ELEETLYKQWTLQQR LEGGGGG-Fc (SEQ 3D NO: 335) L1-18(N) 3.16 MGAQ-Q3^MDPLDELDE3LYEQFMFQQS 3JEGGGGG-Fc (SEQ 3D NO: 336) L1-3(N) ( 3.16 MGAQ- TKT^NPLDELEQTLYEQWTLQHQ LEGGGGG-Fc (SEQ 3D NO: 337) L1-21(N) С ).17 MGAQ- IITFQPLDEL3iETLYYQWLYDQL LEGGGGG-Fc (SEQ 3D NO: 338) LI-CI (N) 0.56 MGAQ- QHFKPLDELEQTLYEQWTLQQR LEGGGGG-Fc (SEQ 3D NO: 339) Li--i9-^7 1.26 MGAQ- QTFQPLDDLEEYLYEQW3RRYH LEGGGGG-Fc {SEQ E> N0r34(c)) H-9(N) 1.62 MGAQ- SKFKPLDELEQTLYEQWTXQHA LEGGGGG-Fc (SEQ 3D NO: 341) Зрелые по аффинности Pbs ("пептитела") из Conl 1 hAng-2:Tie2 IC50 (нМ) Последовательность "пептитела" (Seq Id No:) Conl^t (C) 1.68 M-Fc-GGGGGAQ-SGQLRPCEEIFGCGTQNLAL-LE (SEQ 3D NO: 342) Conl-1 (C) 3.08 M-Fc-GGGGGAQ- AGGMRPYDG> Y1LG?T;PNYDVQA-LE (SEQ 3D NO: 343) Conl-6 (C) 8.60 M-Fc-GGGGGAQ-GQDLRPCEDMFGCGTKDWYG-LE (SEQ 3D NO: 344) Conl-3 (C) 16.42 M-Fc-GGGGGAQ-APGQRPYDGMLGWPTYQPJV-LE (SEQ 3D NO: 345) Conl-2 (C) No 3nhibition M-Fc-GGGGGAQ-QTWDDPCMEIILGPVT^VRRCI-LE (SEQ 3D NO: 346) Conl-5 (C) No Inhibition M-Fc-GGGGGAQ-FGDICRPLECMFGGPIQLCPR-LE (SEQ 3D NO: 347) Parent: Conl (C) 26.00 M-Fc-GGGGGAQ-3SRPC33E3FGGCTYQ-LE (SEQ Ш NO: 348) - 72 - 008248 Зрелые по аффинности Pbs ("пептитела") из 12-9 hAng-2:Tie2 1С50 (нМ) Последовательность "пептитела" (Seq Id No:) 12-9-3 (С) 0.81 M-Fc-GGGGGAQ-LQEWCEGVEDPFTFGCEKQR-LE (SEQ ID NO: 349) 12-9-7 (С) 0.93 M-Fc-GGGGGAQ-^^YCEGmBPFT^GGDKQM-fcE (SEQ ID NO: 350) 12-9-6 (С) 0.95 M-Fc-GGGGGAQ-HQEYCEGMEDPFTFGCEYQG-LE (SEQ ID NO: 351) 12-9-С2 (С) 1.41 M-Fc-GGGGGAQ-LQDYCEGVEDPFTFGCENQR-LE (SEQ ID NO: 352) 12-9-5 (С) 1.56 M-Fc-GGGGGAQ-LIJDYCEGVQDPFTFGCENLD-LE (SEQ ID NO: 353) 12-9-1 (С) 1.84 M-Fc-GGGGGAQ-GFEYCDGMEDPFTFGCDKQT-LE (SEQ ГО NO: 354) 12-9-4 (С) 2.05 M-Fc-GGGGGAQ-AQDYCEGMEDPFTFGCEMQK-LE (SEQ ID NO: 355) 12-9-С1 (С) 2.68 M-Fc-GGGGGAQ-LQDYCEGVEDPFTFGCEKQR-LE (SEQ ID NO: 356) 12-9-2 (С) 8.42 M-Fc-GGGGGAQ-KLEYCDGIVIEDPFTQGCDNQS-UE (SEQ Ш" NO: 357) Parent: 12-9 (С) 15.00 M-Fc-GGGGGAQ-FDYCEGVEDPFTFGCDNH-LE (SEQ ID NO: 358) Пример 9. Шесть примеров anra-Ang2 "пептител" испытывают на активность связывания с huAng2 (R &D Systems, BNO12103A) на BIAcore. Протеин G иммобилизуют на СМ5 датчике в соответствии со стандартным протоколом взаимодействия амина (BIAcore Inc.), а "пептитела" затем инъецируют через поверхность протеина G для захвата (RL ~ 100 Ru). Для испытания связывания между huAng2 и захваченным "пептителом" 0,3-40 нМ huAng2 инъецируют через поверхности захваченных "пептител" и отражающие связывание показания датчиков анализируют, используя программу BIAevaluation (BIAcore Inc.). В табл. 8 суммированы результаты этого эксперимента. Таблица 8 "Пептитело" Lot# KD(M) ka (1/Ms) kd (1/s) Соп4-44 (С) 011702 2.1Е-10 2.9E+05 5.9E-05 Ll-7 (N) 022102 2.4Е-10 3.7E+05 8.7E-05 L1-10(N) 021302 7.7Е-10 1.5E+05 1.1E-04 Ll-21 (N) 021802 2.4Е-10 5.6E+05 1.4E-04 Соп4 (С) 3345677 3.8Е-10 5.3E+05 2.0E-04 2хСоп4 (С) 1К 092501 3.4Е-10 4.8E+05 1.6E-04 Пример 10. Нейтрализационный метод ELISA. Кондиционированные среды человеческого, мышиного, суш и крысиного Ang-2 и человеческого и мышиного Ang-1 разводят в DМЕМ/50 мкг/мл BSA следующим образом: hAng-2-1:64 разведение; mAng-2-l:64 разведение; крысиный Ang-2-без разведения; cyno Ang-2-1:32 разведение; hAng-1-1:4 разведение; и mAng-1-1:4 разведение. Степень разведения каждой из этих кондиционированных сред определяют по их способности связываться с 1 нМ hTie2-Fc (при условии, что в молекуле Tie-2-Fc область Tie-2 содержит только растворимый внеклеточный домен молекулы; R &D Systems, номер в каталоге 313 TI) с величиной 50% максимально достижимого связывания (т.е плато). Микротитрационные планшеты покрывают с помощью 100 мкл разведённых кондиционированных сред. Для Ang-2 по нейтрализационному методу ELISAs возможные анти-Ang-2 "пептитела" (кандидаты) титруют от 2,5 нМ до 0,015 пМ в четырёхкратных разведениях в растворе PBS, содержащем около 1% BSA и около 1 нМ Tie-2 (при условии, что в молекуле Tie-2-Fc область Tie-2 содержит только растворимый внеклеточный домен молекулы; R &D Systems, номер в каталоге 313 TI). Для Ang-1 нейтрализационного ELISAs, возможные (кандидаты) anra-Ang-2 "пептитела" - 73 - 008248 титруют от 1000 нМ до 0,2 пМ в четырёхкратных разведениях в растворе PBS, содержащем около 1% BSA и около 1 нМ Tie-2 (при условии, что в молекуле Tie-2-Fc область Tie-2 содержит только растворимый внеклеточный домен молекулы; R &D Systems, номер в каталоге 313 TI). После добавления примерно 100 мкл раствора "пептитело"/Tie-2 в каждую лунку планшеты инкубируют в течение ночи при комнатной температуре, а затем отмывают 5 раз в PBS, содержащем около 0,1% Tween-20. После отмывания добавляют около 100 мкл на лунку анти-^е-2 антитела (Pharmingen Inc., catalog #557039) до примерной конечной концентрации 1 мкг/мл и планшеты инкубируют около 1 ч при комнатной температуре. Далее около 100 мкл/лунка антитела козы против мышиного IgG-HRP (Pierce Chemical Co., catalog #31432) добавляют в разведении 1:10000 в PBS, содержащем около 1% BSA. Планшеты инкубируют при комнатной температуре примерно 1 ч, после чего их отмывают 5 раз PBS, содержащем около 0,1% Tween-20. Затем добавляют в каждую лунку по 100 мкл субстрата ТМВ (SIGMA, catalog # T8665) и ожидают появления голубого цвета. Затем на спектрофотометре (ридере) прочитывают оптическую плотность при 370 нм. Результаты представлены в табл. 9, см. ниже. Таблица 9 Опосредуемая "пептителом" нейтрализация взаимодействий ангиопоэтин:^е2 hAng-2 mAng-2 rAng-2 cAng-2 hAng-1 m Ang-1 Пептитело IC50 IC50 IC50 ICso ICso ICso (нМ) (нМ) (нМ) (нМ) (нМ) (нМ) 2хСоп4 (С) 0.026 0.035 0.024 0.047 3.0 3.2 Соп4 (С) 0.197 0.289 0.236 0.540 200 300 Соп4-44 0.08 0.16 0.22 (С) Соп4-40 0.20 0.27 0.35 > 1000 (С) L1-7 (N) 0.046 0.063 0.035 0.108 > 1000 > 1000 L1-10(N) 0.06 0.06 0.06 > 1000 Пример 11. Дизайн исследования. Самцов мышей CD-1 весом 20-30 г произвольно разделяют на группы, получающие определённые "пептитела" (2xCon4-C, L1-7-N и L1-21-N). Животным вводят разовую IV (в.в.) болюсную (^38/группа) или разовую SC (подкожную) инъекцию 50 мкг "пептитела" (n=34/группа). Инъекции делают в хвостовую вену и под кожу в плечо для IV и SC введения соответственно. Отбор проб крови и аналитические методы. Пробы крови отбирают для измерения каждой концентрации am^Ang-2 "пептитела" перед введением дозы и через 1, 2, 4, 8, 16, 24, 48, 72, 96, 120, 144, 168, 216, 264, 312 и 336 ч после введения для SC и IV (в. в.) групп. Дополнительные пробы отбирают через 5 и через 30 мин для IV групп. Одновременно берут пробу крови у двух животных и по окончании отбора проб животных умерщвляют. Кровь (примерно 0,50 мл) отбирают пункцией перикарда в полипропиленовые пробирки Микротейнер(r) для сепарации сыворотки. Пробы хранят на льду примерно 20 или до образования сгустка. Сыворотку отделяют от образца крови центрифугированием в течение примерно 10 мин при 2-8°С и хранят примерно при -70°С до момента анализа. Измерение в образцах проводят методом флуоресценции с разрешением по времени (TRF) с нижним пределом квантификации (LLOQ) 100 нг/мл. Микротитрационные планшеты NUNC fluoroMaxisorp покрывают рекомбинантным мышиным белком Ang-2. Затем планшеты блокируют раствором белка с целью уменьшить неспецифическое связывание. Эталонные образцы, образцы для качественного контроля и неизвестные образцы готовят в 10% буфере для анализа мышиной сыворотки и пипеткой вносят в лунки микротитрационных планшетов. "Пептитела" специфически связываются с иммобилизованным Ang-2. После отмывания всех несвязанных веществ (Kirkegaard & Perry Laboratories Inc.) в лунки добавляют биотинилированное моноклональное антитело козы к человеческому IgG (H+L) (Jackson ImmunoResearch Laboratories Inc.). После стадии отмывания для удаления всех несвязанных био-тинилированных моноклональных антител в лунки добавляют стрептавидин, меченый европием, связанный европий выделяют из стрептавидина с помощью кислого раствора, пипеткой вносимого в каждую лунку. Сигнал флуоресценции получают и читают на флуорометрическом ридере Wallac. Анти-Ang-2 " пептитело" анализируют в интервале 0,078-5 мкг/мл. Фармакокинетический анализ. Сложные средние данные зависимости концентрация-время для каждой группы анализируют с помощью "некомпартментального" анализа, используя WinNonlin Professional (Version 3.3; Pharsight Crp., Mountain View, CA). Для РК анализа используют номинальное время выборки, так образцы отбирают в течение 10% номинального времени. Всем концентрациям ниже LLOQ перед РК анализом задают нулевые значения. Рассчитывают следующие РК параметры. - 74 - 008248 T _Ь(2) 1/2--1- Конечный период полужизни (ti/г) рассчитывают как ке\ , где kei обозначает константу ко- нечной скорости первого порядка, рассчитываемой по уравнению линейной регрессии конечной фазы log-линейного затухания. Область (площадь поверхности) под кривой концентрация в сыворотке-время (AUC(0-last)) рассчитывают по линейному/log методу трапеций от времени 0 до last, времени последней поддающейся расчёту концентрации (Clast). Область (площадь поверхности) под кривой от 0 бесконечности (AUC(0-00)) рассчитывают как сумму значений соответствующего AUC(o-iast) и предсказанного Ciast/kei: лиг ^дттг ,?*edictedClast АиЦо-оо ) ~ AUC(o-iast)+ " Абсолютную биодоступность (F) после SC введения рассчитывают как F=AUC(0^SC х\00 AUC^_myV Результаты представлены на фиг. 2. Пример 12. Самкам бестимусных ("голых") мышей инъецируют подкожно 1х107 А431 клеток в день исследования 0. В день 3 Ang-2 "пептитело" 2ХСОП4-С вводят подкожно в дозе 200 мкг/мышь/день. Через регулярные временные интервалы регистрируют объёмы опухолей и вес тела, как показано на фигуре. Наблюдают заметное различие роста опухоли между группой, получающей Ang-2 " пептитело", относительно группы, получающей в качестве контроля носитель, и группы, получающей контрольное "пептитело", (р <0,0001 по сравнению (vs) с каждой контрольной группой при использовании повторного измерения ANOVA, с post hoc (последующим) тестом Шеффе (Scheffe)). Лечение этим "пептителом" не оказывает значительного влияния на вес тела. Результаты представлены на фиг. 3. Пример 13. Кривая роста А431 In vitro. Клетки А431 засевают в 96-луночные планшеты для тканевых культур, по 2000 клеток в лунке, в 200 мкл среды DMEM, дополненной 10% фетальной бычьей сывороткой (FBS). Через 16 ч после посева среду отсасывают. Затем в лунки добавляют следующее (и устанавливают в тройном повторе): 100 мкл/лунка DMEM, 10% FBS, 1 мг/мл негативного контрольного "пептитела" 4883 или "пептитела" TN8-Con4. Такие же комплекты повторяют на 5 планшетах. Среду из одного планшета отсасывают через 24, 48, 72, 96 и 120 ч после обработки. Затем добавляют 100 мкл 10% трихлоруксусной кислоты (ТСА) на лунку и планшеты хранят при 4°С. Всё содержимое планшетов собирают, когда 10% ТСА находится в последнем планшете, как минимум, 4 ч. 10% ТСА отбрасывают и лунки промывают 5 раз водопроводной водой. Затем клетки окрашивают с помощью 100 мкл 0,4% сульфородамином В (Sigma S-9012) в 1% уксусной кислоте (Sigma A-6283) в течение 10 мин при комнатной температуре, а затем 5 раз отмывают 1% уксусной кислотой. Затем планшеты сушат на воздухе. Краситель солюбилизируют, используя 300 мкл 20 мМ незабуференного Tris (pH> 10) в течение 2 ч на роторном шейкере. Затем прочитывают оптическую плотность (OD) при 540 нм на ридере для микротитрационных планшетов. Результаты представлены на фиг. 4. Пример 14. Самкам " голых" мышей подкожно инъецируют 2х106 клеток Colo-205 плюс Матригель (2:1) в день исследования 0. С дня 3 Ang-2 "пептитела" L1-7-N, L1-21-N, СОП4-С и 2ХСОП4-С вводят подкожно в дозе 14 мкг/мышь, дважды в неделю. Антитело против Ang-2 Ab536, 47 мкг/мышь, трижды в неделю вводят в качестве позитивного контроля. Через регулярные временные интервалы регистрируют объёмы опухолей и вес тела. Наблюдают заметное различие роста опухоли между каждой из групп, получающих Ang-2 " пепти-тело", по сравнению с группой, получающей в качестве контроля носитель, и группой, получающей контрольное "пептитело", (р <0,0001 по сравнению (vs) с каждой контрольной группой при использовании повторного измерения ANOVA, с post hoc (последующим) тестом Шеффе (Scheffe)). Лечение этими " пептителами" не оказывает значительного влияния на вес тела (результаты не показаны). Результаты представлены на фиг. 5. Пример 15. Самкам " голых" мышей подкожно инъецируют 2х106 Colo-205 клеток плюс Матригель (2:1) в день исследования 0. Начиная со дня 3, Ang-2 "пептитело" 2ХСОП4-С вводят подкожно в дозах 14, 2,8 и 0,56 мкг/мышь, дважды в неделю. Объёмы опухолей и вес тела записывают через регулярные интервалы, как показано. Наблюдают значительную разницу в росте опухоли между группой, получающей две наивысшие дозы Ang-2 " пептитела", и группами, получающими в качестве контроля носитель и контрольное " пепти-тело" (р=0,003 для промежуточной дозы и р <0,0001 для высокой дозы, при использовании повторного измерения ANOVA, с post hoc (последующим) тестом Шеффе (Scheffe)). Лечение этими "пептителами" не оказывает значительного влияния на вес тела. Пунктирной линией показано уменьшение общего n группы с 10 до 9 вследствие смерти одной из мышей по неизвестным причинам. Результаты представлены на - 75 - 008248 фиг. 6. Пример 16. Affra-Ang-2 "пептитела" против ксенотрансплантатов опухолей Colo-205. Самкам "голых" мышей подкожно инъецируют 2х106 клеток Colo-205 плюс Матригель (2:1) в день исследования 0. С дня 3 Ang-2 "пептитело" 2ХСОП4-С или контрольное "пептитело" вводят подкожно в дозе 350 мкг/день. Опухоли из групп, получавших контрольное "пептитело" (как описано в табл. 5), собирают либо в день 14 (контроль по размеру) или в день 18 (контроль по времени). Затем в день 18 собирают опухоли в группе, получающей 2ХСОП4(С). Через регулярные временные интервалы регистрируют объёмы опухолей, как показано. Наблюдают заметное различие роста опухоли между групп контроля по времени и группой, получающей в качестве контроля носитель, и группой, получающей 2ХСОП4-С, (р=0,0154 при использовании повторного измерения ANOVA, с post hoc (последующим) тестом Шеффе (Scheffe)). Лечение этими "пептителами" не оказывает значительного влияния на вес тела. Опухоли, приготовленные для анализа изображений, рассекают коронально на две половины и одну половину быстро замораживают в OCT (Sakura Finetek USA Inc., Torrance, CA). Криосрезы окрашивают иммуногистохимически, используя CD31 против иммуноглобулинов мыши (catalogue #553370, BD PharMingen, San Diego, CA) no 2 мкг/мл, с DAB в качестве хромогена. Делают цифровые фотографии срезов опухолей объективом с увеличением 20Х. Захватывают четыре поля по "компасным румбам", по десять опухолей на обрабатываемую группу. Систему анализа изображений MetaMorph (Universal Imaging Corporation, Downington, PA) применяют, чтобы увидеть (отразить) в изображении окрашенные CD31 кровеносные сосуды. Площадь CD31-позитивного окрашивания выражают в виде отношения к общей площади опухолевой ткани внутри каждого поля. Результаты представлены на фиг. 7. Пример 17. Самкам "голых" мышей подкожно инъецируют 2х106 Colo-205 клеток плюс Матригель (2:1) в день исследования 0. Лечение с помощью подкожного введения дважды в неделю 350 мкг/мышь Ang-2 "пеп-титела" 2ХСОП4-С или эквивалентного контрольного "пептитела" начинают в день исследования 3, 10 или 15. Объёмы опухолей и вес тела записывают через регулярные интервалы. Наблюдают значительную разницу в росте опухоли между всеми группами, получающими Ang-2 "пептитело", и группой, получающей в качестве контроля носитель (р=0,089 для группы, начавшей получать "пептитело" в день 15, и р <0,0001 для групп, начавших лечение в дни 3 и 10, при использовании повторного измерения ANOVA, с post hoc (последующим) тестом Шеффе (Scheffe)). Лечение этими "пептителами" не оказывает значительного влияния на вес тела. Результаты представлены на фиг. 8 (вес тела не показан). Пример 18. Резюме скоростей объективных ответов (CR) получают, используя антитело АЬ536 в количестве 47 мкг/самка "голой" мыши, вводимое интраперитонеально 3 раза в неделю, или "пептитело" 2ХСОП4(С), вводимое подкожно по схемам многократного введения в исследованиях различной длительности (время введения доз > 10 недель), обоим, А431 и Colo-205, моделям ксенотрансплантатов. CR по данному описанию относится к результату, когда после лечения не остаётся определяемой (измеряемой) опухоли. Результаты представлены на фиг. 9. Пример 19. a) Комбинация РЬ ("пептитела") с Таксотером на Colo-205 опухолевой модели. Самкам "голых" мышей подкожно инъецируют 2х106 Colo-205 клеток плюс Матригель (2:1) в день исследования 0. В день исследования 14 начинают лечение с помощью а)подкожного (s.c.) введения дважды в неделю 350 мкг/мышь Ang-2 "пептитела" 2xCon4-C; b) 20 мг/кг Таксотера по схеме qwx3 i.p.; или с) их комбинации. Объёмы опухолей и вес тела записывают через регулярные интервалы. Наблюдают значительную разницу в росте опухоли между всеми группами и группой, получающей в качестве контроля носитель (р <0,0001 при использовании повторного измерения ANOVA, с post hoc (последующим) тестом Шеффе (Scheffe)). Помимо этого, группа с комбинированной терапией значительно отличается от любой из групп с монотерапией (р <0,0001 vs. 2xCon-4-C и р=0,0122 vs. Таксотера). Пунктирной линией показано уменьшение n группы, с 10 до 9 мышей, вследствие смерти одной из мышей по неизвестной причине. Лечение этими "пептителами" не оказывает значительного влияния на вес тела. Результаты представлены на фиг. 10а. b) Комбинация РЬ с 5-FU на Colo-205 опухолевой модели. Самкам "голых" мышей подкожно инъецируют 2х106 Colo-205 клеток плюс Матригель (2:1) в день исследования 0. В день исследования 14 начинают лечение с помощью а)подкожного (s.c.) введения дважды в неделю 350 мкг/мышь Ang-2 "пептитела" 2ХСОП4-С; b) 50 мг/кг 5-FU по схеме qwx5 i.p.; или с) их комбинации. Объёмы опухолей и вес тела записывают через регулярные интервалы. Наблюдают значительную разницу в росте опухоли между всеми группами и группой, получающей в качестве контроля носитель (р <0,0001 при использовании повторного измерения ANOVA, с post hoc (последующим) тестом Шеффе (Scheffe)). Помимо этого, группа с комбинированной терапией значительно отличается от любой из групп с монотерапией (р=0,0375 vs. 2xCon-4-C и р=0,0453 vs. 5-FU). Временное снижение веса тела наблюдают в ^упш 5-FU (18% в день исследования 20), а также в группе с - 76 - 008248 комбинированной терапией (16% в день исследования 20), с последующим полным восстановлением веса тела. Результаты представлены на фиг. 10b. Пример 20. Модель адъювантного артрита. Самцов крыс Льюиса (120-130 г, Charles River, Wilmington MA) помещают по двое в клетку с дверцей-фильтром в комнате с регулируемой окружающей средой (температура 23±2°С, относительная влажность 50±20%) с соблюдением цикла с 12-часовым световым днём. Животным дают корм для грызунов (Рецептура 8640; Тек Lab, Madison, WI) и отфильтрованную водопроводную воду ad libitum (по желанию). Содержание кальция и фосфора в пище составляет 1,2 и 1,0% соответственно. Адъювантный артрит вызывают разовой инъекцией 0,5 мг термоинактивированных Mycobacterium tuberculosis H37Ra (Difco Laboratories, Detroit, MI), суспендированных в 0,05 мл парафинового масла (Crescent Chemical Co., Hauppauge, NY), интрадермально в основание хвоста. Клиническое начало артрита наступает в день 9, на что указывает опухание задней лапы и трудности с передвижением. За исключением группы, получающей 2ХСОП4(С) (которая начинает получать это "пептитело" с 1 дня после иммунизации), препараты вводят в виде ежедневных подкожных инъекций, начиная с дня 9 после иммунизации (до начала артрита), и продолжают давать вплоть до дня 18. Клинический мониторинг адъювантного артрита. Прогрессирование воспаления оценивают клинически, периодически измеряя объём задней лапы методом "водной" плетизмографии, описанным Feige et al., Cellular Molec. Life Sci., 57:1457-1470 (2000). Подавление воспаления лапы рассчитывают, исходя их площади под кривой (AUC) в соответствии с правилом трапеций по формуле: [1-{(Обработан. AdA)-норм.)