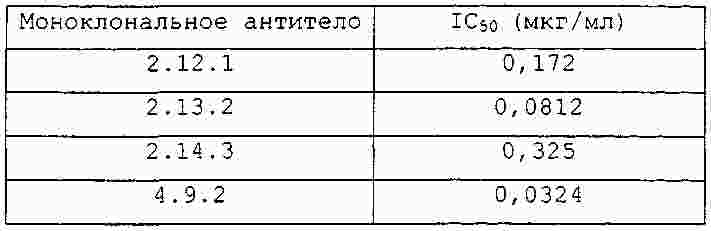
|
Патентная документация ЕАПВ |
|
||
| Запрос: | ea000012079b*\id |
|
Термины запроса в документе Реферат Данное изобретение относится к антителам и их антигенсвязывающим частям, которые специфично связываются с рецептором инсулиноподобного фактора роста I (IGF-IR), который предпочтительно является IGF-IR человека. Изобретение также относится к человеческим анти-IGF-IR-антителам, включая химерные, биспецифичные, дериватизированные, одноцепочечные антитела или части слитых белков. Изобретение также относится к выделенным молекулам тяжелой и легкой цепи иммуноглобулина, полученным из анти-IGF-IR-антител, и молекулам нуклеиновой кислоты, кодирующим такие молекулы. Данное изобретение также относится к способам получения анти-IGF-IR-антител, фармацевтическим композициям, содержащим указанные антитела, и способам применения антител и их композиций для диагностики и лечения. Изобретение также относится к способам генотерапии с использованием молекул нуклеиновой кислоты, кодирующих молекулы тяжелой и/или легкой цепей иммуноглобулина, которые включают человеческие анти-IGF-IR-антитела. Изобретение также относится к способам генотерапии и трансгенным животным, содержащим молекулы нуклеиновой кислоты согласно данному изобретению. Формула [0001] Человеческое моноклональное антитело или его антигенсвязывающая часть, которые специфически связываются с рецептором инсулиноподобного фактора роста I (IGF-IR), где указанное антитело или его часть ингибируют индуцированное IGF-IR фосфорилирование тирозина. [0002] Моноклональное антитело или его антигенсвязывающая часть по п.1, обладающие по меньшей мере одним из следующих свойств: [0003] Моноклональное антитело или его антигенсвязывающая часть по п.2, обладающие всеми указанными свойствами. [0004] Моноклональное антитело или его антигенсвязывающая часть по п.2, которые ингибируют связывание IGF-I или IGF-II с IGF-IR, имеют Kd для IGF-IR, составляющую 8 ×10-9 М или менее, и обладают избирательностью в отношении IGF-IR, которая по меньшей мере в 50 раз превышает их избирательность в отношении рецептора инсулина. [0005] Моноклональное антитело или его антигенсвязывающая часть по п.1 или 2, обладающие по меньшей мере одним из следующих свойств: [0006] Моноклональное антитело или его антигенсвязывающая часть по любому из пп.1-5, ингибирующие связывание IGF-IR с IGF-I или IGF-II с IC50, составляющей менее 100 нМ. [0007] Моноклональное антитело или его антигенсвязывающая часть по любому из пп.1-6, содержащие вариабельную область легкой цепи, которая содержит не более 10 изменений аминокислот по сравнению с аминокислотной последовательностью, кодируемой геном VK A30, A27 или O12 зародышевой линии. [0008] Моноклональное антитело или его антигенсвязывающая часть по любому из пп.1-6, содержащие вариабельную область тяжелой цепи, которая содержит не более 8 изменений аминокислот по сравнению с аминокислотной последовательностью, кодируемой геном VH DP47, DP35, DP71 или VIV-4 зародышевой линии. [0009] Моноклональное антитело или его антигенсвязывающая часть по любому из пп.1-8, содержащие аминокислотную последовательность по меньшей мере одной CDR вариабельной области, выбранной из группы, состоящей из: [0010] Моноклональное антитело или его антигенсвязывающая часть по любому из пп.1-8, содержащие аминокислотную последовательность по меньшей мере одной CDR вариабельной области, выбранной из группы, состоящей из: [0011] Моноклональное антитело или его антигенсвязывающая часть по п.9, содержащие все аминокислотные последовательности CDR вариабельной области, выбранной из группы, состоящей из: [0012] Моноклональное антитело или его антигенсвязывающая часть по п.10, содержащие все аминокислотные последовательности CDR вариабельной области, выбранной из группы, состоящей из: [0013] Моноклональное антитело или его антигенсвязывающая часть по п.11, отличающиеся тем, что вариабельная область легкой цепи антитела содержит аминокислотную последовательность, выбранную из SEQ ID NO:2, SEQ ID NO: 6, SEQ ID NO: 10, SEQ ID NO: 14 и SEQ ID NO: 18, или аминокислотную последовательность, имеющую 1-10 аминокислотных инсерций, делеций или замен в указанных последовательностях. [0014] Моноклональное антитело или его антигенсвязывающая часть по п.11, отличающиеся тем, что вариабельная область тяжелой цепи антитела содержит аминокислотную последовательность, выбранную из SEQ ID NO:4, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 16 и SEQ ID NO: 20, или аминокислотную последовательность, имеющую 1-10 аминокислотных инсерций, делеций или замен в указанных последовательностях. [0015] Моноклональное антитело или его антигенсвязывающая часть по п.1, отличающиеся тем, что указанное антитело выбрано из группы, состоящей из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 и 4.17.3. [0016] Моноклональное антитело по любому из пп.1-15, отличающееся тем, что указанное антитело относится к изотипу IgG, IgM, IgE, IgA или IgD. [0017] Моноклональное антитело или его антигенсвязывающая часть по любому из пп.1-15, представляющие собой Fab-фрагмент, F(ab')2-фрагмент, Fv-фрагмент, одноцепочечное антитело, гуманизированное антитело, химерное антитело или биспецифическое антитело. [0018] Моноклональное антитело по п.1, содержащее тяжелую и легкую цепи, причем аминокислотные последовательности тяжелой цепи и легкой цепи выбраны из группы, состоящей из: [0019] Моноклональное антитело по п.1, содержащее тяжелую и легкую цепи, причем аминокислотные последовательности тяжелой цепи и легкой цепи выбраны из группы, состоящей из: [0020] Моноклональное антитело по п.1, содержащее аминокислотную последовательность, содержащую CDR антител 2.12.1 или 2.13.2, или последовательности CDR указанных антител, имеющие не более 5 консервативных аминокислотных замен. [0021] Моноклинальное антитело, которое специфически связывается с IGF-IR и ингибирует индуцированное IGF-IR фосфорилирование тирозина, содержащее аминокислотную последовательность вариабельной области тяжелой цепи, содержащую CDR1, CDR2 и CDR3 последовательности SEQ ID NO:4, и аминокислотную последовательность вариабельной области легкой цепи, содержащую CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 2. [0022] Моноклональное антитело по п.21, отличающееся тем, что аминокислотная последовательность тяжелой цепи содержит аминокислотную последовательность вариабельной области SEQ ID NO:4 или указанную последовательность, содержащую вплоть до 5 консервативных аминокислотных замен, и аминокислотная последовательность легкой цепи содержит аминокислотную последовательность вариабельной области SEQ ID NO: 2 или указанную последовательность, содержащую вплоть до 5 аминокислотных замен. [0023] Моноклональное антитело по п.22, отличающееся тем, что аминокислотная последовательность тяжелой цепи содержит аминокислотную последовательность вариабельной области SEQ ID NO:4 и аминокислотная последовательность легкой цепи содержит аминокислотную последовательность вариабельной области SEQ ID NO: 2. [0024] Моноклональное антитело по любому из пп.21-23, отличающееся тем, что аминокислотная последовательность тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:49 без сигнальной последовательности и аминокислотная последовательность легкой цепи содержит аминокислотную последовательность SEQ ID NO: 51 без сигнальной последовательности. [0025] Моноклональное антитело, которое специфически связывается с IGF-IR и ингибирует индуцированное IGF-IR фосфорилирование тирозина, содержащее аминокислотную последовательность вариабельной области тяжелой цепи, содержащей CDR1, CDR2 и CDR3 последовательности SEQ ID NO:8, и аминокислотную последовательность вариабельной области легкой цепи, содержащей CDR1, CDR2 и CDR3 последовательности SEQ ID NO: 6. [0026] Моноклональное антитело по п.25, отличающееся тем, что аминокислотная последовательность тяжелой цепи содержит аминокислотную последовательность вариабельной области SEQ ID NO:8 или указанную последовательность, содержащую вплоть до 5 консервативных аминокислотных замен, и аминокислотная последовательность легкой цепи содержит аминокислотную последовательность вариабельной области SEQ ID NO: 6 или указанную последовательность, содержащую вплоть до 5 аминокислотных замен. [0027] Моноклональное антитело по п.26, отличающееся тем, что аминокислотная последовательность тяжелой цепи содержит аминокислотную последовательность вариабельной области SEQ:ID NO: 8 и аминокислотная последовательность легкой цепи содержит аминокислотную последовательность вариабельной области SEQ: ID NO: 6. [0028] Моноклональное антитело по любому из пп.25-27, отличающееся тем, что аминокислотная последовательность тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:45 без сигнальной последовательности и аминокислотная последовательность легкой цепи содержит аминокислотную последовательность SEQ ID NO: 47 без сигнальной последовательности. [0029] Фармацевтическая композиция, содержащая моноклональное антитело или его антигенсвязывающую часть по любому из пп.1-28 и фармацевтически приемлемый носитель. [0030] Фармацевтическая композиция по п.29, дополнительно содержащая антинеопластическое, химиотерапевтическое или противоопухолевое средство. [0031] Способ получения моноклонального анти-IGF-IR-антитела или его антигенсвязывающей части по любому из пп.1-28, предусматривающий следующие стадии: [0032] Выделенная линия клеток, которая продуцирует моноклональное антитело по любому из пп.1-28. [0033] Линия клеток по п.32, отличающаяся тем, что продуцирует моноклональное антитело, выбранное из группы, состоящей из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 и 4.17.3, или антитело, которое имеет те же самые аминокислотные последовательности, что и указанные антитела. [0034] Способ диагностики наличия или локализации опухоли, экспрессирующей IGF-IR, у нуждающегося в этом человека, предусматривающий стадии: [0035] Способ лечения рака у человека, включающий стадию введения человеку антитела или его антигенсвязывающей части по любому из пп.1-28 в количестве, эффективном для лечения указанного рака. [0036] Способ по п.34 или 35, дополнительно предусматривающий стадию введения человеку антинеопластического, противоопухолевого, антиангиогенного или химиотерапевтического средства. [0037] Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, кодирующую тяжелую цепь или ее антигенсвязывающую часть или легкую цепь или ее антигенсвязывающую часть моноклонального антитела по любому из пп.1-28. [0038] Выделенная молекула нуклеиновой кислоты по п.37, содержащая последовательность нуклеиновой кислоты, выбранную из группы, состоящей из: [0039] Вектор, содержащий молекулу нуклеиновой кислоты по любому из пп.37 или 38, при необходимости содержащий последовательность регуляции экспрессии, функционально связанную с молекулой нуклеиновой кислоты. [0040] Клетка-хозяин, содержащая вектор по п.39 или молекулу нуклеиновой кислоты по любому из пп.37 или 38. [0041] Способ получения моноклонального анти-IGF-IR-антитела или его антигенсвязывающей части, предусматривающий культивирование клетки-хозяина по п.40 или линии клеток по п.32 в подходящих условиях и выделение указанного антитела или его антигенсвязывающей части. [0042] Трансгенное животное, отличное от человека, содержащее молекулу нуклеиновой кислоты по любому из пп.37 или 38, экспрессирующее указанную нуклеиновую кислоту. [0043] Способ лечения рака у человека, включающий стадию введения человеку эффективного количества молекулы нуклеиновой кислоты по любому из пп.37 или 38 с обеспечением ее экспрессии. Полный текст патента Данная заявка претендует на приоритет по предварительной заявке США 60/259927, поданной 5 января 2001г. Инсулиноподобный фактор роста (IGF-I) является полипептидом с М.м. 7,5 кД, циркулирующим в плазме в высоких концентрациях и обнаруживаемым в большинстве тканей. IGF-I стимулирует дифференцировку и пролиферацию клеток и необходим для длительной пролиферации большинства типов клеток млекопитающих. Указанные типы клеток, помимо прочих, включают в себя диплоидные фибробласты, эпителиальные клетки, гладкомышечные клетки, Т-лимфоциты, нервные клетки, миелоидные клетки, хондроциты, остеобласты и стволовые клетки костного мозга человека. С целью обзора обширного множества типов клеток, в которых клеточная пролиферация опосредована взаимодействием IGF-I/IGF-I-рецептора, см. Goldring et al., Eukar. Gene Express., 1: 31-326 (1991). Первым этапом пути трансдукции, ведущего к стимулируемой IGF-I пролиферации или дифференцировке клеток, является связывание IGF-I или IGF-II (или инсулина в концентрациях выше физиологических) с рецептором IGF-I. Рецептор IGF-I состоит из субъединиц двух типов: альфа-субъединицы (полностью внеклеточный белок в 130-135 кД, который функционирует при связывании с лигандом) и бета-субъединицы (трансмембранный белок в 95 кД с трансмембранным и цитоплазматическим доменами). IGF-IR относится к семейству тирозинкиназных рецепторов факторов роста (Ullrich et al., Cell 61: 203-212, 1990) и по структуре сходен с рецептором инсулина (Ullrich et al., EMBO J. 5: 2503-2512, 1986). IGF-IR сначала синтезируется в виде одноцепочечного полипептида прорецептора, который затем подвергается процессингу посредством гликозилирования, протеолитического расщепления и ковалентного связывания, образуя зрелый гетеротетрамер с М.м. 460 кД, содержащий две альфа-субъединицы и две бета-субъединицы. Бета-субъединица(цы) обладает активируемой лигандом тирозинкиназной активностью. Указанная активность участвует в путях передачи сигнала, которыми опосредовано действие лиганда, заключающееся в аутофосфорилировании бета-субъединицы и фосфорилировании субстратов IGF-IR. Уровни IGF-I в сыворотке in vivo зависят от присутствия гипофизарного гормона роста (GH). Хотя печень является основным местом зависимого от GH синтеза IGF-I, недавняя работа показала, что большинство нормальных тканей также продуцируют IGF-I. Множество неопластических тканей также могут продуцировать IGF-I. Таким образом, IGF-I может действовать как регулятор нормальной и аномальной пролиферации клеток посредством аутокринного или паракринного, а также эндокринного механизмов. IGF-I и IGF-II связываются с IGF-связывающими белками (IGFBP) in vivo. Доступность свободного IGF для взаимодействия с IGF-IR модулируется IGFBP. Для обзора IGFBP и IGF-I см. Grimberg et al., J. Cell. Physiol. 183: 1-9, 2000. Есть немало доказательств роли IGF-I и/или IGF-IR в поддержании опухолевых клеток in vitro и in vivo. Уровни IGF-IR повышены в опухолях легкого (Kaiser et al., J. Cancer Res. Clin. Oncol. 119: 665-668, 1993; Moody et al., Life Sciences 52: 1161-1173, 1993; Macauley et al., Cancer Res., 50: 2511-2517, 1990), молочной железы (Pollak et al., Cancer Lett. 38: 223-230, 1987; Foekens et al., Cancer Res. 49: 7002-7009, 1989; Cullen et al., Cancer Res. 49: 7002-7009, 1990; Arteaga et al., J. Clin. Invest. 84: 1418-1423, 1989), простаты и толстой кишки (Remaole-Bennet et al., J. Clin. Endocrinol. Metab. 75: 609-616, 1992; Guo et al., Gastroenterol. 102: 1101-1108, 1992). Нерегулируемая экспрессия IGF-I в эпителии простаты ведет к неоплазии у трансгенных мышей (DiGiovanni et al., Proc. Natl. Acad. Sci. USA 97: 3455-60, 2000). Кроме того, оказалось, что IGF-I является аутокринным стимулятором глиом человека (Sandberg-Nordqvist et al., Cancer Res. 53: 2475-2478, 1993), в то время как IGF-I стимулировал рост фибросарком, которые сверхэкспрессировали IGF-IR (Butler et al., Cancer Res. 58: 3021-27, 1998). Кроме того, для субъектов с "высокими нормальными" уровнями IGF-I повышен риск развития общих злокачественных опухолей по сравнению с субъектами с уровнями IGF-I в "низких нормальных" пределах (Rosen et al., Trends Endocrinol. Metab. 10: 136-41, 1999). Многие из указанных типов опухолевых клеток отвечают на IGF-I пролиферативным сигналом в культуре (Nakanishi et al., J. Clin. Invest. 82: 354-359, 1988; Freed et al., J. Mol. Endocrinol. 3: 509-514, 1989), и высказано предположение об аутокринной или паракринной петлях пролиферации in vivo (LeRoith et al., Endocrine Revs. 16: 143-163, 1995; Yee et al., Mol. Endocrinol. 3: 509-514, 1989). Для обзора того, какую роль взаимодействие IGF-I/IGF-I-рецептор играет в росте различных опухолей человека, см. Macaulay, Br. J. Cancer, 65: 311-320, 1992. Повышенные уровни IGF-I также коррелируют с несколькими незлокачественными патологическими состояниями, включая акромегалию и гигантизм (Barkan, Cleveland Clin. J. Med. 65: 343, 347-349, 1998), в то время как аномальное функционирование IGF-I/IGF-I-рецептор вовлечено в псориаз (Wraight et al., Nat. Biotech. 18: 521-526, 2000), атеросклероз и рестеноз гладкой мускулатуры кровеносных сосудов после ангиопластики (Bayes-Genis et al., Circ. Res. 86: 125-130, 2000). Повышенные уровни IGF-I также могут быть проблемой при диабете и его осложнениях, таких как пролиферация микрососудов (Smith et al., Nat. Med. 5: 1390-1395, 1999). Пониженные уровни IGF-I, которые, в частности, могут иметь место в тех случаях, когда уровни GH в сыворотке снижены или когда существует нечувствительность или резистентность к GH, связаны с такими нарушениями, как маленький рост (Laron, Paediatr. Drugs 1: 155-159, 1999), нейропатия, уменьшение мышечной массы и остеопороз (Rosen et al., Trends Endocrinol. Metab. 10: 136-141, 1999). С использованием антисмысловых экспрессирующих векторов или антисмысловых олигонуклеотидов к РНК IGF-IR было показано, что воздействие IGF-IR приводит к ингибированию опосредованного IGF-I или опосредованного IGF-II роста клеток (см., например, Wraight et al., Nat. Biotech. 18: 521-526, 2000). Антисмысловая стратегия оказалась успешной при ингибировании клеточной пролиферации в нескольких нормальных типах клеток и линиях клеток опухолей человека. Рост также можно ингибировать с использованием пептидного аналога IGF-I (Pietrzkowski et al., Cell Growth and Diff. 3: 199-205, 1992; и Pietrzkowski et al., Mol. Cell. Biol., 12: 3883-3889, 1992), или вектора, экспрессирующего антисмысловую РНК к РНК IGF-I (Trojan et al., Science 259: 94-97, 1992). Кроме того, антитела к IGF-IR (Arteaga et al., Breast Canc. Res. Treatm., 22: 101-106, 1992; и Kalebic et al., Cancer Res. 54: 5531-5534, 1994) и доминантные негативные мутанты IGF-IR (Prager et al., Proc. Natl. Acad. Sci. U.S.A. 91: 2181-2185, 1994; Li et al., J. Biol. Chem., 269: 32558-32564, 1994 и Jiang et al., Oncogene 18: 6071-77, 1999) могут реверсировать трансформированный фенотип, ингибировать образование опухолей и индуцировать потерю метастазирующего фенотипа. IGF-I также имеет важное значение в регуляции апоптоза. Апоптоз, который является запрограммированной гибелью клеток, вовлечен в широкое множество процессов развития, включая созревание иммунной и нервной системы. Сделан вывод, что кроме роли в развитии апоптоз также является важной клеточной защитой от образования опухолей (Williams, Cell 65: 1097-1098, 1991; Lane, Nature 362: 786-787, 1993). Подавление программы апоптоза за счет множества генетических повреждений может вносить вклад в развитие и прогрессирование злокачественных новообразований. IGF-I защищает IL-3-зависимые гемопоэтические клетки от апоптоза, индуцированного удалением цитокина (Rodriguez-Tarduchy, G. et al., J. Immunol. 149: 535-540, 1992), и клетки Rat-1/mycER при удалении сыворотки (Harrington, Е., et al., EMBO J. 13: 3286-3295, 1994). Противоапоптозная функция IGF-I имеет важное значение на стадии, следующей после задержки клеточного цикла, а также в клетках, прохождение клеточного цикла которых блокировано этопозидом или тимидином. Демонстрация того, что жизнеспособность регулируемых c-myc фибробластов зависит от IGF-I, свидетельствует о том, что IGF-IR играет важную роль в поддержании опухолевых клеток посредством специфичного ингибирования апоптоза, роль, отличную от пролифератизного действия IGF-I или IGF-IR. Это подобно той роли, которую, как считается, играют другие противоапоптозные гены, такие как bcl-2 в стимулировании жизнеспособности опухолей (McDonnell et al., Cell 57: 79-88, 1989; Hockenberry et al., Nature 348: 334-336, 1990). Защитное действие IGF-I от апоптоза зависит от имеющихся IGF-IR, присутствующих на клетках для взаимодействия с IGF-I (Resnicoff et al., Cancer Res. 55: 3739-3741, 1995). Подтверждение противоапоптозной функции IGF-IR при поддержании опухолевых клеток также получили при исследовании с использованием антисмысловых олигонуклеотидов к IGF-IR, в котором выявлена количественная взаимосвязь между уровнями IGF-IR, степенью апоптоза и канцерогенным потенциалом сингенной опухоли крыс (Rescinoff et al., Cancer Res. 55: 3739-3741, 1995). Обнаружено, что сверхэкспрессированный IGF-IR защищает опухолевые клетки in vitro от апоптоза, индуцированного этопозидом (Sell et al., Cancer Res. 55: 303-306, 1995), и, что еще более поразительно, что снижение уровней IGF-IR ниже уровней дикого типа вызывало массовый апоптоз опухолевых клеток in vivo (Resnicoff et al., Cancer Res. 55: 2463-2469, 1995). Возможная методика индукции апоптоза или ингибирования клеточной пролиферации, связанной с повышенными уровнями IGF-I, повышенными уровнями IGF-II и/или повышенными уровнями рецептора IGF-IR, включает подавление уровней IGF-I или уровней IGF-II или предотвращение связывания IGF-I с IGF-IR. Например, длительно действующий аналог соматостатина октреотид использовали для снижения синтеза и/или секреции IGF. Растворимый IGF-IR использовали для того, чтобы индуцировать апоптоз в опухолевых клетках in vivo и ингибировать образование опухоли в экспериментальной системе на животных (D'Ambrosio et al., Cancer Res. 56: 4013-20, 1996). Кроме того, антисмысловые олигонуклеотиды IGF-IR, пептидные аналоги IGF-I и антитела к IGF-IR использовали для снижения экспрессии IGF-I или IGF-IR (см. выше). Однако ни одно из указанных соединений не подходило для долговременного введения больным людям. Кроме того, несмотря на то, что IGF-I вводили пациентам для лечения низкого роста, остеопороза, пониженной мышечной массы, нейропатии или диабета, связывание IGF-I с IGFBP часто делало лечение IGF-I трудным или неэффективным. Таким образом, принимая во внимание роль, которую играют IGF-I и IGF-IR в таких нарушениях, как рак и другие пролиферативные расстройства, в том случае, когда IGF-I и/или IGF-IR сверхэкспрессированы, и роли, которые играют слишком малые количества IGF-I и IGF-IR при таких нарушениях, как низкий рост и хрупкость костей, в том случае, когда либо IGF-I, и/или IGF-IR недоэкспрессированы, было бы желательно создать антитела к IGF-IR, которые можно использовать либо для ингибирования, либо для стимулирования IGF-IR. Хотя сообщалось, что анти-IGF-IR-антитела были обнаружены у некоторых пациентов с аутоиммунными заболеваниями, ни одно из обнаруженных антител не было очищено и не было показано ни для одного из них, что оно подходит для ингибирования активности IGF-I для диагностических и клинических способов. См., например, Thompson et al., Pediat. Res. 32: 455-459, 1988; Tappy et al., Diabetes 37: 1708-1714, 1988; Weightman et al., Autoimmunity 16: 251-257, 1993; Drexhage et al., Nether. J. of Med. 45: 285-293, 1994. Таким образом, является актуальным получение высокоаффинных анти-IGF-IR-антител человека, которые можно использовать для лечения заболеваний у людей. На фиг. 1А-1С показано сопоставление нуклеотидных последовательностей вариабельных областей легких цепей шести анти-IGF-IR-антител человека по отношению друг к другу и к последовательностям зародышевой линии. На фиг. 1А показано сопоставление нуклеотидных последовательностей вариабельной области легкой цепи (VL) антител 2.12.1 (SEQ ID NO: 1), 2.13.2 (SEQ ID NO: 5), 2.14.3 (SEQ ID NO: 9) и 4.9.2 (SEQ ID NO: 13) друг с другом и с последовательностью V κ A30 зародышевой линии (SEQ ID NO: 39). На фиг. 1В показано сопоставление нуклеотидной последовательности VL антитела 4.17.3 (SEQ ID NO: 17) с последовательностью V κ 012 зародышевой линии (SEQ ID NO: 41). На фиг. 1С показано сопоставление нуклеотидной последовательности VL антитела 6.1.1 (SEQ ID NO: 21) с последовательностью V κ A27 зародышевой линии (SEQ ID NO: 37). Также показаны сопоставления CDR-областей VL из каждого антитела. Консенсусные последовательности для фиг. 1А-1С показаны в SEQ ID NO: 53-55, соответственно. На фиг. 2A-2D показаны сопоставления нуклеотидных последовательностей вариабельных областей тяжелых цепей шести анти-IGF-IR-антител человека по отношению друг к другу и к последовательностям зародышевой линии. На фиг. 2А показано сопоставление нуклеотидной последовательности VH антитела 2.12.1 (SEQ ID NO: 3) с последовательностью VH DP-35 зародышевой линии (SEQ ID NO: 29). На фиг. 2В показано сопоставление нуклеотидной последовательности VH антитела 2.14.3 (SEQ ID NO: 11) с последовательностью VIV-4/4.35 зародышевой линии (SEQ ID NO: 43). На фиг. 2С-1 и 2С-2 показаны сопоставления нуклеотидных последовательностей VH антител 2.13.2 (SEQ ID NO: 7), 4.9.2 (SEQ ID NO: 15) и 6.1.1 (SEQ ID NO: 23) друг с другом и с последовательностью VH DP-47 зародышевой линии (SEQ ID NO: 31). На фиг. 2D показано сопоставление нуклеотидной последовательности VH антитела 4.17.3 (SEQ ID NO: 19) с последовательностью VH DP-71 зародышевой линии (SEQ ID NO: 35). Также показаны сопоставления CDR-областей антител. Консенсусные последовательности для фиг. 2A-2D показаны в SEQ ID NO: 56-59, соответственно. На фиг. 3 показано, что анти-IGF-IR-антитела 2.13.2, 4.9.2 и 2.12.1 ингибируют связывание IGF-I с клетками 3T3-IGF-IR. На фиг. 4 показано, что анти-IGF-IR-антитело 4.9.2 ингибирует индуцированное IGF-I фосфорилирование рецептора тирозина (верхняя панель) и индуцирует понижающую регуляцию IGF-IR на поверхности клеток (нижняя панель). На фиг. 5 показано, что анти-IGF-IR-антитела 2.13.2 и 4.9.2 уменьшают сигнал фосфотирозина IGF-IR в опухолях 3T3-IGF-IR. На фиг. 6 показано, что анти-IGF-IR-антитела 2.13.2 и 4.9.2 уменьшают количество IGF-IR в опухолях 3T3-IGF-IR. На фиг. 7 показано, что анти-IGF-IR-антитело 2.13.2 отдельно (левая панель) или в комбинации с адриамицином (правая панель) ингибирует рост опухоли 3T3-IGF-IR in vivo. На фиг. 8 показана взаимосвязь между уровнями анти-IGF-IR-антитела 2.13.2 в сыворотке и понижающей регуляцией IGF-IR в опухолях 3T3-IGF-IR. На фиг. 9 показано, что многократные дозы анти-IGF-IR-антитела 2.13.2 отдельно или в комбинации с адриамицином ингибируют рост опухоли 3T3-IGF-IR in vivo. На фиг. 10 показано, что анти-IGF-IR-антитело 2.13.2 в комбинации с адриамицином ингибирует рост крупной опухоли in vivo. На фиг. 11 показано, что анти-IGF-IR-антитело 2.13.2 отдельно или в комбинации с 5-дезоксиуридином (5-FU) ингибирует рост опухоли Colo 205 in vivo. На фиг. 12 показано, что многократные дозы анти-IGF-IR-антитела 2.13.2 отдельно или в комбинации с 5-FU ингибируют рост опухоли Colo 205 in vivo. На фиг. 13 показано, что многократные дозы анти-IGF-IR-антитела 2.13.2 отдельно или в комбинации с таксолом ингибируют рост опухоли MCF-7 in vivo. На фиг. 14 показано, что анти-IGF-IR-антитело 2.13.2 отдельно (левая панель) или в комбинации с адриамицином (правая панель) ингибирует рост опухоли MCF-7 in vivo. На фиг. 15 показано, что многократные дозы анти-IGF-IR-антитела 2.13.2 отдельно или в комбинации с тамоксифеном ингибируют рост опухоли MCF-7 in vivo. На фиг. 16 показано, что многократные дозы анти-IGF-IR-антитела 2.13.2 отдельно или в комбинации с ингибитором тирозинкиназы рецептора эпидермального фактора роста (EGF-R) СР-358774 ингибируют рост опухоли А431 in vivo. На фиг. 17 показана фармакокинетическая оценка однократной внутривенной инъекции анти-IGF-IR-антитела 2.13.2 макакам-крабоедам. На фиг. 18 показано, что комбинация анти-IGF-IR-антитела 2.13.2 и адриамицина увеличивает понижающую регуляцию IGF-IR на опухолях 3T3-IGF-IR in vivo. На фиг. 19А показан ряд мутаций в различных областях тяжелой и легкой цепей 2.13.2 и 2.12.1 по сравнению с последовательностями зародышевой линии. На фиг. 19В-Е показаны сопоставления аминокислотных последовательностей тяжелой и легкой цепей антител 2.13.2 и 2.12.1 с последовательностями зародышевой линии, из которых они получены. На фиг. 19В показано сопоставление аминокислотной последовательности тяжелой цепи антитела 2.13.2 (SEQ ID NO: 45) с последовательностью зародышевой линии DP-47 (3-23)/D6-19/JH6 (SEQ ID NO: 46). На фиг. 19С показано сопоставление аминокислотной последовательности легкой цепи антитела 2.13.2 (SEQ ID NO: 47) с последовательностью зародышевой линии A30/Jk2 (SEQ ID NO: 48). На фиг. 19D показано сопоставление аминокислотной последовательности тяжелой цепи антитела 2.12.1 (SEQ ID NO: 49) с последовательностью зародышевой линии DP-35 (3-11)/D3-3/JH6 (SEQ ID NO: 50). На фиг. 19Е показано сопоставление аминокислотной последовательности легкой цепи антитела 2.12.1 (SEQ ID NO: 51) с последовательностью зародышевой линии А30/Jk1 (SEQ ID NO: 52). На фиг. 19В-Е сигнальные последовательности указаны курсивом, CDR подчеркнуты, вариабельные домены выделены жирным шрифтом, мутации каркасной области (FR) выделены знаком плюс ("+") над остатком аминокислоты и мутации CDR выделены звездочкой над остатком аминокислоты. Данное изобретение относится к выделенному антителу или его антигенсвязывающей части, которая связывает IGF-IR, предпочтительно к антителу, которое связывает IGF-IR примата или человека, и более предпочтительно к антителу, которое является антителом человека. Изобретение относится к анти-IGF-IR-антителу, которое ингибирует связывание IGF-I или IGF-II с IGF-IR, а также относится к анти-IGF-IR-антителу, которое активирует IGF-IR. Изобретение относится к фармацевтической композиции, содержащей антитело и фармацевтически приемлемый носитель. Фармацевтическая композиция может дополнительно содержать другой компонент, такой как противоопухолевый агент или реагент для визуализации. Изобретение также относится к диагностическим и терапевтическим способам. Диагностические способы включают способ диагностики наличия или локализации ткани, экспрессирующей IGF-IR, с использованием анти-IGF-IR-антитела. Терапевтический способ включает введение антитела нуждающемуся в этом субъекту, предпочтительно в сочетании с другим терапевтическим средством. Изобретение относится к выделенной линии клеток, такой как гибридома, которая продуцирует анти-IGF-IR-антитело. Изобретение также относится к молекулам нуклеиновых кислот, кодирующих тяжелую и/или легкую цепь анти-IGF-IR-антитела или их антигенсвязывающие части. Изобретение относится к векторам и клеткам-хозяевам, содержащим молекулы нуклеиновых кислот, а также к способам рекомбинантного получения полипептидов, кодируемых молекулами нуклеиновых кислот. Также представлены трансгенные животные, отличные от человека, которые экспрессируют тяжелую и/или легкую цепь анти-IGF-IR-антитела или их антигенсвязывающие части. Изобретение также относится к способу лечения нуждающегося в этом субъекта с помощью эффективного количества молекулы нуклеиновой кислоты, кодирующей тяжелую и/или легкую цепь анти-IGF-IR-антитела или их антигенсвязывающие части. Если не оговорено особо, научные и технические термины, используемые в связи с данным изобретением, будут иметь значения, которые обычно подразумеваются специалистами в данной области. Кроме того, если не подразумевается иное, исходя из контекста, термины в единственном числе будут включать понятие во множественном числе, а термины во множественном числе будут включать единственное число. Как правило, используемая номенклатура и методики, связанные с культурой клеток и тканей, молекулярной биологией, иммунологией, микробиологией, генетикой и химией белков и нуклеиновых кислот и гибридизацией, приведенные в данном описании, являются хорошо известными и обычно используемыми в данной области. Способы и методики согласно данному изобретению обычно осуществляют в соответствии со стандартными способами, хорошо известными в данной области, и так, как описано в различных общих и более конкретных источниках информации, которые цитированы и обсуждаются в данном описании, если не указано особо. См., например, Sambrook et al. Molecular Cloning: A Laboratory Manual, 2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. (1989) и Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates (1992), и Harlow and Lane Antibodies: A Laboratory Manual Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. (1990), которые включены в данное описание в виде ссылки. Ферментативные реакции и способы очистки выполняют согласно описаниям производителя, которые обычно прилагаются в данной области, или как описано в данной заявке. Используемая номенклатура и лабораторные способы, связанные с аналитической химией, химией органического синтеза и медицинской и фармацевтической химией, приведенные в данном описании, хорошо известны и широко используются в данной области. Стандартные способы используют в случае химического синтеза, химического анализа, фармацевтического препарата, композиции и доставки и лечения пациентов. Если не оговорено особо, следует понимать, что следующие термины имеют нижеприведенные значения. Термин "полипептид" охватывает нативные или искусственные белки, белковые фрагменты и полипептидные аналоги последовательности белка. Полипептид может быть мономерным или полимерным. Термин "выделенный белок" или "выделенный полипептид" означает белок или полипептид, которые в силу своей природы или источника происхождения: (1) не связаны с компонентами, связанными в природных условиях, которые сопутствуют ему в нативном состоянии, (2) не содержат других белков из того же самого вида, (3) экспрессируются клеткой другого вида или (4) не встречаются в природе. Таким образом, полипептид, который синтезирован химическим путем или синтезирован в клеточной системе, отличной от клетки, из которой он происходит в естественных условиях, будет "выделенным" от связанных с ним в природе компонентов. Белок также можно сделать в значительной степени свободным от компонентов, связанных с ним в природных условиях, посредством выделения с использованием способов очистки белка, хорошо известных в данной области. Белок или полипептид является "в значительной степени чистым", "в значительной степени гомогенным" или "в значительной степени очищенным" в том случае, когда по меньшей мере примерно 60-75% образца представлено одним видом полипептида. Полипептид или белок может быть мономерным или полимерным. В значительной степени чистый полипептид или белок обычно будет содержать примерно 50, 60, 70, 80 или 90% мас./мас. образца белка, более обычно примерно 95% и предпочтительно будет очищен более чем на 99%. Чистота или гомогенность белка может быть показана рядом способов, хорошо известных в данной области, таких как электрофорез образца белка в полиакриламидном геле с последующей визуализацией отдельной полосы полипептида при окрашивании геля красителем, хорошо известным в данной области. Для некоторых целей более высокое разрешение можно получить с использованием ВЭЖХ или других способов, хорошо известных в данной области для очистки. Термин "полипептидный фрагмент" в используемом в данном описании смысле относится к полипептиду, который имеет делецию по аминоконцу и/или по карбоксильному концу, но в котором сохраняется аминокислотная последовательность, идентичная соответствующим положениям последовательности природного происхождения. Обычно фрагменты имеют длину по меньшей мере из 5, 6, 8 или 10 аминокислот, предпочтительно имеют длину, по меньшей мере равную 14 аминокислотам, более предпочтительно длину, по меньшей мере равную 20 аминокислотам, обычно длину, по меньшей мере равную 50 аминокислотам, еще более предпочтительно длину, по меньшей мере равную 70, 80, 90, 100, 150 или 200 аминокислотам. Термин "аналог полипептида" в используемом в данном описании смысле относится к полипептиду, который состоит из участка по меньшей мере из 25 аминокислот, который в значительной степени идентичен части аминокислотной последовательности и который обладает по меньшей мере одним из следующих свойств: (1) специфичное связывание с IGF-IR при подходящих условиях связывания, (2) способность блокировать связывание IGF-I или IGF-II с IGF-IR или (3) способность уменьшать экспрессию IGF-IR на поверхности клеток или фосфорилирование тирозина in vitro или in vivo. Обычно аналоги полипептида содержат консервативную аминокислотную замену (или инсерцию или делецию) относительно последовательности природного происхождения. Обычно аналоги имеют длину, равную по меньшей мере 20 аминокислотам, предпочтительно длину, равную по меньшей мере 50, 60, 70, 80, 90, 100, 150 или 200 аминокислотам или длиннее, и часто могут иметь длину полноразмерного полипептида природного происхождения. Предпочтительными аминокислотными заменами являются замены, которые: (1) уменьшают чувствительность к протеолизу, (2) уменьшают чувствительность к окислению, (3) изменяют аффинность связывания при образовании белковых комплексов, (4) изменяют аффинности связывания и (4) придают или модифицируют другие физико-химические или функциональные свойства таких аналогов. Аналоги могут включать различные мутеины с последовательностью, отличной от последовательности пептида природного происхождения. Например, можно произвести единичные или множественные аминокислотные замены (предпочтительно консервативные аминокислотные замены) в последовательности природного происхождения (предпочтительно в части полипептида вне домена(нов), образующего внутримолекулярные контакты). Консервативная аминокислотная замена не должна в значительной степени изменять структурные характеристики исходной последовательности (например, замена аминокислоты не должна способствовать разрушению спирали, которая имеется в исходной последовательности, или разрушать другие типы вторичной структуры, которые характеризуют исходную последовательность). Примеры общеизвестных в данной области вторичных и третичных структур полипептидов описаны в Proteins, Structures and Molecular Principles (Creighton, Ed., W.H. Freeman and Company, New York (1984)); Introduction to Protein Structure (C. Branden and J. Tooze, eds., Garland Publishing, New York, N. Y. (1991)); и Thornton et at. Nature 354: 105 (1991), которые включены в данное описание в виде ссылки. Непептидные аналоги широко используют в фармацевтической промышленности в качестве лекарственных средств со свойствами, аналогичными свойствам матричного пептида. Указанные типы непептидного соединения называют "миметиками пептида" или "пептидомиметиками". Fauchere, J. Adv. Drug Res. 15: 29 (1986); Veber and Freidinger TINS p. 392 (1985); и Evans et al. J. Med. Chem. 30: 1229 (1987), которые включены в данное описание в виде ссылки. Такие соединения часто разрабатывают с помощью компьютеризованного молекулярного моделирования. Миметики пептидов, которые структурно сходны с терапевтически полезными пептидами, можно использовать для получения эквивалентного терапевтического или профилактического действия. Как правило, пептидомиметики структурно сходны с образцом полипептида (т.е. полипептидом, который обладает требуемым биохимическим свойством или фармакологической активностью), таким как антитело человека, но имеют одну или более пептидных связей, необязательно замененных связью, выбранной из группы, состоящей из -CH 2 NH-, -CH 2 S-, -СН 2 -СН 2 -, -СН=СН- (цис- и транс-), -СОСН 2 -, -СН(ОН)СН 2 - и -CH 2 SO-, способами, хорошо известными в данной области. Направленную замену одной или более аминокислот консенсусной последовательности D-аминокислотой того же самого типа (например, D-лизином вместо L-лизина) также можно использовать для создания более стабильных пептидов. Кроме того, ограниченные пептиды, содержащие консенсусную последовательность или в значительной степени идентичный вариант консенсусной последовательности, можно создать способами, известными в данной области (Rizo and Gierasch Ann. Rev. Biochem. 61: 387 (1992), включенная в данное описание в виде ссылки), например добавлением внутренних остатков цистеина, способных образовывать внутримолекулярные дисульфидные мостики, которые циклизуют пептид. "Иммуноглобулин" является тетрамерной молекулой. В иммуноглобулине природного происхождения каждый тетрамер состоит из двух идентичных пар полипептидных цепей, при этом каждая пара имеет одну "легкую" (примерно 25 кД) и одну "тяжелую" цепь (примерно 50-70 кД). Аминоконцевая часть каждой цепи включает вариабельную область длиной примерно от 100 до 110 или более аминокислот, главным образом, ответственных за узнавание антигена. Часть карбоксильного конца каждой цепи определяет константную область, главным образом, ответственную за эффекторную функцию. Легкие цепи иммуноглобулина человека классифицируют как легкие цепи κ и λ. Тяжелые цепи подразделяют на μ, Δ, γ, α или ε, и изотипы антител определяют как IgM, IgD, IgG, IgA и IgE, соответственно. В легких и тяжелых цепях вариабельные и константные области связаны областью "J" примерно из 12 или большего количества аминокислот, при этом тяжелая цепь также содержит область "D" еще примерно из 10 или более аминокислот. В общих чертах, см. Fundamental Immunology Ch. 7 (Paul, W., ed., 2nd ed. Raven Press, N. Y. (1989)) (включена в виде ссылки в полном объеме для всех целей). Вариабельные области каждой пары легкая/тяжелая цепь образуют связывающий сайт антитела, так что интактный иммуноглобулин имеет два связывающих сайта. Для цепей иммуноглобулинов обнаружена одна и та же общая структура из относительно консервативных каркасных областей (FR), связанных тремя гипервариабельными областями, также называемыми областями, определяющими комплементарность, или CDR. CDR из двух цепей каждой пары сопоставлены посредством каркасных областей, обеспечивая при этом связывание со специфичным эпитопом. От N-конца к С-концу и легкая, и тяжелая цепи содержат домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Распределение аминокислот в каждом домене соответствует определениям Kabat Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987 and 1991)) или Chothia and Lesk J. Mol. Biol. 196: 901-917 (1987); Chothia et al. Nature 342: 878-883 (1989). "Антитело" относится к интактному иммуноглобулину или к его антигенсвязывающей части, которая конкурирует с интактным антителом за специфичное связывание. Антигенсвязывающие части можно получить способами на основе рекомбинантной ДНК или ферментативным или химическим расщеплением интактных антител. Антигенсвязывающие части включают наряду с другими фрагменты Fab, Fab', F(ab') 2 , Fv, dAb и фрагменты гипервариабельной области (CDR), одноцепочечные антитела (scFv), химерные антитела, диантитела и полипептиды, которые содержат по меньшей мере часть иммуноглобулина, которая достаточна для придания полипептиду способности специфично связывать антиген. В используемом в данном описании смысле антитело, которое названо, например, 2.12.1, 2.13.2, 2.14.3, 4.9.2, 4.17.3 и 6.1.1, является антителом, которое получают из гибридомы с таким же названием. Например, антитело 2.12.1 получают из гибридомы 2.12.1. Fab-фрагмент является моновалентным фрагментом, состоящим из доменов VL, VH, CL и СН I; F(ab') 2 -фрагмент является бивалентным фрагментом, содержащим два Fab-фрагмента, связанных дисульфидным мостиком в области шарнира; Fd-фрагмент состоит из доменов VH и СН1; Fv-фрагмент состоит из доменов VL и VH одного плеча антитела; и dAb-фрагмент (Ward et al., Nature 341: 544-546, 1989) состоит из домена VH. Однсцепочечное антитело (scFv) является антителом, в котором области VL и VH спарены, образуя моновалентные молекулы, посредством синтетического линкера, который дает возможность образовать из них одну белковую цепь (Bird et al., Science 242: 423-426, 1988 и Huston et al., Proc. Natl. Acad. Sci. USA 85: 5879-5883, 1988). Диантитела являются бивалентными, биспецифичными антителами, в которых домены VH и VL экспрессированы в виде одной полипептидной цепи, но с использованием линкера, который слишком короткий, чтобы обеспечить спаривание между двумя доменами на одной и той же цепи, тем самым, усиливая спаривание доменов с комплементарными доменами другой цепи и создавая при этом два антигенсвязывающих сайта (см., например, Holliger, P., et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448, 1993 и Poljak, R.J., et al., Structure 2: 1121-1123, 1994). Один или более CDR можно включить в молекулу либо ковалентно, либо нековалентно, чтобы превратить ее в иммуноадгезин. Иммуноадгезин может содержать CDR в виде части большей по размеру полипептидной цепи, может содержать CDR, ковалентно связанные с другой полипептидной цепью, или может содержать CDR, связанный(ые) нековалентно. CDR дают возможность иммуноадгезину специфично связываться с конкретным представляющим интерес антигеном. Антитело может иметь один или более сайтов связывания. В том случае, если имеется более одного сайта связывания, связывающие сайты могут быть идентичными по отношению друг к другу или могут быть разными. Например, иммуноглобулин природного происхождения имеет два идентичных сайта связывания, одноцепочечное антитело или Fab-фрагмент имеет один сайт связывания, тогда как "биспецифичное" или "бифункциональное антитело" имеет два разных сайта связывания. "Выделенное антитело" является антителом, которое: (1) не связано с компонентами, с которыми оно связано в природных условиях, включая другие связанные в природных условиях антитела, которые являются сопутствующими в его нативном состоянии, (2) не содержит других белков из того же вида, (3) экспрессируется клеткой от другого вида или (4) не встречается в природе. Примеры выделенных антител включают анти-IGF-IR-антитело, которое было аффинно очищено с использованием IGF-IR и является выделенным антителом, анти-IGF-IR-антитело, которое было синтезировано гибридомой или другой линией клеток in vitro, и анти-IGF-IR-антитело человека, полученное из трансгенной мыши. Термин "антитело человека" включает все антитела, которые имеют одну или более вариабельных и константных областей, полученных из последовательностей иммуноглобулина человека. В предпочтительном варианте все вариабельные и константные домены получены из последовательностей иммуноглобулина человека (полностью человеческое антитело). Указанные антитела можно получить множеством способов, которые описаны далее. Гуманизированным антителом является антитело, которое получают из вида, отличного от человека, в котором аминокислоты в каркасных областях и константных доменах тяжелой и легкой цепей были мутированы с тем, чтобы избежать или исключить иммунный ответ у людей. Альтернативно гуманизированное антитело можно получить слиянием константных доменов из антитела человека с вариабельными доменами вида, отличного от человека. Примеры того, как получить гуманизированные антитела, можно найти в патентах США № № 6054297, 5886152 и 5877293. Термин "химерное антитело" относится к антителу, которое содержит одну или более областей из одного антитела и одну или более областей из одного или более других антител. В предпочтительном варианте один или более CDR получают из анти-IGF-IR-антитела человека. В более предпочтительном варианте все CDR получают из анти-IGF-IR-антитела человека. В другом предпочтительном варианте объединяют CDR из более чем одного анти-IGF-IR-антитела человека и собирают в химерное антитело. Например, химерное антитело может содержать CDR1 из легкой цепи первого анти-IGF-IR-антитела человека, которое можно объединить с CDR2 и CDR3 из легкой цепи второго анти-IGF-IR-антитела человека, и CDR из тяжелой цепи можно получить из третьего анти-IGF-IR-антитела. Кроме того, каркасные области можно получить из одного из тех же самых анти-IGF-IR-антител, из одного или более других антител, таких как антитело человека, или из гуманизированного антитела. "Нейтрализующим антителом" или "ингибирующим антителом" является антитело, которое ингибирует связывание IGF-IR с IGF-I в том случае, когда избыток анти-IGF-IR-антитела уменьшает количество IGF-I, связанного с IGF-IR, по меньшей мере примерно на 20%. В предпочтительном варианте антитело уменьшает количество IGF-I, связанного с IGF-IR, по меньшей мере примерно на 40%, более предпочтительно на 60%, еще более предпочтительно на 80% или еще более предпочтительно на 85%. Уменьшение связывания можно измерить способами, известными специалисту в данной области, например так, как измеряют при анализе конкурентного связывания in vitro. Пример измерения уменьшения связывания IGF-I с IGF-IR представлен ниже в примере 4. "Активирующим антителом" является антитело, которое активирует IGF-IR по меньшей мере примерно на 20% при добавлении к клетке, ткани или введении в организм, экспрессирующий IGF-IR. В предпочтительном варианте антитело активирует активность IGF-IR по меньшей мере на 40%, более предпочтительно на 60%, еще более предпочтительно на 80% или еще более предпочтительно на 85%. В более предпочтительном варианте активирующее антитело добавляют в присутствии IGF-I или IGF-II. В другом предпочтительном варианте активность активирующего антитела измеряют посредством количественного определения автофосфорилирования тирозина в IGF-IR. Специалисты в данной области легко могут получить фрагменты или аналоги антител, следуя руководствам данного описания. Предпочтительные амино- и карбоксильные концы фрагментов или аналогов находятся вблизи границ функциональных доменов. Структурные и функциональные домены можно идентифицировать путем сравнения данных о нуклеотидной и/или аминокислотной последовательности с опубликованной или собственной базой данных последовательностей. Предпочтительно используют компьютеризованные способы сравнения для идентификации мотивов последовательностей или рассчитанных конформационных доменов белка, которые имеются в других белках известной структуры и/или функции. Способы идентификации белковых последовательностей, которые укладываются в известную трехмерную структуру, известны. Bowie et al. Science 253: 164 (1991). Термин "резонанс поверхностного плазмона" в используемом в данном описании смысле относится к оптическому явлению, которое позволяет анализировать в режиме реального времени биоспецифичные взаимодействия посредством определения изменений концентраций белка в биочувствительной матрице, например, с использованием системы BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden and Piscataway, N.J.). Дополнительное описание см. в Jonsson, U., et al. (1993) Ann. Biol. Clin. 51: 19-26; Jonsson, U., et al. (1991) Biotechniques 11: 620-627; Johnsson, В., et al. (1995) J. Mol. Recognit. 8: 125-131; и Johnson, В., et al. (1991) Anal. Biochem. 198: 268-277. Термин "K off " относится к константе скорости распада при диссоциации антитела из комплекса антитело/антиген. Термин "K d " относится к константе диссоциации конкретного взаимодействия антитело-антиген. Термин "эпитоп" включает любую белковую детерминанту, способную специфично связываться с иммуноглобулином или рецептором Т-клетки. Эпитопные детерминанты обычно состоят из химически активных поверхностных группировок молекул, таких как боковые цепи аминокислот или сахаров, и обычно имеют специфичные параметры трехмерной структуры, а также специфичные характеристики заряда. Говорят, что антитело специфично связывается с антигеном в том случае, когда константа диссоциации ≤1 мкМ, предпочтительно ≤100 нМ и наиболее предпочтительно ≤10 нМ. В используемом в данном описании смысле названия 20 обычных аминокислот и их сокращения не отличаются от традиционного применения. См. Immunology-A Synthesis (2nd Edition, E.S. Golub and D.R. Gren, Eds., Sinauer Associates, Sunderland, Mass. (1991)), которая включена в данное описание в виде ссылки. Стереоизомеры (например, D-аминокислоты) 20 обычных аминокислот, неприродные аминокислоты, такие как α-, α-дизамещенные аминокислоты, N-алкиламинокислоты, молочная кислота и другие необычные аминокислоты, также могут быть подходящими компонентами полипептидов согласно данному изобретению. Примеры необычных аминокислот включают 4-гидроксипролин, γ-карбоксиглутамат, ε-N,N,N-триметиллизин, ε-N-ацетиллизин, О-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, s-N-метиларгинин и другие подобные аминокислоты и иминокислоты (например, 4-гидроксипролин). В используемом в данном описании обозначении полипептида направление влево является направлением аминоконца, а направление вправо является направлением карбоксильного конца в соответствии со стандартным использованием и правилами. Термин "полинуклеотид", указываемый в данном описании, означает полимерную форму нуклеотидов по меньшей мере длиной 10 оснований, либо рибонуклеотидов, либо дезоксирибонуклеотидов, или модифицированной формы любого типа нуклеотида. Термин включает одноцпочечные и двуцепочечные формы ДНК. Термин "выделенный полинуклеотид" в используемом в данном описании смысле будет означать полинуклеотид геномной, кДНК или синтетической природы или их комбинации, и на основании своего происхождения указанный "выделенный полинуклеотид": (1) не связан с полным или с частью полинуклеотида, в котором "выделенный полинуклеотид" выявляют в природе, (2) функционально связан с полинуклеотидом, с которым не связан в природе, или (3) не встречается в природе в виде части последовательности большего размера. Упоминаемый в данном описании термин "олигонуклеотид" включает нуклеотиды природного происхождения и модифицированные нуклеотиды, связанные вместе посредством олигонуклеотидных связей природного происхождения и неприродного происхождения. Олигонуклеотиды являются подгруппой полинуклеотидов, обычно содержащих в длину 200 оснований или меньше. Предпочтительно олигонуклеотиды имеют длину от 10 до 60 оснований и наиболее предпочтительно длину 12, 13, 14, 15, 16, 17, 18, 19 или 20-40 оснований. Олигонуклеотиды обычно являются одноцепочечными, например, для зондов; хотя олигонуклеотиды могут быть двуцепочечными, например, для применения при конструировании мутантного гена. Олигонуклеотиды согласно изобретению могут быть либо смысловыми, либо антисмысловыми олигонуклеотидами. Упоминаемый в данном описании термин "нуклеотиды природного происхождения" включает дезоксирибонуклеотиды и рибонуклеотиды. Упоминаемый в данном описании термин "модифицированные нуклеотиды" включает нуклеотиды с модифицированными или замещенными группами сахаров и т.п. Упоминаемый в данном описании термин "олигонуклеотидные связи" включает такие связи в олигонуклеотидах, как фосфоротиоатные, фосфородитиоатные, фосфороселеноатные, фосфородиселеноатные, фосфороанилотиоатные, фосфороаниладатные, фосфороамидатные и т.п. См., например, LaPlanche et al. Nucl. Acids Res. 14: 9081 (1986); Stec et al. J. Am. Chem. Soc. 106: 6077 (1984); Stein et al. Nucl. Acids Res. 16: 3209 (1988); Zon et al. Anti-Cancer Drug Design 6: 539 (1991); Zon et al. Oligonucleotides and Analogues: A Practical Approach, pp. 87-108 (F. Eckstein, Ed., Oxford University Press, Oxford England (1991)); Stec et al. патент США № 5151510; Uhlmann and Peyman Chemical Reviews 90: 543 (1990), описания которых включены в данное описание в виде ссылки. При необходимости олигонуклеотид может содержать метку для регистрации. "Функционально связанные" последовательности включают как последовательности регуляции экспрессии, которые непосредственно прилегают к представляющему интерес гену, так и последовательности регуляции экспрессии, которые действуют в транс-положении или на расстоянии, регулируя представляющий интерес ген. Термин "последовательность регуляции экспрессии" в используемом в данном описании смысле относится к полинуклеотидным последовательностям, которые необходимы для воздействия на экспрессию и процессинг кодирующих последовательностей, с которыми они связаны. Последовательности регуляции экспрессии включают соответствующие последовательности инициации транскрипции, терминации, промоторные и энхансерные последовательности; эффективные сигналы процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, которые стабилизируют цитоплазматическую мРНК; последовательности, которые усиливают эффективность трансляции (т.е. консенсусная последовательность Козака); последовательности, которые повышают стабильность белка; и, при желании, последовательности, которые усиливают секрецию белка. Природа таких регуляторных последовательностей различна в зависимости от организма хозяина; у прокариот такие регуляторные последовательности обычно включают промотор, сайт связывания рибосомы и последовательность терминации транскрипции; у эукариот, как правило, такие регуляторные последовательности включают промоторы и последовательности терминации транскрипции. Подразумевается, что термин "регуляторные последовательности" включает, как минимум, все компоненты, присутствие которых необходимо для экспрессии и процессинга, а также может включать дополнительные компоненты, присутствие которых является преимущественным, например лидерные последовательности и последовательности партнеров при слиянии. Подразумевается, что в используемом в данном описании смысле термин "вектор" относится к молекуле нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она была связана. Одним типом вектора является "плазмида", которая относится к кольцевой двуцепочечной петле ДНК, в которую можно лигировать дополнительные участки ДНК. Другим типом вектора является вирусный вектор, где дополнительные участки ДНК можно лигировать в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальное начало репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина и поэтому реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы называют в данном описании "рекомбинантными экспрессирующими векторами" (или просто "экспрессирующими векторами"). В общем, экспрессирующие векторы, применяемые в способах на основе рекомбинантной ДНК, часто имеют форму плазмид. В данном описании термины "плазмида" и "вектор" могут использоваться взаимозаменяемо, так как плазмида является наиболее широко используемой формой вектора. Однако подразумевается, что изобретение включает в себя другие формы экомпрессирующих векторов, такие как вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), которые подходят для выполнения эквивалентных функций. Имеется в виду, что термин "рекомбинантная клетка-хозяин" (или просто "клетка-хозяин") в используемом в данном описании смысле относится к клетке, в которую был введен рекомбинантный экспрессирующий вектор. Следует понимать, что имеется в виду, что такие термины относятся не только к конкретной клетке-объекту, но и к потомству такой клетки. Так как в последующих поколениях могут возникать определенные модификации либо вследствие мутации, либо вследствие влияния окружающей среды, в действительности, такое потомство может быть неидентичным родительской клетке, но еще включено в рамки термина "клетка-хозяин" в используемом в данном описании смысле. Термин "селективно гибридизуется", упоминаемый в данном описании, означает регистрируемо и специфично связывается. Полинуклеотиды, олигонуклеотиды и их фрагменты согласно изобретению селективно гибридизуются с цепями нуклеиновой кислоты в условиях гибридизации и промывки, которые минимизируют существенные количества регистрируемого связывания с неспецифичными нуклеиновыми кислотами. Для достижения условий для селективной гибридизации можно использовать условия "высокой жесткости" или "высокожесткие" условия, которые известны в данной области и обсуждаются в данном описании. Примером условий "высокой жесткости" или "высокожестких" условий является способ инкубирования полинуклеотида с другим полинуклеотидом, при этом один полинуклеотид может быть прикреплен к твердой поверхности, такой как мембрана, в буфере для гибридизации 6Х SSPE или SSC, 50% формамиде, 5Х реагенте Денхардта, 0,5% SDS, 100 мкг/мл денатурированной фрагментированной ДНК спермы лосося при температуре гибридизации 42 °С в течение 12-16 ч с последующей двукратной промывкой при 55 °С с использованием буфера для промывки 1X SSC, 0,5% SDS. См. также Sambrook et al., выше, pp. 9.50-9.55. Термин "процент идентичности последовательностей" в контексте последовательностей нуклеиновых кислот относится к остаткам в двух последовательностях, которые являются теми же самыми при сопоставлении в отношении максимального соответствия. Длина сравнения идентичности последовательностей может быть на протяжении участка по меньшей мере примерно из 9 нуклеотидов, обычно по меньшей мере примерно из 18 нуклеотидов, более обычно по меньшей мере примерно из 24 нуклеотидов, типично по меньшей мере примерно из 28 нуклеотидов, более обычно по меньшей мере примерно из 32 нуклеотидов и предпочтительно по меньшей мере примерно из 36, 48 или большего количества нуклеотидов. В данной области известен ряд различных алгоритмов, которые можно использовать для измерения идентичности нуклеотидных последовательностей. Например, полинуклеотидные последовательности можно сравнивать с использованием FASTA, Gap или Bestfit, которые представляют собой программы в пакете Wisconsin, версия 10.0, Genetics Computer Group (GCG), Madison, Wisconsin. FASTA, которая включает, например, программы FASTA2 и FASTA3, обеспечивает сопоставление и процент идентичности последовательностей в областях наилучшего перекрывания между запрашиваемой и исследуемой последовательностями (Pearson, Methods Enzymol. 