/(Необработан. AdA-норм.)}] х100 Кроме того, ежедневно в течение 9-дневного введения препарата (по схеме) в качестве дополнительного параметра (концевой точки), так как показано, что потеря веса тела идёт параллельно прогрессиро-ванию воспаления суставов в этой подели артрита. Животных умерщвляют в атмосфере СО2 на 18 день. Снижение минеральной плотности костей (BMD) изучают при аутопсии (18 день после иммунизации). Задние лапы удаляют по линии шерсти (непосредственно рядом с голеностопным (скакательным) суставом), погружают в 70% этанол, а затем сканируют в горизонтальном направлении на веерном рентгеновском денситометре (Model QDR-4500А; Hologic, Waltham, MA). См. выше Feige et al. После сканирования прямоугольный корпус (29х25 мм), центрированный по пяточной кости, позиционируют так, чтобы очертить ("оконтурить") анализируемый участок, и с помощью соответствующих алгоритмов (программное обеспечение Hologic) рассчитывают площадь кости, минеральное содержание костей и минеральную плотность костей. Все результаты выражают как среднее ± стандартная ошибка. Значение р 0,05 используют, чтобы описать значительные различия между группами. Тест Kruskal-Wallis ANOVA и Mann-Whitney U. тест, использующий промышленное статистическое программное обеспечение (Statsoft v3.0; Statsoft, Tulsa, OK) проводят, используя клинические данные (непрерывные переменные). Результаты представлены на фиг. 11 а, 11b и 11 с соответственно. Пример 21. Модель ангиогенеза в роговице. Влияние CON4(C) на VEGF-индуцированный ангиогенез у крыс. Ang-2 "пептитело" CON4(C) оценивают на модели ангиогенеза у крыс. Ангиогенез вызывают, имплантируя намоченный в VEGF- (или в BSA-контрольный) найлоновый диск в строму роговицы (n=8/группа). "Пептитело" TN8CON4-C вводят в виде подкожной инъекции в дозе 1,0 или 0,1 мг крыса/день в течение 7 дней. Двум другим группам животных дают ту же дозу негативного контрольного "пептитела" 4883. Всем группам предварительно дают разовую ударную дозу 3,0 или 0,3 мг, что в 3 раза выше поддерживающей дозы 1,0 или 0,1 мг (см. фигуру). Через 7 дней лечения на каждом цифровом изображении роговицы глаза крыс определяют две васкулярные концевые точки; число сосудов, пересекающих среднюю точку (центр) между диском и лимбом, и площадь кровеносных сосудов, лечение с помощью TN8CON4-C заметно подавляет VEGF-индуцированный ангиогенез в зависимости от дозы (р <0,04), тогда как введение контрольного "пептитела" не оказывает заметного влияния ни на одну концевую точку. Вес тела пролеченных животных не свидетельствует о токсичности. Результаты представлены на фиг. 12. Пример 22. Эпитопное картирование. Полноразмерные (аминокислоты 1-495) N-концевые (аминокислоты 1-254) и С-концевые (аминокислоты 255-495) человеческие Ang-2 (hAng-2) белки клонируют в CMV-контролируемый вектор экспрессии млекопитающих с С-концевыми 6xHis хвостами. Три полученных в результате конструкции плюс элемент контроля вектора транзиторно экспрессируют в клетки 293Т. Затем из трансфецированных клеток собирают кондиционированную среду и оценивают уровень Ang-2 в средах методом анти-6х1^ ELISA и Вестерн-блоттингом. Связывающий эпитоп антител против Ang-2 и "пептител" определяют по их способности связываться с тремя вариантами человеческого hAng-2 методом ELISA в соответствии со следующим протоколом: 96-луночный аналитический планшет с высоким связыванием покрывают 100 мкл кондициониро - 77 - 008248 ванной среде на лунку и инкубируют при 37°С в течение 1 ч. Кондиционированные среды отсасывают и планшет блокируют с помощью 200 мкл/лунка 5% BSA в PBS при комнатной температуре в течение 1 ч. Затем отсасывают блокирующий раствор. Добавляют 100 мкл/лунка антитела, "пептитела" или Tie2-Fc количестве 1 мкг/мл в 1% BSA в PBS И инкубируют при комнатной температуре в течение 1 ч. Лунки 4 раза отмывают в 200 мкл 0,1% Tween в PBS. Добавляют 100 мкл/лунка HRP-коньюгированного антитела козы к человеческому IgG или антитела козы к мышиному IgG и инкубируют в течение 45 мин при комнатной температуре. Затем лунки 4 раза отмывают в 200 мкл 0,1% Tween в PBS. Далее добавляют 100 мкл/лунка субстрат ТМВ. OD. прочитывают при 370 нм. Результаты представлены на фиг. 13а, 13b и 13с. Пример 23. Вследствие некоторых ограничений, связанных с чувствительностью, присущей методу анализа BiaCore, аффинность связывания определяют методом анализа Sepidyne KinExA. Связывание 2xCON4-C (Pb5714) с huAng-2 проверяют на KinExA (Sapidyne, Boise, ID). Бусы Reacti-Gel 6x (Pierce, Rockford, IL) предварительно покрывают huAng-2 и блокируют BSA. Образцы 10 и 30 пМ 2xCON4-C инкубируют с различными концентрациями (0,3 пМ - 3 нМ) huAng-2 при комнатной температуре в течение 8 ч прежде, чем пропустить их через покрытые huAng-2 бусы. Количество связанного с бусами "пептитела" определяют с помощью меченого флуоресцентной (Су5) меткой антитела козы против человеческого Fc антитела (Jackson Immuno Research, West Grove, PA). Сигнал (его интенсивность) связывания пропорционален концентрации свободного "пептитела" в равновесном состоянии. Константу равновесной диссоциации (Кз) определяют нелинейной регрессией конкурентных кривых, применяя односайтовую гомогенную модель связывания по двум кривым (программное обеспечение KinEx(tm)). Затем определяют, что KD составляет, примерно, 2 пМ для связывания 2xCON4-C с huAng-2. Как видно на фиг. 14, методом KinExA показано, что аффинность "пептитела" 2хСоп4 к hAng-2 составляет ~2 пМ. Пример 24. ПЭГированные пептиды. L1-7 пептид синтезируют в 431 ABI синтезаторе по стандартному протоколу связывания и двойного связывания остатка 14 (met) с N-концевым остатком 1 (Cys), нумерация идёт от N-конца к С-концу. Конъюгация L1-7 пептида с метоксиполи(этиленгликоль)малеинимидом; синтез конъюгата с MW: 5 КДа; обозначенного "mPEG5K-(Ll-7 Peptide)". Раствор 0,8 мг L1-7 пептида в 400 мкл буфера 1 (20 мМ фосфата, 5 мМ EDTA, рН 6,5) обрабатывают 13,5 мг метоксиполи(этиленгликоль)малеинимида (MW = 5 КДа; Shearwater Corp.); 0,27 мл раствора с концентрацией 50,0 мг/мл в буфере 1. Реакционную смесь инкубируют при 4°С в течение ночи, затем разводят в 1,6 мл буфера А (20 мМ Tris гидрохлорида, рН 7,2) и подвергают диализу в кассете Slide-A-Lyzer (3500 MWCO, Pierce) против того же буфера. Диализованную реакционную смесь очищают ионообменной хроматографией на колонке 1,0 мл HiTrap Q Sepharose HP (Amersham Biosciences Corp.). Пик продукта элюируют в виде двух фракций по 1,0 мл в градиенте от 100% буфера А до 100% буфера В (буфер А + 0,5М NaCl) в объёме, равном 40 объёмам колонки. Объединённые фракции с продуктом упаривают до 250 мкл с содержанием 0,23 мг белка/мл в центрифуге Microsep 1K Centrifugal Device (Pall Life Sciences). Конъюгация L1-7 пептида с 1.11-бисмалеинимидотетраэтиленгликолем; синтез конъюгата, обозначенного TEO4(L1-7 Peptide)2". Раствор 1,0 мг of L1-7 пептида в 500 мкл буфера 1 (20 мМ фосфата, 5 мМ EDTA, рН 6,5) обрабатывают 0,0375 мг 1,11-бисмалеинимидотетраэтиленгликолем (Pierce) (0,375 мл раствора 0,1 мг/мл в буфере 1). Реакционную смесь инкубируют при 4°С в течение 3,33 ч, затем подвергают диализу в кассете Slide-A-Lyzer (3500 MWCO, Pierce) против буфера А (20 мМ Tris гидрохлорида, рН 7,2). Диализованную реакционную смесь очищают ионообменной хроматографией на колонке 1,0 мл HiTrap Q Sepharose HP (Am-ersham Biosciences Corp.). Пик димерного продукта элюируют в виде трёх фракций по 1,0 мл в градиенте от 100%) буфера А до 100% буфера В (буфер А + 0,5 М NaCl) в объёме, равном 40 объёмам колонки. Объединённые фракции с продуктом упаривают до 550 мкл с содержанием 0,12 мг белка/мл в центрифуге Microsep 1K Centrifugal Device (Pall Life Sciences). Конъюгация L1-7 пептида с поли(этиленгликоль)бисмалеинимидом; синтез конъюгата с MW 3,4 КДа, обозначенного "PEG3.4K(Ll-7 Peptide^". К раствору 3,0 мг L1-7 пептида в 1.5 мл буфера 1 (20 мМ фосфата, 5 мМ EDTA, рН 6,5) прибавляют 1,125 мг поли(этиленгликоль)бисмалеинимида (MW=3,4 КДа, Shearwater Corp.); 0,563 мл раствора 2.0 мг/мл в буфере 1. Реакционную смесь инкубируют при 4°С в течение ночи, затем подвергают диализу в кассете Slide-A-Lyzer (3500 MWCO, Pierce) против буфера А (20 мМ Tris гидрохлорида, рН 7,2). Диали-зованную реакционную смесь очищают ионообменной хроматографией на колонке 5,0 мл HiTrap Q Sepharose HP (Amersham Biosciences Corp.). Пик продукта элюируют в виде трёх фракций по 3,0 мл в градиенте от 100% буфера А до 100% буфера В (буфер А + 0,5М NaCl) в объёме, равном 40 объёмам колонки. Объединённые фракции с продуктом упаривают до 850 мкл с содержанием 0,24 мг белка/мл в двух центрифугах Microsep 1K Centrifugal Device (Pall Life Sciences). Результаты MALDI-TOF масс-спектроскопии представлены ниже. - 78 - 008248 Образец# Идентичность Расч. MS Найд. MS L1-7 (неПЭГированный пептид) 3545 3538.7 mPEG5K-(Ll-7 Peptide) 8500 8851 PE04(Ll-7 Peptide)2 7443 7446.29 PEG3.4K(Ll-7 Peptide)2 10550 10552 6882.61 3550.13 Ясно, что нижний индекс "2" для PEG3.4K(L1-7 Peptide)2 и PEO4(L1-7 Peptide)2 указывает, что на одну пептидную цепь приходятся два пептида, по одному на каждом конце полимера. Определение ID50. IC50 ингибирования hAng2:hTie2-Fc для L1-7 свободного и ПЭГированного пептидов определяют нейтрализационным методом ELISA, описанным в примере 2. Для нейтрализационного метода ELISA микротитрационные планшеты, с которыми связывается человеческий Ang-2 полипептид, получают, как описано в примере 2 для аффинного метода ELISA. Возможные (кандидаты) affra-Ang-2 L1-7 ПЭГиро-ванный и свободный пептиды титруют от 1000 до 0,2 пМ в 4-кратном разведении в растворе PBS, содержащем около 1% BSA и около 1 нМ Tie2 (при условии, что молекула Tie-2-Fc представляет собой молекулу, в которой участок Tie-2 содержит только растворимую внеклеточную область молекулы; R &D Systems, № в каталоге 313-TI). После того, как в каждую лунку добавляют около 100 мкл раствора, планшеты инкубируют в течение ночи при комнатной температуре, а затем пять раз отмывают в PBS, содержащем около 0,1% Tween-20. После отмывания в каждую лунку добавляют около 100 мкл антитела против Tie-2 (Pharmingen Inc., № в каталоге 557039) до примерной конечной концентрации 1 мкг в мл, и планшеты инкубируют в течение 1 ч при комнатной температуре. Далее добавляют около 100 мкл/лунка антитела козы против мышиного IgG-HRP (Pierce Chemical CO., № в каталоге 31432) в разведении 1:10000 в PBS, содержащем около 1% BSA. Планшеты инкубируют при комнатной температуре около 1 ч, после чего их отмывают 5 раз PBS, содержащем 0,1% Tween-20. Затем добавляют около 100 мкл/лунка ТМВ субстрата (описанного выше) и оставляют до появления синего цвета. Затем определяют оптическую плотность на спектрофотометре (планшетном ридере) при 370 нм. L1-7 пептиды (C-GGGGG-AQ-TNFMPMDDLEQRLYEQFILQQG-LE) (SEQ ID NO: 359) включают N-концевой цистеин для связывания с ПЭГ и линкер 5Gly. Фланкирующие последовательности AQ и LE присутствуют как в оригинальном (исходном) клоне фага, так и в "пептителе". Результаты IC50 ингибирования hAng-2: Tie2 приведены ниже. Пептид IC50(HM) L1-7 пептид 0.49 mPEG5K-(Ll-7 Peptide) 11.7 PE04(Ll-7 Peptide)2 0.064 PEG3.4K(Ll-7 Peptide)2 0.058 ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность WDPWT (SEQ ID NO: 65) длиной в 5-50 аминокислот, и его физиологически приемлемые соли. 2. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность WDPWTC (SEQ ID NO: 66) и его физиологически приемлемые соли. 3. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность Cz2WDPWT 2 (SEQ ID NO: 67) где z2 представляет собой кислый или нейтральный полярный аминокислотный остаток, и его физиологически приемлемые соли. 4. Полипептид по п.3, содержащий аминокислотную последовательность Cz2WDPWTC (SEQ ID NO: 68) где z2 обозначает кислый или нейтральный полярный аминокислотный остаток, и его физиологически приемлемые соли. 5. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы - 79 - 008248 a1a2a3Ca5WDPWTCa12a13a14 (SEQ ID NO: 69) где а1, а2 и а3, каждый независимо, обозначают аминокислотные остатки; а5 обозначает аминокислотный остаток; а12 отсутствует или обозначает аминокислотный остаток; а13 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; а14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; и его физиологически приемлемые соли. 6. Полипептид по п.5, отличающийся тем, что а1 обозначает V, I, P, W, G, S, Q, N, Е, K, R или Н; а2 обозначает V, Р, М, G, S, Q, D, Е, K, R или Н; а3 обозначает А, V, Р, М, F, T, G, D, Е, K или Н; а8 обозначает А, V, G, Q, N, D или Е; а12 обозначает S, Q, N, D, Е, K или R; а13 обозначает L, Т или Н; а14 обозначает V, L, I, W или М; и его физиологически приемлемые соли. 7. Полипептид по п.5, отличающийся тем, что а1 обозначает Q; а2 обозначает Е; а3 обозначает Е; а5 обозначает D или Е; а12 обозначает D или Е; а13 обозначает Н и а14 обозначает М, и его физиологически приемлемые соли. 8. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы b1b2b3b4b5b6Cb8WDPWTCb15b16b17b18b19b20 (SEQ ID NO: 70) где b1 отсутствует или обозначает аминокислотный остаток; b2 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; b3, b4, b5 и b6, каждый независимо, отсутствуют или обозначают аминокислотные остатки; b8 обозначает аминокислотный остаток; b15 отсутствует или обозначает аминокислотный остаток; b16 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; b17 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; 18 19 20 b18, b19 и b20, каждый независимо, отсутствуют или обозначают аминокислотные остатки; и его физиологически приемлемые соли. 9. Полипептид по п.8, отличающийся тем, что b1 отсутствует или обозначает А, V, L, P, W, F, T, G, S, Q, N, K, R или Н; b2 отсутствует или обозначает А, V, L, I, P, W, M, T, G, S, Y, N, K, R или Н; b3 отсутствует или обозначает A, L, I, P, W, M, T, G, S, Q, N, E, R или Н; b4 обозначает V, I, P, W, G, S, Q, N, Е, K, R или Н; b5 обозначает V, Р, М, G, S, Q, D, Е, K, R или Н; b6 обозначает А, V, Р, М, F, T, G, D, Е, K или Н; b8 обозначает А, V, G, Q, N, D или Е; b15 обозначает S, Q, N, D, Е, K или R; b16 обозначает L, Т или Н; b17 обозначает V, L, I, W или М; b18 отсутствует или обозначает А, V, L, P, W, F, T, G, Y, Q, D, Е или R; b19 отсутствует или обозначает V, L, I, P, T, G, S, Y, Q, N, D, Е или R и b20 отсутствует или обозначает V, L, P, W, M, T, G, S, Y, Q, N, D, K или R; и его физиологически приемлемые соли. 10. Полипептид по п.8, отличающийся тем, что b1 отсутствует или обозначает Р или Т; b2 отсутствует или обозначает I или N; b3 отсутствует или обозначает R или I; - 80 - 008248 b4 обозначает Q; b5 обозначает Е; b6 обозначает Е; b8 обозначает D или Е; b15 обозначает D или Е; b16 обозначает Н; b17 обозначает М; b18 отсутствует или обозначает W или Р; b19 отсутствует или обозначает G или Е и b20 отсутствует или обозначает V или K; и его физиологически приемлемые соли. 11. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбираемую из группы, приведенной в табл. 1, состоящей из SEQ ID NO: 4 и с SEQ ID NO: 76 по SEQ ID NO: 118 включительно, и его физиологически приемлемые соли. Таблица 1 | ПЕПТИД SEQ ID NO ПЕПТИДНАЯ ПОСЛЕДОВА -ТЕЛЬНОСТЬ Соп4-44 PRQEECDWDPWTCEHMWEV Соп4-40 TNIQEECEWDPWTCDHMPGK Соп4-4 WYEQDACEWDPWTCEHMAEV Соп4-31 NRLQEVCEWDPWTCEHMENV Соп4-С5 AATQEECEWDPWTCEHMPRS Соп4-42 JJ№QEGCEWDPWTCEHMFDW Соп4-35 WRQKDCEWDPWTCEHMYVG Соп4-43 SISHEECEWDPWTCEHMQVG Соп4-49 WAAQEECEWDPWTCEHMGRM Соп4-27 TWPQDKCEWDPWTCEHMGST Соп4-48 GHSQEECGWDPWTCEHMGTS Соп4-46 QHWQ^CEWDPWTCDHMPSK СопФ41 NVRQEKCEWDPWTCEHMPVR Соп4-36 KSGQVECNWDPWTCEHMPRN Соп4-34 WTQmCDWDPWTCEHMREW Соп4-28 AWGQEGCDWDPWTCEHMLPM Соп4-39 PVNQEDCEWDPWTCEHMPPM Соп4-25 RAPQEDCEWDPWTCAHMDIK Соп4-50 HGQNMECEWDPWTOEHMFRY Соп4-38 PPJ^QF^CVWDPWTCEHMPLR Соп4-29 RTTQEKCEWDPWTGEHMES Q Соп4-47 QTSQEDCVWDPWTCDHMVSS Соп4-20 QVIGRPCEWDPWTCEHLEGL Соп4^5 WAQQEECAWDPWTCDFTMVGL Соп4-37 100 IJPGQEDCEWDPWTCEHMVRS Соп4-33 101 PMNQVECDWDPWTCEHMPRS АС2-Соп4 102 FGWSHGCEWDPWTCEHMGST Соп4-32 103 KSTQDDCDWDPWTCEHMVGP Соп4-17 104 GPRISTCOWDPWTCEHMDQL Соп4-8 105 STIGDMCEWDPWTCAHMQVT) АС4-Соп4 106 VLGGQGCEWDPWTCRLLQGW Соп4-1 107 YLGGQGCQWDPWTCSHLEDG Соп4-С1 108 TTIGSMCEWDPWTCAHMQGG Соп4-21 109 TKGKSVCQWDPWTCSHMQSG Соп4-С2 TTTGSMCQWDPWTCAHMQGG Соп4-18 111 WVNEWCEWDPWTCNHWDTP Соп4-19 112 WQVGMCQWDPWTCKHMRLQ Соп4-16 113 AVGSQTCEWDPWTCAHLVEV Соп4-11 114 QGMKMFCEWDPWTCAHIVYR Соп4-С4 115 TTIGSMCQWDPWTCEBMQGG Соп4-23 116 TSQRVGCEWDPWTCQELTYT Соп4-15 117 QWSWPPCEWDPWTCQTVWPS Соп4-9 118 GTSPSFCQWDPWTCSHMVQG TN8-Con4* QEECEWDPWTCEHM 12. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пеп - 81 - 008248 тид по пп.1, 5, 8 или 11 и носитель, и его физиологически приемлемые соли. 13. Слитый полипептид по п.12, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 14. Полипептид по пп.1, 5, 8 или 11, который является циклическим. 15. Димер или мультимер полипептидов по пп.1, 5, 8 или 11. 16. Состав полипептида, имеющий формулу (X1)a-F1-(X2)b, и его мультимеры, где F1 обозначает носитель; X1 и X2, каждый независимо, выбирают из -(LV4LVP2; 4LVP4LVP2-(L3)E-P3;H 1 2 3 4LVP4LVP4LVP3-(LVP4; где один или более из Р , Р , Р и Р , каждый независимо, представляют собой полипептид, выбираемый из группы, состоящей из: (a) аминокислотной последовательности WDPWT (SEQ ID NO: 65), отличающейся тем, что указанный полипептид имеет протяжённость 5-50 аминокислот; (b) аминокислотной последовательности WDPWTC (SEQ ID NO: 66), (c) аминокислотной последовательности Cz2WDPWT (SEQ ID NO: 67) и (d) аминокислотной последовательности Cz2WDPWTC (SEQ ID NO: 68), где z2 обозначает кислый или нейтральный полярный аминокислотный остаток; L1, L2, L3 и L4, каждый независимо, обозначает линкеры и а, b, с, d, e и f, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 17. Состав полипептида по п.16, отличающийся тем, что z2 обозначает А, V, G, Q, N, D или Е. 18. Состав полипептида по п.16, отличающийся тем, что один или более из Р1, Р2, Р3 или Р4, каждый независимо, представляет собой полипептид, выбираемый из группы, состоящей из SEQ ID NO: 4 и с SEQ ID NO: 76 по SEQ ID NO: 118 включительно. 19. Состав полипептида по п.16, соответствующий формулам X1-F1 или F1-X2 и его физиологически приемлемые соли. 20. Состав полипептида по п.16 формулы F1-(L1)c-P1 и его физиологически приемлемые соли. 21. Состав полипептида по п.16, соответствующий формуле F1-(L1)c-P1-(L2)d-P2 и его физиологически приемлемые соли. 22. Состав полипептида по п.16, соответствующий формуле P1-(L1)c-F1-(L2)d-P2 и его физиологически приемлемые соли. 23. Состав полипептида по п.16, отличающийся тем, что F1 обозначает домен Fc или его фрагмент. 24. Состав полипептида по п.16, отличающийся тем, что F1 содержит аминокислотную последовательность SEQ ID NO: 60. 25. Полинуклеотид, кодирующий состав полипептида по пп.1, 5, 8 или 11. 26. Вектор экспрессии, содержащий полинуклеотид по п.25. 27. Клетка-хозяин, содержащая вектор экспрессии по п.26. 28. Клетка-хозяин по п.27, отличающаяся тем, что является клеткой прокариот. 29. Клетка-хозяин по п.28, отличающаяся тем что является клеткой Е. coli. 30. Клетка-хозяин по п.27, отличающаяся тем, что является клеткой эукариот. 31. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы Pc2Dc4Lc6c7c8LY (SEQ ID NO: 71) где с2 обозначает нейтральный гидрофобный аминокислотный остаток; с4 обозначает A, D или Е; - 82 - 008248 с6 обозначает кислый аминокислотный остаток; с7 обозначает аминокислотный остаток и с8 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; и его физиологически приемлемые соли. 32. Полипептид по п.31, отличающийся тем, что с2 обозначает L или М. 33. Полипептид по п.31, отличающийся тем, что с6 обозначает D или Е. 34. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы d1d2d3d4Pd6Dd8Ld10d11d12LYd15d16d17d18d19d20d21d22 1 (SEQ ID NO: 72) где d1 отсутствует или обозначает аминокислотный остаток; d2 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d3 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d4 отсутствует или обозначает аминокислотный остаток; d6 обозначает нейтральный гидрофобный аминокислотный остаток; d8 обозначает A, D или Е; d10 обозначает кислый аминокислотный остаток; d11 обозначает аминокислотный остаток; d12 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; d15 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d16 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d17 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d18 отсутствует или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; d19 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; d20 отсутствует или обозначает аминокислотный остаток; d21 отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток; d22 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; и его физиологически приемлемые соли. 35. Полипептид по п.34, отличающийся тем, что: d1 обозначает Т, S, Q, R или Н; d2 обозначает Т, Q, N или K; d3 обозначает F; d4 обозначает М, Q, Е или K; d6 обозначает L или М; d8 обозначает D или Е; d10 обозначает Е; d11 обозначает Q или Е; d12 обозначает Т или R; d15 обозначает Y, D, Е или K; d16 обозначает Q; d17 обозначает W или F; d18 обозначает L, I, М или Т; d19 обозначает L, F или Y; d20 обозначает Q, D или Е; d21 отсутствует или обозначает Q или Н; d22 отсутствует или обозначает A, L, G, S или R; и его физиологически приемлемые соли. 36. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 2, состоящей из SEQ ID NO: 6 и с SEQ ID NO: 119 по SEQ ID NO: 142 включительно, и его физиологически приемлемые соли. - 83 - 008248 Таблица 2 Пептид SEQ ID NO Последовательность L1-1 119 QNYKPLDELDATLYEHF1FHYT L1-2 120 mFTPLDELEQTLYEQWTLQQS L1-3 121 TKFNPLDELEQTLYEQWTLQHQ L1-4 122 VKFPCPLDALEQTLYEHWMFQQA L1-5 123 VKYKPLDELDEILYEQQTFQER L1-7 124 TNFMPMDDLEQRLYEQFILQQG L1-9 125 SKFTKPLDELEQTLYEQWTLQHA L1-10 126 QKFQPLDELEQTLYEQEMLQQA L1-11 127 QNEKPMDELEDTLYKQFLFQHS L1-12 128 YKT^TIJDDIBQTLYEQWTLQHV Ы-13 129 j QEYEPLDELDETLYNQWMFHQR L1-14 130 ! S№N1PLDELEQTLYEQEN4LQHQ L1-15 131 QKYQPLDELDKTLYDQFMLQQG L1-16 132 QKFQPLDELEETLYKQWTLQQR L1-17 133 VKYKPLDELDEWLYHQFTLHHQ L1-18 134 QKTMPLDELDEILYEQFMFQQS L1-19 135 QTFQPLDDLEEYLYEQWIRRYH L1-20 136 EDYMPIJDAIDAQLYEQFILLHG L1-21 137 ! HTFQPLDELEETLYYOWLYDOL L1-22 138 \TOFNP№ELEQTLYEEFLFQHA AC6-L1 139 imT^JDELDATLYEHWILQHS L1-C1 140 QKFKPLDELEQTLYEQWTLQQR L1-C2 ' 141 TKFQPLDELDQTLYEQWTLQQR L1-C3 142 TNFQPLDELDQTLYEQWTLQQR -KFNPLDELEETLYEQFTFQQ - 37. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пептид по пп.31, 34 или 36 и носитель, и его физиологически приемлемые соли. 38. Слитый полипептид по п.37, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 39. Полипептид по пп.31, 34 или 36, который является циклическим. 40. Димер или мультимер полипептидов по пп.31, 34 или 36. 41. Состав полипептида, имеющий формулу (X1)a-F1-(X2)b и его мультимеры, где F1 обозначает носитель; X1 и X2, каждый независимо, выбирают из -(LVP1; -(LVP'-CLVP2; -(L^-P^CL^d-P'-CL^e-P3; и ^^^^^^^^^^^^^^(L3VP3-(L4)RP4; где один или более из Р1, Р2, Р3 и Р4, каждый независимо, представляют собой полипептид по пп.31, 34 или 36; L1, L2, L3 и L4, каждый независимо, обозначают линкеры и а, b, с, d, e и f, каждый независимо, обозначают 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 42. Состав полипептида по п.41, отличающийся тем, что один или более из Р1, Р2, Р3 и Р4, каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 6 и с SEQ ID NO: 119 по SEQ ID NO: 142 включительно. 43. Состав полипептида по п.41, соответствующий формулам X1-F1 или F1-X2 и его физиологически приемлемые соли. 44. Состав полипептида по п.41 формулы F1-(L1)c-P1 - 84 - 008248 и его физиологически приемлемые соли. 45. Состав полипептида по п.41, соответствующий формуле F1-(L1)c-P1-(L2)d-P2 и его физиологически приемлемые соли. 46. Состав полипептида по п.41, соответствующий формуле P1-(L1)c-F1-(L2)d-P2 и его физиологически приемлемые соли. 47. Состав полипептида по п.41, отличающийся тем, что F1 обозначает домен Fc или его фрагмент. 48. Состав полипептида по п.41, отличающийся тем, что F1 содержит аминокислотную последовательность SEQ ID NO: 60. 49. Полинуклеотид, кодирующий полипептид по любому из пп.31, 34 или 36. 50. Вектор экспрессии, содержащий полинуклеотид по п.49. 51. Клетка-хозяин, содержащая вектор экспрессии по п.50. 52. Клетка-хозяин по п.51, отличающаяся тем, что является клеткой прокариот. 53. Клетка-хозяин по п.52, отличающаяся тем что является клеткой Е. coli. 54. Клетка-хозяин по п.51, отличающаяся тем, что является клеткой эукариот. 55. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы RPe3e4e5e6e7G (SEQ ID NO: 73) где е3 обозначает нейтральный полярный аминокислотный остаток; е4 обозначает кислый аминокислотный остаток; е5 обозначает нейтральный полярный или кислый аминокислотный остаток; е6 обозначает нейтральный гидрофобный аминокислотный остаток; е7 обозначает нейтральный гидрофобный аминокислотный остаток; и его физиологически приемлемые соли. 56. Полипептид по п.55, отличающийся тем, что е3 обозначает Y или С. 57. Полипептид по п.55, отличающийся тем, что е4 обозначает D или Е. 58. Полипептид по п.55, отличающийся тем, что е6 обозначает I или М. 59. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы f1 f2f3f4RPf7f8ff10f11 Gf13f14f15f16f17f18f19f20 (SEQ ID NO: 74) где f1 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f2 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f3 обозначает нейтральный полярный или кислый аминокислотный остаток; f4 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f7 обозначает нейтральный полярный аминокислотный остаток; f8 обозначает кислый аминокислотный остаток; f9 обозначает нейтральный полярный или кислый аминокислотный остаток; f10 обозначает нейтральный гидрофобный аминокислотный остаток; f11 обозначает нейтральный гидрофобный аминокислотный остаток; f13 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; f15 обозначает нейтральный полярный аминокислотный остаток; f16 обозначает нейтральный полярный аминокислотный остаток; f17 обозначает нейтральный полярный или кислый аминокислотный остаток; f18 обозначает нейтральный гидрофобный или основной аминокислотный остаток; f19 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и f20 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; и его физиологически приемлемые соли. 60. Полипептид по п.59, отличающийся тем, что f1 обозначает S, А или G; f2 обозначает G, Q или Р; f3 обозначает Q, G или D; f4 обозначает L, М или Q; f7 обозначает С или Y; f8 обозначает Е или D; f9 обозначает Е, G или D; f10 обозначает I или М; f11 обозначает F или L; f13 обозначает С или W; - 85 - 008248 f обозначает G или Р; f15 обозначает Т или N; f16 обозначает Q, Y или K; f17 обозначает N, D или Q; f18 обозначает L, V, W или R; f19 обозначает A, Q, Y или I и f20 обозначает L, A, G или V; и его физиологически приемлемые соли. 61. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 3, состоящей из SEQ ID NO: 3 и с SEQ ID NO: 143 по SEQ ID NO: 148 включительно, и его физиологически приемлемые соли Таблица 3 Пептид SEQ ID NO Последовательность Conl-1 143 AGGMRPYDGMLGWPNYDVQA Conl-2 144 QTWDDPCMHILGPVTWRRCI Conl-3 145 AJPGQPPYDGMLGWPTYQPJV _СопТ4________ 146 SGQLRPCEEIFGCGTQNLAL Conl-5 147 FGDKRPLECMFGGPIQLCPR Con 1-6 . 148 GQDLRPCEDMFGCGTICDWYG Conl 3 i KRPCEEIFGGCTYQ 62. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один полипептид по пп.55, 59 или 61 и носитель и его физиологически приемлемые соли. 63. Слитый полипептид по п.62, отличающийся тем, что указанный носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и олигосахарид. 64. Полипептид по пп.55, 59 или 61, который является циклическим. 65. Димер или мультимер полипептидов по пп.55, 59 или 61. 66. Состав полипептида, имеющий формулу и его мультимеры, где F1 обозначает носитель; X1 и X2, каждый независимо, выбирают из -(LVP1; -(LVP^LVP2; -(L1)C-P1-(L2)D-P2-(L3)e-P3; и ^^^^^^^^^^^^^-(L3)E-P3-(L4)f-P4; где один или более из Р1, Р2, Р3 и Р4, каждый независимо, представляют собой полипептид по пп.55, 59 или 61; L1, L2, L3 и L4, каждый независимо, обозначают линкеры и а, b, с, d, е и f, каждый независимо, обозначают 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 67. Состав полипептида по п.66, отличающийся тем, что один или более из Р1, Р2, Р3 и Р4, каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 3 и с SEQ ID NO: 143 по SEQ ID NO: 148 включительно. 68. Состав полипептида по п.66, соответствующий формулам X1-F1 или F1-X2 и его физиологически приемлемые соли. 69. Состав полипептида по п.66 формулы F1-(L1)c-P1 и его физиологически приемлемые соли. 70. Состав полипептида по п.66, соответствующий формуле F1-(L1)c-P1-(L2)d-P2 и его физиологически приемлемые соли. 71. Состав полипептида по п.66, соответствующий формуле P1-(L1)c-F1-(L2)d-P2 и его физиологически приемлемые соли. - 86 - 008248 72. Состав полипептида по п.66, отличающийся тем, что F1 обозначает домен Fc или его фрагмент. 73. Состав полипептида по п.66, отличающийся тем, что F1 содержит аминокислотную последовательность SEQ ID NO: 60. 74. Полинуклеотид, кодирующий полипептид по любому из пп.55, 59 или 61. 75. Вектор экспрессии, содержащий полинуклеотид по п.74. 76. Клетка-хозяин, содержащая вектор экспрессии по п.75. 77. Клетка-хозяин по п.76, отличающаяся тем, что является клеткой прокариот. 78. Клетка-хозяин по п.77, отличающаяся тем что является клеткой Е. coli. 79. Клетка-хозяин по п.76, отличающаяся тем, что является клеткой эукариот. 80. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы Cg2Gg4g5DPFTg10GCg13 (SEQ ID NO: 75) где g2 обозначает кислый аминокислотный остаток; g4 обозначает нейтральный гидрофобный аминокислотный остаток; g5 обозначает Е, D или Q; g10 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; g13 обозначает кислый остаток; и его физиологически приемлемые соли. 81. Полипептид по п.80, отличающийся тем, что g2 обозначает Е или D. 82. Полипептид по п.80, отличающийся тем, что g4 обозначает V или М. 83. Полипептид по п.80, отличающийся тем, что g10 обозначает F или Q. 84. Полипептид по п.80, отличающийся тем, что g13 обозначает D или Е. 85. Полипептид, способный связываться с Ang-2, содержащий аминокислотную последовательность формулы h1h2h3h4Ch6Gh8h9DPFTh14GCh17h18h19h20 (SEQ ID NO:158) где h1 отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; h2 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h3 обозначает кислый аминокислотный остаток; h4 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h6 обозначает кислый аминокислотный остаток; h8 обозначает нейтральный гидрофобный аминокислотный остаток; h9 обозначает Е, D или Q; h14 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток; h17 обозначает кислый аминокислотный остаток; h18 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; h19 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и h20 отсутствует или обозначает аминокислотный остаток; и его физиологически приемлемые соли. 86. Полипептид по п.85, отличающийся тем, что h1 отсутствует или обозначает A, L, M, G, K или Н; h2 обозначает L, F или Q; h3 обозначает D или Е; h4 обозначает W или Y; h6 обозначает D или Е; h8 обозначает V или М; h14 обозначает F или Q; h17 обозначает D или Е; h18 обозначает М, Y, N или K; h19 обозначает L или Q и h20 отсутствует или обозначает М, Т, G, S, D, K или R; и его физиологически приемлемые соли. 87. Полипептид, способный связываться с Ang-2, содержащий по меньшей мере одну аминокислотную последовательность, выбранную из группы, приведенной в табл. 4, состоящей из SEQ ID NO: 5 или с SEQ ID NO: 149 по SEQ ID NO: 157 включительно, и его физиологически приемлемые соли. - 87 - 008248 Таблица 4 Пептид SEQ ID NO Последовательность 12-9-1 149 GFEYCDGMEDPFTFGCDKQT 12-9-2 150 KLEYCDGMEDPFTQGCDNQS 12-9-3 151 LQEWCEGVEDPFTFGCEKQR 12-9-4 152 AQDYCEGMEDPFTFGCEMQK 12-9-5 153 LLDYCEGVQDPFTFGCENLD 12-9-6 154 HQEYCEGMEDPFTFGCEYQG 12-9-7 155 JN'ILDYCEGMDDPFTFGCDKQM 12-9-С2 156 LQDYCEGVEDPFTFGCENQR 12-9-С1 157 LQDYCEGVEDPFTFGCEKQR 12-9 EDYCEGVEDPFTFGCDNH 88. Слитый полипептид, способный связываться с Ang-2, содержащий по меньшей мере один пептид по пп.80, 85 или 87 и носитель, его физиологически приемлемые соли. 89. Слитый полипептид по п.88, отличающийся тем, что носитель представляет собой по меньшей мере один из группы: Fc область (домен), полиэтиленгликоль, липид, группа холестерина, углевод и оли-госахарид. 90. Полипептид по пп.80, 85 или 87, который является циклическим. 91. Димер или мультимер полипептидов по пп.80, 85 или 87. 92. Состав полипептида, имеющий формулу (X1)a-F1-(X2)b и его мультимеры, где F1 обозначает носитель; каждый независимо, выбирают из -(L')c-P'; -(LVP'-CLVP2; -(L'VP'-CLVP'-CL^e-P3; и 4(L')C-P4LVP4L3)C-P3-(LVP4; где один или более из Р1, Р2, Р3 и Р4, каждый независимо, представляют собой полипептид по пп.80, 85 или 87; L1, L2, L3 и L4, каждый независимо, обозначает линкеры и а, b, с, d, e и f, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из а и b обозначает 1; и его физиологически приемлемые соли. 93. Состав полипептида по п.92, отличающийся тем, что один или более из Р1, Р2, Р3 и Р4, каждый независимо, представляет собой полипептид, выбранный из группы, состоящей из SEQ ID NO: 5 и с SEQ ID NO: 149 по SEQ ID NO: 157 включительно. 94. Состав полипептида по п.92, соответствующий формулам X1-F1 или F1-X2 и его физиологически приемлемые соли. 95. Состав полипептида по п.92 формулы F1-(L1)c-P1 и его физиологически приемлемые соли. 96. Состав полипептида по п.92, соответствующий формуле F1-(L1)c-P1-(L2)d-P2 и его физиологически приемлемые соли. 97. Состав полипептида по п.92, соответствующий формуле P1-(L1)c-F1-(L2)d-P2 и его физиологически приемлемые соли. 98. Состав полипептида по п.92, отличающийся тем, что F1 обозначает домен Fc или его фрагмент. 99. Состав полипептида по п.92, отличающийся тем, что F1 содержит аминокислотную последовательность SEQ ID NO: 60. 100. Полинуклеотид, кодирующий полипептид по любому из пп.80, 85 или 87. 101. Вектор экспрессии, содержащий полинуклеотид по п.100. 102. Клетка-хозяин, содержащая вектор экспрессии по п.101. 103. Клетка-хозяин по п.102, отличающаяся тем, что является клеткой прокариот. 104. Клетка-хозяин по п.103, отличающаяся тем что является клеткой Е. coli. - 88 - 008248 105. Клетка-хозяин по п.102, отличающаяся тем, что является клеткой эукариот. 106. Полипептид в соответствии с любой из последовательностей SEQ ID NO: l, SEQ ID NO: 2, SEQ ID NO: 3 или SEQ ID NO: 4 или его вариант. 107. Полипептид в соответствии с любой из последовательностей SEQ ID NO: 5, SEQ ID NO: 6 или SEQ ID NO: 7 или его вариант. 