183: 63-98 (1990); Pearson, Methods Mol. Biol. 132: 185-219 (2000); Pearson, Methods Enzymol. 266: 227-258 (1996); Pearson, J. Mol. Biol. 276: 71-84 (1998); включенные в данное описание в виде ссылки). Если не оговорено особо, для конкретной программы или алгоритма используют параметры по умолчанию. Например, идентичность последовательностей в процентах между последовательностями нуклеиновых кислот можно определить, используя FASTA с его параметрами по умолчанию (размер слова 6 и фактор NOPAM для матрицы подсчета очков) или используя Gap с его параметрами по умолчанию, которые даны в версии 6.1 GCG, включенной в данное описание в виде ссылки. Упоминание последовательности нуклеиновой кислоты охватывает ее комплемент, если не оговорено особо. Таким образом, следует понимать, что ссылка на молекулу нуклеиновой кислоты, имеющую конкретную последовательность, охватывает комплементарную ей цепь с комплементарной последовательностью. В области молекулярной биологии исследователи используют термины "процент идентичности последовательностей", "процент сходства последовательностей" и "процент гомологии последовательностей" взаимозаменяемо. В данной заявке указанные термины будут иметь одинаковые значения только по отношению к последовательностям нуклеиновых кислот. Термин "значительное сходство" или "значительное сходство последовательностей" по отношению к нуклеиновой кислоте или ее фрагменту свидетельствует о том, что при оптимальном сопоставлении с соответствующими инсерциями или делециями нуклеотидов в другой нуклеиновой кислоте (или комплементарной ей цепи) идентичность последовательности нуклеотидов составляет по меньшей мере примерно 85%, предпочтительно по меньшей мере примерно 90% и более предпочтительно по меньшей мере примерно 95, 96, 97, 98 или 99% оснований нуклеотидов, что измеряют с помощью любого хорошо известного алгоритма идентичности последовательностей, такого как FASTA, BLAST или Gap, которые обсуждаются выше. В применении к полипептидам термин "значительная идентичность" означает, что две пептидные последовательности при оптимальном сопоставлении, таком как с помощью программ GAP или BESTFIT, с использованием параметров пробелов по умолчанию, обладают по меньшей мере 75 или 80% идентичностью по последовательностям, предпочтительно по меньшей мере 90 или 95% идентичностью по последовательностям, еще более предпочтительно по меньшей мере 98 или 99% идентичностью по последовательностям. Предпочтительно положения остатков, которые не являются идентичными, отличаются консервативными заменами аминокислот. "Консервативной заменой аминокислот" является замена, при которой аминокислотный остаток заменяют другим аминокислотным остатком, имеющим боковую цепь (R-группу) со сходными химическими свойствами (например, заряд или гидрофобность). В общем, консервативная аминокислотная замена не будет в значительной степени изменять функциональные свойства белка. В тех случаях, когда две или большее количество аминокислотных последовательностей отличаются друг от друга консервативными заменами, процент идентичности последовательностей или степень сходства можно скорректировать как более высокую, введя поправку на консервативную природу замены. Способы осуществления такой корректировки хорошо известны специалистам в данной области. См., например, Pearson, Methods Mol. Biol. 24: 307-31 (1994), которая включена в данное описание в виде ссылки. Примеры групп аминокислот, которые имеют боковые цепи со сходными химическими свойствами, включают аминокислоты: 1) с алифатическими боковыми цепями: глицин, аланин, валин, лейцин и изолейцин; 2) с алифатическими содержащими гидроксил боковыми цепями: серин и треонин; 3) с амидосодержащими боковыми цепями: аспарагин и глутамин; 4) с ароматическими боковыми цепями: фенилаланин, тирозин и триптофан; 5) с основными боковыми цепями: лизин, аргинин и гистидин; и 6) с серосодержащими боковыми цепями: цистеин и метионин. Предпочтительными группами консервативных аминокислотных замен являются валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин, глутамат-аспартат и аспарагин-глутамин. Альтернативно, консервативной заменой является любое изменение, имеющее положительное значение в матрице логарифмической функции правдоподобия РАМ250, описанной в работе Gonnet et al., Science 256: 1443-45 (1992), включенной в данное описание в виде ссылки. "Умеренно консервативной" заменой является любое изменение, имеющее неотрицательное значение в матрице логарифмической функции правдоподобия РАМ250. Сходство последовательностей в случае полипептидов, которое также называют идентичностью последовательностей, обычно измеряют с использованием компьютерной программы анализа последовательностей. Компьютерная программа анализа белков подбирает сходные последовательности на основе использования критериев сходства, приписываемых различным заменам, делециям и другим модификациям, включая консервативные аминокислотные замены. Например, GCG содержит такие программы, как "Gap" и "Bestfit", которые можно использовать с параметрами по умолчанию для определения гомологии последовательностей или идентичности последовательностей между близкородственными полипептидами, такими как гомологичные полипептиды из разных видов организмов, или между белком дикого типа и его мутеином. См., например GCG, версия 6.1. Полипептидные последовательности также можно сравнивать с применением FASTA, используя параметры по умолчанию или рекомендуемые параметры, программа в GCG версии 6.1. FASTA (например, FASTA2 и FASTA3) осуществляет сопоставления и дает процент идентичности последовательностей областей наилучшего перекрывания между запрашиваемой и исследуемой последовательностями (Pearson (1990); Pearson (2000)). Другим предпочтительным алгоритмом при сравнении последовательности согласно изобретению с базой данных, содержащей большое количество последовательностей из разных организмов, является компьютерная программа BLAST, особенно blastp или tblastn, с использованием параметров по умолчанию. См., например, Altschul et al., J. Mol. Biol. 215: 403-410 (1990); Altschul et al., Nucleic Acids Res. 25: 3389-402 (1997); включенные в данное описание в виде ссылки. Длина сравниваемых в отношении гомологии полипептидных последовательностей, как правило, будет составлять по меньшей мере примерно 16 аминокислотных остатков, обычно по меньшей мере примерно 20 остатков, более обычно по меньшей мере примерно 24 остатка, обычно по меньшей мере 28 остатков и предпочтительно примерно более 35 остатков. В случае поиска в базе данных, содержащей последовательности из большого числа различных организмов, предпочтительно сравнивать аминокислотные последовательности. В используемом в данном описании смысле термин "метка" или "меченый" относится к включению другой молекулы в антитело. В одном варианте меткой является регистрируемый маркер, например включение радиоактивно меченной аминокислоты или присоединение к полипептиду фрагментов биотинила, которые можно регистрировать маркированным авидином (например, стрептавидин, содержащий флуоресцентный маркер или ферментативную активность, которую можно регистрировать оптическими или колориметрическими способами). В другом варианте меткой или маркером может быть терапевтическое средство, например конъюгат лекарственного средства или токсин. Различные способы мечения полипептидов и гликопротеидов известны в данной области и могут быть использованы. Примеры меток полипептидов включают, но не ограничены указанным, следующие метки: радиоизотопы или радионуклиды (например, 3 Н, 14 С, 15 N, 35 S, 90 Y, 99 Тс, 111 In, 125 I, 131 I), флуоресцентные метки (например, ФИТЦ, родамин, люминофоры на основе лантанидов), ферментные метки (например, пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза), хемилюминесцентные маркеры, группы биотинила, предварительно определенные полипептидные эпитопы, узнаваемые вторичным репортером (например, парные последовательности лейциновой молнии, сайты связывания вторичных антител, домены, связывающие металлы, эпитопные метки), магнитные агенты, такие как хелаты гадолиния, токсины, такие как токсин коклюша, таксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, и пуромицин, и их аналоги или гомологи. В некоторых вариантах метки присоединяют посредством спейсерного плеча различной длины, чтобы уменьшить возможные стерические помехи. Термин "агент" в данном описании используют для обозначения химического соединения, смеси химических соединений, биологической макромолекулы или экстракта, полученного из биологических материалов. Термин "фармацевтический агент или лекарственное средство" в данном описании используют по отношению к химическому соединению или композиции, способной индуцировать требуемое терапевтическое действие при правильном введении пациенту. Другие химические термины в данном описании используют согласно традиционному применению в данной области, примеры которого приведены в The McGraw-Hill Dictionary of Chemical Terms (Parker, S., Ed., McGraw-Hill, San Francisco (1985)), включенном в данное описание в виде ссылки). Термин "антинеопластическое средство" в данном описании используют по отношению к агентам, которые обладают функциональным свойством ингибировать развитие или прогрессирование неоплазмы у человека, особенно злокачественного (канцерогенного) повреждения, такого как карцинома, саркома, лимфома или лейкоз. Часто свойством антинеопластических средств является ингибирование метастазов. Термин "пациент" включает человека и субъекты ветеринарии. Антитела человека позволяют избежать некоторые проблемы, связанные с антителами, которые имеют вариабельные и/или константные области мышей или крыс. Присутствие таких полученных от мышей или крыс последовательностей может приводить к быстрому клиренсу антител или может приводить к возникновению у пациента иммунного ответа против антитела. Таким образом, в одном варианте изобретение относится к гуманизированным анти-IGF-IR-антителам. В предпочтительном варианте изобретение относится к полностью человеческим анти-IGF-IR-антителам, полученным благодаря введению генов иммуноглобулинов человека в организм грызуна, так чтобы у грызуна продуцировались полностью человеческие антитела. Более предпочтительны полностью человеческие антитела против IGF-IR человека. Предполагается, что полностью человеческие анти-IGF-IR-антитела минимизируют иммуногенные и аллергенные ответы, свойственные мышиным или полученным из мышей моноклональным антителам (мАт), и таким образом повышают эффективность и безопасность вводимых антител. Можно ожидать, что применение полностью человеческих антител обеспечит значительное преимущество при лечение хронических и рецидивирующих заболеваний у человека, таких как воспаление и рак, при которых могут требоваться многократные введения антител. В другом варианте изобретение относится к анти-IGF-IR-антителу, которое не связывается с комплементом. В предпочтительном варианте анти-IGF-IR-антителом является 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте анти-IGF-IR-антитело включает легкую цепь, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 2, 6, 10, 14, 18 или 22, или один или более CDR из указанных аминокислотных последовательностей. В другом предпочтительном варианте анти-IGF-IR-антитело включает тяжелую цепь, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 4, 8, 12, 16, 20 или 24, или один или более CDR из указанных аминокислотных последовательностей. Антитело может представлять собой молекулу IgG, IgM, IgE, IgA или IgD. В предпочтительном варианте антитело представляет собой IgG и относится к подтипу IgG1, IgG2, IgG3 или IgG4. В более предпочтительном варианте анти-IGF-IR-антитело представлено подклассом IgG2. В другом предпочтительном варианте анти-IGF-IR-антитело относится к тому же классу, что и антитело 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1, которые представляют собой IgG2. Класс и подкласс анти-IGF-IR-антител можно определить любым способом, известным в данной области. В общем, класс и подкласс антитела можно определить с использованием антител, которые специфичны по отношению к конкретному классу и подклассу антитела. Такие антитела коммерчески доступны. Класс и подкласс можно определить посредством ELISA, Вестерн-блота, а также других способов. Альтернативно, класс и подкласс можно определить секвенированием всех или части константных доменов тяжелой и/или легкой цепей антител, сравнением их аминокислотных последовательностей с известными аминокислотными последовательностями иммуноглобулинов различных классов и подклассов и определением класса и подкласса антител. В другом аспекте изобретения анти-IGF-IR-антитело проявляет видовую и молекулярную избирательность. В одном варианте анти-IGF-IR-антитело связывается с IGF-IR человека, макак-крабоедов и макак-резус. В предпочтительном варианте анти-IGF-IR-антитело не связывается с IGF-IR мыши, крысы, морской свинки, собаки или кролика. В другом предпочтительном варианте анти-IGF-IR-антитело не связывается с рецепторами вида обезьян Нового Света, такого как мартышка. Следуя инструкциям описания, можно определить видовую избирательность анти-IGF-IR-антитела с использованием способов, хорошо известных в данной области. Например, можно определить видовую избирательность, используя Вестерн-блот, FACS, ELISA или РИА. В предпочтительном варианте видовую избирательность можно определить с использованием Вестерн-блота. В другом варианте анти-IGF-IR-антитело обладает избирательностью по отношению к IGF-IR, которая по меньшей мере в 50 раз выше, чем избирательность по отношению к рецептору инсулина. В предпочтительном варианте избирательность анти-IGF-IR-антитела более чем в 100 раз выше, чем его избирательность по отношению к рецептору инсулина. В еще более предпочтительном варианте анти-IGF-IR-антитело не проявляет какого-либо заметного специфичного связывания с любым другим белком, отличным от IGF-IR. Избирательность анти-IGF-IR-антитела по отношению IGF-IR можно определить с использованием способов, хорошо известных в данной области, следуя инструкциям описания. Например, избирательность можно определить с использованием Вестерн-блота, FACS, ELISA или РИА. В предпочтительном варианте молекулярную избирательность можно определить с использованием Вестерн-блота. В другом аспекте изобретения анти-IGF-IR-антитела связываются с IGF-IR с высокой аффинностью. В одном варианте анти-IGF-IR-антитело связывается с IGF-IR с K d , равной 1 ×10 -8 М или меньше. В более предпочтительном варианте антитело связывается с IGF-IR с K d либо 1 ×10 -9 М, либо меньше. В еще более предпочтительном варианте антитело связывается с IGF-IR с K d либо 5 ×10 -10 М, либо меньше. В другом предпочтительном варианте антитело связывается с IGF-IR с K d либо 1 ×10 -10 М, либо меньше. В другом предпочтительном варианте антитело связывается с IGF-IR, по существу, с такой же K d , как и антитело, выбранное из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте антитело связывается с IGF-IR, по существу, с такой же K d , как и антитело, которое содержит один или более CDR из антитела, выбранного из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. Еще в одном предпочтительном варианте антитело связывается с IGF-IR, по существу, с такой же K d , как антитело, которое содержит одну из аминокислотных последовательностей, выбранных из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 или 24. В другом предпочтительном варианте антитело связывается с IGF-IR, по существу, с такой же K d , как антитело, которое содержит один или более CDR из антитела, которое содержит одну из аминокислотных последовательностей выбранных из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 или 24. В другом аспекте изобретения анти-IGF-IR-антитело имеет низкую скорость диссоциации. В одном варианте анти-IGF-IR-антитело имеет K off , равную 1 ×10 -4 с -1 или ниже. В предпочтительном варианте K off составляет 5 ×10 -5 с -1 или ниже. В другом предпочтительном варианте K off , по существу, имеет такое же значение, как и в случае антитела, выбранного из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте антитело связывается с IGF-IR, по существу, с такой же K off , как и антитело, которое содержит один или более CDR из антитела, выбранного из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. Еще в одном предпочтительном варианте антитело связывается с IGF-IR, по существу, с такой же K off , как и антитело, которое содержит одну из аминокислотных последовательностей, выбранных из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 или 24. В другом предпочтительном варианте антитело связывается с IGF-IR, по существу, с такой же K off , как и антитело, которое содержит один или более CDR из антитела, которое содержит одну из аминокислотных последовательностей, выбранных из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22. Аффинность связывания и скорость диссоциации анти-IGF-IR-антитела с IGF-IR можно определить любым способом, известным в данной области. В одном варианте аффинность связывания можно измерить с помощью конкурентного ELISA, РИА или резонанса поверхностного плазмона, такого как BIAcore. Скорость диссоциации также можно измерить с помощью резонанса поверхностного плазмона. В более предпочтительном варианте аффинность связывания и скорость диссоциации измеряют с помощью резонанса поверхностного плазмона. В еще более предпочтительном варианте аффинность связывания и скорость диссоциации измеряют с использованием BIAcore. Пример определения аффинности связывания и скорости диссоциации описан ниже в примере 2. Согласно другой цели изобретения анти-IGF-IR-антитело имеет время полужизни, составляющее in vitro или in vivo по меньшей мере 1 день. В предпочтительном варианте антитело или его часть имеет время полужизни, составляющее по меньшей мере 3 дня. В более предпочтительном варианте антитело или его часть имеет время полужизни, составляющее 4 дня или больше. В другом варианте антитело или его часть имеет время полужизни, составляющее 8 дней или больше. В другом варианте антитело или его антигенсвязывающая часть дериватизированы или модифицированы так, что имеют более длительное время полужизни, как обсуждается ниже. В другом предпочтительном варианте антитело может содержать точечные мутации, чтобы увеличить время полужизни в сыворотке, так, как описано в заявке WO 00/09560, опубликованной 24 февраля 2000г. Время полужизни антитела можно измерить любыми способами, известными специалисту в данной области. Например, время полужизни антитела можно измерить с помощью Вестерн-блота, ELISA или РИА на протяжении соответствующего периода времени. Время полужизни антитела можно измерить у любого подходящего животного, например обезьяны, такой как макака-крабоед, примата или человека. Изобретение также относится к анти-IGF-IR-антителу, которое связывается с тем же самым антигеном или эпитопом, что и анти-IGF-IR-антитело человека. Кроме того, изобретение относится к анти-IGF-IR-антителу, которое перекрестно конкурирует с анти-IGF-IR-антителом человека. В предпочтительном варианте анти-IGF-IR-антителом человека является 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте анти-IGF-IR человека содержит один или более CDR из антитела, выбранного из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. Еще в одном предпочтительном варианте анти-IGF-IR человека содержит одну из аминокислотных последовательностей, выбранных из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 или 24. В другом предпочтительном варианте анти-IGF-IR человека содержит один или более CDR из антитела, которое содержит одну из аминокислотных последовательностей, выбранных из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 или 24. В высокопредпочтительном варианте анти-IGF-IR-антителом является другое антитело человека. Определить, связывается ли анти-IGF-IR-антитело с тем же самым антигеном, можно с использованием ряда способов, известных в данной области. Например, можно определить, связывается ли тестируемое анти-IGF-IR-антитело с тем же самым антигеном, используя анти-IGF-IR-антитело для захвата антигена, о котором известно, что он связывается с анти-IGF-IR-антителом, такого как IGF-IR, элюируя антиген из связи с антителом и затем определяя, будет ли тестируемое антитело связываться с элюированным антигеном. Связывается ли антитело с тем же самым эпитопом, что и анти-IGF-IR-антитело, можно определить, связывая анти-IGF-IR-антитело с IGF-IR в условиях насыщения и затем измеряя способность тестируемого антитела связываться с IGF-IR. Если тестируемое антитело способно связываться с IGF-IR одновременно с анти-IGF-IR-антителом, то тестируемое антитело связывается с другим эпитопом, отличным от эпитопа для анти-IGF-IR-антитела. Однако если тестируемое антитело не способно одновременно связываться с IGF-IR, то тестируемое антитело связывается с тем же самым эпитопом, что и анти-IGF-IR-антитело человека. Указанный эксперимент можно выполнить с использованием ELISA, РИА или резонанса поверхностного плазмона. В предпочтительном варианте эксперимент проводят с использованием резонанса поверхностного плазмона. В более предпочтительном варианте используют BIAcore. Также можно определить, конкурирует ли анти-IGF-IR-антитело перекрестно с анти-IGF-IR-антителом. В предпочтительном варианте можно определить, конкурирует ли анти-IGF-IR-антитело перекрестно с другим антителом, используя тот же способ, который используют для измерения того, способно ли анти-IGF-IR-антитело связываться с тем же самым эпитопом, что и другое анти-IGF-IR-антитело. Изобретение также относится к анти-IGF-IR-антителу, которое содержит вариабельные последовательности, кодируемые геном κ человека. В предпочтительном варианте вариабельные последовательности кодируются семейством генов либо V κ А27, А30, либо 012. В предпочтительном варианте вариабельные последовательности кодируются семейством генов V κ А30 человека. В более предпочтительном варианте легкая цепь содержит не более 10 аминокислотных замен по сравнению с V κ A27, А30 или 012 зародышевой линии, предпочтительно не более 6 аминокислотных замен и более предпочтительно не более 3 аминокислотных замен. В предпочтительном варианте аминокислотные замены являются консервативными заменами. SEQ ID NO: 2, 6, 10, 14, 18 и 22 определяют аминокислотные последовательности вариабельных областей 6 легких κ-цепей анти-IGF-IR. SEQ ID NO: 38, 40 и 42 определяют аминокислотные последовательности 3 легких κ-цепей зародышевой линии, из которых получены 6 легких κ-цепей анти-IGF-IR. На фиг. 1А-1С показаны сопоставления нуклеотидных последовательностей вариабельных областей легких цепей 6 анти-IGF-IR-антител друг с другом и с последовательностями зародышевой линии, из которых они получены. Следуя инструкциям данного описания, специалист в данной области сможет определить кодируемую аминокислотную последовательность 6 легких κ-цепей анти-IGF-IR и легких κ-цепей зародышевой линии и определить различия между последовательностями зародышевой линии и последовательностями антител. В предпочтительном варианте VL анти-IGF-IR-антитела содержит такие же аминокислотные замены относительно аминокислотной последовательности зародышевой линии, что и любая одна или более VL антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. Например, VL анти-IGF-IR-антитела может содержать одну или более аминокислотных замен, которые являются такими же, как замены, присутствующие в антителе 2.13.2, другую аминокислотную замену, которая является такой же, как замена, присутствующая в антителе 2.14.3, и другую аминокислотную замену, которая является такой же, как в антителе 4.9.2. Таким образом, можно сочетать и подбирать различные характеристики связывания антител для того, чтобы изменить, например, аффинность антитела в отношении IGF-IR или скорость его диссоциации из комплекса с антигеном. В другом варианте аминокислотные замены осуществляют в таком же положении, как и положения, обнаруживаемые в любом одной или более VL антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1, но не используют ту же самую аминокислоту, а осуществляют консервативные аминокислотные замены. Например, если аминокислотная замена по сравнению с зародышевой линией в одном из антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1 представляет собой глутамат, его можно консервативно заменить аспартатом. Подобным образом, если аминокислотной заменой является серин, его можно консервативно заменить треонином. В другом предпочтительном варианте легкая цепь содержит аминокислотную последовательность, которая является такой же, как аминокислотная последовательность VL 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом высокопредпочтительном варианте легкая цепь содержит аминокислотные последовательности, которые являются такими же, как CDR-области легкой цепи 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте легкая цепь содержит аминокислотную последовательность по меньшей мере из одной CDR-области легкой цепи 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте легкая цепь содержит аминокислотные последовательности CDR из разных легких цепей. В более предпочтительном варианте CDR из разных легких цепей получают из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте легкая цепь содержит аминокислотную последовательность, выбранную из SEQ ID NO: 2, 6, 10, 14, 18 или 22. В другом варианте легкая цепь содержит аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, выбранной из SEQ ID NO: 1, 5, 9, 13, 17 или 21, или последовательностью нуклеиновой кислоты, которая кодирует аминокислотную последовательность, имеющую 1-10 инсерций, делеций или замен аминокислот. Предпочтительно аминокислотные замены являются консервативными аминокислотными заменами. В другом варианте антитело или его часть содержит легкую цепь лямбда. Данное изобретение также относится к анти-IGF-IR-антителу или его части, которое содержит тяжелую цепь человека или последовательность, полученную из тяжелой цепи человека. В одном варианте аминокислотную последовательность тяжелой цепи получают из семейства генов V H DP-35, DP-47, DP-70, DP-71 или VIV-4/4.35 человека. В предпочтительном варианте аминокислотную последовательность тяжелой цепи получают из семейства генов V H DP-47 человека. В более предпочтительном варианте тяжелая цепь содержит не более 8 изменений аминокислот по сравнению с V H DP-35, DP-47, DP-70, DP-71 или VIV-4/4.35 зародышевой линии, более предпочтительно не более 6 изменений аминокислот и еще более предпочтительно не более 3 изменений аминокислот. SEQ ID NO: 4, 8, 12, 16, 20 и 24 представляют аминокислотные последовательности вариабельных областей 6 тяжелых цепей анти-IGF-IR. SEQ ID NO: 30, 32, 34, 36 и 44 представляют аминокислотные последовательности, a SEQ ID NO: 29, 31, 33, 35 и 43, соответственно, представляют нуклеотидные последовательности тяжелых цепей DP-35, DP-47, DP-70, DP-71 и VIV-4, соответственно, зародышевой линии. На фиг. 2A-2D показаны сопоставления аминокислотных последовательностей вариабельной области 6 анти-IGF-IR-антител с соответствующими им последовательностями зародышевой линии. Следуя инструкциям данного описания, специалист в данной области может определить кодируемую аминокислотную последовательность 6 тяжелых цепей анти-IGF-IR и тяжелых цепей зародышевой линии и определить различия между последовательностями зародышевой линии и последовательностями антител. В предпочтительном варианте VH анти-IGF-IR-антитела содержит такие же аминокислотные замены по сравнению аминокислотной последовательностью зародышевой линии, как и любая одна или более VH антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. Подобно тому, как обсуждалось выше, VH анти-IGF-IR-антитела может содержать одну или более аминокислотных замен, которые являются такими же, как замены, присутствующие в антителе 2.13.2, другую аминокислотную замену, которая является такой же, как замена, присутствующая в антителе 2.14.3, и другую аминокислотную замену, которая является такой же, как в антителе 4.9.2. Таким образом, можно сочетать и подбирать различные характеристики связывания антител для того, чтобы изменить, например, аффинность антитела в отношении IGF-IR или скорость его диссоциации из комплекса с антигеном. В другом варианте аминокислотные замены осуществляют в таком же положении, как и положения, обнаруживаемые в любой одной или более VH антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1, но не используют ту же самую аминокислоту, а осуществляют консервативные аминокислотные замены. В другом предпочтительном варианте тяжелая цепь содержит аминокислотную последовательность, которая является такой же, как аминокислотная последовательность VH 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом высокопредпочтительном варианте тяжелая цепь содержит аминокислотные последовательности, которые являются такими же, как CDR-области легкой цепи 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте тяжелая цепь содержит аминокислотную последовательность по меньшей мере из одной CDR-области тяжелой цепи 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте тяжелая цепь содержит аминокислотные последовательности из CDR из разных тяжелых цепей. В более предпочтительном варианте CDR из разных тяжелых цепей получают из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте тяжелая цепь содержит аминокислотную последовательность, выбранную из SEQ ID NO: 4, 8, 12, 16, 20 или 24. В другом варианте тяжелая цепь содержит аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, выбранной из SEQ ID NO: 3, 7, 11, 15, 19 или 23, или последовательностью нуклеиновой кислоты, которая кодирует аминокислотную последовательность, имеющую 