108. Состав полипептида формулы (X1)q-F1-(X2)r и его мультимеры, где F1 обозначает носитель; X1 и X2, каждый независимо, выбирают из -ОЛ-Н1; -ОЛ-Р'-ОЛ-Р2; -(L^S-P^CLV'-CL^U-P3; И 1 2 3 4 4L')S4> 4LVP24L3)U-P34L4)V-P4; где один или более из Р , Р , Р и Р , каждый независимо, представляют собой полипептид, выбранный из группы, состоящей из: (a) аминокислотной последовательности WDPWT (SEQ ID NO: 65), причём указанный полипептид имеет в длину 5-50 аминокислот; (b) аминокислотной последовательности WDPWTC (SEQ ID NO: 66); (c) аминокислотной последовательности Cz2WDPWT (SEQ ID NO: 67), где z2 обозначает кислый или нейтральный полярный аминокислотный остаток; (d) аминокислотной последовательности Cz2WDPWTC (SEQ ID NO: 68), где z2 обозначает кислый или нейтральный полярный аминокислотный остаток; (e) аминокислотной последовательности Рс2Dc4Lc6с7с8LY (SEQ ID NO: 71), где с2 обозначает нейтральный гидрофобный аминокислотный остаток; с4 обозначает A, D или Е; с6 обозначает кислый аминокислотный остаток; с7 обозначает аминокислотный остаток и с8 обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток; (f) аминокислотной последовательности RPe3e4e5e6e7G (SEQ ID NO: 73), где е3 обозначает нейтральный полярный аминокислотный остаток; е4 обозначает кислый аминокислотный остаток; е5 обозначает нейтральный полярный или кислый аминокислотный остаток; е6 обозначает нейтральный гидрофобный аминокислотный остаток и е7 обозначает нейтральный гидрофобный аминокислотный остаток; (g) аминокислотной последовательности Cg2Cg4g5DPFTg10GCg13 (SEQ ID NO: 75), где g2 обозначает кислый аминокислотный остаток; g4 обозначает нейтральный гидрофобный аминокислотный остаток; g5 обозначает нейтральный полярный или кислый аминокислотный остаток; g10 обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток и g13 обозначает кислый остаток; (h) полипептида SEQ ID NO: 1; (i) полипептида SEQ ID NO: 2 и (j) полипептида SEQ ID NO: 7; L1, L2, L3 и L4, каждый независимо, обозначает линкеры и q, r, s, t, u и v, каждый независимо, обозначает 0 или 1, при условии, что по меньшей мере один из q и r обозначает 1; и его физиологически приемлемые соли. 109. Фармацевтическая композиция, содержащая эффективное количество состава полипептида по пп.1, 31, 55 или 80 в смеси с фармацевтически приемлемым носителем. 110. Способ ингибирования нежелательного ангиогенеза у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 111. Способ лечения ангиогенеза у субъекта, заключающийся во введении эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 112. Способ модулирования ангиогенеза у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 113. Способ подавления роста опухоли, характеризующейся нежелательным ангиогенезом, у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 114. Способ лечения рака у млекопитающих, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92 и химиотерапевтического агента. 115. Способ по п.114, отличающийся тем, что химиотерапевтический агент является по меньшей мере одним из 5-FU, СРТ-11 и Таксотер. 116. Способ модулирования по меньшей мере одного нарушения у млекопитающего: сосудистой - 89 - 008248 проницаемости или истечения плазмы, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 117. Способ лечения по меньшей мере одного из заболеваний: реваскулярного заболевания глаза, ожирения, ангиоретикулёмы, гемангиомы, артериосклероза, воспалительного заболевания, воспалительных нарушений, атеросклероза, эндометриоза, опухолевого заболевания, костного заболевания или псориаза, у млекопитающего, заключающийся во введении терапевтически эффективного количества состава полипептида по пп.12, 16, 37, 41, 62, 66, 88 или 92. 700 о -I--ч-ь--1-н-1---I-:-I-1 0 2 4 6 8 10 12 14 16 Дни -"¦-1: Носитель контроль -¦-2: TN8-Con4 (С) (1 мг ) -йг- 3: TN8-Con4 (С) (200 иг) -в- 4: TN8-Con4 (С) (40 иг) -e-5-.TN8-Con4(C) (8 иг)________ Фиг. 1 t1/2 5г 1 01 час AUCinf = 1545 иг* ч/мл Биосовместимость (s.c.) = 96% *1/2 = 49 часов AUCinf = 679 иг* ч/мл Биосовместимость (s.c.) = 95% ^1/2 = 1 05 часов ;AUCinf = 1185 иг" ч/мл Биосовместимость (s.c.) = 124% Фиг. 2 РК "пептитела"у мышей (доза 50 иг) - 90 - 008248 1000 900 f 800 700 V 600 -I 500 " I 400 ° 300 200 100 + 0 -GR 1: Носитель (PBS) -GR2: контрольРЬ (200 ur, qd) -GR 6: 2x-TN8-Cori4C (200 ur, qd) p < 0.0001 -f-4-I-I-\-I-I-I. I II-I-I-I-I-I-f-ь 0 10 Начало введения доз Дни Фиг. 3 "Пептитела"против Ang2 ингибируют рост ксенотрансплантата опухоли А431 2 1.8 1.6 о ю Q О 1.4 1.2 о.8 ^ 0.6 0.4 0.2 0 - Контроль РЬ -Anti-Ang2 РЬ (Соп4-С) - Необработанные (клетки) Дни Фиг. 4 " Пептитело " против Ang2 в подпороговой дозе (Has) 1 мг/мл не оказывает влияния на рост культивированных клеток А431 900 800 700 -i ^600 s 1500 |400 ] 1 300 о 200 100 0 Начало введения доз день 3 | [ I I I | I I l I I I I I I I I I [ I I I I I I I II I I I И | I I I I | II I I | 5 ' 10 15 20 25 30 35 40 45 '50 55 Дни -"- Носитель(РВЗ) -Ф- FC (14 иг, 2х/нед) -•-L1-7-N(14 иг,2х/нед) -0-L1-21-N (14 ' иг,2х/нед) -^-С0П4-С(14 иг,2х/нед) -^г-2хС0П4-С (14 иг,2х/нед) -о- Anti-Ang2 АЬ536 (47 иг, Зх/неД) Фиг. 5 " Пептитела" против Ang2 ингибируют рост опухоли Colo205 - 91 - 008248 2500 2000 & 1500 1000 р = 0.003 р < 0.0001 0 | 5 10 Начало введения доз 20 25 30 Дни после имплантации Фиг. 6 "Пептитела" против Ang2 ингибируют рост опухоли Colo205 Рост опухоли CD31 Плотность Anti-Ang2 РЬ Начало лечения -О Контрольное "пептитело" 350 иг/день (По размеру) -О- Контрольное "пептитело" 350 иг/день (По времени) -#-Antl-Ang2 РЬ (2хСоп4С) 350ш7день Фиг. 7 Лечение "пептителом "против Ang2 снижает плотность CD31 окрашивания трансплантатов опухоли Colo205 1000 т 900 800 + 700 600 -500 400 300 200 100 + 0 -PBS - Control РЬ (350 иг/доза, 2х/нед): Приём доз с дня 3 -2хС0П4С (350 иг/доза, 2х/нед): Прием доз с дня 3 -Control РЬ (350 иг/доза, 2х/нед): Прием доз с дня 10 -2хСоп4С (350 иг/доза, 2х/нед): Приём доз с дня 10 - Control РЬ (350 иг/доза, 2х/нед): Приём доз с дня 15 -2ХСОП4С (350 иг/доза, 2х/нед ): Прием доз с дня 15 р = 0.009 р < 0.0001 р < 0.0001 Дни Фиг. 8 "Пептитела" против Ang2 ингибируют рост опухоли Colo205 вне зависимости от времени начала введения доз - 92 - 008248 Опухоль Исследование Модель Терапия 082801 А431 АЬ 536 100201 А431 АЬ 536 100201 А431 2хСоп4-С 100201 А431 2хСоп4-С 100201 А431 2хСоп4-С 012902 Colo205 АЬ 536 012902 COIO205 2хСоп4-С 031802 Colo205 2хСоп4-С 111901 Colo205 АЬ 536 042602 Colo205 2хСоп4-С Введение доз Длительность Схема приёма * (недели) %CR 46.7 иг , ЗХ/ нед flO 60** 46.7 иг , ЗХ/ нед 40** 200 иг/день 30** 40 иг/день 8 иг/день 46.7 иг, ЗХ/ "ед 140 иг/нед 350 иг , 2Х/ нед 46.7 иг, Зх/нед 14 иг , 2Х/ нед *Во всех исследованиях введение доз начинают в день 3 **Препарат отменяют после достижения CR (полной ремиссии) и повторного роста опухоли не наблюдают. Средняя продолжительность наблюдения 15.3 недель (в интервале 6-27 недель). Фиг. 9 Скорость объективного ответа при длительных исследованиях введения доз 1600 -1 Р <0.0001 Все vs носитель Дни после имплантации -М- GR 1 :Носитель(РВ8): 0.1мл, 2х/нед,8С. GR 2: 2хСоп4-С: 350 иг/мышь, 2х/неД, sc -A- GR 3: Taxotere (20 мг/кг i, IP, Q7Dx3) + PBS (0.1 мл, 2х/нед,ЗС) -О- GR 6: Taxotere (20, мг/кг, IP, Q7Dx3) + 2х Соп4-С ( 350 иг/мышь, 2х/нед , SC) Фиг. 10а Комбинация Pb с Таксотером на модели опухоли Colo205 . р <0.0001 Все vs носитель р=0.0375 2xCon4vs Combo р=0.0453 5-FU vs Combo 30 40 50 : 60 Дни после имплантации GR 1:носитель(РВЗ): 0.1; мл, 2х/неД)ЗС GR 2: 2хСоп4-С: 350 иг/мышь, 2х/нед, sc -акт- GR 4: 5-FU (50 мг/кг, IP, QDx5) + PBS (0.1 мл, 2х/нед,ЗС) -О- GR 7: 5-FU (50 мг/кг, IP, QDx5) + 2х Соп4-С (350 иг/мышь, 2х/нед ) Фиг. 10b Комбинация РЬ с 5-FU на модели опухоли Colo205 - 93 - 008248 1 мг/крыса/день ,' 1 мг/крыса/день "Пептительный" ?х-СОП4С (d1-контроль 18) Фиг. 11а "Пептитела" против Ang2 ингибируют опухание лапы у модели вызванного адъювантом артрита 0.24 0.22 S 0.20 *р <0.05 0.18 0.16 гтп Нормальный Артритный PBS контроль 1 мг/крыса/день °| мг/крыса/день контроль контроль "Пептительный" 2Х-СОП4С контроль (d1-18) Фиг. 11b "Пептитела" против Ang2 ингибируют снижение минеральной плотности костей у модели вызванного адъювантом артрита -¦-: Нормальный контроль ¦Нк- Артритный контроль -¦- PBS контроль -•-1 мг/крыса/день "Пептительный" -О- 1 мг/крыса/день 2x-Con4C (d1 -18) контроль Фиг. 11 с Влияние "пептител" против Ang2 на потерю веса тела у модели вызванного адъювантом артрита - 94 - 008248 и S BSA VEGF VEGF VEGF + + + PBS VEGF C0N4-C C0N4-C C0N4-C 1 мг/крыса 0.1мг/крыса 0.01 мг/крыса BSA VEGF . VEGF VEGF VEGF C0N4-C C0N4-C C0N4-C 1 мг/крыса 0.1 мг/крыса 0.01 мг/крыса Фиг. 12 "Пептитело" CON4-C ингибирует VEGF-индуцированный нормальный ангиогенез Связывание с полноразмерным hAng2 JZU_ Ц} и ол нораз м ерный * pCDNA3 L Фиг. 13 а Эпитопное картирование Связывание с N-конц. hAng2 (аа 1-254) q N-копц. ¦ pCDNA3 r5> Фиг. 13b Эпитопное картирование 2.5 1-5 § 1 0.5 0 Связывание с С-конц. hAng2 (аа 255-495) .ЯГ Q С" КОНЦ.: ¦ pCDNA3 пи . ¦ у 1 ш .1 Hi , Фиг. 13 с Эпитопное картирование - 95 - 008248 120 о -- ¦4- & 10 рМ "пептитело" 8 -20 6.7146-014 1 е-010 1.6е-008 [hAng21 М Фиг. 14 Перечень последовательностей <110> Амген Инк. <120> Агенты, специфически связывающиеся с ангиопоэтином-2 <130> А-801В <140> Ещё не присвоено <141> 2002-10-11 <150> US 60/414,155 <151> 2002-09-27 <150> US 60/328,624 <151> 2001-10-11 <160> 359 <170> Версия Patent In 3.1 <210> 1 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Пептиды, связывающиеся с Ang-2 ; <400> 1 Lys Arq Pro cys Glu Glu Met тгр Gly Gly Cys Asn Tyr Asp 1 5 10 <210> 2 <211> 14 <212> Белок - 96 - 008248 <213> Искусственная последовательность <220> <223> Пептиды, связьшающиеся с Ang-2 <400> 2 His Gin Пе Cys Lys Trp Asp Pro Trp Thr cys Lys His Trp X 5 ±u <210> 3 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, связывающийся с Ang-2 <400> 3 Lys Arg Pro cys Glu Glu Пе Phe Gly Gly Cys Thr Tyr Gin 1 5 10 <210> 4 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, связьюающийся с Ang-2 <400> 4 Gin Glu Glu cys Glu Trp Asp Pro Trp Thr cys Glu His Met 1 5 1° <210> 5 <211> 18 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, связывающийся с Ang-2 - 97 - 008248 <400> 5 Phe Asp туг Cys Glu Gly val Glu Asp Pro Phe Thr Phe Gly cys Asp 15 10 15 Asn His <210> 6 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, связьшающийся с Ang-2 <400> 6 Lys Phe Asn Pro Leu Asp Glu Leu Glu Glu Thr Leu Tyr Glu Gin Phe 1 5 10 15 Thr Phe Gin Gin 20 <210> 7 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Пептиды, связывающиеся с Ang-2 <400> 7 Gin Tyr Gly Cys Asp Gly Phe Leu Tyr Gly cys Met lie Asn 15 10 <210> 8 <211> 67 <212> ДНК <213> Искусственная последовательность <220> <223> Олигонуклеотид - 98 - 008248 - 99 - 008248 <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> (32)..(32) <223> Хаа = Fc <400> 12 Met Gly Ala Gin Lys Phe Asn Pro Leu Asp Glu Leu Glu Glu Thr Leu 15 10 15 Tyr Glu Gin Phe Thr Phe Gin Gin Leu Glu Gly Gly Gly Gly Gly Xaa 20 25 30 <210> 13 <211> 29 <212> Белок <213> Искусственная последовательность "Пептитело", способное связываться с Ang-2 смеш_ признаки (29).. (29) Хаа = Fc <400> 13 Met Lys Phe Asn Pro Leu Asp Glu Leu Glu Glu Thr Leu Tyr Glu Gin 15 10 15 Phe Thr Phe Gin Gin Leu Glu Gly Gly Gly Gly Gly Xaa 20 25 <210> 14 <211> 51 <212> Белок - 100 - 008248 - 101 - 008248 - 102 - 008248 <221> смеш_ признаки <222> (26)..(26) <223> Хаа = Fc <400> 17 Met Gly Ala Gin Gin Glu Glu Cys Glu Trp Asp Pro Trp Thr cys Glu 15 10 15 His Met Leu Glu Gly Gly Gly Gly Gly Xaa 20 25 <210> 18 <211> 45 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> (45)..(45) <223> Хаа = Fc <400> 18 Met Gin Glu Glu Cys Glu Trp Asp Pro Trp Thr cys Glu His Met Gly 1 5 10 15 Ser Gly Ser Ala Thr Gly Gly Ser Gly Ser Thr Ala Ser Ser Gly Ser 20 25 30 Gly Ser Ala Thr His Leu Glu Gly Gly Gly Gly Gly Xaa 35 40 45 <210> 19 <211> 62 <212> Белок <213> Искусственная последовательность <220> - 103 - 008248 <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> (62)..(62) <223> Хаа = Fc <400> 19. Met Gly Ala Gin Gin Glu Glu cys Glu Trp Asp Pro Trp Thr cys Glu 1---- • " 5 ¦ 10 - - 15 His Met Gly Ser Gly Ser Ala Thr Gly Gly Ser Gly Ser Thr Ala ser 20 25 30 Ser Gly Ser Gly ser Ala Thr His Gin Glu Glu cys Glu Trp Asp Pro 35 40 45 Trp Thr Cys Glu His Met Leu Glu Gly Gly Gly Gly Gly xaa 50 55 60 <210> 20 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> (2)..(2) <223> Хаа = Fc <400> 20 Met Хаа Gly Gly Gly Gly Gly Ala Gin Lys Phe Asn Pro Leu Asp Glu 1 5 10 15 Leu Glu Glu Thr Leu туг Glu Gin Phe Thr Phe Gin Gin Leu Glu 20 25 30 <210> 21 <211> 53 - 104 - 008248 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> С2)..(2) <223> хаа = Fc <400> 21 Met хаа Gly Gly Gly Gly Gly Ala Gin Gly ser Gly Ser Ala Thr Gly 15 10 15 Gly ser Gly ser Thr Ala Ser Ser Gly Ser Gly ser Ala Thr His Lys 20 25 30 Phe Asn Pro Leu Asp Glu Leu Glu Glu Thr Leu Tyr Glu Gin Phe Thr 35 40 45 Phe Gin Gin Leu Glu 50 <210> 22 <211> 59 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> (2)..С2) <223> Хаа = Fc <400> 22 Met Хаа Gly Gly Gly Gly Gly Ala Gin Lys Phe Asn Pro Leu Asp Glu 15 10 15 - 105 - 008248 - 106 - 008248 <400> 24 Met Хаа Gly Gly Gly Gly Gly Ala Gin Gly Ser Gly Ser Ala Thr Gly 1 .5 10 15 Gly ser Gly Ser Thr Ala Ser Ser Gly Ser Gly ser Ala Thr His Gin 20 25 30 Glu Glu Cys ..Glu Trp Asp Pro Trp Thr Cys Glu His Met Leu Glu 35 40 45 <210> 25 <211> 61 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> (2)..(2) <223> Хаа = Fc <400> 25 Met Хаа Gly Gly Gly Gly Gly Ala Gin Gin Glu Glu Cys Glu Trp Asp 15 10 15 Pro Trp Thr Cys Glu His Met Gly Ser Gly Ser Ala Thr Gly Gly Ser 20 25 30 Gly ser Thr Ala Ser Ser Gly Ser Gly Ser Ala Thr His Gin Glu Glu 35 40 45 cys Glu Trp Asp Pro Trp Thr cys Glu His Met Leu Glu 50 55 60 <210> 26 <211> 75 <212> Белок <213> Искусственная последовательность - 107 - 008248 - 108 - 008248 Phe Asn Pro Leu Asp Glu Leu Glu Glu Thr Leu Tyr Glu Gin Phe Thr 35 40 45 Phe Gin Gin Gly Gly Gly Gly Gly Gin Glu Glu Cys Glu Trp Asp Pro 50 55 60 Trp Thr Cys Glu His Met Leu Glu 65 70 <210> 28 <211> 30 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> (30)..(30) <223> Хаа = Fc <400> 28 Met Gly Ala Gin Phe Asp туг cys Glu Gly Val Glu Asp Pro Phe Thr 1 5 10 15 Phe Gly cys Asp Asn His Leu Glu Gly Gly Gly Gly Gly Xaa 20 25 30 <210> 29 <211> 26 <212> Белок <213> Искусственная последовательность "Пептитело", способное связываться с Ang-2 смеш_ признаки (26).. (26) - 109 - 008248 <223> Хаа = Fc <400> 29 Met Gly Ala Gin Gin Tyr Gly Cys Asp Gly Phe Leu Tyr Gly cys Met 15 10 15 lie Asn Leu Glu Gly Gly Gly Gly Gly Xaa 20 25 <210> 30 <211> 26 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <2 21> смеш_ признаки <222> (26)..(26) <223> Хаа = Fc <400> 30 Met Gly Ala Gin Lys Arg Pro Cys Glu Glu Met Trp Gly Gly cys Asn 1 5 10 15 Tyr Asp Leu Glu Gly Gly Gly Gly Gly xaa 20 25 <210> 31 <211> 26 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитело", способное связываться с Ang-2 <220> <221> смеш_ признаки <222> (26)..(26) - 110 - 008248 <223> Хаа = Fc <400> 31 Met Gly Ala Gin His Gin He Cys Lys Trp Asp Pro Trp Thr Cys Lys 1 5 10 15 His Trp Leu Glu Gly Gly Gly Gly Gly Xaa 20 25 <210> 32 <211> 26 <212> Белок <213> Искусственная последовательность <400> 32 Met Glv Ala Gin Lvs Ara Pro Cvs Glu Glu lie Phe Gly Gly Cys Thr 1 5' " ' 10 15 Tyr Gin Leu Glu Gly Gly Gly Gly Gly Хаа 20 25 <210> 33 <211> 784 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 33 atgggtgcac agaaattcaa cccgctggac gaactggaag aaactctgta cgaacagttc actttccagc agctcgaggg tggaggcggt ggggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca gttttcctct tccccccaaa acccaaggac - 111 - 008248 accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa 240 gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca 300 a'agccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg 360 caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca 420 gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac 480 accctgcccc catcccggga tgagctgacc aagaaccagg tcagcctgac ctgcctggtc 540 aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 600 aactacaaga ccacgcctcc cgrgcrggac Txcgacggctr ccttcttxct- etacageaag- - - 660 ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat 720 gaggctctgc acaaccacta cacgcagaag agcctctccc tgtctccggg taaataatgg 780 atcc 784 <210> 34 <211> 768 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 acgaacagtt cactttccag 60 gtccaccttg cccagcacct 120 aacccaagga caccctcatg 180 tgagccacga agaccctgag 240 atgccaagac aaagccgcgg 300 tcaccgtcct gcaccaggac 360 aagccctccc agcccccatc 420 cacaggtgta caccctgccc 480 cctgcctggt caaaggcttc 540 agccggagaa caactacaag 600 tctacagcaa gctcaccgtg 660 ccgtgatgca tgaggctctg 720 gtaaataa 768 <210> 35 <400> 34 atgaaattca acccgctgga cagctcgagg gtggaggcgg gaactcctgg ggggaccgtc atctcccgga cccctgaggt gtcaagttca actggtacgt gaggagcagt acaacagcac tggctgaatg gcaaggagta gagaaaacca tctccaaagc ccatcccggg atgagctgac tatcccagcg acatcgccgt accacgcctc ccgtgctgga gacaagagca ggtggcagca cacaaccact acacgcagaa cgaactggaa gaaactctgt tggggacaaa actcacacat agttttcctc ttccccccaa cacatgcgtg gtggtggacg ggacggcgtg gaggtgcata gtaccgtgtg gtcagcgtcc caagtgcaag gtctccaaca caaagggcag ccccgagaac caagaaccag gtcagcctga ggagtgggag agcaatgggc ctccgacggc tccttcttcc ggggaacgtc ttctcatgct gagcctctcc ctgtctccgg - 112 - 008248 <211> 834 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 35 atgaaattca acccgctgga cgaactggaa gaaactctgt acgaacagtt cactttccag 60 cagggatccg grrcrgctac Tzggtggttcc-ggctxcaccg- caagctctgg -ttcaggea-gt - - -120 gcgactcatc tcgagggtgg aggcggtggg gacaaaactc acacatgtcc accttgccca 180 gcacctgaac tcctgggggg accgtcagtt ttcctcttcc ccccaaaacc caaggacacc 240 ctcatgatct cccggacccc tgaggtcaca tgcgtggtgg tggacgtgag ccacgaagac 300 cctgaggtca agttcaactg gtacgtggac ggcgtggagg tgcataatgc caagacaaag 360 ccgcgggagg agcagtacaa cagcacgtac cgtgtggtca gcgtcctcac cgtcctgcac 420 caggactggc tgaatggcaa ggagtacaag tgcaaggtct ccaacaaagc cctcccagcc 480 cccatcgaga aaaccatctc caaagccaaa gggcagcccc gagaaccaca ggtgtacacc 540 ctgcccccat cccgggatga gctgaccaag aaccaggtca gcctgacctg cctggtcaaa 600 ggcttctatc ccagcgacat cgccgtggag tgggagagca atgggcagcc ggagaacaac 660 tacaagacca cgcctcccgt gctggactcc gacggctcct tcttcctcta cagcaagctc 720 accgtggaca agagcaggtg gcagcagggg aacgtcttct catgctccgt gatgcatgag ' 780 gctctgcaca accactacac gcagaagagc ctctccctgt ctccgggtaa ataa 834 <210> 36 <211> 861 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 36 atgggtgcac agaaattcaa cccgctggac gaactggaag aaactctgta cgaacagttc 60 actttccagc agggtggtgg tggtggtggc ggtggtaagt tcaacccact ggatgagctg 120 gaagagactc tgtatgaaca gttcactttc cagcaactcg agggtggagg cggtggggac 180 aaaactcaca catgtccacc