1-10 инсерций, делеций или замен аминокислот. В другом варианте замены являются консервативными аминокислотными заменами. В другом варианте изобретение относится к анти-IGF-IR-антителу, которое ингибирует связывание IGF-I с IGF-IR или связывание IGF-II с IGF-IR. В предпочтительном варианте IGF-IR относится к человеку. В другом предпочтительном варианте анти-IGF-IR-антитело является антителом человека. В другом варианте антитело или его часть ингибирует связывание между IGF-IR и IGF-I с IC 50 , не превышающим 100 нМ. В предпочтительном варианте IC 50 составляет не более 10 нМ. В более предпочтительном варианте IC 50 составляет не более 5 нМ. IC 50 можно измерить любым способом, известным в данной области. Обычно IC 50 можно измерить с помощью ELISA или РИА. В предпочтительном варианте IC 50 измеряют с помощью РИА. В другом варианте изобретение относится к анти-IGF-IR-антителу, которое предотвращает активацию IGF-IR в присутствии IGF-I. В предпочтительном варианте анти-IGF-IR-антитело ингибирует индуцируемое IGF-IR фосфорилирование тирозина, которое происходит после того, как рецептор становится занятым. В другом предпочтительном варианте анти-IGF-IR-антитело ингибирует последующие события в клетке, после того как это происходит. Например, анти-IGF-IR может ингибировать фосфорилирование тирозина Shc и субстрата рецептора инсулина (IRS) 1 и 2, которые все в норме фосфорилируются в том случае, когда клетки обрабатывают IGF-I (Kim et al., J. Biol. Chem. 273: 34543-34550, 1998). Выявить, может ли анти-IGF-IR-антитело предотвращать активацию IGF-IR в присутствии IGF-1, можно посредством определения уровней автофосфорилирования IGF-IR, Shc, IRS-1 или IRS-2 с помощью Вестерн-блоттинга или иммунопреципитации. В предпочтительном варианте уровни автофосфорилирования IGF-IR могут быть определены с помощью Вестерн-блоттинга. См., например, пример 7. В другом аспекте изобретения антитело вызывает понижающую регуляцию IGF-IR в случае клетки, обработанной антителом. В одном варианте IGF-IR интернализуется в цитоплазму клетки. После того, как анти-IGF-IR-антитело связывается с IGF-IR, антитело интернализуется, что показано с помощью конфокальной микроскопии. Не имея намерения связывать с какой-либо теорией, предполагается, что комплекс антитело-IGF-IR интернализуется в лизосому и разрушается. Понижающую регуляцию IGF-IR можно измерить любым способом, известным в данной области, включая иммунопреципитацию, конфокальную микроскопию или Вестерн-блоттинг. См., например, пример 7. В предпочтительном варианте антитело выбрано из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 или 6.1.1 или содержит их тяжелую цепь, легкую цепь или антигенсвязывающую область. Другой аспект данного изобретения включает активирующие анти-IGF-IR-антитела. Активирующее антитело отличается от ингибируюшего антитела, поскольку оно усиливает или заменяет действие IGF-I на IGF-IR. В одном варианте активирующее антитело способно связываться с IGF-IR и вызывать его активацию в отсутствие IGF-I. Указанный тип активирующего антитела, по существу, имитирует IGF-I. В другом варианте активирующее антитело усиливает действие IGF-I на IGF-IR. Указанный тип антитела не активирует IGF-IR сам по себе, а повышает активацию IGF-IR в присутствии IGF-I. Имитирующее анти-IGF-IR-антитело можно легко отличить от усиливающего анти-IGF-IR-антитела с помощью обработки клеток in vitro антителом в присутствии или в отсутствие низких уровней IGF-I. Если антитело способно вызывать активацию IGF-IR в отсутствие IGF-I, например оно увеличивает фосфорилирование тирозина IGF-IR, то антитело является имитирующим антителом. Если антитело не может вызывать активацию IGF-IR в отсутствие IGF-I, но способно количественно усиливать активацию IGF-IR, то антитело является усиливающим антителом. В предпочтительном варианте активирующим антителом является 4.17.3. В другом предпочтительном варианте антитело содержит один или более CDR из 4.17.3. В другом предпочтительном варианте антитело получают либо из одной из двух, либо из обеих последовательностей зародышевой линии 012 (легкая цепь) и/или D71 (тяжелая цепь). Другой вариант изобретения относится к анти-IGF-IR-антителу, которое ингибирует фосфорилирование тирозина IGF-IR и уровни рецептора in vivo. В одном варианте введение анти-IGF-IR-антитела животному вызывает снижение сигнала фосфотирозина IGF-IR в опухолях, экспрессирующих IGF-IR. В предпочтительном варианте анти-IGF-IR-антитело вызывает снижение сигнала фосфотирозина по меньшей мере на 20%. В более предпочтительном варианте анти-IGF-IR-антитело вызывает уменьшение сигнала фосфотирозина по меньшей мере на 60%, более предпочтительно на 50%. В еще более предпочтительном варианте антитело вызывает уменьшение сигнала фосфотирозина по меньшей мере на 40%, более предпочтительно на 30%, еще более предпочтительно на 20%. В предпочтительном варианте антитело вводят примерно за 24 ч до измерения уровней фосфорилирования тирозина. Уровни фосфорилирования тирозина можно измерить любым способом, известным в данной области, таким как способы, описанные ниже. См., например, пример 3 и фиг. 5. В предпочтительном варианте антитело выбрано из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 или 6.1.1 или содержит его тяжелую цепь, легкую цепь или антигенсвязывающую часть. В другом варианте введение анти-IGF-IR-антитела животному вызывает снижение уровней IGF-IR в опухолях, экспрессирующих IGF-IR. В предпочтительном варианте анти-IGF-IR-антитело вызывает снижение уровней рецептора по меньшей мере на 20% по сравнению с необработанным животным. В более предпочтительном варианте анти-IGF-IR-антитело вызывает снижение уровней рецептора по меньшей мере на 60%, более предпочтительно на 50% от уровней рецептора у животных, не подвергнутых лечению. В еще более предпочтительном варианте антитело вызывает уменьшение уровней рецептора по меньшей мере на 40%, более предпочтительно на 30%. В предпочтительном варианте антитело вводят примерно за 24 ч до измерения уровней IGF-IR. Уровни IGF-IR можно измерить любым способом, известным в данной области, таким как способы, описанные ниже. См., например, пример 8 и фиг. 6. В предпочтительном варианте антитело выбрано из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 или 6.1.1 или содержит его тяжелую цепь, легкую цепь или антигенсвязывающую часть. В другом варианте анти-IGF-IR-антитело ингибирует рост опухолевых клеток in vivo. Опухолевую клетку можно получить из любого типа клеток, включая без ограничения эпидермальные, эпителиальные, эндотелиальные клетки, клетки лейкоза, саркомы, множественной миеломы или мезодермальные клетки. Примеры опухолевых клеток включают клетки А549 (немелкоклеточная карцинома легкого), клетки MCF-7, клетки Colo 205, клетки 3T3/IGF-IR и клетки А431. В предпочтительном варианте антитело ингибирует рост опухолевых клеток по сравнению с ростом опухоли у животного, не подвергшегося лечению. В более предпочтительном варианте антитело ингибирует рост опухолевых клеток на 50%. В еще более предпочтительном варианте антитело ингибирует рост опухолевых клеток на 60, 65, 70 или 75%. В одном варианте ингибирование роста опухолевых клеток измеряют по меньшей мере через 7 дней после того, как животным начинают проводить лечение антителом. В более предпочтительном варианте ингибирование роста опухолевых клеток измеряют по меньшей мере через 14 дней после того, как животным начинают проводить лечение антителом. В другом предпочтительном варианте с анти-IGF-IR-антителом животному вводят другое антинеопластическое средство. В предпочтительном варианте антинеопластическое средство способно дополнительно ингибировать рост опухолевых клеток. В еще более предпочтительном варианте антинеопластическим средством является адриамицин, таксол, тамоксифен, 5-фтордезоксиуридин (5-FU) или СР-358774. В предпочтительном варианте совместное введение антинеопластического средства и анти-IGF-IR-антитела ингибирует рост опухолевых клеток по меньшей мере на 50%, более предпочтительно на 60, 65, 70 или 75%, более предпочтительно на 80, 85 или 90% через промежуток, равный 22-24 дням. См., например, фиг. 7 и пример 9. В предпочтительном варианте антитело выбрано из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 или 6.1.1 или содержит его тяжелую цепь, легкую цепь или антигенсвязывающую часть. Другой аспект изобретения относится к анти-IGF-IR-антителу, которое индуцирует гибель клеток. В одном варианте антитело вызывает апоптоз. Антитело может индуцировать апоптоз либо in vivo, либо in vitro. В общем, опухолевые клетки являются более чувствительными к апоптозу, чем нормальные клетки, так что введение анти-IGF-IR-антитела предпочтительно вызывает апоптоз опухолевой клетки по сравнению с нормальной клеткой. В другом варианте введение анти-IGF-IR-антитела уменьшает уровни фермента akt, который вовлечен в фосфатидилинозитол (PI)-киназный путь. PI-киназный путь, в свою очередь, вовлечен в пролиферацию клеток и предотвращение апоптоза. Таким образом, ингибирование akt может вызвать апоптоз. В более предпочтительном варианте антитело вводят in vivo, чтобы вызвать апоптоз IGF-IR-экспрессирующей клетки. В предпочтительном варианте антитело выбрано из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 или 6.1.1 или содержит его тяжелую цепь, легкую цепь или антигенсвязывающую часть. В одном варианте данного изобретения антитела человека получают путем иммунизации животного, отличного от человека, содержащего некоторые или все локусы иммуноглобулина человека, IGF-IR-антигеном. В предпочтительном варианте животным, отличным от человека, является ксеномышь XENOMOUSE ™, которая представляет собой сконструированный штамм мышей, который содержит большие фрагменты локусов иммуноглобулина человека и дефицитна по продуцированию мышиных антител. См., например, Green et al. Nature Genetics 7: 13-21 (1994) и патенты США 5916771, 5939598, 5985615, 5998209, 6075181, 6091001, 6114598 и 6130364. См. также заявку WO 91/10741, опубликованную 25 июля 1991, WO 94/02602, опубликованную 3 февраля 1994, WO 96/34096 и WO 96/33735, которые обе опубликованы 31 октября 1996, WO 98/16654, опубликованную 23 апреля 1998, WO 98/24893, опубликованную 11 июня 1998, WO 98/50433, опубликованную 12 ноября 1998, WO 99/45031, опубликованную 10 сентября 1999, WO 99/53049, опубликованную 21 октября 1999, WO 00/09560, опубликованную 24 февраля 2000, и WO 00/037504, опубликованную 29 июня 2000. XENOMOUSE ™ продуцирует набор полностью человеческих антител, соответствующий набору у взрослого человека, и образует антиген-специфичные мАт человека. XENOMOUSE ™ второго поколения содержит примерно 80% набора антител человека вследствие введения YAC-фрагментов локусов тяжелой цепи и локусов легкой цепи κ человека, имеющих структуру зародышевой линии и размер, измеряемый миллионами оснований. См. Mendez et al. Nature Genetics 15: 146-156 (1997), Green and Jakobovits J. Exp. Med. 188: 483-495 (1998), описания которых включены в данную заявку в виде ссылки. Изобретение также относится к способу получения анти-IGF-IR-антител из организма животного, отличного от человека и отличного от мыши, путем иммунизации трансгенных животных, отличных от человека, которые содержат локусы иммуноглобулинов человека. Таких животных можно получить с использованием способов, описанных непосредственно выше. Способы, описанные в указанных патентах, можно модифицировать, как описано в патенте США 5994619. В предпочтительном варианте животными, отличными от человека, могут быть крысы, овцы, свиньи, козы, крупный рогатый скот или лошади. В другом варианте животными, отличными от человека, содержащими локусы иммуноглобулиновых генов человека, являются животные, которые имеют "мини-локус" иммуноглобулинов человека. В методике на основе мини-локуса осуществляют имитацию экзогенного локуса Ig посредством включения отдельных генов из локуса Ig. Таким образом, из одного или более генов V H , одного или более генов D H , одного или более генов J H , константной области mu и второй константной области (предпочтительно константной области гамма) образуют конструкцию для встраивания в организм животного. Указанная методика описана наряду с другими в патентах США № № 5545807, 5545806, 5625825, 5625126, 5633425, 5661016, 5770429, 5789650, 5814318, 5591669, 5612205, 5721367, 5789215 и 5643763, включенных в данное описание в виде ссылки. Преимуществом методики на основе мини-локуса является скорость, с которой могут быть созданы и введены в животных конструкции, включающие части локуса Ig. Однако возможным недостатком методики на основе мини-локуса является то, что может иметь место недостаточное разнообразие иммуноглобулинов, чтобы поддержать полное развитие В-клеток, так что продукция антител может быть ниже. Для того, чтобы получить анти-IGF-IR-антитело человека, животное, отличное от человека, содержащее некоторые или все локусы иммуноглобулинов человека, иммунизируют IGF-IR-антигеном и из организма животного выделяют антитело или клетку, продуцирующую антитело. IGF-IR-антиген может представлять собой выделенный и/или очищенный IGF-IR и предпочтительно является IGF-IR человека. В другом варианте IGF-IR-антигеном является фрагмент IGF-IR, предпочтительно внеклеточный домен IGF-IR. В другом варианте IGF-IR-антигеном является фрагмент, который содержит по меньшей мере один эпитоп IGF-IR. В другом варианте IGF-IR-антигеном является клетка, которая экспрессирует IGF-IR на своей клеточной поверхности, предпочтительно клетка, которая сверхэкспрессирует IGF-IR на своей клеточной поверхности. Иммунизацию животных можно проводить любым способом, известным в данной области. См., например, Harlow and Lane, Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Press, 1990. В данной области хорошо известны способы иммунизации животных, отличных от человека, таких как мыши, крысы, овцы, козы, свиньи, крупный рогатый скот и лошади. См., например, Harlow and Lane и патент США 5994619. В предпочтительном варианте IGF-IR-антиген вводят с адъювантом, чтобы стимулировать иммунный ответ. Такие адъюванты включают неполный адъювант Фрейнда, RIBI (мурамилдипептиды) или ISCOM (иммуностимулирующие комплексы). Такие адъюванты могут защищать полипептид от быстрого рассеивания посредством депонирования в виде локального отложения, или они могут содержать вещества, которые стимулируют хозяина к секреции факторов, которые являются хемотаксическими для макрофагов и других компонентов иммунной системы. Предпочтительно при введении полипептида схема иммунизации будет включать два или большее количество введений полипептида, проводимых в течение нескольких недель. В примере 1 представлен протокол иммунизации ксеномыши XENOMOUSE ™ полноразмерным IGF-IR человека в фосфатно-солевом буфере. После иммунизации животного IGF-IR-антигеном от животного можно получить антитела и/или клетки, продуцирующие антитела. Сыворотку, содержащую анти-IGF-IR-антитела, получают от животного посредством взятия крови или забоя животного. Можно использовать сыворотку в том виде, как она получена от животного, из сыворотки можно получить иммуноглобулиновую фракцию или из сыворотки можно очистить анти-IGF-IR-антитела. Сыворотка или иммуноглобулины, полученные таким образом, являются поликлональными, что имеет недостатки, поскольку количество антител, которое можно получить, ограничено и поликлональное антитело обладает гетерогенным набором свойств. В другом варианте от иммунизированного животного можно получить иммортализованные гибридомы, продуцирующие антитела. После иммунизации животного умерщвляют, и В-клетки селезенки сливают с иммортализованными клетками миеломы, как хорошо известно в данной области. См., например, Harlow and Lane, выше. В предпочтительном варианте клетки миеломы не секретируют полипептиды иммуноглобулинов (несекретирующая линия клеток). После слияния и селекции с использованием антибиотиков проводят скрининг гибридом, используя IGF-IR, его часть или клетку, экспрессирующую IGF-IR. В предпочтительном варианте начальный скрининг выполняют с использованием твердофазного иммуноферментного анализа (ELISA) или радиоиммуноанализа (РИА), предпочтительно ELISA. Пример скрининга с применением ELISA представлен в заявке WO 00/37504, включенной в данное описание в виде ссылки. В другом варианте клетки, продуцирующие антитела, можно получить от человека, у которого имеется аутоиммунное нарушение и у которого экспрессируются анти-IGF-IR-антитела. Клетки, экспрессирующие анти-IGF-IR-антитела, можно выделить, выделяя лейкоциты и подвергая их флуоресцентно активируемой сортировке клеток (FACS) или посредством пэннинга на чашках, покрытых IGF-IR или его частью. Указанные клетки можно слить с несекретирующей миеломой человека, чтобы получить гибридомы человека, экспрессирующие анти-IGF-IR-антитела человека. В общем, это менее предпочтительный вариант, так как анти-IGF-IR-антитела, вероятно, будут иметь низкую аффинность по отношению к IGF-IR. Гибридомы, продуцирующие анти-IGF-IR-антитело, отбирают, клонируют и подвергают дальнейшему скринингу в отношении требуемых характеристик, включая устойчивый рост гибридомы, высокую продукцию антител и требуемые характеристики антитела, которые обсуждаются далее. Гибридомы можно культивировать и размножать in vivo в сингенных животных, в животных, у которых отсутствует иммунная система, например у "голых" мышей, или в культуре клеток in vitro. Способы отбора, клонирования и размножения гибридом хорошо известны специалистам в данной области. Предпочтительно иммунизированным животным является животное, отличное от человека, которое экспрессирует гены иммуноглобулинов человека, и В-клетки селезенки сливают с миеломой, полученной от того же самого вида животного, отличного от человека. Более предпочтительно иммунизированным животным является ксеномышь XENOMOUSE ™, а линией клеток миеломы является несекретирующая миелома мышей, такая как линия клеток миеломы NSO-bcl2. См., например, пример 1. В одном аспекте изобретение относится к получаемым гибридомам, которые продуцируют анти-IGF-IR-антитела человека. В предпочтительном варианте гибридомы являются мышиными гибридомами, которые описаны выше. В другом предпочтительном варианте гибридомы получают для вида, отличного от человека, отличного от мыши, такого как крысы, овцы, свиньи, козы, крупный рогатый скот или лошади. В другом варианте гибридомы являются гибридомами человека, при этом несекретирующую миелому человека сливают с клеткой человека, экспрессирующей анти-IGF-IR-антитело. Представлены молекулы нуклеиновых кислот, кодирующие анти-IGF-IR-антитела согласно изобретению. В одном варианте молекула нуклеиновой кислоты кодирует тяжелую и/или легкую цепь анти-IGF-IR-иммуноглобулина. В предпочтительном варианте одна молекула нуклеиновой кислоты кодирует тяжелую цепь анти-IGF-IR-иммуноглобулина, а другая молекула нуклеиновой кислоты кодирует легкую цепь анти-IGF-IR-иммуноглобулина. В более предпочтительном варианте кодируемый иммуноглобулин является иммуноглобулином человека, предпочтительно IgG человека. Кодируемая легкая цепь может быть λ-цепью или κ-цепью, предпочтительно κ-цепью. Молекулу нуклеиновой кислоты, кодирующую вариабельную область легкой цепи, можно получить из гена V κ A30, А27 или 012. В предпочтительном варианте легкую цепь получают из гена V κ A30. В другом предпочтительном варианте молекула нуклеиновой кислоты, кодирующая легкую цепь, содержит соединительную область, полученную из J κ1, J κ2 или J κ4. В еще более предпочтительном варианте молекула нуклеиновой кислоты, кодирующая легкую цепь, содержит не более десяти изменений аминокислот по сравнению с геном V κ A30 зародышевой линии, предпочтительно не более шести изменений аминокислот и еще более предпочтительно не более трех изменений аминокислот. Изобретение относится к молекуле нуклеиновой кислоты, которая кодирует вариабельную область легкой цепи (VL), содержащую по меньшей мере три изменения аминокислот по сравнению с последовательностью зародышевой линии, при этом изменения аминокислот идентичны изменениям аминокислот по сравнению с последовательностью зародышевой линии в VL одного из антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. Изобретение также относится к молекуле нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность вариабельной области легкой цепи 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. Изобретение также относится к молекуле нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность одного или более CDR любой из легких цепей 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В предпочтительном варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность всех CDR любой из легких цепей 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность одной из SEQ ID NO: 2, 6, 10, 14, 18 или 22, или содержит последовательность нуклеиновой кислоты одной из SEQ ID NO: 1, 5, 9, 13, 17 или 21. В другом предпочтительном варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность одного или более CDR любой из SEQ ID NO: 2, 6, 10, 14, 18 или 22, или содержит последовательность нуклеиновой кислоты одного или более CDR любой из SEQ ID NO: 1, 5, 9, 13, 17 или 21. В более предпочтительном варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность всех CDR любой из SEQ ID NO: 2, 6, 10, 14, 18 или 22, или содержит последовательность нуклеиновой кислоты всех CDR любой из SEQ ID NO: 1, 5, 9, 13, 17 или 21. Изобретение также относится к молекулам нуклеиновой кислоты, которые кодируют аминокислотную последовательность VL, которая имеет последовательность аминокислот, которая по меньшей мере на 70, 75, 80, 85, 90, 95, 96, 97, 98 или 99% идентична описанной выше VL, в частности VL, которая содержит аминокислотную последовательность одной из SEQ ID NO: 2, 6, 10, 14, 18 или 22. Изобретение также относится к последовательности нуклеиновой кислоты, которая по меньшей мере на 70, 75, 80, 85, 90, 95, 96, 97, 98 или 99% идентична последовательности нуклеиновой кислоты одной из SEQ ID NO: 1, 5, 9, 13, 17 или 21. В другом варианте изобретение относится к молекуле нуклеиновой кислоты, кодирующей VL, которая в условиях высокой жесткости гибридизуется с молекулой нуклеиновой кислоты, кодирующей VL, которая описана выше, в частности молекулой нуклеиновой кислоты, которая содержит последовательность нуклеиновой кислоты, кодирующую аминокислотную последовательность SEQ ID NO: 2, 6, 10, 14, 18 или 22. Изобретение также относится к последовательности нуклеиновой кислоты, кодирующей VL, которая в условиях высокой жесткости гибридизуется с молекулой нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты одной из SEQ ID NO: 1, 5, 9, 13, 17 или 21. Изобретение также относится к молекуле нуклеиновой кислоты, кодирующей вариабельную область тяжелой цепи (VH), полученной из VH-гена DP-35, DP-47, DP-71 или VIV-4/4.35, предпочтительно VH-гена DP-35. В другом предпочтительном варианте молекула нуклеиновой кислоты, кодирующая VH, содержит соединительную область из JH6 или JH5, более предпочтительно JH6. В другом предпочтительном варианте D-участок получен из 3-3, 6-19 или 4-17. В еще более предпочтительном варианте молекула нуклеиновой кислоты, кодирующая VH, содержит не более десяти изменений аминокислот по сравнению с геном DP-47 зародышевой линии, предпочтительно не более шести изменений аминокислот и еще более предпочтительно не более трех изменений аминокислот. В высокопредпочтительном варианте молекула нуклеиновой кислоты, кодирующая VH, содержит по меньшей мере одно изменение аминокислот по сравнению с последовательностью зародышевой линии, при этом изменение аминокислоты идентично изменению аминокислоты по сравнению с последовательностью зародышевой линии в тяжелой цепи одного из антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В еще более предпочтительном варианте VH содержит по меньшей мере три изменения аминокислот по сравнению с последовательностями зародышевой линии, при этом изменения идентичны изменениям по сравнению с последовательностью зародышевой линии в VH одного из антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В одном варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность VH 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность одного или более CDR тяжелой цепи 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В предпочтительном варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотные последовательности всех CDR тяжелой цепи 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом предпочтительном варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность одной из SEQ ID NO: 4, 8, 12, 16, 20 или 24, или содержит последовательность нуклеиновой кислоты одной из SEQ ID NO: 3, 7, 11, 15, 19 или 23. В другом предпочтительном варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотную последовательность одного или более CDR любой из SEQ ID NO: 4, 8, 12, 16, 20 или 24, или содержит последовательность нуклеиновой кислоты одного или более CDR любой из SEQ ID NO: 3, 7, 11, 15, 19 или 23. В предпочтительном варианте молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая кодирует аминокислотные последовательности всех CDR любой из SEQ ID NO: 4, 8, 12, 16, 20 или 24, или содержит последовательность нуклеиновой кислоты всех CDR любой из SEQ ID NO: 3, 7, 11, 15, 19 или 23. В другом варианте молекула нуклеиновой кислоты кодирует аминокислотную последовательность VH, которая по меньшей мере на 70, 75, 80, 85, 90, 95, 96, 97, 98 или 99% идентична одной из аминокислотных последовательностей, кодирующих VH, которые описаны непосредственно выше, в частности VH, которая содержит аминокислотную последовательность одной из SEQ ID NO: 4, 8, 12, 16, 20 или 24. Изобретение также относится к последовательности нуклеиновой кислоты, которая по меньшей мере на 70, 75, 80, 85, 90, 95, 96, 97, 98 или 99% идентична последовательности нуклеиновой кислоты одной из SEQ ID NO: 3, 7, 11, 15, 19 или 23. В другом варианте молекулой нуклеиновой кислоты, кодирующей VH, является молекула, которая в условиях высокой жесткости гибридизуется с последовательностью нуклеиновой кислоты, кодирующей VH, которая описана выше, в частности VH, которая содержит аминокислотную последовательность одной из SEQ ID NO: 4, 8, 12, 16, 20 или 24. Изобретение также относится к последовательности нуклеиновой кислоты, кодирующей VH, которая в условиях высокой жесткости гибридизуется с молекулой нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты одной из SEQ ID NO: 3, 7, 11, 15, 19 или 23. Молекула нуклеиновой кислоты, кодирующая одну из двух или обе полные тяжелую и легкую цепи анти-IGF-IR-антитела или их вариабельные области, можно получить из любого источника, который продуцирует анти-IGF-IR-антитело. Способы выделения мРНК, кодирующей антитело, хорошо известны в данной области. См., например, Sambrook et al. мРНК можно использовать для получения кДНК для применения в полимеразной цепной реакции (ПЦР) или при кДНК-клонировании генов антитела. В одном варианте изобретения молекулы нуклеиновой кислоты можно получить из гибридомы, которая экспрессирует анти-IGF-IR-антитело, которая описана выше, предпочтительно из гибридомы, которая имеет в качестве одного из партнеров слияния клетку трансгенного животного, которая экспрессирует гены иммуноглобулинов человека, такого как XENOMOUSE ™, трансгенного животного, отличного от человека, такого как мышь, или трансгенного животного, отличного от человека и отличного от мыши. В другом варианте гибридому получают из нетрансгенного животного, отличного от человека, которое можно использовать, например, для получения гуманизированных антител. Молекулу нуклеиновой кислоты, кодирующую полную тяжелую цепь анти-IGF-IR-антитела, можно сконструировать посредством слияния молекулы нуклеиновой кислоты, кодирующей вариабельный домен тяжелой цепи или ее антигенсвязывающий домен, с константным доменом тяжелой цепи. Подобным образом молекулу нуклеиновой кислоты, кодирующую легкую цепь анти-IGF-IR-антитела, можно сконструировать посредством слияния молекулы нуклеиновой кислоты, кодирующей вариабельный домен легкой цепи или ее антигенсвязывающий домен, с константным доменом легкой цепи. Молекулы нуклеиновой кислоты, кодирующие цепь VH и VL, можно превратить в полноразмерные гены антитела путем их встраивания в экспрессирующие векторы, уже кодирующие константную область тяжелой цепи и константную область легкой цепи, соответственно, так, чтобы участок VH был функционально связан с участком(ами) константной области тяжелой цепи (СН) в векторе, а участок VL был функционально связан с участком константной области легкой цепи (CL) в векторе. Альтернативно, молекулы нуклеиновой кислоты, кодирующие цепи VH или VL, превращают в полноразмерные гены антитела связыванием, например лигированием, молекулы нуклеиновой кислоты, кодирующей цепь VH, с молекулой нуклеиновой кислоты, кодирующей цепь СН, используя стандартные способы молекулярной биологии. Этого же можно достичь с использованием молекул нуклеиновой кислоты, кодирующих цепи VL и CL. Последовательности генов константных областей тяжелой и легкой цепей человека известны в данной области. См., например, Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed., NIH Publ. No. 91-3242, 1991. Затем молекулы нуклеиновой кислоты, кодирующие полноразмерные тяжелые и/или легкие цепи, можно экспрессировать из клетки, в которую они введены, и выделить анти-IGF-IR-антитело. В предпочтительном варианте нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи, кодирует аминокислотную последовательность SEQ ID NO: 4, 8, 12, 16, 20 или 24, а молекула нуклеиновой кислоты, кодирующая вариабельную область легких цепей, кодирует аминокислотную последовательность SEQ ID NO: 2, 6, 10, 14, 18 или 22. SEQ ID NO: 28 показывает аминокислотную последовательность, a SEQ ID NO: 27 показывает последовательность нуклеиновой кислоты, кодирующую константную область тяжелой цепи анти-IGF-IR-антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 и 6.1.1. SEQ ID NO: 26 показывает аминокислотную последовательность, a SEQ ID NO: 25 показывает последовательность нуклеиновой кислоты, кодирующую константную область легкой цепи анти-IGF-IR-антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 и 6.1.1. Таким образом, в предпочтительном варианте молекула нуклеиновой кислоты, кодирующая константную область тяжелой цепи, кодирует SEQ ID NO: 28, а молекула нуклеиновой кислоты, кодирующая константную область легкой цепи, кодирует SEQ ID NO: 26. В более предпочтительном варианте молекула нуклеиновой кислоты, кодирующая константный домен тяжелой цепи, имеет последовательность нуклеиновой кислоты SEQ ID NO: 27, а молекула нуклеиновой кислоты, кодирующая константный домен, имеет последовательность нуклеиновой кислоты SEQ ID NO: 25. В другом варианте молекулу нуклеиновой кислоты, кодирующую либо тяжелую цепь анти-IGF-IR-антитела или ее антигенсвязывающий домен, либо легкую цепь анти-IGF-IR-антитела или ее антигенсвязывающий домен, можно выделить из животного, отличного от человека и отличного от мыши, которое экспрессирует гены иммуноглобулина человека и было иммунизировано IGF-IR-антигеном. В другом варианте молекулу нуклеиновой кислоты можно выделить из клетки, продуцирующей анти-IGF-IR-антитело, полученной из нетрансгенного животного или из организма пациента-человека, у которого продуцируются анти-IGF-IR-антитела. В качестве способов выделения мРНК из клеток, продуцирующих анти-IGF-IR-антитела, можно использовать стандартные способы выделения, мРНК можно клонировать и/или амплифицировать с использованием ПЦР и способов конструирования библиотек и провести скрининг с использованием стандартных протоколов, чтобы получить молекулы нуклеиновой кислоты, кодирующие тяжелые и легкие цепи анти-IGF-IR-антител. Молекулы нуклеиновой кислоты можно использовать для того, чтобы на основе рекомбинантного способа экспрессировать большие количества анти-IGF-IR-антител, как описано ниже. Молекулы нуклеиновой кислоты также можно использовать для получения химерных антител, одноцепочечных антител, иммуноадгезинов, диантител, мутантных антител и производных антител, которые описаны далее. Если молекулы нуклеиновой кислоты получают от нетрансгенного животного, отличного от человека, молекулы нуклеиновой кислоты можно использовать для гуманизации антител так, как описано ниже. В другом варианте молекулы нуклеиновой кислоты согласно изобретению можно использовать в качестве зондов или праймеров ПЦР для специфичных последовательностей антител. Например, зонд на основе молекулы нуклеиновой кислоты можно использовать в диагностических способах или праймер ПЦР на основе молекулы нуклеиновой кислоты можно использовать для амплификации областей ДНК, которые можно использовать, наряду с прочим, для выделения последовательностей нуклеиновой кислоты для применения при получении вариабельных доменов анти-IGF-IR-антител. В предпочтительном варианте молекулы нуклеиновой кислоты представляют собой олигонуклеотиды. В более предпочтительном варианте олигонуклеотиды получены из высоковариабельных областей тяжелой и легкой цепей представляющего интерес антитела. В еще более предпочтительном варианте олигонуклеотиды кодируют все или часть одного или более CDR. Изобретение относится к векторам, содержащим молекулы нуклеиновой кислоты согласно изобретению, которые кодируют тяжелую цепь или ее антигенсвязывающую часть. Изобретение также относится к векторам, содержащим молекулы нуклеиновой кислоты согласно изобретению, которые кодируют легкую цепь или ее антигенсвязывающую часть. Изобретение также относится к векторам, содержащим молекулы нуклеиновой кислоты, кодирующие слитые белки, модифицированные антитела, фрагменты антител и их зонды. Для того, чтобы экспрессировать антитела или части антител согласно изобретению, ДНК, кодирующие неполные или полноразмерные легкие и тяжелые цепи, полученные, как описано выше, встраивают в экспрессирующие векторы так, чтобы гены были функционально связаны с последовательностями регуляции транскрипции и трансляции. Экспрессирующие векторы включают плазмиды, ретровирусы, космиды, YAC, эписомы, полученные из EBV, и т.п. Ген антитела лигируют в вектор так, чтобы последовательности регуляции транскрипции и трансляции в векторе выполняли свою предназначенную функцию регулирования транскрипции и трансляции гена антитела. Экспрессирующий вектор и последовательности регуляции экспрессии выбирают так, чтобы они были совместимы с используемой клеткой-хозяином для экспрессии. Ген легкой цепи антитела и ген тяжелой цепи антитела можно встроить в отдельные векторы. В предпочтительном варианте оба гена встраивают в один и тот же экспрессирующий вектор. Гены антитела встраивают в экспрессирующий вектор стандартными способами (например, лигированием комплементарных сайтов рестрикции на фрагменте гена антитела и векторе или лигированием тупых концов, если сайты рестрикции отсутствуют). Подходящим вектором является вектор, который кодирует функционально полную последовательность СН или CL иммуноглобулина человека, с соответствующими сайтами рестрикции, сконструированными так, чтобы можно было легко встроить и экспрессировать любую последовательность VH или VL, которые описаны выше. В таких векторах сплайсинг обычно происходит между донорным сайтом сплайсинга во встроенной J-области и акцепторным сайтом сплайсинга, предшествующим С-области человека, а также в областях сплайсинга, которые имеются в экзонах СН человека. Полиаденилирование и терминация транскрипции происходят в нативных сайтах хромосомы ниже кодирующих областей. Рекомбинантный экспрессирующий вектор также может кодировать сигнальный пептид, который способствует секреции цепи антитела из клетки-хозяина. Ген цепи антитела можно клонировать в векторе так, чтобы сигнальный пептид был связан в рамке с аминоконцом гена цепи антитела. Сигнальный пептид может представлять собой сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (т.е. сигнальный пептид из белка, отличного от иммуноглобулина). Кроме генов цепей антител рекомбинантные экспрессирующие векторы согласно изобретению несут регуляторные последовательности, которые регулируют экспрессию генов цепей антител в клетке-хозяине. Специалистам в данной области будет понятно, что конструирование экспрессирующего вектора, включая подбор регуляторных последовательностей, может зависеть от таких факторов, как выбор клетки-хозяина, которую необходимо трансформировать, уровня экспрессии требуемого белка и т.д. Предпочтительные регуляторные последовательности для экспрессии в клетке-хозяине млекопитающего включают вирусные элементы, которые обеспечивают высокие уровни экспрессии белка в клетках млекопитающих, такие как промоторы и/или энхансеры, полученные из ретровирусных LTR, цитомегаловируса (CMV) (такие, как промотор/энхансер CMV), обезьяньего вируса 40 (SV40) (такой, как промотор/энхансер SV40), аденовируса (например, главный поздний промотор аденовируса (AdMLP)), полиомы, и сильных промоторов млекопитающих, таких как нативные промоторы иммуноглобулина и актина. Для дальнейшего знакомства с описанием вирусных регуляторных элементов и их последовательностей см., например, патент США № 5168062, Stinski; патент США № 4510245, Bell et al.; и патент США № 4968615, Schaffner et al. Кроме генов цепей антител и регуляторных последовательностей рекомбинантные экспрессирующие векторы согласно изобретению могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетке-хозяине (например, начала репликации) и гены селективных маркеров. Ген селективного маркера облегчает селекцию клеток-хозяев, в которые был введен вектор (см., например, патенты США № № 4399216, 4634665 и 5179017, все патенты Axel et al.). Например, ген селективного маркера обычно придает клетке-хозяину, в которую был введен вектор, резистентность к лекарственным средствам, таким как G418, гигромицин или метотрексат. Предпочтительные гены селективного маркера включают ген дигидрофолатредуктазы (DHFR) (для применения в клетках-хозяевах dhfr - при селекции/амплификации с метотрексатом) и ген neo (для селекции с помощью G418). Молекулы нуклеиновых кислот, кодирующие тяжелую цепь или ее антигенсвязывающую часть и/или легкую цепь или ее антигенсвязывающую часть анти-IGF-IR-антитела, и векторы, содержащие указанные молекулы нуклеиновых кислот, можно использовать для трансформации подходящей клетки-хозяина млекопитающего. Трансформацию можно осуществлять любым известным способом для введения полинуклеотидов в клетку-хозяин. Способы введения гетерологичных полинуклеотидов в клетки млекопитающих хорошо известны в данной области и включают трансфекцию, опосредованную декстраном, осаждение фосфатом кальция, трансфекцию, опосредованную полибреном, слияние протопластов, электропорацию, инкапсулирование полинуклеотида(тов) в липосомы, биолистическую инъекцию и прямую микроинъекцию ДНК в ядра. Кроме того, молекулы нуклеиновых кислот можно ввести в клетки млекопитающих с помощью вирусных векторов. Способы трансформации клеток хорошо известны в данной области. См., например, патенты США № № 4399216, 4912040, 4740461 и 4959455 (и указанные патенты, таким образом, включены в данное описание в виде ссылки). Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии, хорошо известны в данной области и включают многие иммортализованные линии клеток, доступные из Американской коллекции типов культур (АТСС). Указанные клетки наряду с другими включают клетки яичника китайского хомячка (СНО), NSO, клетки SP2, клетки HeLa, клетки почки детеныша хомячка (ВНК), клетки почки обезьяны (COS), клетки гепатоклеточной карциномы человека (например, Hep G2), клетки А549, клетки 3Т3 и ряд других клеточных линий. Клетки-хозяева млекопитающих включают клетки человека, мыши, крысы, собаки, обезьяны, свиньи, козы, коровы, лошади и хомячка. Особенно предпочтительные линии клеток отбирают посредством определения того, какие линии клеток имеют высокие уровни экспрессии. Другими линиями клеток, которые можно использовать, являются линии клеток насекомых, такие как клетки Sf9, клетки амфибий, бактериальные клетки, клетки растений и клетки грибов. В том случае, когда рекомбинантные экспрессирующие векторы, кодирующие тяжелую цепь или ее антигенсвязывающую часть, легкую цепь и/или ее антигенсвязывающую часть, вводят в клетки-хозяева млекопитающих, антитела получают посредством культивирования клеток-хозяев в течение периода времени, достаточного для того, чтобы обеспечить экспрессию антитела в клетках-хозяевах или более предпочтительно секрецию антитела в культуральную среду, в которой выращивают клетки-хозяева. Антитела можно извлечь из культуральной среды с использованием стандартных способов очистки белка. Кроме того, экспрессию антител согласно изобретению (или других их фрагментов) продуцирующими линиями клеток можно усилить, используя ряд известных способов. Например, широко используемым подходом для усиления экспрессии при некоторых условиях является система экспрессии гена глутаминсинтетазы (система GS). Система GS обсуждается в целом или частично в европейских патентах № № 0216846, 0256055 и 0323997 и европейской патентной заявке № 89303964.4. Возможно, что антитела, экспрессируемые различными линиями клеток или трансгенными животными, будут отличаться друг от друга гликозилированием. Однако все антитела, кодируемые представленными в данном описании молекулами нуклеиновых кислот или содержащие представленные в данном описании аминокислотные последовательности, являются частью данного изобретения независимо от гликозилирования антител. Изобретение также относится к трансгенным животным, отличным от человека, содержащим одну или более молекул нуклеиновой кислоты согласно изобретению, которые можно использовать для получения антител согласно изобретению. Антитела можно получить и извлечь из ткани или жидкостей организма, таких как молоко, кровь или моча, коз, коров, лошадей, свиней, крыс, мышей, кроликов, хомячков или других млекопитающих. См., например, патенты США № № 5827 690, 5756687, 5750172 и 5741957. Как описано выше, трансгенных животных, отличных от человека, которые содержат локусы иммуноглобулинов человека, можно получить иммунизацией IGF-IR или его частью. В другом варианте трансгенных животных, отличных от человека, получают введением в животного одной или более молекул нуклеиновой кислоты согласно изобретению стандартными трансгенными способами. См. Hogan, выше. Трансгенными клетками, используемыми для получения трансгенного животного, могут быть эмбриональные стволовые клетки или соматические клетки. Трансгенные организмы, отличные от человека, могут быть химерными, нехимерными гетерозиготами и нехимерными гомозиготами. См., например, Hogan et al., Manipulating the Mouse Embryo: A Laboratory Manual 2ed., Cold Spring Harbor Press (1999); Jackson et al., Mouse Genetics and Transgenics: A Practical Approach, Oxford University Press (2000); и Pinkert, Transgenic Animal Technology: A Laboratory Handbook, Academic Press (1999). В другом варианте трансгенные организмы, отличные от человека, могут иметь целенаправленное нарушение и замену, которая кодирует представляющую интерес тяжелую цепь и/или легкую цепь. В предпочтительном варианте трансгенные животные содержат и экспрессируют молекулы нуклеиновой кислоты, кодирующие тяжелую и легкую цепи, которые специфично связываются с IGF-IR, предпочтительно с IGF-IR человека. В другом варианте трансгенные животные содержат молекулы нуклеиновой кислоты, кодирующие модифицированное антитело, такое как одноцепочечное антитело, химерное антитело или гуманизированное антитело. Анти-IGF-IR-антитела можно получить в любом трансгенном животном. В предпочтительном варианте животными, отличными от человека, являются мыши, крысы, овцы, свиньи, козы, крупный рогатый скот или лошади. Трансгенное животное, отличное от человека, экспрессирует указанные кодируемые полипептиды в крови, молоке, моче, слюне, слезах, слизи и других жидкостях организма. Изобретение относится к способу получения анти-IGF-IR-антитела или его антигенсвязывающей части, включающему стадии синтеза библиотеки человеческих антител на фаге, скрининга библиотеки с помощью IGF-IR или его части, выделения фага, который связывает IGF-IR, и получения антитела из фага. Один из способов получения библиотеки антител включает стадии иммунизации животного-хозяина, отличного от человека, содержащего локус иммуноглобулина человека, с помощью IGF-IR или его антигенной части, чтобы вызвать иммунный ответ, извлечения клеток из животного-хозяина, клеток, которые ответственны за продукцию антител; выделения РНК их извлеченных клеток, обратной транскрипции РНК для получения кДНК, амплификации кДНК с использованием праймера и встраивания кДНК в вектор для фагового дисплея так, чтобы на фаге экспрессировались антитела. Таким образом можно получить рекомбинантные анти-IGF-IR-антитела согласно изобретению. Рекомбинантные анти-IGF-IR-антитела человека согласно изобретению, дополнительно к описанным в данном изобретении анти-IGF-IR-антителам, можно выделить посредством скрининга рекомбинантной комбинаторной библиотеки антител, предпочтительно библиотеки на основе фагового дисплея scFv, полученного с использованием кДНК VL и VH человека, приготовленной на мРНК, полученной из лимфоцитов человека. Методики получения и скрининга таких библиотек известны в данной области. Имеются коммерчески доступные наборы для создания библиотек на основе фагового дисплея (например, система для антител на основе рекомбинантного фага Pharmacia, № по каталогу 27-9400-01; и набор для фагового дисплея Stratagene SurfZAP ™, № по каталогу 240612). Также существуют другие способы и реагенты, которые можно использовать при создании и скрининге дисплейных библиотек антител (см., например, Ladner et al., патент США № 5223409; Kang et al., заявка PCT № WO 92/18619; Dower et al., заявка PCT № WO 91/17271; Winter et al., заявка РСТ № WO 92/20791; Markland et al., заявка РСТ № WO 92/15679; Breitling et al., заявка PCT № WO 93/01288; McCafferty et al., заявка РСТ № WO 92/01047; Garrard et al., заявка РСТ № WO 92/09690; Fuchs et al. (1991) Bio/Technology 9: 1370-1372; Hay et al. (1992) Hum. Antibod. Hybridomas 3: 81-85; Huse et al. (1989) Science 246: 1275-1281; McCafferty et al., Nature (1990) 348: 552-554; Griffiths et al. (1993) EMBO J. 12: 725-734; Hawkins et al. (1992) J. Mol. Biol. 226: 889-896; Clackson et al. (1991) Nature 352: 624-628; Gram et al. (1992) Proc. Natl. Acad. Sci. USA 89: 3576-3580; Garrad et al. (1991) Bio/Technology 9: 1373-1377; Hoogenboom et al. (1991) Nuc. Acid Res. 19: 4133-4137; и Barbas et al. (1991) Proc. Natl. Acad. Sci. USA 88: 7978-7982). В предпочтительном варианте для выделения анти-IGF-IR-антител человека с требуемыми параметрами описанное в данной заявке анти-IGF-IR-антитело человека сначала используют для отбора последовательностей тяжелой и легкой цепей человека, обладающих сходной связывающей активностью по отношению к IGF-IR, используя способы импринтинга эпитопов, описанные в Hoogenboom et al., заявка РСТ № WO 93/06213. Библиотеки антител, используемые в данном способе, предпочтительно представляют собой scFv-библиотеки, полученные и подвергнутые скринингу, как описано в McCafferty et al., заявка РСТ № WO 92/01047, McCafferty et al., Nature (1990) 348: 552-554; и Griffiths et al., (1993) EMBO J. 12: 725-734. Скрининг библиотек scFv-антител предпочтительно осуществляют с использованием в качестве антигена IGF-IR человека. После того, как выбраны исходные участки VL и VH человека, выполняют эксперименты по "сочетанию и подбору пар", в которых проводят скрининг различных частей исходно выбранных участков VL и VH в отношении связывания IGF-IR, чтобы выбрать предпочтительные комбинации в паре VL/VH. Кроме того, для дальнейшего улучшения качества антитела участки VL и VH в предпочтительной паре(ах) VL/VH можно подвергнуть случайному мутированию предпочтительно в области CDR3 VH и/или VL, в процессе, аналогичном процессу образования соматических мутаций in vivo, ответственных за аффинное созревание антител во время естественного иммунного ответа. Указанное аффинное созревание in vitro можно осуществить посредством амплификации областей VH и VL с использованием ПЦР-праймеров, комплементарных CDR3 VH или CDR3 VL, соответственно, при этом указанные праймеры в определенных положениях "мечены" с помощью случайной смеси четырех оснований нуклеотидов так, чтобы полученные в результате продукты ПЦР кодировали участки VH и VL, в которые были введены случайные мутации в областях CDR3 VH и/или VL. Такие случайно мутированные участки VH и VL можно подвергнуть повторному скринингу в отношении связывания с IGF-IR. После скрининга и выделения анти-IGF-IR-антитела согласно изобретению из рекомбинантной дисплейной библиотеки иммуноглобулинов нуклеиновую кислоту, кодирующую выбранное антитело, можно извлечь из упаковки в фаг (например, из фагового генома) и субклонировать в другие экспрессирующие векторы стандартными способами на основе рекомбинантной ДНК. При желании, нуклеиновую кислоту можно подвергнуть дальнейшей обработке, чтобы создать другие формы антитела согласно изобретению, как описано ниже. Чтобы экспрессировать рекомбинантное антитело человека, выделенное в результате скрининга комбинаторной библиотеки, ДНК, кодирующую антитело, клонируют в рекомбинантном экспрессирующем векторе и вводят в клетки-хозяева млекопитающих, которые описаны выше. Другим аспектом данного изобретения является предоставление механизма, посредством которого можно переключить класс анти-IGF-IR-антитела на другой. В одном аспекте изобретения молекулу нуклеиновой кислоты, кодирующую VL или VH, выделяют с использованием способов, хорошо известных в данной области, так, чтобы они не содержали никаких последовательностей нуклеиновой кислоты, кодирующих CL или СН. Затем молекулу нуклеиновой кислоты, кодирующую VL или VH, функционально связывают с последовательностью нуклеиновой кислоты, кодирующей CL или СН из молекулы иммуноглобулина другого класса. Это можно осуществить с использованием вектора или молекулы нуклеиновой кислоты, которая содержит цепь CL или СН, как описано выше. Например, анти-IGF-IR-антитело, которое исходно представляло собой IgM, можно переключить на другой класс IgG. Кроме того, переключение классов можно использовать для превращения одного подкласса IgG в другой, например IgG1 в IgG2. Предпочтительный способ получения антитела согласно изобретению, содержащего требуемые изотипы, включает стадии выделения нуклеиновой кислоты, кодирующей тяжелую цепь анти-IGF-IR-антитела, и нуклеиновой кислоты, кодирующей легкую цепь анти-IGF-IR-антитела, получения вариабельной области тяжелой цепи, лигирования вариабельной области тяжелой цепи с константным доменом тяжелой цепи требуемого изотипа, экспрессии легкой цепи и лигированной тяжелой цепи в клетке и сбора анти-IGF-IR-антитела требуемого изотипа. Описанные выше молекулы нуклеиновых кислот можно использовать для создания производных антител с использованием методик и способов, известных специалисту в данной области. Как обсуждалось выше в связи с созданием антител человека, получение антител с пониженной иммуногенностью имеет преимущества. В некоторой степени это можно осуществить с использованием способа гуманизации и методики дисплея с применением подходящих библиотек. Будет понятно, что мышиные антитела или антитела из других видов можно гуманизировать или приматизировать с использованием способа, хорошо известного в данной области. См., например, Winter and Harris Immunol Today 14: 43-46 (1993) и Wright et al. Crit. Reviews in Immunol. 12125-168 (1992). Представляющее интерес антитело можно сконструировать способом на основе рекомбинантной ДНК, чтобы заменить домены CH1, CH2, СН3, шарнирный домен и/или каркасный домен соответствующей последовательностью человека (см. WO 92/02190 и патенты США № № 5530101, 5585089, 5693761, 5693792, 5714350 и 5777085). В предпочтительном варианте анти-IGF-IR-антитело можно гуманизировать заменой доменов CH1, CH2, СН3, шарнирного домена и/или каркасного домена соответствующей последовательностью человека при сохранении всех CDR тяжелой цепи, легкой цепи или обеих, тяжелой и легкой, цепей. В другом варианте молекулы нуклеиновой кислоты, векторы и клетки-хозяева можно использовать для получения мутантных анти-IGF-IR-антител. Можно получить мутации антител в вариабельных доменах тяжелой и/или легкой цепей, чтобы изменить связывающие свойства антитела. Например, можно получить мутацию в одной или более областях CDR, чтобы увеличить или уменьшить K d антитела по отношению к IGF-IR, увеличить или уменьшить K off или изменить специфичность связывания антитела. Способы сайт-специфичного мутагенеза хорошо известны в данной области. См., например, Sambrook et al. and Ausubel et al., выше. В предпочтительном варианте осуществляют мутацию в аминокислотном остатке, о котором известно, что он изменен по сравнению с зародышевой линией в вариабельной области анти-IGF-IR-антитела. В более предпочтительном варианте получают одну или более мутаций в аминокислотном остатке, о котором известно, что он изменяется по сравнению с зародышевой линией, в вариабельной области или CDR-области одного из анти-IGF-IR-антител 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом варианте получают одну или более мутаций в аминокислотном остатке, о котором известно, что он изменен по сравнению с зародышевой линией, в вариабельной области или CDR-области, аминокислотная последовательность которых представлена в SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 или 24 или последовательность нуклеиновой кислоты которых представлена в SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21 или 23. В другом варианте молекулы нуклеиновой кислоты подвергают мутированию в одной или более каркасных областях. Можно получить мутацию в каркасной области или константном домене, чтобы увеличить время полужизни анти-IGF-IR-антитела. См., например, WO 00/09560, опубликованную 24 февраля 2000г., включенную в данное описание в виде ссылки. В одном варианте может иметь место одна, три или пять точечных мутаций и не более десяти точечных мутаций. Также можно получить мутацию в каркасной области или константном домене, чтобы изменить иммуногенность антитела, получить сайт для ковалентного или нековалентного связывания с другой молекулой, или для того, чтобы изменить такие свойства, как фиксация комплемента. Мутации можно осуществлять в каждой из каркасных областей, в константном домене и вариабельных областях в одном мутантном антителе. Альтернативно, мутации можно осуществить только в одной из каркасных областей, вариабельных областей или константном домене в одном мутантном антителе. В одном варианте имеется не более десяти изменений аминокислот в одной из VH- или VL-областей мутантного анти-IGF-IR-антитела по сравнению с анти-IGF-IR-антителом до мутивирования. В более предпочтительном варианте имеется не более пяти изменений аминокислот в одной из VH- или VL-областей мутантного анти-IGF-IR-антитела, более предпочтительно не более трех изменений аминокислот. В другом варианте имеется не более пятнадцати изменений аминокислот в константных доменах, более предпочтительно не более десяти изменений аминокислот, еще более предпочтительно не более пяти изменений аминокислот. В другом варианте можно получить слитое антитело или иммуноадгезин, который содержит все или часть анти-IGF-IR-антитела, связанного с другим полипептидом. В предпочтительном варианте с полипептидом связаны только вариабельные области анти-IGF-IR-антитела. В другом предпочтительном варианте домен VH анти-IGF-IR-антитела связан с первым полипептидом, тогда как домен VL анти-IGF-IR-антитела связан со вторым полипептидом, который связан с первым полипептидом таким образом, при котором домены VH и VL могут взаимодействовать друг с другом, образуя связывающий сайт антитела. В другом предпочтительном варианте домен VH отделен от домена VL линкером так, чтобы домены VH и VL могли взаимодействовать друг с другом (см. далее одноцепочечные антитела). Затем антитело VH-линкер-VL связывают с представляющим интерес полипептидом. Слитое антитело применимо для направления полипептида в клетку или ткань, экспрессирующую IGF-IR. Полипептид может представлять собой терапевтическое средство, такое как токсин, фактор роста или другой регуляторный белок, или может являться диагностическим средством, таким как фермент, который можно легко визуализировать, такой как пероксидаза хрена. Кроме того, можно создать слитые антитела, в которых два (или большее количество) одноцепочечных антител связаны друг с другом. Это применяется в том случае, когда требуется создать дивалентное или поливалентное антитело в одной полипептидной цепи или если требуется создать биспецифичное антитело. Чтобы создать одноцепочечное антитело (scFv), фрагменты ДНК, кодирующие VH и VL, функционально связывают с другим фрагментом, кодирующим гибкий линкер, например кодирующим аминокислотную последовательность (Gly 4 -Ser) 3 (SEQ ID NO: 60), так, чтобы последовательности VH и VL могли экспрессироваться в виде непрерывного одноцепочечного белка, содержащего VL- и VH-области, связанные гибким линкером (см., например, Bird et al. (1988) Science 242: 423-426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85: 5879-5883; McCafferty et al., Nature (1990) 343: 552-554). Одноцепочечное антитело может быть моновалентным в том случае, если используют только одну VH и VL, бивалентым, если используют две VH и VL, или поливалентным, если используют больше двух VH и VL. В другом варианте можно получить другие модифицированные антитела, используя молекулы нуклеиновой кислоты, кодирующие анти-IGF-IR. Например, можно получить "каппа-тела" (Ill et al., Protein Eng 10: 949-57 (1997)), "мини-антитела" (Martin et al., EMBO J 13 : 5303-9 (1994)), "диантитела" (Holliger et al., PNAS USA 90: 6444-6448 (1993)) или "Janusins" (Traunecker et al., EMBO J. 10: 3655-3659 (1991) и Traunecker et al. "Janusin: new molecular design for bispecific reagents", Int. J. Cancer Suppl. 7: 51-52 (1992)), используя стандартные способы молекулярной биологии, следуя инструкциям описания. В другом аспекте можно создать химерные и биспецифичные антитела. Можно получить химерное антитело, которое содержит CDR и каркасные области из разных антител. В предпочтительном варианте CDR химерного антитела содержат все CDR вариабельной области легкой цепи или тяжелой цепи анти-IGF-IR-антитела, тогда как каркасные области получены из одного или более разных антител. В более предпочтительном варианте CDR химерного антитела содержат все CDR вариабельных областей легкой цепи и тяжелой цепи анти-IGF-IR-антитела. Каркасные области могут быть из другого вида и в предпочтительном варианте могут быть гуманизированы. Альтернативно, каркасные области могут быть из другого антитела человека. Можно создать биспецифичное антитело, которое специфично связывается с IGF-IR посредством одного связывающего домена, а со второй молекулой посредством второго связывающего домена. Биспецифичное антитело можно получить рекомбинантным молекулярно-биологическим способом или в результате физической конъюгации друг с другом. Кроме того, можно создать одноцепочечное антитело, содержащее более одной VH и VL, которое специфично связывается с IGF-IR и с другой молекулой. Такие биспецифичные антитела можно создать, используя способы, которые хорошо известны, например, в связи с (i) и (ii), см., например, Fanger et al. Immunol Methods 4: 72-81 (1994), и Wright and Harris, выше, и в связи с (iii), см., например, Traunecker et al. Int. J. Cancer (Suppl.) 