ttgcccagca cctgaactcc tggggggacc gtcagttttc 240 ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 300 gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 360 - 113 - 008248 gtggaggtgc gtggtcagcg aaggtctcca cagccccgag caggtcagcc gagagcaatg ggctccttct gtcttcteat tccctgtctc ataatgccaa tcctcaccgt acaaagccct aaccacaggt tgacctgcct ggcagccgga tcctctacag gctccg-tgat-cgggtaaata gacaaagccg cctgcaccag cccagccccc gtacaccctg ggtcaaaggc gaacaactac caagctcacc gcatgaggetcgggaggagc gactggctga atcgagaaaa cccccatccc ttctatccca aagaccacgc gtggacaaga ot-gea-eaaee agtacaacag atggcaagga ccatctccaa gggatgagct gcgacatcgc ctcccgtgct gcaggtggca actaeaegea- <210> 37 <211> 849 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <4О0> 37 atgaaattca acccgctgga cgaactggaa gaaactctgt acgaacagtt cactttccag cagggtggtg gtggtggcgg tggtaagttc aacccactgg atgagctgga agagactctg tatgaacagt tcactttcca gcaactcgag ggtggaggcg gtggggacaa aactcacaca tgtccacctt gcccagcacc tgaactcctg gggggaccgt cagttttcct cttcccccca aaacccaagg acaccctcat gatctcccgg acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac tacacgcaga agagcctctc cctgtctccg ggtaaataa <210> 38 - 114 - 008248 <211> 759 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 38 atgggtgcac agcaggaaga atgcgaatgg gacccatgga cttgcgaaca catgctcgag ggtggaggcg " gtggggacaar aaxtcacara ^gtxraxctt^gcccagcacc- tgaactcctg - - 120 gggggaccgt cagttttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 180 acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 240 aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 300 tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaecagga ctggctgaat 360 ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 420 atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 480 gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 540 gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 600 cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 660 aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 720 tacacgcaga agagcctctc cctgtctccg ggtaaataa 759 <210> 39 <211> 816 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 39 atgcaggaag aatgcgaatg ggacccatgg acttgcgaac acatgggatc cggttctgct 60 actggtggtt ccggctccac cgcaagctct ggttcaggca gtgcgactca tctcgagggt 120 ggaggcggtg gggacaaaac tcacacatgt ccaccttgcc cagcacctga actcctgggg 180 ggaccgtcag ttttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 240 cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 300 tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 360 aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 420 - 115 - 008248 aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 480 tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggat 540 gagctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 600 atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 660 gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 720 tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 780 acgcagaaga gcctctccct gtctccgggt aaataa 816 <210> 40 <211> 867 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 40 atgggtgcac agcaggaaga atgcgaatgg gacccatgga cttgcgaaca catgggatcc 60 ggttctgcta ctggtggttc cggctccacc gcaagctctg gttcaggcag tgcgactcat 120 caggaagaat gcgaatggga cccatggact tgcgaacaca tgctcgaggg tggaggcggt 180 ggggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca 240 gttttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 300 acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 360 gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 420 taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 480 aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 540 aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 600 aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 660 gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 720 tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 780 gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 840 agcctctccc tgtctccggg taaataa 867 <210> 41 <211> 774 <212> ДНК - 116 - 008248 <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 41 atggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca 60 gttttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 120 acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 180 gacggcgtgg aggtgcataa tgcraagara aagcrgcrjgg aggagcagta caacagcacg~ 240- taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 300 aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 360 aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 420 aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 480 gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 540 tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 600 gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 660 agcctctccc tgtctccggg taaaggtgga ggtggtggtg cacagaaatt caacccgctg 720 gacgagctgg aagagactct gtacgaacag tttacttttc aacagctcga gtaa 774 <210> 42 <211> 840 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 42 atggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca 60 gttttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 120 acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 180 gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 240 taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 300 aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 360 aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 420 aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 480 gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 540 - 117 - 008248 tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag agcctctccc tgtctccggg taaaggtgga ggtggtggtg cacagggatc cggttctgct actggtggtt ccggctccac cgcaagctct ggttcaggca gtgcgactca taaattcaac ccgctggacg aactggaaga aactctgtac gaacagttca ctttccagca actcgagtaa <210> 43 <211> 858 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 43 atggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca gttttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag agcctctccc tgtctccggg taaaggtgga ggtggtggtg cacagaaatt caacccgctg gacgaactgg aagaaactct gtacgaacag ttcactttcc agcagggtgg tggtggtggt ggcggtggta agttcaaccc actggatgag ctggaagaga ctctgtatga acagttcact ttccagcaac tcgagtaa <210> 44 <211> 756 <212> ДНК <213> Искусственная последовательность - 118 - 008248 <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 44 atggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca 60 gttttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 120 acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 180 gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 240 taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 300 aagtgcaagg tctccaacaa agccctccca gcccccatcg" agaaaaccat ctccaaagcc" " 360 aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 420 aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 480 gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 540 tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 600 gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 660 agcctctccc tgtctccggg taaaggtgga ggtggtggtg cacagcagga agaatgcgaa 720 tgggacccat ggacttgcga acacatgctc gagtaa 756 <210> 45 <211> 822 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 45 atggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca 60 gttttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 120 acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 180 gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 240 taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 300 aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 360 aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc . 420 aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 480 gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 540 tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 600 gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 660 - 119 - 008248 agcctctccc tgtctccggg taaaggtgga ggtggtggtg cacagggatc cggttctgct actggtggtt ccggctccac cgcaagctct ggttcaggca gtgcgactca tcaggaagaa tgcgaatggg acccatggac ttgcgaacac atgctcgagt aa <210> 46 <211> 864 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 46 atggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca gttttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc . 120 acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 180 gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 240 taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 300 aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 360 aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 420 aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 480 gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 540 tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 600 gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 660 agcctctccc tgtctccggg taaaggtgga ggtggtggtg cacagcagga agaatgcgaa 720 tgggacccat ggacttgcga acacatggga tccggttctg ctactggtgg ttccggctcc 780 accgcaagct ctggttcagg cagcgcgact catcaggaag aatgcgaatg ggacccatgg 840 acttgcgaac acatgctcga gtaa 864 <210> 47 <211> 906 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 - 120 - 008248 <400> 47 atgggtgcac aggaagaatg cgaatgggac ccatggactt gcgaacacat gggtggtggt 60 ggtggtggcg gtggtaaatt caacccgctg gacgaactgg aagaaactct gtacgaacag 120 ttcactttcc agcagggatc cggttctgct actggtggtt ccggctccac cgcaagctct 180 ggttcaggca gtgcgactca tctcgagggt ggaggcggtg gggacaaaac tcacacatgt 240 ccaccttgcc cagcacctga actcctgggg ggaccgtcag ttttcctctt ccccccaaaa 300 cccaaggaca ccctcatgat ctcccggacc cctgaggtca catgcgtggt ggtggacgtg 360 agccacgaag accctgaggt caagttcaac tggtacgtgg acggcgtgga ggtgcataat 420 gccaagacaa agccgcggga ggagcagtac aacagcacgt accgtgtggt cagcgtcctc 480 accgtcctgc accaggactg gctgaatggc aaggagtaca agtgcaaggt ctccaacaaa 540 gccctcccag cccccatcga gaaaaccatc tccaaagcca aagggcagcc ccgagaacca 600 caggtgtaca ccctgccccc atcccgggat gagctgacca agaaccaggt cagcctgacc 660 tgcctggtca aaggcttcta tcccagcgac atcgccgtgg agtgggagag caatgggcag 720 ccggagaaca actacaagac cacgcctcec gtgctggact ccgacggctc cttcttcctc 780 tacagcaagc tcaccgtgga caagagcagg tggcagcagg ggaacgtctt ctcatgctcc 840 gtgatgcatg aggctctgca caaccactac acgcagaaga gcctctccct gtctccgggt 900 aaataa 906 <210> 48 <211> 897 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 48 atggacaaaa ctcacacatg tccaccttgc ccagcacctg aactcctggg gggaccgtca 60 gttttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 120 acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 180 gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 240 taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 300 aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 360 aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 420 aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 480 gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 540 tccgacggct ccttcttcct'ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 600 - 121 - 008248 gggaacgtct tctcatgctc cgtgatgcat gaggctctgc agcctctccc tgtctccggg taaaggtgga ggtggtggtg actggtggtt ccggctccac cgcaagctct ggttcaggca ccgctggacg aactggaaga aactctgtac gaacagttca ggtggtcagg aagaatgcga atgggaccca tggacttgcg <210> 49 <211> 771 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 49 atgggtgcac agttcgacta ctgcgaaggt gttgaagacc cgttcacttt cggttgcgac aaccacctcg agggtggagg cggtggggac aaaactcaca catgtccacc ttgcccagca 120 cctgaactcc tggggggacc gtcagttttc ctcttccccc caaaacccaa ggacaccctc 180 atgatctccc ggacccctga ggtcacatgc gtggtggtgg acgtgagcca cgaagaccct 240 gaggtcaagt tcaactggta cgtggacggc gtggaggtgc ataatgccaa gacaaagccg 300 cgggaggagc agtacaacag cacgtaccgt gtggtcagcg tcctcaccgt cctgcaccag 360 gactggctga atggcaagga gtacaagtgc aaggtctcca acaaagccct cccagccccc 420 atcgagaaaa ccatctccaa agccaaaggg cagccccgag aaccacaggt gtacaccctg 480 cccccatccc gggatgagct gaccaagaac caggtcagcc tgacctgcct ggtcaaaggc 540 ttctatccca gcgacatcgc cgtggagtgg gagagcaatg ggcagccgga gaacaactac 600 aagaccacgc ctcccgtgct ggactccgac ggctccttct tcctctacag caagctcacc 660 gtggacaaga gcaggtggca gcaggggaac gtcttctcat gctccgtgat gcatgaggct 720 ctgcacaacc actacacgca gaagagcctc tccctgtctc cgggtaaata 771 <210> 50 <211> 759 <212> ДНК <213> Искусственная последовательность acaaccacta cacgcagaag 660 cacagggatc cggttctgct 720 gtgcgactca taaattcaac 780 ctttccagca gggtggtggc 840 aacacatgct cgagtaa 897 - 122 - 008248 <400> 50 atgggtgcac agcagtacgg ttgcgacggt tttctgtacg gttgcatgat caacctcgag ggtggaggcg gtggggacaa aactcacaca tgtccacctt gcccagcacc tgaactcctg 120 gggggaccgt cagttttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 180 acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 240 aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 300 tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 360 ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 420 atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 480 gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 540 gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 600 cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 660 aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 720 tacacgcaga agagcctctc cctgtctccg ggtaaataa 759 <210> 51 <211> 759 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 51 atgggtgcac agaaacgccc atgcgaagaa atgtggggtg gttgcaacta cgacctcgag 60 ggtggaggcg gtggggacaa aactcacaca tgtccacctt gcccagcacc tgaactcctg 120 gggggaccgt cagttttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 180 acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 240 aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 300 tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 360 ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 420 atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 480 gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 540 gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 600 cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 660 aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 720 tacacgcaga agagcctctc cctgtctccg ggtaaataa 759 - 123 - 008248 <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 52 atgggtgcac agcaccagat ctgcaaatgg gacccgtgga cctgcaaaca ctggctcgag ggtggaggcg gtggggacaa aactcacaca tgtccacctt gcccagcacc tgaactcctg 120 gggggaccgt cagttttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 180 acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 240 aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 300 tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 360 ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 420 atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 480 gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 540 gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 600 cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 660 aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 720 tacacgcaga agagcctctc cctgtctccg ggtaaataa 759 <210> 53 <211> 759 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая "пептитела", способные связываться с Ang-2 <400> 53 atgggtgcac agaaacgtcc atgcgaagaa atcttcggtg gttgcaccta ccagctcgag 60 ggtggaggcg gtggggacaa aactcacaca tgtccacctt gcccagcacc tgaactcctg 120 gggggaccgt cagttttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 180 acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 240 aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 300 - 124 - 008248 tacaacagca ggcaaggagt atctccaaag gatgagctga gacatcgccg cccgtgctgg aggtggcagc tacacgcaga cgtaccgtgt acaagtgcaa ccaaagggca ccaagaacca tggagtggga actccgacgg aggggaacgt agagcctctc ggtcagcgtc ggtctccaac gccccgagaa ggtcagcctg gagcaatggg ctccttcttc cttctcatgc cctgtctccg ctcaccgtcc aaagccctcc ccacaggtgt acctgcctgg cagccggaga ctctacagca tccgtgatgc ggtaaataa tgcaccagga cagcccccat acaccctgcc tcaaaggctt acaactacaa agctcaccgt atgaggctct ctggctgaat cgagaaaacc cccatcccgg ctatcccagc gaccacgcct ggacaagagc gcacaaccac <210> 54 <211> 25 <212> ДНК <213> Искусственная последовательность <220> <223> Олигонуклеотид <400> 54 cggcgcaact atcggtatca agctg <210> 55 <211> 26 <212> ДНК <213> Искусственная последовательность <220> <223> Олигонуклеотид <400> 55 catgtaccgt aacactgagt ttcgtc <210> 56 <211> 14 <212> Белок <213> Искусственная последовательность 25 <220> <223> Консенсусные мотивы, образованные при использовании библиотеки TN8-LX - 125 - 008248 <220> <221> Смеш_ признаки <222> (7, 12 И )..(14) <223> Хаа относится к любой природной аминокислоте <400> 56 Lys Arg Pro cys Glu Glu xaa тгр Gly Gly cys xaa Tyr xaa 1 5 10 <210> - 57 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Консенсусные мотивы, образованные при использовании библиотеки TN8-LX <220> <221> смеш_признаки <222> С7, 12 И )..(14) <223> Хаа относится к любой природной аминокислоте <400> 57 Lys Arg Pro Cys Glu Glu Xaa Phe Gly Gly Cys Xaa Tyr xaa 15 10 <210> 58 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Консенсусные мотивы, образованные при использовании библиотеки TN8-LX <220> <221> смеш_признаки <222> С1, 2, 3, S и "Э..ОЗЭ <223> Xaa относится к любой природной аминокислоте - 126 - 008248 <400> 58 Хаа Хаа хаа cys хаа тгр Asp Pro Trp Thr cys Glu Xaa Met 1 5 10 <210> 59 <211> 18 <212> Белок <213> Искусственная последовательность <220> <223> Консенсусный мотив, образованный при использовании библиотеки TN12-I <220> <221> Смеш. признаки <222> СЗ, 8, 10-14 И )..