7: 51-52 (1992). В предпочтительном варианте биспецифичное антитело связывается с IGF-IR и с другой молекулой, экспрессируемой на высоком уровне на злокачественных или опухолевых клетках. В более предпочтительном варианте другой молекулой является рецептор erbB2, VEGF, CD20 или EGF-R. В одном варианте описанные выше модифицированные антитела получают, используя одну или более вариабельных областей или одну или более CDR-областей из одного из антител, выбранного из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 или 6.1.1. В другом варианте модифицированные антитела получают, используя одну или более вариабельных областей или одну или более CDR-областей, аминокислотная последовательность которых представлена в SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 или 24 или последовательность нуклеиновой кислоты которых представлена в SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21 или 23. Антитело или часть антитела согласно изобретению можно дериватизировать или связать с другой молекулой (например, другим пептидом или белком). В общем, антитела или их часть дериватизируют так, чтобы дериватизация или мечение не оказывали неблагоприятного воздействия на связывание IGF-IR. Таким образом, подразумевается, что антитела и части антител согласно изобретению включают как интактные, так и модифицированные формы анти-IGF-IR-антител человека, приведенные в данном описании. Например, антитело или часть антитела согласно изобретению могут быть функционально связаны (посредством химического связывания, генетического слияния, нековалентной ассоциации или другим образом) с одной или более молекулярными единицами, такими как другое антитело (например, биспецифичное антитело или диантитело), детектирующее средство, цитотоксическое средство, фармацевтическое средство и/или белок или пептид, который может опосредовать связывание антитела или части антитела с другой молекулой (такой, как коровая область стрептавидина или полигистидиновая метка). Один тип дериватизированного антитела получают посредством перекрестного сшивания двух или более антител (одного и того же типа или разных типов, например, чтобы создать биспецифичные антитела). Подходящие перекрестно-сшивающие агенты включают агенты, которые являются гетеробифункциональными, имеющими две отдельные реакционноспособные группы, разделенные подходящим спейсером (например, сложный эфир мета-малеимидобензоил-N-гидроксисукцинимида), или гомобифункциональными (например, дисукцинимидилсуберат). Такие линкеры доступны из Pierce Chemical Company, Rockford, Ill. Другим типом дериватизированного антитела является меченое антитело. Пригодные детектирующие средства, которыми может быть дериватизирозано антитело или часть антитела согласно изобретению, включают флуоресцентные соединения, включая флуоресцеин, изотиоцианат флуоресцеина, родамин, 5-диметиламин-1-нафталинсульфонилхлорид, фикоэритрин, люминофоры на основе лантанидов и т.п. Антитело также можно метить ферментами, которые применимы для делеции, такими как пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза, глюкозоксидаза и т.п. В том случае, когда антитело метят детектируемым ферментом, его выявляют посредством добавления дополнительных реагентов, которые фермент использует для получения продукта реакции, который можно различить. Например, в том случае, когда присутствует агент пероксидаза хрена, добавление перекиси водорода и диаминобензидина приводит к образованию окрашенного продукта реакции, который можно детектировать. Антитело также можно метить биотином и детектировать с помощью непрямого измерения связывания авидина или стрептавидина. Антитело можно метить магнитным агентом, таким как гадолиний. Антитело также можно метить предварительно определенными полипептидными эпитопами, распознаваемыми вторичным репортером (например, парные последовательности лейциновой молнии, сайты связывания вторичных антител, домены, связывающие металлы, эпитопные метки). В некоторых вариантах метки связаны плечами спейсера различной длины, чтобы уменьшить возможные стерические помехи. Анти-IGF-IR-антитело также можно метить радиоактивно меченной аминокислотой. Радиоактивную метку можно использовать как для диагностических, так и терапевтических целей. Например, радиоактивную метку можно использовать для детекции опухолей, экспрессирующих IGF-IR, с помощью рентгеновского излучения или другими диагностическими способами. Кроме того, радиоактивную метку можно использовать терапевтически в качестве токсина для злокачественных клеток или опухолей. Примеры меток для полипептидов включают без ограничения следующие радиоизотопы или радионуклиды: 3 Н, 14 С, 15 N, 35 S, 90 Y, 99 Tc, 111 In, 125 I, 131 I. Анти-IGF-IR-антитело также можно дериватизирвоать химической группой, такой как полиэтиленгликоль (ПЭГ), метильная или этильная группа или углеводная группа. Указанные группы можно применять для улучшения биологических характеристик антитела, например для увеличения времени полужизни в сыворотке или для увеличения связывания в ткани. Изобретение также относится к фармацевтической композиции для лечения гиперпролиферативного нарушения у млекопитающего, которая содержит терапевтически эффективное количество соединения согласно изобретению и фармацевтически приемлемый носитель. В одном варианте указанная фармацевтическая композиция предназначена для лечения злокачественной опухоли, такой как рак головного мозга, легкого, сквамозных клеток, мочевого пузыря, желудка, поджелудочной железы, молочной железы, головы, шеи, почек, яичника, простаты, ободочной и прямой кишки, пищевода, женских половых органов или щитовидной железы. В другом варианте указанная фармакологическая композиция имеет отношение к незлокачественным гиперпролиферативным нарушениям без ограничения, таким как рестеноз после ангиопластики и псориаз. В другом варианте изобретение относится к фармацевтическим композициям для лечения млекопитающего, которому требуется активация IGF-IR, при этом фармацевтическая композиция содержит терапевтически эффективное количество активирующего антитела согласно изобретению и фармацевтически приемлемый носитель. Фармацевтические композиции, содержащие активирующие антитела, можно использовать для лечения животных, у которых отсутствует достаточное количество IGF-I или IGF-II, или можно использовать для лечения остеопороза, ломкости или расстройств, при которых млекопитающее секретирует слишком мало активного гормона роста или не способно отвечать на гормон роста. Анти-IGF-IR-антитела согласно изобретению можно включать в фармацевтические композиции, подходящие для введения субъекту. Обычно фармацевтическая композиция содержит антитело согласно изобретению и фармацевтически приемлемый носитель. В используемом в данном описании смысле "фармацевтически приемлемый носитель" включает любой и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотоничные и замедляющие всасывание агенты и т.п., которые являются физиологически совместимыми. Примеры фармацевтически приемлемых носителей включают один или более из следующих носителей: воду, физиологический раствор, фосфатно-солевой буфер, декстрозу, глицерин, этанол и т.п., а также их комбинации. Во многих случаях будет предпочтительно включать в композицию агенты для изотоничности, например сахара, полиспирты, такие как маннит, сорбит, или хлорид натрия. Используют фармацевтически приемлемые вещества, такие как увлажняющие или небольшие количества вспомогательных веществ, таких как увлажняющие или эмульгирующие агенты, консерванты или буферы, которые увеличивают срок хранения или эффективность антитела или части антитела. Композиции согласно данному изобретению могут иметь множество форм. Формы включают, например, жидкие, полутвердые и твердые дозированные формы, такие как жидкие растворы (например, растворы, пригодные для инъекций и инфузий), дисперсии или суспензии, таблетки, пилюли, порошки, липосомы и суппозитории. Предпочтительная форма зависит от планируемого способа введения и терапевтического применения. Обычные предпочтительные композиции имеют форму пригодных для инъекций или инфузий растворов, такие как композиции, подобные композициям, используемым для пассивной иммунизации людей другими антителами. Предпочтительным способом введения является парентеральное (например, внутривенное, подкожное, внутрибрюшинное, внутримышечное). В предпочтительном варианте антитело вводят посредством внутривенной инфузии или инъекции. В другом предпочтительном варианте антитело вводят посредством внутримышечной или подкожной инъекции. Терапевтические композиции обычно должны быть стерильными и стабильными в условиях производства и хранения. Композиция может быть приготовлена в виде раствора, микроэмульсии, дисперсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Стерильные инъекционные растворы можно приготовить введением анти-IGF-IR-антитела в требуемом количестве в соответствующий растворитель при необходимости с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, с последующей стерилизацией фильтрованием. Как правило, дисперсии готовят введением активного соединения в стерильный наполнитель, который содержит основную дисперсионную среду и необходимые другие ингредиенты из ингредиентов, перечисленных выше. В случае стерильных порошков для приготовления стерильных инъекционных растворов предпочтительными способами приготовления являются вакуумная сушка и сушка вымораживанием, которые дают порошок активного ингредиента плюс любой требуемый дополнительный ингредиент из его предварительно простерилизованного фильтрованием раствора. Подходящую текучесть раствора можно поддерживать, например, посредством использования покрытия, такого как лецитин, посредством поддержания требуемого размера частиц в случае дисперсии и благодаря использованию поверхностно-активных веществ. Длительное всасывание инъекционных композиций можно вызвать посредством включения в композицию агента, который замедляет всасывание, например солей моностеаратов и желатина. Антитела согласно данному изобретению можно вводить множеством способов, известных в данной области, хотя для многих терапевтических применений предпочтительным путем/способом введения является внутрибрюшинный, подкожный, внутримышечный, внутривенный способ или инфузия. Специалисту в данной области будет понятно, что путь и/или способ введения будет варьировать в зависимости от требуемых результатов. В одном варианте антитела согласно данному изобретению можно вводить в виде однократной дозы или можно вводить в виде многократных доз. В некоторых вариантах активное соединение можно приготовить с носителем, который будет защищать соединение от быстрого высвобождения, а именно в виде композиции контролируемого высвобождения, включая имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Можно использовать биодеградируемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Многие способы приготовления таких композиций запатентованы и, как правило, известны специалистам в данной области. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978. В некоторых вариантах анти-IGF-IR согласно изобретению можно вводить перорально, например, с инертным разбавителем или усваиваемым съедобным носителем. Соединение (и при необходимости другие ингредиенты) также можно заключать в желатиновую капсулу с твердой или мягкой оболочкой, прессовать в таблетки или непосредственно включать в питание субъекта. Для перорального терапевтического введения соединения можно включать вместе с эксципиентами и использовать в форме таблеток для глотания, буккальных таблеток, пилюль, капсул, эликсиров, суспензий, сиропов, пластинок и т.п.. Для того, чтобы соединение согласно данному изобретению ввести другим, отличным от парентерального способом введения, может быть необходимо покрыть соединение или ввести соединение вместе с материалом, предотвращающим его инактивацию. В композиции также можно включить дополнительные активные соединения. В некоторых вариантах анти-IGF-IR согласно изобретению готовят в виде совместной композиции и/или вводят совместно с одним или более дополнительными терапевтическими средствами, такими как химиотерапевтическое средство, антинеопластическое средство или противоопухолевое средство. Например, анти-IGF-IR-антитело можно приготовить в виде совместной композиции и/или ввести совместно с одним или более дополнительными терапевтическими средствами. Указанные средства включают без ограничения антитела, которые связывают другие мишени (например, антитела, которые связывают один или более факторов роста или цитокинов, их рецепторов на клеточной поверхности или IGF-I), IGF-I-связывающие белки, антинеопластические средства, химиотерапевтические средства, противоопухолевые средства, антисмысловые олигонуклеотиды против IGF-IR или IGF-I, пептидные аналоги, которые блокируют активацию IGF-IR, растворимый IGF-IR и/или один или более химических агентов, которые ингибируют продукцию или активность IGF-I, которые известны в данной области, например октреотид. В случае фармацевтической композиции, содержащей активирующее антитело, анти-IGF-IR-антитело можно приготовить в виде композиции с фактором, который увеличивает пролиферацию клеток или предотвращает апоптоз. Такие факторы включают факторы роста, такие как IGF-I, и/или аналоги IGF-I, которые активируют IGF-IR. При такой комбинаторной терапии могут требоваться более низкие дозы анти-IGF-IR-антитела, а также совместно вводимых агентов, позволяя таким образом избежать возможной токсичности или осложнений, связанных с различными случаями монотерапии. В одном варианте вводят антитело и один или более дополнительных терапевтических средств. Фармацевтические композиции согласно изобретению могут включать "терапевтически эффективное количество" или "профилактически эффективное количество" антитела или части антитела согласно изобретению. "Терапевтически эффективное количество" относится к количеству, эффективному при используемых дозах и в течение необходимых периодов времени для достижения требуемого терапевтического результата. Терапевтически эффективное количество антитела или части антитела может варьировать в соответствии с такими факторами, как состояние заболевания, возраст, пол и масса индивидуума и способность антитела или части антитела вызывать требуемый ответ у индивидуума. Терапевтически эффективным количеством также является количество, при котором токсичные или вредные воздействия антитела или части антитела компенсируются терапевтически полезными воздействиями. "Профилактически эффективное количество" относится к количеству, эффективному при используемых дозах и в течение необходимых периодов времени для достижения требуемого профилактического результата. Как правило, в связи с тем, что профилактическую дозу используют у субъектов до заболевания или на более ранней стадии заболевания, профилактически эффективное количество будет меньше, чем терапевтически эффективное количество. Режим дозирования можно скорректировать, чтобы обеспечить оптимальное значение требуемого ответа (например, терапевтического или профилактического ответа). Например, можно вводить однократный болюс, можно вводить несколько отдельных доз в течение периода времени или дозу можно пропорционально уменьшить или увеличить, как того требует терапевтическая ситуация. Фармацевтическую композицию, содержащую антитело или содержащую комбинацию терапевтических средств, включающую антитело и одно или более дополнительных терапевтических средств, можно приготовить в виде однократной или многократных доз. Особенно предпочтительным является приготовление парентеральных композиций в дозированной лекарственной форме для простоты введения и равномерности дозирования. В используемом в данном описании смысле дозированная лекарственная форма относится к физически дискретным единицам, подходящим в качестве однократных доз для видов млекопитающих, которых необходимо лечить; при этом каждая единица содержит предварительно определенное количество активного соединения, рассчитанное для получения требуемого терапевтического действия, в ассоциации с требуемым фармацевтическим носителем. Технические характеристики дозированных форм согласно изобретению продиктованы и непосредственно зависят от: (а) уникальных характеристик активного соединения и конкретного терапевтического или профилактического действия, которое необходимо достичь, и (b) ограничений, свойственных области приготовления такого активного соединения для лечения чувствительности у индизидуумов. Особенно пригодной композицией является 5 мг/мл анти-IGF-IR-антитела в буфере, содержащем 20 мМ цитрат натрия, рН 5,5, 140 мМ NaCl и 0,2 мг/мл полисорбата 80. Примерными неограничивающими пределами терапевтически или профилактически эффективного количества антитела или части антитела согласно изобретению являются 0,1-100 мг/кг, более предпочтительно 0,5-50 мг/кг, более предпочтительно 1-20 мг/кг и еще более предпочтительно 1-10 мг/кг. Следует отметить, что значения доз могут варьировать в зависимости от типа и тяжести состояния, которое необходимо облегчить. Кроме того, понятно, что для любого конкретного субъекта конкретный режим дозирования следует корректировать в течение времени в соответствии с индивидуальной потребностью и профессиональным мнением человека, осуществляющего введение или контролирующего введение композиций, и что указанные в данном описании пределы доз являются только примерными и не предназначены для ограничения пределов или практического применения заявленной композиции. В одном варианте терапевтически или профилактически эффективное количество антитела или его антигенсвязывающей части вводят вместе с одним или более терапевтическими средствами. В другом аспекте изобретение относится к введению анти-IGF-IR-антитела для лечения рака в дозе менее 300 мг в месяц. Другой аспект данного изобретения относится к наборам, содержащим анти-IGF-IR-антитела и фармацевтические композиции, содержащие указанные антитела. Набор может включать кроме антитела или фармацевтической композиции диагностические или терапевтические средства. Набор также может содержать инструкции по применению в диагностическом или терапевтическом способе. В предпочтительном варианте набор содержит антитело или его фармацевтическую композицию и диагностическое средство, которое можно использовать в описанном далее способе. В другом предпочтительном варианте набор содержит антитело или его фармацевтическую композицию и одно или более терапевтических средств, таких как дополнительное антинеопластическое средство, противоопухолевое средство или химиотерапевтическое средство, которое можно использовать в описанном далее способе. Данное изобретение также относится к фармацевтическим композициям для ингибирования аномального роста клеток у млекопитающего, которые содержат некоторое количество соединения согласно изобретению в комбинации с некоторым количеством химиотерапевтического средства, причем количества соединения, соли, сольвата или пролекарства и химиотерапевтического средства вместе эффективны в ингибировании аномального роста клеток. В настоящее время в данной области известны многие химиотерапевтические средства. В одном варианте химиотерапевтические средства выбраны из группы, состоящей из ингибиторов митоза, алкилирующих агентов, антиметаболитов, интеркалирующих антибиотиков, ингибиторов фактора роста, ингибиторов клеточного цикла, ферментов, ингибиторов топоизомераз, средств, понижающих жизнеспособность, модификаторов биологического ответа, антигормонов, например антиандрогенов, и средств, направленных против ангиогенеза. Средства против ангиогенеза, такие как ингибиторы ММР-2 (металлопротеиназа матрикса 2), ингибиторы ММР-9 (металлопротеиназа матрикса 9) и ингибиторы ЦОГ-II (циклооксигеназа II), можно использовать вместе с соединением согласно изобретению. Примеры пригодных ингибиторов ЦОГ-II включают CELEBREX ™ (алекоксиб), вальдекоксиб и рофекоксиб. Примеры пригодных ингибиторов металлопротеиназ матрикса описаны в WO 96/33172 (опубликована 24 октября 1996), WO 96/27583 (опубликована 7 марта 1996), европейской патентной заявке № 97304971.1 (подана 8 июля 1997), европейской патентной заявке № 99308617.2 (подана 29 октября 1999), WO 98/07697 (опубликована 26 февраля 1998), WO 98/03516 (опубликована 29 января 1998), WO 98/34918 (опубликована 13 августа 1998), WO 98/34915 (опубликована 13 августа 1998), WO 98/33768 (опубликована 6 августа 1998), WO 98/30566 (опубликована 16 июля 1998), европейской патентной публикации 606046 (опубликована 13 июля 1994), европейской патентной публикации 931788 (опубликована 28 июля 1999), WO 90/05719 (опубликована 31 мая 1990), WO 99/52910 (опубликована 21 октября 1999), WO 99/52889 (опубликована 21 октября 1999), WO 99/29667 (опубликована 17 июня 1999), международной заявке РСТ № PCT/IB98/01113 (поданной 21 июля 1998), европейской патентной заявке № 99302232.1 (поданной 25 марта 1999), патентной заявке Великобритании № 9912961.1 (поданной 3 июня 1999), предварительной патентной заявке США № 60/148464 (поданной 12 августа 1999), патенте США 5863949 (выданном 26 января 1999), патенте США 5861510 (выданном 19 января 1999) и европейской патентной публикации 780386 (опубликованной 25 июня 1997), которые все включены в данное описание в виде ссылки в полном объеме. Предпочтительными ингибиторами ММР являются ингибиторы, которые не вызывают артралгии. Более предпочтительными являются ингибиторы, которые селективно ингибируют ММР-2 и/или ММР-9 по отношению к другим металлопротеиназам матрикса (т.е. ММР-1, ММР-3, ММР-4, ММР-5, ММР-6, ММР-7, ММР-8, ММР-10, ММР-11, ММР-12 и ММР-13). Некоторыми конкретными примерами ингибиторов ММР, пригодных в данном изобретении, являются AG-3340, RO 32-3555, RS 13-0830 и соединения, перечисленные в следующем списке: 3-[[4-(4-фторфенокси)бензолсульфонил]-(1-гидроксикарбамоилциклопентил)амино]пропионовая кислота; гидроксиамид 3-экзо-3-[4-(4-фторфенокси)бензолсульфониламино]-8-окса-бицикло[3,2,1]октан-3-карбоновой кислоты; гидроксиамид (2R,3R)-1-[4-(2-хлор-4-фторбензилокси)бензолсульфонил]-3-гидрокси-3-метилпиперидин-2-карбоновой кислоты; гидроксиамид 4-[4-(4-фторфенокси)бензолсульфониламино]тетрагидропиран-4-карбоновой кислоты; 3-[[4-(4-фторфенокси)бензолсульфонил]-(1-гидроксикарбамоилциклобутил)амино]пропионовая кислота; гидроксиамид 4-[4-(4-хлорфенокси)бензолсульфониламино]тетрагидропиран-4-карбоновой кислоты; гидроксиамид (R)-3-[4(4-хлорфенокси)бензолсульфониламино]тетрагидропиран-3-карбоновой кислоты; гидроксиамид (2R,3R)-1-[4-(4-фтор-2-метилбензилокси)бензолсульфонил]-3-гидрокси-3-метилпиперидин-2-карбоновой кислоты; 3-[[4-(4-фторфенокси)бензолсульфонил]-(1-гидроксикарбамоил-1-метилэтил)амино]пропионовая кислота; 3-[[4-(4-фторфенокси)бензолсульфонил]-(4-гидроксикарбамоилтетрагидропиран-4-ил)амино]пропионовая кислота; гидроксиамид 3-экзо-3-[4-(4-хлорфенокси)бензолсульфониламино]-8-оксаицикло[3,2,1]октан-3-карбоновой кислоты; гидроксиамид 3-эндо-3-[4-(4-фторфенокси)бензолсульфониламино]-8-оксаицикло[3,2,1]октан-3-карбоновой кислоты; и гидроксиамид (R)-3-[4-(4-фторфенокси)бензолсульфониламино]тетрагидрофуран-3-карбоновой кислоты; и фармацевтически приемлемые соли и сольваты указанных соединений. Соединение согласно изобретению также можно использовать вместе с ингибиторами сигнальной трансдукции, такими как агенты, которые могут ингибировать ответы EGF-R (рецептор эпидермального фактора роста), такие как EGF-R-антитела, EGF-антитела и молекулы, которые являются ингибиторами EGF-R; ингибиторами VEGF (васкулярный эндотелиальный фактор роста), такими как рецепторы VEGF и молекулы, которые могут ингибировать; и ингибиторами рецептора erbB2, такими как органические молекулы или антитела, которые связываются с рецептором erbB2, например HERCEPTIN ™ (Genentech, Inc.). Ингибиторы EGF-R описаны, например, в WO 95/19970 (опубликованной 27 июля 1995), WO 98/14451 (опубликованной 9 апреля 1998), WO 98/02434 (опубликованной 22 января 1998) и патенте США 5747498 (выданном 5 мая 1998), и такие вещества можно использовать в данном изобретении, как описано в данной заявке. EGFR-ингибирующие агенты включают, но не ограничены указанным, моноклональные антитела С225 и анти-EGFR 22 мАт (ImClone Systems Incorporated), ABX-EGF (Abgenix/Cell Genesys), EMD-7200 (Merck KgaA), EMD-5590 (Merck KgaA), MDX-447/H-477 (Medarex Inc. и Merck KgaA) и соединения ZD-1834, ZD-1838 и ZD-1839 (AstraZeneca), PKI-166 (Novartis), PKI-166/CGP-75166 (Novartis), PTK 787 (Novartis), CP 701 (Cephalon), лефлюномид (Pharmacia/Sugen), CI-1033 (Warner Lambert Parke Davis), CI-1033/PD 183805 (Warner Lambert Parke Davis), CL-387785 (Wyeth-Ayerst), BBR-1611 (Boehringer Mannheim GmbH/Roche), наамидин А (Bristol Myers Squibb), RC-3940-II (Pharmacia), BIBX-1382 (Boehringer Ingelheim), OLX-103 (Merck and Co.), VRCTC-310 (Ventech Research), токсин, слитый с EGF (Seragen Inc.), DAB-389 (Seragen/Lilgand), ZM-252808 (Imperial Cancer Research Fund), RG-50864 (INSERM), LFM-A12 (Parker Hughes Cancer Center), WHI-P97 (Parker Hughes Cancer Center), GW-282974 (Glaxo), KT-8391 (Kyowa Hakko) и вакцина EGF-R (York Medical/Centro de Immunologia Molecular (CIM)). В данном изобретении можно использовать указанные и другие EGF-R-ингибируюшие агенты. Ингибиторы VEGF, например SU-5416 и SU-6668 (Sugen Inc.), SH-268 (Scbering) и NX-1838 (NeXstar) также можно комбинировать с соединением согласно данному изобретению. Ингибиторы VEGF описаны, например, в WO 99/24440 (опубликованной 20 мая 1999), международной РСТ-заявке PCT/IB99/00797 (поданной 3 мая 1999), WO 95/21613 (опубликованной 17 августа 1995), WO 99/61422 (опубликованной 2 декабря 1999), патенте США 5834504 (выданном 10 ноября 1998), WO 98/50356 (опубликованной 12 ноября 1998), патенте США 5883113 (выданном 16 марта 1999), патенте США 5886020 (выданном 23 марта 1999), патенте США 5792783 (выданном 11 августа 1998), WO 99/10349 (опубликованной 4 марта 1999), WO 97/32856 (опубликованной 12 сентября 1997), WO 97/22596 (опубликованной 26 июня 1997), WO 98/54093 (опубликованной 3 декабря 1998), WO 98/02438 (опубликованной 22 января 1998), WO 99/16755 (опубликованной 8 апреля 1999) и WO 98/02437 (опубликованной 22 января 1998), и все указанные публикации включены в данное описание в виде ссылки в полном объеме. Другими примерами некоторых конкретных ингибиторов VEGF, применимых в данном изобретении, являются IM862 (Cytran Inc.); моноклональное анти-VEGF-антитело Genentech, Inc.; и ангиозим, синтетический рибозим из Ribozyme and Chiron. Указанные и другие ингибиторы VEGF можно использовать в данном изобретении, как описано в данной заявке. Кроме того, с соединением согласно данному изобретению можно комбинировать ингибиторы рецептора erbB2, такие как GW-282974 (Glaxo Wellcome plc) и моноклональные антитела AR-209 (Aronex Pharmaceuticals Inc.) и 2В-1 (Chiron), например такие, которые указаны в WO 98/02434 (опубликованной 22 января 1998), WO 99/35146 (опубликованной 15 июля 1999), WO 99/35132 (опубликованной 15 июля 1999), WO 98/02437 (опубликованной 22 января 1998), WO 97/13760 (опубликованной 17 апреля 1997), WO 95/19970 (опубликованной 27 июля 1995), патенте США 5587458 (выданном 24 декабря 1996), и патенте США 5877305 (выданном 2 марта 1999), и все указанные публикации включены в данное описание в виде ссылки в полном объеме. Ингибиторы рецептора erbB2, применимые в данном изобретении, также описаны в предварительной патентной заявке США № 60/117341, поданной 27 января 1999, предварительной патентной заявке США № 60/117346, поданной 27 января 1999, которые включены в данное описание в полном объеме в виде ссылки. Согласно данному изобретению соединения ингибиторов рецептора erbB2 и вещества, описанные в вышеуказанных заявках РСТ, патентах США и предварительных патентных заявках США, а также другие соединения и вещества, которые ингибируют рецептор erbB2, можно использовать с соединением согласно данному изобретению. Агенты, понижающие жизнеспособность, включают анти-IGF-IR-антитела и антиинтегриновые агенты, такие как антиинтегриновые антитела. Анти-IGF-IR-антитела можно использовать для выявления IGF-IR в биологическом образце in vitro или in vivo. Анти-IGF-IR-антитела можно использовать в обычном иммуноанализе, включая без ограничения ELISA, РИА, FACS, иммуногистохимию тканей, Вестерн-блот или иммунопреципитацию. Анти-IGF-IR-антитела согласно изобретению можно использовать для выявления IGF-IR человека. В другом варианте анти-IGF-IR-антитела можно использовать для выявления IGF-IR приматов Старого Света, таких как макаки-крабоеды и макаки-резус, шимпанзе и человекообразные обезьяны. Изобретение относится к способу выявления анти-IGF-IR в биологическом образце, включающему осуществление контакта биологического образца с анти-IGF-IR-антителом согласно изобретению и выявление связанного антитела, связанного с анти-IGF-IR для выявления IGF-IR в биологическом образце. В одном варианте анти-IGF-IR-антитело непосредственно метят детектируемой меткой. В другом варианте анти-IGF-IR-антитело (первое антитело) не метят, а метят второе антитело или другую молекулу, которая может связывать анти-IGF-IR-антитело. Как хорошо известно специалисту в данной области, в качестве второго антитела выбирают антитело, которое способно специфично связывать конкретный вид и класс первого антитела. Например, если анти-IGF-IR-антитело является IgG человека, то вторичным антителом может быть антитело против IgG человека. Другие молекулы, которые могут связываться с антителами, включают без ограничения белок А и белок G, и оба указанных белка коммерчески доступны, например, из Pierce Chemical Co. Подходящие метки для антитела или вторичного антитела описаны выше и включают различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, магнитные агенты и радиоактивные вещества. Примеры подходящих ферментов включают пероксидазу хрена, щелочную фосфатазу, β-галактозидазу или ацетилхолинэстеразу; примеры подходящих комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных веществ включают умбеллиферон, флуоресцеин, изотиоцианат флуоресцеина, родамин, дихлортриазиниламин-флуоресцеин, дансилхлорид или фикоэритрин; примером люминесцентного вещества является люминол; примером магнитного агента является гадолиний; и примеры подходящих радиоактивных материалов включают 125 I, 131 I, 35 S или 3 Н. В альтернативном варианте можно проводить анализ IGF-IR в биологическом образце посредством конкурентного иммуноанализа, используя стандарты IGF-IR, меченные детектируемым веществом, и немеченое анти-IGF-IR-антитело. В данном анализе смешивают биологический образец, меченые стандарты IGF-IR и анти-IGF-IR-антитело и определяют количество меченого стандарта IGF-IR, связанного с немеченым антителом. Количество IGF-IR в биологическом образце обратно пропорционально количеству меченого стандарта IGF-IR, связанного с анти-IGF-IR-антителом. Описанный выше иммуноанализ можно использовать для ряда целей. В одном варианте анти-IGF-IR-антитела можно использовать для выявления IGF-IR в клетках в культуре клеток. В предпочтительном варианте анти-IGF-IR-антитела можно использовать для определения уровня фосфорилирования тирозина, аутофосфорилирования тирозина IGF-IR и/или количества IGF-IR на поверхности клеток после обработки клеток различными соединениями. Указанный способ можно использовать для тестирования соединений, которые можно применять для того, чтобы активировать или ингибировать IGF-IR. В указанном способе один образец клеток обрабатывают тестируемым соединением в течение некоторого периода времени, тогда как другой образец оставляют необработанным. Если необходимо измерить аутофосфорилирование тирозина, клетки лизируют и фосфорилирование тирозина IGF-IR измеряют с использованием описанного выше иммуноанализа или как описано в примере 3, где используют ELISA. Если необходимо измерить общий уровень IGF-IR, клетки лизируют и общий уровень IGF-IR измеряют, используя один из иммуноанализов, описанных выше. Предпочтительным иммуноанализом для определения фосфорилирования тирозина IGF-IR или для измерения общих уровней IGF-IR является ELISA или Вестерн-блот. Если необходимо измерить только уровень IGF-IR на поверхности клеток, клетки не лизируют и уровни IGF-IR на поверхности клеток измеряют, используя один из иммуноанализов, описанных выше. Предпочтительный иммуноанализ для определения уровней IGF-IR на поверхности клеток включает стадии мечения белков клеточной поверхности детектируемой меткой, такой как биотин или 125 I, иммунопреципитации IGF-IR с помощью анти-IGF-IR-антитела и затем детекции меченого IGF-IR. Другим предпочтительным иммуноанализом для определения локализации IGF-IR, например уровней на поверхности клеток, является анализ с использованием иммуногистохимии. Такие способы, как ELISA, РИА, Вестерн-блот, иммуногистохимия, мечение на поверхности клеток интегральных мембранных белков и иммунопреципитация, хорошо известны в данной области. См., например, Harlow and Lane, выше. Кроме того, иммуноанализ можно проводить в большем масштабе для высокопроизводительного скрининга для того, чтобы тестировать большое количество соединений либо в отношении активации, либо ингибирования IGF-IR. Анти-IGF-IR-антитела согласно изобретению также можно использовать для определения уровней IGF-IR в ткани или в клетках, полученных из ткани. В предпочтительном варианте тканью является пораженная ткань. В более предпочтительном варианте тканью является опухоль или биопсия опухоли. В предпочтительном варианте способа ткань или биопсию ткани извлекают из организма пациента. Затем ткань или биопсию используют в иммуноанализе, чтобы определить, например, уровни IGF-IR, уровни IGF-IR на поверхности клеток, уровни фосфорилирования тирозина IGF-IR или локализацию IGF-IR способами, которые обсуждались выше. Способ можно использовать для определения того, экспрессирует ли опухоль IGF-IR на высоком уровне. Описанный выше диагностический способ можно использовать для определения того, экспрессирует ли опухоль высокие уровни IGF-IR, которые свидетельствуют о том, что опухоль будет хорошо отвечать на лечение анти-IGF-IR-антителом. Диагностический способ также можно использовать для определения того, является ли опухоль потенциально канцерогенной в том случае, если она экспрессирует высокие уровни IGF-IR, или доброкачественной в том случае, если она экспрессирует низкие уровни IGF-IR. Кроме того, диагностический способ также можно использовать для определения того, приводит ли лечение анти-IGF-IR-антителом (см. ниже) к тому, что опухоль экспрессирует более низкие уровни IGF-IR и/или экспрессирует более низкие уровни аутофосфорилирования тирозина и, следовательно, антитело можно использовать для определения того, будет ли лечение успешным. В общем, способ определения того, снижает ли анти-IGF-IR-антитело фосфорилирование тирозина, включает стадии измерения уровня фосфорилирования тирозина в представляющей интерес клетке или ткани, инкубации клетки или ткани с анти-IGF-IR-антителом или его антигенсвязывающей частью, затем повторное измерение уровня фосфорилирования тирозина в клетке или ткани. Фосфорилирование тирозина IGF-IR или другого белка(ков) можно измерить. Диагностический способ также можно использовать для определения того, что ткань или клетка не экспрессирует достаточно высокие уровни IGF-IR или достаточно высокие уровни активированного IGF-IR, что может быть причиной карликовости, остеопороза или диабета у индивидуума. Диагноз, свидетельствующий о том, что уровни IGF-IR или активного IGF-IR слишком малы, можно использовать для лечения активирующими анти-IGF-IR-антителами, IGF-I или другими терапевтическими средствами для повышения уровней или активности IGF-IR. Антитела согласно данному изобретению также можно использовать при локализации in vivo тканей и органов, которые экспрессируют IGF-IR. В предпочтительном варианте анти-IGF-IR-антитела можно использовать для локализации опухолей, экспрессирующих IGF-IR. Преимущество анти-IGF-IR-антител согласно данному изобретению состоит в том, что они не будут вызывать иммунный ответ при введении. Способ включает стадию введения анти-IGF-IR-антитела или его фармацевтической композиции пациенту, нуждающемуся в таком диагностическом тестировании, и стадию, на которой пациента подвергают анализу для визуального определения локализации тканей, экспрессирующих IGF-IR. Визуализационный анализ хорошо известен в области медицины и включает без ограничения рентгеновский анализ, визуализацию на основе магнитного резонанса (MRI) или компьютерную томографию (СЕ). В другом варианте способа пациента не подвергают визуализационному анализу, а получают от пациента биопсию, чтобы определить, экспрессирует ли представляющая интерес ткань IGF-IR. В предпочтительном варианте анти-IGF-IR-антитела можно метить детектируемым агентом, который можно визуализировать в организме пациента. Например, антитело можно метить контрастным веществом, таким как барий, который можно использовать для рентгеновского анализа, или магнитным контрастным веществом, таким как хелат гадолиния, который можно использовать для MRI или СЕ. Другие агенты для мечения включают без ограничения радиоизотопы, такие как 99 Тс. В другом варианте анти-IGF-IR-антитело будет немеченым и его будут визуализировать посредством введения второго антитела или другой детектируемой молекулы, которая может связывать анти-IGF-IR-антитело. В другом варианте изобретение относится к способу ингибирования активности IGF-IR посредством введения анти-IGF-IR-антитела нуждающемуся в этом пациенту. Терапевтически можно использовать любой из описанных в данной заявке типов антител. В предпочтительном варианте анти-IGF-IR-антитело является антителом человека, химерным или гуманизированным антителом. В другом предпочтительном варианте IGF-IR является рецептором человека, а пациентом является пациент-человек. Альтернативно пациентом может быть млекопитающее, которое экспрессирует IGF-IR, с которым перекрестно реагирует анти-IGF-IR-антитело. Антитело можно вводить млекопитающему, отличному от человека, экспрессирующему IGF-IR, с которым антитело перекрестно реагирует (т.е. примату или макаке-крабоеду или макаке-резус) для ветеринарных целей или в качестве животной модели заболевания человека. Такие модели на животных могут быть пригодны для оценки терапевтической эффективности антител согласно данному изобретению. В используемом в данном описании смысле подразумевается, что термин "расстройство, при котором активность IGF-IR вредна" включает заболевания и другие расстройства, при которых показано, что наличие высоких уровней IGF-IR у субъекта, страдающего от расстройства, является или предположительно является либо ответственным за патофизиологию расстройства, либо фактором, который участвует в обострении расстройства. Таким образом, заболеванием, при котором вредны высокие уровни активности IGF-IR, является заболевание, при котором предполагается, что ингибирование активности IGF-IR уменьшит симптомы и/или прогрессирование расстройства. Признаками таких расстройств могут быть, например, увеличение уровней IGF-IR на поверхности клеток или повышенное аутофосфорилирование тирозина IGF-IR в пораженных клетках или тканях субъекта, страдающего расстройством. Повышение уровней IGF-IR можно, например, выявить с использованием анти-IGF-IR-антитела, как описано выше. В предпочтительном варианте анти-IGF-IR-антитело можно вводить пациенту, у которого имеется опухоль, экспрессирующая IGF-IR. Опухоль может представлять собой солидную опухоль или может быть несолидной опухолью, такой как лимфома. В более предпочтительном варианте анти-IGF-IR-антитело можно вводить пациенту, у которого имеется опухоль, экспрессирующая IGF-IR, которая является злокачественной. В еще более предпочтительном варианте анти-IGF-IR-антитело вводят пациенту, у которого имеется опухоль легкого, молочной железы, простаты или ободочной кишки. В высокопредпочтительном варианте способ приводит к тому, что опухоль не увеличивается по массе или объему или уменьшается по массе или объему. В другом варианте способ вызывает интернализацию IGF-IR на опухоли. В предпочтительном варианте антитело выбрано из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 или 6.1.1 или содержит их тяжелую цепь, легкую цепь или антигенсвязывающую область. В другом предпочтительном варианте анти-IGF-IR-антитело можно вводить пациенту, у которого экспрессируются несоответствующие высокие уровни IGF-J. В данной области известно, что экспрессия IGF-I на высоком уровне может приводить к ряду распространенных злокачественных опухолей. В более предпочтительном варианте анти-IGF-IR-антитело вводят пациенту с раком простаты, глиомой или фибросаркомой. В еще более предпочтительном варианте способ приводит к остановке аномальной пролиферации злокачественной опухоли или приводит к тому, что опухоль не увеличивается по массе или объему или уменьшается по массе или объему. В одном варианте указанный способ относится к лечению рака, такого как рак головного мозга, сквамозных клеток, мочевого пузыря, желудка, поджелудочной железы, молочной железы, головы, шеи, пищевода, простаты, ободочной и прямой кишки, легкого, почек, яичника, женских половых органов или щитовидной железы. Пациенты, которых можно лечить соединениями согласно изобретению в соответствии со способами согласно данному изобретению, включают, например, пациентов, у которых диагностировали наличие рака легкого, рака кости, рака поджелудочной железы, рака кожи, рака головы и шеи, кожной или внутриглазной меланомы, рака матки, рака яичника, рака прямой кишки, рака анальной области, рака желудка, рака ободочной кишки, рака молочной железы, опухолей женских половых органов (например, маточных сарком, карциномы фаллопиевых труб, карциномы эндометрия, карциномы шейки матки, карциномы влагалища или карциномы вульвы), болезни Ходжкина, рака пищевода, рака тонкого кишечника, рака эндокринной системы (например, рака щитовидной, паращитовидной или надпочечниковой желез), сарком мягких тканей, рака мочеиспускательного канала, рака пениса, рака простаты, хронического или острого лейкоза, солидных опухолей в детском возрасте, лимфоцитарных лимфом, рака мочевого пузыря, рака почки или мочеточника (например, карциномы клеток почек, карциномы почечной лоханки) или неоплазмы центральной нервной системы (например, первичной лимфомы ЦНС, опухоли спинного мозга, глиом ствола головного мозга или аденом гипофиза). Антитело можно вводить однократно, но более предпочтительно его вводят многократно. Антитело можно вводить от 3 раз в сутки до 1 раза каждые 6 месяцев. Схема введения может быть такой, как 3 раза в сутки, 2 раза в сутки, 1 раз в сутки, 1 раз каждые 2 дня, 1 раз каждые 3 дня, 1 раз еженедельно, 1 раз каждые 2 недели, 1 раз ежемесячно, 1 раз каждые 2 месяца, 1 раз каждые 3 месяца и 1 раз каждые 6 месяцев. Антитело можно вводить посредством перорального, мукозального, буккального, интраназального, ингаляционного, внутривенного, подкожного, внутримышечного, парентерального, внутриопухолевого или местного пути. Антитело можно вводить в место, удаленное от места опухоли. Антитело также можно вводить непрерывно через мини-насос. Антитело можно вводить однократно, по меньшей мере двукратно или по меньшей мере в течение периода времени вплоть до того, как состояние подвергают лечению, облегчают или излечивают. Как правило, антитело будет вводиться до тех пор, пока будет присутствовать опухоль, при условии, что антитело вызывает остановку роста опухоли или злокачественной опухоли или уменьшает ее массу или объем. Как правило, антитело будет вводиться в виде части фармацевтической композиции, которая описана выше. Доза антитела, как правило, будет в пределах 0,1-100 мг/кг, более предпочтительно 0,5-50 мг/кг, более предпочтительно 1-20 мг/кг и еще более предпочтительно 1-10 мг/кг. Концентрацию антитела в сыворотке можно измерить любым способом, известным в данной области. См., например, пример 17 далее. Антитело также можно вводить профилактически, чтобы предотвратить возникновение рака или опухоли. Особенно полезным это может быть для пациентов, которые имеют "высокий уровень IGF-I в норме", так как показано, что для таких пациентов существует более высокая степень риска развития обычно встречающихся раков. См. Rosen et al., выше. В другом аспекте анти-IGF-IR-антитело можно вводить совместно с другими терапевтическими средствами, такими как антинеопластические лекарственные средства или молекулы, пациенту с гиперпролиферативным расстройством, таким как злокачественная опухоль или опухоль. В одном аспекте изобретение относится к способу лечения гиперпролиферативного расстройства у млекопитающего, включающему введение указанному млекопитающему терапевтически эффективного количества соединения согласно изобретению в комбинации с противоопухолевым агентом, выбранным из группы, состоящей из ингибиторов митоза, алкилирующих агентов, антиметаболитов, интеркалирующих агентов, ингибиторов фактора роста, ингибиторов клеточного цикла, ферментов, ингибиторов топоизомераз, модификаторов биологического ответа, антигормонов, ингибиторов киназ, ингибиторов металлопротеаз матрикса, генетических терапевтических средств и антиандрогенов, но не ограниченных указанным. В более предпочтительном варианте антитело можно вводить с антинеопластическим средством, таким как адриамицин или таксол. В другом предпочтительном варианте антитело или комбинацию терапевтических средств назначают наряду с лучевой терапией, химиотерапией, фотодинамической терапией, хирургией или другой иммунотерапией. Еще в одном предпочтительном варианте антитело будет вводиться с другим антителом. Например, анти-IGF-IR-антитело можно вводить с антителом или другим агентом, о котором известно, что он ингибирует пролиферацию клеток опухоли или злокачественной опухоли, например антителом или агентом, который ингибирует рецептор erbB2, EGF-R, CD20 или VEGF. Совместное введение антитела и дополнительного терапевтического средства (комбинаторная терапия) охватывает введение фармацевтической композиции, содержащей анти-IGF-IR-антитело и дополнительное терапевтическое средство, и введение двух или большего количества отдельных фармацевтических композиций, из которых одна содержит анти-IGF-IR-антитело, а другая(ие) содержит дополнительное терапевтическое средство(ва). Кроме того, хотя совместное введение или комбинаторная терапия, как правило, означает, что антитело и дополнительные терапевтические средства вводят одновременно друг с другом, она также включает в себя случаи, при которых антитело и дополнительные терапевтические средства вводят в разное время. Например, антитело можно вводить 1 раз каждые 3 дня, тогда как дополнительное терапевтическое средство вводят 1 раз ежедневно. Альтернативно, антитело можно вводить до или после лечения расстройства дополнительным терапевтическим средством. Подобным образом, введение анти-IGF-IR-антитела можно осуществлять до или после другой терапии, такой как лучевая терапия, химиотерапия, фотодинамическая терапия, хирургия или другая иммунотерапия. Антитело и одно или более дополнительных терапевтических средств (комбинаторная терапия) можно вводить однократно, двукратно или, по меньшей мере, в течение периода времени вплоть до того, как состояние подвергают лечению, облегчают или излечивают. Предпочтительно комбинаторную терапию проводят многократно. Комбинаторную терапию можно назначать от 3 раз в сутки до 1 раза каждые 6 месяцев. Схема введения может быть такой, как 3 раза в сутки, 2 раза в сутки, 1 раз в сутки, 1 раз каждые 2 дня, 1 раз каждые 3 дня, 1 раз еженедельно, 1 раз каждые 2 недели, 1 раз ежемесячно, 1 раз каждые 2 месяца, 1 раз каждые 3 месяца и 1 раз каждые 6 месяцев, или можно вводить непрерывно через мини-насос. Комбинаторную терапию можно проводить посредством перорального, мукозального, буккального, интраназального, ингаляционного, внутривенного, подкожного, внутримышечного, парентерального, внутриопухолевого или местного пути. Комбинаторную терапию можно назначать в место, удаленное от места опухоли. Как правило, комбинаторная терапия будет назначаться до тех пор, пока будет присутствовать опухоль, при условии, что антитело вызывает остановку роста опухоли или злокачественной опухоли или уменьшает ее массу или объем. Еще в одном варианте анти-IGF-IR-антитело метят радиоактивной меткой, иммунотоксином или токсином, или антитело является слитым белком, содержащим токсичный пептид. Анти-IGF-IR-антитело или слитый белок анти-IGF-IR-антитела направляет радиоактивную метку, иммунотоксин, токсин или токсичный пептид к опухолевой или злокачественной клетке, экспрессирующей IGF-IR. В предпочтительном варианте радиоактивная метка, иммунотоксин, токсин или токсичный пептид интернализуется после того, как анти-IGF-IR-антитело связывается с IGF-IR на поверхности опухолевой или злокачественной клетки. В другом аспекте анти-IGF-IR-антитело можно использовать терапевтически, чтобы индуцировать апоптоз специфичных клеток у пациента, который в этом нуждается. Во многих случаях клетками, представляющими собой мишени для апоптоза, являются канцерогенные или опухолевые клетки. Таким образом, в предпочтительном варианте изобретение относится к способу индуцирования апоптоза посредством введения терапевтически эффективного количества анти-IGF-IR-антитела нуждающемуся в этом пациенту. В предпочтительном варианте антитело выбрано из 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 или 6.1.1 или содержит их тяжелую цепь, легкую цепь или антигенсвязывающую область. В другом аспекте анти-IGF-IR-антитело можно использовать для лечения неканцерогенных состояний, при которых высокие уровни IGF-I и/или IGF-IR были связаны с неканцерогенным состоянием или заболеванием. В одном варианте способ включает стадию введения анти-IGF-IR-антитела пациенту, у которого имеется неканцерогенное патологическое состояние, вызванное или усиленное высокими уровнями IGF-I и/или IGF-IR или высокими уровнями их активности. В предпочтительном варианте неканцерогенным патологическим состоянием является акромегалия, гигантизм, псориаз, атеросклероз, рестеноз гладкой мускулатуры кровеносных сосудов или несоответствующая пролиферация микрососудов, такая как пролиферация, обнаруженная как осложнение при диабете, особенно на глаза. В более предпочтительном варианте анти-IGF-IR-антитело замедляет прогрессирование неканцерогенного патологического состояния. В более предпочтительном варианте анти-IGF-IR-антитело останавливает или реверсирует, по меньшей мере, частично неканцерогенное патологическое состояние. В другом аспекте изобретение относится к способу введения активирующего анти-IGF-IR-антитела нуждающемуся в этом пациенту. В одном варианте активирующее антитело или фармацевтическую композицию вводят нуждающемуся в этом пациенту в количестве, эффективном для того, чтобы увеличить активность IGF-IR. В более предпочтительном варианте актизирующее антитело способно восстанавливать нормальную актизность IGF-IR. В другом предпочтительном варианте активирующее антитело можно вводить пациенту, который страдает низкорослостью, невропатией, снижением мышечной массы или остеопорозом. В другом предпочтительном варианте активирующее антитело можно вводить с одним или более другими факторами, которые увеличивают пролиферацию клеток, предотвращают апоптоз или увеличивают активность IGF-IR. Такие факторы включают факторы роста, такие как IGF-I и/или аналоги IGF-I, которые активируют IGF-IR. В предпочтительном варианте антитело выбрано из 4.17.3 или содержит его тяжелую цепь, легкую цепь или антигенсвязывающую часть. Молекулы нуклеиновой кислоты согласно данному изобретению можно вводить нуждающемуся в этом пациенту с помощью генотерапии. Терапию можно проводить либо in vivo, либо ex vivo. В предпочтительном варианте пациенту вводят молекулы нуклеиновой кислоты, кодирующие как тяжелую цепь, так и легкую цепь. В более предпочтительном варианте молекулы нуклеиновой кислоты вводят так, чтобы они были стабильно интегрированы в хромосому В-клеток, поскольку указанные клетки являются специализированными клетками для продукции антител. В предпочтительном варианте В-клетки-предшественники трансфицируют или инфицируют ex vivo и снова трансплантируют нуждающемуся в этом пациенту. В другом варианте В-клетки-предшественники или другие клетки инфицируют in vivo, используя вирус, о котором известно, что он инфицирует клетки представляющего интерес типа. Обычные векторы, используемые для генотерапии, включают липосомы, плазмиды или вирусные векторы, такие как ретровирусы, аденовирусы и аденоассоциированные вирусы. После инфицирования либо in vivo, либо ex vivo уровни экспрессии антител можно контролировать посредством отбора образца у пациента, подвергаемого лечению, и используя любой иммуноанализ, известный в данной области и обсуждаемый в данном описании. В предпочтительном варианте генотерапевтический способ включает стадии введения эффективного количества выделенной молекулы нуклеиновой кислоты, кодирующей тяжелую цепь или ее антигенсвязывающую часть антитела человека или его части, и экспрессии молекулы нуклеиновой кислоты. В другом варианте генотерапевтический способ включает стадии введения эффективного количества выделенной молекулы нуклеиновой кислоты, кодирующей легкую цепь или ее антигенсвязывающую часть антитела человека или его части, и экспрессии молекулы нуклеиновой кислоты. В более предпочтительном варианте генотерапевтический способ включает стадии введения эффективного количества выделенной молекулы нуклеиновой кислоты, кодирующей тяжелую цепь или ее антигенсвязывающую часть антитела человека или его части, и эффективного количества выделенной молекулы нуклеиновой кислоты, кодирующей легкую цепь или ее антигенсвязывающую часть антитела человека или его части, и экспрессии молекул нуклеиновой кислоты. Генотерапевтический способ также может включать стадию введения другого противоракового средства, такого как таксол, тамоксифен, 5-FU, адриамицин или СР-358774. Для лучшего понимания данного изобретения приведены следующие примеры. Указанные примеры приведены только в целях иллюстрации, и их никоим образом не следует считать ограничивающими объем изобретения. Пример 1. Создание гибридом, продуцирующих анти-IGF-IR-антитело. Антитела согласно изобретению получают, отбирают и анализируют следующим образом. Ксеномышей XENOMICE ™ 8-10-недельного возраста иммунизировали внутрибрюшинно или в подушечку их задней лапы либо внеклеточным доменом IGF-IR человека (10 мкг/дозу/мышь), либо клетками 3T3-IGF-IR или 300.19-IGF-IR, которые представляют собой две трансфицированные линии клеток, которые экспрессируют IGF-IR человека на своих плазматических мембранах (10 ×10 6 клеток/дозу/мышь). Указанную дозу повторяли 5-7 раз в течение периода времени от 3 до 8 недель. За 4 дня до слияния мышам вводили последнюю инъекцию внеклеточного домена IGF-IR человека в PBS. Лимфоциты селезенки и лимфатических узлов иммунизированных мышей сливали с несекретирующей линией клеток миеломы P3-X63-Ag8.653 и подвергали селекции на HAT, как описано ранее (Galfre and Milstein, Methods Enzymol. 73: 3-46, 1981). Получали панель гибридом, каждая из которых секретировала специфичные по отношению к IGF-IR антитела IgG2 κ человека. Для дальнейшего исследования отобрали семь гибридом, продуцирующих моноклональные антитела, специфичные по отношению к IGF-IR, и обозначили 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2, 4.17.3 и 6.1.1. Гибридомы 2.12.1, 2.13.2, 2.14.3, 3.1.1, 4.9.2 и 4.17.3 депонированы в американской коллекции типов культур (АТСС), 10801 University Boulevard, Manassas, VA 20110-2209, 12 декабря 2000г. под следующими регистрационными номерами: Пример 2. Определение констант аффинности (Kd) полностью человеческих моноклональных анти-IGF-IR-антител с помощью BIAcore. Авторы выполнили измерения аффинности очищенных антител с помощью резонанса поверхностного плазмона, используя прибор BIAcore 3000, следуя протоколам производителя. Протокол 1. Для выполнения кинетических анализов белок-А иммобилизовали на поверхностях сенсорных чипов BIAcore. Затем сенсорный чип использовали для захвата анти-IGF-IR-антител согласно данному изобретению. Различные концентрации внеклеточного домена IGF-IR инъецировали на сенсорный чип и анализировали кинетики связывания и диссоциации при взаимодействиях анти-IGF-IR-антител и внеклеточного домена IGF-IR. Данные оценивали с помощью общей подгонки Ленгмюра 1:1, используя модели смещения базовой линии, имеющимися в компьютерной программе BIAevaluation, предоставляемой BIAcore. Протокол 2. Измерения BIAcore выполняли, по существу, так, как описано Fagerstam et al. "Detection of antigen-antibody interactions by surface plasmon resonance. Applications to epitope mapping." J. Mol. Recog. 3: 208-214 (1990). В табл. 1 перечислены результаты измерения аффинности для типичных анти-IGF-IR-антител согласно данному изобретению: Кинетические анализы свидетельствуют о том, что антитела, полученные согласно изобретению, обладают высокой аффинностью и значительными константами связывания для внеклеточного домена IGF-IR. Пример 3. Опосредованное антителом ингибирование индуцированного IGF-I фосфорилирования IGF-IR. Авторы изобретения проводили эксперименты ELISA для того, чтобы определить, способны ли антитела согласно данному изобретению блокировать опосредованную IGF-I активацию IGF-IR. Опосредованную IGF-I активацию IGF-IR детектировали по повышенному фосфорилированию тирозина, связанного с рецептором. Авторы изобретения готовили иммобилизирующие планшеты для ELISA добавлением 100 мкл блокирующего буфера (3% бычий сывороточный альбумин [БСА] в забуференном трисом физиологическом растворе [TBS]) в каждую лунку 96-луночных планшетов ReactiBind, покрытых белком G (Pierce), и планшеты инкубировали при встряхивании в течение 30 мин при комнатной температуре. Авторы изобретения разводили пан-специфичное анти-IGF-IR-антитело кролика SC-713 (Santa Cruz) в блокирующем буфере до концентрации 5 мкг/мл и в каждую лунку добавляли по 100 мкл разведенного антитела. Авторы изобретения инкубировали планшеты при встряхивании в течение 60-90 мин при комнатной температуре. Затем авторы изобретения 5 раз промывали планшеты буфером для промывки (TBS+0,1% твин 20) и осторожно промокали оставшийся буфер на бумажные полотенца. Указанным планшетам не позволяли высыхать до добавления лизата. Авторы изобретения помещали клетки NIH-3T3, трансфицированные IGF-IR (5 ×10 4 /мл), в 100 мкл среды роста (среда DMEM с высоким содержанием глюкозы с добавлением L-глутамина (0,29 мг/мл), 10% инактивированной нагреванием FBS и 500 мкг/мл каждого из антибиотиков - генетицина, пенициллина и стрептомицина) в 96-луночные планшеты с U-образным дном. Авторы изобретения инкубировали планшеты при 37 °С, 5% СО 2 в течение ночи, давая возможность клеткам прикрепиться. Авторы изобретения среду декантировали из планшетов и заменяли ее 100 мкл свежей среды на лунку. Для тестирования авторы изобретения разводили потенциальные анти-IGF-IR-антитела до пятикратной требуемой конечной концентрации в среде роста и добавляли 25 мкл на лунку. Все образцы готовили в трех повторах. Затем авторы изобретения инкубировали планшеты при 37 °С в течение 1 ч. Авторы изобретения стимулировали клетки 25 мкл/лунку 600 нг/мл IGF-1 (приготовленном в среде роста) и инкубировали планшеты при комнатной температуре в течение 10 мин. Затем авторы изобретения декантировали среду, переворачивая планшеты и осторожно промокая на бумажные полотенца, и лизировали адгезированные клетки добавлением 50 мкл лизирующего буфера (50 мМ HEPES, рН 7,4, 10 мМ EDTA, 150 мМ NaCl, 1,5 мМ MgCl 2 , 1,6 мМ NaVO 4 , 1% тритон Х-100, 1% глицерин при добавлении непосредственно перед использованием 1 таблетки ингибитора протеазы, не содержащего EDTA [Roche Molecular Sciences] на 50 мл) и перемешивая в течение 5 мин при комнатной температуре. В каждую лунку авторы изобретения добавляли 200 мкл буфера для разведения (50 мМ HEPES, рН 7,4, 1,6 мМ NaVO 4 ) и перемешивали пипетированием вверх и вниз. Авторы изобретения переносили 100 мкл лизата из каждой лунки в каждую лунку иммобилизирующего планшета для ELISA, приготовленного, как описано выше, и инкубировали при осторожном встряхивании в течение 2 ч при комнатной температуре. Авторы изобретения удаляли лизат клеток, переворачивая планшеты, промывали планшеты 5 раз буфером для промывки и промокали на бумажные полотенца. Авторы изобретения добавляли по 100 мкл на лунку pTYR-специфичного антитела (HRP-PY54), разведенного в блокирующем буфере до концентрации 0,2 мкг/мл, и планшеты инкубировали при встряхивании в течение 30 мин при комнатной температуре. Затем авторы изобретения 5 раз промывали указанные планшеты буфером для промывки и промокали на бумажных полотенцах. Авторы изобретения детектировали связывание HRP-PY54-антитела, добавляя по 100 мкл на лунку раствора субстрата пероксидазы ТМВ (Kirkegaard and Perry), и инкубировали со встряхиванием до окрашивания (примерно 2-10 мин). Авторы изобретения останавливали реакцию окрашивания добавлением по 100 мкл на лунку раствора для остановки ТМВ (Kirkegaard and Perry). Затем авторы изобретения встряхивали планшеты в течение 10 с при комнатной температуре, чтобы перемешать раствор, и проводили количественное определение посредством измерения OD 450 нм . В табл. 2 и на фиг. 4 показаны результаты данного эксперимента, выполненного с несколькими антителами согласно изобретению. Результаты данного эксперимента демонстрируют способность антител согласно данному изобретению блокировать опосредованную IGF-I активацию IGF-IR, о чем свидетельствует повышенное фосфорилирование тирозина, связанного с рецептором. Кроме того, полученные результаты можно использовать для количественного