СД8) <223> Хаа относится к любой природной аминокислоте <400> 59 Trp ser Хаа cys Ala Trp Phe Xaa Gly Xaa Xaa Xaa Xaa Xaa Cys Arg 15 10 15 Arg xaa <210> 60 <211> 227 <212> Белок <213> Искусственная последовательность <220> <223> Fc человеческого IgG l <400> 60 Asp Lys Thr His Thr cys pro pro Cys Pro Ala Pro Glu Leu Leu Gly 15 10 15 Gly Pro Ser val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met 20 25 30 lie ser Arg Thr Pro Glu Val Thr Cys val val Val Asp Val Ser His 35 40 45 - 127 - 008248 Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val 50 55 60 His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gin Tyr Asn ser Thr туг 65 70 75 80 Arg Val val ser Val Leu Thr Val Leu His Gin Asp Trp Leu Asn Gly 85 90 95 Lys Glu туг Lys cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro lie 100 105 110 Glu Lys Thr lie ser Lys Ala Lys Gly Gin Pro Arg Glu Pro Gin Val 115 120 125 Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gin Val Ser 130 135 140 Leu Thr cys Leu Val Lys Gly Phe Tyr Pro Ser Asp lie Ala val Glu 150 145 155 160 Trp Glu ser Asn Gly Gin Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro 165 170 175 Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val 180 185 190 Asp Lys Ser Arg Trp Gin Gin Gly Asn Val Phe Ser Cys Ser Val Met 195 200 205 His Glu Ala Leu His Asn His Tyr Thr Gin Lys Ser Leu Ser Leu ser 210 215 220 Pro Gly Lys 225 <210> 61 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Консенсусные мотивы, образованные при использовании библиотеки TN8-LX <220> <221> смеш_признаки - 128 - 008248 <222> 0--3, 5, 7, 12, 13 И).. (14) <223> -^аа охносится к любой природной аминокислоте Хаа Хаа хаа Cys хаа Asp Хаа туг Тгр туг cys Хаа Хаа хаа 1 5 10 <210> 62 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Консенсусные мотивы, образованные при использовании библиотеки TN8-IX <220> <221> смеш_признаки <222> (1-3, 5, 7, 12, 13 И ).•(14) <223> Xaa относится к любой природной аминокислоте Хаа Хаа Хаа cys Хаа Asp Хаа туг Thr туг Cys Хаа Хаа Хаа 15 10 <210> 63 <21Т> 14 <212> Белок <213> Искусственная последовательность <220> <223> Консенсусные мотивы, образованные при использовании библиотеки TN8-IX <220> <221> смеш_признаки <222> (1-3, 5, 7, 12, 13 И").. (14) <223> Хаа относится к любой природной аминокислоте Хаа Хаа Хаа Cys Хаа Asp Хаа Phe Trp Туг Cys Хаа Хаа Хаа 15 10 <210> 64 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Консенсусные мотивы, образованные при использовании библиотеки TN8-LX <220> <221> смеш_признаки <222> (1-3, 5, 7, 12, 13 И)..(14) <223> Хаа относится к любой природной аминокислоте <400> 64 Хаа хаа Хаа cys Хаа Asp Хаа Phe Thr Туг cys xaa Хаа хаа 1 5 10 <210> 65 <211> 5 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 65 Trp Asp Pro тгр Thr <210> 66 <211> 6 <212> Белок <213> Искусственная последовательность <220> - 129 - 008248 <223> Полипептид, способный связываться с Ang-2 <400> 66 Тгр Asp Pro тгр Thr Cys 1 5 <210> 67 <211> 7 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа обозначает кислый или нейтральный полярный аминокислотный остаток <400> 67 Cys Хаа Тгр Asp Pro Тгр Thr <210> 68 <211> 8 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа обозначает кислый или нейтральный полярный аминокислотный остаток <4О0> 68 cys хаа Trp Asp Pro Trp Thr Cys <210> 69 <211> 14 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <223> - смеш_признаки <222> С1, 2 И)..(3) <223> Xaa, каждый независимо, обозначает аминокислотный остаток <220> <221> смеш признаки <222> (5)..(5) <223> Хаа обозначает аминокислотный остаток <220> <221> смеш_признаки <222> (12)..(12) <223> :Хаа отсутствует или обозначает аминокислотный остаток <220> <221> смеш_признаки <222> (13) . . (13) <223> Хаа отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток <221> смеш_признаки <222> (14).. (14) <223> Хаа обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток <400> 69 Хаа хаа Xaa cys хаа Trp Asp Pro тгр Thr cys Хаа Хаа Хаа - 130 - 008248 15 10 <210> 70 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <221> смеш_признаки <222> С1 И )..(15) <223> Хаа отсутствует или обозначает аминокислотный остаток <220> <221> смеш_признаки <222> С2 И)..(16) <223> Хаа отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток <220> <221> смеш_признаки <222> (3-6, 18, 19 И )..(20) <223> Хаа, каждый независимо, отсутствуют или обозначают аминокислотные остатки <220> <221> смеш_гфизнаки <222> (8)..(8) <223> Хаа обозначает аминокислотный остаток <220> <221> смеш_признаки <222> (17)..(17) <223> ^аа 0ТСУтствУет или обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток <400> 70 Хаа Хаа xaa Хаа xaa xaa cys Хаа тгр Asp Pro тгр Thr cys xaa хаа 15 Ю 15 Хаа хаа хаа Хаа 20 <210> 71 <211> 10 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <221> смеш признаки <222> (2)..(2) <223> Хаа обозначает нейтральный гидрофобный аминокислотный остаток <220> <221> смеш_признаки <222> (4)..(4) <223> Хаа обозначает A, D или Е. <220> <221> смеш_признаки <222> (6),,(6) <223> Хаа обозначает кислый аминокислотный остаток <220> <221> смеш_признаки <222> (7)..С7) <223> Хаа обозначает аминокислотный остаток - 131 - 008248 <221> смеш_признаки <222> C8)•.(8) <223> Xaa обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток. <400> 71 pro Хаа Asp Хаа Leu Хаа Хаа Хаа Leu Туг 15 10 <210> 72 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <221> смеш_признаки <222> (1, 4 И )..(20) <223> Хаа отсутствует или обозначает аминокислотный остаток. <220> <221> смеш_признаки <222> (2, 15, 16 И).. (21) <223> Хаа отсутствует или обозначает нейтральный полярный, кислый или основной аминокислотный остаток. <220> <221> смеш_признаки <222> (3, 17 и').. (18) <223> Хаа отсутствует или обозначает нейтральный гидрофобный или нейтральный1 полярный аминокислотный остаток. <220> <221> смеш_признаки <222> (6).. (6) <223> Хаа обозначает нейтральный гидрофобный аминокислотный остаток. <220> <221> смеш_признаки <222> (8)..(8) <223> Хаа обозначает A, D или Е. <220> <221> смеш_признаки <222> (10)..(10) <223> Хаа обозначает кислый аминокислотный остаток. <220> <221> смешпризнаки <222> (11) ..(11) <223> Хаа обозначает аминокислотный остаток. <220> <221> смеш_признаки <222> (12)..(12) <223> Хаа обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток. <220> <221> смешпризнаки <222> (19 И )..(22) <223> Хаа отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток. <400> 72 Хаа Хаа Хаа Хаа pro Хаа Asp Хаа Leu хаа Хаа Хаа Leu Туг Хаа Хаа 15 10 15 Хаа хаа Хаа Хаа Хаа Хаа 20 - 132 - 008248 <211> 8 <2i2> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <221> смеш_признаки <222> (3)..(3) <223> Хаа обозначает нейтральный полярный аминокислотный остаток. <220> <221> смеш_признаки <222> (4)..С4) <223> Хаа обозначает кислый аминокислотный остаток. <220> <221> смеш_признаки <222> (5)..С5) <223> Хаа обозначает нейтральный полярный или кислый аминокислотный остаток. <220> <221> смеш_признаки <222> (6 И )..(7) <223> Хаа обозначает нейтральный гидрофобный аминокислотный остаток. <4О0> 73 Arq pro хаа Хаа Хаа хаа Хаа Gly 1 5 <210> 74 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <221> смеш_признаки <222> С1, 2, 4, 13, 14, 19 И )..С20) <223> Хаа. обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток. <220> <221> смеш признаки <222> СЗ, 9 И).. (17) <223> Хаа обозначает нейтральный полярный или кислый аминокислотный остаток. <220> <221> смеш_признаки <222> (7, 15 И).. (16) <223> Хаа обозначает нейтральный полярный аминокислотный остаток. <220> <221> смеш_признаки <222> (8)..(8) <223> Хаа обозначает кислый аминокислотный остаток. <220> <221> смешпризнаки <222> (10 И 1) .. (И) <223> Хаа обозначает нейтральный гидрофобный аминокислотный остаток. <220> <221> смеш_признаки <222> (18) .. (18) <223> Хаа обозначает нейтральный гидрофобный или основной аминокислотный остаток. <400> 74 - 133 - 008248 Хаа Хаа Хаа xaa Arg Pro xaa Хаа хаа xaa Хаа Gly Хаа Хаа Хаа Хаа 1 5 10 15 <210> 75 <211> 13 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа обозначает кислый аминокислотный остаток; <220> <221> смеш_признаки <222> (4)..(4) <223> ^аа обозначает нейтральный гидрофобный аминокислотный остаток. <220> <221> смеш_признаки <222> (5)..(5) <223> хаа обозначает Е, D или Q. <220> <221> смеш_признаки <222> (10)..(10) <223> Хаа обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток. <220> <221> смеш_признаки <222> (13)..(13) - 134 - 008248 <220> <223> Полипептад, способный связываться с Ang-2 <400> 78 тгр туг Glu Gin Asp Ala Cys Glu Trp Asp Pro Trp Thr Cys Glu His 1 5 10 15 Met Ala Glu Val 20 <210> 79 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 79 Asn Arg Leu Gin Glu val cys Glu Trp Asp Pro тгр Thr cys Glu His 15 10 15 Met Glu Asn Val 20 <210> 80 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 80 Ala Ala Thr Gin Glu Glu cys Glu Trp Asp pro тгр Thr cys Glu His 1 5 10 15 Met Pro Arg Ser 20 <210> 81 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 81 Leu Arg His Gin Glu Gly cys Glu Trp Asp Pro Trp Thr cys Glu His 15 10 15 Met Phe Asp Trp ZD <210> 82 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 82 Val Pro Arg Gin Lys Asp cys Glu Trp Asp Pro Trp Thr cys Glu His 15 10 15 Met Tyr Val Gly 20 <210> 83 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 83 Ser Пе Ser His Glu Glu Cys Glu Trp Asp Pro Trp Thr cys Glu His 1 5 10 15 Met Gin Val Gly 20 <210> 84 - 135 - 008248 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 84 тгр Ala Ala Gin Glu Glu cys Glu Trp Asp Pro Trp Thr cys Glu His 1 • 5 - - - - 10- - - - - - - 15 Met Gly Arg Met 20 <210> 85 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 85 Thr тгр Pro Gin Asp Lys Cys Glu Trp Asp Pro Trp Thr cys Glu His 15 10 15 Met Gly Ser Thr 20 <210> 86 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 86 Gly His ser Gin Glu Glu cys Gly Trp Asp Pro тгр Thr cys Glu His 1 5 10 15 Met Gly Thr ser <210> 87 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 87 Gin His Trp Gin Glu Glu cys Glu Trp Asp Pro Trp Thr cys Asp His 1 5 10 15 Met Pro ser Lys 20 <210> 88 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 88 Asn Val Arg Gin Glu Lys cys Glu Trp Asp Pro тгр Thr Cys Glu His 1 5 10 15 Met Pro val Arg 20 <210> 89 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьвзаться с Ang-2 <400> 89 Lys Ser Gly Gin Val Glu cys Asn Trp Asp Pro Trp Thr Cys Glu His - 136 - 008248 Met Pro Arg Asn 20 <210> 90 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <4О0> 90 val Lys Thr Gin Glu His cys Asp Trp Asp Pro Trp Thr cys Glu His 1 5 10 . хэ Met Arg Glu Trp 20 <210> 91 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 91 Ala Trp Gly Gin Glu Gly cys Asp Trp Asp Pro Trp Thr Cys Glu His 15 10 Met Leu Pro Met 20 <210> 92 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 - 137 - 008248 <220> <223> Полипепгид, способный связываться с Ang-2 <400> 95 pro Arg Leu Gin Glu Glu Cys Val тгр Asp pro Trp Thr cys Glu His 15 10 15 Met Pro Leu Arg 20 <210> 96 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 96 Arg Thr Thr Gin Glu Lys Cys Glu Trp Asp Pro Trp Thr Cys Glu His 15 10 15 Met Glu Ser Gin 20 <210> 97 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 97 Gin Thr ser Gin Glu Asp cys val Trp Asp Pro Trp Thr cys Asp His 15 10 15 Met val ser ser 20 <210> 98 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 98 Gin Val Пе Gly Arg Pro cys Glu Trp Asp Pro Trp Thr Cys Glu His 15 10 15 Leu Glu Gly Leu 20 <210> 99 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 99 тгр Ala Gin Gin Glu Glu cys Ala Trp Asp Pro тгр Thr cys Asp His 15 10 15 Met Val Gly Leu 20 <210> 100 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 100 Leu Pro Gly Gin Glu Asp cys Glu Trp Asp Pro Trp Thr cys Glu His 15 10 15 Met val Arg Ser 20 <210> 101 - 138 - 008248 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 101 pro Met Asn Gin val Glu cys Asp тгр Asp Pro тгр Thr Cys Glu His 1 5 10 15 Met Pro Arg Ser 20 <210> 102 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 102 Phe Gly Trp ser His Gly cys Glu Trp Asp Pro Trp Thr cys Glu His 15 10 15 Met Gly Ser Thr 20 <210> 103 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> ЮЗ Lys ser Thr Gin Asp Asp cys Asp Trp Asp Pro Trp Thr cys Glu His 15 10 15 Met val Gly Pro <210> 104 <211> 20 <212> Белок <213> Искусственная последовательность <223> Полипептид, способный связываться с Ang-2 <400> 104 Gly pro Arg Пе ser Thr cys Gin Trp Asp Pro Trp Thr cys Glu His 1 5 10 .15 Met Asp Gin Leu 20 <210> 105 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 105 Ser Thr lie Gly Asp Met Cys Glu Trp Asp Pro Trp Thr Cys Ala His 1 5 10 15 Met Gin Val Asp 20 <210> 106 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 106 val Leu Gly Gly Gin Gly cys Glu Trp Asp Pro Trp Thr cys Arg Leu - 139 - 008248 <400> 109 Thr Lys Gly Lys ser Val cys Gin Trp Asp Pro тгр Thr cys ser His 15 10 15 Met Gin Ser Gly 20 <210> 110 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 110 Thr Thr He Gly Ser Met Cys Gin Trp Asp Pro Trp Thr cys Ala His Met Gin Gly Gly 20 <210> 111 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 111 Trp Val Asn Glu Val Val Cys Glu Trp Asp Pro Trp Thr Cys Asn His 15 10 15 Trp Asp Thr Pro 20 <210> 112 <211> 20 <212> Белок <213> Искусственная последовательность - 140 - 008248 <220> <223> Полипептид, способный связываться с Ang-2 <4О0> 112 Val val Gin val Gly Met Cys Gin Trp Asp pro Trp Thr cys Lys His 15 10 15 Met Arg Leu Gin 20 <210> 113 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 113 Ala val Gly ser Gin Thr cys Glu Trp Asp Pro Trp Thr cys Ala His 1 5 10 15 Leu Val Glu val 20 <210> 114 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 114 Gin Gly Met Lys Met Phe cys Glu Trp Asp Pro Trp Thr cys Ala His 1 5 10 15 lie val Tyr Arg 20 <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 115 Thr Thr lie Gly Ser Met Cys Gin Trp Asp Pro Trp Thr cys Glu His 15 10 15 Met Gin Gly Gly 20 <210> 116 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 116 Thr Ser Gin Arg Val Gly cys Glu Trp Asp Pro Trp Thr Cys Gin His 15 10 15 Leu Thr Tyr Thr 20 <210> 117 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 117 Gin тгр ser тгр Pro Pro cys Glu Trp Asp Pro Trp Thr Cys Gin Thr 1 5 10 15 Val Trp Pro Ser 20 <210> 118 - 141 - 008248 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 118 Gly Thr ser Pro ser phe Cys Gin Trp Asp Pro Trp Thr Cys Ser His i ----- " 5 - - - - - - -10 ----- 15 Met val Gin Gly 20 <210> 119 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 119 Gin Asn Tyr Lys Pro Leu Asp Glu Leu,Asp Ala Thr Leu Tyr Glu His 1 5 10 lb Phe lie Phe His Tyr Thr 20 <210> . 120 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 120 Leu Asn Phe Thr Pro Leu Asp Glu Leu Glu Gin Thr Leu Tyr Glu Gin ! 5 10 15 Trp Thr Leu Gin Gin Ser <210> 121 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 121 Thr Lys Phe Asn Pro Leu Asp Glu Leu Glu Gin Thr Leu туг Glu Gin 1 5 10 .15 Trp Thr Leu Gin His Gin 20 <210> 122 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 122 val Lys Phe Lys Pro Leu Asp Ala Leu Glu Gin Thr Leu Tyr Glu His 15 10 15 Trp Met Phe Gin Gin Ala 20 <210> 123 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 123 Val Lys туг Lys Pro Leu Asp Glu Leu Asp Glu lie Leu туг Glu Gin - 142 - 008248 <400> 126 Gin Lys Phe Gin Pro Leu Asp Glu Leu Glu Gin Thr Leu Tyr Glu Gin 15 10 15 Phe Met Leu Gin Gin Ala 20 <210> 127 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 127 Gin Asn Phe Lys Pro Met Asp Glu Leu Glu Asp Thr Leu Tyr Lys Gin 1 5 10 15 Phe Leu Phe Gin His ser <210> 128 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 128 Tyr Lys Phe Thr Pro Leu Asp Asp Leu Glu Gin Thr Leu Tyr Glu Gin 15 10 15 Trp Thr Leu Gin His val 20 <210> 129 <211> 22 <212> Белок <213> Искусственная последовательность - 143 - 008248 <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 129 Gin Glu Туг Glu pro Leu Asp Glu Leu Asp Glu Thr Leu Tyr Asn Gin 15 10 15 Trp Met Phe His Gin Arg 20 <210> 130 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 130 Ser Asn Phe Met Pro Leu Asp Glu Leu Glu Gin Thr Leu Tyr Glu Gin 1 5 10 15 Phe Met Leu Gin His Gin 20 <210> 131 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 131 Gin Lys Tyr Gin Pro Leu Asp Glu Leu Asp Lys Thr Leu Tyr Asp Gin 15 10 15 Phe Met Leu Gin Gin Gly 20 <210> 132 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 132 Gin Lys Phe Gin Pro Leu Asp Glu Leu Glu Glu Thr Leu Tyr Lys Gin 15 10 15 Trp Thr Leu Gin Gin Arg 20 <210> 133 <2H> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 133 Val Lys туг Lys Pro Leu Asp Glu Leu Asp Glu Trp Leu Tyr His Gin 1 5 10 15 Phe Thr Leu His His Gin 20 <210> 134 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 134 Gin Lys Phe Met Pro Leu Asp Glu Leu Asp Glu lie Leu Tyr Glu Gin 1 5 10 15 Phe Met Phe Gin Gin ser 20 <210> 135 - 144 - 008248 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 135 Gin Tbr Phe Gin Pro Leu Asp Asp Leu Glu Glu Tyr Leu Tyr Glu Gin 1 "' T ---- - - -- ю----- " " 15 Trp lie Arg Arg Tyr His 20 <210> 136 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 136 Glu Asp Tyr Met Pro Leu Asp Ala Leu Asp Ala Gin Leu Tyr Glu Gin 15 10 15 Phe lie Leu Leu His Gly 20 <210> 137 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 137 His Thr Phe Gin Pro Leu Asp Glu Leu Glu Glu Thr Leu туг Tyr Gin 15 10 15 Trp Leu Tyr Asp Gin Leu <210> 138 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 138 Туг Lys Phe Asn pro Met Asp Glu Leu Glu Gin Thr Leu Tyr Glu Glu 1 5 10 15 Phe Leu Phe Gin His Ala 20 <210> 139 <2U> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 139 Thr Asn Tyr Lys pro Leu Asp Glu Leu Asp Ala Thr Leu Tyr Glu His 1 .5 10 15 Trp lie Leu Gin His ser 20 <210> 140 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 140 Gin Lys Phe Lys pro Leu Asp Glu Leu Glu Gin Thr Leu Tyr Glu Gin - 145 - 008248 Тгр Thr Leu Gin Gin Arg 20 <210> 141 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 141 Thr Lys Phe Gin Pro Leu Asp Glu Leu Asp Gin Thr Leu Tyr Glu Gin 15 10 15 Trp Thr Leu Gin Gin Arg 20 <210> 142 <211> 22 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 142 Thr Asn Phe Gin Pro Leu Asp Glu Leu Asp Gin Thr Leu Tyr Glu Gin 1 5 10 15 Trp Thr Leu Gin Gin Arg <210> 143 <211> 20 <212> Белок <213> Искусственная последовательность <220> <22з> Полипептид, способный связьшаться с Ang-2 <400> 143 Ala Gly Gly Met Arg Pro туг Asp Gly Met Leu Gly Trp Pro Asn Tyr 15 10 хь Asp Val Gin Ala 20 <210> 144 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 144 Gin Thr Trp Asp Asp Pro cys Met His lie Leu Gly Pro val Thr Trp 1 5 10 1:> Arg Arg Cys lie <210> 145 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 145 Ala Pro Gly Gin Arg Pro Tyr Asp Gly Met Leu Gly Trp Pro Thr Tyr 15 10 15 Gin Arg lie Val 20 <210> 146 <211> 20 <212> Белок <213> Искусственная последовательность - 146 - 008248 <220> <223> Полипептид, способный связываться с Ang-2 <400> 146 ser Gly Gin Leu Arg Pro Cys Glu Glu lie Phe Gly cys Gly Thr Gin 15 10 " Asn Leu Ala Leu 20 <210> 147 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 147 Phe Gly Asp Lys Arg Pro Leu Glu cys Met Phe Gly Gly Pro lie Gin 15 10 J-b Leu Cys Pro Arg 20 <210> 148 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способньш связьшаться с Ang-2 <400> 148 Gly Gin Asp Leu Arg Pro cys Glu Asp Met Phe Gly cys Gly Thr Lys 15 10 15 Asp тгр туг Gly <210> 149 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 149 Gly Phe Glu Tyr cys Asp Gly Met Glu Asp Pro phe Thr Phe Gly cys 15 10 15 Asp Lys Gin Thr <210> 150 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связываться с Ang-2 <400> 150 Lys Leu Glu Tyr Cys Asp Gly Met Glu Asp Pro Phe Thr Gin Gly Cys Asp Asn Gin Ser 20 <210> 151 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 151 Leu Gin Glu Trp cys Glu Gly Val Glu Asp Pro Phe Thr Phe Gly cys 1 5 10 15 Glu Lys Gin Arg 20 <210> 152 - 147 - 008248 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способньш связываться с Ang-2 <400> 152 Ala GLnJ^_BT_Q?s_Glu Gly Met Glu Asp_Pro Phe Thr Phe Gly cys 1 5 "10" Glu Met Gin Lys 20 <210> 153 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 153 Leu Leu Asp туг Cys Glu Gly Val Gin Asp Pro Phe Thr Phe Gly cys 1 5 10 хэ Glu Asn Leu Asp 20 <210> 154 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 154 His Gin Glu Tyr cys Glu Gly Met Glu Asp Pro Phe Thr Phe Gly cys 1 5 10 х:з Glu Tyr Gin Gly <210> 155 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 155 Met Leu Asp Tyr Cys Glu Gly Met Asp Asp pro Phe Thr Phe Gly Cys 15 10 15 Asp Lys Gin Met 20 <210> 156 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способный связьшаться с Ang-2 <400> 156 Leu Gin Asp Tyr cys Glu Gly val Glu Asp Pro Phe Thr Phe Gly cys 1 5 10 15 Glu Asn Gin Arg 20 <210> 157 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способньш связьшаться с Ang-2 <400> 157 Leu Gin Asp туг Cys Glu Gly Val Glu Asp pro Phe Thr Phe Gly cys - 148 - 008248 Glu Lys Gin Arg 20 <210> 158 <211> 20 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, способньш связьшаться с Ang-2 <220> <221> смеш_признаки <222> (1).. СО <223> Хаа отсутствует или обозначает нейтральный гидрофобный, нейтральный полярный или основной аминокислотный остаток. <220> <221> смеш_признаки <222> (2, 4, 14)..(19) <223> Хаа обозначает нейтральный гидрофобный или нейтральный полярный аминокислотный остаток. <220> <221> смеш_признаки <222> (3, 6).. (17) <223> Хаа обозначает кислый аминокислотный остаток. <220> <221> смеш_признаки <222> (8) .. (8) <223> Хаа обозначает нейтральный гидрофобный аминокислотный остаток. <220> <221> смеш_признаки <222> (9)..(9) <223> Хаа обозначает Е, D или Q. <220> <221> смеш_признаки <222> (18).. (18) <223> Хаа обозначает нейтральный гидрофобный, нейтральньш полярный или основной аминокислотный остаток. <220> <221> смеш_признаки <222> (20) .. (20) <223> Хаа отсутствует или обозначает аминокислотный остаток. <400> 158 Хаа Хаа Хаа Хаа cys Хаа Gly Хаа Хаа Asp Pro Phe Thr xaa Gly Cys 15 10 15 Xaa Xaa Xaa xaa 20 <210> 159 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. <400> 159 " ccgatccgtc aggaagaatg cgactgggac ccgtggacct gcgaacacat gtgggaagtt 60 <210> 160 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 160 - 149 - 008248 accaacatcc aggaagaatg cgaatgggac ccgtggacct gcgaccacat gccgggtaaa <210> 161 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связываться с Ang-2. <400> 161 tggtacgaac aggacgcttg cgaatgggac ccgtggacct gcgaacacat ggctgaagtt <210> 162 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 162 aaccgtctgc aggaagtttg cgaatgggac ccgtggacct gcgaacacat ggaaaacgtt <210> 163 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 163 gctgctaccc aggaagaatg cgaatgggac ccgtggacct gcgaacacat gccgcgttcc <210> 164 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 164 ctgcgtcacc aggaaggttg cgaatgggac ccgtggacct gcgaacacat gttcgactgg <210> 165 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 165 gttccgcgtc agaaagactg cgaatgggac ccgtggacct gcgaacacat gtacgttggt <210> 166 <211> 60 <212> ДНК <213> Искусственная последовательность <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 166 tgggctgctc aggaagaatg cgaatgggat ccgtggactt gcgaacacat gggtcgtatg 60 <210> 167 <211> 60 <212> ДНК <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 167 acttggccgc aggacaaatg cgaatgggat ccgtggactt gcgaacacat gggttctact <210> 168 <211> 60 <212> ДНК <213> Искусственная последовательность - 150 - 008248 <223> ДНК, кодирующая пептид, способньш связываться с Ang-2. <400> 168 ggtcactccc aggaagaatg cggttgggac ccgtggacct gcgaacacat gggtacgtcc <210> 169 <211> 60 <2i2> --ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 169 cagcactggc aggaagaatg cgaatgggac ccgtggacct gcgaccacat gccgtccaaa <210> 170 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 170 ^ ^ aacgttcgtc aggaaaaatg cgaatgggac ccgtggacct gcgaacacat gccggttcgt <210> 171 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 171 aaatccggtc aggttgaatg caactgggac ccgtggacct gcgaacacat gccgcgtaac <210> 172 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. gttaaaaccc aggaacactg cgactgggac ccgtggacct gcgaacacat gcgtgaatgg <210^-1Z3 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. gcttggggtc aggaaggttg cgactgggac ccgtggacct gcgaacacat gctgccgatg <210> 174 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. ccggttaacc aggaagactg cgaatgggac ccgtggacct gcgaacacat gccgccgatg <210> 175 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. cgtgctccgc aggaagactg cgaatgggac ccgtggacct gcgctcacat ggacatcaaa - 151 - 008248 <210> 176 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. cacggtcap"acarggaatg cgaatgggac ccgtggacct gcgaacacat gttccgttac---60 <210> 177 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. <210> 182 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. ccgcgtctgc aggaagaatg cgtttgggac ccgtggacct gcgaacacat gccgctgcgt <210> 178 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. cgtaccaccc aggaaaaatg cgaatgggac ccgtggacct gcgaacacat ggaatcccag <210> 179 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 179 cagacctccc aggaagactg cgtttgggac ccgtggacct gcgaccacat ggtttcctcc <210> 180 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <400> 180 caggttatcg gtcgtccgtg cgaatgggac ccgtggacct gcgaacacct ggaaggtctg 60 <210> 181 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. <400> 181 tgggctcagc aggaagaatg cgcttgggac ccgtggacct gcgaccacat ggttggtctg 60 <400> 182 ctgccgggtc aggaagactg cgaatgggac ccgtggacct gcgaacacat ggttcgttcc 60 <210> 183 <211> 60 <212> ДНК <213> Искусственная последовательность - 152 - 008248 <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. <400> 183 ccgatgaacc aggttgaatg cgactgggac ccgtggacct gcgaacacat gccgcgttcc <210> 184 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 184 ttcggttggt ctcacggttg cgaatgggat ccgtggactt gcgaacacat gggttctacc <210> 185 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 185 aaatccaccc aggacgactg cgactgggac ccgtggacct gcgaacacat ggttggtccg <210> 186 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 186 ggtccgcgta tctccacctg ccagtgggac ccgtggacct gcgaacacat ggaccagctg <210> 187 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 187 tccaccatcg gtgacatgtg cgaatgggac ccgtggacct gcgctcacat gcaggttgac 60 <210> 188 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. <400> 188 gttctgggtg gtcagggttg cgaatgggac ccgtggacct gccgtctgct gcagggttgg 60 <210> 189 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 189 gttctgggtg gtcagggttg ccagtgggac ccgtggacct gctcccacct ggaagacggt 60 <210> 190 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. <400> 190 accaccatcg gttccatgtg cgaatgggac ccgtggacct gcgctcacat gcagggtggt 60 <210> 191 - 153 - 008248 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 191 accaaaggta aatccgtttg ccagtgggac ccgtggacct gctcccacat gcagtccggt 60 <210> 192 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 192 accaccatcg gttccatgtg ccagtgggac ccgtggacct gcgctcacat gcagggtggt 60 <210> 193 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 193 tgggttaacg aagttgtttg cgaatgggac ccgtggacct gcaaccactg ggacaccccg 60 <210> 194 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 194 gttgttcagg ttggtatgtg ccagtgggac ccgtggacct gcaaacacat gcgtctgcag <210> 195 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> Дик, кодирующая пептид, способньш связьшаться с Ang-2. <400> 195 gctgttggtt cccagacctg cgaatgggac ccgtggacct gcgctcacct ggttgaagtt <210> 196 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 196 cagggtatga aaatgttctg cgaatgggac ccgtggacct gcgctcacat cgtttaccgt 60 <210> 197 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 197 accaccatcg gttccatgtg ccagtgggac ccgtggacct gcgaacacat gcagggtggt 60 <210> 198 <211> 60 <212> ДНК <213> Искусственная последовательность <220> - 154 - 008248 <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 198 acctcccagc gtgttggttg cgaatgggac ccgtggacct gccagcacct gacctacacc 60 <210> 199 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 199 cagtggtcct ggccgccgtg cgaatgggac ccgtggacct gccagaccgt ttggccgtcc 60 <210> 200 <211> 50 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 200 ggtacctccc cgtccttctg ccagtgggac ccgtggacct gctcccacat ggttcagggt 60 <210> 201 <211> 42 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 201 caggaagaat gcgaatggga cccatggact tgcgaacaca tg 42 <210> 202 <211> 66 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 202 cagaactaca aaccgctgga cgaactggac gctaccctgt acgaacactt catcttccac <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 204 accaaattca acccgctgga cgaactggaa cagaccctgt acgaacagtg gaccctgcag caccag <210> 205 <211> 66 <212> ДНК <213> Искусственная последовательность <220> <223> ; ДНК, кодирующая пептид, способньш связьшаться с Ang-2. <400> 205 <210> 203 <211> 66 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 203 ctgaacttca ccccgctgga cgaactggaa cagaccctgt acgaacagtg gaccctgcag bo cagtcc <210> 204 <211> 66 <212> ДНК <213> Искусственная последовательность - 155 - 008248 - 156 - 008248 <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 212 aggaatacga accgctggac gaactggacg aaaccctgta caaccagtgg atgttccacc agcgt <210> 213 <211> 66 <212> ДНК ^213> i_ Искусственная последовательность <220> <223> дик, кодирующая пептид, способный связываться с Ang-2. <400> 213 tccaacttca tgccgctgga cgaactggaa cagaccctgt acgaacagtt catgctgcag caccag <210> 214 <211> 66 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 214 cagaaatacc agccgctgga cgaactggac aaaaccctgt acgatcagtt catgctgcag cagggt <210> 215 <211> 66 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 215 cagaaattcc agccgctgga cgaactggaa gaaaccctgt acaaacagtg gaccctgcag cagcgt - 157 - 008248 <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 219 gaagactaca tgccgctgga cgctctggac gctcagctgt acgaacagtt catcctgctg cacggt <210> 220 _ <2!L1> . _66 . <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 220 cacaccttcc agccgctgga cgaactggaa gaaaccctgt actaccagtg gctgtacgac cagctg <210> 221 <211> 66 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связываться с Ang-2. <400> 221 tacaaattca acccgatgga cgaactggaa cagaccctgt acgaagaatt cctgttccag cacgct <210> 222 <211> 66 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 222 - 158 - 008248 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 226 aaattcaacc cgctggacga gctggaagag actctgtacg aacagtttac ttttcaacag 60 <210> - 227 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 227 gctggtggta tgcgtccgta cgacggtatg ctgggttggc cgaactacga cgttcaggct 60 <210> 228 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связываться с Ang-2. <400> 228 cagacttggg acgatccgtg catgcacatt ctgggtccgg ttacttggcg tcgttgcatc 60 <210> 229 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <4О0> 229 gctccgggtc agcgtccgta cgacggtatg ctgggttggc cgacctacca gcgtatcgtt 60 <210> 230 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. *;400> _ 23Q________________________ " tccggtcagc tgcgtccgtg cgaagaaatc ttcggftgcg дтатхсадая-х^лддс^д ш <210> 231 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. ttcggtgaca aacgtccgct ggaatgcatg ttcggtggtc cgatccagct gtgcccgcgt <210> 232 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связываться с Ang-2. ggtcaggacc tgcgtccgtg cgaagacatg ttcggttgcg gtaccaaaga ctggtacggt <210> 233 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. - 159 - 008248 <400> 233 ggtttcgaat actgcgacgg tatggaagac ccgttcacct tcggttgcga caaacagacc <210> 234 <211> 60 <212> ДНК <213> Искусственная последовательность <22ffe <223> ДНК, кодирующая пептид, способньш связьшаться с Ang-2. <400> 234 aaactggaat actgcgacgg tatggaagac ccgttcaccc agggttgcga caaccagtcc <210> 235 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 235 ctgcaggaat ggtgcgaagg tgttgaagac ccgttcacct tcggttgcga aaaacagcgt <2X0> 236 <211> 60 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 236 gctcaggact actgcgaagg tatggaagac ccgttcacct tcggttgcga aatgcagaaa <210> 237 <211> 60 <212> ДНК <213> Искусственная последовательность - 160 - 008248 <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 241 ctgcaggact actgcgaagg tgttgaagac ccgttcacct tcggttgcga aaaacagcgt 60 <210> 242 <211> 54 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <40Q> 242 ttcgactact gcgaaggtgt tgaagacccg ttcactttcg gctgtgataa ccac <210> 243 <211> 250 <212> ДНК <213> Искусственная последовательность <220> <223> ДНК, кодирующая пептид, способный связьшаться с Ang-2. <400> 243 Met Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu 15 10 15 Gly Gly Pro ser Val Phe Leu Phe pro Pro Lys Pro Lys Asp Thr Leu 20 25 30 Met He Ser Arg Thr Pro Glu val Thr cys Val Val val Asp Val Ser 35 40 45 His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr val Asp Gly Val Glu 50 . 55 60 Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gin туг Asn Ser Thr 65 70 75 80 Tyr Arg Val val Ser Val Leu Thr Val Leu His Gin Asp Trp Leu Asn 85 90 95 Gly Lys Glu Tyr Lys cys Lys Val ser Asn Lys Ala Leu Pro Ala Pro 100 Ю5. 110 Пе Glu Lys Thr He Ser Lys Ala Lys Gly Gin Pro Arg Glu Pro Gin 115 120 12b val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gin Val 130 135 140 ser Leu Thr cys Leu val Lys Gly Phe Tyr Pro ser Asp He Ala.Val Glu тгр Glu ser Asn Gly Gin Pro Glu Asn Asn туг Lys Thr Thr Pro 165 170 х'э Pro val Leu Asp Ser Asp Gly ser Phe Phe Leu туг Ser Lys Leu Thr val Asp Lys ser Arg Trp Gin Gin Gly Asn val Phe Ser cys ser val 195 200 ^u:> Met His Glu Ala Leu His Asn His туг Thr Gin Lys ser Leu ser Leu 210 215 220 ser Pro Gly Lys Gly Gly Gly Gly Gly cys Thr Ala Gly туг His Trp 225 230 235 Asn Ser Asp cys Glu cys Cys Arg Arg Asn 245 "U <210> 244 <211> 29 <212> ДНК <213> Искусственная последовательность <220> <223> Олигонуклеотид <400> 244 caaacgaatg gatcctcatt aaagccaga <210> 245 <2ll> 42 <212> ДНК <213> Искусственная последовательность - 161 - 008248 Искусственная последовательность 162 008248 <220> <223> Олигонуклеотид <400> 250 cys Ala Cys Ala Gly Thr Gly cys Ala cys Ala Gly Gly Gly Thr Asn 1 5 10 15 Asn Lys Ala Ala Lys Thr Thr Lys Ala Ala Lys cys Cys Lys cys Thr 20 25 30 -Lys-Gly-Ala_ Lys.Gly. Ala Lys Cys Thr Lys Gly Ala Lys Gly Ala Lys 35 40 " 45 Ala cys Lys cys Thr Lys Thr Ala Lys Gly Ala Lys cys Ala Lys Thr 50 55 60 Thr Lys Ala cys Lys Thr Thr Lys cys Ala Lys cys Ala Lys Asn Asn 65 70 75 80 Lys cys Ala Thr Thr cys Thr cys Thr cys Gly Ala Gly Ala Thr 85 90 95 <210> 251 <211> 91 <212> Белок <213> Искусственная последовательность <220> <223> Олигонуклеотид <400> 251 cys Ala cys Ala Gly Thr Gly cys Ala Cys Ala Gly Gly Gly Thr Asn 15 10 15 Asn Lys Asn Asn Lys Asn Asn Lys cys Ala Lys Gly Ala Lys Gly Ala 20 25 30 Lys Thr Gly Cys Gly Ala Lys Thr Gly Lys Gly Ala Lys cys Cys Lys Thr Gly Lys Ala cys Lys Thr Gly cys Gly Ala Lys Cys Ala Lys Ala 50 55 60 Thr Lys Asn Asn Lys Asn Asn Lys Asn Asn Lys Cys Ala Thr Thr Cys 65 70 75 80 Thr Cys Thr cys Gly Ala Gly Ala Thr cys Ala 85 9Q Lys Gly Ala Lys Gly Ala Lys cys Thr Lys Gly Ala Lys Gly Ala Lys 163 008248 - 164 - 008248 <210> 260 <211> 31 <212> Белок <213> Искусственная последовательность <220> <2-2-3> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> хаа = Fc <400> 260 Met Хаа Gly Gly Gly Gly Gly Ala Gin Thr Asn lie Gin Glu Glu cys 15 10 !5 Glu Trp Asp Pro Trp Thr Cys Asp His Met Pro Gly Lys Leu Glu 20 25 30 <210> 261 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <22l> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 261 Met хаа Gly Gly Gly Gly Gly Ala Gin Trp Tyr Glu Gin Asp Ala cys 1 5 10 15 Glu Trp Asp Pro Trp Thr Cys Glu His Met Ala Glu val Leu Glu 20 25 30 <210> 262 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> хаа = Fc <400> 262 Met Хаа Gly Gly Gly Gly Gly Ala Gin Asn Arg Leu Gin Glu Val Cys 15 io 15 Glu Trp Asp Pro Trp Thr cys Glu His Met Glu Asn Val Leu Glu 20 25 30 <210> 263 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2) .. С.2) <223> Хаа = Fc <400> 263 Met xaa Gly Gly Gly Gly Gly Ala Gin Ala Ala Thr Gin Glu Glu cys 15 10 15 Glu тгр Asp Pro тгр Thr cys Glu His Met Pro Arg Ser Leu Glu - 165 - 008248 <210> 264 <21Л> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2).-С2) <223> Хаа = Fc <400> 264 Met хаа Gly Gly Gly Gly Gly Ala Gin Leu Arg His Gin Glu Gly cys Glu Trp Asp Pro Trp Thr cys Glu His Met Phe Asp Trp Leu Glu 20 25 30 <2X0> 265 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 265 Met хаа Gly Gly Gly Gly Gly Ala Gin val Pro Arg Gin Lys Asp cys 1 5 10 15 Glu Trp Asp Pro Trp Thr Cys Glu His Met Tyr val Gly Leu Glu 20 25 30 <210> 266 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа = Fc <400> 266 Met хаа Gly Gly Gly Gly Gly Ala Gin ser He ser His Glu Glu cys Glu Trp Asp Pro Trp Thr cys Glu His Met Gin val Gly Leu Glu 20 25 30 <210> 267 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2D..(2) <223> Хаа = Fc <400> 267 Met хаа Gly Gly Gly Gly Gly Ala Gin Trp Ala Ala Gin Glu Glu Cys 1 5 10 15 Glu Trp Asp Pro Trp Thr cys Glu His Met Gly Arg Met Leu Glu - 166 - 008248 <210> 268 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> - "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc Met хаа Gly Gly Gly Gly Gly Ala Gin Thr Trp Pro Gin Asp Lys cys 1 5 10 15 Glu Tro ASD Pro TrD Thr cvs Glu His Met Gly Ser Thr Leu Glu ' ' 20 " 25 30 <210> 269 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc Met хаа Gly Gly Gly Gly Gly Ala Gin Gly His ser Gin Glu Glu cys 15 10 15 Gly Trp-Asp Pro Trp Thr cys Glu His Met Gly Thr Ser Leu Glu 20 25 30 <210> 270 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> Хаа = Fc Met хаа Gly Gly Gly Gly Gly Ala Gin Gin His Trp Gin Glu Glu Cys 1 5 10 15 Glu Trp Asp Pro тгр Thr Cys Asp His Met Pro ser Lys Leu Glu 20 25 30 <210> 271 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> хаа = Fc <400> 271 Met Xaa Gly Gly Gly Gly Gly Ala Gin Asn Val Arg Gin Glu Lys cys 1.5 10 15 Glu Trp Asp Pro Trp Thr cys Glu His Met Pro val Arg Leu Glu - 167 - 008248 <210> 272 <211> 31 <212> Белок <213> Искусственная последовательность <220> "223-> - "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc Met Хаа Gly Gly Gly Gly Gly Ala Gin Lys ser Gly Gin Val Glu cys 15 10 15 Asn тгЪ ASD Pro TTD Thr cvs Glu His Met"Pro Arg Asn Leu Glu 20 " ' 25 30 <210> 273 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <22l> смеш_признаки <222> (2)..