определения относительной эффективности антител согласно данному изобретению. Пример 4. Опосредованное антителом блокирование связывания IGF-I/IGF-IR. Авторы изобретения проводили эксперименты ELISA, чтобы количественно оценить способность антител согласно изобретению ингибировать связывание IGF-I с IGF-IR в анализе, основанном на клетках. Авторы высевали трансфицированные IGF-IR клетки NIH-3Т3 (5 ×10 4 /мл) в 100 мкл DMEM с высоким содержанием глюкозы с добавлением L-глутамина (0,29 мг/мл), 10% инактивированной нагреванием FBS и 500 мкг/мл каждого из антибиотиков генетицина, пенициллина и стрептомицина в 96-луночные планшеты с U-образным дном. Затем авторы изобретения инкубировали планшеты при 37 °С, 5% CO 2 в течение ночи, давая возможность клеткам прикрепиться. Затем авторы изобретения сливали среду из планшетов и заменяли 100 мкл свежей среды на лунку. Для тестирования авторы изобретения разбавляли антитела в среде для анализа (DMEM с высоким содержанием глюкозы с добавлением L-глутамина, 10% инактизированной нагреванием FBS, 200 мкг/мл БСА и 500 мкг/мл каждого из антибиотиков - генетицина, пенициллина и стрептомицина) до требуемой конечной концентрации и добавляли по 50 мкл на лунку. Все образцы готовили в трех повторах. Затем авторы изобретения инкубировали планшеты при 37 °С в течение 10 мин. Авторы изобретения разбавляли [ 125 I]-IGF-I до концентрации 1 мкКи/мл в среде для анализа и добавляли в планшет по 50 мкл на лунку. В качестве контроля фоновой радиоактивности авторы изобретения добавляли холодный IGF-I до конечной концентрации 100 нг/мл. Авторы изобретения инкубировали планшеты в течение 10 мин при 37 °С, среду декантировали осторожным промоканием на бумажные полотенца и дважды промывали средой для анализа. Затем авторы изобретения лизировали клетки добавлением 50 мкл 0,1н. NaOH, 0,1% SDS и встряхиванием планшетов в течение 5 мин при комнатной температуре. Затем авторы изобретения переносили образцы в планшет для сцинтилляции, добавляли 150 мкл OptiPhase Supermix и регистрировали сигнал в счетчике Wallac Micro-Beta. В табл. 3 и на фиг. 3 показаны результаты данного эксперимента, выполненного с тремя типичными антителами согласно изобретению. Данный эксперимент показывает, что антитела согласно изобретению специфично ингибируют связывание [ 125 I]-IGF-I с клетками, сверхэкспрессирующими IGF-IR. Пример 5. Исследование по картированию эпитопов. Показав, что антитела согласно изобретению распознают IGF-IR, авторы изобретения провели исследование по картированию эпитопов нескольких антител согласно изобретению. В частности, авторы изобретения сосредоточили эксперименты на антителах 2.12.1, 2.13.2, 2.14.3 и 4.9.2. Авторы изобретения провели конкурентные исследования BIAcore, чтобы определить, связываются ли антитела согласно данному изобретению с одним и тем же или с разными местами на молекуле IGF-IR. Авторы изобретения связывали внеклеточный домен (ECD) IGF-IR с сенсорным чипом BIAcore, как описано выше в примере 2. Авторы изобретения связывали первое антитело согласно изобретению с данным IGF-IR, связанным с сенсорным чипом, в условиях насыщения. Затем авторы изобретения измеряли способность следующих вторичных антител согласно изобретению конкурировать с первичным антителом за связывание с IGF-IR. Указанный способ позволил авторам изобретения определить антитела согласно данному изобретению с разными связывающими группами. Авторы изобретения проводили указанный эксперимент с антителами 2.12.1, 2.13.2, 2.14.3 и 4.9.2. Авторы изобретения отметили, что 2.13.2 и 4.9.2 конкурируют за один и тот же сайт во внеклеточном домене IGF-IR. Другие антитела 2.12.1 и 2.14.3 связываются с сайтами IGF-IR, которые отличаются друг от друга и от сайта, связываемого 2.13.2 и 4.9.2. Пример 6. Видовая перекрестная реактивность антител согласно изобретению. Для того, чтобы определить видовую перекрестную реактивность антител согласно изобретению, авторы изобретения выполнили несколько экспериментов, включая иммунопреципитацию, опосредованное антителом блокирование индуцированного IGF-I фосфорилирования рецептора и FACS-анализ. Чтобы выполнить эксперименты по иммунопреципитации, авторы изобретения высевали клетки в среду DMEM с высоким содержанием глюкозы с добавлением L-глутамина (0,29 мг/мл), 10% инактивированной нагреванием FBS и 500 мкг/мл каждого из антибиотиков - генетицина, пенициллина и стрептомицина, до слияния на 50% во флаконах Т25. Затем авторы изобретения добавляли 100 мкл антитела согласно изобретению в забуференном физиологическом растворе Хенкса (HBSS; Gibco BRL) при концентрации 1 мкг/мл. Авторы изобретения инкубировали планшеты в течение 30 мин при 37 °С в инкубаторе и затем стимулировали клетки IGF-I при концентрации 100 нг/мл в течение 10 мин при комнатной температуре. Авторы изобретения лизировали клетки в буфере RIPA (Harlow and Lane, выше) и иммунопреципитировали IGF-IR с помощью 2 мкг пан-специфичного анти-IGF-IR-антитела SC-713 (Santa Cruz) плюс агарозные шарики с белком А в течение 1 ч при 4 °С. Авторы изобретения осаждали шарики и 3 раза промывали PBS/T (PBS+0,1% твин-20) и затем кипятили шарики в 40 мкл буфера Лэммли, содержащего 5% β-МЭ. Затем образцы, приготовленные, как описано выше, анализировали с помощью Вестерн-блота. Авторы изобретения наносили 12 мкл каждого образца на дорожку на 4-10% градиентные гели Novex ™ и разгоняли в буфере 1X MES (Novex ™). Гели разгоняли при 150 В в течение 1 ч или при 200 В примерно в течение 30 мин. Затем авторы изобретения осуществляли перенос с геля на мембрану в буфере для переноса Novex ™ с 10% метанола либо в течение ночи при 100 мА, либо в течение 1-1,5 ч при 250 мА. Затем авторы изобретения давали возможность мембране полностью высохнуть и блокировали при комнатной температуре TBS (забуференный трисом физиологический раствор рН 8,0), содержащим Superblock (Pierce Chemical Co.). Авторы изобретения добавляли антитело для блоттинга IGF-IR - SC713 (Santa Cruz), чтобы выявить иммунопреципитированный IGF-IR. Указанный эксперимент выполняли с антителами согласно изобретению, в частности 2.12.1, 2.13.2, 4.17.3 и 4.9.2, на клетках из разных животных. Авторы изобретения обнаружили, что антитела 2.12.1, 2.13.2 и 4.9.2 способны связывать IGF-IR человека, но не рецептор собаки, морской свинки или кролика. Кроме того, указанные антитела способны связываться с IGF-IR COS7 и Rhesus, которые получены от обезьян Старого Света, но не с IGF-IR мартышки, которая является обезьяной Нового Света. Указанные эксперименты свидетельствуют о том, что антитела являются высокоспецифичными. После обнаружения того, что антитела согласно изобретению узнают IGF-IR обезьян Старого Света, авторы изобретения также тестировали их способность блокировать связывание IGF-I/IGF-IR в клетках, полученных из указанных обезьян Старого Света. Авторы изобретения высевали клетки в среду DMEM с высоким содержанием глюкозы с добавлением L-глутамина, 10% инактивированной нагреванием FBS и 500 мкг/мл каждого из антибиотиков - генетицина, пенициллина и стрептомицина - до слияния на 50% во флаконах Т25. Затем авторы изобретения добавляли антитело согласно изобретению или среду без антитела в качестве контроля и стимулировали клетки IGF-I при концентрации 100 нг/мл в течение 10 мин при комнатной температуре. После стимуляции авторы изобретения лизировали клетки и иммунопреципитировали IGF-IR с помощью пан-специфичного IGF-IR-антитела SC713, как описано выше. Затем авторы изобретения проводили Вестерн-блот-анализ, как описано выше, используя антитело HRP-PY54, чтобы выявить фосфорилированный тирозин в активированном IGF-IR. Авторы изобретения обнаружили, что антитела согласно данному изобретению, в частности 2.13.2 и 4.9.2, могут блокировать индуцированное IGF-I фосфорилирование IGF-IR как в клетках COS7, так и в клетках Rhesus. IC 50 для измеряемого ингибирования составляло 0,02 и 0,005 мкг/мл для IGF-IR COS7 и Rhesus, соответственно. Авторы изобретения выполняли FACS-анализ, чтобы определить аффинность антител согласно изобретению по отношению к IGF-IR разных животных, в частности обезьян Старого Света, описанных выше. Авторы изобретения инкубировали аликвоты клеток человека и обезьян (5 ×10 5 ) в течение 1 ч на льду с возрастающими концентрациями биотинилированных анти-IGF-IR-антител согласно изобретению или с биотинилирозанным антителом против гемоцианина слизня (KLH) (Abgenix) в качестве негативного контроля. Затем авторы изобретения инкубировали образцы в течение 30 мин на льду с конъюгированным со стрептавидином RPE (фикоэритрин). Авторы изобретения измеряли связывание посредством проточной цитометрии и анализировали гистограммы интенсивности флуоресценции (F12-H) против количества клеток (счет), используя компьютерную программу CellQuest. Авторы изобретения рассчитывали связывание (K d ) для каждого антитела на основе графиков средней интенсивности флуоресценции против концентрации антитела. В большинстве экспериментов авторы изобретения измеряли связывание в культивируемых клетках MCF-7 человека и клетках культуры ткани либо макак-резус, либо макак-крабоедов. Авторы изобретения определяли истощение антитела, измеряя связывание в диапазоне концентраций клеток. Авторы изобретения выполняли вышеупомянутый FACS-анализ, чтобы тестировать способность антител согласно изобретению, в частности 2.13.2 и 4.9.2, связываться с клетками человека, макак-резус и макак-крабоедов. Авторы изобретения регистрировали половину максимального связывания (K d ), составляющую 0,1 мкг/мл для всех тестированных клеточных линий. Пример 7. Понижающая регуляция рецептора IGF-I. Эксперименты по блокированию авторы изобретения выполняли, по существу, так, как описано выше в примере 4, вплоть до добавления [ 125 I]-меченного IGF-I. В указанной точке авторы изобретения кипятили клетки в 40 мкл буфера Лэммли, содержащего 50% МЭ. Затем авторы изобретения анализировали образцы посредством Вестерн-блот-анализа, как описано выше в примере 6, и зондировали блоты с помощью пан-специфичного IGF-IR-антитела SC713, чтобы количественно определить уровни IGF-IR, и с помощью HRP-PY54-антитела, чтобы определить уровни фосфорилированного тирозина в активированном IGF-IR. Как обнаружено ранее (пример 3), авторы изобретения наблюдали блокирование индуцированного IGF-I фосфорилирования IGF-IR после обработки клеток антителом согласно данному изобретению (фиг. 4). Кроме того, авторы изобретения наблюдали, что после блокирования индуцированного IGF-I фосфорилирования следовала понижающая регуляция IGF-IR в указанных клетках. См., например, фиг. 4. Уровни IGF-IR максимально снижались через 16 ч после стимуляции IGF-I в присутствии антитела согласно изобретению. Пример 8. Влияние антител согласно изобретению на IGF-IR in vivo. Авторы изобретения определяли, будет ли проявляться in vivo влияние антител согласно изобретению на IGF-IR, которое описано в предыдущих примерах. Авторы изобретения индуцировали опухоли у бестимусных мышей в соответствии с опубликованными способами (V.A. Pollack et al., "Inhibition of epidermal growth factor receptor-associated tyrosine phosphorylation in human carcinomas with CP-358774: Dynamics of receptor inhibition in situ and antitumor effects in athymic mice", J. Pharmacol. Exp. Ther. 291: 739-748 (1999). Вкратце, авторы изобретения инъецировали трансфицированные IGF-IR клетки NIH-3Т3 (5 ×10 6 ) подкожно бестимусным мышам (nu/nu) 3-4-недельного возраста с 0,2 мл препарата Matrigel. Затем авторы изобретения внутрибрюшинно инъецировали мышам антитело согласно изобретению после образования определяемых опухолей (т.е. объемом примерно 400 мм 3 ). Через 24 ч авторы изобретения извлекали опухоли, гомогенизировали их и определяли уровень IGF-IR. Чтобы определить уровни IGF-IR, авторы изобретения разбавляли антитело SC-713 в блокирующем буфере до конечной концентрации мкг/мл и добавляли по 100 мкл в каждую лунку планшета (Pierce), покрытого антикроличьими антителами козы Reacti-Bind (GAR). Авторы изобретения инкубировали планшеты при комнатной температуре в течение 1 ч при перемешивании и затем промывали планшеты 5 раз буфером для промывки. Затем авторы изобретения взвешивали образцы опухолей, которые были приготовлены, как описано выше, и гомогенизировали их в лизирующем буфере (1 мл/100 мг). Авторы изобретения разбавляли 12,5 мкл экстракта опухоли лизирующим буфером до конечного объема 100 мкл и добавляли его в каждую лунку 96-луночного планшета. Авторы изобретения инкубировали планшеты при комнатной температуре при встряхивании в течение 1-2 ч и затем 5 раз промывали планшеты буфером для промывки. Затем авторы изобретения добавляли в каждую лунку по 100 мкл HRP-PY54 или биотинилированного анти-IGF-IR-антитела в блокирующем буфере и инкубировали при комнатной температуре и встряхивании в течение 30 мин. Затем авторы изобретения промывали планшеты 5 раз буфером для промывки и проявляли планшеты. Авторы изобретения проявляли планшеты, используя в качестве зонда HRP-PY54, добавляя в лунку по 100 мкл субстрата ТМВ Microwell, и останавливали развитие окраски добавлением 100 мкл 0,9М H 2 SO 4 . Затем авторы изобретения количественно оценивали сигнал, встряхивая в течение 10 с и измеряя OD 450 нм . Сигнал нормализовали по отношению к общему белку. Авторы изобретения проявляли планшеты, используя в качестве зонда анти-IGF-IR-антитело, посредством добавления в каждую лунку 100 мкл стрептавидина-HRP, разведенного в блокирующем буфере, инкубируя при комнатной температуре и встряхивании в течение 30 мин и затем продолжая обработку, как описано для HRP-PY54. Авторы изобретения обнаружили, что внутрибрюшинная инъекция антитела согласно изобретению, в частности 2.13.2 и 4.9.2, приводила к ингибированию активности IGF-IR, измеряемому по снижению (как фосфотирозина IGF-IR (фосфорилированного IGF-IR), так и) общего белка IGF-IR (фиг. 6). Кроме того, авторы изобретения также наблюдали уменьшение количества фосфотирозина IGF-IR (фосфорилированного IGF-IR) (фиг. 5). Не имея намерения связать с какой-либо теорией, полагают, что сниженные уровни фосфотирозина IGF-IR могут быть следствием пониженных уровней белка IGF-IR in vivo после лечения антителом или могут быть следствием комбинации пониженных уровней белка IGF-IR и снижения фосфорилирования тирозина в IGF-IR, которое имеет место вследствие блокирования активации лигандом (например, IGF-I или IGF-II). Кроме того, указанное ингибирование зависит от дозы инъецированного антитела (фиг. 6). Полученные данные свидетельствуют о том, что антитела согласно изобретению способны действовать на мишень IGF-IR in vivo аналогично действию, наблюдаемому in vitro. Пример 9. Ингибирование роста (TGI) 3T3/IGF-IR-клеточных опухолей. Авторы изобретения проверяли, будут ли анти-IGF-IR-антитела согласно изобретению функционировать как ингибиторы роста опухолей. Авторы изобретения индуцировали опухоли, как описано выше (пример 8), и, когда формировались устанавливаемые пальпируемые опухоли (т.е. 250 мм 3 , в течение 6-9 дней), авторы изобретения обрабатывали мышей однократной внутрибрюшинной инъекцией антитела в дозе 0,20 мл. Авторы изобретения измеряли размер опухоли штангенциркулями по двум диаметрам каждый третий день и рассчитывали объем, используя формулу (длина ×[ширина] 2 )/2, используя способы, разработанные Geran, et al., "Protocols for screening chemical agents and natural products against animal tumors and other biological systems", Cancer Chemother. Rep. 3: 1-104. В том случае, когда авторы изобретения проводили данный анализ с антителом согласно изобретению, они обнаружили, что обработка только антителом 2.13.2 ингибировала рост опухолей, индуцированных клетками NIH-3T3, трансфицированными IGF-IR (фиг. 7 левая панель). Кроме того, при комбинированных исследованиях с использованием однократной дозы 7,5 мг/кг внутривенно доставляемого адриамицина авторы изобретения наблюдали, что введение однократной дозы 2.13.2 усиливало эффективность адриамицина, известного ингибитора опухолевого роста. При комбинировании адриамицина с антителом согласно изобретению 2.13.2 показано замедление роста на 7 дней по сравнению с лечением одним антителом или адриамицином (фиг. 7, правая панель). Пример 10. Взаимосвязь уровней антител и понижающей регуляции IGF-IR. Опухоли индуцировали у "голых" мышей, как описано в примере 8. Затем мышей обрабатывали 125 мкг 2.13.2 путем внутрибрюшинной инъекции, как описано в примере 8. Опухоли извлекали и измеряли уровни IGF-IR с помощью ELISA, как описано в примере 8. На фиг. 8 показаны уровни антитела 2.13.2 в сыворотке и уровни рецептора IGF-IR в течение времени. Эксперимент свидетельствует о том, что происходит понижающая регуляция IGF-IR антителом и что степень ингибирования IGF-IR пропорциональна по дозе концентрации антитела в сыворотке. Пример 11. Ингибирование роста опухолей 3T3/IGF-IR многократными дозами антитела в комбинации с адриамицином. Опухоли индуцировали у "голых" мышей, как описано в примере 9. Мышей с устанавливаемыми подкожными опухолями объемом примерно 250 мм 3 обрабатывали на 1, 8, 15 и 22 день различными количествами антитела 2.13.2 (в/б) или 7,5 мг/кг адриамицина (в/в) либо в виде отдельных агентов, либо в комбинации, как описано в примере 9. На фиг. 9 показан размер опухолей в зависимости от различных обработок в течение времени. Эксперимент показывает, что обработка анти-IGF-IR-антителом 1 раз каждые 7 дней ингибирует рост клеток опухоли и усиливает ингибирование роста клеток опухоли в комбинации с адриамицином, известным ингибитором опухолей. Пример 12. Ингибирование роста крупных опухолей. Опухоли индуцировали у "голых" мышей, как описано в примере 9. Мышей с крупными устанавливаемыми подкожными опухолями по объему немного меньше 2000 мм 3 обрабатывали на 1 и 8 день различными количествами антитела 2.13.2 (в/б) или 7,5 мг/кг адриамицина (в/в) либо в виде отдельных агентов, либо в комбинации, как описано в примере 9. На фиг. 10 показан размер опухолей в зависимости от различных обработок в течение времени. Группы животных в контроле с использованием только антитела и только адриамицина погибали на 5 день, когда размер опухоли превышал 2000 мм 3 . Эксперимент показывает, что обработка анти-IGF-IR-антителом в комбинации с адриамицином является высокоэффективной против крупных опухолей при введении многократных доз. Пример 13. Ингибирование роста опухолей клеток ободочной и прямой кишки. Опухоли индуцировали у "голых" мышей, как описано в примере 9, за исключением того, что использовали клетки Colo 205 (АТСС CCL 222). Клетки Colo 205 являются клетками аденокарциномы ободочной и прямой кишки человека. Мышей с устанавливаемыми подкожными опухолями объемом примерно 250 мм 3 обрабатывали различными количествами антитела 2.13.2 (в/б) или 100 мг/кг 5-фтордезоксиуридина (5-FU, в/в) либо в виде отдельных агентов, либо в комбинации, как описано в примере 9. На фиг. 11 показан размер опухоли в зависимости от различных обработок в течение времени. Эксперимент показывает, что обработка анти-IGF-IR-антителом, вводимым однократно, ингибирует рост злокачественных клеток ободочной и прямой кишки человека при введении в виде отдельного агента и усиливает эффективность 5-FU, известного ингибитора опухолей. Мышей с устанавливаемыми опухолями Colo 205 обрабатывали на 1, 8, 15 и 22 день 500 мкг 2.13.2 (в/б), 100 мг/кг 5-FU (в/в) или их комбинацией. На фиг. 12 показан размер опухолей в зависимости от различных обработок в течение времени. Эксперимент показывает, что обработка анти-IGF-IR-антителом, вводимым 1 раз каждые 7 дней, ингибирует рост клеток рака ободочной и прямой кишки человека и усиливает эффективность 5-FU. Пример 14. Ингибирование роста опухолей клеток рака молочной железы. "Голым" мышам, описанным в примере 8, имплантировали биодеградируемые таблетки эстрогена (0,72 мг 17- β-эстрадиола/таблетку, высвобождение в течение 60 дней; Innovative Research of America). Через 48 ч у "голых" мышей индуцировали опухоли, по существу, как описано в примере 9, за исключением того, что использовали клетки MCF-7 (АТСС НТВ-22). Клетки MCF-7 являются эстрогензависимыми клетками карциномы молочной железы человека. Мышей с устанавливаемыми подкожными опухолями объемом примерно 250 мм 3 обрабатывали 50 мкг антитела 2.13.2 (в/б) на 1, 4, 7, 10, 13, 16, 19 и 22 день (q3d ×7) или 6,25 мг/кг таксола (в/б) на 1, 2, 3, 4, 5 день (q1d ×5) либо в виде отдельных агентов, либо в комбинации, по существу, как описано в примере 9. На фиг. 13 показан размер опухолей в зависимости от различных обработок в течение времени. Эксперимент показывает, что обработка самим анти-IGF-IR-антителом ингибирует рост клеток рака молочной железы человека при введении 1 раз каждые 3 дня, а также усиливает эффективность таксола, известного ингибитора рака молочной железы, при введении в комбинации. Мышей, имеющих устанавливаемые опухоли из клеток MCF-7, как описано непосредственно выше, обрабатывали в 1 день различными количествами антитела 2.13.2 (в/б) отдельно или с 3,75 мг/кг адриамицина (в/в), по существу, как описано в примере 9. На фиг. 14 показан размер опухолей в зависимости от различных обработок в течение времени. Эксперимент показывает, что однократная обработка самим анти-IGF-IR-антителом ингибирует рост клеток рака молочной железы человека и усиливает эффективность адриамицина, известного ингибитора опухолей. Мышей, имеющих устанавливаемые опухоли из клеток MCF-7, как описано непосредственно выше, обрабатывали 250 мкг антитела 2.13.2 (в/б) в 1, 8, 15 и 23 день или биодеградируемой таблеткой тамоксифена (25 мг/таблетку, свободное основание, высвобождение в течение 60 дней, Innovative Research of America) либо в виде отдельных агентов, либо в комбинации, по существу, как описано в примере 9. Таблетку тамоксифена имплантировали в 1 день после установления опухоли. На фиг. 15 показан размер опухоли в зависимости от различных обработок в течение времени. Эксперимент показывает, что обработка анти-IGF-IR-антителом, вводимым 1 раз каждые 7 дней, ингибирует рост злокачественной опухоли молочной железы человека сам по себе и усиливает эффективность тамоксифена, известного ингибитора опухолей. Пример 16. Ингибирование роста опухолей клеток эпидермоидной карциномы. Опухоли индуцировали у "голых" мышей, по существу, как описано в примере 9, за исключением того, что использовали клетки А431 (АТСС CRL 1555). Клетки А431 являются клетками эпидермоидной карциномы человека, которые сверхэкспрессируют EGFR. Мышей с установленными подкожными опухолями объемом примерно 250 мм 3 обрабатывали в 1, 8, 15, 22 и 29 день 500 мкг антитела 2.13.2 (в/б) или обрабатывали 1 раз в день в течение 27 дней 10 мг/кг СР-358774, вводимым перорально (п/о) либо в виде отдельных агентов, либо в комбинации, как описано в примере 9. СР-358774 описан в патенте США 5747498 и Moyer et al., Cancer Research 57: 4838-4848 (1997), включенных в данное описание в виде ссылки. На фиг. 16 показан размер опухолей в зависимости от различных обработок в течение времени. Эксперимент показывает, что обработка анти-IGF-IR-антителом усиливает эффективность СР-358774, известного ингибитора тирозинкиназы EGF-R, при ингибировании роста опухоли эпидермоидной карциномы человека. Пример 17. Фармакокинетика анти-IGF-IR-антител in vivo. Чтобы оценить фармакокинетику анти-IGF-IR-антител, макакам-крабоедам внутривенно инъецировали 3,30 или 100 мг/кг антитела 2.13.2 в ацетатном буфере. Сыворотку обезьян собирали в разных временных точках и определяли концентрации анти-IGF-IR-антитела у обезьян в течение периода вплоть до 10-недельных уровней. Чтобы количественно оценить уровни функциональных антител в сыворотке, внеклеточный домен IGF-IR человека (IGF-I-sR, R &D Systems, номер в каталоге 391GR) связывали с 96-луночными планшетами. В анализируемые планшеты добавляли сыворотку обезьян (разведенную от 1:100 до 1:15000) так, чтобы каждый образец находился в линейных пределах стандартной кривой, и инкубировали в условиях, при которых любое анти-IGF-IR-антитело связывалось бы с IGF-I-sR. После промывки планшетов в планшеты добавляли меченое антитело против IgG человека и инкубировали в условиях, при которых антитело против IgG человека связывалось бы с анти-IGF-IR-антителом. Затем планшеты промывали и проявляли и использовали стандартную контрольную кривую и подгонки с использованием линейной регрессии, чтобы определить количество анти-IGF-IR-антител. На фиг. 17 показана концентрация 2.13.2 в сыворотке в течение времени. Эксперимент показывает, что время полужизни анти-IGF-IR-антитела составляет от 4,6 до 7,7 дней и имеет объем распределения 74-105 мл/кг. Кроме того, эксперимент показывает, что вводимые количества у обезьян пропорциональны по дозе, что свидетельствует о том, что анти-IGF-IR-антитело заняло любые доступные сайты связывания IGF-IR в организме даже при самой низкой дозе 3 мг/кг. Пример 18. Комбинаторная терапия анти-IGF-IR-антителом и адриамицином приводит к понижающей регуляции IGF-IR in vivo. Опухоли индуцировали у "голых" мышей, как описано в примере 9. Мышей с установленными подкожными опухолями объемом примерно 400 мм 3 обрабатывали однократной инъекцией 250 мкг антитела 2.13.2 (в/б) или 7,5 мг/кг адриамицина (в/в) либо в виде отдельных агентов, либо в комбинации, как описано в примере 9. Через 72 ч после введения агентов опухоли экстрагировали, как описано в примере 8, и равные количества экстрактов опухолей подвергали электрофорезу в полиакриламидном геле с додецилфосфатом натрия (SDS-ПААГ) и Вестерн-блот-анализу с использованием анти-IGF-IR-антитела SC-713 (Santa Cruz). На фиг. 18 показаны количества IGF-IR в опухолевых клетках у контрольных животных (первые три дорожки каждой панели), у животных, которых обрабатывали только антителом (верхняя панель), у животных, которых обрабатывали только адриамицином (средняя панель), и у животных, которых обрабатывали антителом и адриамицином (нижняя панель). Каждая дорожка представляет равные количества белка из отдельных опухолей от отдельных мышей. Эксперимент показывает, что обработка только адриамицином незначительно влияет на уровни IGF-IR и что при обработке только антителом наблюдается некоторое снижение уровней IGF-IR. Неожиданно при обработке адриамицином вместе с антителом наблюдается резкое снижение уровней IGF-IR, что свидетельствует о том, что адриамицин и антитело вызывают значительную понижающую регуляцию уровней IGF-IR. Все публикации и заявки на выдачу патентов, цитированные в данном описании, включены в данное описание в виде ссылки так же, как в случае, когда для каждой отдельной публикации или заявки на выдачу патента было бы конкретно и отдельно указано, что они включены в виде ссылки. Хотя в целях четкого понимания вышеприведенное изобретение было описано подробно посредством иллюстраций и примеров, специалисты в данной области легко поймут в свете инструкций данного изобретения, что могут быть осуществлены некоторые изменения и модификации изобретения без отхода от сути и без превышения объема прилагаемой формулы изобретения. Заявители: компания "Пфайзер Инк." и др. Дата подачи: 20 декабря 2001г. (20.12.01). Название: "Антитела к рецептору инсулиноподобного фактора роста I". Справ. № агента: ABX-PF/2 РСТ. Все указания, относящиеся к депонированному биологическому материалу, содержатся в описании. Ниже приведены дополнительные указания, которые не должны входить в описание, и их следует рассматривать как "отдельные указания". Они относятся только к экспертному решению. Дополнительные указания, приведенные ниже, относятся к депонированному биологическому материалу, который упоминается как "гибридома 4.9.2." в описании на стр. 75, строки 8-18. Материал депонирован в следующем депозитарии: "Американская коллекция типовых культур" (АТСС) 10801 Юниверсити Булевард, Манассас, штат Виржиния 20110-2209 Соединенные Штаты Америки 12 декабря 2000г., под номером РТА 2789. Дополнительные указания. В отношении Канады: Касательно Канады, до выдачи патента Канады или до даты, когда по заявке будет вынесено решение об отказе, или если заявка изъята из рассмотрения и больше не подлежит восстановлению или отозвана, как оговорено Правилами 107 и 108 Патентных Правил в соответствии с Законом Канады о патентах, образцы депонированного биологического материала будут доступны только в виде образца, предоставляемого независимому эксперту, назначенному Комиссионером (Правило 104(4)). В отношении Европатента: Касательно Европатента, до публикации указания о выдаче Европатента или до истечения 20 лет от даты подачи заявки, если по заявке было вынесено решение об отказе или заявка была отозвана или считается отозванной, как оговорено в Правиле 28(3) Правил по применению Европейской патентной конвенции, образцы депонированного биологического материала будут предоставляться только посредством выдачи образца эксперту, назначенному лицом, направившим запрос (Правило 28(4) ЕРС). В отношении Финляндии: Касательно Финляндии, до публикации указания о выдаче патента Национальным советом по патентам и регистрациям или в течение 20 лет от даты подачи заявки, если в результате рассмотрения заявки патент выдан не был, образцы депонированного биологического материала будут предоставляться только эксперту в данной области. В отношении Великобритании: Касательно Великобритании, заявитель настоящим уведомляет о своем желании, чтобы образцы депонированного биологического материала предоставлялись только эксперту в данной области. В отношении Исландии: Касательно Исландии, пока патент не будет выдан Исландским патентным ведомством или Исландским патентным ведомством не будет принято решение, в результате рассмотрения патентной заявки, об отказе в выдаче патента, образцы депонированного биологического материала могут выдаваться только эксперту в данной области. В отношении Швеции: Касательно Швеции, пока заявка не будет представлена на изучение общественности (Шведским патентным ведомством) или пока Шведским патентным ведомством не будет принято окончательное решение по заявке (без предоставления заявки на изучение общественности), образцы депонированного биологического материала могут выдаваться только эксперту в данной области. В отношении Сингапура: Заявитель настоящим уведомляет о своем желании, чтобы образцы вышеуказанной культуры предоставлялись только экспертам в соответствии с параграфом 3 Четвертой таблицы Патентных правил 1995г. |