(2) <223> хаа = Fc <400> 273 Met Хаа Gly Gly Gly Gly Gly Ala Gin Val Lys Thr Gin Glu His cys 15 10 15 Asp Trp Asp pro Trp Thr Cys Glu His Met Arg Glu Trp Leu Glu 20 25 30 <210> 274 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> - "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> Хаа = FC <400> 274 Met Хаа Gly Gly Gly Gly Gly Ala Gin Ala тгр Gly Gin Glu Gly cys 1 5 10 J-i Asp тгр Asp Pro тгр Thr cys Glu His Met Leu pro Met Leu Glu <210> 275 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)-.(2) <223> Хаа = Fc <400> 275 Met xaa Gly Gly Gly Gly Gly Ala Gin Pro Val Asn Gin Glu Asp cys 15 10 15 Glu Trp Asp Pro Trp Thr cys Glu His Met Pro Pro Met Leu Glu - 168 - 008248 <210> 276 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> - "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc Met Хаа Gly Gly Gly Gly Gly Ala Gin Arg Ala Pro Gin Glu Asp Cys 15 10 15 Glu Trp Asp Pro Trp Thr cys Ala His Met Asp lie Lys Leu Glu 20 25 30 <210> 277 <211> 31 <212> ; Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2).. С2) <223> Хаа = Fc Met xaa Gly Gly Gly Gly Gly Ala Gin His Gly Gin Asn Met Glu Cys 15 10 15 Glu Trp Asp Pro Trp Thr cys Glu Hi5 Met Phe Arg Tyr Leu Glu 20 25 30 <210> 278 <211> 31 <212> Белок <213> Искусственная последовательность <220> <:223> _- "Пептитела", способные связьшаться с Ang-2 <220> <221> смешпризнаки <222> С2)..С2) <223> Хаа = Fc <400> 278 Met Хаа Gly Gly Gly Gly Gly Ala Gin Pro Arg Leu Gin Glu Glu cys val Trp Asp Pro Trp Thr cys Glu His Met Pro Leu Arg Leu Glu <210> 279 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2),.(2) <223> xaa = Fc <400> 279 Met Хаа Gly Gly Gly Gly Gly Ala Gin Arg Thr Thr Gin Glu Lys cys 15 1° Х5 Glu тгр Asp Pro Trp Thr cys Glu His Met Glu Ser Gin Leu Glu - 169 - 008248 <210> 280 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> - "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> Хаа = Fc Met Хаа Gly Gly Gly Gly Gly Ala Gin Gin Thr ser Gin Glu Asp cys 1 5 10 15 Val Trp Asp Pro Trp Thr cys Asp His Met Val Ser Ser Leu Glu 20 25 30 <210> 281 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смешмпризнаки <222> (2)..С2 <223> Хаа = F <400> 281 Met Хаа Gly Gly Gly Gly Gly Ala Gin Gin Val lie Gly Arg Pro. cys 1 5 10 15 Glu Trp Asp Pro Trp Thr Cys Glu His Leu Glu Gly Leu Leu Glu 20 25 30 <210> 282 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> L "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc Met хаа Gly Gly Gly Gly Gly Ala Gin тгр Ala Gin Gin Glu Glu cys 15 10 15 Ala тгр Asp pro Trp Thr cys Asp His Met val Gly Leu Leu Glu 20 25 30 <210> 283 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 283 Met Xaa Gly Gly Gly Gly Gly Ala Gin Leu Pro Gly Gin Glu Asp Cys 15 10 15 Glu Trp Asp Pro тгр Thr cys Glu His Met val Arg Ser Leu Glu - 170 - 008248 <210> 284 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> - - "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 284 Met Хаа Gly Gly Gly Gly Gly Ala Gin Pro Met Asn Gin Val Glu cys 15 10 15 Asp Trp Asp Pro Trp Thr cys Glu His Met Pro Arg Ser Leu Glu 20 25 30 <210> 285 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 285 Met Хаа Gly Gly Gly Gly Gly Ala Gin Phe Gly тгр ser His Gly cys 1 5 10 15 Glu Trp Asp Pro тгр Thr Cys Glu His Met Gly Ser Thr Leu Glu 20 25 30 <210> 286 <211> 31 <212> Белок <213> Искусственная последовательность <220> , <223^ - "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2) .. (2) <223> Хаа = Fc <400> 286 Met Хаа Gly Gly Gly Gly Gly Ala Gin Lys Ser Thr Gin Asp Asp cys 15 10 15 Asp Trp Asp Pro Trp Thr cys Glu His Met Val Gly Pro Leu Glu 20 25 30 <210> 287 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 287 Met хаа Gly Gly Gly Gly Gly Ala Gin Gly Pro Arg lie ser Thr cys 1 . 5 10 15 Gin Trp Asp Pro Trp Thr cys Glu His Met Asp Gin Leu Leu Glu. - 171 - 008248 <210> 290 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 290 М 1 Met хаа Gly Gly Gly Gly Gly Ala Gin val Leu Gly Gly Gin Gly cys Gin Trp Asp Pro Trp Thr cys Ser His Leu Glu Asp Gly Leu Glu 20 25 30 <210> 291 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> Хаа = FC <400> 291 Met Хаа Gly Gly Gly Gly Gly Ala Gin Thr Thr lie Gly ser Met cys 15 10 lb Glu Trp Asp Pro Trp Thr cys Ala His Met Gin Gly Gly Leu Glu - 172 - 008248 <210> 292 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> хаа = Fc <400> 292 Met Хаа Gly Gly Gly Gly Gly Ala Gin Thr Lys Gly Lys ser val cys 15 10 15 Gin Trp Asp Pro Trp Thr cys ser His Met Gin ser Gly Leu Glu 20 25 30 <210> 293 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> Хаа = Fc <400> 293 Met xaa Gly Gly Gly Gly Gly Ala Gin Thr Thr lie Gly ser Met Cys 1 5 10 15 Gin Trp Asp pro тгр Thr cys Ala His Met Gin Gly Gly Leu Glu 20 25 30 <210> 294 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <22l> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 294 Met Хаа Gly Gly Gly Gly Gly Ala Gin Trp val Asn Glu val val cys 1 5 10 15 Glu Trp Asp Pro Trp Thr Cys Asn His Trp Asp Thr Pro Leu Glu 20 25 30 <210> 295 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> Хаа = Fc <400> 295 Met Хаа Gly Gly Gly Gly Gly Ala Gin val Val Gin val Gly Met cys 1 5 io 15 Gin Trp Asp Pro Trp Thr cys Lys His Met Arg Leu Gin Leu Glu - 173 - 008248 <210> 296 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222* С2)..(2) <223> Хаа = Fc Met Хаа Gly Gly Gly Gly Gly Ala Gin Ala Val Gly Ser Gin Thr Cys 1 5 10 15 Glu Trp Asp Pro Trp Thr cys Ala His Leu val Glu Val Leu Glu 20 25 30 <210> 297 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смешпризнаки <222> (2)..С2) <223> Хаа = Fc <400> 297 Met xaa Gly Gly Gly Gly Gly Ala Gin Gin Gly Met Lys Met phe cys 1 5 10 15 Glu Trp Asp Pro Trp Thr cys Ala His lie val Tyr Arg Leu Glu 20 25 30 <210> 298 <2U> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 298 Met хаа Gly Gly Gly Gly Gly Ala Gin Thr Thr lie Gly ser Met cys 1 5 10 15 Gin Trp Asp Pro Trp Thr cys Glu His Met Gin Gly Gly Leu Glu <210> 299 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2).. С2) <223> хаа = Fc <400> 299 Met Хаа Gly Gly Gly Gly Gly Ala Gin Thr Ser Gin Arg Val Gly Cys 15 10 15 Glu Trp Asp Pro Trp Thr Cys Gin His Leu Thr Tyr Thr Leu Glu - 174 - 008248 <210> 300 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 300 М < 1 Met xaa Gly Gly Gly Gly Gly Ala Gin Gin Trp ser Trp Pro Pro cys Glu Trp Asp Pro Trp Thr cys Gin Thr Val Trp Pro ser Leu Glu 20 25 5U <210> 301 <211> 31 <212> Белок <213: Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = FC Met Хаа Gly Gly Gly Gly Gly Ala Gin Gly Thr Ser Pro ser Phe cys <400> 301 Gin Trp Asp Pro Trp Thr cys ser His Met Val Gin Gly Leu Glu 20 25 30 <210> 302 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 302 Met Хаа Gly Gly Gly Gly Gly Ala Gin Thr Gin Gly Leu His Gin cys 1 5 10 lb Glu Trp Asp Pro Trp Thr Cys Lys val Leu тгр Pro Ser Leu Glu 20 25 30 <210> 303 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)-.С2) <223> Хаа = Fc <400> 303 Met Хаа Gly Gly Gly Gly Gly Ala Gin val тгр Arg Ser Gin val cys 1 5 10 I5 Gin Trp Asp Pro Trp Thr Cys Asn Leu Gly Gly Asp Trp Leu Glu - 175 - 008248 <210> 304 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> хаа = Fc <400> 304 Met хаа Gly Gly Gly Gly Gly Ala Gin Asp Lys He Leu Glu Glu cys Gin Trp Asp Pro тгр Thr cys Gin Phe Phe туг Gly Ala Leu Glu <210> 305 <211> 31 <212> Белок <213> Искусственная последовательность <220> "Пептитела", способные связываться с Ang-2 <223: <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 305 Met Хаа Gly Gly Gly Gly Gly Ala Gin Ala Thr Phe Ala Arg Gin cys Gln тгр Asp Pro Trp Thr cys Ala Leu Gly Gly Asn Trp Leu Glu 20 25 <210> 306 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <223> смеш_признаки <222> С2)..(2) <223> Хаа = Fc <400> 306 Met xaa Gly Gly Gly Gly Gly Ala Gin Gly Pro Ala Gin Glu Glu cys 15 10 J-b Glu Trp Asp Pro Trp Thr cys Glu pro Leu pro Leu Met Leu Glu <210> 307 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 307 Met Хаа Gly Gly Gly Gly Gly Ala Gin Arg Pro Glu Asp Met cys 15 10 I5 Gin Trp Asp pro Trp Thr тгр Hi's Leu Gin Gly Tyr Cys Leu Glu - 176 - 008248 <210> 308 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа = Fc <400> 308 Met Хаа Gly Gly Gly Gly Gly Ala Gin Leu тгр Gin Leu Ala val cys 15 10 15 Gin Trp Asp Pro Gin Thr cvs ASP His Met Gly Ala Leu Leu Glu '20 ' 25 ' 30 <210> 309 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> хаа = Fc <400> 309 Met Xaa Gly Gly Gly Gly Gly Ala Gin Thr Gin Leu Val ser Leu Cys 15 io 15 Glu Trp Asp Pro Trp Thr cys Arg Leu Leu ASP Gly Trp Leu Glu 20 25 30 - 177 - 008248 <210> 312 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа = Fc <400> 312 Met Xaa Gly Gly Gly Gly Gly Ala Gin Phe Gly Trp ser Hi's Gly Cys 1 5 10 15 Glu Trp Asp Pro тгр Thr cys Arg Leu Leu Gin Gly Trp Leu Glu 20 25 30 <210> 313 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 313 Met Хаа Gly Gly Gly Gly Gly Ala Gin Trp Pro Gin Thr Glu Gly cys 1 5 10 15 Gin Trp Asp Pro Trp Thr cys Arg Leu Leu His Gly Trp Leu Glu 20 25 30 <210> 314 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = FC <400> 314 Met Хаа Gly Gly Gly Gly Gly Ala Gin Pro Asp Thr Arg Gin Gly Cys 1 5 10 15 Gin Trp Asp Pro Trp Thr Cys Arg Leu Tyr Gly Met Trp Leu Glu 20 25 30 <210> 315 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)-.С2) <223> Хаа = Fc <400> 315 Met Хаа Gly Gly Gly Gly Gly Ala Gin Thr Trp Pro Gin Asp Lys cys 1 5 10 15 Glu Trp Asp Pro Trp Thr Cys Arg Leu Leu Gin Gly Trp Leu Glu - 178 - 008248 <210> 316 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа = Fc <400> 316 Met Хаа Gly Gly Gly Gly Gly Ala Gin Asp Lys lie Leu Glu Glu cys 15 io 15 Glu Trp Asp Pro Trp Thr cys Arg Leu Leu Gin Gly Trp Leu Glu 20 25 30 <210> 317 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 317 Met Хаа Gly Gly Gly Gly Gly Ala Gin Ala Ala Thr Gin Glu Glu Cys 1 5 10 15 Glu Trp Asp pro Trp Thr Cys Arg Leu Leu Gin Gly Trp Leu Glu 20 25 30 <210> 318 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (34) . . (34) <223> Хаа = Fc <400> 318 Met Gly Ala Gin Thr Asn Phe Met Pro Met Asp Asp Leu Glu Gin Arg 1 Г 5 10 15 Leu Tvr Glu Gin Phe He Leu Gin Gin Glv Leu Glu Gly Gly Gly Gly ' 20 25 30 Gly Xaa <210> 319 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (34)..(34) <223> Хаа = Fc <400> 319 Met Gly Ala Gin Thr Asn Tyr Lys Pro Leu Asp Glu Leu Asp Ala Thr - 179 - 008248 - 180 - 008248 <223> "Пептитела", способные связываться с Ang-2 <220> <223> смеш_признаки <222> (34) .. (34) <223> Хаа = Fc <400> 323 Met Gly Ala Gin Gin Glu туг "Glu Pro xeir Asp sTu t.^ Asp -Gl-u-Thr 1 ' 5 10 15 Leu Tyr Asn Gin Trp Met Phe His Gin Arg Leu Glu Gly Gly Gly Gly 20 25 30 Gly xaa <210> 324 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <223> смеш_признаки <222> (34) .. (34) <223> Хаа = Fc <400> 324 Met Gly Ala Gin Val Lys Tyr Lys Pro Leu Asp Glu Leu Asp Glu lie 15 io 15 Leu туг Glu Gin Gin Thr phe Gin Glu Arg Leu Glu Gly Gly Gly Gly 20 25 30 Gly Xaa <210> 325 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш признаки <222> (34) .. С34) <223> Хаа = Fc <400> 325 Met Gly Ala Gin Thr Lys Phe Gin Pro Leu Asp Glu Leu Asp Gin Thr 15 10 15 Leu Tyr Glu Gin Trp Thr Leu Gin Gin Arg Leu Glu Gly Gly Gly Gly 20 25 30 Gly xaa <210> 326 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (34) .. (34) <223> Хаа = Fc <400> 326 Met Gly Ala Gin Thr Asn Phe Gin Pro Leu Asp Glu Leu Asp Gin Thr 1 5 10 15 Leu Tyr Glu Gin Trp Thr Leu Gin Gin Arg Leu Glu Gly Gly Gly Gly 20 25 30 - 181 - 008248 - 182 - 008248 <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (34)..(34) <223> Хаа = Fc <400> 330 Met Gly Ala Gin Gin Asn Tyr Lys pro Leu Asp Glu Leu Asp Ala Thr 1 5 10 15 Leu Tyr Glu His Phe lie Phe His Tyr Thr Leu Glu Gly Gly Gly Gly 20 25 30 Gly Xaa <210> 331 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (34)..(34) <223> Хаа = Fc <400> 331 Met Gly Ala Gin val Lys Phe Lys Pro Leu Asp Ala Leu Glu Gin Thr 15 10 15 Leu Tyr Glu His Trp Met Phe Gin Gin Ala Leu Glu Gly Gly Gly Gly 20 25 30 Gly Xaa <210> 332 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (34) .. (34) <223> , Хаа = FC <400> 332 Met Gly Ala Gin Glu Asp Tyr Met Pro Leu Asp Ala Leu Asp Ala Gin 1 5 10 15 Leu Tyr Glu Gin Phe lie Leu Leu His Gly Leu Glu Gly Gly Gly Gly 20 25 30 Gly Xaa <210> 333 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (34) .. (34) <223> Хаа = Fc <4О0> 333 Met Gly Ala Gin Tyr Lys Phe Asn Pro Met Asp Glu Leu Glu Gin Thr 15 10 15 - 183 - 008248 Leu туг Glu Glu Phe Leu Phe Gin His Ala Leu Glu Gly Gly Gly Gly 20 25 30 Gly Xaa <210> 334 <211> 34 <212> Белок <213> " Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С34)..(34) <223> Хаа = Fc <400> 334 Met Gly Ala Gin Ser Asn Phe Met pro Leu Asp Glu Leu Glu Gin Thr 15 10 15 Leu Tyr Glu Gin Phe Met Leu Gin His Gin Leu Glu Gly Gly Gly Gly 20 25 30 Gly Xaa <210> 335 <211> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (34)..(34) <223> Хаа = Fc - 184 - 008248 - 185 - 008248 <210> 341 <21Т> 34 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С34).-С34) <223> Хаа = Fc <400> 341 Met Gly Ala Gin ser Lys phe Lys Pro Leu Asp Glu Leu Glu Gin Thr 1 5 10 ^ Leu туг Glu Gin Trp Thr Leu Gin His Ala Leu Glu Gly Gly Gly Gly 20 25 su Gly xaa <210> 342 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа = Fc <400> 342 Met Хаа Gly Gly Gly Gly Gly Ala Gin Ser Gly Gin Leu Arg Pro cys 1 5 10 15 Glu Glu lie Phe Gly cys Gly Thr Gin Asn Leu Ala Leu Leu Glu 20 25 30 <210> 343 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смёш_признаки <222> (2)..С2) <223> Хаа = Fc Met Хаа Gly Gly Gly Gly Gly Ala Gin Ala Gly Gly Met Arg Pro Tyr 15 10 15 Asp Gly Met Leu Gly Trp Pro Asn Tyr Asp Val Gin Ala Leu Glu 20 25 30 <210> 344 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> хаа = Fc <400> 344 Met Хаа Gly Gly Gly Gly Gly Ala Gin Gly Gin Asp Leu Arg Pro Cys - 186 - 008248 Glu Asp Met Phe Gly Cys Gly Thr Lys Asp Trp Tyr Gly Leu Glu 20 25 30 <210> 345 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc Met Хаа Glv Glv Glv Glv Glv Ala Gin Ala Pro Gly Gin Arg Pro Tyr 1 5 10 15 Asp Gly Met Leu Gly Trp Pro Thr Tyr Gin Arg lie Val Leu Glu 20 25 30 <210> 346 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа = Fc <400> 346 Met Хаа Gly Gly Gly Gly Gly Ala Gin Gin Thr Trp Asp Asp Pro Cys Met His lie Leu Gly Pro Val Thr Trp Arg Arg Cys lie Leu Glu 20 25 30 <210> 347 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа = Fc Met Хаа Gly Gly Gly Gly Gly Ala Gin Phe Gly Asp Lys Arg Pro Leu 1 5 10 15 Glu cys Met Phe Gly Gly Pro lie Gin Leu cys Pro Arg Leu Glu 20 25 30 <210> 348 <211> 25 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> . С2)..(2) <223> Хаа = Fc <400> 348 Met Хаа Gly Gly Gly Gly Gly Ala Gin Lvs Arg Pro cys Glu Glu lie - 187 - 008248 Phe Gly Gly Cys Thr Tyr Gin Leu Glu 20 25 <210> 349 <211> 31 <2i2> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> С2)..(2) <223> Хаа = Fc <400> 349 Met Хаа Gly Gly Gly Gly Gly Ala Gin Leu Gin Glu Trp Cys Glu Gly 15 10 15 val Glu Asp Pro Phe Thr Phe Gly Cys Glu Lys Gin Arg Leu Glu 20 25 30 <210> 350 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2) .. С2) <223> Хаа = Fc <400> 350 Met Хаа Gly Gly Gly Gly Gly Ala Gin Met Leu Asp Tyr Cys Glu Gly Met Asp Asp Pro Phe Thr Phe Gly cys Asp Lys Gin Met Leu Glu 20 25 30 <210> 351 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2)..С2) <223> Хаа = Fc <400> 351 Met Хаа Gly Gly Gly Gly Gly Ala Gin His Gin Glu Tyr cys Glu Gly 15 10 15 Met Glu Asp Pro Phe Thr Phe Gly Cys Glu туг Gin Gly Leu Glu 20 25 30 <210> 352 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2) • • С2) <223> Хаа = Fc <400> 352 Met Хаа Gly Gly Gly Gly Gly Ala Gin Leu Gin Asp Tyr cys Glu Gly - 188 - 008248 Val Glu Asp Pro Phe Thr Phe Gly cys Glu Asn Gin Arg Leu Glu 20 25 30 <210> 353 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 353 Met хаа Gly Gly Gly Gly Gly Ala Gin Leu Leu Asp Tyr Cys Glu Gly Val Gin Asp Pro Phe Thr Phe Gly Cys Glu Asn Leu Asp Leu Glu 20 25 30 <210> 354 <211> 31 <212> Белок <2l3> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 354 Met Хаа Gly Gly Gly Gly Gly Ala Gin Gly Phe Glu Tyr Cys Asp Gly 1 5 10 15 Met Glu Asp pro Phe Thr Phe Gly cys Asp Lys Gin Thr Leu Glu 20 25 30 <210> 355 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> (2)..(2) <223> Хаа = Fc Met xaa Gly Gly Gly Gly Gly Ala Gin Ala Gin Asp Tyr cys Glu Gly 1 5 10 15 Met Glu Asp Pro Phe Thr Phe Gly Cys Glu Met Gin Lys Leu Glu 20 25 30 <210> 356 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связьшаться с Ang-2 <220> <221> смеш_признаки <222> С2).-С2) <223> Хаа = Fc <400> 356 Met Хаа Gly Gly Gly Gly Gly Ala Gin Leu Gin Asp туг cys Glu Gly - 189 - 008248 val Glu Asp Pro Phe Thr Phe Gly cys Glu Lys Gin Arg Leu Glu 20 25 30 <210> 357 <211> 31 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 357 Met Хаа Gly Gly Gly Gly Gly Ala Gin Lys Leu Glu Tyr Cys Asp Gly 1 5 10 15 Met Glu Asp Pro Phe Thr Gin Gly Cys Asp Asn Gin Ser Leu Glu 20 25 30 <210> 358 <211> 29 <212> Белок <213> Искусственная последовательность <220> <223> "Пептитела", способные связываться с Ang-2 <220> <221> смеш_признаки <222> (2)..С2) <223> Хаа = Fc <400> 358 Met Хаа Gly Gly Gly Gly Gly Ala Gin Phe Asp Tyr cys Glu Gly Val Glu Asp Pro Phe Thr Phe Gly cys Asp Asn His Leu Glu 20 25 <210> 359 <211> 32 <212> Белок <213> Искусственная последовательность <220> <223> Полипептид, связывающийся с Ang-2 <400> 359 Cys Gly Gly Gly Gly Gly Ala Gin Thr Asn Phe Met Pro Met Asp Asp 1 5 10 15 Leu Glu Gin Arg Leu Tyr Glu Gin Phe lie Leu Gin Gin Gly Leu Glu 20 25 30 Евразийская патентная организация, ЕАПВ Россия, 109012, Москва, Малый Черкасский пер., 2/6 - 190 - |